
Probiotics inhibit nuclear factor-κB and induce heat shock proteins in colonic epithelial cells...
14
Probiotics Inhibit Nuclear Factor-B and Induce Heat Shock Proteins in Colonic Epithelial Cells Through Proteasome Inhibition ELAINE O. PETROF,* KEISHI KOJIMA,* MARK J. ROPELESKI, ‡ MARK W. MUSCH,* YUN TAO,* CLAUDIO DE SIMONE, § and EUGENE B. CHANG* *Department of Medicine, University of Chicago, Chicago, Illinois; ‡ Department of Medicine, Queen’s University, Kingston, Ontario, Canada; and § University of L’Aquila, L’Aquila, Italy Background & Aims: The extent and severity of muco- sal injury in inflammatory bowel diseases are deter- mined by the disequilibrium between 2 opposing pro- cesses: reparative and cytoprotective mechanisms vs. inflammation-induced injury. Probiotics may provide clinical benefit by ameliorating colitis; however, their mechanisms of action remain largely unknown. Our objective was to investigate microbial– epithelial in- teractions that could explain the beneficial therapeu- tic effects of probiotics. Methods: The effect of VSL#3- conditioned media on the nuclear factor-B pathway in young adult mouse colonic epithelial cells was assessed by using monocyte chemoattractant pro- tein-1 enzyme-linked immunosorbent assays; IB, IB, and p105 immunoblot analysis; and nuclear factor-B luciferase reporter gene and proteasome assays. Effects on heat shock proteins were deter- mined by electrophoretic mobility shift assay and im- munoblot for heat shock proteins 25 and 72 in young adult mouse colonic cells. Cytoprotection against ox- idant injury was determined by chromium 51 release and filamentous and globular actin assays. Results: VSL#3 produces soluble factors that inhibit the chy- motrypsin-like activity of the proteasome in gut epi- thelial cells. Proteasome inhibition is an early event that begins almost immediately after exposure of the epithelial cells to the probiotic-conditioned media. In addition, these bacteria inhibit the proinflammatory nuclear factor-B pathway through a mechanism dif- ferent from the type III secretory mechanisms de- scribed for other nonpathogenic enteric flora. They also induce the expression of cytoprotective heat shock proteins in intestinal epithelial cells. Conclusions: The resulting inhibition of nuclear fac- tor-B and increased expression of heat shock pro- teins may account for the anti-inflammatory and cy- toprotective effects reported for probiotics and may be a novel mechanism of microbial– epithelial inter- action. These effects seem to be mediated through the common unifying mechanism of proteasome in- hibition. T here is growing interest in the use of probiotics, defined as ingestible microorganisms with health benefits beyond their intrinsic nutritive value. Probiotics have been used to treat a variety of gastrointestinal ailments, including inflammatory bowel disease (IBD), 1 irritable bowel syndrome, 2 and pouchitis, 3,4 as well as rotavirus and antibiotic-associated diarrhea. 5–7 Although little is known about their mechanisms of action, probi- otics seem to have protective, trophic, and anti-inflam- matory effects on bowel mucosa. Proposed mechanisms include the production of ammonia, hydrogen perox- ide, 8 –10 and bacteriocins, 11–13 which inhibit the growth of pathogenic bacteria, the competition for adhesion sites on intestinal epithelia, 14,15 and an adjuvant-like stimu- lation of the immune system against pathogenic organ- isms. 16 However, the exact mechanisms by which probi- otics act to protect against intestinal inflammation have yet to be fully elucidated. The probiotic VSL#3 (com- posed of Streptococcus thermophilus and several species of Lactobacillus and Bifidobacteria) attenuates intestinal in- flammation in the interleukin-10 knockout mouse model of enterocolitis 17 and has been shown to improve the clinical outcome of chronic intestinal inflammation in clinical trials. 3 In a randomized, double-blinded, place- bo-controlled trial of recurrent pouchitis, patients as- signed to receive placebo all relapsed within 4 months, whereas only 15% assigned to the probiotic treatment arm relapsed. 3 In addition to maintenance therapy, VSL#3 as a prophylactic treatment may help to prevent Abbreviations used in this paper: AMC, 7-amino-4-methylcoumarin; CM, conditioned medium; F actin, filamentous actin; G actin, globular actin; HSF, heat shock transcription factor; hsp, heat shock protein; IFN, interferon; MCP, monocyte chemoattractant protein; MSIE, mu- rine small-intestine epithelial cells; NF, nuclear factor; TBS, Tris-buff- ered saline; TNF, tumor necrosis factor; YAMC, young adult mouse colon. © 2004 by the American Gastroenterological Association 0016-5085/04/$30.00 doi:10.1053/j.gastro.2004.09.001 GASTROENTEROLOGY 2004;127:1474 –1487
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Probiotics inhibit nuclear factor-κB and induce heat shock proteins in colonic epithelial cells...

PPI
EC*a
BsmcicmottciatIfammaiaVmtteanfsasCtttbath
GASTROENTEROLOGY 2004;127:1474–1487
robiotics Inhibit Nuclear Factor-�B and Induce Heat Shockroteins in Colonic Epithelial Cells Through Proteasome
nhibition
LAINE O. PETROF,* KEISHI KOJIMA,* MARK J. ROPELESKI,‡ MARK W. MUSCH,* YUN TAO,*LAUDIO DE SIMONE,§ and EUGENE B. CHANG*Department of Medicine, University of Chicago, Chicago, Illinois; ‡Department of Medicine, Queen’s University, Kingston, Ontario, Canada;
nd §University of L’Aquila, L’Aquila, ItalyTbhairlomiioolioypLfloccbswaV
CaIrec
ackground & Aims: The extent and severity of muco-al injury in inflammatory bowel diseases are deter-ined by the disequilibrium between 2 opposing pro-
esses: reparative and cytoprotective mechanisms vs.nflammation-induced injury. Probiotics may providelinical benefit by ameliorating colitis; however, theirechanisms of action remain largely unknown. Our
bjective was to investigate microbial–epithelial in-eractions that could explain the beneficial therapeu-ic effects of probiotics. Methods: The effect of VSL#3-onditioned media on the nuclear factor-�B pathwayn young adult mouse colonic epithelial cells wasssessed by using monocyte chemoattractant pro-ein-1 enzyme-linked immunosorbent assays; I�B�,�B�, and p105 immunoblot analysis; and nuclearactor-�B luciferase reporter gene and proteasomessays. Effects on heat shock proteins were deter-ined by electrophoretic mobility shift assay and im-unoblot for heat shock proteins 25 and 72 in young
dult mouse colonic cells. Cytoprotection against ox-dant injury was determined by chromium 51 releasend filamentous and globular actin assays. Results:SL#3 produces soluble factors that inhibit the chy-otrypsin-like activity of the proteasome in gut epi-
helial cells. Proteasome inhibition is an early eventhat begins almost immediately after exposure of thepithelial cells to the probiotic-conditioned media. Inddition, these bacteria inhibit the proinflammatoryuclear factor-�B pathway through a mechanism dif-erent from the type III secretory mechanisms de-cribed for other nonpathogenic enteric flora. Theylso induce the expression of cytoprotective heathock proteins in intestinal epithelial cells.onclusions: The resulting inhibition of nuclear fac-or-�B and increased expression of heat shock pro-eins may account for the anti-inflammatory and cy-oprotective effects reported for probiotics and maye a novel mechanism of microbial–epithelial inter-ction. These effects seem to be mediated throughhe common unifying mechanism of proteasome in-
ibition.here is growing interest in the use of probiotics,defined as ingestible microorganisms with health
enefits beyond their intrinsic nutritive value. Probioticsave been used to treat a variety of gastrointestinalilments, including inflammatory bowel disease (IBD),1
rritable bowel syndrome,2 and pouchitis,3,4 as well asotavirus and antibiotic-associated diarrhea.5–7 Althoughittle is known about their mechanisms of action, probi-tics seem to have protective, trophic, and anti-inflam-atory effects on bowel mucosa. Proposed mechanisms
nclude the production of ammonia, hydrogen perox-de,8–10 and bacteriocins,11–13 which inhibit the growthf pathogenic bacteria, the competition for adhesion sitesn intestinal epithelia,14,15 and an adjuvant-like stimu-ation of the immune system against pathogenic organ-sms.16 However, the exact mechanisms by which probi-tics act to protect against intestinal inflammation haveet to be fully elucidated. The probiotic VSL#3 (com-osed of Streptococcus thermophilus and several species ofactobacillus and Bifidobacteria) attenuates intestinal in-ammation in the interleukin-10 knockout mouse modelf enterocolitis17 and has been shown to improve thelinical outcome of chronic intestinal inflammation inlinical trials.3 In a randomized, double-blinded, place-o-controlled trial of recurrent pouchitis, patients as-igned to receive placebo all relapsed within 4 months,hereas only 15% assigned to the probiotic treatment
rm relapsed.3 In addition to maintenance therapy,SL#3 as a prophylactic treatment may help to prevent
Abbreviations used in this paper: AMC, 7-amino-4-methylcoumarin;M, conditioned medium; F actin, filamentous actin; G actin, globularctin; HSF, heat shock transcription factor; hsp, heat shock protein;FN, interferon; MCP, monocyte chemoattractant protein; MSIE, mu-ine small-intestine epithelial cells; NF, nuclear factor; TBS, Tris-buff-red saline; TNF, tumor necrosis factor; YAMC, young adult mouseolon.
