
Compound Probiotics Improve the Diarrhea Rate and ... - MDPI
Upload
independentCategory
view
5download
0
International Journal of Food Sciences and Nutrition,August 2010; 61(5): 473–496
Cancer-preventing attributes of probiotics: an update
MANOJ KUMAR1, ASHOK KUMAR2, RAVINDER NAGPAL3, DHEERAJMOHANIA4, PRADIP BEHARE1, VINOD VERMA5, PRAMODKUMAR1,DEV PODDAR1, P. K. AGGARWAL1, C. J. K. HENRY6, SHALINI JAIN7 &HARIOM YADAV7
1Dairy Microbiology Division, National Dairy Research Institute, Karnal (Haryana), India,2Department of Zoology, M.L.K. P.G. College, Balrampur (U.P.), India, 3Department ofBiotechnology, Lovely Professional University, Phagwara, Punjab, India, 4Animal BiochemistryDivision, National Dairy Research Institute, Karnal (Haryana), India, 5AgResearch Ltd,Ruakura Research Centre, Reproductive Technologies, East Street, Hamilton, New Zealand,6School of Biological and Molecular Sciences, Oxford Brookes University, Oxford, UK, and7National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health,Bethesda, Maryland, USA
AbstractCancer is a serious global public health problem. Cancer incidence and mortality have beensteadily rising throughout the past century in most places of the world. There are severalepidemiological evidences that support a protective role of probiotics against cancer. Lactic acidbacteria and their probioactive cellular substances exert many beneficial effects in the gastro-intestinal tract, and also release various enzymes into the intestinal lumen and exert potentialsynergistic (LAB) effects on digestion and alleviate symptoms of intestinal malabsorption.Consumption of fermented dairy products with LAB may elicit anti-tumor effects. These effectsare attributed to the inhibition of mutagenic activity, the decrease in several enzymes implicatedin the generation of carcinogens, mutagens, or tumor-promoting agents, suppression of tumors,and epidemiology correlating dietary regimes and cancer. Specific cellular components in lacticacid bacteria seem to induce strong adjuvant effects including modulation of cell-mediatedimmune responses, activation of the reticulo-endothelial system, augmentation of cytokinepathways, and regulation of interleukins and tumor necrosis factors. Studies on the effect ofprobiotic consumption on cancer appear promising, since recent in vitro and in vivo studies haveindicated that probiotic bacteria might reduce the risk, incidence and number of tumors of thecolon, liver and bladder. The protective effect against cancer development may be ascribed tobinding of mutagens by intestinal bacteria, may suppress the growth of bacteria that convertprocarcinogens into carcinogens, thereby reducing the amount of carcinogens in the intestine,reduction of the enzymes b-glucuronidase and b-glucosidase and deconjugation of bile acids, ormerely by enhancing the immune system of the host. There are isolated reports citing thatadministration of LAB results in increased activity of anti-oxidative enzymes or by modulatingcirculatory oxidative stress that protects cells against carcinogen-induced damage. Theseinclude glutathione-S-transferase, glutathione, glutathione reductase, glutathione peroxidase,superoxide dismutase and catalase. However, there is no direct experimental evidence forcancer suppression in human subjects as a result of the consumption of probiotic cultures in
Correspondence: Dr Hariom Yadav, National Institute of Diabetes and Digestive and Kidney Diseases, Diabetes Branch,5W5872, CRC, Bld 10, Bethesda, MD 20892, USA. E-mail: [email protected]
ISSN 0963-7486 print/ISSN 1465-3478 online � 2010 Informa UK LtdDOI: 10.3109/09637480903455971
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

fermented or unfermented dairy products, but there is a wealth of indirect evidence based largelyon laboratory studies.
Keywords: Probiotics, cancer, prebiotics, synbiotics, functional food
Introduction
Probiotics are defined as live microbial food ingredients (supplements) that conferbeneficial health effects to the host upon ingestion in adequate amounts (Salminen andvon Wright 1998). The concept of probiotics evolved at the turn of the twentiethcentury from a hypothesis first proposed by Nobel Prize-winning Russian scientist ElieMetchnikoff, who propounded that the long and healthy life of Bulgarian peopleresulted from their consumption of fermented milk products (Bibel 1998). He believedthat, when consumed, the fermenting bacillus (Lactobacillus) positively influenced themicrobiota of the colon and, decreased their toxic activities. Several lactic acid bacteriamay help prevent initiation of cancer (Table I). It appears that LAB can reduce thelevels of colon enzymes that convert pro-carcinogens to carcinogens. Specifically, LABcan reduce levels of the enzymes b-glucuronidase, nitro-reductase, and azo-reductase.LAB may also be involved in the direct reduction of pro-carcinogens; for example, bytaking up nitrites and by reducing the levels of secondary bile salts (Fernandes andShahani 1990). Changes in enzyme activity in humans have been observed withLactobacillus acidophilus and Bifidobacterium bifidum (Marteau et al. 1990), andLactobacillus rhamnosus GG (LGG) (Goldin and Gorbach 1984). Animal studiesshowed fewer tumors in those exposed to a carcinogen in the presence of LGGcompared with the animals exposed to the carcinogen without the benefit of LGG(Goldin 1996). Human epidemiological reports have indicated that populationsingesting fermented dairy products have a decreased risk of colon cancer (Kampmanet al. 1994). An anti-tumor effect has been reported by oral intake of LAB in in vitrostudies but, since colon carcinogenesis is a multistage process, the mechanisms ifvalidated remain to be elucidated (Hirayama and Rafter 1999). A preventive effect onmalignancy development could be mediated by different mechanisms involved indetoxification of carcinogens.
In vitro studies have shown that, in general, live cells of probiotic bacteria possessedhigher anti-mutagenic activity. Intake of fermented milk by (L. acidophilus andB. bifidum) influenced gut flora enzymes, like b-glucuronidase, b-glucosidase andnitro-reductase (Wollowski et al. 2001). L. acidophilus and B. bifidum decreased theactivity of nitro-reductase and b-glucuronidase but increased that of b-glucosidase(Marteau et al. 1990). This could be an advantage since b-glucosidase may releaseflavonoids that have anti-mutagenic, anti-oxidative and immune-stimulatory effects(Stoner and Mukhtar 1995). Most animal and human studies do indicate that feedingcertain LAB decreases fecal enzyme levels that may be involved in formation ofcarcinogens. Also, beneficial effects can be attributed to immune-potentiating effectsby LAB strains. One specific effect was shown by heat-killed Lactobacillus plantarumL-137, which restored the inhibited IL-12 production in DBA/2 mice with tumors(Murosaki et al. 2000). The anti-tumor effects were found to be due to the activation ofmacrophage by LC9018 (Okawa et al. 1993). Conjugated linoleic acid has beenhighlighted recently because of its effect to reduce carcinogenesis, atherosclerosisand body fats (Chin et al. 1992). Mycotoxins are well-known contaminants of grainsand other food raw materials in which Ochratoxin A is carcinogenic, genotoxic,
474 M. Kumar et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

immunosuppressive and nephrotoxic (Petzinger and Ziegler 2000). In some reports,strains of Lactobacillus and Lactococcus were shown to be resistant to ochratoxinA (5–10 microgram/disc), and just one strain of L. helveticus was sensitive (0.1microgram/disc) (Piotrowska and Zakowska 2005). Furthermore, all strains testedreduced the amount of ochratoxin A in the growth medium as measured after 120 h. L.acidophilus CH-5, LGG and a couple of other strains reduced the initial level ofochratoxin A by more than 50%. Intake of such LAB may potentially reduce carci-nogenic/genotoxic effects of consumption of food contaminated by ochratoxin A.Aflatoxin B1 (AFB1), a mycotoxin produced by the common fungi Aspergillus flavusand Aspergillus parasiticus, is an established human hepato-carcinogen (Vamio et al.1993). Various strains of LAB isolated from either dairy products or healthy humanmicrobiota showed that two probiotic L. rhamnosus strains, LGG and LC705, werefound to be highly efficient in binding a range of mycotoxins, including AFB1. Earlier,a single viable bacterium was found to bind > 107 AFB1 molecules (Haskard et al.2000).
Protective and prevention of cancer occurrence
Liver cancer is the sixth most commonly diagnosed cancer in the world, and the thirdmost common cause of death from cancer, according to Cancer Research UK. Despite
Table I. Potential probiotic bacteria having cancer-preventing properties.
Probiotic bacteria References
B. polyfermenticus Jeong et al. (2003), Park et al. (2004, 2007)B. longum ATCC 15708 Challa et al. (1997), Vanderhoof (2001), Biasco et al. (1991), Pool-Zobel
et al. (1996), Rowland et al. (1998), Reddy and Rivenson (1993), Singhet al. (1997), Reddy (1998), Haskard et al. (2000), Lin and Chang (2000)
Lactobacillus GG Goldin et al. (1996), Drisko et al. (2003), Femia et al. (2002), Caro et al.(2005), Ahotupa et al. (1996), Fang and Polk (2002), Lam et al. (2007)
L. casei (LC9018) Aso et al. (1995), Lidbeck et al. (2000), Okawa et al. (1993)L. casei Shirota Matsumoto et al. (2009)L. acidophilus Lidbeck et al. (1992), Biffi et al. (1997),Vanderhoof (2001), Biasco et al.
(1991), Pool-Zobel et al. (1996), Arimochi et al. (1997), Goldin et al.(1980), Haskard et al. (2000)
L. gasseri Vanderhoof (2001), Pool-Zobel et al. (1996)L. confusus, Vanderhoof (2001), Pool-Zobel et al. (1996)S. thermophilus Vanderhoof (2001), Pool-Zobel et al. (1996)B. breve ATCC 15700 Vanderhoof (2001), Pool-Zobel et al. (1996), Onoue et al.
(1997), Chalova et al. (2008)L. delbrueckeii Rafter (1995)B. infantis Biffi et al. (1997)B. lactis Femia et al. (2002), Sekine et al. (1995)L. plantarum L-137,L. plantarum JCM 1551,L. plantarum KLAB21,L. plantarum 2592
Murosaki et al. (2000), Ando et al. (2004), Rhee and Park,(2001), Kruszewska et al. (2002), Khanafari et al. (2007)
P. pentosaceus 16:1 Kruszewska et al. (2002)L. paracasei F19 Kruszewska et al. (2002)L. reuteri Iyer et al. (2008)B. adoleascentis ATCC 15703 Chalova et al. (2008)VSL#3 Otte et al. (2009)
Cancer-preventing attributes of probiotics 475
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

