
Primate diversity, habitat preferences, and population density estimates in Noel Kempff Mercado...
15
American Journal of Primatology 46:197–211 (1998) © 1998 Wiley-Liss, Inc. Primate Diversity, Habitat Preferences, and Population Density Estimates in Noel Kempff Mercado National Park, Santa Cruz Department, Bolivia ROBERT B. WALLACE, 1,2 * R. LILIAN E. PAINTER, 1,2 AND ANDREW B. TABER 1 1 Wildlife Conservation Society, 185th Street and Southern Boulevard, Bronx, New York 2 Evolutionary Psychology and Behavioural Ecology Group, Department of Biology, University of Liverpool, Liverpool, England This report documents primate communities at two sites within Noel Kempff Mercado National Park in northeastern Santa Cruz Department, Bolivia. Diurnal line transects and incidental observations were employed to survey two field sites, Lago Caiman and Las Gamas, providing infor- mation on primate diversity, habitat preferences, relative abundance, and population density. Primate diversity at both sites was not particularly high, with six observed species: Callithrix argentata melanura, Aotus azarae, Cebus apella, Alouatta caraya, A. seniculus, and Ateles paniscus chamek. Cebus showed no significant habitat preferences at Lago Caiman and was also more generalist in use of forest strata, whereas Ateles clearly preferred the upper levels of structurally tall forest. Callithrix argentata melanura was rarely encountered during surveys at Lago Caiman, where it preferred low vine forest. Both species of Alouatta showed restricted habitat use and were sympatric in Igapo forest in the Lago Caiman area. The most abundant primate at both field sites was Ateles, with density estimates reaching 32.1 individuals/km 2 in the lowland forest at Lago Caiman, compared to 14.1 individuals/km 2 for Cebus. Both Ateles and Cebus were absent from smaller patches of gallery forest at Las Gamas. These densities are compared with estimates from other Neotropical sites. The diversity of habitats and their different floristic composi- tion may account for the numerical dominance of Ateles within the primate communities at both sites. Am. J. Primatol. 46:197–211, 1998. © 1998 Wiley-Liss, Inc. Key words: primate diversity; habitat use; density; Bolivia INTRODUCTION Line transects have been extensively used throughout the forested tropics as a means of calculating the relative abundance and estimating the density of a wide variety of large (>1 kg) mammals [e.g., Emmons, 1984; Koster & Hart, 1988; Contract grant sponsor: Wildlife Conservation Society. *Correspondence to: Robert B. Wallace, Evolutionary Psychology and Behavioural Ecology Group, School of Biological Sciences, Nicholson Building, University of Liverpool, Liverpool L69 3BX, UK. E- mail: [email protected] Received 18 October 1997; revision accepted 12 May 1998
Transcript of Primate diversity, habitat preferences, and population density estimates in Noel Kempff Mercado...

American Journal of Primatology 46:197–211 (1998)
© 1998 Wiley-Liss, Inc.
Primate Diversity, Habitat Preferences, and PopulationDensity Estimates in Noel Kempff Mercado National Park,Santa Cruz Department, Bolivia
ROBERT B. WALLACE,1,2* R. LILIAN E. PAINTER,1,2 AND ANDREW B. TABER11Wildlife Conservation Society, 185th Street and Southern Boulevard, Bronx, New York2Evolutionary Psychology and Behavioural Ecology Group, Department of Biology,University of Liverpool, Liverpool, England
This report documents primate communities at two sites within NoelKempff Mercado National Park in northeastern Santa Cruz Department,Bolivia. Diurnal line transects and incidental observations were employedto survey two field sites, Lago Caiman and Las Gamas, providing infor-mation on primate diversity, habitat preferences, relative abundance, andpopulation density. Primate diversity at both sites was not particularlyhigh, with six observed species: Callithrix argentata melanura, Aotusazarae, Cebus apella, Alouatta caraya, A. seniculus, and Ateles paniscuschamek. Cebus showed no significant habitat preferences at Lago Caimanand was also more generalist in use of forest strata, whereas Ateles clearlypreferred the upper levels of structurally tall forest. Callithrix argentatamelanura was rarely encountered during surveys at Lago Caiman, whereit preferred low vine forest. Both species of Alouatta showed restrictedhabitat use and were sympatric in Igapo forest in the Lago Caiman area.The most abundant primate at both field sites was Ateles, with densityestimates reaching 32.1 individuals/km2 in the lowland forest at LagoCaiman, compared to 14.1 individuals/km2 for Cebus. Both Ateles andCebus were absent from smaller patches of gallery forest at Las Gamas.These densities are compared with estimates from other Neotropicalsites. The diversity of habitats and their different floristic composi-tion may account for the numerical dominance of Ateles within theprimate communities at both sites. Am. J. Primatol. 46:197–211, 1998.© 1998 Wiley-Liss, Inc.
Key words: primate diversity; habitat use; density; Bolivia
INTRODUCTIONLine transects have been extensively used throughout the forested tropics as
a means of calculating the relative abundance and estimating the density of awide variety of large (>1 kg) mammals [e.g., Emmons, 1984; Koster & Hart, 1988;
Contract grant sponsor: Wildlife Conservation Society.
*Correspondence to: Robert B. Wallace, Evolutionary Psychology and Behavioural Ecology Group,School of Biological Sciences, Nicholson Building, University of Liverpool, Liverpool L69 3BX, UK. E-mail: [email protected]
Received 18 October 1997; revision accepted 12 May 1998

198 / Wallace et al.
Karanth & Sunquist, 1992; White, 1994], particularly arboreal primates [e.g.,Whitesides et al., 1988; Thomas, 1991]. However, it is widely recognized thatthis method provides at best only reasonable density estimates and longer termobservational studies, based on the identification of all individuals in an area,are the only way to accurately assess population densities in a given area [Defler& Pintor, 1985; Brockelman & Ali, 1987]. A sampling effort which yields at least40 and preferably 60–80 encounters for a given target species has been recom-mended [Burnham et al., 1980]. Nevertheless, useful comparisons between areasand/or habitat types have been possible with fewer encounters, and line transectshave proved a popular compromise with many researchers due to the rapidity inwhich a relatively large area can be surveyed.
Although Bolivia has recently been the focus of several primatological fieldstudies, much of this attention has been directed toward true Amazonian forestsin the northern regions of the country, particularly in Pando Department, whereprimate diversity is high and includes the endangered Goeldi’s monkey (Callimicogoeldii) [Freese et al., 1982; Kohlhaas, 1988; Buchanan-Smith, 1991]. In con-trast, primate diversity and distribution are largely undocumented in the exten-sive eastern regions of lowland Bolivia [but see an unpublished 1985 report tothe New York Zoological Society by A.D. Brown and D.I. Rumiz; Braza & Garcia,1988; Wallace et al., in preparation]. Here we present data concerning primatecommunities at two field sites in Noel Kempff Mercado National Park, Depart-ment Santa Cruz, which were collected as part of a general mammalian survey[Painter et al., in preparation].