© 2004 by the American Gastroenterological Association0016-5085/04/$30.00
doi:10.1053/j.gastro.2004.09.001

ta
Ippsamfjictscad
(iourccepmfspufrtmt
paeesiaowIdmi
clat
tNt“tmttefietupr
ctdaptcas(d
tlochaNs(B5Bf
November 2004 PROBIOTICS IN COLONIC EPITHELIAL CELLS 1475
he onset of acute pouchitis in the year after ileal pouch/nal anastomosis after colectomy for ulcerative colitis.4
Current concepts regarding the etiopathogenesis ofBD suggest that there is a disequilibrium between therocesses of cytoprotection and wound healing and theroinflammatory pathways, the net result of which is atate of proinflammatory overactivity and resultant dam-ge to the intestinal mucosa.18,19 Central to preservingucosal integrity is maintenance of epithelial barrier
unction, as evidenced by the fact that altered tightunction structure resulting in impaired barrier functions thought to contribute to the clinical sequelae of ul-erative colitis.20 Through the use of sense and antisenseransfection experiments, it has been shown that heathock proteins (hsps) play a central role in providingytoprotection to epithelial cells, as illustrated by theirbility to protect epithelial barrier function under con-itions of oxidative stress.21,22
Activation of the proinflammatory nuclear factorNF)-�B pathway is thought to be a key molecular eventnvolved in the pathogenesis of IBD.23–26 Administrationf antisense oligonucleotides targeting the NF-�B sub-nit p65 was more effective than steroid treatment ineducing inflammation in 2 different murine models ofolitis.27 Immunohistochemical studies have shown thatolonic biopsy samples from patients with Crohn’s dis-ase display increased expression of the NF-�B subunit65 in areas of active inflammation.28 In the noninflam-atory state, NF-�B is held in its inactive, cytosolic
orm complexed to the inhibitory protein I�B. Once aignal is received to activate NF-�B, its inhibitor I�B ishosphorylated and targeted for degradation by thebiquitin–proteasome pathway. The release of NF-�Brom inhibition and its translocation to the nucleusesults in the transcriptional activation of a broad spec-rum of cytokine and chemokine genes, cell adhesionolecules, and immunoreceptors—all important media-
ors of the inflammatory response.23–26
Inducible hsps belong to a family of highly conservedroteins that play an important role in protecting cellsgainst physiological and pathogenic stressors in thenvironment. Under conditions of stress—such as heat,xposure to heavy metals and toxins, ischemia/reperfu-ion injury, or oxidative stress from inflammation—hspnduction is both rapid and robust. Induction of hsps bymild stress confers protection against subsequent insultr injury that would otherwise lead to cell death. Thisell-described phenomenon is known as stress tolerance.29
n intestinal epithelial cells, inducible hsps convey aegree of cytoprotection against stressors such as inflam-atory cell-derived oxidants and preserve the integrity of
ntestinal epithelial cell barrier function under hostile u
onditions.18,30,31 Induction of hsps in intestinal epithe-ial cells prolongs viability under conditions of stress30
nd preserves tight junctions as measured by transepi-helial resistance.31
This report shows that probiotic bacteria attenuate theumor necrosis factor (TNF)-�–mediated induction ofF-�B activation in intestinal epithelial cells and induce
he expression of cytoprotective hsps, thus affecting 2arms” of current IBD models. These beneficial effects onhe gut mucosa seem to stem from a common mechanismediated by proteasome inhibition. Proteasome inhibi-
ion by VSL#3-conditioned media (CM) is an early eventhat begins almost immediately after exposure of thepithelial cells to the probiotic-CM. This study identi-ed a novel mechanism by which bacteria may exertffects on the physiology of the intestinal epitheliumhrough the inhibition of proteasome function; this mayltimately alter programs of epithelial gene responses toathogenic factors derived from the luminal microenvi-onment of the gastrointestinal tract.
Materials and Methods
Probiotic Bacterial Culture and Generationof Conditioned Media
The probiotic formulation, VSL#3 (VSL Pharmaceuti-als, Gaithersburg, MD), contains Streptococcus salivarius subsphermophilus, Lactobacillus casei, L plantarum, L acidophilus, Lelbrueckii subsp. bulgaricus, Bifidobacteria longum, B infantis,nd B breve at a concentration of 5 � 1011 lyophilized bacteriaer gram. VSL#3 (batch 2034-A2) was grown to a concentra-ion of approximately 2 � 1014 (as determined by colonyounts) in phenol red–free RPMI 1640 medium for 16 hoursnd then centrifuged (3000g at 4°C for 10 minutes). Theupernatant (CM) was then passed through a 0.22-�m filterMillipore, Bedford, MA) to remove all bacterial cells andebris. Aliquots of CM were stored at �80°C until further use.
Tissue Culture
Young adult mouse colon (YAMC) cells are a condi-ionally immortalized mouse colonic intestinal epithelial celline derived from the Immortimouse that expresses a transgenef a temperature-sensitive SV40 large T antigen (tsA58) underontrol of an interferon (IFN)-�–sensitive portion of the majoristocompatibility complex class II promoter.32 The cells weregenerous gift of Dr. R. Whitehead (Vanderbilt University,ashville, TN). YAMC cells were maintained under permis-
ive conditions (33°C) in RPMI 1640 medium with 5%vol/vol) fetal bovine serum, 5 U/mL murine IFN-� (GibcoRL, Grand Island, NY), 50 �g/mL streptomycin, and0 U/mL penicillin, supplemented with ITS� Premix (BDiosciences, Bedford, MA). Under nonpermissive (nontrans-
ormed) conditions at 37°C in the absence of IFN-�, these cells
ndergo differentiation and develop mature epithelial cell
fpt
hmmafttNRdeftPwecm
mMIaNpkwaYwwVla6tiBtccE
eomwo
wcrtac
3odcNpfros5Nippfmdfidf6w1pofbtt
tdl1aHbsfw(oG
1476 PETROF ET AL. GASTROENTEROLOGY Vol. 127, No. 5
unctions and properties, including tight junction formation,olarity, microvillar apical membranes, and transport func-ions.
Cells were plated in permissive conditions, and after 24ours of growth at 33°C to allow for cell attachment, theedium was replaced with IFN-free media, and cells wereoved to 37°C (nonpermissive conditions) for 24 hours to
llow development of the differentiated colonocyte phenotypeor all experiments. Unless otherwise specified, cells werereated with VSL#3-CM (1:10 dilution) overnight and werehen used the following day in various experiments. ForF-�B luciferase reporter assays, murine TNF-� (Peprotech,ocky Hill, NJ) at a concentration of 50 ng/mL was addedirectly 6 hours before harvest. Heat shock controls werexposed to 42°C for 23 minutes and allowed to recover at 37°Cor 2 hours before harvest. MG132-treated control cells werereated for 2 hours with 25 �mol/L of MG132 (Biomol,lymouth Meeting, PA) at 37°C before harvest unless other-ise specified. Two other cell lines, murine small-intestine
pithelial cells (MSIE; a small-intestine YAMC counterpartell line) and 3T3 fibroblasts, were used in this study and wereaintained as previously described.33
Nuclear Factor-�B Luciferase Assay
NF-�B luciferase assays were performed with the Pro-ega Dual Luciferase Reporter 1000 Assay System (Promega,adison, WI), and plasmids were transfected by using Trans-
T LT-1 polyamine transfection reagent (Mirus, Madison, WI)ccording to the manufacturer’s instructions. Briefly, 2 �g ofF-�B response element–driven firefly luciferase reporter
lasmid (Clontech, Palo Alto, CA) and 0.2 �g of thymidineinase promoter–driven Renilla reporter plasmid (Promega)ere mixed with leukotriene-1 polyamine transfection re-
gent. After formation of complexes, the solution was added toAMC cells at 33°C and allowed to incubate overnight. Cellsere then placed at 37°C in IFN-�–free media. After cellsere grown in nonpermissive (nontransformed) conditions,SL#3-CM was added to each well at a dilution of 1:10 and
eft overnight, unless otherwise specified. Murine TNF-� wasdded at 50 ng/mL the next morning, and cells were harvestedhours later. NF-�B luciferase assays were performed by using
he Dual Luciferase Assay according to the manufacturer’snstructions (Promega), and luminescence was measured in aerthold Lumat 9047 Luminometer (Oakridge, TN). Co-
ransfection with thymidine kinase–Renilla, which displaysonstitutive low levels of activity, was used as an internalontrol against which to normalize the NF-�B luciferase data.xperiments were performed in triplicate.
Chromium-Release Assay for Cell Viability
YAMC cells were grown in 24-well plates and wereither left untreated (control) or were treated with VSL#3-CMvernight. Cells were loaded with chromium 51 (51Cr; 50 �Ci/L; Sigma Chemical Co., St. Louis, MO) for 60 minutes,ashed, and incubated in media with 0.6 mmol/L of the
xidant monochloramine (NH2Cl) to induce cell injury. Media rere harvested after 60 minutes, and the 51Cr remaining in theells was extracted with 1N HNO3 for 4 hours. The 51Cr in theeleased and cellular fractions was counted by liquid scintilla-ion spectroscopy. The 51Cr released was calculated as themount released divided by the amount released plus theellular remainder.
Globular and Filamentous Actin Assay
Confluent YAMC cell monolayers were switched to7°C in IFN-�–free medium and treated with VSL#3-CMvernight. Before assessment, cells were treated with phalloi-in (30 �g/mL for 2 hours; Molecular Probes, Eugene, OR),ytochalasin D (10 �g/mL for 15 minutes), or the oxidantH2Cl (0.6 mmol/L for 30 minutes).30,31 Cells were rinsed in
hosphate-buffered saline, harvested, and centrifuged (14,000gor 20 seconds at room temperature), and the pellets wereesuspended in 200 �L of 30°C lysis buffer [1 mmol/L aden-sine triphosphate, 50 mmol/L piperazine-N,N=-bis(2-ethane-ulfonic acid) pH 6.9, 50 mmol/L NaCl, 5 mmol/L MgCl2,
mmol/L ethylene glycol-bis(�-aminoethyl ether)-N,N,N=,=-tetraacetic acid, 5% (vol/vol) glycerol, 0.1% (vol/vol) Non-
det P-40, Tween 20, and Triton X-100, containing completerotease inhibitor cocktail]. Cells were homogenized by gentlyipetting up and down 10 times and were incubated at 30°Cor 10 minutes. Samples were centrifuged at 100,000g for 60inutes at 30°C, and the supernatants were removed for
etermination of globular actin (G actin). Pellets containinglamentous actin (F actin) were resuspended in 200 �L of 4°Cistilled water with 1 �mol/L cytochalasin D and left on iceor 60 minutes. Then 20 �L of each extraction was removed,�L of 3� Laemmli stop solution was added, and the samplesere heated to 65°C for 10 minutes. Samples were resolved by2.5% sodium dodecyl sulfate-polyacrylamide gel electro-horesis and immediately transferred to polyvinylidene diflu-ride membranes. After transfer, analysis of actin was per-ormed with a polyclonal antiactin antiserum by Westernlotting (Cytoskeleton, Denver, CO). Because the F-actin frac-ion has been depolymerized, only the monomeric 45-kilodal-on form is observed on the Western blots.
Electrophoretic Mobility Shift Assay
YAMC cells were either treated with VSL#3-CM (forimes varying from 15 minutes to 6 hours) or heat-shocked asescribed previously. Whole-cell extracts were prepared inysis buffer (25% vol/vol glycerol, 420 mmol/L NaCl,.5 mmol/L MgCl2, 0.2 mmol/L ethylenediaminetetraaceticcid [EDTA], 0.5 mmol/L dithiothreitol, and 20 mmol/LEPES, pH 7.4, with complete protease inhibitor cocktail)
y freezing once in a dry ice/alcohol bath, thawing on ice,hearing gently with a pipet tip, and centrifuging at 50,000gor 5 minutes at 4°C. Cell extract containing 10 �g of proteinas mixed with phosphorus-32–labeled heat shock element
HSE) oligonucleotide (containing 4 tandem inverted repeatsf the heat shock element [nGAAn]; CTAGAAGCTTCTA-AAGCTTCTAG) and 0.5 �g of poly(dI-dC) in binding
eaction buffer (final concentrations: 20 mmol/L Tris [pH 7.4],

1empbpwataCmbu
HK(pmclwpu
atapic
mSaH0omanoome(aa
aiaoppsAibc
otmvbMT5[aswwactIbUpibtimTmI
ecct
November 2004 PROBIOTICS IN COLONIC EPITHELIAL CELLS 1477
00 mmol/L NaCl, 1 mmol/L EDTA, and 10% vol/vol glyc-rol). The binding reaction was allowed to incubate for 25inutes at 25°C and then analyzed on a 4% nondenaturing
olyacrylamide gel run in 0.5� Tris-Borate-EDTA (TBE)uffer. Gels were dried and autoradiographed to detect DNA–rotein complexes. For supershift experiments, YAMC cellsere incubated with VSL#3-CM for 6 hours before harvest,
nd 1 �g of specific antibodies to either heat shock transcrip-ion factor (HSF)-1 (SPA-950; Stressgen, Victoria, BC, Can-da) or HSF-2 (sc-8062X; Santa Cruz Biotechnology, Santaruz, CA) was preincubated with cell extracts at 25°C for 30inutes before the HSE-binding reaction. After this preincu-
ation, the binding reaction and analysis were performed assual.
