
Enzymology of the thermophilic ascomycetous fungus Thermoascus aurantiacus
11
Review Enzymology of the thermophilic ascomycetous fungus Thermoascus aurantiacus Michel BRIENZO, Valdeir ARANTES, Adriane M. F. MILAGRES* Department of Biotechnology, Escola de Engenharia de Lorena University of Sa ˜o Paulo – USP Lorena, Estrada Municipal do Campinho, s/n – CP 116, 12602-810 Lorena/SP, Brazil article info Article history: Received 28 February 2008 Received in revised form 6 April 2009 Accepted 21 April 2009 Keywords: Amylases Cellulases Enzyme application Pectinases Thermoascus aurantiacus Xylanases abstract Different strains of the thermophilic ascomycetous fungus Thermoascus aurantiacus have been reported in the literature to produce high levels of a variety of industrial interest enzymes (i.e. amylases, cellulases, pectinases and xylanases), which have been shown to be remarkably stable over a wide range of temperatures and appear to have tremendous commercial potential. Most studies on enzyme production by T. aurantiacus are carried out in chemically defined liquid medium, under conditions suitable for induction of a particular enzyme. A few studies have investigated the production of some enzymes by T. aurantiacus by solid-state fermentation, using lignocellulosic materials. The present review focuses on the enzymes produced by T. aurantiacus, their main kinetic parameters, and the effect of different culture conditions on production and enzyme activity. It also provides a view of the possible applications of T. aurantiacus enzymes, considering that this thermophilic fungus could comprise a potential source of thermostable enzymes. ª 2009 The British Mycological Society. Published by Elsevier Ltd. All rights reserved. 1. Introduction Thermoascus aurantiacus is a thermophilic fungus that belongs to the ascomycetous class, first isolated from selfheating hay by Hugo Miehe in 1907 as reported by Cooney and Emerson (1964). T. aurantiacus has a bright orange color, elliptical asco- spores, and no asexual stage (Maheshwari et al., 2000), although in the past the description given by Cooney and Emerson (1964) depicted it as having an asexual stage. So far, about 19 T. auran- tiacus strains have been classified (Table 1). Optimum and maximum temperatures for T. aurantiacus ascospore germination have been reported to be about 47.5 C and 60 C, respectively (Deploey, 1995), which were higher than those reported for hyphal growth (40–45 C and 55 C, respectively) (Cooney and Emerson, 1964). The minimum temperature for ascospore germination (32.5 C– 35 C) (Deploey, 1995), is also higher than the minimum for hyphal growth (20 C–25 C) (Cooney and Emerson, 1964). In addition, the percentage of germination has been shown to be slightly affected by culture age (Deploey, 1995). Noack (1920) studied the behavior of T. aurantiacus sub- jected to anaerobiosis and found that the withdrawal of oxygen severely affected its respiration and growth. He also investigated the effects of subminimal temperatures and * Corresponding author. Tel.: þ55 1231595000; fax: þ55 1231533165. E-mail addresses: [email protected] (M. Brienzo), [email protected] (V. Arantes), [email protected] (A. M.F. Milagres). journal homepage: www.elsevier.com/locate/fbr fungal biology reviews 22 (2008) 120–130 1749-4613/$ – see front matter ª 2009 The British Mycological Society. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.fbr.2009.04.001
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Enzymology of the thermophilic ascomycetous fungus Thermoascus aurantiacus

f u n g a l b i o l o g y r e v i e w s 2 2 ( 2 0 0 8 ) 1 2 0 – 1 3 0
journa l homepage : www.e lsev ie r . com/ loca te / fbr
Review
Enzymology of the thermophilic ascomycetous fungusThermoascus aurantiacus
Michel BRIENZO, Valdeir ARANTES, Adriane M. F. MILAGRES*
Department of Biotechnology, Escola de Engenharia de Lorena University of Sao Paulo – USP Lorena, Estrada Municipal do Campinho,
s/n� – CP 116, 12602-810 Lorena/SP, Brazil
a r t i c l e i n f o
Article history:
Received 28 February 2008
Received in revised form
6 April 2009
Accepted 21 April 2009
Keywords:
Amylases
Cellulases
Enzyme application
Pectinases
Thermoascus aurantiacus
Xylanases
* Corresponding author. Tel.: þ55 12315950E-mail addresses: [email protected]
(A. M.F. Milagres).1749-4613/$ – see front matter ª 2009 The Bdoi:10.1016/j.fbr.2009.04.001
a b s t r a c t
Different strains of the thermophilic ascomycetous fungus Thermoascus aurantiacus have
been reported in the literature to produce high levels of a variety of industrial interest
enzymes (i.e. amylases, cellulases, pectinases and xylanases), which have been shown to
be remarkably stable over a wide range of temperatures and appear to have tremendous
commercial potential. Most studies on enzyme production by T. aurantiacus are carried
out in chemically defined liquid medium, under conditions suitable for induction of
a particular enzyme. A few studies have investigated the production of some enzymes
by T. aurantiacus by solid-state fermentation, using lignocellulosic materials. The present
review focuses on the enzymes produced by T. aurantiacus, their main kinetic parameters,
and the effect of different culture conditions on production and enzyme activity. It also
provides a view of the possible applications of T. aurantiacus enzymes, considering that
this thermophilic fungus could comprise a potential source of thermostable enzymes.
ª 2009 The British Mycological Society. Published by Elsevier Ltd. All rights reserved.
1. Introduction 47.5 �C and 60 �C, respectively (Deploey, 1995), which were
Thermoascus aurantiacus is a thermophilic fungus that belongs
to the ascomycetous class, first isolated from selfheating hay
by Hugo Miehe in 1907 as reported by Cooney and Emerson
(1964). T. aurantiacus has a bright orange color, elliptical asco-
spores, and no asexual stage (Maheshwari et al., 2000), although
in the past the description given by Cooney and Emerson (1964)
depicted it as having an asexual stage. So far, about 19 T. auran-
tiacus strains have been classified (Table 1).
Optimum and maximum temperatures for T. aurantiacus
ascospore germination have been reported to be about
00; fax: þ55 1231533165.p.br (M. Brienzo), ara
ritish Mycological Society
higher than those reported for hyphal growth (40–45 �C and
55 �C, respectively) (Cooney and Emerson, 1964). The
minimum temperature for ascospore germination (32.5 �C–
35 �C) (Deploey, 1995), is also higher than the minimum for
hyphal growth (20 �C–25 �C) (Cooney and Emerson, 1964). In
addition, the percentage of germination has been shown to
be slightly affected by culture age (Deploey, 1995).
Noack (1920) studied the behavior of T. aurantiacus sub-
jected to anaerobiosis and found that the withdrawal of
oxygen severely affected its respiration and growth. He also
investigated the effects of subminimal temperatures and
[email protected] (V. Arantes), [email protected]
. Published by Elsevier Ltd. All rights reserved.

Table 1 – Classified strains of T. aurantiacus
Strain References
T. aurantiacus 179-5 Martins et al., 2002
T. aurantiacus 26904 Feldman et al., 1988
T. aurantiacus 34115 Feldman et al, 1988
T. aurantiacus A-131 Kawamori et al, 1987
T. aurantiacus ATCC 204492 Milagres et al, 2004
T. aurantiacus BJT 190 Alam et al., 1994
T. aurantiacus BT 2079 Gomes et al., 2007
T. aurantiacus CBMAI-756 Martins et al., 2007
T. aurantiacus IF0 9862 Wang et al., 1998
T. aurantiacus IFO 31693 Ko et al., 2005b
T. aurantiacus IFO 9748 Hong et al., 2003a
T. aurantiacus II S96 Katapodis and Christakopoulos, 2004
T. aurantiacus IMI 173038 Katapodis and Christakopoulos, 2004
T. aurantiacus IMI 216529 Parry et al., 2001
T. aurantiacus Miehe Ortner et al., 2006
T. aurantiacus NBRC 31693 Ko et al., 2005a
T. aurantiacus SL 16 W Kongbuntad et al., 2006
T. aurantiacus TUB F43 Nampoothiri et al., 2004
T. aurantiacus var. levisporus Upahyay et al., 1984
Hydrolases of Thermoascus aurantiacus 121
rewarming and observed that when an actively growing
culture of T. aurantiacus was cooled just a little lower than
the minimum temperature for ascospore germination, its
respiration after 2 d declined only minimally, but those
cultures kept at 21 �C for 24 h stopped respiring. When the
culture medium was rewarmed to 46 �C, practically no respi-
ration was observed, showing the low resistance to lower
temperature. However, until more information is available,
it should not necessarily be assumed that mycelial cultures
of thermophilic fungi cannot be stored under refrigeration or
at subminimal temperatures without loss of viability.
T. aurantiacus has been shown to grow efficiently on ligno-
cellulosic biomass from agro-industrial and forest wastes
(Da Silva et al., 2005; Kalogeris et al., 2003; Milagres et al.,
2004; Santos et al., 2003; Yu et al., 1987). The good growth adap-
tation on such materials of high abundance and low cost is an
attractive and promising alternative for production of
enzymes.
