
Functionally diverse multiple xylanases of thermophilic fungus Myceliophthora sp. IMI 387099
7
Enzyme and Microbial Technology 35 (2004) 460–466 Functionally diverse multiple xylanases of thermophilic fungus Myceliophthora sp. IMI 387099 A.K. Badhan a , B.S. Chadha a,∗ , K.G. Sonia a , H.S. Saini a , M.K. Bhat b a Department of Microbiology, Guru Nanak Dev University, Amritsar 143005, Punjab, India b Food Materials Science Division, Institute of Food Research, Norwich Research Park, Colney, Norwich NR4 7UA, UK Received 9 June 2004; accepted 8 July 2004 Abstract The paper reports the multiplicity of xylanase in a newly isolated thermophilic fungus Myceliophthora sp. IMI 387099. The expression of different xylanase isoforms and level was found to be dependent on substrate type, as well as culture conditions. The zymograms developed from proteins resolved on PAGE and IEF showed rice straw as the best inducer. Of the 10 xylanases produced in presence of rice straw, the major xylanases were of acidic pI. Each of the 10 xylanases was functionally diverse and showed different specificity and catalytic activity against a variety of substituted/unsubstituted xylan and aryl substituted substrates. The major acidic xylanases showed no preferential activity towards xylan (substituted/unsubstituted), however, the basic and moderately acidic xylanases showed higher activities towards substituted xylans (LWX, RAX, WAX). © 2004 Elsevier Inc. All rights reserved. Keywords: Myceliophthora sp.; Xylanase isoforms; Zymograms; Substrate specificity 1. Introduction Xylan is a major hemicellulose constituent of hardwood, softwood and straws [1], and after cellulose, is the next most abundant renewable polysaccharide [2]. Xylan is het- erogeneous polysaccharide consisting of a homopolymeric backbone of 1,4-linked--d-xylopyranose units and short chain branches including O-acetyl, -l-arabinofuranosyl, - d-glucuronyl residues [3]. The side chain branches, position and their composition differ in hemi-cellulosic fraction de- Abbreviations: BWX, birch wood xylan; OSX, oat spelt xylan; LWX, larch wood xylan; RAX, rye arabino xylan; WAX, wheat arabino xy- lan; CMC, carboxy methyl cellulose; MGX, 4-O-methylglucouranoxylan; DA, debranched arabinan; pNPX, p-nitrophenyl--d-xylopyranoside; pNPG, p-nitrophenyl--d-glucopyranoside; pNPC, p-nitrophenyl--d- cellobioside; pNP--l-Araf, p-nitrophenyl--l-arabinofuranoside; pNP-- l-Arap, p-nitrophenyl--l-arabinopyranoside; pNPMan, p-nitrophenyl--d- mannopyranoside; pNPGa, p-nitrophenyl--d-galactoside; RS, rice straw; CC, corncobs; BAG, baggase; WB, wheat bran; WS, wheat straw ∗ Corresponding author. Tel.: +91 183 2450938; fax: +91 183 2258820. E-mail address: [email protected] (B.S. Chadha). rived from different plant sources. The complexity of xylan requires the action of multiple xylanases with overlapping yet different specificities to effect extensive hydrolysis [4].A variety of microorganisms are reported to produce endoxy- lanases that can degrade -1,4-xylan in a random fashion yielding a series of linear and branched oligosaccharide frag- ments [5,6]. These xylanases also differ in the number of sub-sites and the location of the active sites among the sub- sites, which plays an important role in xylo-oligosaccharide hydrolysis [5]. Several microorganisms including thermophilic fungi viz. Talaromyces byssochlamydoides, Melanocarpus albomyces, Humicola insolens (syn. Scytalidium thermophilum), Chaetomium thermophile, produce multiple endoxylanases as a strategy to hydrolyze the xylan effectively [7–11]. Different isoforms of xylanases apparently cooperate to enhance hydrolysis of complex xylan in lignocelluloses. Literature cites many examples of xylanase multiplicity and classifying them as major and minor xylanases with respect to their expression levels as identified by clear bands on xylanase zymograms [12]. The purification and 0141-0229/$ – see front matter © 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.enzmictec.2004.07.002
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Functionally diverse multiple xylanases of thermophilic fungus Myceliophthora sp. IMI 387099

Enzyme and Microbial Technology 35 (2004) 460–466
Functionally diverse multiple xylanases of thermophilic fungusMyceliophthorasp. IMI 387099
A.K. Badhana, B.S. Chadhaa,∗, K.G. Soniaa, H.S. Sainia, M.K. Bhatb
a Department of Microbiology, Guru Nanak Dev University, Amritsar 143005, Punjab, Indiab Food Materials Science Division, Institute of Food Research, Norwich Research Park, Colney, Norwich NR4 7UA, UK
Received 9 June 2004; accepted 8 July 2004
Abstract
The paper reports the multiplicity of xylanase in a newly isolated thermophilic fungusMyceliophthorasp. IMI 387099. The expression ofdifferent xylanase isoforms and level was found to be dependent on substrate type, as well as culture conditions. The zymograms developedfrom proteins resolved on PAGE and IEF showed rice straw as the best inducer. Of the 10 xylanases produced in presence of rice straw, themajor xylanases were of acidic pI. Each of the 10 xylanases was functionally diverse and showed different specificity and catalytic activitya ntial activityt substitutedx©
K
1
smebcda
llDpclmC
laning
oxy-nfrag-r ofsub-
ride
viz.s
es
e toes.