Preparation of Cell Lysates
Cells were washed twice and then scraped in ice-coldEPES buffered saline (HBS) (150 mmol/L NaCl, 5 mmol/LCl, and 10 mmol/L HEPES, pH 7.4). Cells were pelleted
14,000g for 20 seconds at room temperature) and then resus-ended in ice-cold lysis buffer (10 mmol/L Tris [pH 7.4], 5mol/L MgCl2, and 50 U/mL deoxyribonuclease and ribonu-
lease, plus complete protease inhibitor cocktail; Roche Mo-ecular Biochemicals, Indianapolis, IN). Protein concentrationsere determined with the bicinchoninic acid procedure.34 Forroteasome assays, samples were stored immediately at �80°Cntil use.
Monocyte Chemoattractant Protein-1Enzyme-Linked Immunosorbent Assay
YAMC cells were grown and treated with VSL#3-CMnd subsequently treated with TNF-�, as described previously,o stimulate NF-�B activation. Supernatants were harvestednd tested for the production of monocyte chemoattractantrotein (MCP)-1 by using a mouse MCP-1 enzyme-linkedmmunosorbent assay kit (Pierce Endogen, Rockford, IL) ac-ording to the manufacturer’s instructions.
Proteasome Assay
Proteasome activity from cell lysates was deter-ined by using a 20S Proteasome assay kit (Calbiochem,
an Diego, CA). Lysate containing 20 �g of protein wasdded to the proteasome assay reaction buffer (25 mmol/LEPES and 0.5 mmol/L EDTA, pH 7.6) activated with
.03% (wt/vol) sodium dodecyl sulfate, and then 10 �mol/Lf the substrate suc-leu-leu-val-tyr–7-amino-4-methylcou-arin (AMC) was added. Proteasome chymotrypsin-like
ctivity was determined by measuring the fluorogenic sig-al generated by cleavage of AMC from the peptide moietyf the suc-leu-leu-val-tyr-AMC proteasome substrate. Flu-rescence (excitation 380 nm and emission 460 nm) waseasured every minute for the first 10 minutes and then
very 15 minutes thereafter in a Hitachi F-2000 fluorometerHitachi, Naka, Japan). Cells were treated with MG132 aspositive inhibitor control at a concentration of 25 �mol/L,
nd untreated cells were treated with dimethyl sulfoxide as cvehicle control for MG132. Experiments were performedn triplicate. In addition, proteasome assays were performeds described previously by using fluorescent substrates rec-gnized and cleaved by the other protease activities of theroteasome, the trypsin-like activity, and the post-glutamyleptide hydrolyzing (PGPH) or caspase-like activity. Theubstrates used for these experiments were Bz-Val-Gly-Arg-MC and Z-Leu-Leu-Glu-AMC. A different proteasome
nhibitor, lactacystin, was used as an inhibitor controlecause of its ability to inhibit both the trypsin-like andaspase-like activities of the proteasome.
Western Blot Analysis
Twenty micrograms of protein per lane was resolvedn 12.5% sodium dodecyl sulfate-polyacrylamide gel elec-rophoresis and transferred in 1� Towbin buffer (25mol/L Tris, 192 mmol/L glycine [pH 8.8], and 15%
ol/vol methanol) onto polyvinylidene difluoride mem-ranes (Polyscreen; PerkinElmer NEN, Boston, MA).33
embranes were blocked in 5% (wt/vol) nonfat milk inris-buffered saline (TBS)-Tween (TBS [150 mmol/L NaCl,mmol/L KCl, and 10 mmol/L Tris, pH 7.4] with 0.05%
vol/vol] Tween 20). For antiubiquitin, anti-IKB�, andnti-p105 blots, membranes were blocked in 3% bovineerum albumin (Fisher, Pittsburgh, PA). Primary antibodyas added to TBS-Tween and incubated overnight at 4°Cith a specific anti-hsp25 antibody (SPA801; Stressgen),
nti-hsp72 antibody (SPA 810; Stressgen), anti– heat shockognate 73 antibody (SPA 815; Stressgen), anti-I�B� an-ibody (sc-1643; Santa Cruz Biotechnology), anti–phospho�B� antibody (sc-8404; Santa Cruz), anti-ubiquitin anti-ody (PW 8810; Affiniti Research Products Ltd., Exeter,K), anti-IKB� antibody (sc-945; Santa Cruz), or anti-105 antibody (sc-114; Santa Cruz). Blots were then washedn TBS-Tween 5 times for 10 minutes at room temperatureefore incubation with peroxidase-conjugated secondary an-ibodies (Jackson Immunoresearch Labs, Inc., Fort Wash-ngton, PA). Membranes were washed (5 times for 10inutes) in TBS-Tween followed by a final wash in TBS (noween). Blots were visualized with an enhanced chemilu-inescence system reagent (Supersignal; Pierce, Rockford,
L).
Statistical Analysis
Experiments were repeated a minimum of 3 to 6 timesach. Values are expressed as mean SEM. Where multipleomparisons were made, analysis of variance with Bonferroni’sorrection was used to assess the significance of differences be-ween groups. P .05 was considered statistically significant.
Results
Probiotics Inhibit Nuclear Factor-�BActivation in Intestinal Epithelial Cells
To determine whether the bacteria in VSL#3 se-
rete factors possessing anti-inflammatory activity, the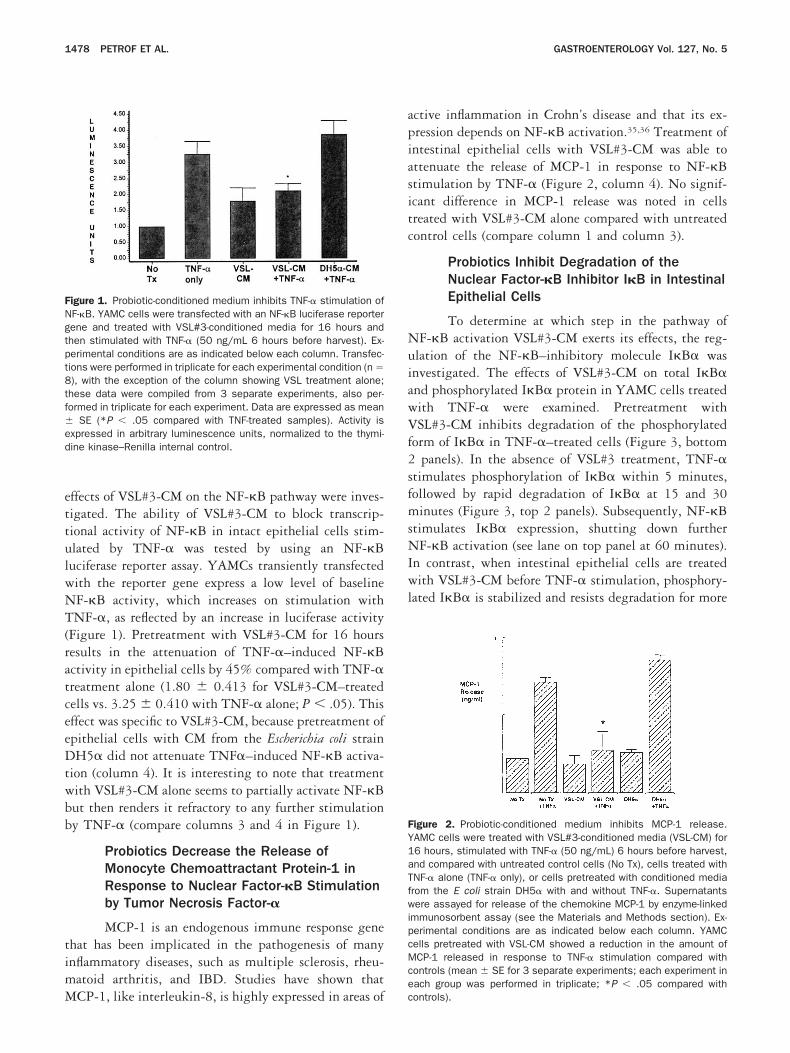
ettulwNT(ratceeDtwbb
timM
apiasitc
NuiawVf2sfmsNIwl
FY1aTfwipcMce
FNgtpt8tfed
1478 PETROF ET AL. GASTROENTEROLOGY Vol. 127, No. 5
ffects of VSL#3-CM on the NF-�B pathway were inves-igated. The ability of VSL#3-CM to block transcrip-ional activity of NF-�B in intact epithelial cells stim-lated by TNF-� was tested by using an NF-�Buciferase reporter assay. YAMCs transiently transfectedith the reporter gene express a low level of baselineF-�B activity, which increases on stimulation withNF-�, as reflected by an increase in luciferase activity
Figure 1). Pretreatment with VSL#3-CM for 16 hoursesults in the attenuation of TNF-�–induced NF-�Bctivity in epithelial cells by 45% compared with TNF-�reatment alone (1.80 0.413 for VSL#3-CM–treatedells vs. 3.25 0.410 with TNF-� alone; P .05). Thisffect was specific to VSL#3-CM, because pretreatment ofpithelial cells with CM from the Escherichia coli strainH5� did not attenuate TNF�–induced NF-�B activa-
ion (column 4). It is interesting to note that treatmentith VSL#3-CM alone seems to partially activate NF-�But then renders it refractory to any further stimulationy TNF-� (compare columns 3 and 4 in Figure 1).
Probiotics Decrease the Release ofMonocyte Chemoattractant Protein-1 inResponse to Nuclear Factor-�B Stimulationby Tumor Necrosis Factor-�
MCP-1 is an endogenous immune response genehat has been implicated in the pathogenesis of manynflammatory diseases, such as multiple sclerosis, rheu-atoid arthritis, and IBD. Studies have shown that
igure 1. Probiotic-conditioned medium inhibits TNF-� stimulation ofF-�B. YAMC cells were transfected with an NF-�B luciferase reporterene and treated with VSL#3-conditioned media for 16 hours andhen stimulated with TNF-� (50 ng/mL 6 hours before harvest). Ex-erimental conditions are as indicated below each column. Transfec-ions were performed in triplicate for each experimental condition (n �), with the exception of the column showing VSL treatment alone;hese data were compiled from 3 separate experiments, also per-ormed in triplicate for each experiment. Data are expressed as mean
SE (*P .05 compared with TNF-treated samples). Activity isxpressed in arbitrary luminescence units, normalized to the thymi-ine kinase–Renilla internal control.
