
Prevalence of Listeria monocytogenes in fresh and fermented Italian sausages and ribotyping of...
7
Prevalence of Listeria monocytogenes in fresh and fermented Italian sausages and ribotyping of contaminating strains Alessandra De Cesare a, ⁎ , Renzo Mioni b , Gerardo Manfreda a a Department of Food Science, Alma Mater Studiorum — University of Bologna, Via S. Giacomo 9, 40126 Bologna, Italy b Istituto Zooprofilattico Sperimentale delle Venezie, V.le dell'Università 10, 35020 Legnaro (PD), Italy Abstract Listeria monocytogenes has been detected in fresh as well as dry and semidry fermented sausages, rendering preparation and consumption of these products as a potential risk to human health. The aims of this study were (1) to evaluate the L. monocytogenes prevalence in 288 fresh and 237 fermented sausages produced in northern Italy; (2) to quantify the average pathogen Most Probable Number (MPN) per g of sausage; (3) to evaluate the sausage strain genetic diversity by automated PvuII ribotyping; and (4) to predict the pathogenicity lineage of these isolates determining their DuPont Identification Library Codes (DUP-IDs) by EcoRI ribotyping. The overall prevalence of L. monocytogenes in the sampled sausages was 28.2%. The percentage of L. monocytogenes positive fresh sausages was significantly higher than that of fermented sausages (i.e. 38.9 vs 15.2%), which had a pathogen load always lower than 10 MPN/g. In contrast, 16.1% of fresh sausages were contaminated by 10 to 100 MPN/g and 20.5% had more than 100 MPN/g. PvuII successfully discriminated sausage isolates with a Simpson's numerical index of discrimination of 0.637. A total of 12 and 9 different PvuII ribogroups were identified among 47 fresh and 24 fermented randomly selected sausage strains, respectively. Six of those ribogroups were shared between strains contaminating both kinds of sausages. According to the evaluation of the strain DUP-IDs, the majority of the isolates investigated in this study were part of the type II L. monocytogenes pathogenicity lineage, but type I lineage strains were identified among fermented sausage isolates. In conclusion, L. monocytogenes prevalence in Italian sausages was estimated to be around 28.2%. However, 84.2% of the samples were contaminated by less than 100 MPN of L. monocytogenes per g and the majority of L. monocytogenes contaminating strains would be classified in the type II pathogenicity lineage, including serotypes 1/2a, 1/2c and 3a. © 2007 Elsevier B.V. All rights reserved. Keywords: Listeria monocytogenes; Sausages; Prevalence; Enumeration; Ribotyping 1. Introduction In Italy, there are many different types of fresh and fer- mented sausages, many of which are only marketed locally or regionally (Comi et al., 2005). The microbiological and physi- cochemical properties of these products influence their contam- ination by L. monocytogenes, which was detected on product surfaces of Italian sausages at an incidence rate of about 9% (Cantoni et al., 1988) and, subsequently, within sausages at a rate ranging between 25 and 32% (Barbuti et al., 1989; Cantoni et al., 1990; Comi et al., 1990; Cantoni, 1991). Fresh sausages are cooked before consumption but they represent a source of cross contamination during meal preparation. On the contrary, finished dry and semidry fermented sausages are ready-to-eat products and their contamination by L. monocytogenes renders the consumption of these sausages as potential food safety risks (Levine et al., 2001; Moore, 2004). Fresh sausage contamination is due to the presence of L. monocytogenes in raw pork meat (Thévenot et al., 2005) as well as in food processing environments, whereby the pathogen is capable of surviving for long periods (Salvat et al., 1995; Lou Available online at www.sciencedirect.com International Journal of Food Microbiology 120 (2007) 124 – 130 www.elsevier.com/locate/ijfoodmicro ⁎ Corresponding author. Tel.: +39 051 209 6766; fax: +39 051 209 6764. E-mail address: [email protected] (A. De Cesare). 0168-1605/$ - see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.ijfoodmicro.2007.06.009
Transcript of Prevalence of Listeria monocytogenes in fresh and fermented Italian sausages and ribotyping of...

Available online at www.sciencedirect.com
biology 120 (2007) 124–130www.elsevier.com/locate/ijfoodmicro
International Journal of Food Micro
Prevalence of Listeria monocytogenes in fresh and fermented Italiansausages and ribotyping of contaminating strains
Alessandra De Cesare a,⁎, Renzo Mioni b, Gerardo Manfreda a
a Department of Food Science, Alma Mater Studiorum — University of Bologna, Via S. Giacomo 9, 40126 Bologna, Italyb Istituto Zooprofilattico Sperimentale delle Venezie, V.le dell'Università 10, 35020 Legnaro (PD), Italy
Abstract
Listeria monocytogenes has been detected in fresh as well as dry and semidry fermented sausages, rendering preparation and consumption ofthese products as a potential risk to human health. The aims of this study were (1) to evaluate the L. monocytogenes prevalence in 288 fresh and237 fermented sausages produced in northern Italy; (2) to quantify the average pathogen Most Probable Number (MPN) per g of sausage; (3) toevaluate the sausage strain genetic diversity by automated PvuII ribotyping; and (4) to predict the pathogenicity lineage of these isolatesdetermining their DuPont Identification Library Codes (DUP-IDs) by EcoRI ribotyping.
The overall prevalence of L. monocytogenes in the sampled sausages was 28.2%. The percentage of L. monocytogenes positive fresh sausageswas significantly higher than that of fermented sausages (i.e. 38.9 vs 15.2%), which had a pathogen load always lower than 10 MPN/g. In contrast,16.1% of fresh sausages were contaminated by 10 to 100 MPN/g and 20.5% had more than 100 MPN/g.