these figures, the cancer remains relatively rare, with 18,500 new cases in the USAevery year, and about 3,000 in the UK. The highest incidences of the disease are in eastand southeast Asia, particularly China, and for this reason the current researcherslooked at the effects of probiotic supplements on markers for the disease. The biggestrisk factor for the disease is said to be chronic hepatitis B virus infection, butconsumption of foods (contaminated with aflatoxins) are also established causes ofliver cancer.
AFB1 is classified as a group I carcinogen in humans by International Agency forResearch on Cancer. However, the exact mechanisms of AFB1 hepatocarcinogenesishave not been fully elucidated. But some studies have suggested that oncogenes arecritical molecular targets for AFB1, and AFB1 causes characteristic genetic changes inthe p53 tumor suppressor gene and ras proto-oncogenes. The hepatic carcinogenAFB1 is metabolized in the liver by at least four different P450s, all of which exhibitlarge inter-individual differences in the expression levels. These differences could affectthe individual risk of hepatocellular carcinoma. Some researchers provided additionalevidence that reactive oxygen species (ROS) and oxidative DNA damage may beinvolved in AFB1-induced p53 and ras mutations. AFB1–glutathione (GSH) conju-gation is the major pathway for the detoxification of aflatoxin metabolites (Figure 1).This reaction is catalyzed by glutathione-S-transferase (GST) and plays a major role inmodulation of AFB1 adduct formation to nuclear DNA. Changes recorded in hepaticGST activity during development of rats can alter the balance between AFB1–GSHconjugation and AFB1–DNA adduct formation. Some probiotic bacterial strainssuccessfully bind mycotoxins; L. rhamnosus binds away AFB1 in vivo, thus reducingbio-absorption of the toxin from the gut, while L. acidophilus and Bifidobacterium longumneutralize AFB1 and AFM1 by a similar binding mechanism (Haskard et al. 2000).Epidemiological studies in Africa and Asia (where high levels of exposure occur) linkAFB1 exposure and hepato-cellular carcinomas (Bulatao-Jayme et al. 1982; Bosch andMunoz 1988), and the risk is synergistic with hepatitis viral infections. The mechanismof action for AFB1 mutagenicity begins with metabolic activation by CYP3A4,CYP3A5 and/or CYP1A2 (Garner et al. 1972; Reynolds et al. 1987) that forms anexo-8,9-epoxide and subsequent adduct formation and DNA damage (Essigmann et al.
Aspergillus spp.
Aflatoxin B1
Aflatoxin-exo8,9-epoxide
CYP 1A2CYP 3A4
DNA
Alatoxin-N7-Guanine
Genotoxicity
Aflatoxin-S-G
AFB1-mercapturate
Aflatoxin-dihydrodiol
Aflatoxin dialdehyde
Aflatoxin-Albumin
Inhibition-Probioticby binding mechanism
Detoxification by antioxidantsproduced by probiotics
Figure 1. Mechanism of action of probiotics during liver cancer progression induced by AFB1.
476 M. Kumar et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

1977; Iyer et al. 1994). This damage has been shown in vitro to cause guaninenucleotide substitutions (Lilleberg et al. 1992) specifically to codon 249 of the p53gene (Aguilar et al. 1993). Mutations in specific oncogenes and tumor-suppressorgenes (e.g. K-ras, p53) are involved in tumor development in the colon and liver, asthey occur at high frequencies in human colon and liver cancers (Erdman et al. 1997).The expression levels of these genes can be measured and used to demonstrate theeffect of various dietary components on tumor formation in colonic mucosa.
Addition of the probiotic B. longum to the diet of rats was shown to exert a stronganti-tumor activity on colonic mucosa by reducing the expression level of ras-p21expression and cell proliferation (Singh et al. 1997; Reddy 1998). LGG administrationdetermined the upregulation and downregulation of 334 and 92 genes, respectively,shown by microarray analysis, and further real-time polymerase chain reaction con-firmed the reliability of the analysis—it mainly affects the expression of genes involvedin immune response and inflammation (transforming growth factor-beta and TNFfamily members, cytokines, nitric oxide synthase 1, defensin alpha 1), apoptosis, cellgrowth and cell differentiation (cyclins and caspases, oncogenes), cell–cell signaling(intracellular adhesion molecules), cell adhesion (cadherins), signal transcription andtransduction (Caro et al. 2005). Colorectal cancer represents a major public healthproblem accounting for over 1 million cases and about 0.5 million deaths worldwide(Chau and Cunningham 2006). Survival from colon cancer at 5 years has been found tovary demographically and estimated to be 65% in North America, 54% in WesternEurope, 34% in Eastern Europe and 30% in India (Parkin et al. 2005). Dietaryinterventions and natural bioactive supplements have now been extensively studied toreduce the risks of colon cancer as a cause of prevention instead of cure. Postulatedmechanisms include reduction in the activity of several cancer-causing bacteria,des-mutagenic and anti-carcinogenic properties (Collins and Gibson 1999; Chauand Cunningham 2006). Probiotics have been found by several researchers to decreasefecal concentrations of enzymes and secondary bile salts, and to reduce absorption ofharmful mutagens that may contribute to colon carcinogenesis (Rafter 1995). Otherstudies suggest that normal intestinal flora can influence carcinogenesis by producingenzymes (glycosidase, B-glucuronidase, azoreductase, and nitroreductase) thattransform precarcinogens into active carcinogens (Goldin et al. 1980; Goldin1990; Marteau et al. 1990; Ling et al. 1994; Pedrosa et al. 1995). Certain probioticsmay protect the host from this activity. L. acidophilus and Lactobacillus casei supple-mentation in humans helped to decrease levels of these enzymes, as shown by fecalspecimens (Hayatsu and Hayatsu 1993; Lee and Salminen 1995; Lidbeck et al. 1992).In animal studies, the aforementioned bacterial enzymes have been suppressed withthe administration of LGG (Drisko et al. 2003). Other LAB have shown similarresults; however, the relationship between enzyme activity and cancer risk needsfurther investigation.
Several mechanisms have been proposed as to how LAB may inhibit colon cancer,including enhancing the host’s immune response, altering the metabolic activity of theintestinal microbiota, binding and degrading carcinogens, producing antimutageniccompounds, and altering the physiochemical conditions in the colon (Hirayama andRafter 2000) (Figure 2). During carcinogenesis in the colon, aberrant crypts can berecognized as early neoplastic lesions in both rodents and humans (Bird 1995).A number of natural chemopreventive agents or medicinal plants that inhibit thedevelopment of aberrant crypt foci (ACF) have been demonstrated to prevent colon
Cancer-preventing attributes of probiotics 477
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

cancer in rodents (Finley et al. 2000; Ohishi et al. 2002), thereby suggesting that ACFassays in the rodent colon can be employed as good biomarkers in colon carcinogen-esis. Colon carcinogenesis is also known to be a pathological consequence of persistentoxidative stress, resulting in DNA damage and mutations in cancer-associated genes,cycle arrest or cell death, and mutations and regeneration in which the cellularoverproduction of reactive oxygen molecules and reactive nitrogen species havebeen implicated (Payne et al. 1999; Bartsch and Nair 2002; Korenaga et al.2002; Nair et al. 2002) In an in vitro experiment, Bacillus polyfermenticus exertedantioxidative, antigenotoxic, and anticarcinogenic effects (Jeong et al. 2003; Park et al.2004). Consumption of large quantities of dairy products such as yoghurt andfermented milk containing Lactobacillus or Bifidobacterium may be related to a lowerincidence of colon cancer (Shahani and Ayebo 1980). Oral administration of LAB hasbeen shown to effectively reduce DNA damage, induced by chemical carcinogens, ingastric and colonic mucosa in rats. By the comet assay, L. acidophilus, Lactobacillusgasseri, Lactobacillus confusus, Streptococcus thermophilus, Bifidobacterium breve and B.longum were antigenotoxic toward N¢-nitro-N-nitrosoguanidine (MNNG) (Pool-Zobelet al. 1996). These bacteria were also protective toward 1,2-dimethylhydrazine(DMH)-induced genotoxicity. Metabolically active L. acidophilus cells, as well as anacetone extract of the culture, prevented MNNG-induced DNA damage, while heat-treated L. acidophilus was not antigenotoxic. Goldin et al. (1996) showed that a specificstrain of L. casei subsp. rhamnosus designated GG can interfere with the initiation orearly promotional stages of DMH-induced intestinal tumor genesis and that thiseffect is most pronounced for animals fed a high-fat diet. Feeding of L. acidophilusand B. longum suppressed the formation of ACF and tumor incidence, induced byazoxymethane (AOM) (Kulkarni and Reddy 1994; Arimochi et al. 1997; Singh et al.1997; Challa et al. 1997; Rowland et al. 1998) or DMH (Abdelali et al.
Steps leading to cancer
DMH (Procarcinogen)
Carcinogen (Azoxymethanemethyldiazoniumion)
Reactions with cellular targets (colonocytes)
Carcinogen-DNA adduct
Neoplastic manifestation
Cancer (Adenocarcinoma)
Probiotics bybinding mechanism
Probiotics byantioxidants
Inhibition and detoxification
Figure 2. Mechanism of action of probiotics during colon cancer progression.
478 M. Kumar et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

1995; Gallaher et al. 1996). In addition, it has been reported that colonization ofbacteria with an ability to produce genotoxic compounds and high b-glucuronidaseactivity enhanced progression of ACF induced by DMH in rats, and that the additionalcolonization of B. breve reduced the number of ACF with four or more crypts/focus andcrypt multiplicity that are reliable predictors of malignancy (Onoue et al. 1997). AOM-induced colon tumor development was also suppressed with a decrease in colonicmucosal cell proliferation and tumor ornithine decarboylase and ras-p21 activities(Hirayama and Rafter 2000). There was a report on the anti-tumorigenic activity ofthe prebiotic inulin, enriched with oligofructose, in combination with the probioticsL. rhamnosus and Bifidobacterium lactis in the AOM-induced colon carcinogenesis ratmodel (Femia et al. 2002). Reddy and Rivenson (1993) reported that lyophilizedcultures of B. longum administered in the diet to rats inhibited liver, colon andmammary tumors, induced by the food mutagen 2-amino-3-methyl-3H-imidazo(4,5-f) quinoline (IQ). Goldin and Gorbach (1980) showed that dietary supplementsof L. acidophilus not only suppressed the incidence of DMH-induced colon carcino-genesis but also increased the latency period in rats. Feeding of fermented milkincreased the survival rate of rats with chemically induced colon cancer (Shackelfordet al. 1983). B. polyfermenticus was also determined to protect against the DNA damageinduced by MNNG, a direct-acting carcinogen with alkylating properties in CHO-K1cells and in human lymphocytes, and was also determined to inhibit the growth ofCaco-2 human colon cancer cells in a dose-dependent manner, as shown by the resultsof an MTT assay. These in vitro results of B. polyfermenticus were confirmed in thepresent in vivo animal study. Our current results indicate clearly that the dietary feedingof B. polyfermenticus (3.1 x 108 cfu/day) effectively suppresses the occurrence of colonicACF induced by DMH when administered 1 week prior to treatment with thecarcinogen (Park et al. 2007). Using AOM-induced aberrant crypt foci in rats,Reddy et al. (1997) found that a stimulated growth of bifidobacteria in the coloncould lead to the inhibition of colon carcinogenesis. Reduction in GST activity isassociated with an increased risk for Colorectal Cancer (CRC) (Szarka et al. 1995).The use of lactulose separately or in conjunction with B. longum has been demonstratedto significantly increase GST levels in rat colon (Challa et al. 1997). These results led tothe suggestion that probiotics could suppress colon cancer.
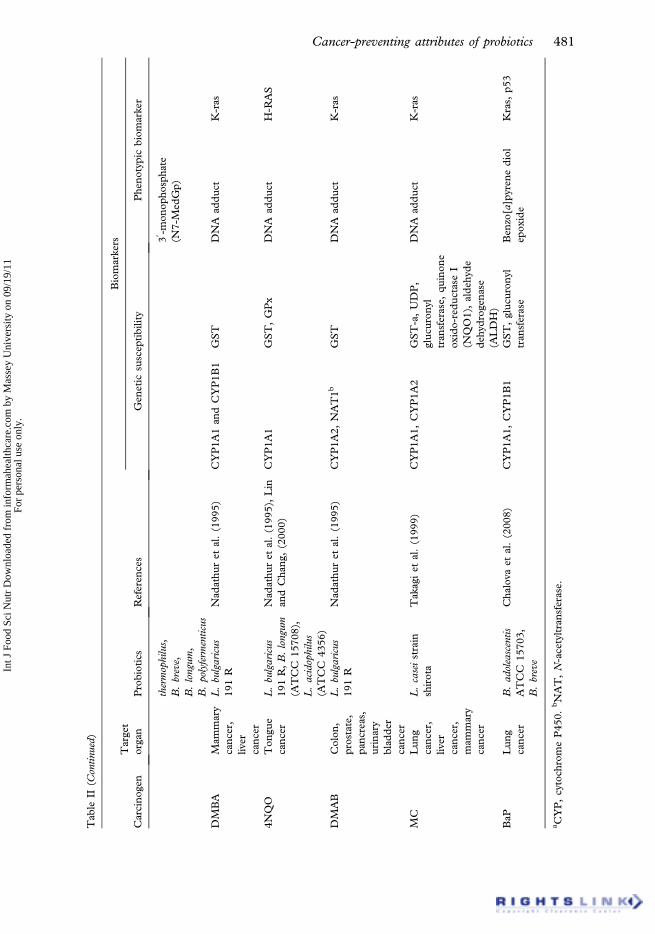
There are 57 cytochrome P450s encoded in the human genome, mainly catalyzingthe metabolism of steroids, bile acids, eicosanoids, drugs and xenobiotic chemicals.However, some of the P450s are also active carcinogens. Past epidemiological researchhas shown increased risk of colon cancer in individuals with high P450-1A2 activity.The metabolic activation of food-borne heterocyclic amines to colon carcinogens inhumans is hypothesized to occur via N-oxidation followed by O-acetylation to form theN-acetoxy arylamine that binds to DNA to give carcinogen-DNA adducts. These stepsare catalyzed by hepatic cytochrome P450-1A2 and N-acetyltransferase-2 (NAT-2),respectively (Lang et al. 1994) (Table II). It has been postulated that probiotics such asBifidobacterium could lower the risks of colon cancer, by producing metabolites thatcould affect the mixed-function of P450s and subsequently affect the conversion ofazoxymethane from proximate to ultimate carcinogen (Campbell and Hayes 1976).GST is an important family of phase II detoxification enzymes, which play a crucial rolein protecting the colon mucosa from dietary carcinogens.
Burns and Rowland (2000) suggested that increasing the amount of LAB in thecolon decreases the ability of microbiota to produce carcinogens. A randomized,
Cancer-preventing attributes of probiotics 479
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