CP-1, like interleukin-8, is highly expressed in areas of c
ctive inflammation in Crohn’s disease and that its ex-ression depends on NF-�B activation.35,36 Treatment ofntestinal epithelial cells with VSL#3-CM was able tottenuate the release of MCP-1 in response to NF-�Btimulation by TNF-� (Figure 2, column 4). No signif-cant difference in MCP-1 release was noted in cellsreated with VSL#3-CM alone compared with untreatedontrol cells (compare column 1 and column 3).
Probiotics Inhibit Degradation of theNuclear Factor-�B Inhibitor I�B in IntestinalEpithelial Cells
To determine at which step in the pathway ofF-�B activation VSL#3-CM exerts its effects, the reg-
lation of the NF-�B–inhibitory molecule I�B� wasnvestigated. The effects of VSL#3-CM on total I�B�nd phosphorylated I�B� protein in YAMC cells treatedith TNF-� were examined. Pretreatment withSL#3-CM inhibits degradation of the phosphorylated
orm of I�B� in TNF-�–treated cells (Figure 3, bottompanels). In the absence of VSL#3 treatment, TNF-�
timulates phosphorylation of I�B� within 5 minutes,ollowed by rapid degradation of I�B� at 15 and 30
inutes (Figure 3, top 2 panels). Subsequently, NF-�Btimulates I�B� expression, shutting down furtherF-�B activation (see lane on top panel at 60 minutes).
n contrast, when intestinal epithelial cells are treatedith VSL#3-CM before TNF-� stimulation, phosphory-
ated I�B� is stabilized and resists degradation for more
igure 2. Probiotic-conditioned medium inhibits MCP-1 release.AMC cells were treated with VSL#3-conditioned media (VSL-CM) for6 hours, stimulated with TNF-� (50 ng/mL) 6 hours before harvest,nd compared with untreated control cells (No Tx), cells treated withNF-� alone (TNF-� only), or cells pretreated with conditioned mediarom the E coli strain DH5� with and without TNF-�. Supernatantsere assayed for release of the chemokine MCP-1 by enzyme-linked
mmunosorbent assay (see the Materials and Methods section). Ex-erimental conditions are as indicated below each column. YAMCells pretreated with VSL-CM showed a reduction in the amount ofCP-1 released in response to TNF-� stimulation compared with
ontrols (mean SE for 3 separate experiments; each experiment inach group was performed in triplicate; *P .05 compared with
ontrols).
tmotniuyVY2
TpapsiumlimCiecoew
tVtbtdmtps
variVtpM(t
FmcbVast
FrmhtddtdI
November 2004 PROBIOTICS IN COLONIC EPITHELIAL CELLS 1479
han 2 hours (Figure 3, lower bottom panel). Further-ore, the amount of total I�B� never declines through-
ut the period of TNF-� stimulation, thus suggestinghat VSL#3-CM inhibits pathways of I�B� degradationormally associated with TNF-� stimulation of gut ep-thelial cells. Whereas VSL#3-CM had some basal (yetndefined) regulatory effect at the level of I�B phosphor-lation, as shown in Figure 3 (lane 1), no effect ofSL#3-CM alone was seen on MCP-1 secretion byAMC cells in the absence of TNF-�, as shown in Figure(column 3).
Probiotics Do Not Inhibit Ubiquitination inIntestinal Epithelial Cells
The lack of I�B� degradation in response toNF-� in VSL#3-CM–treated cells implies that therobiotic-CM interferes with 1 or more downstreamctivation events, namely, the steps of ubiquitination orroteasomal degradation of I�B�. A nonpathogenictrain of Salmonella typhimurium has been reported tonhibit degradation of I�B� through blockade of ubiq-itination.37 These effects of S typhimurium may be 1ethod by which gastrointestinal flora is able to modu-
ate the immune system and thus live in symbiosis withts eukaryotic host. To test whether a similar mechanismay be involved in the inhibition of NF-�B by VSL#3-M, immunoblot analyses with anti-ubiquitin antibod-
es were performed on total cell protein from intestinalpithelial cells treated with VSL#3-CM (Figure 4). Inontrast to what might have been expected on the basisf the ability of nonpathogenic S typhimurium to inhibitpithelial ubiquitin ligase, treatment of epithelial cells
igure 3. Probiotic-conditioned medium stabilizes and prevents deg-adation of I�B�. YAMC cells were treated with VSL#3-conditionededia for 16 hours and then stimulated with TNF-� (50 ng/mL) andarvested at the times indicated. (Upper panel) TNF-� stimulatesransient phosphorylation of I�B� (5 minutes) and is associated withecreased total I�B� (5–15 minutes) as I�B� is targeted for degra-ation. (Bottom panel) Pretreatment of YAMC cells with VSL#3-condi-ioned media inhibits the effects of TNF-� on I�B�, thus preventing itsegradation. Note the persistence of the phosphorylated form of�B�.
ith VSL#3-CM does not result in a decrease of ubiqui- p
inated proteins compared with untreated cells. In fact,SL#3-CM treatment actually results in the accumula-
ion of certain ubiquitinated proteins. One mechanismy which this may occur is through inhibition of pro-easome function, which results in accumulation of un-egraded, ubiquitinated proteins.38 Although not as dra-atic as the effects of the proteasome inhibitor MG132,
he amount and the pattern of increase in ubiquitinatedroteins on VSL#3-CM treatment are similar to what iseen with thermal stress.
Probiotics Inhibit Proteasome Activity inIntestinal Epithelial Cells
Proteasome inhibitors that block NF-�B acti-ation through inhibition of I�B� degradation havelready been described.39 Because probiotic treatmentesults in both attenuation of NF-�B activity andnhibition of I�B degradation, we tested the effect ofSL#3-CM on proteasome activity (Figure 5A). Ex-
racts from cells treated with VSL#3-CM were com-ared with untreated controls, cells treated withG132 (a potent proteasome inhibitor), and DH5�
E coli) CM for proteasome activity. Epithelial cellsreated with VSL#3-CM display markedly lower levels
igure 4. Global ubiquitination is not inhibited by VSL#3-conditionededia. Immunoblot analysis of ubiquitinated proteins from YAMC
ells after treatment with VSL-CM for 16 hours shows that globallockade of ubiquitination does not occur when cells are treated withSL-CM. MG132, a compound known to inhibit proteasome functionnd increase the accumulation of ubiquitinated proteins, is alsohown, as is thermal stress (HS). The pattern of ubiquitinated pro-eins observed after VSL-CM treatment most closely resembles the
attern seen with thermal stress.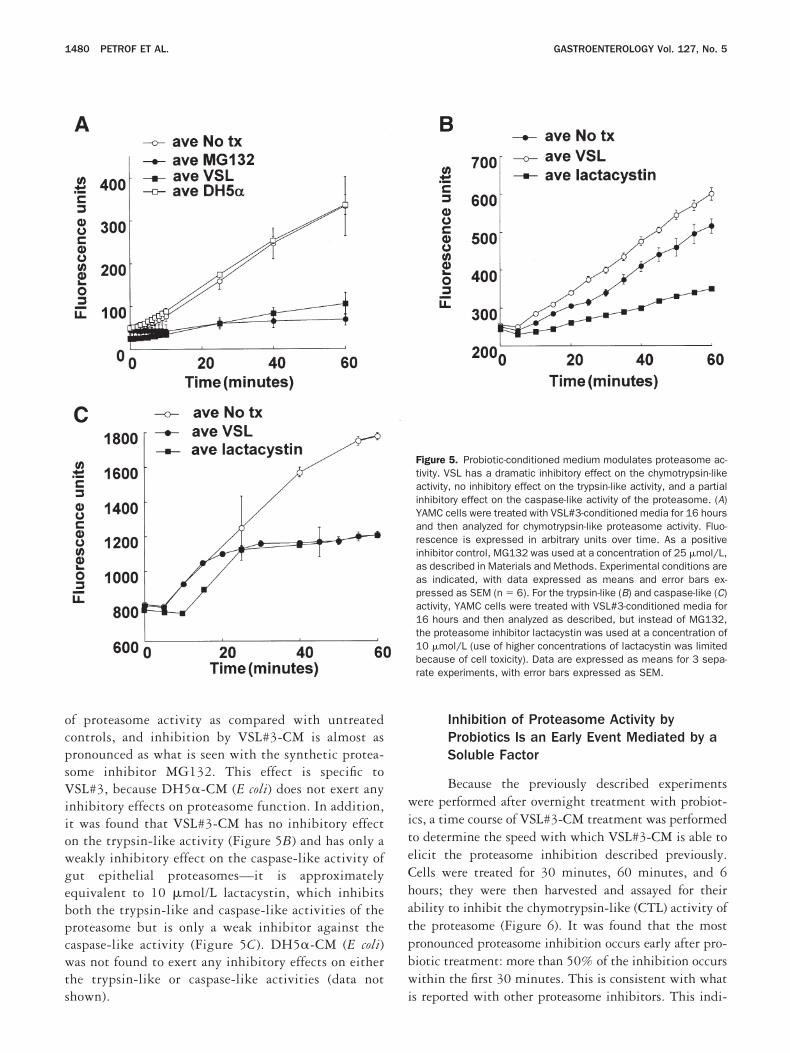
ocpsViiowgebpcwts
witeChatpbw
1480 PETROF ET AL. GASTROENTEROLOGY Vol. 127, No. 5
f proteasome activity as compared with untreatedontrols, and inhibition by VSL#3-CM is almost asronounced as what is seen with the synthetic protea-ome inhibitor MG132. This effect is specific toSL#3, because DH5�-CM (E coli) does not exert any