PvuII successfully discriminated sausage isolates with a Simpson's numerical index of discrimination of 0.637. A total of 12 and 9 differentPvuII ribogroups were identified among 47 fresh and 24 fermented randomly selected sausage strains, respectively. Six of those ribogroups wereshared between strains contaminating both kinds of sausages. According to the evaluation of the strain DUP-IDs, the majority of the isolatesinvestigated in this study were part of the type II L. monocytogenes pathogenicity lineage, but type I lineage strains were identified amongfermented sausage isolates.
In conclusion, L. monocytogenes prevalence in Italian sausages was estimated to be around 28.2%. However, 84.2% of the samples werecontaminated by less than 100 MPN of L. monocytogenes per g and the majority of L. monocytogenes contaminating strains would be classified inthe type II pathogenicity lineage, including serotypes 1/2a, 1/2c and 3a.© 2007 Elsevier B.V. All rights reserved.
Keywords: Listeria monocytogenes; Sausages; Prevalence; Enumeration; Ribotyping
1. Introduction
In Italy, there are many different types of fresh and fer-mented sausages, many of which are only marketed locally orregionally (Comi et al., 2005). The microbiological and physi-cochemical properties of these products influence their contam-ination by L. monocytogenes, which was detected on productsurfaces of Italian sausages at an incidence rate of about 9%
⁎ Corresponding author. Tel.: +39 051 209 6766; fax: +39 051 209 6764.E-mail address: [email protected] (A. De Cesare).
0168-1605/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.ijfoodmicro.2007.06.009
(Cantoni et al., 1988) and, subsequently, within sausages at a rateranging between 25 and 32% (Barbuti et al., 1989; Cantoni et al.,1990; Comi et al., 1990; Cantoni, 1991).
Fresh sausages are cooked before consumption but theyrepresent a source of cross contamination during mealpreparation. On the contrary, finished dry and semidryfermented sausages are ready-to-eat products and theircontamination by L. monocytogenes renders the consumptionof these sausages as potential food safety risks (Levine et al.,2001; Moore, 2004).
Fresh sausage contamination is due to the presence ofL. monocytogenes in raw pork meat (Thévenot et al., 2005) aswell as in food processing environments, whereby the pathogenis capable of surviving for long periods (Salvat et al., 1995; Lou

125A. De Cesare et al. / International Journal of Food Microbiology 120 (2007) 124–130
and Yousef, 1999; Tompkin et al., 1999; Tompkin, 2002). Infermented and dried meat products L. monocytogenes is able tosurvive due to its high tolerance to environmental stress factors,including low pH conditions and high salt concentrations(Farber and Peterkin, 1991).
Listeriosis has been classified in Italy as a notificable diseasesince 1993 with 283 reported cases between 1993 to 2000. Inparticular, extensive outbreaks occurred in 1993 and 1997. Thefirst outbreak was attributed to the L. monocytogenes serotype1/2b, electrophoretic type 16, phage type 1967:10:43, in acontaminated rice salad and caused gastroenteritis amongimmunocompetent adults who attended a supper held in a privatehome (Salamina et al., 1996). The second outbreak, involving thestudents and the staff of two primary schools, was attributed to aL. monocytogenes serotype 4b contaminant in corn and tuna fishsalad, which caused a febrile illness with gastroenteritis (Aureliet al., 2000). Moreover, sporadic invasive cases were attributed todifferent foods including a cooked meat pork product named“Testa in cascetta” (Cantoni et al., 1989; Gianfranceschi et al.,2003). Outside Italy other outbreaks related tomeat products havebeen reported; in 1998 a multistate listeriosis outbreak, attributedto the consumption of hot dogs, caused 100 illnesses and 21deaths (CDC, 1999). Another multistate listeriosis outbreakoccurred in 2002 from the consumption of turkey deli products,resulting in 53 illnesses and 11 deaths (CDC, 2002).
A few virulent strains of L. monocytogenes are known tocolonise processing plant environments, contaminate foods andcause listeriosis (Tompkin, 2002). Therefore, the ability todifferentiate isolates of L. monocytogenes is particularly impor-tant for tracking transmission of pathogenic strains within foodprocessing plants and developing more effective interventionstrategies to prevent human illness. The molecular subtypingmethods that can be used to differentiate L. monocytogenes havebeen reviewed by Wiedmann (2002). These include (i) PCR-based methods, e.g. randomly amplified polymorphic DNA(RAPD) (Lawrence et al., 1993; Yoshida et al., 1999) andrepetitive sequence-based PCR (Jersek et al., 1999); (ii) restrictiondigestion-based methods, e.g. ribotyping (Bruce et al., 1995;Swaminathan et al., 1996) and pulsed field gel electrophoresis(PFGE) (Brosch et al., 1996; Graves and Swaminathan, 2001;Gravesen et al., 2000); (iii) amplification–restriction methods,e.g. amplified fragment length polymorphism (AFLP) (Guerraet al., 2002) and PCR-restriction fragment length polymorphism(PCR-RFLP) (Jeffers et al., 2001); (iv) DNA sequence-basedmethods, e.g. multilocus sequence typing (MLST) Maiden et al.,1998; Chan et al., 2001).
Among these methods EcoRI ribotyping on the RiboPrinter®has been shown to be an effective tool for subtyping isolates(Allerberger and Fritschel, 1999; Gendel and Ulaszek, 2000).However, a promising restriction enzyme to discriminate strainsby ribotyping, even within the same clonal groups, is representedby PvuII (De Cesare et al., 2001). Ribotypes appear to be goodmarkers for L. monocytogenes clones that vary in virulence forparticular host species (Jeffers et al., 2001). Moreover, thepotential of using DuPont Identification Library Codes (DUP-IDs) for serotype and lineage classification prediction has beendemonstrated (Nadon et al., 2001). Three serotypes are associated
with many sporadic cases of listeriosis (i.e. 1/2a, 1/2b and 4b)and serotype 4b has been linked to nearly all outbreaks oflisteriosis in Europe (Jay, 1996; Kathariou, 2002).