MATERIALS AND METHODSStudy Area
Noel Kempff Mercado National Park is situated on the Brazilian Shield geo-logical formation in northeastern Santa Cruz Department, Bolivia. The ItenezRiver defines the park’s eastern and northern edges and marks the border withneighboring Brazil. Largely comprised of the Serrania de Huanchaca, a Precam-brian table-mountain or mesa rising 200–400 m above the surrounding lowlands,the park covers 1,500,000 ha and contains an array of markedly differing habitattypes, including open pampas, cerradâo forest, gallery forest, and various formsof tropical lowland forest. This lowland forest is broadly classified as Precam-brian shield humid forest [Killeen et al., 1993]. While no meteorological dataexist for the park itself, the region is characterized by a marked dry season inthe austral winter, a mean annual temperature of 24o C, and annual precipita-tion of approximately 1,500 mm [Killeen, 1996].
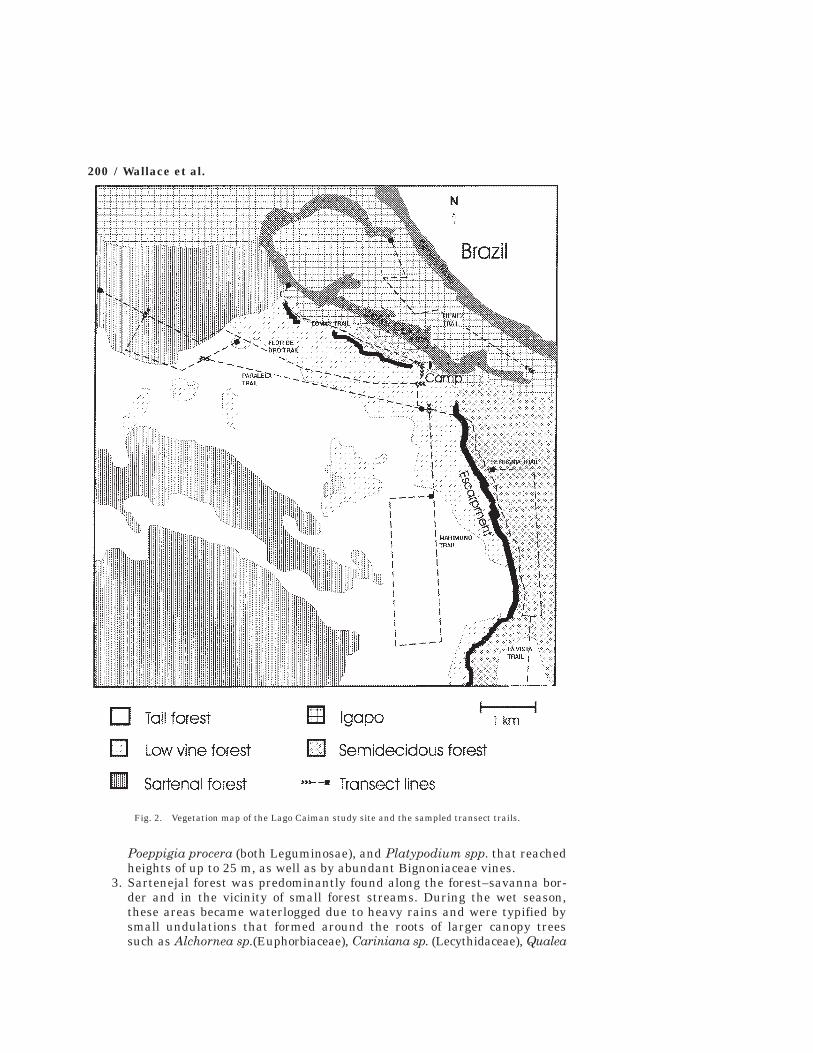
Research was conducted at two sites within the park (see Fig. 1). For 15months between September 1991 and November 1992 research was based at LagoCaiman (lat. 13°, 36′ S, lon. 60°, 55′ W), a large oxbow lake situated at the baseof the northern tip of the Huanchaca escarpment and approximately 21 km upthe Itenez River from the Flor de Oro tourist center. Trails totaling 46.6 km weredistributed across an approximately 50 km2 area (see Fig. 2) and passed througha variety of distinct habitats. The following habitat types were recognized alongthe trails:
1. Tall forest was found on slightly undulating terrain with relatively deepand well-drained clay soils at the base of the escarpment. Emergent andcanopy tree species such as Amburana cearensis, Parkia pendula,

Primate Density in Northeastern Bolivia / 199
Enterolobium sp.(all Leguminosae), Pseudolmedia spp., Brosimum spp.,Clarisia racemosa (all Moraceae), Qualea spp., and Erisma uncinatum (bothVochysiaceae) were typical and attained heights of up to 40 m. Palms suchas Socratea exorrhiza and Euterpe precatoria, as well as smaller trees suchas Theobroma speciosum (Sterculiaceae), Hirtella sp.(Chrysobalanaceae),Inga spp., Platypodium spp.(both Leguminosae), and Pourouma spp(Moraceae), were common in the subcanopy.
2. Low vine forest was found on well-drained slopes at the base of the es-carpment. This habitat was characterized by large canopy and emergenttrees such as Pseudolmedia spp., Casearia gossypiosperma (Flacourtiaceae),Sloanea spp.(Elaeocarpaceae), Celtis chipii (Ulmaceae), Sclerolobium spp.,
Fig. 1. Map of Noel Kempff Mercado National Park and locations of the two field sites: Lago Caiman andLas Gamas.
200 / Wallace et al.
Poeppigia procera (both Leguminosae), and Platypodium spp. that reachedheights of up to 25 m, as well as by abundant Bignoniaceae vines.
3. Sartenejal forest was predominantly found along the forest–savanna bor-der and in the vicinity of small forest streams. During the wet season,these areas became waterlogged due to heavy rains and were typified bysmall undulations that formed around the roots of larger canopy treessuch as Alchornea sp.(Euphorbiaceae), Cariniana sp. (Lecythidaceae), Qualea
Fig. 2. Vegetation map of the Lago Caiman study site and the sampled transect trails.

Primate Density in Northeastern Bolivia / 201
sp., Sloanea spp., Guatteria spp. (Annonaceae), Maquira sp. (Moraceae),Pourouma spp., Pseudolmedia spp., and Brosimum spp. that reached heightsof 35 m. Palms such as Euterpe precatoria and Socratea exorrhiza were com-mon in the subcanopy stratum.