citywithlear
d
0d
gainst a variety of substituted/unsubstituted xylan and aryl substituted substrates. The major acidic xylanases showed no prefereowards xylan (substituted/unsubstituted), however, the basic and moderately acidic xylanases showed higher activities towardsylans (LWX, RAX, WAX).2004 Elsevier Inc. All rights reserved.
eywords: Myceliophthorasp.; Xylanase isoforms; Zymograms; Substrate specificity
. Introduction
Xylan is a major hemicellulose constituent of hardwood,oftwood and straws[1], and after cellulose, is the nextost abundant renewable polysaccharide[2]. Xylan is het-rogeneous polysaccharide consisting of a homopolymericackbone of 1,4-linked-�-d-xylopyranose units and shorthain branches includingO-acetyl,�-l-arabinofuranosyl,�--glucuronyl residues[3]. The side chain branches, positionnd their composition differ in hemi-cellulosic fraction de-
Abbreviations:BWX, birch wood xylan; OSX, oat spelt xylan; LWX,arch wood xylan; RAX, rye arabino xylan; WAX, wheat arabino xy-an; CMC, carboxy methyl cellulose; MGX, 4-O-methylglucouranoxylan;A, debranched arabinan; pNPX,p-nitrophenyl-�-d-xylopyranoside;NPG, p-nitrophenyl-�-d-glucopyranoside; pNPC,p-nitrophenyl-�-d-ellobioside; pNP-�-l-Araf, p-nitrophenyl-�-l-arabinofuranoside; pNP-�--Arap,p-nitrophenyl-�-l-arabinopyranoside; pNPMan,p-nitrophenyl-�-d-annopyranoside; pNPGa,p-nitrophenyl-�-d-galactoside; RS, rice straw;C, corncobs; BAG, baggase; WB, wheat bran; WS, wheat straw∗ Corresponding author. Tel.: +91 183 2450938; fax: +91 183 2258820.E-mail address:[email protected] (B.S. Chadha).
rived from different plant sources. The complexity of xyrequires the action of multiple xylanases with overlappyet different specificities to effect extensive hydrolysis[4]. Avariety of microorganisms are reported to produce endlanases that can degrade�-1,4-xylan in a random fashioyielding a series of linear and branched oligosaccharidements[5,6]. These xylanases also differ in the numbesub-sites and the location of the active sites among thesites, which plays an important role in xylo-oligosacchahydrolysis[5].
Several microorganisms including thermophilic fungiTalaromyces byssochlamydoides, Melanocarpus albomyce,Humicola insolens (syn. Scytalidium thermophilum),Chaetomium thermophile,produce multiple endoxylanasas a strategy to hydrolyze the xylan effectively[7–11].Different isoforms of xylanases apparently cooperatenhance hydrolysis of complex xylan in lignocellulosLiterature cites many examples of xylanase multipliand classifying them as major and minor xylanasesrespect to their expression levels as identified by cbands on xylanase zymograms[12]. The purification an
141-0229/$ – see front matter © 2004 Elsevier Inc. All rights reserved.oi:10.1016/j.enzmictec.2004.07.002

A.K. Badhan et al. / Enzyme and Microbial Technology 35 (2004) 460–466 461
characterization of the major xylanases is often reported,however the nature and relevance of minor xylanases remainelusive owing to obvious difficulties in their purification.
This study reports the induction of multiple forms of xy-lanases in a thermophilic fungusMyceliophthorasp. IMI387099 as well as characterizing these xylanases on the ba-sis of their expression level, pI and substrate specificity tohighlight their functional diversity.
