
Microbial characteristics of methanogenic sludge consortia developed in thermophilic U ASB reactors
7
Appl Microbiol Biotechnol (1993) 39:654-660 © Springer-Verlag 1993 Microbial characteristics of methanogenic sludge consortia developed in thermophUic UASB reactors Shigeki Uemura ~, Hideki Harada 2 1 Research Division, Komatsu Ltd., 1200, Manda, Hiratsuka-shi, Kanagawa 254, Japan 2 Department of Civil Engineering, Nagaoka University of Technology, 1603-1, Kamitomioka, Nagaoka-shi, Niigata 940-21, Japan Received: 18 September 1992/Accepted: 8 March 1993 Abstract. The effect of temperature on granulation and microbial interaction of anaerobic sludges grown in thermophilic upflow anaerobic sludge bed (UASB) reactors was investigated at two different tempera- tures, 55°C (Run 1) and 65°C (Run 2). Each run con- sisted of two phases. Phase 1 was conducted by feeding acetate for a period of 200 days. In Phase 2, both reac- tors were fed a mixture of acetate and sucrose for a further 100 days. During Phase 1, no granulation oc- curred in the sludge of either run. Microscopic obser- vation revealed that the predominant methanogen was Methanothrix in Run 1, whereas Methanobacterium- like bacteria existed to a significant extent in Run 2. The acetate-utilizing methanogenic activity of both sludges increased with increasing test temperature in the range 55-65°C. Since the acetate-grown sludges ex- hibited far higher H2-utilizing methanogenic activity than acetate-utilizing methanogenic activity, it is sug- gested that a syntrophic association of acetate-oxidiz- ing bacteria with hydrogenotrophic methanogens was responsible for a considerable portion of the overall acetate elimination in thermophilic anaerobic sludge. During Phase 2, granules coated with either fil- amentous bacteria or cocci-type bacteria (both pre- sttmably acid-forming bacteria) were successfully es- tablished in Run 1 and Run 2, respectively. Since the acetate-utilizing methanogenic activities of the granu- lar sludges were four to five times higher than those of the acetate-grown sludges (Phase 1), the co-existence of these "coating bacteria" appeared to contribute to the enclosing of acetate consumers inside granules. Introduction In recent years a number of upflow anaerobic sludge bed (UASB) reactors have been increasingly used for the anaerobic treatment of a variety of industrial Correspondence to: S. Uemura waste,waters. However, most of the UASB reactors designed and constructed so far have adopted the me- sophilic process, and there are at present very few ther- mophilic UASB reactors of a practical scale in opera- tion. This lack of popularity of the thermophilic UASB process may be attributed to the difficulty in accom- plishing granulation of thermophilic anaerobes, com- pared with the mesophilic process. It is well known that granules that settle well are commonly composed of long or short filaments of mul- ticellular-rod organisms, presumably Methanothrix (Hulshoff Pol et al. 1983). On the other hand, when Methanosarcina is the predominant methanogen as an acetate-utilizing methane-producing bacteria (A-MPB) in a sludge consortium, it is subject to washout from the reactor, and leads to process failure (Endo 1986; Weigant and de Man 1986). Therefore, it should be re- membered that the predominance of Methanothrix is essential for granulation in mesophilic as well as ther- mophilic UASB processes. However, some researchers tried but failed to make Methanothrix predominate in their thermophilic anae- robic high-rate reactors, i.e., not only UASB reactors but also anaerobic fixed-bed reactors. The superiority of therrnophilic treatment over mesophilic treatment was not established in their experiments due to a pre- dominance of Methanosarcina (Endo 1986; Kennedy and van den Berg 1982). Their results suggested that Methanosarcina tended to grow more preferentially in thermophilic anaerobic digestion. Therefore, the pre- ferential propagation of Methanothrix in a competition between these two A-MPB may be a key factor for suc- cessful performance of a thermophilic UASB reactor. According to the results of thermophilic acetoclastic methanogens reported so far, the optimum tempera- ture for Methanosarcina ranges from 50°C to 58°C (Zinder and Mah 1979; Olivier et al. 1984), whereas that for Methanothrix lies in the rather high ranges be- tween 55°C and 65°C (Zinder et al. 1987; Kamagata and Mikami 1991; Nozhevnikova and Chudina 1985). Furthermore, thermophilic Methanosarcina tends to drastically lose its activity at around 65°C, at which
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Microbial characteristics of methanogenic sludge consortia developed in thermophilic U ASB reactors

Appl Microbiol Biotechnol (1993) 39:654-660
© Springer-Verlag 1993
Microbial characteristics of methanogenic sludge consortia developed in thermophUic UASB reactors Shigeki Uemura ~, Hideki Harada 2