nhibitory effects on proteasome function. In addition,t was found that VSL#3-CM has no inhibitory effectn the trypsin-like activity (Figure 5B) and has only aeakly inhibitory effect on the caspase-like activity ofut epithelial proteasomes—it is approximatelyquivalent to 10 �mol/L lactacystin, which inhibitsoth the trypsin-like and caspase-like activities of theroteasome but is only a weak inhibitor against theaspase-like activity (Figure 5C). DH5�-CM (E coli)as not found to exert any inhibitory effects on either
he trypsin-like or caspase-like activities (data not
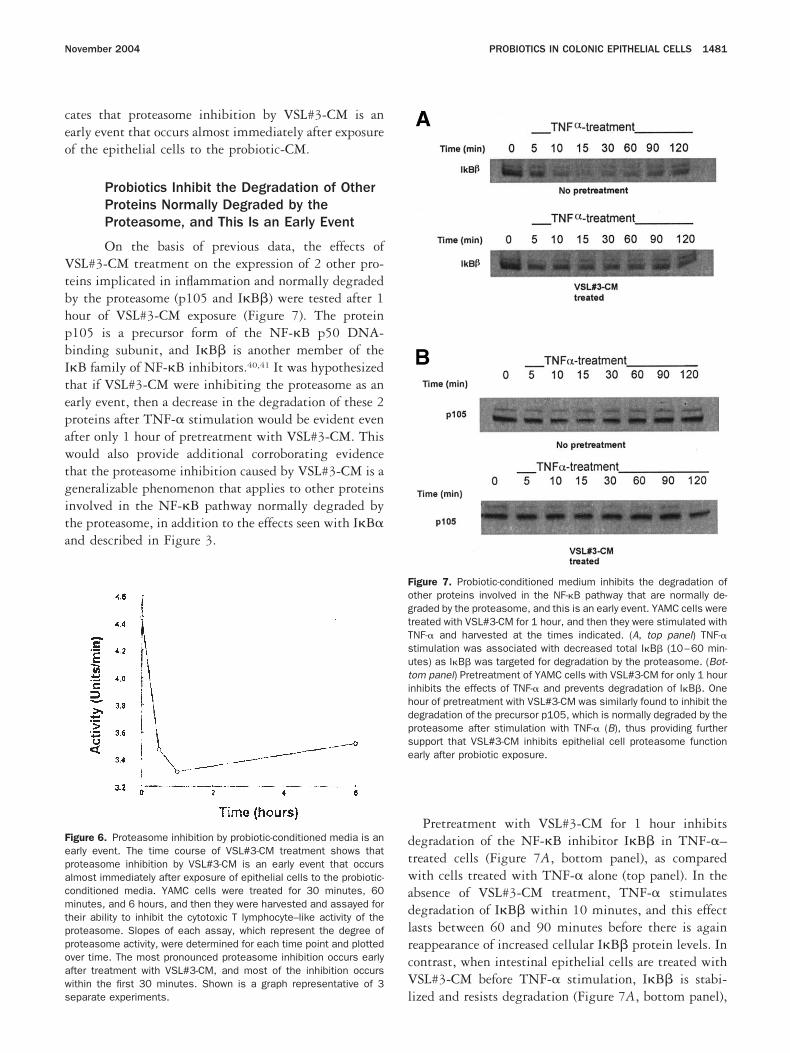
hown). iInhibition of Proteasome Activity byProbiotics Is an Early Event Mediated by aSoluble Factor
Because the previously described experimentsere performed after overnight treatment with probiot-
cs, a time course of VSL#3-CM treatment was performedo determine the speed with which VSL#3-CM is able tolicit the proteasome inhibition described previously.ells were treated for 30 minutes, 60 minutes, and 6ours; they were then harvested and assayed for theirbility to inhibit the chymotrypsin-like (CTL) activity ofhe proteasome (Figure 6). It was found that the mostronounced proteasome inhibition occurs early after pro-iotic treatment: more than 50% of the inhibition occursithin the first 30 minutes. This is consistent with what
Figure 5. Probiotic-conditioned medium modulates proteasome ac-tivity. VSL has a dramatic inhibitory effect on the chymotrypsin-likeactivity, no inhibitory effect on the trypsin-like activity, and a partialinhibitory effect on the caspase-like activity of the proteasome. (A)YAMC cells were treated with VSL#3-conditioned media for 16 hoursand then analyzed for chymotrypsin-like proteasome activity. Fluo-rescence is expressed in arbitrary units over time. As a positiveinhibitor control, MG132 was used at a concentration of 25 �mol/L,as described in Materials and Methods. Experimental conditions areas indicated, with data expressed as means and error bars ex-pressed as SEM (n � 6). For the trypsin-like (B) and caspase-like (C)activity, YAMC cells were treated with VSL#3-conditioned media for16 hours and then analyzed as described, but instead of MG132,the proteasome inhibitor lactacystin was used at a concentration of10 �mol/L (use of higher concentrations of lactacystin was limitedbecause of cell toxicity). Data are expressed as means for 3 sepa-rate experiments, with error bars expressed as SEM.
s reported with other proteasome inhibitors. This indi-
ceo
VtbhpbItepawtgita
dtwadlrcV
Fepacmtppoaws
FogtTsutihdpse
November 2004 PROBIOTICS IN COLONIC EPITHELIAL CELLS 1481
ates that proteasome inhibition by VSL#3-CM is anarly event that occurs almost immediately after exposuref the epithelial cells to the probiotic-CM.
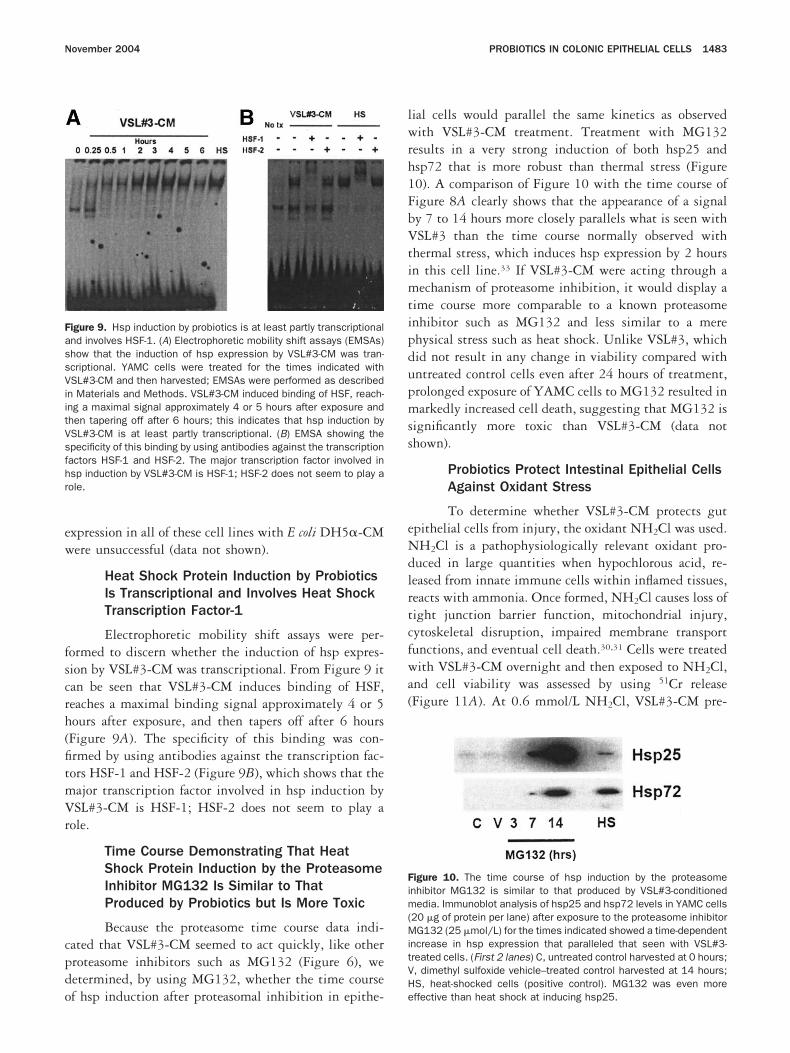
Probiotics Inhibit the Degradation of OtherProteins Normally Degraded by theProteasome, and This Is an Early Event
On the basis of previous data, the effects ofSL#3-CM treatment on the expression of 2 other pro-
eins implicated in inflammation and normally degradedy the proteasome (p105 and I�B�) were tested after 1our of VSL#3-CM exposure (Figure 7). The protein105 is a precursor form of the NF-�B p50 DNA-inding subunit, and I�B� is another member of the�B family of NF-�B inhibitors.40,41 It was hypothesizedhat if VSL#3-CM were inhibiting the proteasome as anarly event, then a decrease in the degradation of these 2roteins after TNF-� stimulation would be evident evenfter only 1 hour of pretreatment with VSL#3-CM. Thisould also provide additional corroborating evidence
hat the proteasome inhibition caused by VSL#3-CM is aeneralizable phenomenon that applies to other proteinsnvolved in the NF-�B pathway normally degraded byhe proteasome, in addition to the effects seen with I�B�nd described in Figure 3.
igure 6. Proteasome inhibition by probiotic-conditioned media is anarly event. The time course of VSL#3-CM treatment shows thatroteasome inhibition by VSL#3-CM is an early event that occurslmost immediately after exposure of epithelial cells to the probiotic-onditioned media. YAMC cells were treated for 30 minutes, 60inutes, and 6 hours, and then they were harvested and assayed for
heir ability to inhibit the cytotoxic T lymphocyte–like activity of theroteasome. Slopes of each assay, which represent the degree ofroteasome activity, were determined for each time point and plottedver time. The most pronounced proteasome inhibition occurs earlyfter treatment with VSL#3-CM, and most of the inhibition occursithin the first 30 minutes. Shown is a graph representative of 3
leparate experiments.
Pretreatment with VSL#3-CM for 1 hour inhibitsegradation of the NF-�B inhibitor I�B� in TNF-�–reated cells (Figure 7A, bottom panel), as comparedith cells treated with TNF-� alone (top panel). In the
bsence of VSL#3-CM treatment, TNF-� stimulatesegradation of I�B� within 10 minutes, and this effectasts between 60 and 90 minutes before there is againeappearance of increased cellular I�B� protein levels. Inontrast, when intestinal epithelial cells are treated withSL#3-CM before TNF-� stimulation, I�B� is stabi-
igure 7. Probiotic-conditioned medium inhibits the degradation ofther proteins involved in the NF-�B pathway that are normally de-raded by the proteasome, and this is an early event. YAMC cells werereated with VSL#3-CM for 1 hour, and then they were stimulated withNF-� and harvested at the times indicated. (A, top panel) TNF-�timulation was associated with decreased total I�B� (10–60 min-tes) as I�B� was targeted for degradation by the proteasome. (Bot-om panel) Pretreatment of YAMC cells with VSL#3-CM for only 1 hournhibits the effects of TNF-� and prevents degradation of I�B�. Oneour of pretreatment with VSL#3-CM was similarly found to inhibit theegradation of the precursor p105, which is normally degraded by theroteasome after stimulation with TNF-� (B), thus providing furtherupport that VSL#3-CM inhibits epithelial cell proteasome functionarly after probiotic exposure.
ized and resists degradation (Figure 7A, bottom panel),

cSeaptwt7fsp
boi1tdciibciUapt
btnab(lgeactgTittu
tc
assa3tV
Fdtcau(iYifMfcwfwaVdicT
1482 PETROF ET AL. GASTROENTEROLOGY Vol. 127, No. 5
onsistent with what was seen with I�B� in Figure 3.imilar findings were observed with the precursor mol-cule p105 (Figure 7B). Although less dramatic, thedministration of TNF-� in the absence of probioticretreatment results in a decrease in p105 cellular pro-ein from 10 to 30 minutes (Figure 7B top panel),hereas a much smaller decrease in p105 is observed in
he cells pretreated with VSL#3-CM for 1 hour (FigureB bottom panel). Taken together, these results provideurther substantiation that VSL#3-CM inhibits protea-ome function and that this event occurs early afterrobiotic exposure.