2. Materials and methods
2.1. Enzyme production
Different natural lignocellulosics (agro-residues) namely,rice straw, wheat straw, baggase, corncob and wheatbran were used as carbon sources for the solid substrateproduction of xylanases. Medium used for productioncontained (NH4)2SO4, 0.3%; KH2PO4, 0.3%; ammoniumacetate, 0.6%; pH, 7.5. Each flask containing 5 g of substrate(particle size 2–8 mm) and 15 ml of production medium wasinoculated with 2 ml (2× 107) of spore suspension preparedfrom 5 days old culture grown on YpSS medium. SSF wascarried out at 45◦C for 5 days. The crude enzyme wasextracted by addition of 50 ml acetate buffer (50 mM, pH5.0) and then filtered and centrifuged. The clear supernatantsw 3).T umn(
2
E)w ingM mli[ o-e thei elc linec nine0 orica ssingw and2
2
tedb oser ibedb sist 0 for3 for3
2
anew d for
24–48 h at 4◦C in 500�l sodium citrate buffer (50 mM; pH6.0) containing 0.02% sodium azide. The eluted proteins fromthe gels in each fraction were assayed for xylanase activityagainst birch wood xylan.
2.5. Substrate specificity
The xylanase activity was determined in different gelfractions against birch wood xylan. The xylanase active peakfractions were also assayed for specificity against differentsubstrates including polysaccharides (BWX, OSX, LWX,RAX, WAX, CMC, MGX, DA) as well as pNP derivatives(pNPX, pNPC, pNP-�-l-Araf, pNP-�-l-Arap, pNPMan,pNPGa). Activities on polysaccharides were determined byincubating 1 ml reaction mixture containing 100�l enzyme,500�l of 1% substrate and 400�l of citrate buffer (50 mM,pH 6.0) at 50◦C for 1 h. The reducing sugars releasedwere quantified using DNS at 540 nm. Activity on pNPderivatives was determined by incubating 100�l reactionmixture containing 25�l enzyme, 25�l of substrate (3 mM)and 50�l of citrate buffer (50 mM, pH 6.0) at 50◦C for 2 h.The reaction was stopped by adding 0.4 M glycine-NaOHbuffer, pH 10.8. The color developed was read at 405 nmusing ELISA reader (Multiskan, Labsystems).
3
3f
el-os axi-
Fd rates,r
ere subjected to lyophilization (HETO, Drywinnerhe lyophilized sample was desalted using PD-10 colPharmacia) and used in further experiments.
.2. Electrophoresis and Isoelectric focussing
Native-polyacrylamide gel electrophoresis (PAGas performed in 7.5% gel with 4% stacking gel usini-Protean II system of Biorad by method of Laem
13]. Total protein (70�g) was loaded in each well. Islectric focussing (IEF) was performed according to
nstruction provided by Novex, using 5% acrylamide gontaining 2.4% broad pH range (3.5–10.0) amphoarrier ampholyte. The cathode buffer contained argi.35% (w/v) and lysine 0.29% (w/v), whereas, phosphcid (10 mM) was used as anode buffer. Isoelectrofocuas carried out for 1 h each at constant voltage of 10000 V followed with 500 V for 30 min.
.3. Activity staining
Xylanase activity in native and IEF gels was detecy activity staining (Zymogram technique) with agareplica containing covalently dyed RBB-xylan as descry Biely et al. [14]. Upon completion of electrophore
he gels were incubated in sodium acetate buffer pH 6.0 min and then overlaid on RBB-xylan containing gel0 min–1 h at 50◦C.
.4. Quantification of xylanase in IEF gel
After fractionating the protein on IEF, the gel in each las sliced (1.25 mm thickness). Each slice was incubate
. Results
.1. Effect of carbon source on production of multipleorms of xylanases
The results inFig. 1show the xylanase zymograms devped after native PAGE. It was observed thatMyceliophthorap., when cultured under SSF, on rice straw induced m
ig. 1. Pattern of xylanolytic enzyme system produced byMyceliophthorauring growth (SSF) in the presence of various lignocellulosic substesolved by native PAGE and detected with RBB-xylan overlay.