Tab
leII.Examples
ofcarcinog
ens,
theirbiom
arkers
andpo
tentialprob
iotics
that
redu
cetheireffects.
Target
organ
Biomarkers
Carcino
gen
Probiotics
Referen
ces
Gen
etic
suscep
tibility
Phe
notypicbiom
arker
Activatingen
zymewith
gene
tic
polymorph
isms
Detox
ifyingen
zyme
withgene
tic
polymorph
isms
Add
uct
Gen
emutation
AFB1
Liver
L.rham
nosus,
L.acidophilus,
B.longum
Haskard
etal.(200
0)CYP3A
4a,CYP3A
5,CYP1A
2Glutathione
tran
sferases,
gluc
uron
yltran
sferases
Add
ucts
inDNA,
albu
min,he
mog
loblin
andurinarymetab
olites
p53mutation
atco
don24
9AGG
toAGT
IQColon
,breast
B.longum
,S.thermophilus,
B.an
imalis
Red
dyan
dRiven
son
(199
3),Tavan
etal.(200
2)CYP1A
2,NAT1b
GST,GPx,
NAT2
N-(de
oxygua
nosin-
8-yl)-2-am
ino-3-
methy
limidazo-[4,5-f]
quinoline(dG-C
8-IQ
)
k-ras
MEIQ
Colon
,breast
S.thermophilus,
B.an
imalis
Red
dyan
dRiven
son
(199
3),Tavan
etal.(200
2)CYP1A
2,NAT1b
GST,GPx,
NAT2,
gluc
uron
yltran
sferase
C-8
guan
ine
addu
cts
Kras
PhIP
Colon
,breast
S.thermophilus,
B.an
imalis
Red
dyan
dRiven
son
(199
3),Tavan
etal.(200
2)CYP1A
2,NAT1b
GST,GPx,
NAT2
DNA
addu
ctan
dad
ductsin
albu
min
C-8
guan
inead
ducts
P53
,K-ras
Trp-P-2
Liver,
colon
B.longum
Red
dyan
dRiven
son(199
3)CYP1A
2,CYP1A
1,an
dCYP2B
GST
DNA
addu
ctKras
DM
HColon
L.acidophilus,
L.caseisubsp.
rham
nosus,
B.breve
Goldinet
al.(198
0,19
96),
Ono
ueet
al.(199
7)CYP1A
1,CYP2E
1GST,gluc
uron
yltran
sferases
O6-Methy
lgua
nine
(O6-M
eGua
)ad
duct
Kras,
c-myc
AOM
Colon
L.acidophilus,
B.longum
Arimoc
hiet
al.(199
7),
Kulkarniet
al.(199
4),
Cha
llaet
al.(199
7),
Row
land
etal.(199
8)
CYP1A
1,CYP2E
1GST,gluc
uron
yltran
sferases
O6-Methy
lgua
nine
(O6-M
eGua
)ad
duct
Kras,
c-myc
MNNG
Colon
L.acidophilus,
L.gasseri,
L.confusus,S.
Poo
let
al.(199
6),
Parket
al.(200
7),
Caldini
etal.(200
5)
CYP1A
2,CYP1A
3GST
N7-M
ethy
lde
oxygua
nosine
Kras
480 M. Kumar et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.
Tab
leII
(Contin
ued)
Target
organ
Biomarkers
Carcino
gen
Probiotics
Referen
ces
Gen
etic
suscep
tibility
Phe
notypicbiom
arker
thermophilus,
B.breve,
B.longum
,B.polyferm
enticus
3¢-m
onop
hospha
te(N
7-Med
Gp)
DM
BA
Mam
mary
canc
er,
liver
canc
er
L.bulgaricus
191R
Nad
athu
ret
al.(199
5)CYP1A
1an
dCYP1B
1GST
DNA
addu
ctK-ras
4NQO
Ton
gue
canc
erL.bulgaricus
191R,B.longum
(ATCC
1570
8),
L.acidophilus
(ATCC
4356
)
Nad
athu
ret
al.(199
5),Lin
andCha
ng,(200
0)CYP1A
1GST,GPx
DNA
addu
ctH-R
AS
DM
AB
Colon
,prostate,
panc
reas,
urinary
blad
der
canc
er
L.bulgaricus
191R
Nad
athu
ret
al.(199
5)CYP1A
2,NAT1b
GST
DNA
addu
ctK-ras
MC
Lun
gcanc
er,
liver
canc
er,
mam
mary
canc
er
L.caseistrain
shirota
Takagiet
al.(199
9)CYP1A
1,CYP1A
2GST-a,UDP,
gluc
uron
yltran
sferase,
quinon
eox
ido-redu
ctaseI
(NQO1),alde
hyde
dehy
drog
enase
(ALDH)
DNA
addu
ctK-ras
BaP
Lun
gcanc
erB.ad
oleascentis
ATCC
1570
3,B.breve
Cha
lova
etal.(200
8)CYP1A
1,CYP1B
1GST,gluc
uron
yltran
sferase
Ben
zo[a]pyren
ediol
epox
ide
Kras,
p53
a CYP,cytoch
romeP45
0.bNAT,N-acetyltransferase.
Cancer-preventing attributes of probiotics 481
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

controlled study by Aso and Akazan (1992) demonstrated that the recurrence ofbladder tumors was delayed with daily intake of L. casei. They performed another studythat was larger (125 patients) and placebo-controlled, and found that L. casei reducedthe recurrence of tumors in all patients except those with more than one recurrenttumor (Aso et al. 1995). In an animal model with DMH-induced colon cancer, it wasshown that LGG significantly reduced the incidence of colon tumors (Goldin et al.1996). LAB administered to animals have been shown to prevent carcinogen-inducedpreneoplastic tumors and lesions (Wollowski et al. 2001) and also reduced the growthand viability of the HT-29 human colon cancer line (Hirayama and Rafter 2000). L.gasseri, L. confusus, S. thermophilus, B. breve, B. longum, and L. acidophilus strainsshowed an antigenotoxic effect after MNNG administration (Vanderhoof 2001), andL. acidophilus, L. confusus, S. thermophilus, L. delbrueckeii, L. gasseri, B. longum, and B.breve inhibited DNA damage from DMH (Rafter 1995).
Supplementation of the chow with 0.5% lyophilized B. longum caused pronouncedinhibition of the incidence of 3-amino-1-methyl-5H-pyrido[4,3-6]indole (Trp-P-2)-induced liver and colon tumors (i.e. 80% and 100% reduction in tumor frequencies,respectively) (Reddy and Rivenson 1993); however, the concentration of tryptophanpyrolysates in meats are much lower than those of other heterocyclic amines (HCAs)(Felton et al. 2000). Fermented milks prepared by S. thermophilus or Bifidobacteriumanimalis inhibited the induction of preneoplastic lesions (ACF in the colon) andDNA migration caused by a HCA mixture containing IQ, 2-amino-3,4-dimethyl-3H-imidazo[4,5-f]quinoline (MeIQ) and 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP) (ratio 1:1:1) (Tavan et al. 2002). Anti-mutagenicity of an acetoneextract of a L. bulgaricus 191R fermented yoghurt observed a significant dose-depen-dent anti-mutagenic activity against several mutagens including MNNG,4-nitroquinoline-N-oxide, 3,2-dimethyl-4-aminobiphenyl, 9,10-dimethyl-1,2-benz[a]anthracene and Trp-P2. This study suggests that a metabolite of the LAB may beresponsible for their anti-carcinogenic properties (Nadathur et al. 1995). Bifidobacter-ium infantis and L. acidophilus bacteria strains inhibited the growth of the MCF7 breastcancer cell line (Biffi et al. 1997). Studies have suggested that the host’s immuneresponse may be stimulated by B. infantis, leading to tumor suppression or regression(Hirayama and Rafter 2000). The metabolic activity of the intestinal microbiota mayalso be altered with administration of LAB (Goldin and Gorbach 1984). The alterationof the physicochemical conditions in the colon may influence colon cancer (Modleret al. 1990), and suggest that reducing the intestinal pH may prevent the growth ofputrefactive bacteria. In a 3-month study, L. acidophilus and B. bifidum were admin-istered to patients with colonic adenomas. The result was a decrease in fecal pH andcell proliferative activity in the upper colon (Biasco et al. 1991). The mechanisms of thelinks of probiotics to anti-tumor activity are not completely clear, but offer usefulpotential material for future cancer studies.
Possible modes of actions of probiotics: as evidenced by various probioticstrains
Reduction of reactive oxygen species load
Oxidative stress results when the balance between the production of ROS overrides theantioxidant capability of the target cell; oxidative damage from the interaction of
482 M. Kumar et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