Probiotics Induce Heat Shock Proteins inIntestinal Epithelial Cells
It has been shown that the enteric flora (luminalacteria of the colon) in the gut influence the expressionf epithelial hsps.33,42 Proteasome inhibitors are potentnducers of hsp expression through activation of HSF-.43 On the basis of these data, we sought to corroboratehese findings by determining whether subsequent in-uction of hsp expression occurred. YAMC cells wereocultured with the probiotic VSL#3 to test its ability tonduce hsp expression. By immunoblot analysis, VSL#3nduced hsp25 and hsp72 expression in YAMC cellseginning after 6 hours (Figure 8A). Expression of theonstitutively expressed hsp heat shock cognate 73, serv-ng as a loading control, was not affected by VSL#3.ntreated cells left in culture for 48 hours do not mount
n hsp response, showing that the effect is specific to therobiotic treatment and is not a time-dependent charac-eristic of the cells grown in culture.
To determine whether the effects produced by VSL#3acteria require viable bacteria and direct physical con-act (e.g., as is necessary for type III secretion mecha-isms) or are secreted bacterial products, VSL#3 wasdded to YAMC cells, as filter-sterilized CM or sonicatedacterial pellets, in a concentration-dependent mannerFigure 8B). Although live gram-negative bacteria andipopolysaccharide, a cell wall component found only inram-negative bacteria, can induce hsp expression inpithelial cells,33 the bacteria that comprise VSL#3 arell gram-positive organisms, and, thus, none of themontains lipopolysaccharide. It was therefore of interesto discern whether any cell wall components of theseram-positive organisms possess hsp-inducing potential.he hsp induction produced by VSL#3 could be elicited
n a dose-response fashion with the CM alone, indicatinghat neither direct contact nor live bacteria are necessaryo elicit this response. Sonicated organisms (pellet) are
nable to induce the heat shock response, indicating that she inducing factors are likely secreted products and notell wall components.
Examination of 2 other cell lines revealed that thebility of probiotic-CM to induce hsp72 expression ispecific to epithelial cells (Figure 8C). MSIE is a murinemall-intestine epithelial cell line; these cells respond insimilar fashion to YAMC cells. In contrast, although
T3 fibroblasts are able mount a heat shock response tohermal stress, they do not respond to treatment withSL#3-CM, suggesting that the effects of VSL#3-CM are
igure 8. Hsp25 and hsp72 expression is induced by probiotics,oes not involve cell wall components, and is specific to epithelial cellypes. (A) Immunoblot analysis of levels of hsp25 and hsp72 in YAMCells after exposure to VSL#3 bacteria for the times indicated showstime-dependent increase in hsp expression. (Last 2 lanes) 48h,
ntreated controls harvested at 48 hours; HS, heat-shocked cellspositive controls). Hsc73 (heat shock cognate 73) served as a load-ng control. (B) Immunoblot analysis of levels of hsp25 and hsp72 inAMC cells after exposure to VSL#3-conditioned media (CM) or son-
cated organisms at the concentrations of bacteria indicated (colony-orming units [cfu] per milliliter). Bacteria were grown as described inaterials and Methods and were then separated into either a CM
raction or sonicated pellet (Pellet). A concentration-dependent in-rease in hsp expression can be seen on exposure to VSL#3-CM thatas not seen with sonicated pellet, thus indicating that the active
actors produced by the bacteria are secreted into CM and are not cellall components. Hsc73 served as a loading control. (C) Immunoblotnalysis of hsp72, comparing different cell lines after exposure toSL#3-CM for 16 hours (20 �g of protein per lane). VSL#3-CM in-uces a robust hsp72 response in both colonic (YAMC) and small-ntestinal (MSIE) epithelial cells that is not seen in 3T3 fibroblastells, suggesting that the probiotic effect is specific to epithelial cells.hermal stress (HS) served as a positive control.
pecific to epithelial cell types. Attempts to induce hsp
ew
fscrh(fitmVr
cpdo
lwrh1FbVtimtipdupmss
eNdlrtcfwa(
Fim(MitVH
FassViitVsfhr
November 2004 PROBIOTICS IN COLONIC EPITHELIAL CELLS 1483
xpression in all of these cell lines with E coli DH5�-CMere unsuccessful (data not shown).
Heat Shock Protein Induction by ProbioticsIs Transcriptional and Involves Heat ShockTranscription Factor-1
Electrophoretic mobility shift assays were per-ormed to discern whether the induction of hsp expres-ion by VSL#3-CM was transcriptional. From Figure 9 itan be seen that VSL#3-CM induces binding of HSF,eaches a maximal binding signal approximately 4 or 5ours after exposure, and then tapers off after 6 hoursFigure 9A). The specificity of this binding was con-rmed by using antibodies against the transcription fac-ors HSF-1 and HSF-2 (Figure 9B), which shows that theajor transcription factor involved in hsp induction bySL#3-CM is HSF-1; HSF-2 does not seem to play a
ole.
Time Course Demonstrating That HeatShock Protein Induction by the ProteasomeInhibitor MG132 Is Similar to ThatProduced by Probiotics but Is More Toxic
Because the proteasome time course data indi-ated that VSL#3-CM seemed to act quickly, like otherroteasome inhibitors such as MG132 (Figure 6), weetermined, by using MG132, whether the time course
igure 9. Hsp induction by probiotics is at least partly transcriptionalnd involves HSF-1. (A) Electrophoretic mobility shift assays (EMSAs)how that the induction of hsp expression by VSL#3-CM was tran-criptional. YAMC cells were treated for the times indicated withSL#3-CM and then harvested; EMSAs were performed as described
n Materials and Methods. VSL#3-CM induced binding of HSF, reach-ng a maximal signal approximately 4 or 5 hours after exposure andhen tapering off after 6 hours; this indicates that hsp induction bySL#3-CM is at least partly transcriptional. (B) EMSA showing thepecificity of this binding by using antibodies against the transcriptionactors HSF-1 and HSF-2. The major transcription factor involved insp induction by VSL#3-CM is HSF-1; HSF-2 does not seem to play aole.
f hsp induction after proteasomal inhibition in epithe- e
ial cells would parallel the same kinetics as observedith VSL#3-CM treatment. Treatment with MG132
esults in a very strong induction of both hsp25 andsp72 that is more robust than thermal stress (Figure0). A comparison of Figure 10 with the time course ofigure 8A clearly shows that the appearance of a signaly 7 to 14 hours more closely parallels what is seen withSL#3 than the time course normally observed with
hermal stress, which induces hsp expression by 2 hoursn this cell line.33 If VSL#3-CM were acting through a
echanism of proteasome inhibition, it would display aime course more comparable to a known proteasomenhibitor such as MG132 and less similar to a merehysical stress such as heat shock. Unlike VSL#3, whichid not result in any change in viability compared withntreated control cells even after 24 hours of treatment,rolonged exposure of YAMC cells to MG132 resulted inarkedly increased cell death, suggesting that MG132 is
ignificantly more toxic than VSL#3-CM (data nothown).
Probiotics Protect Intestinal Epithelial CellsAgainst Oxidant Stress
To determine whether VSL#3-CM protects gutpithelial cells from injury, the oxidant NH2Cl was used.H2Cl is a pathophysiologically relevant oxidant pro-
uced in large quantities when hypochlorous acid, re-eased from innate immune cells within inflamed tissues,eacts with ammonia. Once formed, NH2Cl causes loss ofight junction barrier function, mitochondrial injury,ytoskeletal disruption, impaired membrane transportunctions, and eventual cell death.30,31 Cells were treatedith VSL#3-CM overnight and then exposed to NH2Cl,
nd cell viability was assessed by using 51Cr releaseFigure 11A). At 0.6 mmol/L NH2Cl, VSL#3-CM pre-
igure 10. The time course of hsp induction by the proteasomenhibitor MG132 is similar to that produced by VSL#3-conditionededia. Immunoblot analysis of hsp25 and hsp72 levels in YAMC cells
20 �g of protein per lane) after exposure to the proteasome inhibitorG132 (25 �mol/L) for the times indicated showed a time-dependent
ncrease in hsp expression that paralleled that seen with VSL#3-reated cells. (First 2 lanes) C, untreated control harvested at 0 hours;, dimethyl sulfoxide vehicle–treated control harvested at 14 hours;S, heat-shocked cells (positive control). MG132 was even more
ffective than heat shock at inducing hsp25.
tprow
VcwffnawNdtocd1GwciolFagu
cstptatm
lpoIpaiurow(tp
amj
Fpspw( . Ima
1484 PETROF ET AL. GASTROENTEROLOGY Vol. 127, No. 5
reatment resulted in a mild but statistically significantrotective effect, decreasing the NH2Cl-stimulated 51Crelease and improving epithelial cell viability in the facef oxidant injury by approximately one third comparedith controls treated with NH2Cl alone (P .05).As another functional readout, the ability of
SL#3-CM treatment to protect epithelial cells againstytoskeletal damage from oxidant stress was determinedith F and G actin assays.44 F actin performs important
unctions involved in the maintenance of cellular scaf-olding and shape and acts as an anchoring point forumerous integral membrane proteins. Nevertheless, thectin cytoskeleton is particularly vulnerable to injury,hich can result in cellular compromise. Exposure toH2Cl causes rapid dissociation of F actin.31 Hence, we
etermined whether VSL#3-CM treatment would pro-ect the integrity of cytoskeletal F actin in the face ofxidant stress. NH2Cl treatment alone (0.6 mmol/L)auses a disruption of F-actin filaments, as shown by aecrease in F actin and an increase in G actin (Figure1B). By itself, VSL#3-CM has little effect on the F and
actin distribution. However, YAMC cells pretreatedith VSL#3-CM before NH2Cl showed significantly less
hange in the F and G actin distribution, again indicat-ng that this probiotic exerts some protection againstxidant injury. As positive and negative controls, phal-oidin (which binds and stabilizes the barbed ends of-actin filaments, thus increasing the amount of F actin)nd cytochalasin D (an F actin–disrupting agent thatreatly increases the amount of G actin) were used (Fig-
igure 11. Probiotic-conditioned medium protects epithelial cells agarotects YAMC cells from oxidant injury. YAMC cells were treated withtimulated with monochloramine (NH2Cl; 0.6 mmol/L) for 60 minutesSE for 3 separate experiments; each experiment in each group was
revents oxidant-induced actin depolymerization from F to G actin. YAMere then treated with the oxidant NH2Cl (0.6 mmol/L; 60 minutes) a
G) and filamentous (F) actin as described in Materials and Methods
re 11B). t
Discussion
This report, which describes intestinal epithelialell proteasome inhibition by probiotic bacteria, repre-ents a novel paradigm of microbial–epithelial cell in-eraction. We show that the beneficial effects of somerobiotics on the gut mucosa may be due to a combina-ion of decreased inflammation via blockade of NF-�Bctivity and increased cytoprotection through hsp induc-ion, both of which are likely mediated through a com-on link stemming from inhibition of the proteasome.Current models of IBD suggest that there is disequi-
ibrium between the cytoprotective and wound-healingrocesses and the inflammatory pathways, the net resultf which culminates in damage to the intestinal mucosa.t has been shown by others that administration of theroteasome inhibitor MG132 increases production of hspnd that HSF-1 is essential for the expression of induc-ble hsps on proteasome inhibition; e.g., hsp72 up-reg-lation will not occur in the absence of HSF-1.45 Ouresults confirmed not only that VSL#3-CM was capablef inducing hsp expression, but also that hsp inductionas occurring, at least in part, at the transcriptional level
Figure 9) through activation of HSF-1. This is consis-ent with what has been reported by others studyingroteasome inhibition.The cytoprotective effects of Hsps on epithelial cells
re well recognized,18,21,22,30,31,46 and VSL#3-CM treat-ent confers a degree of protection against oxidant in-
ury (Figure 11), most likely through its ability to induce
oxidant stress. (A) Chromium release assay showing that VSL#3-CM#3-CM for 16 hours. Cells were labeled with 51Cr for 60 minutes andthe ratio of released 51Cr to intracellular 51Cr was determined (meanormed in triplicate; *P .05 compared with controls). (B) VSL#3-CMlls were treated with VSL#3-CM for 16 hours, when appropriate, andwith untreated control (Con) cells. Cells were processed for globularges shown are representative of 3 separate experiments.