462 A.K. Badhan et al. / Enzyme and Microbial Technology 35 (2004) 460–466
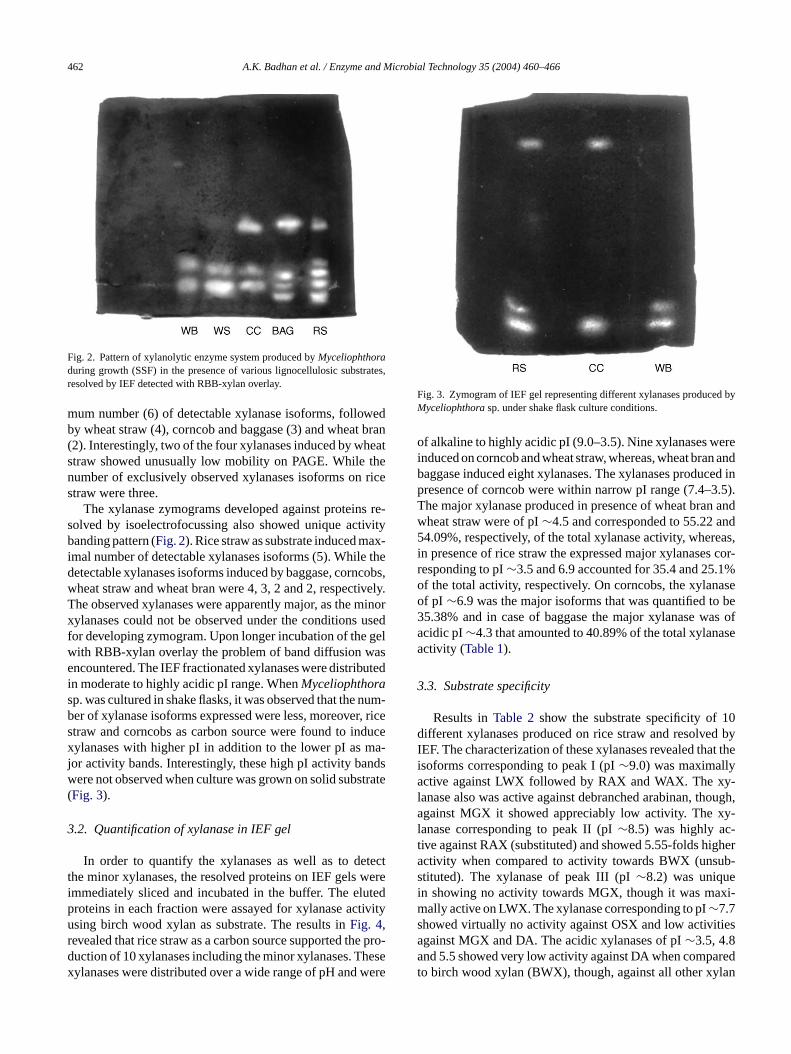
Fig. 2. Pattern of xylanolytic enzyme system produced byMyceliophthoraduring growth (SSF) in the presence of various lignocellulosic substrates,resolved by IEF detected with RBB-xylan overlay.
mum number (6) of detectable xylanase isoforms, followedby wheat straw (4), corncob and baggase (3) and wheat bran(2). Interestingly, two of the four xylanases induced by wheatstraw showed unusually low mobility on PAGE. While thenumber of exclusively observed xylanases isoforms on ricestraw were three.
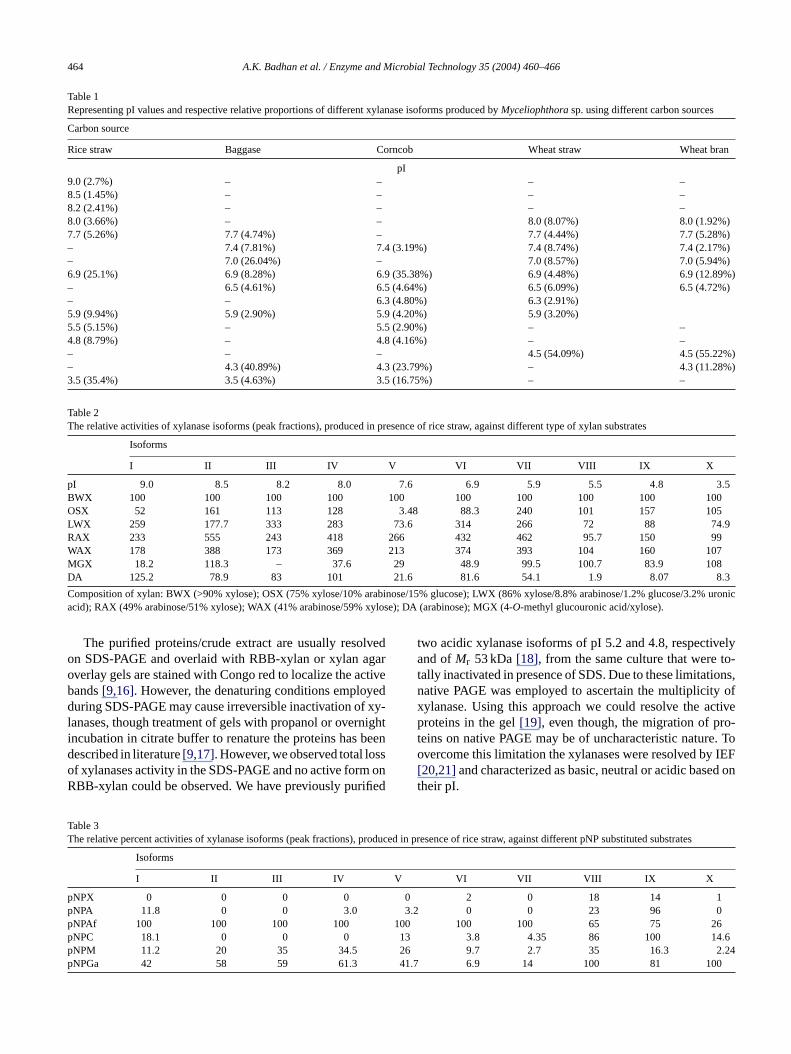
The xylanase zymograms developed against proteins re-solved by isoelectrofocussing also showed unique activitybanding pattern (Fig. 2). Rice straw as substrate induced max-imal number of detectable xylanases isoforms (5). While thedetectable xylanases isoforms induced by baggase, corncobs,wheat straw and wheat bran were 4, 3, 2 and 2, respectively.The observed xylanases were apparently major, as the minorxylanases could not be observed under the conditions usedfor developing zymogram. Upon longer incubation of the gelwith RBB-xylan overlay the problem of band diffusion wasencountered. The IEF fractionated xylanases were distributedin moderate to highly acidic pI range. WhenMyceliophthorasp. was cultured in shake flasks, it was observed that the num-ber of xylanase isoforms expressed were less, moreover, ricestraw and corncobs as carbon source were found to inducexylanases with higher pI in addition to the lower pI as ma-jor activity bands. Interestingly, these high pI activity bandswere not observed when culture was grown on solid substrate(Fig. 3).
3
tectt werei utedp tivityur e prod hesex were
Fig. 3. Zymogram of IEF gel representing different xylanases produced byMyceliophthorasp. under shake flask culture conditions.
of alkaline to highly acidic pI (9.0–3.5). Nine xylanases wereinduced on corncob and wheat straw, whereas, wheat bran andbaggase induced eight xylanases. The xylanases produced inpresence of corncob were within narrow pI range (7.4–3.5).The major xylanase produced in presence of wheat bran andwheat straw were of pI∼4.5 and corresponded to 55.22 and54.09%, respectively, of