The following sections describe the main physico-chemi-
cal characteristics of hydrolytic enzymes produced by T. aur-
antiacus. The considerable biotechnological potential of most
of these enzymes is also briefly discussed.
Cellulases
Cellulose, the major constituent of plant cell walls, consists
of an unbranched polymer of cellobiose, and is one of the
most abundant organic compounds in the biosphere. Cellu-
lose crystallinity and the large amount of hydrogen bonds
are two essential factors affecting the biodegradation of
native cellulose; a group of enzymes acting in a synergistic
manner is required to facilitate complete cleavage of the
cellulose bonds to form glucose. Cellulose hydrolyzing
enzymes (cellulases) are divided into three major groups:
endo-1,4-b-D-glucanases (EC 3.2.1.4), exo-1,4-b-D-glucanases
(EC 3.2.1.91) (or cellobiohydrolase) and b-glucosidases (EC
3.2.1.21), all three attacking b-1,4-glycosidic bonds.
T. aurantiacus can produce all cellulolytic enzymes required
for complete degradation of cellulose to glucose. However, as
reported for some other well known cellulase-producing
microorganisms such as Trichoderma reesei, Termonospora fusca
and Aspergillus niger endo-1,4-b-D-glucanase is the main cellu-
lase activity produced by T. aurantiacus (Himmel et al., 1999).
Efforts to increase the performance of cellulase mixtures for
the hydrolysis of pretreated biomass have sometimes focused
on the selection of enzymes with high thermal stability to
enable operations at higher temperatures, which implies
possibilities for prolonged storage increased and reduce risks
of contamination. A summary of some relevant published
data for the properties of cellulases from different strains of
T. aurantiacus is given in Table 2.
b-glucosidaseb-glucosidase production by T. aurantiacus is highly depen-
dent on the culture conditions. Thus, larchwood xylan, blot-
ting paper, bagasse, pectin (Khandke et al., 1989a), enhance
the production of b-glucosidase. It was suggested that in
T. aurantiacus more than one type of b-glucosidase exists
and have important role in the formation of endo and exo-
glucanase inducers from cellulose (Feldman et al., 1988; De
Palma-Fernandez et al., 2002; Tong et al., 1980). In general,
its b-glucosidases show high stability, half lives at 70 �C of
23.5 h (Gomes et al., 2000), optimum temperature and pH
between 65 �C and 80 �C, and 4.5 �C and 6 �C, respectively
(Hong et al., 2006; Parry et al., 2001). Leite et al. (2007) observed
a first-order kinetic for thermo inactivation of b-glucosidase
from T. aurantiacus 179-5 when exposed to temperatures
higher than 80 �C.
Kalogeris et al. (2003) identified b-glucosidases from T. aur-
antiacus IMI 216529 having a pI of 3.9. An extracellular b-gluco-
sidase purified from T. aurantiacus IMI 216529 has a molecular
weight of 120 kDa (based on gel permeation) hydrolyzed disac-
charides with b,1-1, b,1-6 and b,1-2 glucosidic bonds. The pNP-
cellooligosaccharides were used to demonstrate that the
enzyme cleaves one glucose unit at a turn from the non-
reducing end (Parry et al., 2001). A high degree of N-terminal
amino acid sequence identity between T. aurantiacus b-gluco-
sidase and other b-glucosidases of family 3 suggested that the
enzyme could be a member of glycoside hydrolase family 3
(Parry et al., 2001).
The synergism in a mixed culture of T. aurantiacus and
A. niger on oat straw semi-solid fermentation resulted in
high levels of b-glucosidase activity (500 IU/g), there were
2.5-fold higher than the activity observed when A. niger was
grown in monoculture (Stoilova et al., 2005).
A b-glucosidase gene (bgl2) of T. aurantiacus IFO 9748 was
isolated and expressed in Pichia pastoris KM71H. The recombi-
nant enzyme retained 50 % of its activity in the presence of
1.39 M of glucose, while the wild enzyme was inhibited at
low glucose concentrations.
Endo-1,4-b-D-glucanaseEndoglucanases cleave randomly b-glycosidic bonds in b-1,4-
glucan chains to produce free chain ends. Different T. auran-
tiacus strains have been shown to produce endoglucanases
with pIs (around 3.5–3.7) (Kalogeris et al., 2003; Parry et al.,
2002). In general, endoglucanases from T. aurantiacus display
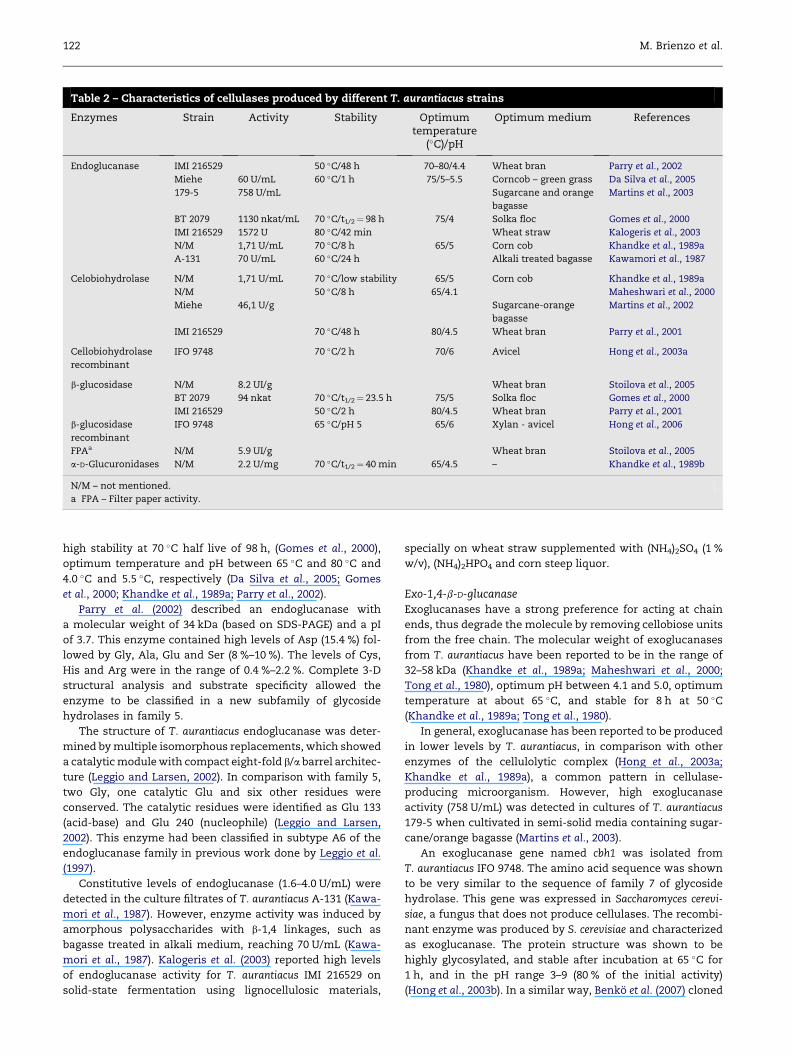
Table 2 – Characteristics of cellulases produced by different T. aurantiacus strains
Enzymes Strain Activity Stability Optimumtemperature
(�C)/pH
Optimum medium References
Endoglucanase IMI 216529 50 �C/48 h 70–80/4.4 Wheat bran Parry et al., 2002
Miehe 60 U/mL 60 �C/1 h 75/5–5.5 Corncob – green grass Da Silva et al., 2005
179-5 758 U/mL Sugarcane and orange
bagasse
Martins et al., 2003
BT 2079 1130 nkat/mL 70 �C/t1/2¼ 98 h 75/4 Solka floc Gomes et al., 2000
IMI 216529 1572 U 80 �C/42 min Wheat straw Kalogeris et al., 2003
N/M 1,71 U/mL 70 �C/8 h 65/5 Corn cob Khandke et al., 1989a
A-131 70 U/mL 60 �C/24 h Alkali treated bagasse Kawamori et al., 1987
Celobiohydrolase N/M 1,71 U/mL 70 �C/low stability 65/5 Corn cob Khandke et al., 1989a
N/M 50 �C/8 h 65/4.1 Maheshwari et al., 2000
Miehe 46,1 U/g Sugarcane-orange
bagasse
Martins et al., 2002
IMI 216529 70 �C/48 h 80/4.5 Wheat bran Parry et al., 2001
Cellobiohydrolase
recombinant
IFO 9748 70 �C/2 h 70/6 Avicel Hong et al., 2003a
b-glucosidase N/M 8.2 UI/g Wheat bran Stoilova et al., 2005
BT 2079 94 nkat 70 �C/t1/2¼ 23.5 h 75/5 Solka floc Gomes et al., 2000
IMI 216529 50 �C/2 h 80/4.5 Wheat bran Parry et al., 2001
b-glucosidase
recombinant
IFO 9748 65 �C/pH 5 65/6 Xylan - avicel Hong et al., 2006
FPAa N/M 5.9 UI/g Wheat bran Stoilova et al., 2005
a-D-Glucuronidases N/M 2.2 U/mg 70 �C/t1/2¼ 40 min 65/4.5 – Khandke et al., 1989b
N/M – not mentioned.
a FPA – Filter paper activity.