The objectives of this study were to evaluate the Listeriamonocytogenes prevalence and populations in fresh andfermented sausages and to ribotype contaminating strains toevaluate their level of genetic diversity as well as to predict theirpathogenicity lineages.
2. Materials and methods
2.1. Sampling
A total of 525 samples, represented by 288 fresh and 237fermented sausages, collected from July 2003 troughFebruary 2004, were tested for the presence and populationsof L. monocytogenes. All sausages were sampled in 6 differ-ent regions located in northern Italy. In particular, 439 werecollected in Veneto and 86 in other areas, such as Lombardia,Trentino Alto Adige, Emilia Romagna, Friuli Venezia Giulia andPiemonte. Both fermented as well as fresh sausages were col-lected in large retail chains (N=227), butcher shops (N=259)and other locations (N=39) such as farming holidays.
2.2. Detection of L. monocytogenes in fresh and fermentedsausages
A 25 g fraction, aseptically taken from the central portion ofeach sausage, was added to 225 ml of Listeria Enrichment broth(Oxoid,Milan, Italy) supplementedwith Listeria Primary SelectiveEnrichment Supplement (UVM1) (Oxoid). Samples were homo-genised in a stomacher bag for 60 s at normal speed and incubatedat 32±1 °C for 24 h. A 0.1 ml portion of the first enrichment brothwas inoculated into 9.9 ml of Fraser broth (Oxoid) for a secondenrichment at 32±1 °C for 24 h twice. At the end of eachincubation period the second enrichment broth was streakedonto ALOA agar plates (Microgen, Camberly, UK) and incubatedat 37±1 °C for 48 h. The identification of L. monocytogenes-likecolonies was confirmed by automated EcoRI ribotyping.
2.3. Enumeration of L. monocytogenes in fresh and fermentedsausages
L. monocytogenes was enumerated using the Most ProbableNumber (MPN) procedure when the samples were found to bepositive following qualitative analysis.
Briefly, a 10 g fraction, aseptically taken from the central portionof each sausage, was added to 90 ml of sterile Peptone Water(Oxoid). Samples were homogenised in a stomacher bag for 60 s atnormal speed and serially diluted (i.e. 1:10, 1:100 and 1:1000) insterile Peptone Water. One ml from each dilution was inoculatedinto 9ml of Fraser broth (Oxoid) and incubated at 32±1 °C for 24 htwice. At the end of each incubation period the enrichment brothwas streaked onto ALOA agar plates (Microgen) then incubated at37±1 °C for 48 h. The presence of L. monocytogenes-like colonieswas evaluated and the MPN estimated. MPN results werecategorized by dividing the samples into four categories: samples

Fig. 1. PvuII ribotyping profiles of the L. monocytogenes isolates.
126 A. De Cesare et al. / International Journal of Food Microbiology 120 (2007) 124–130
contaminated by less than 3MPNper g, by 3 to 9MPNper g, by 10to 100 MPN per g, and by more than 100 MPN/g.
2.4. Strain selection and automated ribotyping
A total of 71 randomly selected strains, isolated from 47 freshand 24 fermented sausages were ribotyped according to themanufacturer's instructions using the RiboPrinter® (Qualicon,Inc. Wilmington, DE, USA) (Bruce, 1996). All isolates wereribotyped using PvuII; moreover, 1 up to 16 strains classified inthe different PvuII ribogroups were ribotyped using EcoRI foridentifying their DuPont Identification Library Codes (DUP-IDs).
The ribotype pattern for each isolate was automaticallycompared to patterns stored in the RiboPrinter® database. Theidentification of an isolate was predicted when the corre-sponding EcoRI pattern matched with a similarity ≥0.86 oneof the patterns from DuPont's RiboPrinter® Identification li-brary, labelled as DUP-ID. The characterisation consisted ofcombining EcoRI, and PvuII profiles, within a similarity rangeof ≥0.93 as calculated using the RiboPrinter®'s proprietaryalgorithm, to form a dynamic ribotype or ribogroup that re-flected the genetic relatedness of the isolates (Bruce, 1996).
The profiles collected in this study were labelled with al-phanumeric codes, such as RIBO1::KHB::PvuII 153-216-S-7,which define the strain ribotype or ribogroup label. These codesincorporate the RiboPrinter®'s library in which the strain hasbeen included (i.e. RIBO1::KHB::PvuII), the instrument number(i.e. 153) and the proper strain ribotype or ribogroup (i.e. 216 S7).
2.5. Simpson's index of discrimination (SID)
The suitability of PvuII ribotyping for differentiation ofL. monocytogenes sausage isolates was determined usingthe Simpson's numerical index (SID) as described by Hunterand Gaston (1988). This index represents the probability thattwo randomly chosen isolates would be distinguished by PvuIIautomated ribotyping. As the numerical index approaches themaximum value of one (representing 100% discriminatory
Table 1Distribution of the samples in each MPN contamination category as a functionof sausage type, sampling area and collection point
N. samplescontainingb3 MPN/g(%)
N. samplescontaining3–9 MPN/g(%)
N. samplescontaining10–100 MPN/g(%)
N. samplescontainingN100 MPN/g(%)
Fermentedsausages
24 (66.7) 12 (33.3) 0 0
Fresh sausage 49 (43.8) 22 (19.6) 18 (16.1) 23 (20.5)P⁎ 0.016 0.090 – –
Veneto 61 (48.8) 28 (22.4) 16 (12.8) 20 (16)Other areas 12 (52.2) 6 (26.1) 2 (8.7) 3 (13)P 0.76 0.70 0.58 0.72
Butcher shops 39 (52.7) 13 (17.6) 9 (12.1) 13 (17.6)Large retail chains 31 (47) 19 (28.8) 8 (12.1) 8 (12.1)Other locations 3 (37.5) 2 (25) 1 (12.5) 2 (25)P 0.63 0.29 0.99 0.51
⁎Pb0.05 was considered statistically significant.
ability), the higher the probability that the technique will beable to discriminate between two unrelated isolates.