1 Research Division, Komatsu Ltd., 1200, Manda, Hiratsuka-shi, Kanagawa 254, Japan 2 Department of Civil Engineering, Nagaoka University of Technology, 1603-1, Kamitomioka, Nagaoka-shi, Niigata 940-21, Japan
Received: 18 September 1992/Accepted: 8 March 1993
Abstract. The effect of temperature on granulation and microbial interaction of anaerobic sludges grown in thermophilic upflow anaerobic sludge bed (UASB) reactors was investigated at two different tempera- tures, 55°C (Run 1) and 65°C (Run 2). Each run con- sisted of two phases. Phase 1 was conducted by feeding acetate for a period of 200 days. In Phase 2, both reac- tors were fed a mixture of acetate and sucrose for a further 100 days. During Phase 1, no granulation oc- curred in the sludge of either run. Microscopic obser- vation revealed that the predominant methanogen was Methanothrix in Run 1, whereas Methanobacterium- like bacteria existed to a significant extent in Run 2. The acetate-utilizing methanogenic activity of both sludges increased with increasing test temperature in the range 55-65°C. Since the acetate-grown sludges ex- hibited far higher H2-utilizing methanogenic activity than acetate-utilizing methanogenic activity, it is sug- gested that a syntrophic association of acetate-oxidiz- ing bacteria with hydrogenotrophic methanogens was responsible for a considerable portion of the overall acetate elimination in thermophilic anaerobic sludge. During Phase 2, granules coated with either fil- amentous bacteria or cocci-type bacteria (both pre- sttmably acid-forming bacteria) were successfully es- tablished in Run 1 and Run 2, respectively. Since the acetate-utilizing methanogenic activities of the granu- lar sludges were four to five times higher than those of the acetate-grown sludges (Phase 1), the co-existence of these "coating bacteria" appeared to contribute to the enclosing of acetate consumers inside granules.
Introduction
In recent years a number of upflow anaerobic sludge bed (UASB) reactors have been increasingly used for the anaerobic treatment of a variety of industrial
Correspondence to: S. Uemura
waste,waters. However, most of the UASB reactors designed and constructed so far have adopted the me- sophilic process, and there are at present very few ther- mophilic UASB reactors of a practical scale in opera- tion. This lack of popularity of the thermophilic UASB process may be attributed to the difficulty in accom- plishing granulation of thermophilic anaerobes, com- pared with the mesophilic process.
It is well known that granules that settle well are commonly composed of long or short filaments of mul- ticellular-rod organisms, presumably Methanothrix (Hulshoff Pol et al. 1983). On the other hand, when Methanosarcina is the predominant methanogen as an acetate-utilizing methane-producing bacteria (A-MPB) in a sludge consortium, it is subject to washout from the reactor, and leads to process failure (Endo 1986; Weigant and de Man 1986). Therefore, it should be re- membered that the predominance of Methanothrix is essential for granulation in mesophilic as well as ther- mophilic UASB processes.
However, some researchers tried but failed to make Methanothrix predominate in their thermophilic anae- robic high-rate reactors, i.e., not only UASB reactors but also anaerobic fixed-bed reactors. The superiority of therrnophilic treatment over mesophilic treatment was not established in their experiments due to a pre- dominance of Methanosarcina (Endo 1986; Kennedy and van den Berg 1982). Their results suggested that Methanosarcina tended to grow more preferentially in thermophilic anaerobic digestion. Therefore, the pre- ferential propagation of Methanothrix in a competition between these two A-MPB may be a key factor for suc- cessful performance of a thermophilic UASB reactor.
According to the results of thermophilic acetoclastic methanogens reported so far, the optimum tempera- ture for Methanosarcina ranges from 50°C to 58°C (Zinder and Mah 1979; Olivier et al. 1984), whereas that for Methanothrix lies in the rather high ranges be- tween 55°C and 65°C (Zinder et al. 1987; Kamagata and Mikami 1991; Nozhevnikova and Chudina 1985). Furthermore, thermophilic Methanosarcina tends to drastically lose its activity at around 65°C, at which

655
thermophil ic Methanothrix still maintains its activity sufficiently (Zinder 1988). Based on this background, we conducted a compara t ive exper iment of thermo- philic U A S B process at 55°C and at 65°C, in order to investigate the effect of cultivation t empera tu re on the microbial-ecological behaviour of thermophil ic sludges. This study also focused on a syntrophic asso- ciation of acetate-oxidizing bacter ia (AOB) with hy- drogen-utilizing methane-producing bacter ia (H- MPB), which is supposed to be significant under ther- mophil ic conditions (Zinder and Koch 1984), and with respect to the role of thermophil ic acidogens in granule formation.