instVSL
, andperfC ce
long
he expression of hsps. Hsp 72 binds and stabilizes

capbseebscs
pchbdnVbpVeadHtatt
tn�nhsueoIiomtNdwucaa
tpCT
eVd(ciip(aa(ihntebo
pacVtlirtsfmsletoawwstleio
November 2004 PROBIOTICS IN COLONIC EPITHELIAL CELLS 1485
ritical cellular proteins, thus preventing their denatur-tion, and by chromium release assay has been shown torotect other intestinal epithelial cell lines against injuryy NH2Cl.31 Hsp25 is an actin-stabilizing agent, pre-erves cytoskeletal and tight junction functions, andxerts its cytoprotective effects more through cytoskel-tal stabilization by binding to actin filaments and sta-ilizing F actin.47 The results of the F/G actin assayuggest that VSL#3-CM confers a protective effect onytoskeletal integrity that is compatible with the actin-tabilizing functions of hsp25.
Clearly, the induction of hsps bestows a protection andreservation of functions vital to intestinal epithelialells. However, we hypothesized that the induction ofsps alone may not be sufficient to explain the observedeneficial effects of VSL#3 during colitis. Although in-uction of hsps would provide cytoprotection, it mightot fully explain the decrease in inflammation caused bySL#3 that has been reported by others.17,48,49 It shoulde noted that other cytoprotective effects mediated byathways independent of hsps may also be operative inSL#3-treated cells. In addition to its cytoprotective
ffects, VSL#3-CM inhibits NF-�B activation (Figure 1),nd pathways involving Akt-mediated cytoprotection in-uced by probiotics have been reported previously.50
ence, the cytoprotective effects seen with VSL#3-reated cells may be due not only to Hsp induction butlso to activation of other cytoprotective NF-�B geneargets and pathways, which in turn may be contributingo the observed cytoprotective effects reported here.
We have shown that VSL#3-CM inhibits NF-�B ac-ivation (Figure 1) and is capable of inhibiting endoge-ous immune response genes normally activated by NF-B, such as MCP-1 (Figure 2). Although otheronpathogenic bacteria that inhibit NF-�B through in-ibition of I�B degradation have been described, they doo by directly modulating ubiquitin ligase activity andse a type III secretion mechanism to interact with thepithelial cell.37 Because our studies show an attenuationf NF-�B activation in the absence of live bacteria, a typeII secretion mechanism is not implicated. Furthermore,t has been shown that cellular ubiquitination decreasesn direct-contact treatment with nonpathogenic S typhi-urium, which in turn results in decreased degradation of
he NF-�B inhibitor I�B and, hence, attenuation of theF-�B response.37 In our case, I�B degradation was also
ecreased (Figure 3), but through a different mechanism,hich, along with evidence that certain cellular pools ofbiquitinated proteins are increased (Figure 4), impli-ates proteasome inhibition as the major mechanism ofction. This was further confirmed by the proteasome
ssay (Figure 5). On the basis of our data, we speculate phat a portion of phosphorylated I�B� remains com-lexed to NF-�B in epithelial cells treated with VSL#3-M, which prevents subsequent activation of NF-�B byNF-�.The temporal data from the proteasome time course
xperiments suggest that proteasome inhibition bySL#3-CM is an early event that occurs almost imme-iately, long before hsp protein induction is detectedcompare the time course in Figure 6 with the timeourse in Figure 8A). Consistent with this is the fact thatnhibition of the degradation of other proteins importantn the NF-�B inflammatory response (such as I�B� and105) occurs after only 1 hour of exposure to VSL#3-CMFigure 7). MG132, a potent proteasome inhibitor thatcts on the proteasome within minutes, was tested for itsbility to induce hsps and was found to take several hoursFigure 10), again suggesting that proteasome inhibitions an early event that occurs long before the induction ofsps. Although we recognize that these correlations doot provide conclusive proof, taken together they suggesthat proteasome inhibition by VSL#3-CM is an earlyvent that results in the inhibition of NF-�B throughlockade of I�B degradation and that hsp inductionccurs afterward as a later event.
It is interesting to note that, unlike the effects of theroteasome inhibitor MG132, which results in massiveccumulation of ubiquitinated proteins and ultimately inell death, the increase in ubiquitination caused bySL#3 treatment is restricted to only a select few pro-
eins and, furthermore, that VSL#3 does not cause cel-ular toxicity. This suggests that VSL#3 does not resultn large-scale inhibition of proteasome function butather acts by specifically inhibiting only particular fea-ures of proteasome function, leaving the rest of protea-ome functions intact. In support of this hypothesis is theact that, although VSL#3-CM strongly inhibits the chy-otrypsin-like activity on the proteasome assay, it pos-
esses only weak activity against the caspase-like proteo-ytic function of the proteasome and has no inhibitoryffect on its trypsin-like activity. This would suggesthat it does not globally inhibit all proteolytic functionsf the proteasome but rather displays some specificity orffinity for some functions more than others. Treatmentith VSL#3-CM is well tolerated by the epithelial cells,hich is not the case with most of the synthetic protea-
ome inhibitors. We speculate that VSL#3-CM is not asoxic to cells as is the synthetic inhibitor MG132 oractacystin, for example, likely as a result of this differ-ntial affinity, which likely allows some normal function-ng of the proteasome to continue while the degradationf certain proteins such as I�B is blocked. This may in
art explain why the pattern of accumulated ubiquiti-
nmIfiPnbeubtmptsmsm
itsmgtmftcrbtab
1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
1486 PETROF ET AL. GASTROENTEROLOGY Vol. 127, No. 5
ated proteins is different from what is observed with theore powerful and toxic synthetic proteasome inhibitors.
n this respect, VSL#3 may act in a similar but distinctashion from the peptide PR39, a selective proteasomenhibitor isolated from pig intestine.39 Administration ofR39 results in accumulation of only select ubiquiti-ated proteins and results in less toxicity than MG132,ut in contrast to VSL#3, PR39 has no effect on hspxpression. This mammalian peptide, PR39, is up-reg-lated in areas of inflammation and blocks inflammationy inhibiting the degradation of I�B� through interac-ion with the �7 subunit of the proteasome, but it leavesost other proteasome functions unaffected.39 Whether
robiotic-derived proteasome inhibitors act similarlyhrough the targeted binding and inhibition of specificubunits of the proteasome still remains to be deter-ined, and efforts are currently under way to purify the
ecreted products of the probiotic to further study theirechanism of proteasome inhibition.It is increasingly apparent that microbial–epithelial
nteractions are extremely important in maintaining in-estinal functional integrity. Microarray analysis hashown that exposure of intestinal epithelial cells to com-ensal bacteria results in up-regulation of a vast array of
enes involved in functions as diverse as host lipid me-abolism, angiogenesis, micronutrient absorption, gutotility, and maintenance of intestinal epithelial barrier
unction.51 By inhibiting epithelial cell proteasome ac-ivity, VSL#3 displays a novel mechanism by whicholonizing nonpathogenic probiotic bacteria are able toegulate epithelial cell responses to their environment tooth increase cytoprotection and decrease inflammation,hus providing a net benefit for the intestinal mucosa andlikely mechanism for the therapeutic effects of these
acteria described in the clinical setting.
References1. Gionchetti P, Rizzello F, Venturi A, Campieri M. Probiotics in
infective diarrhoea and inflammatory bowel diseases. J Gastro-enterol Hepatol 2000;15:489–493.
2. Niedzielin K, Kordecki H, Birkenfeld B. A controlled, double-blind,randomized study on the efficacy of Lactobacillus plantarum299V in patients with irritable bowel syndrome. Eur J Gastroen-terol Hepatol 2001;13:1143–1147.
3. Gionchetti P, Rizzello F, Venturi A, Brigidi P, Matteuzzi D, Bazzoc-chi G, Poggioli G, Miglioli M, Campieri M. Oral bacteriotherapy asmaintenance treatment in patients with chronic pouchitis: a dou-ble-blind, placebo-controlled trial. Gastroenterology 2000;119:305–309.
4. Gionchetti P, Rizzello F, Helwig U, Venturi A, Lammers KM, BrigidiP, Vitali B, Poggioli G, Miglioli M, Campieri M. Prophylaxis ofpouchitis onset with probiotic therapy: a double-blind, placebo-controlled trial. Gastroenterology 2003;124:1202–1209.
5. Isolauri E, Juntunen M, Rautanen T, Sillanaukee P, Koivula T. A
human Lactobacillus strain (Lactobacillus casei sp strain GG)promotes recovery from acute diarrhea in children. Pediatrics1991;88:90–97.
6. Majamaa H, Isolauri E, Saxelin M, Vesikari T. Lactic acid bacteriain the treatment of acute rotavirus gastroenteritis. J Pediatr Gas-troenterol Nutr 1995;20:333–338.
7. Arvola T, Laiho K, Torkkeli S, Mykkanen H, Salminen S, MaunulaL, Isolauri E. Prophylactic Lactobacillus GG reduces antibiotic-associated diarrhea in children with respiratory infections: a ran-domized study. Pediatrics 1999;104:e64.
8. Kullisaar T, Zilmer M, Mikelsaar M, Vihalemm T, Annuk H, KairaneC, Kilk A. Two antioxidative lactobacilli strains as promising pro-biotics. Int J Food Microbiol 2002;72:215–224.
9. Annuk H, Shchepetova J, Kullisaar T, Songisepp E, Zilmer M,Mikelsaar M. Characterization of intestinal lactobacilli as putativeprobiotic candidates. J Appl Microbiol 2003;94:403–412.