2.6. Statistical analysis
The datawere analysedwith the Statgraphics® package (ver. 5.1)(StatSoft, Inc.). The comparisons between the L. monocytogenesprevalence in fresh and fermented sausages, as well as betweensamples produced in different areas (Veneto vs other productionareas) or delivered in different retail points (large retail chains vsbutcher shops vs other locations) were analysed by ANOVA usingbi or tri-factorialmodels.Moreover, themeanvalueswere comparedusing the Scheffè test. P≤0.05 was considered statisticallysignificant.
3. Results
3.1. L. monocytogenes prevalence
A total of 148 out of 525 samples were positive forL. monocytogenes, with an overall prevalence rate of 28.2%.The percentage of L. monocytogenes positive fresh sausages wassignificantly higher than that of fermented sausages (i.e. 38.9 vs15.2%). Furthermore, the percentage of positive sausagescollected in the Veneto region was not significantly differentfrom the percentage of positive samples collected in other sam-pling areas (28.5 vs 26.7%). Finally, significant differences werenot detected among the percentage of positive sausages dis-tributed in large retail chains, butcher shops and other locations(29.1, 28.5 and 20.5% respectively).
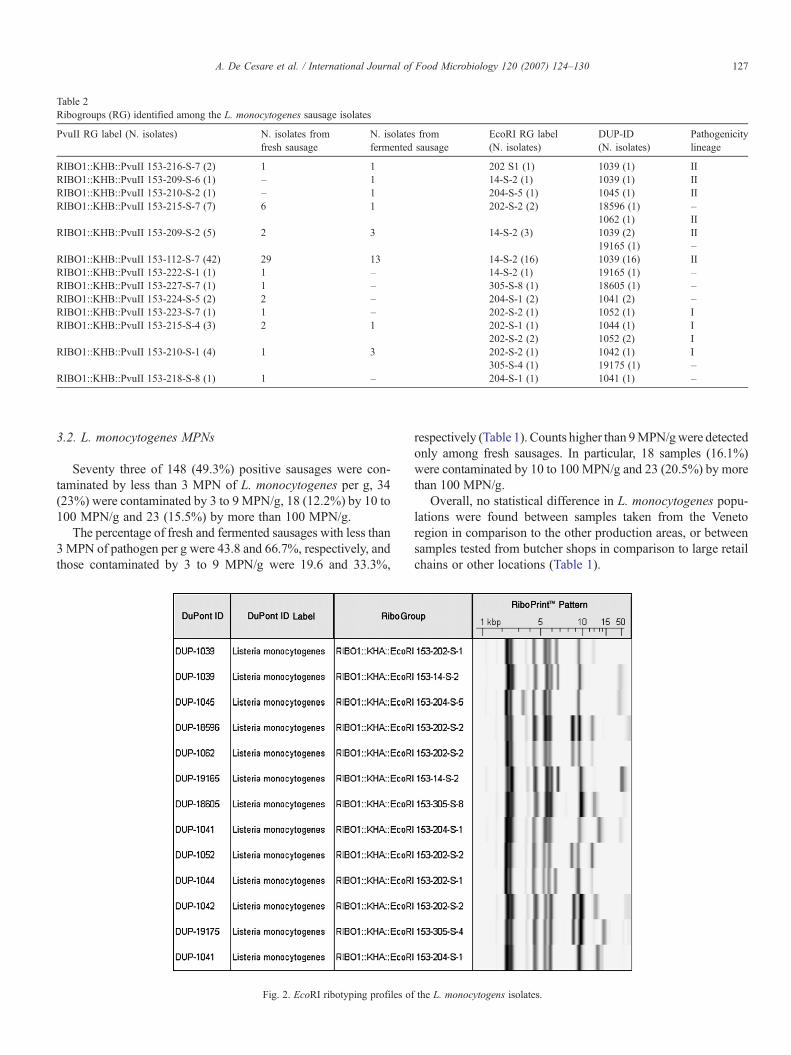
Table 2Ribogroups (RG) identified among the L. monocytogenes sausage isolates
PvuII RG label (N. isolates) N. isolates fromfresh sausage
N. isolates fromfermented sausage
EcoRI RG label(N. isolates)
DUP-ID(N. isolates)
Pathogenicitylineage
RIBO1::KHB::PvuII 153-216-S-7 (2) 1 1 202 S1 (1) 1039 (1) IIRIBO1::KHB::PvuII 153-209-S-6 (1) – 1 14-S-2 (1) 1039 (1) IIRIBO1::KHB::PvuII 153-210-S-2 (1) – 1 204-S-5 (1) 1045 (1) IIRIBO1::KHB::PvuII 153-215-S-7 (7) 6 1 202-S-2 (2) 18596 (1) –
1062 (1) IIRIBO1::KHB::PvuII 153-209-S-2 (5) 2 3 14-S-2 (3) 1039 (2) II
19165 (1) –RIBO1::KHB::PvuII 153-112-S-7 (42) 29 13 14-S-2 (16) 1039 (16) IIRIBO1::KHB::PvuII 153-222-S-1 (1) 1 – 14-S-2 (1) 19165 (1) –RIBO1::KHB::PvuII 153-227-S-7 (1) 1 – 305-S-8 (1) 18605 (1) –RIBO1::KHB::PvuII 153-224-S-5 (2) 2 – 204-S-1 (2) 1041 (2) –RIBO1::KHB::PvuII 153-223-S-7 (1) 1 – 202-S-2 (1) 1052 (1) IRIBO1::KHB::PvuII 153-215-S-4 (3) 2 1 202-S-1 (1) 1044 (1) I
202-S-2 (2) 1052 (2) IRIBO1::KHB::PvuII 153-210-S-1 (4) 1 3 202-S-2 (1) 1042 (1) I
305-S-4 (1) 19175 (1) –RIBO1::KHB::PvuII 153-218-S-8 (1) 1 – 204-S-1 (1) 1041 (1) –
127A. De Cesare et al. / International Journal of Food Microbiology 120 (2007) 124–130
3.2. L. monocytogenes MPNs
Seventy three of 148 (49.3%) positive sausages were con-taminated by less than 3 MPN of L. monocytogenes per g, 34(23%) were contaminated by 3 to 9 MPN/g, 18 (12.2%) by 10 to100 MPN/g and 23 (15.5%) by more than 100 MPN/g.