the total xylanase activity, whereas,in presence of rice straw the expressed major xylanases cor-responding to pI∼3.5 and 6.9 accounted for 35.4 and 25.1%of the total activity, respectively. On corncobs, the xylanaseof pI ∼6.9 was the major isoforms that was quantified to be35.38% and in case of baggase the major xylanase was ofacidic pI∼4.3 that amounted to 40.89% of the total xylanaseactivity (Table 1).
3.3. Substrate specificity
Results inTable 2show the substrate specificity of 10different xylanases produced on rice straw and resolved byIEF. The characterization of these xylanases revealed that theisoforms corresponding to peak I (pI∼9.0) was maximallyactive against LWX followed by RAX and WAX. The xy-lanase also was active against debranched arabinan, though,against MGX it showed appreciably low activity. The xy-lanase corresponding to peak II (pI∼8.5) was highly ac-t ghera ub-s ei xi-ms iesaa redt lan
.2. Quantification of xylanase in IEF gel
In order to quantify the xylanases as well as to dehe minor xylanases, the resolved proteins on IEF gelsmmediately sliced and incubated in the buffer. The elroteins in each fraction were assayed for xylanase acsing birch wood xylan as substrate. The results inFig. 4,evealed that rice straw as a carbon source supported thuction of 10 xylanases including the minor xylanases. Tylanases were distributed over a wide range of pH and
-
ive against RAX (substituted) and showed 5.55-folds hictivity when compared to activity towards BWX (unstituted). The xylanase of peak III (pI∼8.2) was uniqun showing no activity towards MGX, though it was ma
ally active on LWX. The xylanase corresponding to pI∼7.7howed virtually no activity against OSX and low activitgainst MGX and DA. The acidic xylanases of pI∼3.5, 4.8nd 5.5 showed very low activity against DA when compa
o birch wood xylan (BWX), though, against all other xy

A.K. Badhan et al. / Enzyme and Microbial Technology 35 (2004) 460–466 463
Fig. 4. Fractionation ofMyceliophthorasp. IMI 387099 xylanolytic components grown on different lignocellulosic substrates, by isoelectric focussing inpolyacrylamide gel.
substrate types the activities were comparable. Very low ac-tivity against xyloglucan (Tamarind) was observed (data notshown) in xylanases IX and X.