0. Ocana VS, Pesce de Ruiz Holgado AA, Nader-Macias ME. Selec-tion of vaginal H2O2-generating Lactobacillus species for probi-otic use. Curr Microbiol 1999;38:279–284.
1. Cleveland J, Montville TJ, Nes IF, Chikindas ML. Bacteriocins:safe, natural antimicrobials for food preservation. Int J FoodMicrobiol 2001;71:1–20.
2. Paraje MG, Albesa I, Eraso AJ. Conservation in probiotic prepa-rations of Lactobacillus with inhibitory capacity on other species.New Microbiol 2000;23:423–431.
3. Braude AI, Siemienski JS. The influence of bacteriocins on resis-tance to infection by gram-negative bacteria. II. Colicin action,transfer of colicinogeny, and transfer of antibiotic resistance inurinary infections. J Clin Invest 1968;47:1763–1773.
4. Lee YK, Lim CY, Teng WL, Ouwehand AC, Tuomola EM, SalminenS. Quantitative approach in the study of adhesion of lactic acidbacteria to intestinal cells and their competition with enterobac-teria. Appl Environ Microbiol 2000;66:3692–3697.
5. Lee YK, Puong KY, Ouwehand AC, Salminen S. Displacement ofbacterial pathogens from mucus and Caco-2 cell surface bylactobacilli. J Med Microbiol 2003;52:925–930.
6. Maassen CB, van Holten-Neelen C, Balk F, den Bak-GlashouwerMJ, Leer RJ, Laman JD, Boersma WJ, Claassen E. Strain-depen-dent induction of cytokine profiles in the gut by orally adminis-tered Lactobacillus strains. Vaccine 2000;18:2613–2623.
7. Madsen K, Cornish A, Soper P, McKaigney C, Jijon H, YachimecC, Doyle J, Jewell L, De Simone C. Probiotic bacteria enhancemurine and human intestinal epithelial barrier function. Gastro-enterology 2001;121:580–591.
8. Chang E. Intestinal epithelial function and response to mucosalinjury in inflammatory bowel diseases. In: Kirsner J, ed. Inflam-matory bowel diseases. 5th ed. Philadelphia: Saunders, 1999:3–19.
9. Podolsky DK. Inflammatory bowel disease. N Engl J Med 2002;347:417–429.
0. Schmitz H, Barmeyer C, Fromm M, Runkel N, Foss HD, BentzelCJ, Riecken EO, Schulzke JD. Altered tight junction structurecontributes to the impaired epithelial barrier function in ulcerativecolitis. Gastroenterology 1999;116:301–309.
1. Ropeleski MJ, Tang J, Walsh-Reitz MM, Musch MW, Chang EB.Interleukin-11-induced heat shock protein 25 confers intestinalepithelial-specific cytoprotection from oxidant stress. Gastroen-terology 2003;124:1358–1368.
2. Urayama S, Musch MW, Retsky J, Madonna MB, Straus D, ChangEB. Dexamethasone protection of rat intestinal epithelial cellsagainst oxidant injury is mediated by induction of heat shockprotein 72. J Clin Invest 1998;102:1860–1865.
3. Neurath MF, Becker C, Barbulescu K. Role of NF-kappaB inimmune and inflammatory responses in the gut. Gut 1998;43:856–860.
4. Jobin C, Sartor RB. NF-kappaB signaling proteins as therapeutictargets for inflammatory bowel diseases. Inflamm Bowel Dis
2000;6:206–213.
2
2
2
2
2
3
3
3
3
3
3
3
3
3
3
4
4
4
4
4
4
4
4
4
4
5
5
oMu
tdtsKAeR
November 2004 PROBIOTICS IN COLONIC EPITHELIAL CELLS 1487
5. Schmid RM, Adler G. NF-kappaB/rel/IkappaB: implications ingastrointestinal diseases. Gastroenterology 2000;118:1208–1228.
6. Boone DL, Lee EG, Libby S, Gibson PJ, Chien M, Chan F, MadoniaM, Burkett PR, Ma A. Recent advances in understanding NF-kappaB regulation. Inflamm Bowel Dis 2002;8:201–212.
7. Neurath MF, Pettersson S, Meyer zum Buschenfelde KH, StroberW. Local administration of antisense phosphorothioate oligonu-cleotides to the p65 subunit of NF-kappa B abrogates estab-lished experimental colitis in mice. Nat Med 1996;2:998–1004.
8. Neurath MF, Fuss I, Schurmann G, Pettersson S, Arnold K, Muller-Lobeck H, Strober W, Herfarth C, Buschenfelde KH. Cytokinegene transcription by NF-kappa B family members in patients withinflammatory bowel disease. Ann N Y Acad Sci 1998;859:149–159.
9. Parsell DA, Lindquist S. The function of heat-shock proteins instress tolerance: degradation and reactivation of damaged pro-teins. Annu Rev Genet 1993;27:437–496.
0. Musch MW, Ciancio MJ, Sarge K, Chang EB. Induction of heatshock protein 70 protects intestinal epithelial IEC-18 cells fromoxidant and thermal injury. Am J Physiol 1996;270:C429–C436.
1. Musch MW, Sugi K, Straus D, Chang EB. Heat-shock protein 72protects against oxidant-induced injury of barrier function of hu-man colonic epithelial Caco2/bbe cells. Gastroenterology 1999;117:115–122.
2. Whitehead RH, VanEeden PE, Noble MD, Ataliotis P, Jat PS.Establishment of conditionally immortalized epithelial cell linesfrom both colon and small intestine of adult H-2Kb-tsA58 trans-genic mice. Proc Natl Acad Sci U S A 1993;90:587–591.
3. Kojima K, Musch MW, Ren H, Boone DL, Hendrickson BA, Ma A,Chang EB. Enteric flora and lymphocyte-derived cytokines deter-mine expression of heat shock proteins in mouse colonic epithe-lial cells. Gastroenterology 2003;124:1395–1407.
4. Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH,Provenzano MD, Fujimoto EK, Goeke NM, Olson BJ, Klenk DC.Measurement of protein using bicinchoninic acid. Anal Biochem1985;150:76–85.
5. Reinecker HC, Loh EY, Ringler DJ, Mehta A, Rombeau JL, Mac-Dermott RP. Monocyte-chemoattractant protein 1 gene expres-sion in intestinal epithelial cells and inflammatory bowel diseasemucosa. Gastroenterology 1995;108:40–50.
6. MacDermott RP. Chemokines in the inflammatory bowel dis-eases. J Clin Immunol 1999;19:266–272.
7. Neish AS, Gewirtz AT, Zeng H, Young AN, Hobert ME, Karmali V,Rao AS, Madara JL. Prokaryotic regulation of epithelial responsesby inhibition of IkappaB-alpha ubiquitination. Science 2000;289:1560–1563.
8. Voges D, Zwickl P, Baumeister W. The 26S proteasome: a mo-lecular machine designed for controlled proteolysis. Annu RevBiochem 1999;68:1015–1068.
9. Gao Y, Lecker S, Post MJ, Hietaranta AJ, Li J, Volk R, Li M, SatoK, Saluja AK, Steer ML, Goldberg AL, Simons M. Inhibition ofubiquitin-proteasome pathway-mediated I kappa B alpha degra-dation by a naturally occurring antibacterial peptide. J Clin Invest
2000;106:439–448.0. Sears C, Olesen J, Rubin D, Finley D, Maniatis T. NF-kappa Bp105 processing via the ubiquitin-proteasome pathway. J BiolChem 1998;273:1409–1419.
1. Ghosh S, Karin M. Missing pieces in the NF-kappaB puzzle. Cell2002;109(suppl):S81–S96.
2. Beck SC, Paidas CN, Mooney ML, Deitch EA, De Maio A. Pres-ence of the stress-inducible form of hsp-70 (hsp-72) in normal ratcolon. Shock 1995;3:398–402.
3. Pirkkala L, Alastalo TP, Zuo X, Benjamin IJ, Sistonen L. Disruptionof heat shock factor 1 reveals an essential role in the ubiquitinproteolytic pathway. Mol Cell Biol 2000;20:2670–2675.
4. White JR, Naccache PH, Sha’afi RI. Stimulation by chemotacticfactor of actin association with the cytoskeleton in rabbit neutro-phils. Effects of calcium and cytochalasin B. J Biol Chem 1983;258:14041–14047.
5. Kawazoe Y, Nakai A, Tanabe M, Nagata K. Proteasome inhibitionleads to the activation of all members of the heat-shock-factorfamily. Eur J Biochem 1998;255:356–362.
6. Liu TS, Musch MW, Sugi K, Walsh-Reitz MM, Ropeleski MJ,Hendrickson BA, Pothoulakis C, Lamont JT, Chang EB. Protectiverole of HSP72 against Clostridium difficile toxin A-induced intes-tinal epithelial cell dysfunction. Am J Physiol Cell Physiol 2003;284:C1073–C1082.
7. Mounier N, Arrigo AP. Actin cytoskeleton and small heat shockproteins: how do they interact? Cell Stress Chaperones 2002;7:167–176.
8. Lammers KM, Helwig U, Swennen E, Rizzello F, Venturi A, Cara-melli E, Kamm MA, Brigidi P, Gionchetti P, Campieri M. Effect ofprobiotic strains on interleukin 8 production by HT29/19A cells.Am J Gastroenterol 2002;97:1182–1186.
9. Ulisse S, Gionchetti P, D’Alo S, Russo FP, Pesce I, Ricci G,Rizzello F, Helwig U, Cifone MG, Campieri M, De Simone C.Expression of cytokines, inducible nitric oxide synthase, andmatrix metalloproteinases in pouchitis: effects of probiotic treat-ment. Am J Gastroenterol 2001;96:2691–2699.
0. Yan F, Polk DB. Probiotic bacterium prevents cytokine-inducedapoptosis in intestinal epithelial cells. J Biol Chem 2002;277:50959–50965.
1. Hooper LV, Wong MH, Thelin A, Hansson L, Falk PG, Gordon JI.Molecular analysis of commensal host-microbial relationships inthe intestine. Science 2001;291:881–884.
Received November 24, 2003. Accepted August 12, 2004.Address requests for reprints to: Eugene B. Chang, MD, Department
f Medicine, The University of Chicago, 5841 S. Maryland Avenue,C6084, Chicago, Illinois 60637. e-mail: [email protected]; fax: (773) 702-2281.Supported by National Institutes of Health grant DK47722, Diges-
ive Disease Research Core Center grant DK42086, the Broad Foun-ation, the Crohn’s and Colitis Foundation of America, and the Gas-rointestinal Research Foundation of Chicago (to E.B.C.). E.O.P. isupported by National Institutes of Health grant K08 DK064840-01..K. is supported by a grant from the Crohn’s and Colitis Foundation ofmerica. M.J.R. is supported by the Canadian Association of Gastro-nterology, Novartis Canada, and the Canadian Institutes for Healthesearch.
A special thanks to Dr. Sanford Grossman for his support.



