The percentage of fresh and fermented sausages with less than3 MPN of pathogen per g were 43.8 and 66.7%, respectively, andthose contaminated by 3 to 9 MPN/g were 19.6 and 33.3%,
Fig. 2. EcoRI ribotyping profiles of
respectively (Table 1). Counts higher than 9MPN/gwere detectedonly among fresh sausages. In particular, 18 samples (16.1%)were contaminated by 10 to 100 MPN/g and 23 (20.5%) by morethan 100 MPN/g.
Overall, no statistical difference in L. monocytogenes popu-lations were found between samples taken from the Venetoregion in comparison to the other production areas, or betweensamples tested from butcher shops in comparison to large retailchains or other locations (Table 1).
the L. monocytogens isolates.

128 A. De Cesare et al. / International Journal of Food Microbiology 120 (2007) 124–130
3.3. Automated ribotyping results
Thirteen different PvuII ribotyping profiles, characterised by4 to 8 bands, with a molecular weight ranging between 2.5 and39 kbp, were identified among the 71 isolates tested (Fig. 1).The similarity between these profiles ranged between 11 and89%. The only exception was represented by profilescorresponding to ribogroups 112-S-7 and 224-S-5, with asimilarity of 95%. However, these two profiles were considereddifferent because the second one had an additional band at4.5 kbp (Fig. 1).
Eleven profiles were associated with the 47 strains isolatedfrom fresh sausages and 8 profiles with the 24 strains isolatedfrom fermented sausages (Table 2). Six PvuII ribogroups (i.e.216-S-7, 215-S-7, 209-S-2, 112-S-7, 215-S-4, 210-S-1) wereshared between strains isolated from both fresh and fermentedsausages, whereas 7 ribogroups were associated with strainscollected only from fresh (i.e. 222-S-1, 227-S-7, 224-S-5, 223-S-7, 218-S-8) or fermented (i.e. 209-S-6, 210-S-2) sausages(Table 2). The most frequently identified ribogroup, labelled as112 S7, was identified in 59.1% of the isolates. The suitabilityof PvuII ribotyping to discriminate among the isolates collectedin this study was shown by a SID value of 0.637.
The identification results obtained by EcoRI ribotypingconfirmed the presumptive classification of the isolates withinthe species L. monocytogenes. The isolates tested by EcoRIribotyping were classified within 7 different ribogroups,characterised by 7 to 9 bands, with molecular weights rangingbetween 2.3 and 39 kbp (Fig. 2), and with a similarity rangingbetween 61 and 90%.
Isolates with the same EcoRI ribogroup were classified in upto 5 different PvuII ribogroups. For instance, the 6 isolates withthe EcoRI profile 202-S-2 were characterised by 5 different PvuIIribogroups (i.e. 210-S-1 (N=1); 215-S-4 (N=2); 215-S-7 (N=2);223-S-7 (N=1). In contrast, only two PvuII ribogroups (i.e. 215-S-4 and 210-S-1) included isolates with 2 different EcoRIribogroups (Table 2).
Overall, 11 different DUP-IDs were related to the isolatestested by EcoRI ribotyping. Up to 4 different DUP-IDs wereassociated with isolates with the same EcoRI ribogroup(Table 2). This finding is not unusual because the DUP-IDsand ribogroups are calculated using two different algorithms inthe RiboPrinter®. According to Nadon et al. (2001), six of theDUP-IDs identified in this study are related to one of thepathogenicity lineages known for L. monocytogenes (Table 2).In particular, DUP-IDs 1039, 1045 and 1062 are classified in thetype II pathogenicity lineage, whereas DUP-IDs 1042, 1044 and1052 are classified in the type I lineage.
4. Discussion
The last summary report on trends and sources of zoo-noses, zoonotic agents and antimicrobial resistance in theEuropean Union (www.efsa.eu.int/science/monitoring_zoo-noses/reports) showed that in 2005 there were 1439 casesof listeriosis which represents an incidence of 0.3 cases per100,000 population.
Although L. monocytogenes is the most frequent foodbornepathogen isolated from fermented sausages, as documented bythe U. S. Department of Agriculture, Food Safety andInspection Service (USDA-FSIS) monitoring program, clinicallisteriosis cases have yet to be linked to the consumption offermented sausages contaminated by L. monocytogenes(USDA-FSIS, 2001). However, the presence of L. monocyto-genes in these products is a relevant issue for conductingmicrobiological risk assessment and may result in significanteconomic losses for the food industry.
The prevalence of L. monocytogenes in the fermentedsausages tested in this study (i.e. 15.2%) was slightly higher incomparison to those detected by Thévenot et al. (2005) inFrance (10%) and Cordano and Rocourt (2001) in Chile(10.6%). However, in our study as well as in the French study,the populations of L. monocytogenes found per g of fermentedsausage were lower than 10 CFU. This low pathogen load wasdetected in 63.4% of the fresh sausages tested, with theremaining 16.1% contaminated by 10 to 100 MPN/g and 20.5%of the samples with more than 100 MPN/g. According to otherpublished studies, the lower incidence of L. monocytogenespositive samples detected among fermented sausages incomparison to fresh sausages is attributed to the adverseenvironment created by a pH of about 5.3–6.2, low wateractivity (about 0.80) and high salt concentration used duringproduct fermentation and drying (Bunèiæ et al., 1991).