4. Igapo forest was found on either side of the Itenez River. During the wetseason (November–March), this forest was flooded, giving rise to a fairlyopen understory in the dry season. The canopy in this habitat was charac-terized by Macrolobium acaciifolium, Acosmium nitens (both Leguminosae),Ficus guianensis, Ficus trigona (both Moraceae), Hevea brasiliensis(Euphorbiaceae), and Cochlospermum orinicense (Cochlospermaceae) thatreached heights of up to 30 m. The understory included Bactris gasipaes(Palmae), Mouriri apiranga (Melastomataceae), and Salacia elliptica(Hippocrateaceae).
5. Semideciduous forest was a broad habitat classification representing theupland forests of the study site including cerradâo forest. This was thedominant vegetation found on the top of the northern regions of theHuanchaca escarpment as well as on lower surrounding hills. The canopyof this habitat ranged between 10 and 20 m and included species such asSchinopsis sp. (Leguminosae), Jacaranda sp. (Bignoniaceae), Sterculiaapetala (Sterculiaceae), and Crepidospermum sp. (Burseraceae). The un-derstory included Talisia esculenta (Sapindaceae), Roupala montana(Proteaceae), and Abuta grandiflora (Menispermaceae).
The second field site, Las Gamas (lat. 14°, 48′ S, lon. 60°, 23′ W), was around anabandoned airstrip situated on top of the table-mountain in the southern region ofthe park and was visited for 5 weeks during March/April 1993. Trails were locatedin three patches of gallery forest and totaled 10.3 km in length (see Fig. 3). Two ofthese gallery forest patches, patch A (approximately 41 ha) and patch B (approxi-mately 50 ha), were relatively small, isolated, and surrounded by open grasslandand a variety of cerrado scrub vegetation types. The larger third patch, patch C(approximately 1,535 ha), was located on the edge of the escarpment, with one sidebordering the lowland forest below. The canopy of this patch often reached 30 m andwas characterized by Aspidosperma cf. marcgravianum (Apocynaceae), Erismauncinatum, and Qualea sp. [Arroyo Padilla, 1995]. Other dominant tree species in-cluded Miconia chrysophylla (Melastomataceae), Pseudolmedia cf. rigida, and anunidentified Lauraceae [Arroyo Padilla, 1995]. The understory was typified byPsychotria sp. (Rubiaceae), Cordia nodosa (Boraginaceae), Vitex sp. (Verbenaceae),and Ouratea sp. (Ochnaceae) [Arroyo Padilla, 1995].
MethodsPopulation density estimates were determined using line transect methodol-
ogy [Burnham et al., 1980; Brockelman & Ali, 1987]. Transects were conductedby one to three observers in fair weather conditions along established trails. AtLago Caiman, 503.5 km of line transect were conducted between 6:00 and 11:15h and 15:00 and 18:00 h. At Las Gamas, 50.4 km were walked between 6:30 and12:15 h. Details regarding the location, length, and replication of transect trailsand habitats present on each trail are listed in Table I. Although each transectwalk did not necessarily total the trail length, efforts were made to sample allsections of the trails evenly. Each transect was independent because at least 2days passed before a trail was resampled. The rate at which transects were walkedranged between 1 and 3 km/h and depended on trail conditions. Periods of walk-

202 / Wallace et al.
Fig. 3. Vegetation map of the Las Gamas study site and the three sampled gallery forest patches.
ing were regularly interspersed with brief listening stops to increase the prob-ability of detecting cryptic primate groups.
The following information was recorded for all primate groups encountered ontransect: species, group size (and, where possible, age/sex composition), date and timedetected, observation duration, transect position, habitat type, perpendicular distancefrom the transect trail to the estimated geometric centroid of the group, and verticalposition of the group in forest strata. Only “good” observations were used for estimat-

Primate Density in Northeastern Bolivia / 203
ing primate group sizes. Good observations were those in which the observer gained aprolonged view of the troop, allowing an accurate count with a reasonable certaintythat no additional unobserved individuals were present. The vertical position of eachprimate troop encountered was classified as in either the upper levels of the forest(emergents, upper and middle canopy), medium levels (subcanopy and understory), orlower levels (shrub layer) or as terrestrial. Additionally, all nontransect incidental ob-servations were noted and appropriate parameters recorded.
Transect information was analyzed using the DISTANCE computer programwhich employs Fourier Series transformations [Laake et al., 1991; Buckland et al.,1993]. The independence of observations on a given transect was maintained byexcluding any observation of the same species that occurred within 200 m of anoriginal observation. Due to problems of data heaping, perpendicular distances weregrouped for analysis, which provided more accurate fits to the various estimatormodels used in the DISTANCE analysis. Heaping was largely a result of the diffi-culty in accurately assessing, in dense forest vegetation, the geometric centroid of anobserved primate group. Similarly, as seemed appropriate for each primate species,perpendicular distance data were truncated to remove outliers from the analysis.
To determine the proportion of habitat types occurring along the transect trails,a habitat classification, using the habitats described above, was assigned every 100m along each trail. Due to logistical constraints and the desire to assess specifichabitat types, some transect trails were placed within certain habitats and not dis-tributed in a random fashion. Thus, the proportion of habitat occurring along thetrails does not necessarily reflect the relative abundance of the Igapo forest andsemideciduous forest habitats within the study area. Statistical analysis follows Siegeland Castellan [1988], and significance levels were set at the P < 0.05 level.
RESULTSPrimate Diversity
During the study, the following primate species were registered at LagoCaiman: silvery marmoset (Callithrix argentata melanura), owl monkey (Aotus
TABLE I. Details of Transect Routes, Sampling Effort, and Habitat Types at LagoCaiman and Las Gamas*
Number of Trail Total distanceTrail name walks length (km) Habitat types censused
Lago CaimanFlor de Oro 25 7.6 85.36 Low vine, tall, and sartenejal forestsParalela 23 7.9 110.16 Low vine, tall, and sartenejal forestsMarimono 27 10.5 116.11 Low vine, tall, and sartenejal forestsSerrania 24 10 98.18 Semideciduous and low vine forestsLomas 13 2.9 34.7 Semideciduous forestLa Vista 5 3.1 13.34 Semideciduous forestItenez 13 4.6 45.65 Igapo forestTotal 130 46.6 503.5
Las GamasPatch A 4 1.02 4.08 Gallery forestPatch B 4 2.01 4.02 Gallery forestPatch C 18 7.25 42.3 Gallery forest
Total 26 10.28 50.4
*Each transect walk did not necessarily total the trail length; thus, the product of trail length and numberof walks does not always equal the total distance censused for each trail.