reactive oxygen with critical cellular macromolecules may occur. ROS may interactwith and modify cellular protein, lipid, and DNA, which results in altered target cellfunction. The accumulation of oxidative damage has been implicated in both acute andchronic cell injury including possible participation in the formation of cancer. Oxidativestress occurs in a cell or tissue when the concentration of ROS generated exceeds theantioxidant capability of that cell (Sies 1991). ROS can be produced both endoge-nously and exogenously (Figure 3). Endogenous oxidative stress can be the result ofnormal cellular metabolism and oxidative phosphorylation. The metabolism of sub-stances by the P450 enzyme system generates oxygen free radicals through normal orfutile cycling mechanisms (Parke and Ioannides. 1990). Exogenous sources of ROScan also impact on the overall oxidative status of a cell. Drugs, hormones, and otherxenobiotic chemicals can produce ROS by either direct or indirect mechanisms (Trushand Kensler 1991; Halliwell 1996). Alternatively, oxidative stress can also occur whenthere is a decrease in the antioxidant capacity of a cell. Non-enzymatic antioxidantlevels (vitamin E, vitamin C, GSH, etc.) and enzymatic antioxidant levels (superoxidedismutase, GSH peroxidase, and catalase) in the cell can be decreased throughmodification in gene expression, decreased in their uptake in the diet, or can beoverloaded in ROS production, which creates a net increase in the amount of oxygenfree radicals present in the cell (Vuillaume 1987; Barber and Harris 1994). Severalhuman chronic disease states, including cancer, have been associated with oxidativestress produced through either an increased free radical generation and/or a decreasedantioxidant level in the target cells and tissues (Trush and Kensler 1991; Rice-Evans andBurdon 1993). Also it is well known that LGG and LGG-fermented milk were potentscavengers of superoxide anion and inhibitors of lipid peroxidation reactions in vitro(Ahotupa et al. 1996). Glutathione peroxidase (GPx) and glutathione-S-transferase
Endogenous source ofROS
mitochondriacytochrome P450
peroxisomesinflammatary cells
Exogenous source ofROS
radiationozone
hyperoxiaxenobiotics
Protein damage(DNA repair
proteins,Caspases)
DNA damage andmutation8-oxo-dG
nitrosamines8-nitroguanineS-nitrosothiol
DNA-strand breako
Lipidperoxidation
4HNE
MDA
Arachidonicacid cascade
Eicosanoids
Cellproliferation
Probioticbacteria
Source
Enz
ymat
ic(S
OD
, CA
T, G
SH
Per
oxid
ase)
Non
enzy
mat
ic(V
it E
, GS
H, V
it C
)
Cel
l mem
bran
e
Figure 3. Effect of probiotics on ROS production that are produced by endogenous and exogenous sources.
Cancer-preventing attributes of probiotics 483
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

(GST) are detoxification/biotransformation enzymes that are involved in the detoxifi-cation of toxic substances such as xenobiotics, carcinogens, free radicals and peroxidesby conjugating these substances with GSH (Hayes and Pulford 1995). Since the ultimatecarcinogenic form of DMH and AFB1 is a toxic electrophile (methyldiazonium ion andcarbonium ion) and exo-8,9-epoxide, GPx and GST take considerable significance inpromoting carcinogen detoxification (Beckett and Hayes, 1993). ROS are producedduring passage of nutrients through the gastrointestinal tract. The natural production ofhost antioxidants decreases rostrally. It is well known that oxidative damage forms part inthe pathogenesis of cancer, cirrhosis, atherosclerosis and other chronic diseases.B. longum (ATCC 15708) and L. acidophilus (ATCC 4356) showed antioxidative activity,inhibiting linoleic acid peroxidation by 28–48% and also showed the ability to scavengethe a-diphenyl-b-picrylhydrazyl free radical, scavenging 21–52%. The intact cells ofthese two intestinal bacteria demonstrated a high inhibitory effect on the cytotoxicity of4-nitroquinoline-N-oxide (4NQO). Cytotoxicity of 4NQO was reduced by L. acidophilusby approximately one-half, and by almost 90% by B. longum. Nevertheless, no inhibitionof cytoxicity was observed for intracellular cell-free extracts of 109 cells of B. longum andL. acidophilus (Lin and Chang 2000). Pediococcus pentosaceus 16:1 and L. plantarum 2592produced antioxidants after 18 h growth corresponding to 100 microgram vitamin C,and L. paracasei F19 slightly less but another L. paracasei did not exert antioxidativeactivity, again emphasizing that these characteristics are strain dependent (Kruszewskaet al. 2002). In a recent study, obligatory homofermentative lactobacilli produced highantioxidant activity whereas this was highly strain dependent among facultatively andobligate heterofermentative lactobacilli (Annuk et al. 2003).
Binding/adsorption of carcinogens
Several carcinogens such as heterocyclic amines and AFB1 are reportedly adsorbed orbound in vitro by LAB and other intestinal bacteria. A concomitant decrease inmutagenicity is often reported. (Orrhage 1994; Bolognani et al. 1997). In-depthinvestigations have also showed that cultured milk possessed des-mutagenicity andthis activity increased with increasing numbers of viable cells, indicating that probioticscould play an important role in the inhibition of mutagenicity (Usman 1998). Thya-garaja and Hosono (1993) found that probiotic isolated from ‘idly’, a traditional cerealpulse product of India, could exert des-mutagenicity on various spice mutagens,heterocyclic amines and aflatoxins. Subsequent studies on the des-mutagenicityproperties of probiotics suggested that the des-mutagenic substances may reside inthe cellular envelope of the bacterial cell wall (Singh et al. 1997). A cell wall preparationof B. infantis was found to inhibit tumor activity in mouse peritoneal cells (Sekine et al.1995), while a cell wall preparation of heat-killed L. casei (LC9018) was found toinduce immunity against tumor induction in a randomized, controlled and compar-ative study involving 223 patients with stage III cervical cancer. Also there are somepotential probiotics, Bifidobacterium adoleascentis ATCC 15703 and B. breve ATCC15700, that exhibited higher antimutagenicity against benzo[a]pyrene (BaP) andsodium azide (Chalova et al. 2008). Strains of B. lactis were shown to express anti-mutagenic properties, probably linked to cell wall constituents. The anti-mutageniceffect occurred even after acid and bile treatment, mimicking the gastrointestinaltransport; interestingly, enhanced in the presence of whole milk (Lo et al. 2004).One mechanism for this effect could be binding of mutagens, as heterocyclic aromatic
484 M. Kumar et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

amines were shown to be bound to the cell wall of certain bacteria such as B. longumand other LAB, and thereby be detoxified (Orrhage et al. 1994). However, the anti-mutagenic effect of L. plantarum KLAB21 was mediated by three glycoproteins that aresecreted extracellularly (Rhee and Park 2001).
Antigenotoxic activity of several Lactobacillus species against 4-nitroquinoline-1-oxide was shown in in vitro tests, whereas only one L. acidophilus strain inhibitedMNNG. All strains also showed anti-mutagenic properties and were viable after thetests (Caldini et al. 2005). Also, mutagens were suggested to be bound to the cell wallof probiotics.
Effect on production of bacterial enzymes and metabolites
Oral administration of some LAB/fermented milks to animals and humans leads to adecrease in certain metabolites and enzymes purported to be involved in synthesis oractivation of carcinogens, genotoxins and tumor promoters such as b-glucuronidase,b-glucosidase, nitrate reductase and ammonia. Such changes in enzyme activitiesconcomitantly suppressed tumors. This would appear to be due to the low specificactivity of these enzymes in LAB (Saito and Rowland 1992). Such changes in enzymeactivity or metabolite concentration have been suggested to be responsible for thedecreased level of pre-neoplastic lesions or tumors seen in carcinogen-treated rats givenprebiotics and probiotics (Reddy and Rivenson 1993; Rowland et al. 1998).
Stimulation of host enzymes involved in carcinogen inactivation
There are isolated reports citing that administration of LAB results in increased activityof antioxidative enzymes or processes that protect cells against carcinogen-induceddamage. These include GST, GSH, GSH reductase, and GSH peroxidase, whichconsequently reduce the apoptotic rate and the necrobiotic changes in liver. Suchmechanisms of protection could be effective against a wide range of dietary carcinogenspossibly influencing several cancer sites. Lowering of hepatic GSH levels and enhancedlipid peroxidation was studied in animals dosed with AFB1 and other carcinogens. Itmay be useful to employ hepatic lipid peroxidation as a biochemical marker duringAFB1-initiated hepato-carcinogenesis (Corrales et al. 1992; Aguilar-Delfin et al.1996; Gasso et al. 1996). Many food-borne carcinogens such as heterocyclic aminesand polycyclic aromatic hydrocarbons are known to be conjugated to GSH, whichappears to result in inactivation. The enzyme involved, GST, is found in the liver and inother tissues including the gut. A study of the effect of B. longum and lactulose on AOM-induced ACF in the colon showed that the activity of GSH in the colonic mucosa wasinversely related to the ACF numbers (Challa et al. 1997). Such a mechanism ofprotection would be effective against a wide range of dietary carcinogens that the variousLAB can inhibit genotoxicity of dietary carcinogens. The degree of inhibition wasstrongly species dependent. For example, Pool-Zobel et al. (1993) demonstrated thatL. casei and L. lactis inhibited the mutagenic activity of nitrosated beef by over 85%,whereas L. confusus and Lactobacillus sake had no effect. Pre-treatment of mice withL. casei 1 h before the administration of genotoxins can lead to a moderate reduction(P < 0.05) in the genotoxicity of pro-carbazine and urethane (Abraham and Paul 2001).Using the technique of single-cell microgel electrophoresis (Comet assay), Pool-Zobelet al. (1993) investigated the ability of range of species of LAB to inhibit DNA damage in
Cancer-preventing attributes of probiotics 485
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

the colon mucosa of rats treated with the carcinogens MNNG or DMH. All the strains oflactobacilli and bifidobacteria tested—L. acidophilus (isolated from a yoghurt sample), L.gasseri, L. confusus, B. breve and B. longum—prevented MNNG-induced DNA damagewhen given at a dose of 1010 cells/kg body weight 8 h before feeding the carcinogen. Inmost cases the DNA damage was reduced to a level similar to that in untreated rats. S.thermophilus was not as effective as the other LAB strains. The comet assay has also beenused to evaluate the effect of a prebiotic, lactulose on DNA damage in the colonicmucosa. Rats that were fed a diet containing 3% lactulose and given DMH exhibited lessDNA damage in colon cells than similarly treated animals fed a sucrose diet. In the latteranimals, the percentage of cells with severe DNA damage comprised 33% of the total,compared with only 12.6% in the lactulose-fed rats (Rowland et al. 1998).
Enhancing the host’s immune response
In animal and human studies with probiotic treatments (L. casei, L. acidophilus, orB. bifidus) shown to influence several aspects of immune function, enhancing secretoryIgA production was observed in which L. casei is most effective in stimulating secretoryIgA (Perdigon et al. 1991) and increasing the systemic immune response in malnour-ished animals (Perdigon 1992). There are several other examples of probiotics (L. casei,LGG, and other strains) that can affect cytokine production and promote a non-specific immune response by enhancing phagocytosis of pathogens (Erickson andHubbard 2000). Another study showed that mice fed LAB had higher splenocyteproliferation in response to mitogens for T cells and B cells (De Simone et al.1993; Erickson and Hubbard 2000). LAB shows anti-tumor activity against sarcoma180, a transplantable mouse tumor (Yokokura et al. 1981). One explanation for tumorsuppression by LAB may be mediated through an immune response of the host. Sekineet al. (1985) suggested that B. infantis stimulates the host-mediated response, leading totumor suppression or regression. In addition, there are studies to suggest that LAB playan important role and function in the host’s immunoprotective system by increasingspecific and non-specific mechanisms to have an anti-tumor effect (Kato et al.1983; De Simone et al. 1993; Schiffrin et al. 1995). L. casei Shirota (LcS) has beenshown to have potent anti-tumor and anti-metastatic effects on transplantable tumor cellsand to suppress chemically induced carcinogenesis in rodents. Also, intra-pleuraladministration of LcS into tumor-bearing mice has been shown to induce the productionof several cytokines, such as IFN-g, IL-1 and TNF-a, in the thoracic cavity of mice,resulting in the inhibition of tumor growth and increased survival (Matsuzaki 1998). Anadditional study has indicated that oral administration of BLP, a preparation of viableL. casei YIT 9018, potentiated systemic immune responses that modified T-cell func-tions in tumor-bearing mice (Kato et al. 1994). It has also been demonstrated thatB. longum and B. animalis promote the induction of inflammatory cytokines (IL-6,TNF-a) in mouse peritoneal cells (Sekine et al. 1994). Suppression of colon tumorgrowth by yoghurt in DMH-treated mice was associated with suppression of theinflammatory immune response (Wollowski et al. 2001). Many case studies have shownthat consumption of fermented milk resulted in reduction of risk of breast, liver andcolon cancer (Aso and Akazan 1992).
Among them, LcS showed especially high potency. LcS is a strain of LAB that hasbeen selected for its specific biological activity in humans. This strain is not directlycytotoxic to tumor cells in vitro; it has been postulated that its anti-tumor action may be
486 M. Kumar et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