Materials and methods
UASB reactor. Two UASB reactors with a 5.5 1 working volume were used (Fig. 1). Each reactor was equipped with a water-jack- et around the column, in which hot water was cycled to keep the inner temperatures at 55°C (Run 1) and at 65°C (Run 2). Five litres of mesophilic digested sewage sludge containing 23.8 g vola- tile suspended solids (VSS) and 57.7 g suspended solids were seeded in each reactor.
Media. The Complete experiments of Run i and Run 2 consisted of two phases. During the first phase (as Phase 1, days 1-200), 3000mg chemical oxygen demand (COD)-1-1 of acetate was used as a major organic component of the influent. In the second phase (Phase 2), the reactors were operated by feeding a mixture of acetate and sucrose (80% :20% as COD) with a strength of 3000 mg COD .1-2. Yeast extract at 100 mg COD .1-1 was added to the influent for both of the phases. An addition of inorganic salts and trace elements was made according to Wiegant and Let- tinga (1985).
Methanogenic activity test. The methanogenic activities were de- termined with serum vial bottles with a volume of 120 ml. For the acetate-utilizing methanogenic activity (A-MA) test, a 30 ml sludge suspension was cultivated in advance overnight in the vial to eliminate external substrate. And then the basal salts, reducing
12 I / -10
l , IIII
" ~ 1 ' 1 | 1''4 "A L ~ 6
Fig. 1. Experimental set-up of the upflow anaerobic sludge blan- ket (UASB) reactor 1, substrate tank; 2, water coder; 3, feed pump; 4, heater; 5, recirculation pump; 6, influent; 7, water jack- et; 8, temperature sensor; 9, perforated cones; 10, effluent; 11, gas-solid separator; 12, temperature monitor; 13, water seal; 14, wet-test gas meter
agent (NazS" 9H20, 200 mg. 1-1 as a vial final concentration), and resazurin (1 mg.1-1) were added to the vials. A 5 ml concen- trated acetate solution was subsequently added to the vials (3000 mg COD .1-1). The vials were then placed in water baths kept at 50°C, 60°C and 65°C. Methane production was estimated by frequent measurement of gas production and gas composition at regular intervals.
For determination of Hz-utilizing methanogenic activity (H- MA), 20 ml sludge mixture was added to the vials, of which the headspace was filled with Hz/CO2 (80% :20%, v/v) gas mixture.
Hydrogen production test (BES test). The BES test procedure was, in principle, the same as that of the A-MA test, except for the addition of 100 mM 2-bromoethanesulphonate (2-BES) as a methanogenic inhibitor. Taking into account that even a small amount of H2 accumulation causes an adverse effect on acetate conversion to H2, the headspace gas was replaced with N2 gas by passing it through a syringe to the vial bottles at once after each gas sampling. The test was conducted at 65°C.
Analyses. The total organic carbon (TOC) was determined by a TOC analyser (Shimadzu TOC-500). The contents of methane and hydrogen in biogas were determined by a thermal conductiv- ity detector (TCD) gas chromatograph (Shimadzu GC-14A), equipped with a 3 m × 2 mm (ID) stainless-steel column with Mo- lecular Sieve 5A (80/100 mesh). Argon gas was used as a carrier gas at a column pressure of 1 kg/cm 2. The column and detector were kept at 40°C and 70°C, respectively.
Microscopy. Samples for scanning electron microscope (SEM) observation were fixed with 2.5% glutaraldehyde in 0.1 M caco- dylate buffer for 2h. After post-fixation with 2% of osmic acid for i h, samples were dehydrated in a graded acetone series and replaced with n-pentyl acetate, then dried with a critical-point drier. Observations were made with an Hitachi-S800 SEM oper- ating at an accelerating voltage of 15 kV.