122 M. Brienzo et al.
high stability at 70 �C half live of 98 h, (Gomes et al., 2000),
optimum temperature and pH between 65 �C and 80 �C and
4.0 �C and 5.5 �C, respectively (Da Silva et al., 2005; Gomes
et al., 2000; Khandke et al., 1989a; Parry et al., 2002).
Parry et al. (2002) described an endoglucanase with
a molecular weight of 34 kDa (based on SDS-PAGE) and a pI
of 3.7. This enzyme contained high levels of Asp (15.4 %) fol-
lowed by Gly, Ala, Glu and Ser (8 %–10 %). The levels of Cys,
His and Arg were in the range of 0.4 %–2.2 %. Complete 3-D
structural analysis and substrate specificity allowed the
enzyme to be classified in a new subfamily of glycoside
hydrolases in family 5.
The structure of T. aurantiacus endoglucanase was deter-
mined by multiple isomorphous replacements, which showed
a catalytic module with compact eight-fold b/a barrel architec-
ture (Leggio and Larsen, 2002). In comparison with family 5,
two Gly, one catalytic Glu and six other residues were
conserved. The catalytic residues were identified as Glu 133
(acid-base) and Glu 240 (nucleophile) (Leggio and Larsen,
2002). This enzyme had been classified in subtype A6 of the
endoglucanase family in previous work done by Leggio et al.
(1997).
Constitutive levels of endoglucanase (1.6–4.0 U/mL) were
detected in the culture filtrates of T. aurantiacus A-131 (Kawa-
mori et al., 1987). However, enzyme activity was induced by
amorphous polysaccharides with b-1,4 linkages, such as
bagasse treated in alkali medium, reaching 70 U/mL (Kawa-
mori et al., 1987). Kalogeris et al. (2003) reported high levels
of endoglucanase activity for T. aurantiacus IMI 216529 on
solid-state fermentation using lignocellulosic materials,
specially on wheat straw supplemented with (NH4)2SO4 (1 %
w/v), (NH4)2HPO4 and corn steep liquor.
Exo-1,4-b-D-glucanaseExoglucanases have a strong preference for acting at chain
ends, thus degrade the molecule by removing cellobiose units
from the free chain. The molecular weight of exoglucanases
from T. aurantiacus have been reported to be in the range of
32–58 kDa (Khandke et al., 1989a; Maheshwari et al., 2000;
Tong et al., 1980), optimum pH between 4.1 and 5.0, optimum
temperature at about 65 �C, and stable for 8 h at 50 �C
(Khandke et al., 1989a; Tong et al., 1980).
In general, exoglucanase has been reported to be produced
in lower levels by T. aurantiacus, in comparison with other
enzymes of the cellulolytic complex (Hong et al., 2003a;
Khandke et al., 1989a), a common pattern in cellulase-
producing microorganism. However, high exoglucanase
activity (758 U/mL) was detected in cultures of T. aurantiacus
179-5 when cultivated in semi-solid media containing sugar-
cane/orange bagasse (Martins et al., 2003).
An exoglucanase gene named cbh1 was isolated from
T. aurantiacus IFO 9748. The amino acid sequence was shown
to be very similar to the sequence of family 7 of glycoside
hydrolase. This gene was expressed in Saccharomyces cerevi-
siae, a fungus that does not produce cellulases. The recombi-
nant enzyme was produced by S. cerevisiae and characterized
as exoglucanase. The protein structure was shown to be
highly glycosylated, and stable after incubation at 65 �C for
1 h, and in the pH range 3–9 (80 % of the initial activity)
(Hong et al., 2003b). In a similar way, Benko et al. (2007) cloned

Hydrolases of Thermoascus aurantiacus 123
an exoglucanase from T. aurantiacus and expressed it in T. ree-
sei Rut-C30 using the cbhl promoter.
Applications of thermostable cellulasesThe significant industrial importance of cellulases was prob-
ably peaked during the 1990’s, mainly within the textile, deter-
gent and cellulosic pulp industries. In the current industrial
processes, cellulolytic enzymes are employed in: (i) clarifica-
tion of juices and wines; (ii) detergents causing color bright-
ening and softening; (iii) pretreatment of biomass to
improve nutritional quality of forage; and (iv) pretreatment
of industrial wastes (Bhat, 2000; Haki and Rakshit, 2003).
For instance, the biopolishing process of cotton in the
textile industry requires cellulases stable at high tempera-
ture (Ando et al., 2002). During this process, the cellulases
act on small fiber ends that protrude from the fabric surface,
where the mechanical action removes these fibers and
polishes the fabrics (Sreenath et al., 1996). Cellulases can
improve drainage of recycled pulps by hydrolyzing the amor-
phous hydrophilic cellulose, which is the main constituent of
the fines formed during refining (Oksanen et al., 2000). Endo-
glucanase treatment can also decrease viscosity and increase
the alkaline solubility of dissolving pulp (Rahkamo et al.,
1998).
Nowadays, the conversion of lignocellulosic into ferment-
able sugar is the main focus for biofuel production. In this
matter, the lignocellulosic conversion has clear advantages
when thermostable cellulases are employed, as well as low
activity losses occur during processing, even at elevated
temperature often used in raw material pre-treatments.
Some studies have shown that for high conversion efficiency,
a long reaction time is required (Sassner et al., 2008; Zhu et al.,
2008; Borjesson et al., 2007; Ohgren et al., 2007), so, considering
that most studies were performed near 50 �C, the use of ther-
mostable enzymes, such as T. aurantiacus cellulases would
guarantee that enzymes would still remain stable until the
end of the process, improving significantly the hydrolysis
process. In general, thermostable enzymes have enjoyed
much attention because of their advantages in usage under
industrial conditions since mass transfer is facilitated at
higher temperatures due to decreased fluid viscosity; contam-
ination risks are reduced and enzymes can be purified and
stored at room temperature without inactivation (Krahe
et al., 1996). However, due to low specific growth, rates large
scale commercial cultivation of thermophiles for enzyme
production remains an economical challenge (Santos et al.
2003).
Hemicellulases
Hemicelluloses are the second most abundant polysaccha-
rides present in the cell walls of herbaceous and woody plants.
Xylans of many plant materials are heteropolysaccharides
with homopolymeric backbone chains of 1,4-linked b-D-xylo-
pyranose units. Besides xilose, xylans may contain arabinose,
glucuronic acid or its 4-0-methyl ether, and acetic, ferulic, and
p-coumaric acids. The frequency and composition of
branches are dependent on the source of xylanare the most
abundant hemicellulose-type polysaccharides constituent in
plants (Ward and Moo-Young, 1989).
Synergistic effects among hemicellulases have been docu-
mented in many works (Coughlan and Hazlewood, 1993;
Biely et al., 1986; Tenkanen et al., 1992). The key factors
that influence xylan hydrolysis are chain length and degree
of substitution (Li et al., 2000). The main enzymes involved
in hemicellulose degradation are endo-b-1,4-xylanase (EC
3.2.1.8) and b-xylosidase (EC 3.2.1.37) (acting in the main
chain), and enzymes that act on the substituted groups,
called accessory enzymes, such as ferulic acid esterase (EC
3.1.1.73), p-coumaric acid esterase (EC 3.1.1.-), acetylxylan
esterase (EC 3.1.1.6), a-arabinofuranosidase (EC 3.2.1.55) and
a-glucuronidase (EC 3,2,1,131) (Beg et al., 2001; Polizeli et al.,
2005).
T. aurantiacus can produce most of the hemicellulolytic
enzymes, however, endoxylanase is the main enzyme
detected in its culture. Many well known hemicellulase-
producing microorganisms, such as A. niger (Kang et al.,
2004; Yuan et al., 2005) and T. reesei (Tenkanen et al., 1992)
possess complete xylan-degrading enzyme systems. Some
characteristics of T. aurantiacus hemicellulases are described
in the following sections, and additional detailed properties
of hemicellulases from various strains of T. aurantiacus are
presented in Table 3.
Endo-b-1,4-xylanaseEndo-b-1,4-xylanases cleave the internal xylan chain and
release short xylooligosaccharides (Shallom and Shoham,
2003). T. aurantiacus endoxylanases are in the range of
33–39.7 kDa (Kalogeris et al., 2001; Viswamitra et al., 1993),
and have been shown to be highly active and thermo-
stable. T. aurantiacus C 436 produces endoxylanases that
are stable at 50 �C for 12 weeks, retaining 90 % of the initial
activity (Yu et al., 1987). Alam et al. (1994) determined that
the strain BJT 190 retains 80 % of initial activity after incu-
bation for 15 d at 55 �C.