mediated by augmentation of the host’s immune system (Kato et al. 1981) and alsoexhibited marked anti-tumor activity against human malignant cancer cells in clinicaltrials (Aso and Akazan 1992; Aso et al. 1995). This idea stimulated further studies ofthe anti-tumor and immunoregulatory actions of LcS in various experimental models(Matsuzaki et al. 1985, 1988; Miake et al. 1985), and it has been demonstrated that oraladministration of LcS has beneficial effects in both humans and animals. It has alsobeen reported that LcS induced recovery of host immune responses that were decreasedby treatment with carcinogens and augmented the natural killer activity and T-cellfunctions of host immune cells. After LcS is ingested by the host, it is incorporatedinto M cells in Peyer’s patches and digested to form active components. In Peyer’spatches, macrophages or dendritic cells that phagocytosed LcS gained the ability toproduce several cytokines, especially TNF. The components of LcS digested in Peyer’spatches were then recognized through Toll-like receptor 2 in antigen-presenting cells,resulting in the production of several cytokines that elicited varied responses in hostimmune cells (Takeshi et al. 2007). LcS exerted a strong inhibitory effect on carcino-genesis in mice through the regulation of host immune cells (Takagi et al. 1999) in a3-methylcholanthrene (MC)-induced carcinogenesis model (Noguchi et al. 1996) thatinduced various tumors such as colon, liver, lung, uterine cervix, and mammary glandcancer models (Rao and Hussain 1988; Baral andMaity 1992). Oral feeding of mice withLcS inhibits MC-induced tumorigenesis by modulating the host immune responses thatare disrupted during MC carcinogenesis. A possible mechanism of the prevention of thecarcinogenesis is the activation of natural killer (NK) cells. NK cells are large granularlymphocytes derived from bone marrow, and these cells display non-MHC-restrictedcytotoxicity against a variety of tumors. It is well recognized that NK cells act as cytolyticeffector cells of the innate immune system. Oral feeding of LcS to MC-treated micerendered their NK cells tumoricidal in terms of both quality and quantity, resulting in thesuppression of tumor incidence (Takagi et al. 2001). Other possible mechanism of actionin which effector cells that may respond to LcS are dendritic cells. Dendritic cells arethought to be one of the most important types of cells involved in the presentation ofseveral antigens and in the production of cytokines (Banchereau and Steinman1998; Rescigno et al. 1999). Recent studies showed Lactobacillus strain-specific activityin prevention of murine tumorigenesis and in induction of IL-12 release by bonemarrow-derived dendritic cells in vitro (Takagi et al. 2008). Recently, Matsumotoet al. (2009) described the role of the LAB strain LcS for prevention of colon cancerby targeting immune system. They reported that L. casei produces polysaccharide, whichsuppresses colon carcinogenesis by suppressing synthesis of IL-6 and signal transducerand activator of transcription 3 in a mouse model.
These findings suggest that treatment with probiotics has the potential to ameliorateor prevent tumorigenesis through modulation of the host’s immune system, specificallycellular immune responses. Elucidating the precise mechanism of cancer prevention byprobiotic bacteria may help identify a novel and effective molecular target for cancerprevention.
Production of conjugated linoleic acid from castor oil
Conjugated linoleic acid (CLA) 4 is a term defining a group of positional (e.g. 7:9,9:11, 10:12, and 11:13) and geometric (i.e. cis or trans) isomers of linoleic acid(C18, cis-9:cis-12) that have been shown to exert numerous health benefits, including
Cancer-preventing attributes of probiotics 487
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

anti-atherogenic, anti-diabetic, anti-inflammatory and anticarcinogenic properties(Maggiora et al. 2004). In vitro, CLA inhibits the growth of HT-29 and Caco-2 coloncancer cells (Lampen et al. 2005). CLA-treated SW480 colonic tumor cells possessincreased caspase-3 and caspase-9 activities and reduced Bcl-2 expression comparedwith controls (Miller et al. 2002). CLA has been shown to reduce the incidence ofcolonic, skin, mammary, and prostate carcinogenesis in animal models (Belury 2002).Rats supplemented with CLA showed reduced incidence of colonic tumors andincreased apoptotic indices in response to the administering of DMH (Kim andPark 2003). Some probiotic strains of bacteria have demonstrated protective effectsagainst tumor production (Sekine et al. 1995; Biffi et al. 1997; O’Mahony et al.2001; Saikali et al. 2004). L. acidophilus and B. longum have been shown to reduceincidence of colonic tumors and aberrant crypt foci, respectively, in animal models(Rowland et al. 1996; McIntosh et al. 1999). A cocktail of probiotic strains was recentlydemonstrated to increase the colonic apoptotic index in normal rats (Linsalata et al.2005). Several strains of bacteria that are considered to have probiotic effects(i.e. lactobacilli and bifidobacteria) are capable of converting linoleic acid to CLA.In this study, we investigated a previously unreported mechanism of probiotic action:the production of CLA. We demonstrated that probiotic strains in VSL3 (L. casei,L. plantarum, L. acidophilus, L. delbrueckii subsp. bulgaricus, B. infantis, B. breve,B. longum, S. salivarius subsp. thermophilus) have the capacity to convert linoleicacid to CLA, inducing the upregulation of PPARg, a reduction in cancer cell viability,and the induction of apoptosis. This linoleic acid-conjugating capacity is maintainedin vivo (Julia et al. 2006). One LAB strain, L. plantarum JCM 1551, was shown toefficiently produce conjugated linoleic acid from castor oil in the presence of lipase,which is an interesting ‘side effect’ of anti-tumor activities of LAB (Ando et al. 2004).
By regulation of apoptosis
Apoptosis is a genetically regulated active process abolishing cell populations in bothphysiological and pathological processes. Impaired apoptosis, either due to expressionof oncogenes or mutations of tumor suppressor genes, leads to an uncontrolledaccumulation of malignant cells, and eventually to cancer.
Caspase-3 is one of the cysteine proteases that play a major role in the execution ofapoptosis (Nicholson 1999). A number of genetic and biochemical studies suggest thatcaspase activation is essential for the occurrence of the apoptotic phenotype of celldeath (Janicke et al. 1998) A variety of caspase substrates are involved in the regulationof DNA structure, repair and replication. Caspase-3 substrate cleavage has beenobserved under oxidative stress in different pathological conditions (Nicholson andThornberry 1997). Aflatoxins are one of the most dangerous mycotoxins known, owingto their high toxicity to both animals and human. AFB1, a metabolite of Aspergillusflavus, is a potent hepatotoxic and hepatocarcinogenic mycotoxin. One of manifesta-tions of AFB1-induced toxicity is oxidative stress (Souza et al. 1999). Recently, it isaccepted that oxidative stress is an apoptosis inducer (Chandra et al 2000). Manyagents that induce apoptosis are either oxidants or stimulators of cellular oxidativemetabolism. Conversely, many inhibitors of apoptosis have antioxidant activities orenhance cellular antioxidant defenses (Freeman and Grapo 1982). The GSH redoxcycle is an important component of the antioxidant machinery in cells. In normal cells,a primary defense against oxidative damage is provided by antioxidants such as GSH
488 M. Kumar et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

and the onset of apoptosis is associated with a fall in intracellular GSH in numerouscellular systems (Ratan et al. 1994).
There are isolated reports citing that administration of LAB results in increasedactivity of antioxidative enzymes or process that protect cells against carcinogen-induced damage. These include GST, GSH, GSH reductase, and GSH peroxidase,which consequently reduce the apoptotic rate and the necrobiotic changes inliver. Such mechanisms of protection could be effective against a wide range ofdietary carcinogens possibly influencing several cancer sites. Some probiotics(e.g. LGG) prevent cytokine-induced apoptosis in intestinal epithelial cell models.Also it is well known that LGG and LGG-fermented milk are potent scavengers ofsuperoxide anion and inhibitors of lipid peroxidation reactions in vitro (Ahotupa et al.1996).
Culture of LGG with either mouse or human colon cells activates the anti-apoptoticAkt/protein kinase B. This model probiotic also inhibits activation of the pro-apoptoticp38/mitogen-activated protein kinase by TNF, IL-1a or IFN-g. Furthermore, productsrecovered from LGG culture broth supernatant showed concentration-dependentactivation of Akt and inhibition of cytokine-induced apoptosis. These observationssuggest a novel mechanism of communication between probiotics and colon epitheliathat increases survival of intestinal cells normally found in an environment ofpro-apoptotic cytokines (Fang and Polk 2002). LGG enhanced gastric ulcer healingvia attenuation of the cell apoptosis to cell proliferation ratio and an increase inangiogenesis. Regulators of these processes such as ornithine decarboxylase, B-celllymphoma 2 (Bcl-2), vascular endothelial growth factor and epidermal growth factorreceptor are likely to be involved in the healing action of LGG for gastric ulcer(Lam et al. 2007). B. longum is a probiotic, known for its beneficial effects to thehuman gut and even for its immune-modulatory and anti-tumor activities. Recently,many studies have indicated an intimate relation between probiotic bacteria and thegut mucosa and its influence on human cellular homeostasis. Bacteria were incubatedwith Caco-2 cells to investigate apoptotic deletion. Co-cultures of Caco-2 cells withadherent strains (B12 and B18) of B. longum induced DNA fragmentation, indicatingthat it can induce apoptotic deletion of Caco-2 cells. Thus it helps in restoring theecology of damaged colon tissues. Lactobacillus reuteri secretes factors that potentiateapoptosis in myeloid leukaemia-derived cells induced by TNF, as indicated byintracellular esterase activity, terminal deoxynucleotidyl transferase-mediateddeoxyuridine triphosphate nick end-labeling assays and poly(ADP-ribose) polymer-ase cleavage. L. reuteri downregulated nuclear factor-kB-dependent gene productsthose mediate cell proliferation (Cox-2, cyclin D1) and cell survival (Bcl-2, Bcl-xL).Moreover Otte et al. (2009) demonstrated that a probiotic combination call VSL#3significantly suppressed the COX-2 expression in a cancer cell line (Colo320 andSW480 intestinal epithelial cells). L. reuteri suppressed TNF-induced nuclearfactor-kB activation, including nuclear factor-kB-dependent reporter geneexpression, in a dose-dependent and time-dependent manner. L. reuteri mayregulate cell proliferation by promoting apoptosis of activated immune cells viainhibition of IkBa ubiquitination and enhancing pro-apoptotic mitogen-activatedprotein kinase signaling. An improved understanding of L. reuteri-mediated effectson apoptotic signaling pathways may facilitate development of future probiotic-based regimens for prevention of colorectal cancer and inflammatory bowel disease(Iyer et al. 2008).
Cancer-preventing attributes of probiotics 489
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