Results and discussion
Acetate-feeding experiments (Phase 1)
The volumetr ic loading rates of both runs were in- creased stepwise by shortening hydraulic re tent ion t ime ( H R T ) when the effluent total organic carbon (TOC) level at tained less than 100 m g . l -a. For Run 1 (at 55°C), the volumetr ic T O C loading rate reached 12.1 kg T O C . m - 3 , d a y -1 on the 127th day with 90.6% T O C removal , but afterwards a severe washout of sludge occurred and was followed by process failure. Despi te subsequent reduction of the loading rate in Run 1 to 5-6 kg T O C . m - 3 . d a y -1, there was no sign of the recovery of process performance . The process per formance of Run 2 (65°C) was not be t ter than that of Run 1 throughout Phase 1, as opposed to our expec- tation. The T O C loading ra te reached in Run 2 was no greater than 2-4 kg T O C . m - 3 . d a y -~, and the T O C removal efficiency hovered be tween 50% and 80% (data not shown). Dur ing Phase 1 of bo th runs, no dis- tinctive granule format ion took place due to the con- tinuous sludge washout.
A significant extent of rod- type bacter ia with a length of 2 i~m was present in bo th effluents. Judging f rom the presence of characteristic gas vacuoles inside the cells (Zinder e t al. 1987; Kamaga ta and Mikami 1991; Nozhevnikova and Chudina 1984) and peculiar

656
2a, b. Scanning electron mi- crographs of thermophilic sludge grown at 55°C (Run 1: a) and at 65°C (Run 2: b), re- spectively, during Phase 1: bar, 3 ixm
cell morphology, this bacterium can be identified as a thermophilic Methanothrix (data not shown).
Figure 2a presents a scanning electron micrograph of the sludge retained in Run i reactor. No presence of Methanosarcina was confirmed in the sludge of either run by routine SEM observation. On the other hand, the prevalence of Methanothrix-like bacteria in the form of dispersed growth was observed in both runs, not as often found in the mesophilic granules, which usually contain long filaments. Kamagata and Mikami (1990) reported that there are two morphotypes of Me- thanothrix. One is the "aggregating type" with a ten- dency to form a cotton-like conglomeration by entan- glement of long filaments, and the other is the "dis- persing type", which tends to form short chains com- posed of a few cells. In our 55°C cultivation, the latter "dispersing" type of Methanothrix proliferated even though the propagation of Methanosarcina was suc- cessfully suppressed. As a consequence, the sludges were susceptible to washout because granules that set- tled well did not form, so the process performance was not able to attain a satisfactory level. Figure 2b shows the SEM observation of the sludge cultivated at 65°C (Run 2). Methanothrix existed to a lesser extent, and instead, a bacterium resembling Methanobacterium thermoautotrophicum appeared in abundance. As in
Table 1. Methanogenic activities of the thermophilic sludges grown on acetate
A-MA H-MA
T (° C) 55 60 65 60
Run 1 0.53 (0.03) 0.54 (0.01) 0.68 (0.01) 1.62 (0.05) Run 2 0.10 (0.01) 0.16 (0.01) 0.18 (0.01) 1.55 (0.09)
Figures in parentheses represent standard deviation of duplicate or triplicate activity determinations: A-MA, acetate-utilizing me- thanogenic activity (kg CH4-COD.kg VSS -l . day -1); H-MA, hydrogen-utilizing methanogenic activity (kg CH4-COD.kg VSS-1. day-l); VSS, volatile suspended solids