204 / Wallace et al.
azarae), brown capuchin monkey (Cebus apella), black howler monkey (Alouattacaraya), red howler monkey (Alouatta seniculus), and black spider monkey (Atelespaniscus chamek). Four additional primate species were registered on the Bra-zilian side of the Itenez River during the course of this study: dusky titi monkey(Callicebus brunneus), bare-eared squirrel monkey (Saimiri ustus), monk sakimonkey (Pithecia irrorata), and red-faced bearded saki monkey (Chiropotesalbinasus) [Wallace et al., 1996]. At the Las Gamas study site, the following pri-mates were registered: Aotus azarae, Cebus apella, Alouatta caraya, and Atelespaniscus chamek.
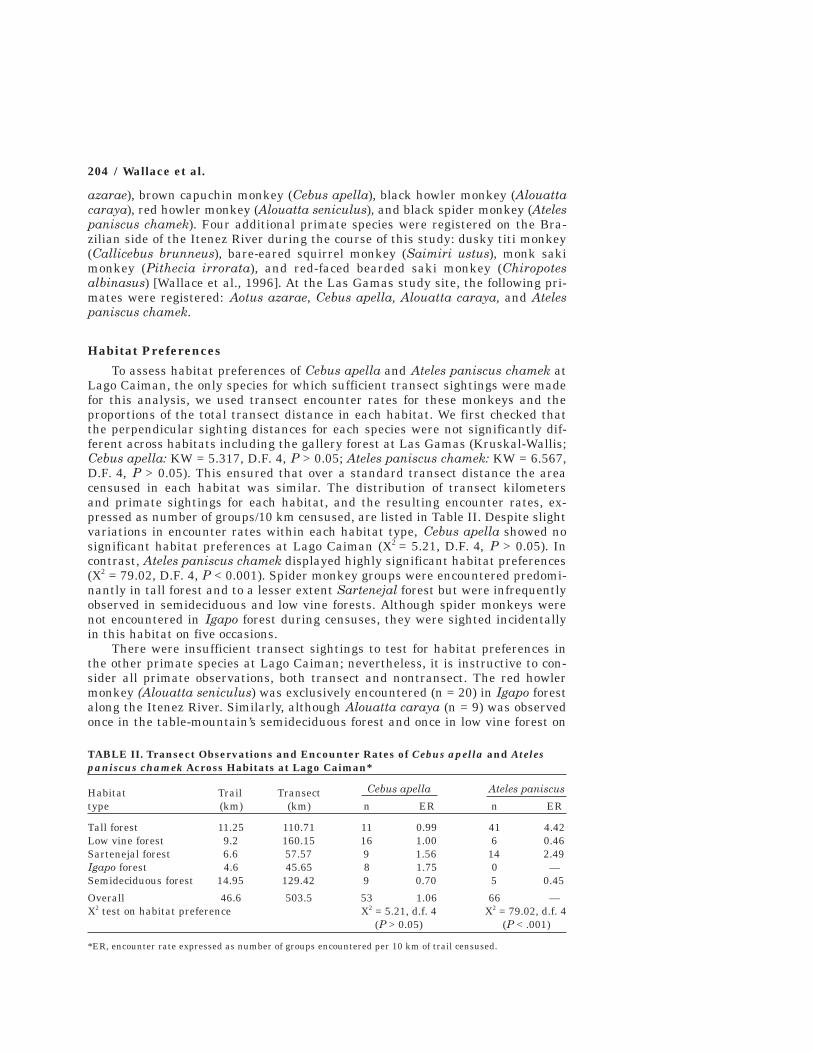
Habitat PreferencesTo assess habitat preferences of Cebus apella and Ateles paniscus chamek at
Lago Caiman, the only species for which sufficient transect sightings were madefor this analysis, we used transect encounter rates for these monkeys and theproportions of the total transect distance in each habitat. We first checked thatthe perpendicular sighting distances for each species were not significantly dif-ferent across habitats including the gallery forest at Las Gamas (Kruskal-Wallis;Cebus apella: KW = 5.317, D.F. 4, P > 0.05; Ateles paniscus chamek: KW = 6.567,D.F. 4, P > 0.05). This ensured that over a standard transect distance the areacensused in each habitat was similar. The distribution of transect kilometersand primate sightings for each habitat, and the resulting encounter rates, ex-pressed as number of groups/10 km censused, are listed in Table II. Despite slightvariations in encounter rates within each habitat type, Cebus apella showed nosignificant habitat preferences at Lago Caiman (X2 = 5.21, D.F. 4, P > 0.05). Incontrast, Ateles paniscus chamek displayed highly significant habitat preferences(X2 = 79.02, D.F. 4, P < 0.001). Spider monkey groups were encountered predomi-nantly in tall forest and to a lesser extent Sartenejal forest but were infrequentlyobserved in semideciduous and low vine forests. Although spider monkeys werenot encountered in Igapo forest during censuses, they were sighted incidentallyin this habitat on five occasions.
There were insufficient transect sightings to test for habitat preferences inthe other primate species at Lago Caiman; nevertheless, it is instructive to con-sider all primate observations, both transect and nontransect. The red howlermonkey (Alouatta seniculus) was exclusively encountered (n = 20) in Igapo forestalong the Itenez River. Similarly, although Alouatta caraya (n = 9) was observedonce in the table-mountain’s semideciduous forest and once in low vine forest on
TABLE II. Transect Observations and Encounter Rates of Cebus apella and Atelespaniscus chamek Across Habitats at Lago Caiman*
Habitat Trail Transect Cebus apella Ateles paniscus
type (km) (km) n ER n ER
Tall forest 11.25 110.71 11 0.99 41 4.42Low vine forest 9.2 160.15 16 1.00 6 0.46Sartenejal forest 6.6 57.57 9 1.56 14 2.49Igapo forest 4.6 45.65 8 1.75 0 —Semideciduous forest 14.95 129.42 9 0.70 5 0.45Overall 46.6 503.5 53 1.06 66 —X2 test on habitat preference X2 = 5.21, d.f. 4 X2 = 79.02, d.f. 4
(P > 0.05) (P < .001)
*ER, encounter rate expressed as number of groups encountered per 10 km of trail censused.

Primate Density in Northeastern Bolivia / 205
one of its slopes, seven (77.8%) sightings of this species were in Igapo forest inthe Lago Caiman vicinity. These two congeners had sympatric distribution inthis region: at Flor de Oro both species frequented Igapo forest on the oppositebank of the Itenez River, and on three occasions groups of each species wereobserved in close proximity, once in adjacent Ficus sp. trees. For Callithrixargentata melanura, low vine forest appeared to be the most important habitat,accounting for 77.8% of all sightings (n = 9). Incidental nocturnal observationsregistered Aotus azarae in tall forest and low vine forest at Lago Caiman.