Xylanases from different strains of T. aurantiacus have been
shown to be optimally active at about 70–75 �C and around pH
5 (Alam et al., 1994; Gomes et al., 2000; Yu et al., 1987). More-
over, Da Silva et al. (2005) reported that xylanase from T. auran-
tiacus CBMAI-756 isolated in Brazil was stable in a wide range
of pH (3–9) at room temperature.
Only two isozymes active on xylan, one with a pI 6.8 and
the other with a pI of 5.5, have been reported to be produced
by T. aurantiacus IMI 216529 (Kalogeris et al., 2001). This may
imply that in general T. aurantiacus does not produce many
xylanases isozymes, thus differ from other filamentous fungi
that have demonstrated a great capability for secreting a wide
range of xylanases.
Solid-state fermentation of T. aurantiacus has been shown
to produce maximum xylanase activities in a short period of
time (Kalogeris et al., 1998; Martins et al., 2007; Santos et al.,
2003; Souza et al., 1999). T. aurantiacus ATCC 204492 cultivated
on sugarcane bagasse displayed xylanase production depen-
dent on the aeration (Santos et al., 2003). Other variables
such as nitrogen and carbon sources have also been reported
to affect xylanase production by T. aurantiacus, as observed for
T. aurantiacus Miehe 216529 (Kalogeris et al., 1998). For T. auran-
tiacus ATCC 204492, the use of sugarcane bagasse with 80 %
initial moisture content was proven ideal for the xylanase
activity, whereas inoculum level and cultivation time were

Table 3 – Characteristics of hemicellulases produced by different T. aurantiacus strains
Enzymes Strain Activity Stability Optimumtemperature
(�C)/pH
Optimum medium References
Endoxylanase N/M 45.5 UI/g Wheat bran Stoilova et al., 2005
Miehe 107 U/mL 60 �C/1 h 75/5 Corncob – green grass Da Silva et al., 2005
ATCC 204492 1597 UI/g SCBa Milagres et al., 2004
N/M 500 U/g SCBaþ rice bran Santos et al., 2003
IMI 216529 6193 U/g t1/241 min/80 �C 70/4.5 Wheat straw Kalogeris et al., 1998
N/M 705.7 U/g 55 �C/15 d 70/5 Wheat bran Alam et al., 1994
C 436 575.9 U/mL 50 �C/84 d 75/5 Oat spelt xylan Yu et al., 1987
179-5 1288.8 U/g Sugarcane-orange
bagasse
Martins et al., 2003
b-xylosidase BT 2079 5 nkat/mL 75/5 Solka floc Gomes et al., 2000
ATCC 204492 27 UI/g SCBa Milagres et al., 2004
BT 2079 3479 nkat/mL Solka floc Gomes et al., 2000
C 436 1.63 U/mL t1/236 min/70 �C 75/5 Oat spelt xylan Yu et al., 1987
b-mannanase BT 2079 Low level Solka floc Gomes et al., 2000
b-manosidase N/M 10 nkat/mL 50 �C/5 h 76/2.5–3 Bean gum Gomes et al., 2007
b-galactosidase BT 2079 Low level Solka floc Gomes et al., 2000
a-galactosidase BT 2079 Low level Solka floc Gomes et al., 2000
a-arabinofuranosidase N/M 1083.5 nkat/g 50 �C/48 h 70/4 Sugar beet Roche et al., 1994
a-glucuronidase N/M 2.2 U/mg protein t1/240 min/70 �C 65/4.5 Khandke et al., 1989b
N/M – not mentioned.
a SCB – Sugarcane baggase.
124 M. Brienzo et al.
of lower significant effect on the production of enzyme
activity (Souza et al., 1999).
Commonly, endoxylanases are classified by substrate
specificity in family 10 or 11 based on hydrophobic cluster
analysis and amino acid sequence homologies. Enzymes
from family 10, in contrast to endoxylanase of family 11,
exhibit low substrate specificity and is capable of attacking
the xylosidic linkage next to the branch and towards the
non-reducing end. Kalogeris et al. (2001) concluded based on
the catalytic properties that the major endoxylanase purified
of T. aurantiacus belongs to glycosyl hydrolases of family 10.
Vardakou et al. (2003) revealed that xylanase from T. aurantia-
cus hydrolyzed two times more efficiently insoluble water-
unextractable arabinoxylans than the family 11 enzyme
showing better capability of cleaving glycosidic linkages in
the xylan main chain closer to the substituents. In spite of
higher molecular mass, xylanses of family 10 have smaller
substrate binding sites than xilanases of family 11 (Biely
et al., 1997).
b-xylosidaseThere are only a few reports dealing with b-xylosidase from
T. aurantiacus. It appears that b-xylosidase is produced at
lower levels by this fungus compared to some other hemicel-
lulolytic enzymes, such as endoxylanases. A study on the
ability of fourteen substrates, such as beech and birch wood
xylan and methyl b-D-xylopyranoside, for the production of
hemicellulases by T. aurantiacus indicated that only low levels
of constitutive b-xylosidase were produced by T. aurantiacus,
even in the presence of cited inducers (Gomes et al., 1994).
On the other hand, in the presence of wheat straw, a b-xylosi-
dase with an acidic pI (3.5) was induced (40 UI/g) in cultures of
the T. aurantiacus IMI 216529 (Kalogeris et al., 1998; Kalogeris
et al., 1999).
a-L-arabinofuranosidasea-L-arabinofuranosidases hydrolyze non-reducing arabinofur-
anose residues in short oligosaccharides and branched
arabinoxylans substituted at positions 2 and 3 on b-D-xylopyr-
anosyl (De Vries et al., 2000).
Low levels of exo-a-L-arabinofuranosidase were detected in
cultures of T. aurantiacus from solid-state fermentation of
leached sugar beet pulp based medium (Gomes et al., 2000).
Moreover, Roche et al. (1994) reported that the optimal
medium conditions for maximum arabinofuranosidase
production were sugar beet pulp at 77.8 % humidity supple-
mented with mineral solution and yeast extract (1.2 %) at pH
4.5. This arabinofuranosidase showed optimum temperature
and pH at 70 �C and 4 �C, respectively, and was stable for
48 h at 50 �C.
a-D-glucuronidasea-D-glucuronidases cleave the a-1,2-glycosidic bond of the
4-O-methyl-D-glucuronic acid side chain of xylans (Puls et al.,
1987).
A purified a-D-glucuronidase from T. aurantiacus with a single
polypeptide chain and molecular weight of 118 kDa (based on
SDS-PAGE), cleaved the a,1-2 linkage between 4-O-methyl-a-D-
glucuronic acid and the xylose residue in xylan and several glu-
curono-xylooligosaccharides. The enzyme was stable at 70 �C
with a half live of 40 min, optimum temperature and pH at
65 �C and 4.5 �C, respectively (Khandke et al., 1989b).
b-1,4-MannanaseMannan is composed of b-1,4-linked mannopyranose and glu-
copryranose units, which in softwood is present as acetylated
galactoglucomanans. b-1,4-mannanase (EC 3.2.1.78) cleaves
randomly within the 1,4-b-mannan (McCleary and Mathen-
son, 1986).

Hydrolases of Thermoascus aurantiacus 125
Araujo and Ward (1990) investigated the effect of carbon
and nitrogen sources on mannanase production by eight ther-
mophilic fungi. Mannose has displayed the greatest inducer
effect on mannose production by T. aurantiacus, and sodium
nitrate was the best nitrogen source. Gomes et al. (2007) eval-
uated different carbon and nitrogen sources and found that
the best sources for mannanase production (10 nktal/mL) by
T. aurantiacus were locust bean gum and soymeal. According
to Gomes and Steiner (2000) the enzyme was much more ther-
mostable than other mannanases from some other strains of
T. aurantiacus. In another work Gomes et al., 2000 also reported
simultaneous production of mannanase under optimized
conditions for xylanase and endoglucanase production by
T. aurantiacus BT 2079. Mannanase production was about 232
and 75-fold lower than xylanase and endoglucanase, respec-
tively, and similar to the level of a-galactosidase.
b-mannosidaseb-mannosidase (EC 3.2.1.25) catalyzes the hydrolysis of
mannose units from the non-reducing end of mannosides.
b-mannosidase is commonly produced at low levels by
various microorganisms and a similar pattern has also been
described for T. aurantiacus BT 2079 (Guy et al., 1997). Gomes
et al., 2007 detected a b-mannosidase in cultures of T. aurantia-
cus of 99.9 kDa (UREA-PAGE electrophoresis), pI 4.8, and stable
at 50 �C for 5 h. Among different inducers tested locust bean
gum (15 g/L), induced the highest b-mannosidase activity. In
addition, the enzyme also displayed transglycosylation
activity by transferring the mannosyl group from p-nitro-
phenyl-b-D-mannopyranoside to methyl-a-D-mannopyrano-
side for the synthesis of disaccharide.