Run 1, Methanosarcina was not observed at all throughout the Run 2 experiment.
Table 1 presents the methanogenic activities of sludges taken from both reactors on the 185th day. The A-MAs of the sludge in both runs showed a tendency to increase slightly but consistently with increasing ac- tivity-test temperature. An interesting finding in Table 1 is that the H-MA was significantly greater than A- MA even though the sludge was cultivated on acetate solely for a long period (almost for 6 months). It is no- teworthy that at 60°C the H-MA of Run 1 was three
Table 2. Methanogenic activities of the acetate-grown anaerobic sludges
T H-MA A-MA H-MA a (°C)
A-MA
Sludge type Reference
Mesophilic 30 0.18 1.16 0.16 35 0.11 2.10 0.05
Thermophilic 55 0.24 1.16 0.21 60 1.62 0.54 3.0 65 1.55 0.16 9.7
Granular Granular Granular Non-granular Non-granular
Dolfing and Mulder (1985) Harada et al. (1988) Schmidt and Ahring (1990) This study (Run 1) This study (Run 2)
a The ratio of H-MA to A-MA
657
80
40
2O
'-6 E o
(D -20
-Z,O
-60
-80
-1 -2 -3 -& -5 -6 -7 tog (pH2)
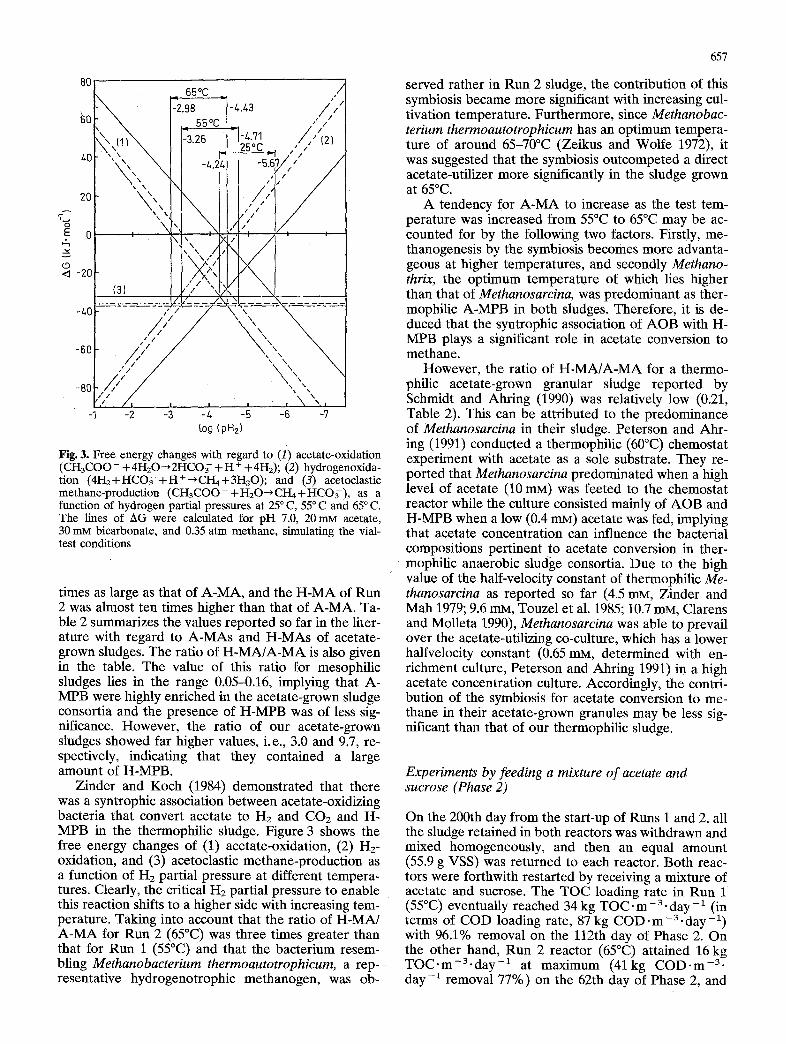
Fig. 3. Free energy changes with regard to (1) acetate-oxidation (CH3COO - + 41-{20 ~2HCO ;- + H + + 4H2); (2) hydrogenoxida- tion (4H2+HCO~-+H+~CI-I4+3H20); and (3) acetoclastic methane-production (CH3COO-+H20~CH4+HCO;-), as a function of hydrogen partial pressures at 25 ° C, 55°C and 65 ° C. The lines of AG were calculated for pH 7.0, 20 mM acetate, 30 mM bicarbonate, and 0.35 atm methane, simulating the vial- test conditions
times as large as that of A-MA, and the H-MA of Run 2 was almost ten times higher than that of A-MA. Ta- ble 2 summarizes the values reported so far in the liter- ature with regard to A-MAs and H-MAs of acetate- grown sludges. The ratio of H-MA/A-MA is also given in the table. The value of this ratio for mesophilic sludges lies in the range 0.05-0.16, implying that A- MPB were highly enriched in the acetate-grown sludge consortia and the presence of H-MPB was of less sig- nificance. However, the ratio of our acetate-grown sludges showed far higher values, i.e., 3.0 and 9.7, re- spectively, indicating that they contained a large amount of H-MPB.
Zinder and Koch (1984) demonstrated that there was a syntrophic association between acetate-oxidizing bacteria that convert acetate to H2 and CO2 and H- MPB in the thermophilic sludge. Figure 3 shows the free energy changes of (1) acetate-oxidation, (2) Hz- oxidation, and (3) acetoclastic methane-production as a function of H2 partial pressure at different tempera- tures. Clearly, the critical H2 partial pressure to enable this reaction shifts to a higher side with increasing tem- perature. Taking into account that the ratio of H-MA/ A-MA for Run 2 (65°C) was three times greater than that for Run 1 (55°C) and that the bacterium resem- bling Methanobacterium thermoautotrophicum, a rep- resentative hydrogenotrophic methanogen, was ob-
served rather in Run 2 sludge, the contribution of this symbiosis became more significant with increasing cul- tivation temperature. Furthermore, since Methanobac- terium thermoautotrophicum has an optimum tempera- ture of around 65-70°C (Zeikus and Wolfe 1972), it was suggested that the symbiosis outcompeted a direct acetate-utilizer more significantly in the sludge grown at 65°C.