Although at Las Gamas only one forest habitat type was surveyed, censuseswere conducted in separate patches of gallery forest which differed both in sizeand geography. In the two smaller and isolated patches (approximately 50 ha),primate groups were never encountered on transects and overall were only ob-served on three occasions: Alouatta caraya was observed twice in patch B, andAotus azarae was heard calling once in patch A. In the third and larger patch C(approximately 1,535 ha), which also bordered the lowland forest below the es-carpment, Cebus apella was observed once during censuses, with a resulting en-counter rate of 0.24 groups/10 km censused, and five times overall. Ateles paniscuschamek was encountered on 15 occasions during censuses, an encounter rate of3.55 groups/10 km censused, and on 28 occasions overall. Although samples aresmall, it appears that both these species were absent in the smaller and isolatedgallery forest patches. However, if we include the transect kilometers from thesepatches, resulting encounter rates drop to 0.2 groups/10 km for Cebus apella and2.98 groups/10 km for Ateles paniscus.
Vertical StrataClear differences in forest strata use existed among the primate species at
the study sites. Callithrix argentata melanura (n = 8) was exclusively encoun-tered in the middle levels of the forest. Aotus azarae (n = 9) also showed a prefer-ence for this forest stratum, with 77.8% of sightings at this level and the remainingtwo observations in the upper strata. Cebus apella (n = 114) seems to be muchmore of a generalist, with 43.9% of sightings in the upper strata and 50% in themiddle levels of the forest. Additionally, on six occasions Cebus apella was ob-served in the shrub layer and once on the ground in semideciduous forest. Thetwo Alouatta species were also found predominantly in the middle and upperlevels of the forest, although A. seniculus (n = 20) was once observed drinkingfrom a beach on the Itenez River in the dry season. Spider monkeys (n = 175)clearly preferred emergent trees and upper and middle canopy strata of the for-est, with upper levels accounting for 90.9% of sightings and the remaining obser-vations in the middle stratum.
Average Group SizeThe observed mean group size for Callithrix argentata melanura was 6.2
individuals (n = 5, SD ±1.9, range 4–9). Aotus azarae had a mean group size ofthree individuals (n = 3, SD ±1.7, range 2–5). The two species of howler monkeyshowed similar mean group sizes: Alouatta caraya at four individuals (n = 3, SD±2.6, range 1–6) and A. seniculus at 3.3 individuals (n = 18, SD ±2, range 1–7).Cebus apella had the largest mean group size of the primate species examined,with nine individuals (n = 47, SD ±5.7, range 1–25). Ateles paniscus chamekshowed highly variable subgroup size, with a mean of 6.2 individuals (n = 96, SD±5.3, range 1–30). For Ateles paniscus chamek, we also calculated the mean adult/

206 / Wallace et al.
subadult subgroup size by eliminating dependent offspring from the analysis.This resulted in a mean adult/subadult subgroup size of 4.5 individuals; thus,juveniles and infants made up approximately 28.4% of the total population inthe Lago Caiman vicinity.
In addition, for those species with sufficient encounters, we examined varia-tions in mean group size across the recognized habitat types at Lago Caimanand Las Gamas. Cebus apella showed no significant differences in group sizebetween habitats (Kruskal-Wallis; KW = 6.603, D.F. 5, P > 0.05). For Atelespaniscus, however, habitat type had a significant effect on observed subgroupsize (Kruskal-Wallis; KW = 12.133, D.F. 4, P< 0.02). In low vine forest, the meansubgroup size for Ateles paniscus was 4.6 individuals (n = 7, SD ±4, range 1–13),in tall forest the mean was 5.8 individuals (n = 55, SD ±4.2, range 1–20), and ingallery forest the mean was 3.6 individuals (n = 16, SD ±2.2, range 1–7). Sartenejalforest displayed the highest mean subgroup size, with 10.7 individuals (n = 12,SD ±8.5, range 2–30), with semideciduous forest approaching this at 7.5 indi-viduals (n = 4, SD ±6.6, range 2–17).
Population Density EstimatesPopulation density estimates were possible only for Cebus apella and Ateles
paniscus due to insufficient transect sightings for the other primate species withinthe park. Since the encounter rates for Cebus apella were not significantly differ-ent between habitats, we calculated the overall density at the Lago Caiman studysite using transect encounters in all habitats. Perpendicular distance data weretruncated at 36 m on either side of the transect, which resulted in a total of 53sightings. Of the various estimator models tested within the DISTANCE pro-gram, the uniform cosine model provided the best fit to the data and resulted inan overall density estimate of 14.1 Cebus apella individuals/km2 (confidence lim-its, 7.8–25.5).
Due to significant differences in encounter rates between habitats, an over-all population density estimate was not possible for Ateles paniscus. Instead, weestimated the density of spider monkeys for three amalgamated lowland habitattypes for which the trails had been randomly placed: low vine forest, Sartenejalforest and tall forest. The perpendicular distance data were truncated at 45 m oneither side of the transect, which resulted in a total of 61 sightings. Again theuniform cosine model provided the best fit to the data, resulting in a densityestimate of 32.1 Ateles paniscus individuals/km2 of lowland forest (confidencelimits, 21.9–47.0). This results in a density estimate for spider monkey adults/subadults of 23 per km2, calculated by subtracting the proportion of juveniles.
DISCUSSIONIn terms of species number and composition, the primate community within
Noel Kempff Mercado National Park is similar to other documented areas ineastern Santa Cruz Department [Braza & Garcia, 1988; Wallace et al., in prepa-ration]. Callithrix was observed infrequently at Lago Caiman (0.1 groups en-countered per 10 km censused), as is the case at several nearby locations [Wallaceet al., in preparation]. Their apparent preference for lower vine forests and meangroup size in this region is comparable to several other studies on congeners[Stevenson & Rylands, 1988]. The Aotus genus is known to occur in a variety ofhabitats in the Neotropics, and the mean group size of three individuals at LagoCaiman is similar to other reported sites [Robinson et al., 1987].

Primate Density in Northeastern Bolivia / 207
The apparent absence of the two howler monkey species from the majority ofthe habitats considered is surprising given Alouatta’s well-documented habitatadaptability [Crockett & Eisenberg, 1987] and numerical dominance in severalNeotropical primate communities with few species [Emmons, 1984]. Interestingly,sloths also appear scarce and were not observed at either field site. The lack ofarboreal folivores within the nonriverine forests may be a feature of the Brazil-ian Shield, given their absence or scarcity in other nearby areas [Painter et al.,in preparation]. Possible explanations for this absence are the floristic composi-tion of these forests and/or poor nutrient background of the soils of this region[PLUS-CORDECRUZ, 1994], which may limit folivory as a dietary niche. Simi-larly, Alouatta seniculus is reported as widespread in Colombia but rare awayfrom rivers and lake shores [Defler, 1994]. Recently, Peres [1997] demonstratedthat distance from a major white water river, forest structure, seasonality, andcorrelates of soil fertility were useful in predicting the density of howler mon-keys in Brazilian Amazonia once hunting pressure had been controlled for andalso noted the lack of other arboreal folivores in low density howler monkey sites.