A SID value of 0.637 was achieved for the sausage isolatesgenotyped using PvuII ribotyping. This discrimination level isappreciable considering the low level of diversity characterisingthe L. monocytogenes populations that contaminate Italian foodproducts (Gianfranceschi et al., 2003). Moreover, it supports theresults published by other authors showing the suitability ofautomated ribotyping as molecular tool to trace sources andtransmission routes of L. monocytogenes (Norton et al., 2001;Thumothe et al., 2004). In comparison to other well-establishedgenotyping methods, such as PFGE, PCR-RFLP and RAPD,automated ribotyping is often considered the DNA subtypingmethod of choice for many large-scale studies as well as forindustrial applications as other techniques are more time-consuming, labour-intensive and requires more personnel with ahigher level of technical expertise (Wiedmann, 2002). Alternativetechniques, such as PCR single strand conformational polymor-phism (SSCP), AFLP and MLST are currently becomingestablished as typing methods for L. monocytogenes but notmany field data has been collected so far (Gasanov et al., 2005).
In this study, PvuII as well as EcoRI ribogroups sharedbetween fresh and fermented sausage strains have beenidentified. The fermented sausage strains with ribogroupsidentical to those of the fresh sausage isolates may be thoseable to survive the adverse conditions (i.e. low pH, low wateractivity and high salt concentration) present during sausageripening; their serotypes, physiology and genetic properties willbe investigated in the near future and compared to those ofisolates unable to survive during the ripening process.
In this study, isolates with the same EcoRI ribogroup wereclassified in up to 5 different PvuII ribogroups. This observationcan be explained considering the higher discriminatory ability

129A. De Cesare et al. / International Journal of Food Microbiology 120 (2007) 124–130
of PvuII in comparison to EcoRI. According to De Cesare et al.(2001), PvuII is able to differentiate L. monocytogenes strainsbelonging even to the same clonal group. Less frequentlyobserved is the classification of strains with the same PvuIIribogroups (i.e. 215-S4 and 210-S-1) but different EcoRIribogroups. These results might indicate that these strains mayhave additional EcoRI cleavage sites in comparison to theaverage.
Rasmussen et al. (1995) demonstrated that L. monocytogenescan be divided into three genetic lineages based on sequencedetermination of the virulence genes hly, iap and flaA.Wiedmann et al. (1997) confirmed the classification ofL. monocytogenes into three genetically distinct lineagesthrough ribotyping and PCR-RFLP of the virulence gene hly.This classification has also been supported by microarray data(Zhang et al., 2003). Notably, all human epidemic isolatesclustered in lineage I (Wiedmann et al., 1997); isolates fromhuman sporadic cases were distributed between lineages Iand II, while animal isolates appeared to encompass a morediverse population which spanned all three lineages (Wiedmannet al., 1997). Ribotypes appear to be good markers forL. monocytogenes clones that vary in virulence for particularhost species (Jeffers et al., 2001). A specific correlation betweensome DUP-IDs and L. monocytogenes lineages has beendemonstrated (Nadon et al., 2001). According to Nadon et al.(2001), L. monocytogenes isolates with DUP-IDs 1039, 1045and 1062 should belong to the type II pathogenicity lineage,whereas those with DUP-IDs 1042, 1044 and 1052 belong tothe type I lineage. Within the second pathogenicity lineage,strains with DUP-ID 1039 should have serotypes 1/2a, 1/2c or3a, whereas those with DUP-IDs 1045 and 1062 shouldencompass serotype 1/2a. Furthermore, within the type Ipathogenicity lineage, strains with DUP-ID 1042 should becomprised of serotypes 1/2b, 3b, 3c, 4b or 4c; those with DUP-ID 1044 should contain serotypes 1/2a or 4b and those withDUP-ID 1052 should contain serotypes 1/2b, 3b or 4b. Acomplete estimation of the isolate distribution in eachpathogenicity lineage cannot be performed because DUP-IDs18596, 19165, 18605, 1041 and 19175 have not been linked toany lineage so far. However, the majority of the sausage strainsseem to be part of the II pathogenicity lineage and wereclassified in the DUP-ID 1039.
In conclusion, the results of this study showed that theprevalence of L. monocytogenes contamination in Italiansausages produced and sold in northern Italy is around 28.2%.Moreover, a higher prevalence of L. monocytogenes positivesamples was found in fresh sausages than among fermentedproducts. The population of L. monocytogenes per g of sausagewas less than 10 MPN in all fermented sausages tested and in63.4% of fresh sausages. Moreover, among fresh sausages,16.1% were contaminated by less than 100 MPN/g and 20.5%by more than 100 MPN/g.
According to the strain DUP-IDs, it can be assumed thatthe majority of the isolates genotyped in this study belongedto the type II pathogenicity lineage. However, some of thestrains classified in the type I lineage were isolated fromfermented sausages and showed ribogroups identified also in
fresh products. These isolates will be further investigated toidentify the physiological and genetic properties enablingthem to survive during the ripening process because theirpresence in ready to eat fermented sausages should be care-fully avoided.
References
Allerberger, F., Fritschel, S.J., 1999. Epidemiological typing of Austrian Listeriamonocytogenes isolates by automated ribotyping. Journal of MicrobiologicalMethods 35, 237–244.
Aureli, P., Fiorucci, G.C., Caroli, D., Marchiaro, G., Novara, O., Leone, L.,Salmaso, S., 2000. An outbreak of febrile gastroenteritis associated withcorn contaminated with Listeria monocytogenes. New England Journal ofMedicine 342, 1236–1241.