A tendency for A-MA to increase as the test tem- perature was increased from 55°C to 65°C may be ac- counted for by the following two factors. Firstly, me- thanogenesis by the symbiosis becomes more advanta- geous at higher temperatures, and secondly Methano- thrix, the optimum temperature of which lies higher than that of Methanosarcina, was predominant as ther- mophilic A-MPB in both sludges. Therefore, it is de- duced that the syntrophic association of AOB with H- MPB plays a significant role in acetate conversion to methane.
However, the ratio of H-MA/A-MA for a thermo- philic acetate-grown granular sludge reported by Schmidt and Ahring (1990) was relatively low (0.21, Table 2). This can be attributed to the predominance of Methanosarcina in their sludge. Peterson and Ahr - ing (1991) conducted a thermophilic (60°C) chemostat experiment with acetate as a sole substrate. They re- ported that Methanosarcina predominated when a high level of acetate (10 raM) was feeted to the chemostat reactor while the culture consisted mainly of AOB and H-MPB when a low (0.4 mM) acetate was fed, implying that acetate concentration can influence the bacterial compositions pertinent to acetate conversion in ther- mophilic anaerobic sludge consortia. Due to the high value of the half-velocity constant of thermophilic Me- thanosarcina as reported so far (4.5 raM, Zinder and Mah 1979; 9.6 mM, Touzel et al. 1985; 10.7 mM, Clarens and Molleta 1990), Methanosarcina was able to prevail over the acetate-utilizing co-culture, which has a lower halfvelocity constant (0.65 mM, determined with en- richment culture, Peterson and Ahring 1991) in a high acetate concentration culture. Accordingly, the contri- bution of the symbiosis for acetate conversion to me- thane in their acetate-grown granules may be less sig- nificant than that of our thermophilic sludge.
Experiments by feeding a mixture of acetate and sucrose (Phase 2)
On the 200th day from the start-up of Runs i and 2, all the sludge retained in both reactors was withdrawn and mixed homogeneously, and then an equal amount (55.9 g VSS) was returned to each reactor. Both reac- tors were forthwith restarted by receiving a mixture of acetate and sucrose. The TOC loading rate in Run 1 (55°C) eventually reached 34 kg TOC. m-3 "day-1 (in terms of COD loading rate, 87 kg C O D . m - 3 - d a y -1) with 96.1% removal on the 112th day of Phase 2. On the other hand, Run 2 reactor (65°C) attained 16 kg T O C ' m - 3 " d a y -1 at maximum (41kg C O D . m -3. day -1 removal 77%) on the 62th day of Phase 2, and

658
Fig. 4a, b. Scanning electron micrographs of granular sludge grown on a mixture of acetate and sucrose (Phase 2) at 55°C (Run 1). a Whole view: bar 300 ~rn. b Cross- sectional view: bar 3 ~m
Fig. 5a, b. Scanning electron micrographs of granule sur- faces, grown on a mixture of acetate and sucrose (Phase 2) at 55°C (Run 1: a) and 65°C (Run 2: b): bar, 3 p~m. Note a copious amount of extracellu- lar biopolymer indicated by the arrows
then the performance worsened gradually (data not shown). In both runs, the sludges began to form gran- ules between 25 and 35 days after the restart-up of Phase 2, and complete granular beds were established on around the 60th day. The granules obtained from both reactors ranged from 0.5 mm to 1.0 mm in diam- eter, and those from Run 2 (65°C) tended to be more fragile and slightly smaller than those from Run 1 (55°C). Thus, due to the rapid granulation in both reac- tors, very high loading rates comparable to the results reported previously (for example, on acetate, 129 kg COD.m-3-day -1 with removals of 88%, at 55°C, Wiegant 1986) could be established.
Figure 4a and b represent the whole view and the cross-sectional view of a granule grown in Run 1. Me- thanothrix in a state of dispersed growth predominated
throughout the granule, and another type of rod with a length of 1.5-2 ~m was also observed. Unfortunately, granules taken from Run 2 were so friable that the in- side could not be examined by SEM.