Conclusions
Although there is no direct evidence for the prevention or treatment of cancermalignancies in humans using probiotics, numerous in vivo animal studies have beencarried out. These have demonstrated that probiotics are capable of reducing theincidence of tumor formation and aberrant crypt formation, thereby suppressing bac-terial enzyme activities and reducing DNA damage. Mechanistic studies indicate thatprobiotics may exert anticarcinogenic properties by altering colonic metabolism, degrad-ing carcinogens, producing antimutagenic compounds, and enhancing host immuneresponses and by preventing cytokine-induced apoptosis of colonic epithelial cells.Therefore LAB offer potential as chemo-protective agents and thus further researchis clearly needed to quantify the beneficial effects for prevention of human cancer.
Declaration of interest:The authors report no conflicts of interest. The authors aloneare responsible for the content and writing of the paper.
References
Abdelali H, Cassand P, Soussotte V, Daubeze M, Bouley C, Narbonne JF. 1995. Effect of dairy products oninitiation of precursor lesions of colon cancer in rats. Nutr Cancer 24:121–123.
Abraham SK, Paul J. 2001. Protective effects of lactic acid bacteria against genotoxicity in mice. Curr Sci80:1310-1312.
Aguilar F, Hussain SP, Cerutti P. 1993. Aflatoxin B1 induces the transversion of G! T in codon 249 of thep53 tumor suppressor gene in human hepatocytes. Proc Natl Acad Sci USA 90:8586–8590.
Aguilar-Delfin I, Lopez-Barrera F, Hernandez-Munoz R. 1996. Selective enhancement of lipid peroxidationin plasma membrane in two experimental models of liver regeneration: Partial hepatectomy and acuteCCl4 administration. Hepatology 2(4):657–662.
Ahotupa M, Saxelin M, Korpela R. 1996. Antioxidant properties of lactobacillus GG. Nutr Today Suppl31:51–52.
Ando A, Ogawa J, Kishino S, Shimizu S. 2004. Conjugated linoleic acid production from castor oil byLactobacillus plantarum JCM 1551. Enzyme Microb Technol 35:40–45.
Annuk H, Shchepetova J, Kullisaar T, Songisepp E, Zilmer M, Mikelsaar M. 2003. Characterization ofintestinal lactobacilli as putative probiotic candidates. J Appl Microbiol 94:403–412.
Arimochi H, Kinouchi T, Kataoka K, Kuwahara T, Ohnishi Y. 1997. Effect of intestinal bacteria onformation of azoxymethane-induced aberrant crypt foci in the rat colon. Biochem Biophys Res Commun238:753–757.
Aso Y, Akazan H. 1992. Prophylactic effect of a Lactobacillus casei preparation on the recurrence of superficialbladder cancer. BLP Study Group. Urol Int 49:125–129.
Aso Y, Akaza H, Kotake T, Tsukamoto T, Imai K, Naito S 1995. Preventive effect of a Lactobacillus caseipreparation on the recurrence of superficial bladder cancer in a double-blind trial. The BLP Study Group.Eur Urol 27:104–109.
Banchereau J, Steinman RM. 1998. Dendritic cells and the control of immunity. Nature 392:245–252.Baral RN, Maity P. 1992. Induction of colorectal cancer in rats by 20-methylcholanthrene. Cancer Lett
61:177–183.Barber DA, Harris SR. 1994. Oxygen free radicals and antioxidants: A review. Am Pharm 34:26–35.Bartsch H, Nair J. 2002. Potential role of lipid peroxidation derived DNA damage in human colon
carcinogenesis: Studies on exocyclic base adduct as stable oxidative stress markers. Cancer DetectPrev 26:308–312.
Beckett GJ, Hayes JD. 1993. Glutathione S-transferases: biomedical applications. Adv Clin Chem 30:281–380.
Belury MA. 2002. Inhibition of carcinogenesis by conjugated linoleic acid: Potential mechanisms of action.J Nutr 132:2995–2998.
Biasco G, Paganelli GM, Brandi G, Brillanti S, Lami F, Callegari C, Gizzi G. 1991. Effect of Lactobacillusacidophilus and Bifidobacterium bifidum on rectal cell kinetics and fecal pH. Ital J Gastroenterol 23:142–147.
Bibel DJ. 1998. Elie Metchnikoff’s bacillus of long life. ASM News 54:661–665.
490 M. Kumar et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

Biffi A, Coradini D, Larsen R, Riva L, Di Fronzo G. 1997. Antiproliferative effect of fermented milk on thegrowth of a human breast cancer cell line. Nutr Cancer 28:93–99.
Bird RP. 1995. Role of aberrant crypt foci in understanding the pathogenesis of colon cancer. Cancer Lett93:55–71.
Bolognani F, Rumney CJ, Rowland IR. 1997. Influence of carcinogen binding by lactic acid producingbacteria on tissue distribution and in-vivo mutagenicity of dietary carcinogens. Food Chem Toxicol35:535–545.
Bosch FX, Munoz N. 1988. Prospects for epidemiological studies on hepatocellular cancer as a model forassessing viral and chemical interactions. IARC Sci Publ 89:427–438.
Bulatao-Jayme J, Almero EM, Castro MC, Jardeleza MT, Salamat LA. 1982. A case–control dietary study ofprimary liver cancer risk from aflatoxin exposure. Int J Epidemiol 11:112–119.
Burns AJ, Rowland IR. 2000. Anti-carcinogenicity of probiotics and prebiotics. Curr Issues Intest Microbiol1:13–24.
Caldini G, Trotta F, Villarini M, Moretti M, Pasquini R, Scassellati-Sforzolini G, Cenci G. 2005. Screeningof potential lactobacilli antigenotoxicity by microbial and mammalian cell-based tests. Int. J FoodMicrobiol 102:37–47.
Campbell TC, Hayes JR. 1976. The effect of quantity and quality of dietary protein on drug metabolism. FedProc 35:2470–2474.
Challa A, Rao DR, Chawan CB, Shackelford L. 1997. Bifidobacterium longum and lactulose suppressazoxymethane-induced colonic aberrant crypt foci in rats. Carcinogenesis 18:517–521.
Chalova VI, Lingbeck JM, Kwon YM, Ricke SC. 2008. Extracellular antimutagenic activities of selectedprobiotic Bifidobacterium and Lactobacillus spp. as a function of growth phase. J Environ Sci Health Part B43:193–198.
Chandra J, Samali A, Orrenius S. 2000. Triggering and modulation of apoptosis by oxidative stress. FreeRadic Biol Med 29:323–333.
Chau I, Cunningham D. 2006. Adjuvant therapy in colon cancer—what, when and how? Ann Oncol17:1347–1359.
Chin SE, Liu W, Storkson JM, Ha YL, Pariza MW. 1992. Dietary sources of conjugated dienoic isomers oflinoleic acid, a newly recognized class of anticarcinogens. J Food Comp Anal 5:185–197.
Collins MD, Gibson GR. 1999. Probiotics, prebiotics, and synbiotics: approaches for modulating themicrobial ecology of the gut. Am J Clin Nutr 69:1052S-1057S.
Corrales F, Gimenez A, Alvarez L, Caballeria J, Pajares MA, Andreu H, Pares A, Mato JM, Rodes J. 1992.S-adenosylmethionine treatment prevents carbon tetrachloride-induced S-adenosylmethionine synthetaseinactivation and attenuates liver injury. Hepatology 16:1022–1027.
De Simone C, Vesely R, Bianchi SB, Salvadori B, Jirillo E. 1993. The role of probiotics in modulation of theimmune system in man and in animals. Int J Immunother 9:23–28.
Di Caro S, Tao H, Grillo A, Elia C, Gasbarrini G, Sepulveda AR, Gasbarrini A. 2005. Effects of LactobacillusGG on genes expression pattern in small bowel mucosa. Digest Liver Dis 37:320–332.
Drisko JA, Giles CK, Bischoff BJ. 2003. Probiotics in health maintenance and disease prevention. Alt MedRev 8:143–155.
Erdman SH, Wu HD, Hixson LJ, Ahnen DJ, Gerner EW. 1997. Assessment of mutations in Ki-ras andp53 in colon cancers from azoxymethane- and dimethylhydrazine-treated rats. Mol Carcinogen 19:137–144.
Erickson KL, Hubbard NE. 2000. Probiotic immuno-modulation in health and disease. J Nutr 130:403S–409S.
Essigmann JM, Croy RG, Nadzan AM, Busby WF Jr, Reinhold VN, Buchi G, Wogan GN. 1977. Structuralidentification of the major DNA adduct formed by aflatoxin B1 in-vitro. Proc Natl Acad Sci USA 74:1870–1874.
Fang Y, Polk DB. 2002. Probiotic bacterium prevents cytokine-induced apoptosis in intestinal epithelial cells.J Biol Chem 277:50959-50965.
Felton JS, JagerstadM, KnizeMG, Skog K,Wakabayashi K. 2000. Contents in foods, beverages and tobacco.In: Nagao M, Sugimura T, editors. Food borne carcinogenesis: Heterocyclic amines. New York: Wiley.pp 5–29.
Femia AP, Luceri C, Dolara P, Giannini A, Biggeri A, Salvadori M, Clune Y, Collins KJ, Paglierani M,Caderni G. 2002. Antitumorigenic activity of the prebiotic inulin enriched with oligofructose incombination with the probiotics Lactobacillus rhamnosus and Bifidobacterium lactis on azoxymethane-induced colon carcinogenesis in rats. Carcinogenesis 23:1953–1960.
Cancer-preventing attributes of probiotics 491
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