Barbuti, S., Ghisi,M., Campanini,M., 1989. Listeria in prodotti carnei: isolamento,incidenza e caratteristiche di sviluppo. Industria Conserve 64, 221–224.
Brosch, R., Brett, M., Catimel, B., Luchansky, J.B., Ojeniyi, B., Rocourt, J.,1996. Genomic fingerprinting of 80 strains from the WHO multicenterinternational typing study of Listeria monocytogenes via pulsed-field gelelectrophoresis (PFGE). International Journal of Food Microbiology 32,343–355.
Bruce, J.L., Hubner, R.J., Cole, E.M., McDowell, C.I., Webster, J.A., 1995. Setsof EcoRI fragments containing ribosomal RNA sequences are conservedamong different strains of Listeria monocytogenes. Proceedings of theNational Academy of Sciences 92, 5229–5233.
Bruce, J., 1996. Automated system rapidly identifies and characterises micro-organisms in foods. Food Technology 50, 77–78.
Bunèiæ, S., Paunovic, L., Radisic, D., 1991. The fate of Listeria monocytogenesin fermented sausages and in vacuum-packaged frankfurters. Journal ofFood Protection 54, 413–417.
Cantoni, C., 1991. Controllo delle listerie negli stabilimenti della carne.Industrie Alimentari 30, 271–273.
Cantoni, C., Balzaretti, C., Valenti, M., 1989. A case of Listeria monocytogeneshuman infection associated with consumption of “Testa in cascetta” (cookedmeat pork product). Archivio Veterinario Italiano 40, 141–142.
Cantoni, C., Valenti, M., Comi, G., 1988. Listeria in formaggi e salumi. IndustrieAlimentari 27, 859–861.
Cantoni, C., Valenti, M., d'Aubert, S., 1990. Listeria spp. in alcuni alimenti eproblematiche relative alla sua presenza. Ingegneria Alimentare 6, 22–23.
Centers for Disease Control and Prenevtion, 1999. Update: multistate outbreakof listeriosis—United States, 1998–1999. Morbidity and Mortality WeeklyReport 47, 1117–1118.
Centers for Disease Control and Prenevtion, 2002. Public heath dispatch:outbreak of listeriosis—Northeastern United States, 2002. Morbidity andMortality Weekly Report 51, 950–951.
Chan,M.,Maiden,M.C.J., Spatt, B.G., 2001.Database-drivenmultilocus sequencetyping (MLST) of bacterial pathogens. Bioinformatics 17, 1077–1083.
Comi, G., Maifreni, M., Cantoni, C., D'Aurelio, F., Valenti, M., 1990. Conta diListeria spp. in alimenti mediante metodo diretto e metodo M.P.N. (mostprobable number). Industrie Alimentari 29, 985–990.
Comi, G., Urso, R., Iacumin, L., Rantsiou, K., Cattaneo, P., Cantoni, C., Cocoli, L.,2005. Characterisation of naturally fermented sausages produced in the NorthEast of Italy. Meat Science 69, 381–392.
Cordano, A.M., Rocourt, J., 2001. Occurrence of Listeria monocytogenes infood in Chile. International Journal of Food Microbiology 70, 175–178.
De Cesare, A., Dambaugh, T.R., Bruce, J., Guerzoni, M.E., Wiedmann, M.,2001. Automated ribotyping using different enzymes to improve discrim-ination of Listeria monocytogenes, with particular focus on serotype 4bstrains. Journal of Clinical Microbiology 39, 3002–3005.
Farber, J.M., Peterkin, P.I., 1991. Listeria monocytogenes, a foodbornepathogen. Microbiology Review 55, 476–511.
Gasanov, U., Hughes, D., Hansbro, P.M., 2005. Methods for the isolation andidentification of Listeria spp. and Listeria monocytogenes: a review. FEMSMicrobiology Reviews 29, 851–875.
Gendel, S.M., Ulaszek, J., 2000. Ribotype analysis of strain distribution inListeria monocytogenes. Journal of Food Protection 63, 179–185.

130 A. De Cesare et al. / International Journal of Food Microbiology 120 (2007) 124–130
Gianfranceschi, M., Gattuso, A., Tartaro, S., Aureli, P., 2003. Incidence ofListeria monocytogenes in food and environmental samples in Italy between1990 and 1999: serotype distribution in food, environmental and clinicalsamples. European Journal of Epidemiology 18, 1001–1006.
Graves, L.M., Swaminathan, B., 2001. PulseNet standardized protocol forsubtyping Listeria monocytogenes by macrorestriction and pulsed-field gelelectrophoresis. International Journal of Food Microbiology 65, 55–62.
Gravesen, A., Jacobsen, T., Moller, P.L., Hansen, F., Larse, A.G., Knochel, S.,2000. Genotyping of Listeria monocytogenes: comparison of RAPD, ITSand PFGE. International Journal of Food Microbiology 57, 43–51.
Guerra, M.M., Bernardo, F., McLauchlin, J., 2002. Amplified fragment lengthpolymorphism (AFLP) analysis of Listeria monocytogenes. Systematic andApplied Microbiology 25, 456–461.
Hunter, P.R., Gaston, M.A., 1988. Numerical index of discriminatory ability oftyping systems: an application of Simpson's index of diversity. Journal ofClinical Microbiology 26, 2465–2466.
Jay, J.M., 1996. Prevalence of Listeria spp. in meat and poultry products. FoodControl 7, 209–214.
Jeffers, G.T., Bruce, J.L.,McDonough, P.L., Scarlett, J., Boor, K.J.,Wiedmann,M.,2001. Comparative genetic characterization of Listeria monocytogenes isolatesfrom human and animal listeriosis cases. Microbiology 147, 1095–1104.
Jersek, B., Gilot, P., Gubina, M., Klun, N., Mehle, J., Tcherneva, E., Rijpens, N.,Herman, L., 1999. Typing of Listeria monocytogenes strains by repetitiveelement sequence-based PCR. Journal of Clinical Microbiology 37,103–109.