The habitat adaptability of Cebus apella is thought to be related to its gener-alist diet; capuchins opportunistically consume fruit, flowers, leaves, palm nuts,and insects [Terborgh, 1983]. The population density estimate (14.1 individuals/km2), mean group size, and variability in group size of Cebus apella at LagoCaiman is similar to previous studies [see Robinson & Janson, 1987; Janson,1988]. Nevertheless, the Lago Caiman density estimate is considerably less thanthe 38.9 individuals/km2 reported during less extensive surveys in the southernregions of the park [Braza & Garcia, 1988]. For Cebus and Ateles, the size of thetwo smaller gallery forest patches at Las Gamas may prevent groups of a viablesize taking hold [see Lovejoy et al., 1986]. The third patch was considerably largerand floristically different [T. Killeen, personal communication] and bordered thecontinuous forest below the escarpment, with some areas being forested steepslopes. Seasonal movements between the continuous forest below and the uplandgallery forest would be theoretically possible, especially as we have observed Atelesand Alouatta caraya climbing 200 m cliffs in the Lago Caiman vicinity. It isimportant to underline the brevity of the field work at Las Gamas and the limi-tations this imposes on data interpretation. Clearly, Ateles was abundant in patchC during the surveys; however, this abundance may be a seasonal phenomenon.
The preference for the tall forest and structurally similar Sartenejal habitatsdisplayed by Ateles at Lago Caiman mirrors the results from previous studies inColombia and Peru [Klein & Klein, 1976; Terborgh, 1983]. In Surinam, vanRoosmalen [1985] describes Ateles as preferring high forest with only occasionalforays into other forest types when a particularly abundant fruit resource is sea-sonally available. The overall observed mean subgroup size of 6.2 individuals, or4.5 adults/subadults, is higher than reported for longer term studies on conge-ners in Colombia, Guatemala, Surinam, Peru, and Costa Rica [McFarlandSymington, 1988a; Chapman, 1990]. In addition, proportions of juveniles/infantsreported at three of the above sites are similar to that presented here [vanRoosmalen, 1985; McFarland Symington 1988b; Chapman, 1990]. The variabilityin observed subgroup size and the differences in subgroup size between habitatsare as expected considering the well-documented fission–fusion social system ofthis genus [van Roosmalen, 1985; Chapman, 1990; McFarland Symington 1988c].
The lowland forest density estimate for Ateles at Lago Caiman of 32.1 indi-viduals/km2 is one of the highest reported in the literature [McFarland Symington,1988a]. The only study which documented a higher density estimate, 84.5 indi-viduals/km2, was conducted in the southern regions of Noel Kempff Mercado Na-

208 / Wallace et al.
tional Park [Braza & Garcia, 1988]. Braza and Garcia [1988] fail to detail theanalytical methods used to reach this estimate, but, due to the small number ofencounters in the study (n = 11), their estimate should be treated with caution.Regardless, the subgroup encounter rates reported for Lago Caiman (1.86 for thethree lowland habitats) and those calculated from Braza and Garcia [1988] (11encounters in 45 km of transect, giving 2.44 groups/10 km) are similar. Giventhat this region represents a southern limit for this genus, these encounter ratesare perhaps surprising, especially when one considers that in nearby areas en-counter rates are appreciably lower than those reported for the park [Wallace etal., in preparation].
McFarland Symington [1988a] assessed several factors which might explaindifferences in Ateles population densities among four Neotropical sites and sug-gested that while community effects such as the abundance of arboreal frugivo-rous competitors and potential predators may be contributory factors, they failedto explain the observed variation. Noel Kempff Mercado National Park holds afull complement of larger predators, with jaguar (Panthera onca), puma (Pumaconcolor), harpy eagle (Harpia harpyja), and crested eagle (Morphnus guianensis)all present [Killeen, 1996]. Thus, while we have no data regarding the relativeabundances of these predators, it seems that a lack of predation is an unlikelyexplanation for the high Ateles densities within the park.
In terms of primate community structure, the Guatemalan and Costa Ricanstudy sites which have high densities of Ateles and low primate diversity[McFarland Symington 1988a, Chapman 1990] appear similar to Lago Caiman.These observations suggest that the number of frugivorous primate competitorspresent in an area may be inversely correlated with spider monkey densities.However, as McFarland Symington [1988a] noted, the Peruvian site rather con-tradicts this hypothesis, as it features high spider monkey densities and a richprimate assemblage.
Floristic diversity, habitat productivity, and related factors such as the amountand seasonality of rainfall also failed to explain population densities [McFarlandSymington, 1988a]. Initially, this may appear surprising, since in general foodavailability is considered the ultimate limiting factor on primate population den-sity, though other factors, such as access to water, climate, disease, and preda-tion, are certainly important [Dunbar, 1987]. However, as Cant [1980] has argued,this limitation is not a reflection of the overall annual productivity of a habitat,which may far exceed the energetic requirements of the resident primate com-munity [e.g., Coelho et al., 1976]. Rather, populations are constrained by resourcedensity at times of relative food scarcity. These relative resource droughts maybe seasonal in nature or products of rarer “ecological crunch” events [cf. Wiens,1977], such as the lack of fruiting in certain “keystone” resources [cf. Terborgh,1986], massive forest fruit failures, or severe climatic conditions [Dittus, 1977;Cant, 1980; Foster, 1982; Terborgh, 1986]. The fruit resources which make upthe majority of spider monkey diets are relatively scarce compared to other po-tential primate dietary constituents, ephemeral in nature, and patchily distrib-uted, and they are known to seasonally fluctuate in density [Leigh & Windsor,1982; Howe, 1984; Richard, 1985].
Given the apparent importance of keystone resources for tropical forestfrugivores, we tentatively suggest that the diversity of floristically different habi-tats available at the Lago Caiman site offers a year-round abundance of fruitresources. Indeed, all semideciduous forest observations of Ateles occurred in a 2month period (October–November 1992), suggesting a seasonal use of this habi-tat. This explanation may also account for the larger Ateles subgroup sizes ob-

Primate Density in Northeastern Bolivia / 209
served in less preferred habitats. Preliminary results from a recently completedecological study of a spider monkey community at Lago Caiman suggest thatseasonal use of different habitats is an important feature of spider monkey ecol-ogy in this region [Wallace, unpublished data]. This argument would also applyto the Las Gamas site and Braza and Garcia’s [1988] surveys in the south of thepark and may go some way in explaining the clear numerical and biomass domi-nance of Ateles within the primate communities at these sites.