The results inTable 3showed that xylanases correspond-ing to peaks I, V–X were active against pNPC with no orvery little activity against pNPX whereas, the xylanases, II,III, and IV, showed no activity against these two substrates.The basic and moderately acidic xylanases showed relativelyhigher activities towards pNP-�-l-Araf, and consequentlyalso were more active against debranched arabinan. Though,these isoforms showed very less activity against the pyranoseform (pNP-�-l-Arap). The peaks corresponding to acidic pIshowed low activities against pNP-�-l-Araf and DA, whencompared to other xylanases. The xylanase VIII (pI∼5.5)
was catalytically versatile being active against a wide rangeof aryl-substituted substrates.
4. Discussion
The production of multiple isoforms of xylanase by a novelthermophilic fungal strainMyceliophthorasp. IMI 387099 isbeing reported here. The induction of xylanases was found tobe dependent on type and composition of carbon source usedfor culturing. Similar observations have been made earlier inthermophilic fungus,Melanocarpus albomycesIIS86[9,15].We observed that method of culturing (SSF/shake flask) alsoinfluenced the induction of specific xylanases.
464 A.K. Badhan et al. / Enzyme and Microbial Technology 35 (2004) 460–466
Table 1Representing pI values and respective relative proportions of different xylanase isoforms produced byMyceliophthorasp. using different carbon sources
Carbon source
Rice straw Baggase Corncob Wheat straw Wheat bran
pI9.0 (2.7%) – – – –8.5 (1.45%) – – – –8.2 (2.41%) – – – –8.0 (3.66%) – – 8.0 (8.07%) 8.0 (1.92%)7.7 (5.26%) 7.7 (4.74%) – 7.7 (4.44%) 7.7 (5.28%)– 7.4 (7.81%) 7.4 (3.19%) 7.4 (8.74%) 7.4 (2.17%)– 7.0 (26.04%) – 7.0 (8.57%) 7.0 (5.94%)6.9 (25.1%) 6.9 (8.28%) 6.9 (35.38%) 6.9 (4.48%) 6.9 (12.89%)– 6.5 (4.61%) 6.5 (4.64%) 6.5 (6.09%) 6.5 (4.72%)– – 6.3 (4.80%) 6.3 (2.91%)5.9 (9.94%) 5.9 (2.90%) 5.9 (4.20%) 5.9 (3.20%)5.5 (5.15%) – 5.5 (2.90%) – –4.8 (8.79%) – 4.8 (4.16%) – –– – – 4.5 (54.09%) 4.5 (55.22%)– 4.3 (40.89%) 4.3 (23.79%) – 4.3 (11.28%)3.5 (35.4%) 3.5 (4.63%) 3.5 (16.75%) – –
Table 2The relative activities of xylanase isoforms (peak fractions), produced in presence of rice straw, against different type of xylan substrates
Isoforms
I II III IV V VI VII VIII IX X
pI 9.0 8.5 8.2 8.0 7.6 6.9 5.9 5.5 4.8 3.5BWX 100 100 100 100 100 100 100 100 100 100OSX 52 161 113 128 3.48 88.3 240 101 157 105LWX 259 177.7 333 283 73.6 314 266 72 88 74.9RAX 233 555 243 418 266 432 462 95.7 150 99WAX 178 388 173 369 213 374 393 104 160 107MGX 18.2 118.3 – 37.6 29 48.9 99.5 100.7 83.9 108DA 125.2 78.9 83 101 21.6 81.6 54.1 1.9 8.07 8.3
Composition of xylan: BWX (>90% xylose); OSX (75% xylose/10% arabinose/15% glucose); LWX (86% xylose/8.8% arabinose/1.2% glucose/3.2% uronicacid); RAX (49% arabinose/51% xylose); WAX (41% arabinose/59% xylose); DA (arabinose); MGX (4-O-methyl glucouronic acid/xylose).