Kathariou, S., 2002. Listeria monocytogenes virulence and pathogenicity, a foodprospective. Journal of Food Protection 65, 1811–1829.
Lawrence, L.M., Harvey, J., Gilmour, A., 1993. Development of a randomamplification of polymorphicDNA typingmethod forListeriamonocytogenes.Applied and Environmental Microbiology 59, 3117–3119.
Levine, P., Rose, B., Green, S., Ransom, G., Hill, W., 2001. Pathogen testing ofready-to-eat meat and poultry products collected at federally inspectedestablishments in the United States 1990 to 1999. Journal of Food Protection64, 1188–1193.
Lou, Y., Yousef, A.E., 1999. Characteristics of Listeria monocytogenesimportant to food processors, In: Ryser, E.T., Marth, E.H. (Eds.), Listeria,listeriosis and food safety, 2nd ed. rev. Marcel Decker, Inc., New York,pp. 131–234 (And expanded).
Maiden, M.C.J., Bygraves, J.A., Feil, E., Morelli, G., Russell, J.E., Urwin, R.,Zhang, Q., Zhou, J., Zurth, K., Caugant, D.A., Feavers, I.M., Achtman, M.,Spratt, B.G., 1998. Multilocus sequence typing: a portable approach to theidentification of clones within populations of pathogenic microorganisms.Proceedings of the National Academy of Sciences 95, 3140–3145.
Moore, J.E., 2004. Gastrointestinal outbreaks associated with fermented meats.Meat Science 67, 565–568.
Nadon, C.A., Woodward, D.L., Young, C., Rodgers, F.G., Wiedmann, M., 2001.Correlations between molecular subtyping and serotyping of Listeriamonocytogenes. Journal of Clinical Microbiology 39, 2704–2707.
Norton, D.M., McCamey, M.A., Gall, K.L., Scarlett, J.M., Boor, K.J.,Wiedmann, M., 2001. Molecular studies on the ecology of Listeriamonocytogenes in the smoked fish processing industry. Applied andEnvironmental Microbiology 67, 198–205.
Rasmussen, O.F., Skouboe, P., Dons, L., Rosen, L., Olsen, J.E., 1995. Listeriamonocytogenes exists in at least three evolutionary lines: evidence fromflagellin, invasive associated protein and listeriolysin O gene. Microbiology141, 2053–2061.
Salamina, G., Dalle Donne, E., Niccolini, A., Poda, G., Cesaroni, D., Bucci, M.,Fini, R., Maldini, M., Schuchat, A., Swaminathan, B., Bibb, W., Rocourt, J.,Binkin, N., Salmaso, S., 1996. A foodborne outbreak of gastroenteritisinvolving Listeria monocytogenes. Epidemiology and Infection 117,429–436.
Salvat, G., Toquin, M., Michel, Y., Colin, P., 1995. Control of Listeriamonocytogenes in the delicatessen industries: the lessons of a listeriosisoutbreak in France. International Journal of Food Microbiology 25, 75–81.
Swaminathan, B., Hunter, S.B., Desmarchelier, P.M., Gerner-Smidt, P.,Graves, L.M., Harlander, S., Hubner, R., Jacquet, C., Pedersen, R.,Reineccius, K., Ridley, A., Saunders, N.A., Webster, J.A., 1996. WHO-sponsored international collaborative study to evaluate methods forsubtyping Listeria monocytogenes: restriction fragment length polymorph-isms (RFLP) analysis using ribotyping and Southern hybridization withtwo probes derived from L. monocytogenes chromosome. InternationalJournal of Food Microbiology 32, 263–278.
Thévenot, D., Delignette-muller, M.L., Christieans, S., Vernozy-Rozand, C.,2005. Prevalence of Listeria monocytogenes in 13 dried sausage processingplants and their products. International Journal of Food Microbiology 102,85–94.
Thumothe, J., Nightingale, K.K., Gall, K., Scott, V.N., Wiedmann, M., 2004.Tracking of Listeria monocytogenes in smoked fish processing plants.Journal of Food Protection 67, 328–341.
Tompkin, R.B., 2002. Control of Listeria monocytogenes in the food processingenvironment. Journal of Food Protection 65, 709–725.
Tompkin, R.B., Scott, V.N., Bernard, D.T., Sveum, W.H., Gombas, K.S., 1999.Guidelines to prevent post-processing contamination from Listeriamonocytogenes. Dairy, Food and Environmental Sanitation 19, 551–562.
U.S. Department of Agriculture, Food Safety and Inspection Service, 2001.Performance standards for the production of processed meats and poultryproducts; proposed rule. Federal Register 66, 12590–12636.
Wiedmann, M., 2002. Molecular subtyping methods for Listeria monocytogenes.Journal of AOAC International 85, 524–531.
Wiedmann,M., Bruce, J.K., Keating, C., Johnson,A.,McDonough, P.L., Batt, C.A.,1997. Ribotypes and virulence gene polymorphism suggest three distinctListeria monocytogenes lineages with differences in pathogenic potential.Infection and Immunity 65, 2707–2716.
Yoshida, T., Takeuchi, M., Sato, M., Hirai, K., 1999. Typing of Listeriamonocytogenes by random amplified polymorphic DNA (RAPD) finger-printing. Journal of Veterinary Medical Science 61, 850–860.
Zhang, C., Zhang, M., Ju, J., Nietfeld, J., Wise, J., Terry, P.M., Olson, M.,Kachman, S.D., Wiedmann, M., Samadpour, M., Benson, A.K., 2003.Genome diversification in phylogenetic lineages I and II of Listeriamonocytogenes: identification of segments unique to lineage II populations.Journal of Bacteriology 185, 5573–5584.




