CONCLUSIONS1. Primate diversity and species composition at Noel Kempff Mercado Na-
tional Park was typical of the northeastern regions of Santa Cruz Department,Bolivia.
2. Callithrix argentata melanura was infrequently encountered at LagoCaiman and preferred low vine forest. Incidental observations registered Aotusazarae in low vine, tall, and gallery forests of the park.
3. The park includes two species of Alouatta which displayed narrow habitatuse at Lago Caiman. Alouatta seniculus was exclusively encountered in Igapoforest, where it is sympatric with A. caraya. Alouatta caraya was the only diur-nal species encountered in the smaller patches of gallery forest at Las Gamas.
4. At Lago Caiman, Cebus apella showed a generalized use of habitat andvertical strata. The population density estimate for this site was 14.1 individu-als/km2. Cebus was infrequently encountered in the larger patch of gallery forestat Las Gamas.
5. The population density estimate of 32.1 individuals/km2 for Ateles paniscuschamek in lowland forest at Lago Caiman is one of the highest reported in theliterature. Ateles clearly preferred tall and structurally similar Sartenejal forestsand was predominantly encountered in the upper levels of the forest. The en-counter rate for Ateles at Las Gamas was also particularly high in the largerpatch of gallery forest.
6. The diversity of floristically different habitats is suggested as an explana-tion for the numerical dominance of Ateles within the primate communities atboth field sites.
ACKNOWLEDGMENTSThe research was funded as part of a grant to A. Taber from the Wildlife Con-
servation Society (WCS). Considerable logistical support was gratefully received fromF.A.N. (Fundación Amigos de la Naturaleza) at the administrative level in SantaCruz and while in the field from all those resident at Flor de Oro, particularly thepark guards. Thanks also go to Robin Foster, who visited the Lago Caiman field siteand provided invaluable information regarding the vegetation of the predominanthabitat types. John Robinson and four anonymous reviewers provided valuable com-ments on earlier drafts of the manuscript. Assistance in the field was capably pro-vided by several Bolivian undergraduate students, particularly Jose-Carlos Herrera.
REFERENCES
Arroyo Padilla, L. Estructura y composiciónde una isla de bosque y un bosque degalería en el Parque Nacional Noel KempffMercado. Graduate thesis, UniversidadAutónoma Gabriel Rene Moreno, SantaCruz, 1995.
Braza, F.; Garcia, J.E. Rapport preliminairesur les singes de la region montagneusede Huanchaca, Bolivie. FOLIA PRIMA-TOLOGIA 49:182–186, 1988.
Brockelman, W.Y.; Ali, R. Methods of sur-veying and sampling forest primate popu-

210 / Wallace et al.lations. Pp. 23–62 in PRIMATE CONSER-VATION IN THE TROPICAL RAIN FOR-EST. C.W. Marsh; R.A. Mittermeier, eds.New York, Alan R. Liss, 1987.
Buchanan-Smith, H. Field observations ofGoeldi’s monkey, Callimico goeldii, innorthern Bolivia. FOLIA PRIMATOLOGIA57:102–105, 1991.
Buckland, S.T.; Anderson, D.R.; Burnham,K.P.; Laake, J.L. DISTANCE SAMPLING:ESTIMATING ABUNDANCE OF BIO-LOGICAL POPULATIONS. London, Chap-man & Hall, 1993.
Burnham, K.P.; Anderson, D.R.; Laake, J.L.Estimation of density from line transectsampling biological populations. WILD-LIFE MONOGRAPHS 72: 1–202, 1980.
Cant, J.G.H. What limits primates? PRI-MATES 21:538–544, 1980.
Chapman, C. Association patterns of spidermonkeys: The influence of ecology and sexon social organization. BEHAVIORALECOLOGY AND SOCIOBIOLOGY 26:409–414, 1990.
Coehlo, A.M.; Bramblett, C.A.; Quick, L.A.;Bramblett, S.S. Resource availability andpopulation density in primates: A socio-bioenergetic analysis of the energy bud-gets of Guatemalan howler and spidermonkeys. PRIMATES 17:63–80, 1976.
Crockett, C.M.; Eisenberg, J.F. Howlers:Variations in group size and demography.Pp. 54–68 in PRIMATE SOCIETIES. B.B.Smuts; D.L. Cheney; R.M. Seyfarth; R.W.Wrangham; T.T. Struthsaker, eds. Chicago,University of Chicago Press, 1987.
Defler, T.R. La conservacion de primates enColombia. TRIANCA 5:255–287, 1994.
Defler, T.R.; Pintor, D. Censusing primatesby transect in a forest of known primatedensity. INTERNATIONAL JOURNAL OFPRIMATOLOGY 6:243–259, 1985.
Dittus, W.P.J. The social regulation of popu-lation density and age-sex distribution inthe toque monkey. BEHAVIOUR 63:281–322, 1977.
Dunbar, R.I.M. Demography and reproduc-tion. Pp. 240–249 in PRIMATE SOCIETIES.B.B. Smuts; D.L. Cheney; R.M. Seyfarth;R.W. Wrangham; T.T. Struthsaker, eds. Chi-cago, University of Chicago Press, 1987.
Emmons, L.H. Geographical variations indensities and diversities of non-flyingmammals in Amazonia. BIOTROPICA16:210–222, 1984.
Foster, R.B. Famine on Barro Colorado Island.Pp. 201–212 in THE ECOLOGY OF ATROPICAL FOREST: SEASONALRHYTHMS AND LONG TERM CHANGES.E.G. Leigh; A.S. Rand; D.M. Windsor, eds.Washington, DC, Smithsonian InstitutionPress, 1982.
Freese, C.H.; Heltne, P.G.; Castro, R.N.;
Whitesides, G. Patterns and determinantsof monkey densities in Peru and Bolivia,with notes on distributions. INTERNA-TIONAL JOURNAL OF PRIMATOLOGY3:53–90, 1982.
Howe, H.F. Implications of seed dispersalby animals for tropical reserve manage-ment. BIOLOGICAL CONSERVATION30:261–281, 1984.
Janson, C.H. Food competition in brown ca-puchin monkeys (Cebus apella): Quantita-tive effects of group size and treeproductivity. BEHAVIOUR 105:53–76, 1988.
Karanth, K.U.; Sunquist, M.E. Populationstructure, density and biomass of largeherbivores in the tropical forests of Naga-rahole, India. JOURNAL OF TROPICALECOLOGY 8:21–35, 1992.
Killeen, T.J. HISTORIA NATURAL ANDBIODIVERSIDAD DE PARQUE NA-CIONAL “NOEL KEMPFF MERCADO,”SANTA CRUZ, BOLIVIA. Plan de Manejo—Componente Cientifico. Santa Cruz, Bolivia,Museo de Historia Natural Noel KempffMercado and Missouri Botanical Garden,1996.