The purified proteins/crude extract are usually resolvedon SDS-PAGE and overlaid with RBB-xylan or xylan agaroverlay gels are stained with Congo red to localize the activebands[9,16]. However, the denaturing conditions employedduring SDS-PAGE may cause irreversible inactivation of xy-lanases, though treatment of gels with propanol or overnightincubation in citrate buffer to renature the proteins has beendescribed in literature[9,17]. However, we observed total lossof xylanases activity in the SDS-PAGE and no active form onRBB-xylan could be observed. We have previously purified
Table 3The relative percent activities of xylanase isoforms (peak fractions), produced in presence of rice straw, against different pNP substituted substrates
Isoforms
I II III IV V VI VII VIII IX X
pNPX 0 0 0 0 0 2 0 18 14 1pNPA 11.8 0 0 3.0 3.2 0 0 23 96 0pNPAf 100 100 100 100 100 100 100 65 75 26pNPC 18.1 0 0 0 13 3.8 4.35 86 100 14.6pNPM 11.2 20 35 34.5 26 9.7 2.7 35 16.3 2.24pNPGa 42 58 59 61.3 41.7 6.9 14 100 81 100
two acidic xylanase isoforms of pI 5.2 and 4.8, respectivelyand ofMr 53 kDa[18], from the same culture that were to-tally inactivated in presence of SDS. Due to these limitations,native PAGE was employed to ascertain the multiplicity ofxylanase. Using this approach we could resolve the activeproteins in the gel[19], even though, the migration of pro-teins on native PAGE may be of uncharacteristic nature. Toovercome this limitation the xylanases were resolved by IEF[20,21]and characterized as basic, neutral or acidic based ontheir pI.

A.K. Badhan et al. / Enzyme and Microbial Technology 35 (2004) 460–466 465
The multiple forms of xylanases are usually designatedas major and minor xylanases based on the intensity of theclear bands on the RBB-xylan. Whereas, this may not bethe exact conclusion as commercial RBB-xylan is preparedusing MGX (SIGMA) as substrate, which may not be the pre-ferred substrate for the xylanase being assayed. Interestinglythe major xylanases of acidic pI characterized on the basisof observed activity bands on RBB-xylan were active againstMGX during enzyme assay of the eluted fractions. This pointmust, therefore, be carefully considered because some of ap-parently defined minor xylanases, on the basis of in-situ lowactivity bands on RBB-xylan, in fact could be highly activeon other xylan types as shown inTable 2.
The slicing of the IEF gels and incubating each frac-tion in a suitable buffer for overnight or longer incubationensured proper elution of proteins. Similar approach hasbeen reported to quantify the multiple forms of endoglu-canases fromPenicillium pinophilum[22]. The activity ofeach fraction against birch wood xylan helped us to iden-tify the xylanase isoforms peaks. The relative activity ofeach of the constituent xylanase in the enzyme complex alsopointed to the differential expression of each isoforms andwas dependent upon the substrate type. It has been recog-nized that characterization of single xylanase or with singlesubstrate is inadequate for demonstrating the contribution ofif ea-s uted)a on ofx ryingl -l ylan,w drol-y n theb e asc fam-i ses.A cat-a[
5
1 laythe
2 fer-m as
3 hasof
es.pu-
ates
Acknowledgement
The financial support in the form of ISIS fellowshipgranted by BBSRC and DBT government of India for car-rying out this work is duly acknowledged.
References
[1] Timeli TE. Recent progress in the chemistry of wood hemicellulose.Wood Sci Technol 1967;1:45–70.
[2] Biely P. Microbial xylanolytic systems. Trends Biotechnol1985;3:286–90.
[3] Whistler RL, Richards EL. Hemicellulose. In: Pigman W, HortonD, editors. The carbohydrate, vol. 2a. New York: Academic Press;1970. p. 447–69.
[4] Dekker RFH, Richards GN. Hemicellulases: their occurrence, pu-rification, properties and mode of action. Adv Carbohydr ChemBiochem 1976;32:277–352.
[5] Kormelink FJM, Gruppen H, Vietor RJ, Voragen AGJ. Modeof action of the xylan-degrading enzyme fromAspergillusawamori on alkali-extractable cereal arabinoxylans. Carbohydr Res1993;249:355–67.
[6] Bhat MK, Hazlewood GP. Enzymology and other characteristics ofcellulases and xylanases. In: Bedford MR, Partridge CC, editors. En-zymes in farm animal nutrition. Wallingford, UK: CAB International;2001. p. 11–60.
[7] Hayashida SK, Ohta, Mo K, Wood AGJ. Xylanases ofTalaromycesin8.eir88.de-t
[ ca-
[ iza-.
[ev
[ mbly
[ led T,nic
88. p.
[ and
[ ll P,xyl A8.