Ortner et al. (2006) studied the characteristics of regioselec-
tivity and acceptor tolerance in transglycosylation reaction by
culture filtrates from T. aurantiacus, and observed that the
structure of the acceptor caused difference in the anomeric
configuration bond when b-mannopyranosyl fluoride was
employed as the donor.
Applications of thermostable hemicellulasesThe pool of hemicellulases, especially endoxylanases, b-
xylosidases, and the accessory enzymes (b-fructofuranosi-
dase, a-L-arabinosidase, b-mannosidase, etc) can be used
for the saccharification of the hemicellulosic fraction of
lignocellulosic materials for sugar production, which can
then be converted to chemicals and fuels in a similar
manner to that of cellulases in the conversion of cellulosic
materials. For instance, Cardona and Sanchez (2007) appli-
cation have shown that ethanol can be produced from
xylose in a simultaneous process, in which xylose is initially
isomerized and then fermented.
During the production of cellulosic pulp, lignin is removed
by a multistep bleaching process, in which the addition of
xylanases has proved to be an interesting and efficient alter-
native to improve pulp bleaching, besides collaborating with
environment preservation (Wong and Saddler, 1992, 1993; Vii-
kari et al., 1993). For example, in kraft processes, the wood
chips are cooked at a high temperature and alkaline pH. These
processes cause xylan dissolution which precipitates on the
surface of cellulose microfibrils, acting like a barrier to the
chemical removal of lignin. Alternatively, xylanase can
efficiently remove xylan chains exposing lignin to the action
of bleaching reagents. For this purpose, the most appropriate
xylanases would be thermostable and alkaline active xyla-
nases (Battan et al., 2007; Jacques et al., 2000). In this viewpoint,
T. aurantiacus xylanases appears to fulfill these requirements,
and so, would seem to have great potential for biobleaching of
cellulosic pulps.
Besides xylanases, hemicellulases are comprised of
several other enzymes whose potential have not been fully
explored. Arabinofuranosidase can be used as auxiliary in
the conversion of hemicellulose substrate to ferment
sugars. In the some way, it can be used in improvement
of animal feedstock digestibility, hydrolysis of grape mono-
terpenyl a-arabinofuranosylglucosides to increase aroma
during wine making, and clarification and thinning of juices
(Saha, 2000).
Amylases
Plants as cereal grains (e.g. wheat, rice, corn, sorghums, oats,
barley) store glucose as the polysaccharide starch. Amylases
can be divided into two classes, endoamylases (EC 3.2.1.1)
and exoamylases (EC 3.2.1.3). Enzymes belonging to the
second group, the exoamylases, either exclusively cleave
a,1-4-glycosidic bonds such as b-amylase (E.C.3.2.1.2) or cleave
a,1-6-glycosidic bonds like amyloglucosidase or glucoamylase
(EC 3.2.1.3) and a-glucosidase (EC 3.2.1.20) (Fogarty and kelly,
1990).
Only limited information is found in literature about
amylases of T. aurantiacus. It was reported that small quanti-
ties of amylase was produced in extracellular growth media
(Adams, 1992). However, some characteristics of T. aurantiacus
amylases are described below and some others are given in
Table 4.
Endo amylase and ExoamylaseEndo amylase (or a-amylase) hydrolyzes a-1,4 bond randomly
in amylose and amylopectin releasing maltooligosaccharides,
while exoamylases hydrolyzes the penultimate a-1,4 bond
starting from non-reducing end releasing maltose. (Guzman-
Maldonado and Peredes-lopez, 1995).
Ohno et al. (1998) used starch as carbon source for amylase
production by T. aurantiacus IFO 31693 and detected the
production of both exo (75 kDa) and endoamylase (56 kDa).
The authors observed different stability behavior for both
enzymes. The endo was more stable (60 �C/3 h) and showed
better activity at higher temperature (70 �C) than the exoamy-
lase (65 �C).
A mixed culture of T. aurantiacus and A. niger on oat straw
semi-solid fermentation resulted in higher levels of a-amylase
activity if compared to the activity detected in monoculture
(Stoilova et al., 2005).
a-glucosidasea-glucosidases are exoenzymes that hydrolyze the glucosidic
bond a-1,4 and/or a-1,6 in short oligosaccharides. They are
divided into three types: type I hydrolyzes heterogeneous
substrates (e.g. sucrose); type II hydrolyzes mainly maltose
and isomaltose and shows low activity on heterogeneous
substrates; and type III has the same specificity as type II,

Table 4 – Properties of pectinases and amylases produced by T. aurantiacus
Enzyme Strain Activity/Stability Temp opt (�C) pH opt Optimum medium References
Pectin Ko et al., 2005b
Polygalacturonase 179-5 43 U/g. Stable in the acidic
to neutral pH range and at
60 �C for 1 h
65 10.5–11.0 Wheat bran or orange
bagasse
Martins et al., 2002
Pectin lyase 179-5 40180 U/g. Stable at acidic
pH and at 60 �C for 5 h
65 5.0 sugarcane bagasseþorange bagasse
Martins et al., 2002
Pectin esterase IFO 31693 0.156 U/mL Pectin Ko et al., 2005b
Endoamylase IFO 31693 60 �C for 3 h 70 4.5 Starch Ohno et al., 1998
Exoamylase IFO 31693 65 5.0 Starch Ohno et al., 1998
126 M. Brienzo et al.
but also hydrolyzes polysaccharides such as amylose (Frand-
sen and Svensson, 1998).
Carvalho et al. (2006) detected a-glucosidase in cultures of
T. aurantiacus 179-5 grown in solid-state fermentation using
wheat bran as substrate. The enzyme showed specific activity
for oligomers of maltose, however starch, dextrins (2–7 G) and
a-PNPG was very slowly hydrolyzed, suggesting that the
enzyme is a a-glucosidase type II. It also showed to be very
thermostable, with half-life of 15 min at 80 �C, and stable in
a wide range of pH (3–10).
Applications of thermostable amylasesProcessed starch is mainly used for glucose, maltose, and
oligosaccharide production, but a number of other products/
intermediates can also be produced via cyclodextrins (Turner
et al., 2007).
Starch processing is usually performed in a two-step
hydrolysis process of liquefaction and saccharification.
During liquefaction starch is gelatinized by thermal treatment
requiring high temperature. A thermostable a-amylase is
added before the heat treatment. Consequently, a highly ther-
mostable enzyme is required to be active during the whole
procedure. T. aurantiacus has an extracellular a-amylase
enzyme that shows promising characteristics for applications
in the starch industry due to its high thermostability in the
absence of metal ions and is active even at pH 4.0 (Ohno
et al., 1998). The saccharification of the remaining oligosaccha-
rides to produce a hydrolyzate syrup with more than 95 %
glucose is done by using exo-acting glucoamylase and pullula-
nase that efficiently hydrolyzes a,1-6-glycosidic bonds (Maarel
et al., 2002).
Pectinases
Pectins are the third main structural polysaccharide group of
plant cell walls. It is abundant in sugar beet pulp and fruit,
e.g. in citrus fruit and apple, where it can form up to half of
the polymeric content of the cell wall (Brummell, 2006). It is
composed mainly of linearly connected b-1,4-D-galacturonic
acid units and their methyl esters, interrupted in places by
1,2-linked L-rhamnose units (Willats et al., 2001).
For complete hydrolytic degradation of this component,
a set of enzymes, including endo-polygalacturonases (EC
3.2.1.15), exo-polygalacturonases (EC 3.2.1.67), lyases (EC
4.2.2.10) and pectin esterases (EC 3.1.1.11) (Gummadi and
Panda, 2003) are required.
The characteristics of some of the pectinases produced by
T. aurantiacus are described below and some others are shown
in Table 4.
PolygalacturonasesPolygalacturonases (PGases) catalyze the hydrolytic cleavage
of the polygalacturonic acid chain through introduction of
water across the oxygen bridge. They are the most extensively
studied enzyme among the family of pectinolytic enzymes.
The PGases involved in the hydrolysis of pectinaceous
substances are endo-PGase and exo-PGase. Endo-PGases are
widely distributed among fungi, bacteria and many yeasts,
whereas exo-PGases occur less frequently (Jayani et al., 2005).
It is generally agreed that the optimum medium for the
enhanced production of extracellular pectinase is that con-
taining pectic materials as an inducer (Martins et al., 2002).
T. aurantiacus 179-5 grown in solid-state fermentation
using wheat bran or orange bagasse produced the maximum
activity (43 U/g), whereas a decrease of PGase production
was observed when mixed to sugarcane bagasse. Exo-PGase
had optimum activity at pH 5.0, and was stable at pH values
of 7.0–8.0 and maintained 85 % of its activity at pH 4.0–6.5.
The optimum temperature was 65 �C, and at 70 �C, 88 % of
maximum activity was retained. According to the authors,
exo-PGase production by T. aurantiacus 179-5 was high
compared to those reported for pectinolytic microorganisms,
such as A. niger, A. foedidus, Penicillum italicum and Penicillium
frequentans (Martins et al., 2002).