Killeen, T.J.; García, E.; Beck, S.G. GUÍADE ÁRBOLES DE BOLIVIA. La Paz,Herbario Nacional de Bolivia and MissouriBotanical Garden, 1993.
Klein, L.L.; Klein, D.J. Neotropical pri-mates, aspects of habitat usage, popula-tion density, and regional distribution inLa Macarena, Columbia. Pp. 70–78 inNEOTROPICAL PRIMATES: FIELDSTUDIES AND CONSERVATION. R.W.Thorington; P.G. Heltne, eds. Washington,DC, National Academy of Sciences, 1976.
Kohlhaas, A.K. Primate populations innorthern Bolivia. PRIMATE CONSERVA-TION 9:93–97, 1988.
Koster, S.H.; Hart, J.A. Methods of estimat-ing ungulate populations in tropical for-ests. AFRICAN JOURNAL OF ECOLOGY26:117–126, 1988.
Laake, J.L.; Buckland, S.T.; Anderson, D.R.;Burnham, K.P. DISTANCE USER’S GUIDE.Fort Collins, Colorado Cooperative Fish andWildlife Research Unit, Colorado State Uni-versity, 1991.
Leigh, E.G.; Windsor, D.S. Forest produc-tion and regulation of primary consumerson Barro Colorado Island. Pp. 111–122 inTHE ECOLOGY OF A TROPICAL FOR-EST: SEASONAL RHYTHMS AND LONGTERM CHANGES. E.G. Leigh; A.S. Rand;D.S. Windsor, eds. Washington, DC, Smith-sonian Institution Press, 1982.
Lovejoy, T.E.; Bierregard, R.O., Jr.; Rylands,A.B.; Malcolm, J.R.; Quintela, C.E.; Harper,L.H.; Brown, K.S.; Powell, A.H.; Powell,G.V.N.; Schubart, H.O.R.; Hays, M.B. Edgeand other effects of isolation on Amazon for-

Primate Density in Northeastern Bolivia / 211
est fragments. Pp. 257–285 in CONSERVA-TION BIOLOGY: THE SCIENCE OF SCAR-CITY AND DIVERSITY. M.E. Soule, ed.Sunderland, MA, Sinauer, 1986.
McFarland Symington, M. Environmentaldeterminants of population densities inAteles. PRIMATE CONSERVATION 9:74–79, 1988a.
McFarland Symington, M. Demography,ranging patterns, and activity budgets ofblack spider monkeys (Ateles paniscuschamek) in the Manu National Park, Peru.AMERICAN JOURNAL OF PRIMATOL-OGY 15:45–67, 1988b.
McFarland Symington, M. Food competitionand foraging party size in the black spi-der monkey (Ateles paniscus chamek).BEHAVIOUR 105:117–134, 1988c.
Peres, C.A. Effects of habitat quality andhunting pressure on arboreal folivore den-sities in Neotropical forests: A case studyof howler monkeys (Alouatta spp.). FOLIAPRIMATOLOGIA 68:199–222, 1997.
PLUS-CORDECRUZ. PLAN DE USO DESUELOS TOTAL DEL DEPARTAMENTODE SANTA CRUZ. Santa Cruz, Bolivia,Cordecruz, 1994.
Richard, A.F. PRIMATES IN NATURE. NewYork, W.H. Freeman and Company, 1985.
Robinson, J.G.; Janson, C.H. Capuchins,squirrel monkeys, and Atelines: Socioeco-logical convergence with Old World pri-mates. Pp. 69–82 in PRIMATE SOCIETIES.B.B. Smuts; D.L. Cheney; R.M. Seyfarth;R.W. Wrangham; T.T. Struthsaker, eds. Chi-cago, University of Chicago Press, 1987.
Robinson, J.G.; Wright, P.C.; Kinzey, W.G.Monogamous Cebids and their relatives:Intergroup calls and spacing. Pp. 44–53 inPRIMATE SOCIETIES. B.B. Smuts; D.L.Cheney; R.M. Seyfarth; R.W. Wrangham;T.T. Struthsaker, eds. Chicago, Universityof Chicago Press, 1987.
Siegel, S.; Castellan, N.J., Jr. NONPARAMET-RIC STATISTICS FOR THE BEHAVIORAL
SCIENCES. New York, McGraw-Hill Inter-national Edition, 1988.
Stevenson, M.R.; Rylands, A.B. The marmo-sets, genus Callithrix. Pp. 131–222 inECOLOGY AND BEHAVIOUR OF NEO-TROPICAL PRIMATES, VOL. 2. R.A.Mittermeier; A.B. Rylands; A. Coimbra-Filho; G.A.B. Fonseca, eds. Washington,DC, World Wildlife Fund, 1988.
Terborgh, J. FIVE NEW WORLD PRI-MATES: A STUDY IN COMPARATIVEECOLOGY. Princeton, Princeton Univer-sity Press, 1983.
Terborgh, J. Keystone plant resources in thetropical forest. Pp. 330–344 in CONSER-VATION BIOLOGY: THE SCIENCE OFSCARCITY AND DIVERSITY. M.E. Soule,ed. Sunderland, MA, Sinauer, 1986.
Thomas, S.C. Population densities andpatterns of habitat use among anthro-poid primates of the Ituri forest, Zaire.BIOTROPICA 23:68–83, 1991.
van Roosmalen, M.G.M. Habitat preferences,diet, feeding strategy, and social organiza-tion of the black spider monkey (Ateles p.paniscus Linnaeus 1758) in Surinam. ACTAAMAZONICA 15:238 pp., 1985.
Wallace, R.B.; Painter, R.L.E.; Taber, A.B.;Ayres, M. Notes on a distributional riverboundary and southern range extensions fortwo species of Amazonian primates. NEO-TROPICAL PRIMATES 4:149–151, 1996.
White, L.J.T. Biomass of rain forest mam-mals in the Lope Reserve, Gabon. JOUR-NAL OF ANIMAL ECOLOGY 63:499–512,1994.
Whitesides, G.H.; Oates, J.F.; Green, S.M.;Kluberdanz, R.P. Estimating primate den-sities from transects in a West African rainforest: A comparison of techniques. JOUR-NAL OF ANIMAL ECOLOGY 57:345–367,1988.
Wiens, J.A. On competition and variable en-vironments. AMERICAN SCIENTIST 65:590–597, 1977.














!['Pie Memorie' [An Unknown Motet by Noel Bauldeweyn]](https://static.fdokumen.com/doc/165x107/6334bb0b6c27eedec605dd06/pie-memorie-an-unknown-motet-by-noel-bauldeweyn.jpg)