[ f xy-
[ tivegus
[ f an
ndividual enzymes to the xylanolytic systems[12]. There-ore, the catalytic potential of each of the xylanase was mured against different xylan (substituted and unsubstits well as pNP-substituted compounds. The compositiylan substrates used for assaying xylanases was of vaevel of heterogeneity (Table 2). In general, the alkaline xyanases demonstrated higher specificity for substituted xhereas the acidic xylanases were more versatile in hysis of both substituted and unsubstituted substrates Oasis of relatively higher activity towards pNP-cellobiosompared to pNPX, xylanase I, V–X can be classified asly 10 xylanases, while others may be family 11 xylanas a rule, only the endoxylanases of family 10 canlyze hydrolysis of cellulase substrate, aryl-�-d-cellobioside
23].
. Conclusions
. In addition to substrate type, culturing conditions pan important role in expression and level of each ofmultiple xylanases produced.
. The substrate specificity, catalytic activity against difent substrate and pI of each of xylanase reveal thedistinct xylanase.
. The use of electrophoretic techniques (IEF/PAGE)been found useful in studying the functional diversitymultiple forms of xylanases including minor xylanasThis data can form the basis for devising strategies torify the minor xylanases using suitable xylan substrfor identifying active fractions during purification.
bussochlamydoides. In: Kellogg T, Partridge ST, editors. Methodsenzymology, vol. 160. London: Academic Press; 1988. p. 675–
[8] Maheshwari R, Bharadwaj G, Bhat MK. Thermophilic fungi: thphysiology and enzymes. Microbiol Mol Biol Rev 2000;63:461–
[9] Saraswat V, Bisaria SV. Biosynthesis of xylanolytic and xylanbranching enzymes inMelanocarpus albomycesIIS 68. J FermenBioeng 1997;83:352–7.
10] Dusterhoft EM, Linseen VAJM, Voragen AGJ, Beldman G. Purifition, characterization and properties of two xylanases fromHumicolainsolens. Enzyme Microb Technol 1997;20:437–45.
11] Ganju RK, Vithayathil PJ, Murthy SK. Purification and charactertion of two xylanase fromChaetomium thermophilevar. coprophileCan J Microbiol 1989;35:836–42.
12] Wong KKY, Tan LUL, Saddler JN. Multiplicity of�-14-xylanasein microorganisms: function and applications. Microbiol R1988;52:305–17.
13] Laemmli UK. Cleavage of structural proteins during the asseof the head of bacteriophage T4. Nature 1970;227:680–5.
14] Biely PD, Mislovicova, Toman B. Remazol Brilliant Blue-A solubchromogenic substrate for xylanases. In: Enzymology A, WooKellogg ST, editors. Remazol Brilliant Blue-A soluble chromogesubstrate for xylanases, vol. 160. London: Academic Press; 19536–41.
15] Morag E, Bayer EA, Lamed R. Relationship of cellulosomalnon cellulosomal xylanases ofClostridium thermocellumto cellulosedegrading enzymes. J Bacteriol 1990;172:6098–105.
16] Chavez R, Schachter K, Navarro C, Peirano A, Aguirre C, Buet al. Differences in expression of two endo-xylanases genes (and Xyl B) from Penicillum purpurogenum. Gene 2002;293:161–
17] Lee YE, Lowe SE, Zeikus JG. Regulation and characterization olanolytic enzymes ofThermoanaerobacterium saccharolyticumB6A-RI. Appl Environ Microbiol 1993;59:763–71.
18] Chadha BS, Ajay BK, Mellon F, Bhat MK. Two endoxylanases acand stable at alkaline pH from the newly isolated thermophilic funMyceliophthorasp. IMI 387099. J Biotechnol 2004;104:227–37.
19] Montiel MD, Hernandez M, Rodriguez J, Arias ME. Evaluation oendo�-mannanase produced byStreptomyces ipomoeaCECT 3341

466 A.K. Badhan et al. / Enzyme and Microbial Technology 35 (2004) 460–466
for the biobleaching of pine Kraft pulps. Appl Microbiol Biotechnol2002;58:67–72.
[20] Sachstehner A, Nidetzky B, Kulbe DK, Haltrich D. Induction ofmannanase, xylanase and endoglucanase activities inSclerotium rolf-sii. Appl Environ Microbiol 1998;64:594–600.
[21] Gomez de Segura B, Durand R, Fevre M. Multiplicity and expressionof xylanases in the rumen fungusNeocallimastix frontalis. FEMSMicrobiol Lett 1998;164:47–53.
[22] Bhat MK, Wood TM. Multiple forms of endo-1,4-�-d-glucanase inthe extracellular cellulase ofPencillium pinophilum. Biotechnol Bio-eng 1989;33:1242.
[23] Tilbeurgh H, Claeyssens M. Detection and differentiation of cellulasecomponent using low molecular mass fluorogenic substrate. FEMSMicrobiol Lett 1985;187:283–8.




