Among forty thermophilic fungi screened for the produc-
tion of polygalacturonases, most of them did not show detect-
able activity, however, one isolate of T. aurantiacus produced
pectinases in medium containing 5 % (w/v) pectin, which
was markedly enhanced when citrus peel was used instead
of purified pectin. Polygalacturonase from T. aurantiacus was
one of the most stable, showing achieve at 65 �C with a half-
life of 20 min at 55 �C (Maheshwari et al., 2000).
Recently, an unusual polygalacturonase was isolated from
5-day culture filtrates of T. aurantiacus CBMAI-756 and purified
by gel filtration and ion-exchange chromatography (Martins
et al., 2007). Only one form of PG (30 kDa) was detected in
culture medium, which exhibited both endo and exo-PG activ-
ities. The enzyme was maximally active at pH 5.5 and 60 �C–
65 �C. The enzyme was 100 % stable at 50 �C for 1 h and
showed a half-life of 10 min at 60 �C. PG was also shown to
be stable at pH 5.0–5.5 and maintained 33 % of initial activity
at pH 9.0. Metal ions, such as Znþ2, Mnþ2, and Hgþ2, inhibited

Hydrolases of Thermoascus aurantiacus 127
50, 75 and 100 % of enzyme activity, respectively. According to
the authors, the purified enzyme showed to be less thermo-
stable than unpurified enzyme, suggesting the fungi may
produce some thermostabilizer factor.
Pectin lyasesPectin lyases (PL) produce a trans-eliminative split of the
pectic polymer, breaking the glycosidic linkages at C-4 and
simultaneously eliminating H from C-5, producing a D 4:5
unsaturated products (Jayani et al., 2005).
Martins et al. (2002) reported the growth of T. aurantiacus
179-5 and PL production in solid-state fermentation using
wheat bran, orange bagasse and mixtures of these materials
with sugarcane bagasse, and found that the medium con-
taining pectin, in their case orange bagasse, afforded higher
PL production than those with wheat bran. However, the
highest yields of PL activity were obtained by the addition
of sugarcane bagasse to orange bagasse. Maximal PL
activity (40.18 U/g) was obtained in a medium composed of
10 % (w/v) sugarcane and 90 % (w/v) orange bagasse. PL
exhibited maximal activity at pH 10.5–11.0 and 65 �C, retain-
ing 88 % of maximum activity at 70 �C. The authors also
observed that PL production by T. aurantiacus 179-5 was
high compared to other pectinolytic microorganisms, such
as A. niger, A. foedidus, P. italicum and P. frequentans (Martins
et al., 2002).
Applications of thermostable pectinasesPectinases have been applied in fruit juice clarification, juice
extraction, manufacture of pectin free starch, refinement of
vegetable fibers, degumming of natural fibers, waste-water
treatment, curing of coffee, cocoa and tobacco and as an
analytical tool in the assessment of plant products (Turner
et al., 2007). High temperatures are commonly employed in
such processes for different proposes. In most of these
processes the material submitted to heating needs to be
cooled down before enzymatic treatment. Thus, the use of
thermostable pectinases, such as the pectinases produced
by T. aurantiacus, could eliminate the cooling down step,
decreasing the final process costs. In other applications, it
can be more convenient to use thermostable enzymes, partic-
ularly when using substrates (which can also be other natu-
rally occurring glycoside-containing molecules with similar
linkages as in pectin) that are poorly soluble at ambient
temperatures, such as naringin and rutin, present in fruits
(Birgisson et al., 2004).
2. Concluding remarks
T. aurantiacus produces a large pool of enzymes of industrial
interest, i.e. amylases, cellulases, xylanases, etc., which have
been shown to be remarkably stable over a wide range of
temperature and pH. Many studies with T. aurantiacus have
been carried out on lignocellulosic materials, which were
shown to induce or increase production of several enzymes.
The mechanism involved is poorly understood, although it is
well established that the species secrete different types of
hydrolytic enzymes. The annotated gene set of T. aurantiacus,
transcript patterns, and comparative analyses with other
thermophilic fungi will advance our understanding of these
complex mechanisms involved in lignocellulose conversions.
Enzymes of T. aurantiacus are potentially applicable in
a wide range of industrial process mainly due to their extraor-
dinary operational stability at high temperatures and dena-
turant tolerance. Although such enzymes are described for
their ability in the chemical, food, pharmaceutical, paper,
textile and other industries, they are however, still today,
often limited for applications in small scale and testing of
such applications are under investigations (Gomes et al.,
2000; Milagres et al., 2003).
Recently, increasing attention has been devoted to the
utilization of biomass, especially lignocellulosic materials in
various industrial fields, as a possible alternative to fossil
resources because of their abundance, low cost and renew-
ability. The use of T. aurantiacus in biorefinery and for enzyme
production, especially lignocellulolytic enzymes, have gained
a great deal of interest as T. aurantiacus has good growth adap-
tation on lignocellulosic materials such as those originated
from agro-industrial and forest waste, and its enzymes ther-
mostability has tremendous advantages in biorefinery when
compared to the temperatures and pressures required for
chemicals conversion.
The study of T. aurantiacus has added greatly to our under-
standing of thermostable enzyme production and biochem-
ical features. It can be seen as a fungus of great importance
to bio-industrial processes due to its high thermal stability
and its enzymatic systems. However, much more work is
required to fully explore its biotechnological potential, espe-
cially in regard to the molecular and genetic fields.
r e f e r e n c e s
Adams PR, 1992. Growth and amylase production of Thermoascusaurantiacus Miehe. Biotechnology and Applied Biochemistry 15:311–313.
Alam M, Gomes I, Mohiuddin G, Hoq MM, 1994. Production andcharacterization of thermostable xylanases by Thermoascuslanuginosus and Thermoascus aurantiacus grown on lignocellu-loses. Enzyme and Microbial Technology 16: 298–302.
Ando S, Ishida H, Kosugi Y, Ishikawa K, 2002. Hyperthermostableendoglucanase from Pyrococcus horikoshi. Applied and Environ-mental Microbiology 68: 430–433.
Araujo A, Ward OP, 1990. Extracellular mannanases and galacta-nases from selected fungi. Journal of Industrial Microbiology andBiotechnology 6: 171–178.
Battan B, Sharma J, Dhimana SS, Kuhad RC, 2007. Enhancedproduction of cellulase-free thermostable xylanase by Bacilluspumilus ASH and its potential application in paper industry.Enzyme and Microbial Technology 41: 733–739.
Beg QK, Kapoor M, Mahajan L, Hoondal GS, 2001. Microbial xyla-nase and their applications: a review. Applied and Microbiologyand Biotechnology 56: 326–338.
Benko Z, Drahos E, Szengyel Z, Puranen T, Vehmaanpera J,Reczey K, 2007. Thermoascus aurantiacus CBHI/Cel7A produc-tion in Trichoderma reesei on alternative carbon sources. AppliedBiochemistry and Biotechnology 137–140: 195–204.
Bhat MK, 2000. Cellulases and related enzymes in biotechnology.Biotechnology Advances 18: 355–383.
Biely P, Mackenzie CR, Puls J, Schneider H, 1986. Cooperativity ofesterases and xylanases in the enzymatic degradation ofacetyl xylan. Biotechnology 4: 731–733.

128 M. Brienzo et al.
Biely P, VrSanskh M, Tenkanen M, Kluepfel D, 1997. Endo-P – 1,4-xylanase families: differences in catalytic properties. Journal ofBiotechnology 57: 151–166.
Birgisson H, Hreggvidsson GO, Fridjonsson OH, Mort A,Kristjansson JK, Mattiasson B, 2004. Two new thermostablea-L-rhamnosidases from a novel thermophilic bacterium.Enzyme and Microbial Technology 34: 561–571.
Borjesson J, Engqvist M, Sipos B, Tjerneld F, 2007. Effect ofpoly(ethylene glycol) on enzymatic hydrolysis and adsorptionof cellulase enzymes to pretreated lignocellulose. Enzyme andMicrobial Technology 41: 186–195.
Brummell DA, 2006. Cell wall disassembly in ripening fruit.Functional Plant Biology 33: 103–119.
Cardona CA, Sanchez OJ, 2007. Fuel ethanol production: processdesign trends and integration opportunities. Bioresource Tech-nology 98: 2415–2457.
Carvalho AFA, Goncalves AZ, Da Silva R, Gomes E, 2006. A specificshort dextrin-hydrolyzing extracellular glucosidase from thethermophilic fungus Thermoascus aurantiacus 179-5. The Journalof Microbiology 44: 276–283.
Cooney DG, Emerson R, 1964. Thermophilic Fungi. An account oftheir biology, activities and classification. The Quarterly Reviewof Biology 40: 292–293.
Coughlan MP, Hazlewood GP, 1993. b-1,4-D-Xylan-degradingsystems: biochemistry, molecular biology and applications.Biotechnology and Applied Biochemistry 17: 259–289.
Da Silva R, Lago EL, Merheb CW, Macchione MM, Park YK,Gomes E, 2005. Production of xylanase and CMCase on solidstate fermentation in different residues by Thermoascus aur-antiacus. Brazilian Journal of Microbiology 36: 235–241.