Figure 5a and b show SEM observations of micro- bial populations present on the surfaces of granules in Run ! and Run 2, respectively. In the granules from Run 1 (55°C), long and thin filamentous bacteria, which were apparently different from Methanothrix , prevailed over the surfaces by intertwining with each other to form a web-like coating (Fig. 5a). The surfaces of granules from Run 2 (65°C) abounded with cocci- type bacteria that were excreting a copious amount of extracellular biopolymer, as observed in Fig. 5b. The coating of those bacteria on the surfaces of granules from both runs appears to be somehow responsible for

659
Table 3. Acetate-utilizing methanogenic activity (A-MA) of gran- ules grown on a mixture of acetate and sucrose
A-MA
T (° C) 55 60 65
Run 1 1.29 (0.01) 2.00 (0.16) 2.71 (0.05) Run 2 0.76 (0.01) 0.90 (0.04) 0.90 (0.01)
Figures in parentheses represent standard deviation of duplicate or triplicate activity determinations
granule formation. The total amount of biomass pres- ent in the reactors was determined at the end of the experiments. The Run 1 reactor contained twice as much biomass as the Run 2 reactor (124 g VSS in Run 1 and 68 g VSS in Run 2). The proliferation of the former filamentous-type coating bacteria seemed to be more favorable for proper granulation than that of the latter coating bacteria, and consequently led to the achievement of a high loading rate in Run 1. Since both types of coating bacteria appeared in the sludge consortia after starting to feed sucrose to the reactors, they were presumably acid-forming bacteria. A similar phenomenon - a propagation of filamentous bacteria thoroughly coating granule surfaces - was also ob- served in another type of granulation process, i.e., in a multi-staged reverse-flow bioreactor (MRB; Takahashi and Kyousai 1988) for sewage treatment.
Table 3 represents the A-MA of both sludges on the 87th day of Phase 2. As in Phase 1 (Table 1), the A- MA had a tendency to increase with increasing test temperature. Compared with one at 65°C in Phase 1, the A-MA increased four times in Run i and five times in Run 2. This increases suggests that the coating bac- teria were capable of efficiently enclosing acetate-con- suming bacteria, i.e., Methanothrix and the syntrophs of AOB with H-MPB, within a granule.
Test of hydrogen formation from acetate
Since our acetate-grown sludges exhibited high H-MA relative to A-MA, a serum-vial test was conducted to verify the capability of sludge for producing H2 from acetate. The test was carried out using the sludge har- vested from the Run i reactor at the end of Phase 2. A supplementary vial without addition of acetate was also shaken as a reference, but gas production was very small compared with that of the vial with acetate. Dur- ing 22h of cultivation, 2.6 ml 1-12 accumulated in the acetate-added vial. Even though only a trace of Hz ac- cumulated in the headspace (because the test was con- ducted under conditions that were thermodynamically unfavorable for acetate conversion to H2 on account of the addition of 2-BES), the result validated the exis- tence of the bacterium that was able to perform acetate conversion to H2.
Conclus ion
The following conclusions can be drawn from this study:
1. 'It demonstrated that in thermophilic anaerobic process, there are two pathways for methane produc- tion from acetate. A part of the acetate eliminated was directly converted to methane by acetoclasfic metha- nogens, and the remainder was firstly converted to H2 and CO2, and subsequently to methane by hydrogeno- trophic methanogens. With increasing cultivation tem- perature, the contribution of the latter pathway be- came more significant.
2. The acetate-utilizing methanogenic activity of the sludge cultivated at 55°C and at 65°C increased with increasing test temperature in the range 55-65°C. It is suggested that the optimum temperature for methane production from acetate was dependent not merely on cultivation temperature but also on the microbial pop- ulation composition regarding acetate degradation in- volving acetotrophic methanogens as well as a symbio- sis between acetate-oxidizing (hydrogen-producing) bacteria and hydrogenotrophic methanogens.
3. When fed an acetate solution (Phase 1), no gran- ulation occurred either at 55°C or at 65°C over 200 days of operation. However, after starting to feed a mixture of acetate and sucrose, complete granular sludge beds were established in both runs within 2 months. Accordingly, the loading rates reached 34 kg TOC" m -3- day - 1 at 55°C, and 16 kg TOC. m-3. day - ~ at 65°C. Two types of acid-forming bacteria prevailed all over the granule surfaces. The long-filamentous type at 55°C had a stronger mechanical firmness, and led to higher sludge retainment. The co-existence of the former type seems to be preferable for granulation.