Fernandes CF, Shahani KM. 1990. Anticarcinogenic and immunological properties of dietary lactobacilli.J Food Protect 53:704–710.
Finley JW, Davis CD, Feng Y. 2000. Selenium from high-selenium broccoli protects rats from colon cancer.J Nutr 130:2384–2389.
Freeman BA, Grapo JD. 1982. Biology of disease: Free radical and tissue injury. Lab Invest 47:412–426.Gallaher DD, Stallings WH, Blessing LL, Busta FF, Brady LJ. 1996. Probiotics, cecal microbiota, and
aberrant crypts in the rat colon. J Nutr 126:1362–1371.Garner RC, Miller EC, Miller JA. 1972. Liver microsomal metabolism of aflatoxin B1 to a reactive derivative
toxic to Salmonella typhimurium TA 1530. Cancer Res 32:2058–2066.Gasso M, Rubio M, Varela G, Cabre M, Caballeria J, Alonso E, Deulofem R, Camps J, Gimenez A, Pajares
M, Pares A, et al 1996. Effects of S-adenosylmethionine on lipid peroxidation and liver fibrogenesis incarbon tetrachloride-induced cirrhosis. J Hepatol 25:200–205.
Goldin B. 1996. The metabolic activity of the intestinal microbiota and its role in colon cancer. Nutr Today31:24S–27S.
Goldin BR. 1990. Intestinal microbiota: Metabolism of drugs and carcinogens. Ann Med 22:43–48.Goldin BR, Gorbach SL. 1980. Effect of Lactobacillus acidophilus dietary supplements on
1,2-dimethylhydrazine dihydrochloride-induced intestinal cancer in rats. J Natl Cancer Inst 64:263–265.Goldin BR, Gorbach SL. 1984. The effect of milk and lactobacillus feeding on human intestinal bacterial
enzyme activity. Am J Clin Nutr 39:756–761.Goldin BR, Swenson L, Dwyer J, Sexton M, Gorbach SL. 1980. Effect of diet and Lactobacillus acidophilus
supplements on human fecal bacterial enzymes. J Natl Cancer Inst 64:255–261.Goldin BR, Gualtieri LJ, Moore RP. 1996. The effect of Lactobacillus GG on the initiation and promotion of
DMH-induced intestinal tumours in the rat. Nutr Cancer 25:197–204.Halliwell B. 1996. Mechanisms involved in the generation of free radicals. Pathol Biol 44:6–13.Haskard C, Binnion C, Ahokas J. 2000. Factors affecting the sequestration of aflatoxin by Lactobacillus
rhamnosus strain GG. Chem Biol Interact 128:39–49.Hayatsu H, Hayatsu T. 1993. Suppressing effect of Lactobacillus casei administration on the urinary
mutagenicity arising from ingestion of fried ground beef in the human. Cancer Lett 73:173–179.Hayes JD, Pulford DJ. 1995. The glutathione S-transferase supergene family: Regulation of GST and
contribution of the isoenzymes to cancer chemoprevention and drug resistance. Crit Rev Biochem MolBiol 31:445–600.
Hirayama K, Rafter J. 1999. The role of lactic acid bacteria in colon cancer prevention: Mechanisticconsiderations. Ant V Leeuwenhoek 76:391–394.
Hirayama K, Rafter J. 2000. The role of probiotic bacteria in cancer prevention. Microbes Infect 2:681–686.Iyer C, Kosters A, Sethi G, Kunnumakkara AB, Aggarwal BB, Versalovic J. 2008. Probiotic Lactobacillus
reuteri promotes TNF-induced apoptosis in human myeloid leukemia-derived cells by modulation ofNF-kB and MAPK signalling. Cellular Microbiol 10:1442–1452.
Iyer RS, Coles BF, Raney KD, Their R, Guengerich FP, Harris TM. 1994. DNA adduction by the potentcarcinogen aflatoxin B-1-mechanistic studies. J Am Chem Soc 116:1603–1609.
Janicke RU, Spregart ML, Wati MR, Porter AG. 1998. Caspase-3 is required for DNA fragmentation andmorphological changes associated with apoptosis. J Biol Chem 273:9357–9360.
Jeong HY, Kim TH, Park JS, Kim KT, Paik HD. 2003. Antioxidative and cholesterol-reducing activity ofBacillus polyfermenticus SCD. Korean J Biotechol Bioeng 18:371–376.
Julia B, Ewaschuk JW, Walker H, Diaz H, Madsen KL. 2006. Bioproduction of conjugated linoleic acid byprobiotic bacteria occurs in-vitro and in-vivo in mice. J Nutr 136:1483–1487.
Kampman E, Goldbohm RA, van den Brandt PA. 1994. Fermented dairy products, calcium and colorectalcancer in the Netherlands cohort study. Cancer Res 54:3186–3190.
Kato I, Kobayashi S, Yokokura T, Mutai M. 1981. Antitumor activity of Lactobacillus casei in mice. Gann72:517–523.
Kato I, Yokokura T, Mutai M. 1983. Macrophage activation by Lactobacillus casei in mice. MicrobiolImmunol 27:611–618.
Kato I, Endo K, Yokokura T, 1994. Effects of oral administration of Lactobacillus casei on antitumorresponses induced by tumor resection in mice. Int J Immunopharmacol 16:29–36.
Khanafari A, Soudi H, Miraboulfathi M, Osbo RK. 2007. An in vitro investigation of aflatoxin B1 biologicalcontrol by Lactobacillus plantarum. Pakistan J Biol Sci 10:2553–2556.
Kim KH, Park HS. 2003. Dietary supplementation of conjugated linoleic acid reduces colon tumor incidencein DMH-treated rats by increasing apoptosis with modulation of biomarkers. Nutrition 19:772–777.
492 M. Kumar et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

Korenaga D, Takesue F, Kido K, Yasuda M, Inutsuka S, Honda M, Nagahama S. 2002. Impairedantioxidant defense system of colonic tissue and cancer development in dextran sulfate sodium-induced colitis in mice. J Surg Res 102:144–149.
Kruszewska D, Lan J, Lorca G, Yanagisawa N, Marklinder I, Ljungh A. 2002. Selection of lactic acid bacteriaas probiotic strains by in-vitro tests. Microb Ecol Health Dis 29:37–49.
Kulkarni N, Reddy BS. 1994. Inhibitory effect of Bifidobacterium longum cultures on the azoxymethaneinduced aberrant crypt foci formation and fecal bacterial b-glucuronidase. Proc Soc Exp Biol Med207:278–283.
Lam EKY, Tai EKK, Koo MWL, Wong HPS, Wu WKK, Yu L, So WHL, Cho CH. 2007. Enhancement ofgastric mucosal integrity by Lactobacillus rhamnosus GG. Life Sci 80:2128–2136.
Lampen A, Leifheit M, Voss J, Nau H. 2005. Molecular and cellular effects of cis-9, trans-11-conjugatedlinoleic acid in enterocytes: Effects on proliferation, differentiation, and gene expression. Biochim BiophysActa 1735:30–40.
Lang NP, Butler MA, Massengill J, Lawson M, Stotts RC, Maurer-Jensen M, Kadlubar FF. 1994. Rapidmetabolic phenotypes for acetyltransferase and cytochrome P4501A2 and putative exposure to food-borneheterocyclic amines increase the risk for colorectal cancer or polyps. Cancer Epidemiol Biomark Prevent3:675–682.
Lee YK, Salminen S. 1995. The coming age of probiotics. Trends Food Sci Technol 6:241–245.Lidbeck A, Nord CE, Gustafsson JA, Rafter J. 1992. Lactobacilli, anticarcinogenic activities and human
intestinal microbiota. Eur J Cancer Prev 1:341–353.Lilleberg SL, Cabonce MA, Raju NR, Wagner LM, Kier LD. 1992. Alterations in the p53 tumor suppressor
gene in rat liver tumors induced by aflatoxin B1. Prog Clin Biol Res 376:203–222.Lin MY, Chang FJ. 2000. Antioxidative effect of intestinal bacteria Bifidobacterium longum ATCC 15708 and
Lactobacillus acidophilus ATCC 4356. Dig Dis Sci 45:1617–1622.Ling WH, Korpela R, Mykkanen H, Salminen S, Hanninen O. 1994. Lactobacillus strain GG
supplementation decreases colonic hydrolytic and reductive enzyme activities in healthy female adults.J Nutr 124:18–23.
Linsalata M, Russo F, Berloco P, Valentini AM, Caruso ML, De Simone C, Barone M, Polimeno L, Di LeoA. 2005. Effects of probiotic bacteria (VSL#3) on the polyamine biosynthesis and cell proliferation ofnormal colonic mucosa of rats. In Vivo 19:989–995.
Lo PR, Yu RC, Chou CC, Huang EC. 2004. Determinations of the antimutagenic activities of severalprobiotic bifidobacteria under acidic and bile conditions against benzo[a]pyrene by a modified Ames test.Int J Food Microbiol 93:249–257.
Maggiora M, Bologna M, Ceru MP, Possati L, Angelucci A, Cimini A, Miglietta A, Bozzo F, Margiotta C,Muzio G, Canuto RA. 2004. An overview of the effect of linoleic and conjugated-linoleic acids on thegrowth of several human tumor cell lines. Int J Cancer 112:909–919.
Marteau P, Rochart P, Flourie B, Pellier P, Santos L, Desjeux JF, Rambaud JC. 1990. Effect of chronicingestion of a fermented dairy product containing Lactobacillus acidophilus and Bifidobacterium bifidum onmetabolic activities of the colonic flora in humans. Am J Clin Nutr 52:685–688.
Matsuzaki T. 1998. Immunomodulation by treatment with Lactobacillus casei strain Shirota. Int J FoodMicrobiol 41:133–140.
Matsuzaki T, Yokokura T, Azuma I. 1985. Anti-tumor activity of Lactobacillus casei on Lewis lung carcinomaand line-10 hepatoma in syngeneic mice and guinea pigs. Cancer Immunol Immunother 20:18–22.
Matsuzaki T, Yokokura T, Mutai M. 1988. Antitumor effect of intrapleural administration of Lactobacilluscasei in mice. Cancer Immunol Immunother 26:209–214.
McIntosh GH, Royle PJ, Playne MJ. 1999. A probiotic strain of L. acidophilus reduces DMH-induced largeintestinal tumors in male Sprague–Dawley rats. Nutr Cancer 35:153–159.
Miake S, Nomoto K, Yokokura T, Yoshikai Y, Mutai M, Nomoto K. 1985. Protective effect of Lactobacilluscasei on Pseudomonas aeruginosa infection in mice. Infect Immun 48:480–485.
Miller A, Stanton C, Devery R. 2002. Cis 9, trans 11- and trans 10, cis 12-conjugated linoleic acid isomersinduce apoptosis in cultured SW480 cells. Anticancer Res 22:3879–3887.
Modler GW, McKellar RC, Yaguchi M. 1990. Bifidobacteria and bifidogenic factors. Can Inst Food SciTechnol J 23:29–41.
Murosaki S, Muroyama K, Yamamoto Y, Yoshikai Y, 2000. Antitumor effect of heat-killed Lactobacillusplantarum L-137 through restoration of impaired interleukin-12 production in tumor-bearing mice.Cancer Immunol Immunother 49:157–164.
Cancer-preventing attributes of probiotics 493
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