De Palma-Fernandez ER, Gomes E, da Silva R, 2002. Purificationand characterization of two beta-glucosidases from the ther-mophilic fungus Thermoascus aurantiacus. Folia Microbiologica47: 685–690.
De Vries RP, Kester HC, Poulsen CH, Benen JA, Visser J, 2000.Synergy between enzymes from Aspergillus involved indegradation of plant cell wall polysaccharides. CarbohydrateResearch 327: 401–410.
Deploey JJ, 1995. Some factors affecting the germination of Ther-moascus aurantiacus ascospores. Mycologia 87: 362–365.
Feldman KA, Lovett JS, Tsao GT, 1988. Isolation of the cellulaseenzymes from the thermophilic fungus Thermoascus aurantia-cus and regulation of enzyme production. Enzyme and MicrobialTechnology 10: 262–272.
Fogarty WM, Kelly CT, 1990. Recent advances in microbial anal-yses. In: Fogarty WM, Kelly CT (eds), Microbial Enzymes andBiotechnology. Elsevier Science Publishers, London, pp. 71–132.
Frandsen T, Svensson B, 1998. Plant a-glucosidases of theglycoside hydrolase family 31. Molecular properties,substrate specificity, resection mechanism and comparisonwith family members of different origins. Plant MolecularBiology 37: 1–13.
Gomes DJ, Gomes J, Steiner W, 1994. Factors influencing theinduction of endo-xylanase by Thermoascus aurantiacus. Journalof Biotechnology 33: 87–94.
Gomes DJ, Steiner W, 2000. Production and partial characheteri-zation of endo-mannanase from the thermophilic fungusThermoascus aurantiacus. Dhaka University Journal of BiologicalSciences. 9: 51–59.
Gomes I, Gomes J, Gomes DJ, Steiner W, 2000. Simultaneousproduction of high activities of thermostable endoglucanaseand b-glucosidase by the wild thermphilic fungus Ther-moascus aurantiacus. Applied Microbiology and Biotechnology 53:461–468.
Gomes J, Terler K, Kratzer R, Kainz E, Steiner W, 2007. Productionof thermostable b-mannosidase by a strain of Thermoascusaurantiacus: isolation, partial purification and characterizationof the enzyme. Enzyme and Microbial Technology 40: 969–975.
Gummadi SN, Panda T, 2003. Purification and biochemical prop-erties of microbial pectinases – a review. Process Biochemistry38: 987–996.
Guy DD, Carol MM, Pascal L, Kimberley NP, Robert MK, 1997.Purification and characterization of extremely thermostableb-Mannanase, b-Mannosidase, and a-Galactosidase from thehyperthermophilic Eubacterium Thermotoga neapolitana. Appliedand Environmental Microbiology 63: 169–177.
Guzman-Maldonado H, Peredes-Lopez O, 1995. Amylolyticenzymes and products derived from starch: a review. CriticalReview in Food Science and Nutrition 35: 373–403.
Haki GD, Rakshit SK, 2003. Developments in industrially impor-tant thermostable enzymes: a review. Bioresource Technology89: 17–34.
Himmel ME, Ruth MF, Wyman CE, 1999. Cellulase for commodityproducts from cellulosic biomass. Current Opinion in Biotech-nology 10: 358–363.
Hong J, Tamaki H, Kumagai H, 2006. Unusual hydrophobic linkerregion of b-glucosidase (BGLII) from Thermoascus aurantiacus isrequired for hyper-activation by organic solvents. AppliedMicrobiology and Biotechnology 73: 80–88.
Hong J, Tamaki H, Yamamoto K, Kumagai H, 2003a. Cloning ofa gene encoding thermostable cellobiohydrolase from Ther-moascus aurantiacus and its expression in yeast. Applied Micro-biology and Biotechnology 63: 42–50.
Hong J, Tamaki H, Yamamoto K, Kumagai H, 2003b. Cloning ofa gene encoding thermo-stable endo-b-1,4-glucanase fromThermoascus aurantiacus and its expression in yeast. Biotech-nology Letters 25: 657–661.
Jacques G, Frederic DL, Joste LB, Viviane B, Bart D, Fabrizio G,Benoit G, Jean-Marie F, 2000. An additional aromatic interac-tion improves the thermostability and thermophilicity ofa mesophilic family 11 xylanase: structural basis and molec-ular study. Protein Science 9: 466–475.
Jayani RS, Saxena S, Gupta R, 2005. Microbial pectinolyticenzymes: a review. Process Biochemistry 40: 2931–2944.
Kalogeris E, Christakopoulos P, Kekos D, Macris BJ, 1998. Studieson solid-state production of thermostable endoxylanasesfrom Thermoascus aurantiacus. Characterization of twoisozymes. Journal of Biotechnology 60: 155–163.
Kalogeris E, Christakopoulos P, Kekos D, Macris BJ, 1999. Mode ofaction of a minor xilanase from Thermoascus aurantiacus onpolysaccharides and model substrates. Journal of Bioscience andBioengineering 87: 819–821.
Kalogeris E, Christakopoulos P, Vrsanska M, Kekos D, Biely P,Macris BJ, 2001. Catalytic properties of the endoxylanase Ifrom Thermoascus aurantiacus. Journal of Molecular Catalysis B:Enzymatic 11: 491–501.
Kalogeris E, Christakopoulos P, Katapodis P, Alexiou A, Vlachou S,Kekos D, 2003. Production and characterization of cellulolyticenzymes from the thermophilic fungus Thermoascus aurantia-cus under solid state cultivation of agricultural wastes. ProcessBiochemistry 38: 1099–1104.
Kang SW, Park YS, Lee JS, Hong SI, Kim SW, 2004. Production ofcellulases and hemicellulases by Aspergillus niger KK2 fromlignocellulosic biomass. Bioresource Technology 91: 153–156.
Katapodis P, Christakopoulos P, 2004. Induction and partialcharacterization of intracellular b-fructofuranosidase fromThermoascus aurantiacus and its application in the synthesis of6-kestose. World Journal of Microbiology and Biotechnology 20:667–672.
Kawamori M, Takayama K, Takasawa S, 1987. Production ofcellulose by thermophilic fungus. Thermoascus aurantiacusA-131. Agricultural and Biological Chemistry 51: 647–654.
Khandke KM, Vithayathil PJ, Murthy SK, 1989a. Purification ofxylanase, b-glucosidase, endocellulase, and exocellulase froma thermophilic fungus, Thermoascus aurantiacus. Archives ofBiochemistry and Biophysics 274: 491–500.

Hydrolases of Thermoascus aurantiacus 129
Khandke KM, Vithayathil PJ, Murthy SK, 1989b. Purification andcharacterization of an a-D-glucuronidase from a thermophilicfungus, Thermoascus aurantiacus. Archives of Biochemistry andBiophysics 274: 511–517.
Ko HS, Fujiwara H, Yokoyama Y, Ohno N, Amachi S,Shinoyama H, Fujii T, 2005b. Inducible production of alcoholoxidase and catalase in a pectin medium by Thermoascus aur-antiacus IFO 31693. Journal of Bioscience and Bioengineering 99:290–292.
Ko HS, Yokoyama Y, Ohno N, Okadome M, Amachi S,Shinoyama H, Fujii T, 2005a. Purification and characterizationof intracellular and extracellular, thermostable and alkali-tolerant alcohol oxidases produced by a thermophilic fungus,Thermoascus aurantiacus NBRC 31693. Journal of Bioscience andBioengineering 99: 348–353.
Kongbuntad W, Saenphet K, Khanongnuch C, Lumyong S, 2006.Determination of toxic effects of crude xylanase derived fromThermoascus aurantiacus SL16W by hematology and bloodbiochemistry in albino rats. Pakistan Journal of Biological Sciences9: 2255–2260.
Krahe M, Antranikian G, Mairkl H, 1996. Fermentation of extrem-ophilic microorganisms. FEMS Microbiology Reviews 18: 271–285.
Leggio LL, Larsen S, 2002. The 1.62 a structure of Thermoascusaurantiacus endoglucanase: completing the structural pictureof subfamilies in glycoside hydrolaze family 5. FEBS Letters523: 103–108.
Leggio LL, Parry NJ, Van Beeumen J, Claeyssens M, Bhat MK,Pickersgill RW, 1997. Crystallization and preliminary X-rayanalysis of the major endoglucanase from Thermoascus aur-antiacus. Acta Crystallographica 53: 599–604.
Leite RSR, Gomes E, Silva R, 2007. Characterization and compar-ison of thermostability of purified b-glucosidases from a mes-ophilic Aureobasidium pullulans and a thermophilicThermoascus aurantiacus. Process Biochemistry 42: 1101–1106.
Li K, Azadic P, Collins R, Tolan J, Kim JS, Eriksson KL, 2000. Rela-tionships between activities of xylanases and xylan struc-tures. Enzyme and Microbial Technology 27: 89–94.