References
Clarens M, Moletta R (1990) Kinetic studies of acetate fermenta- tion by Methanosarcina sp. MSTA-1. Appl Microbiol Biotech 33: 239-244
Dolfing J, Mulder JW (1985) Comparison of methane production rate and coenzyme F420 content of methanogenic consortia in anaerobic granular sludge. Appl Environ Microbiol 49:1142- 1145
Endo G (1986) Process performance of upflow anaerobic sludge blanket reactors and formation of granular sludge by metha- nogens (in Japanese). Hakkokogaku kaishi 64:207--210
Harada H, Endo G, Toya Y, Momonoi K (1988) High rate per- formance and its related characteristics of granulated sludges in UASB reactors treating various wastewaters. In: Tiche A, Rozzi A (ed) Proceedings of the 5th International Symposium on Anaerobic Digestion, Bologna, Italy, 22-26 May, 1988. Monduzzi Editore S.p.A., Bologna; Italy, pp 1011-1020
Hulshoff Pol LW, Zeeuw WJ de, Velzeboer CTM, Lettinga G (1983) Granulation in UASB reactors. Water Sci Technol 15:291-304
Kamagata Y, Mikami E (1990) Some characteristics of two mor- photypes of Methanothrix soehngenii from mesophilic anae- robic digester. J Ferment Bioeng 70:272-274
Kamagata Y, Mikami E (1991) Isolation and characterization of a novel thermophilic Methanosaeta strain. Int J Syst Bacteriol 41 : 191-196

660
Kennedy KJ, Berg L van den (1982) Thermophilic downflow sta- tionary fixed reactors for methane production from bean blanching waste. Biotechnol Lett 4:171-176
Nozhevnikova AN, Chudina I (1984) Morphology of the thermo- philic acetate methane bacterium Methanothrix thermoaceto- phila sp nov Microbiology 53:618--624
Olivier B, Lombardo A, Albagnac G (1984) Isolation and charac- terization of a new thermophilie Methanosarcina strain (strain MP). Ann Microbiol (Inst Pasteur) 135 B:187-198
Peterson SP, Ahring BK (1991) Acetate-oxidation in a thermo- philic anaerobic sewage-sludge digester: the importance of non-acetoclastic methanogenesis from acetate. FEMS Micro- biol Ecol 86:149-158
Schmidt JE, Ahring BK (1990) Acetate- and hydrogen metabol- ism in intact and distintegrated granules from an acetate-fed, 55°C, UASB reactor. Appl Microbiol Biotechnol 35:681-685
Takahashi M, Kyousai S (1988) Development of multi-stage re- versing-flow bioreactor (MRB) for wastewater treatment. Water Sci Technol 20:361-367
Touzel JP, Petroff D, Albagnac G (1985) Isolation and character- ization of a new thermophilic Methanosarcina, the strain CHTI 55. Syst Appl Microbiol 6:66-71
Wiegant WM (1986) Thermophilic anaerobic wastewater treat- ment. Proceedings of the NVA/EWPCA conference on Anae-
robic Treatment - a Grown-Up Technology, 15-19 Septem- ber, 1986, Amsterdam, Padoc, Wageningen, The Netherlands. pp 115-127
Wiegant WM, Lettinga G (1985) Thermophilic anaerobic diges- tion of sugars in upflow anaerobic sludge blanket reactors. Biotechnol Bioeng 27:1603--1607
Wiegant WM, Man AW de (1986) Granulation of biomass in thermoplfilic upflow anaerobic sludge blanket reactors treat- ing acidified wastewaters. Biotechnol Bioeng 28:718-727
Zeikus JG, Wolfe RS (1972) Methanobacterium thermoautotro- phicus sp n, an anaerobic autotrophic extreme thermophile. J Bacteriol 109:707-713
Zinder SH (1988) Conversion of acetic acids to methane by ther- mophiles. In: Hall ER, Hobson PN (ed) Anaerobic digestion 1988. Pergamon Press, Oxford, pp 1-12
Zinder SH, Koch M (1984) Non-acetoclastic methanogenesis from acetate: acetate-oxidation by a thermophilic acetotro- phic syntrophic coeulture. Arch Microbiol 138:263-272
Zinder SH, Mah RB (1979) Isolation and characterization of a thermophilic strains of Methanosarcina unable to use H2-CO2 for methanogenesis. Appl Environ Microbiol 38:996-1008
Zinder SH, Anguish T, Lobo A1 (1987) Isolation and characteri- zation of a thermophilic acetotrophic strain of Methanothrix. Arch Microbiol 146: 315-322




