Matsumoto S, Hara T, Nagaoka M, Mike A, Mitsuyama K, Sako T, YamamotoM, Kado S, Takada T. 2009.A component of polysaccharide peptidoglycan complex on Lactobacillus induced an improvement ofmurine model of inflammatory bowel disease and colitis-associated cancer. Immunology 128:e170–e180.
Nadathur S, Gould S, Bakalinsky A. 1995. Antimutagenicity of an acetone extract of yoghurt. Mutat Res334:213–224.
Nair S, Norkus EP, Hertan H, Pitchumoni CS. 2002. Serum and colon mucosa micronutrient antioxidants:Differences between adenomatous polyp patients and controls. Am J Gastroenterol 96:3400–3405.
Nicholson DW. 1999. Caspase structure, proteolytic substrates and function during apoptotic cell death. CellDeath Diff 6:1028–1042.
Nicholson DW, Thornberry NA. 1997. Caspases: Killer proteases. Trends Biochem Sci 22:299–306.Noguchi Y, Jungbluth A, Richards EC, Old LJ. 1996. Effect of interleukin 12 on tumor induction by
3-methylcholanthrene. Proc Natl Acad Sci USA 93:11798–11801.O’Mahony L, Feeney M, O’Halloran S, Murphy L, Kiely B, Fitzgibbon J, Lee G, O’Sullivan G, Shanahan F,
Collins JK. 2001. Probiotic impact on microbial flora, inflammation and tumour development in IL-10knockout mice. Aliment Pharmacol Ther 15:1219–1225.
Ohishi T, Kishimoto Y, Miura N, Shiota G, Kohri T, Hara Y, Hasegawa J, Isemura M. 2002. Synergisticeffects of (–)-epigallocatechin gallate with sulindac against colon carcinogenesis of rats treated withazoxymethane. Cancer Lett 177:49–56.
Okawa T, Niibe H, Arai T, Sekiba K, Noda K, Takeuchi S, Hashimoto S, Ogawa N. 1993. Effect of LC9018combined with radiation therapy on carcinoma of the uterine cervix. A phase III, multicenter, randomized,controlled study. Cancer 72:1949-1954.
Onoue M, Kado S, Sakaitani Y, Uchida K, Morotomi M. 1997. Specific species of intestinal bacteriainfluence the induction of aberrant crypt foci by 1,2-dimethylhydrazine in rats. Cancer Lett 113:179–186.
Orrhage K, Sillerstrom E, Gustafsson JA, Nord CE, Rafter J. 1994. Binding of mutagenic heterocyclic aminesby intestinal and lactic acid bacteria. Mutat Res 311:239–248.
Otte JM, Mahjurian-Namari R, Brand S, Werner I, Schmidt WE, Schmitz F. 2009. Probiotics regulate theexpression of COX-2 in intestinal epithelial cells. Nutr Cancer 61:103–113.
Park E, Kim KT, Kim CJ, Paik HD. 2004. Anticarcinogenic and antigenotoxic effects of Bacilluspolyfermenticus. J Microbiol Biotechnol 14:852–858.
Park E, Jeon GI, Park JS, Paik HD. 2007. A probiotic strain of Bacillus polyfermenticus reduces DMH inducedprecancerous lesions in F344 male rat. Biol Pharm Bull 30(3):569–574.
Parke DV, Ioannides C. 1990. Role of cytochrome P-450 in mouse liver tumor production. Prog Clin BiolRes 331:215–230.
Parkin DM, Bray F, Ferlay J, Pisani P. 2005. Global cancer statistics 2002. CA Cancer J Clin 55:74–108.Payne CM, Bernstein C, Bernstein H, Gerner EW, Garewal H. 1999. Reactive nitrogen species in colon
carcinogenesis. Antioxid Redox Signal 1:449–467.Pedrosa MC, Golner BB, Goldin BR, Barakat S, Dallal GE, Russell RM. 1995. Survival of yogurt-containing
organisms and Lactobacillus gasseri (ADH) and their effect on bacterial enzyme activity in thegastrointestinal tract of healthy and hypochlorhydric elderly subjects. Am J Clin Nutr 61:353–359.
Perdigon G. 1992. Probiotics and the immune state. In: Fuller R, editor. Probiotics: The scientific basis.London: Chapman and Hall. p 145.
Perdigon G, Alvarez S, Pesce de Ruiz Holgado A. 1991. Immunoadjuvant activity of oral Lactobacillus casei:Influence of dose on the secretory immune response and protective capacity in intestinal infections. J DairyRes 58:485–496.
Petzinger E, Weidenbach A. 2000. Mycotoxins in the food chain: The role of ochratoxins (2002) LivestockProduction Science, 76, pp 245–250.
Petzinger E, Ziegler K. 2000. Ochratoxin A from a toxicological perspective. J Vet Pharmacol Ther. 23:91–98.
Piotrowska M, Zakowska Z. 2005. The elimination of Ochratoxin A by lactic acid bacteria strains. Polish JMicrobiol 54:279–286.
Pool-Zobel BL, Munzner R, Holzaapfel H. 1993. Antigenotoxic properties of lactic acid bacteria in theS. typhimurium mutagenicity assay. Nutr Cancer 20:261–270.
Pool-Zobel BL, Neudecker C, Domizlaff I, Ji S, Schillinger U, Rumney C, Moretti M, Vilarini I, Scasellati-Sforzolini R, Rowland IR. 1996. Lactobacillus and Bifidobacteriummediated antigenotoxicity in the colon ofrats. Nutr Cancer 26:365–380.
Rafter JJ. 1995. The role of lactic acid bacteria in colon cancer prevention. Scand J Gastroenterol 30:497–502.Rao AR, Hussain SP. 1988. Modulation of methylcholanthrene-induced carcinogenesis in the uterine cervix
of mouse by indomethacin. Cancer Lett 43:15–19.
494 M. Kumar et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

Ratan RR, Murphy TH, Baraban JM. 1994. Oxidative stress induces apoptosis in embryonic corticalneurons. J Neurochem 62:376–379.
Reddy BS. 1998. Prevention of colon cancer by pre- and probiotics: Evidence from laboratory studies. Br JNutr 80:S219–S223.
Reddy BS, Hamid R, Rao CV. 1997. Effect of dietary oligofructose and inulin on colonic preneoplasticaberrant crypt foci inhibition. Carcinogenesis 18:1371–1374.
Reddy BS, Rivenson A. 1993. Inhibitory effect of Bifidobacterium longum on colon, mammary, and livercarcinogenesis induced by 2-amino-3-methylimidazo[4,5-f]quinoline, a food mutagen. Cancer Res53:3914–3918.
Rescigno M, Granucci F, Citterio S, Foti M, Ricciardi-Castagnoli P. 1999. Coordinated events duringbacteria-induced DC maturation. Immunol Today 20:200–203.
Reynolds SH, Stowers SJ, Patterson RM, Maronpot RR, Aaronson SA, Anderson MW. 1987. Activatedoncogenes in B6C3F1 mouse liver tumors: Implications for risk assessment. Science 237:1309–1316.
Rhee CH, Park HD. 2001. Three glycoproteins with antimutagenic activity identified in Lactobacillusplantarum KLAB21. Appl Environ Microbiol 67:3445–3449.
Rice-Evans C, Burdon R. 1993. Free radical–lipid interactions and their pathological consequences. ProgLipid Res 32:71–110.
Rowland IR, Bearne CA, Fischer R, Pool-Zobel BL. 1996. The effect of lactulose on DNA damage inducedby DMH in the colon of human flora-associated rats. Nutr Cancer 26:37–47.
Rowland IR, Rumney CJ, Coutts JT, Lievense LC. 1998. Effect of Bifidobacterium longum and inulinon gut bacterial metabolism and carcinogen-induced aberrant crypt foci in rats. Carcinogenesis19:281–285.
Saikali J, Picard C, Freitas M, Holt P. 2004. Fermented milks, probiotic cultures, and colon cancer. NutrCancer 49:14–24.
Saito YT, Rowland IR. 1992. Effects of soybean oligosaccharides on the human gut microbiota in in-vitroculture. Microb Ecol Health Dis 5:105–110.
Salminen S, Von Wright A. 1998. Current probiotics—safety assured? Microbial Ecol Health Dis 10:68–77.Schiffrin EJ, Rochat F, Link-Amster H, Aeschlimann JM, Donnet-Hughes A. 1995. Immunomodulation of
human blood cells following the ingestion of lactic acid bacteria. J Dairy Sci 78:491–496.Sekine K, Toida T, Saito M, Kuboyama M, Kawashima T, Hashimoto Y. 1985. A new morphologically
characterized cell wall preparation (whole peptidoglycan) from Bifidobacterium infantis with a higherefficacy on the regression of an established tumor in mice. Cancer Res 45:1300–1307.
Sekine K, Kawashima T, Hashimoto, Y. 1994. Comparison of the TNF-a levels induced by human-derivedBifidobacterium longum and rat-derived Bifidobacterium animalis in mouse peritoneal cells. BifidobacteriaMicrobiota 13:79–89.
Sekine K, Ohta J, Onishi M, Tatsuki T, Shimokawa Y, Toida T, Kawashima T, Hashimoto Y. 1995. Analysisof antitumor properties of effector cells stimulated with a cell wall preparation (WPG) of Bifidobacteriuminfantis. Biol Pharm Bull 18:148–153.
Shackelford LA, Rao DR, Chawan CB, Pulusani SR. 1983. Effect of feeding fermented milk on the incidenceof chemically-induced colon tumours in rats. Nutr Cancer 5:159–164.
Shahani KM, Ayebo AD. 1980. Role of dietary lactobacilli in gastrointestinal microecology. Am J Clin Nutr33:2448–2457.
Sies H. 1991. Oxidative stress: Introduction. In: Sies H, editor. Oxidative stress: Oxidants and antioxidants.San Diego, CA: Academic Press. pp 15–22.
Singh J, Rivenson A, Tomita M, Shimamura S, Ishibashi N, Reddy BS. 1997. Bifidobacterium longum, a lacticacid-producing intestinal bacterium inhibits colon cancer and modulates the intermediate biomarkers ofcolon carcinogenesis. Carcinogenesis 18:833–841.
Souza MF, Tome AR, Rao VS. 1999. Inhibition by the bioflavonoid ternatin of aflatoxin B1-induced lipidperoxidation in rat liver. J Pharm Pharmacol 51:125–129.
Stoner GD, Mukhtar H, 1995. Polyphenols as cancer chemopreventive agents. J Cell Biochem 22:169–180.Szarka CE, Pfeiffer GR, Hum ST, Everley LC, Balshem AM, Moore DF, Litwin S, Goosenberg EB, Frucht
H, Engstrom PF. 1995. Glutathione S-transferase activity and glutathione S-transferase mu expression insubjects with risk for colorectal cancer. Cancer Res 55:2789–2793.
Takagi A, Matsuzaki T, Sato M, Nomoto K, Morotomi M, Yokokura T. 1999. Inhibitory effect of oraladministration of Lactobacillus casei on 3-methylcholanthrene-induced carcinogenesis in mice. MedMicrobiol Immunol (Berl) 188:111–116.
Takagi A, Matsuzaki T, Sato M, Nomoto K, Morotomi M, Yokokura T. 2001. Enhancement of natural killercytotoxicity delayed murine carcinogenesis by a probiotic microorganism. Carcinogenesis 22:599–605.
Cancer-preventing attributes of probiotics 495
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.

Takagi A, Ikemura H, Matsuzaki T, Sato M, Nomoto K, Morotomi M, Yokokura T. 2008. Relationshipbetween the in-vitro response of dendritic cells to Lactobacillus and prevention of tumorigenesis in themouse. J Gastroenterol 43:661–669.
Takeshi M, Akimitsu T, Haruo I, Tetsuya M, Teruo Y. 2007. Intestinal microbiota: Probiotics andautoimmunity. J Nutr 137:798S–802S.
Tavan E, Cayuela C, Antoine JM, Trugnan G, Chaugier C, Cassand P. 2002. Effects of dairy products onheterocyclic aromatic amine-induced rat colon carcinogenesis. Carcinogenesis 23:477–483.
Thyagaraja N, Hosono A. 1993. Antimutagenicity of lactic acid bacteria from ‘Idly’ against food relatedmutagens. J Food Protect 56:1061–1066.
Trush MA, Kensler TW. 1991. An overview of the relationship between oxidative stress and chemicalcarcinogenesis. Free Radic Biol Med 10:201–209.
Usman HA. 1998. Desmutagenicity of milk cultured with Lactobacillus acidophilus strains against mutagenicheated tauco. Food Chem Toxicol 36:805–810.
Vanderhoof JA. 2001. Probiotics: Future directions. Am J Clin Nutr 73:1152S–1155S.Vuillaume M. 1987. Reduced oxygen species, mutation, induction and cancer initiation. Mutat Res
186:43–72.Wollowski I, Rechkemmer G, Pool-Zobel BL. 2001. Protective role of probiotics and prebiotics in colon
cancer. Am J Clin Nutr 73:451S–455S.Yokokura T, Kato I, Mutai M. 1981. Antitumor effect of Lactobacillus casei (LC 9018). In: Mitsuoka T,
editor. Intestinal flora and carcinogenesis. Tokyo: Gakkai-Syuppan Center. pp 72–88.
496 M. Kumar et al.
Int J
Foo
d Sc
i Nut
r D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y M
asse
y U
nive
rsity
on
09/1
9/11
For
pers
onal
use
onl
y.
Copyright © 2022 FDOKUMEN




