Maarel MJEC, van der Veen B, Uitehaag JCM, Leemhuis H,Dijkhuizen L, 2002. Preperties and applications of starch-con-verting enzymes of the a-amylase family. Journal of Biotech-nology 94: 137–155.
Maheshwari R, Bharadwaj G, Bhat MK, 2000. Thermophilic fungi:their physiology and enzymes. Microbiology and MolecularBiology Reviews 64: 461–488.
Martins ES, Silva D, Leite RSR, Gomes E, 2007. Purification andcharacterization of polygalacturonase produced by Thermoas-cus aurantiacus CBMAI-756 in submerged fermentation. Antonievan Leeuwenhoek 91: 291–299.
Martins, E.S., Silva, D., Da Silva, R., Gomes, E 2003. Perfil Enzi-matico produzido pelo Fungo Termofılico Thermoascusaurantiacus atraves de Fermentacao em Estado Solido eFracionamento de uma Poligalacturonase Termoestavel. XIVSimposio Nacional de Fermentacoes (SINAFERM),Florianopolis.
Martins ES, Silva D, Da Silva R, Gomes E, 2002. Solid stateproduction of thermostable pectinases from thermophilicThermoascus aurantiacus. Process Biochemistry 37: 949–954.
McCleary BV, Matheson NK, 1986. Enzymic analysis of polysac-charide Structure – review. Advances in Carbohydrate Chemistryand Biochemistry 44: 147–276.
Milagres AMF, Santos E, Piovan T, Roberto IC, 2004. Production ofxylanase by Thermoascus aurantiacus from sugar cane bagasse inan aerated growth fermentor. Process Biochemistry 39: 1387–1391.
Noack K, 1920. Der Betriebstoffwechsel der thermophile Pilze.Jahrbucher Fur Wissenschaitliche Botanik 59: 593–648.
Ohgren K, Bura R, Saddler J, Zacchi G, 2007. Effect of hemicellu-lose and lignin removal on enzymatic hydrolysis of steampretreated corn stover. Bioresource Technology 98: 2503–2510.
Ohno N, Fxjklida H, Wano H, Kasamijra M, Seidioyau H, Fum T,1998. Amylases produced by a thermophilic fungus, Ther-moascus aurantiacus, and some of their properties. Journal ofFermentation and Bioengineering 76: 3–4.
Oksanen T, Pere J, Paavilainen L, Buchert J, Viikari L, 2000. Treat-ment of recycled kraft pulps with Trichoderma reesei hemicellu-lases and cellulases. Journal of Biotechnology 78: 39–48.
Ortner J, Albert M, Terler K, Steiner W, Dax K, 2006. Transglyco-sylation reactions with a crude culture filtrate from Ther-moascus aurantiacus. Carbohydrate Research 327: 483–487.
Parry NJ, Beever DE, Owen E, Nerinckx W, Claaeyssens M,Beeumen JV, 2002. Biochemical characterization and mode ofaction of a thermostable endoglucanase purified from Thermoas-cus aurantiacus.ArchivesofBiochemistryandBiophysics404: 243–253.
Parry NJ, Beever DE, Owen E, Vandenberghe I, Van Beeumen J,Bhat MK, 2001. Biochemical characterization and mechanismof action of a thermostable b-glucosidase purified from Ther-moascus aurantiacus. Biochemical Journal 353: 117–127.
Polizeli MLTM, Rizzatti ACS, Monti R, Terenzi HF, Jorge JA,Amorim DS, 2005. Xylanases from fungi: properties andindustrial applications. Applied Microbiology and Biotechnology67: 577–591.
Puls J, Schmidt O, Granzow C, 1987. a-Glucuronidase in twomicrobial xylanolytic systems. Enzyme and Microbial Technology9: 83–88.
Rahkamo L, Viikari L, Buchert J, 1998. Enzymatic and alkalinetreatments of hardwood dissolving pulp. Cellulose 5: 79–88.
Roche N, Desgranges C, Durand A, 1994. Study on the solid-stateproduction of a thermostable alpha b-L-arabinofuranosidaseof Thermoascus aurantiacus on sugar beet pulp. Journal ofBiotechnology 38: 43–50.
Saha BC, 2000. a-L-Arabinofuranosidases: biochemistry, molec-ular biology and application in biotechnology. BiotechnologyAdvances 18: 403–423.
Santos E, Piovan T, Roberto IC, Milagres AMF, 2003. Kinetics of thesolid state fermentation of sugarcane bagasse by Thermoascusaurantiacus for the production of xylanase. Biotechnology Letters25: 13–16.
Sassner P, Martensson CG, Galbe M, Zacchi G, 2008. Steampretreatment of H2SO4-impregnated Salix for the productionof bioethanol. Bioresource Technology 99: 137–145.
Shallom D, Shoham Y, 2003. Microbial hemicellulases. CurrentOpinion in Microbiology 6: 219–228.
Souza MCO, Roberto IC, Milagres AMF, 1999. Solid state fermen-tation for xylanase production by Thermoascus aurantiacususing response surface methodology. Applied Microbiology andBiotechnology 52: 768–772.
Sreenath HK, Shah AB, Yang VW, Gharia MM, Jeffries TW, 1996.Enzymatic polishing of jute/cotton blended fabrics. Journal ofFermentation and Bioengineering 81: 18–20.
Stoilova IS, Gargova SA, Krastanov AI, 2005. Production ofenzymes by culture from mycelial fungi in solid-statefermentation. Biotechnology and Bioengineering 19: 103–108.
Tenkanen M, Puls J, Poutanen K, 1992. Two major xylanase ofTrichoderma reesei. Enzyme and Microbial Technology 14: 566–574.
Tong CC, Cole AL, Shepherd MG, 1980. Purification and propertiesof the cellulases from the thermophilic fungus Thermoascusaurantiacus. Biochemical Journal 191: 83–94.
Turner P, Mamo G, Karlsson EN, 2007. Review: potential andutilization of thermophiles and thermostable enzymes inbiorefining. Microbial Cell Factories 6: 1–23.
Upahyay JM, Farmelo MS, Goetz SG, Melan MA, 1984. A newvariety of a thermophilic mold, Thermoascus aurantiacus var.levisporus. Mycopathologia 87: 71–80.
Vardakou M, Katapodis P, Samiotaki M, Kekos D, Panayotou G,Christakopoulos P, 2003. Mode of action of family 10 and 11endoxylanases on water-unextractable arabinoxylan. Interna-tional Journal of Biological Macromolecules 33: 129–134.

130 M. Brienzo et al.
Viikari L, Tenkanen M, Buchert J, Ratto M, Bailey M, Siika-Aho M, Linko M, 1993. Hemicellulases for industrial appli-cations. In: Saddler JN (ed), Bioconversion of Forest and Agri-cultural Plant Residues. CAB International, Wallingford, UK,pp. 131–182.
Viswamitra MA, Bhanumoorthy P, Ramakumar S, Manjula MV,Vithayathil PJ, Murthy SK, Naren AP, 1993. Crystallization andpreliminary x-ray diffraction analysis of crystals of Thermoascusaurantiacus xylanase. Journal of Molecular Biology 232: 987–988.
Wang H, Tokusige Y, Shinoyama H, Fujii H, Urakam T, 1998.Purification and characterization of a thermostable catalasefrom culture broth of Thermoascus aurantiacus. Journal ofFermentation and Bioengineering 85: 169–173.
Ward OP, Moo-Young M, 1989. Enzymatic degradation of cell-walland related plant polysaccharides. Critical Reviews in Biotech-nology 8: 237–274.
Willats WGT, Mccartney L, Mackie W, Knox JP, 2001. Pectin: cellbiology and prospects for functional analysis. Plant MolecularBiology 47: 9–27.
Wong KKY, Saddler JN, 1992. Trichoderma xylanases, their prop-erties and application. In: Visser J, Beldman G, Kuster-vanSomeren MA, Voragen AGJ (eds), Xylans and Xylanases. Prog-ress in Biotechnology 7: 171–186, Elsevier: Amsterdam, TheNetherlands.
Wong KKY, Saddler JN, 1993. Applications of hemicellulases inthe food, feed, and pulp and paper industries. In:Coughlan MP, Hazlewood GP (eds), Hemicellulose and Hemicel-lulases. Portland Press, London, pp. 127–143.
Yu EKC, Tan LUL, Chan MKH, Deschatelets L, Saddler JN, 1987.Production of thermostable xylanase by a thermophilicfungus, Thermoascus aurantiacus. Enzyme and Microbial Tech-nology 9: 16–24.
Yuan Q, Wang J, Zhang H, Qian Z, 2005. Effect of temperature shifton production of xylanase by Aspergillus niger. ProcessBiochemistry 40: 3255–3257.
Zhu L, O’dwyer JP, Chang VS, Granda CB, Holtzapple MT, 2008.Structural features affecting biomass enzymatic digestibility.Bioresource Technology 99: 3817–3828.




















