
Pressure-inactivated yellow fever 17DD virus: Implications for vaccine development
6
Journal of Virological Methods 150 (2008) 57–62 Contents lists available at ScienceDirect Journal of Virological Methods journal homepage: www.elsevier.com/locate/jviromet Pressure-inactivated yellow fever 17DD virus: Implications for vaccine development Luciane P. Gaspar a,∗ , Ygara S. Mendes b , Anna M.Y. Yamamura a , Luiz F.C. Almeida a , Elena Caride a , Rafael B. Gonc ¸ alves b,1 , Jerson L. Silva b , Andr ´ ea C. Oliveira b , Ricardo Galler a , Marcos S. Freire a a Programa de Vacinas Virais, Instituto de Tecnologia em Imunobiol´ ogicos, Fundac ¸˜ ao Oswaldo Cruz, Rio de Janeiro, RJ 21045-900, Brazil b Programa de Biologia Estrutural, Instituto de Bioqu´ ımica M´ edica and Centro Nacional de Ressonˆ ancia Magn´ etica Nuclear de Macromol´ eculas Jiri Jonas, Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ 21941-590, Brazil article info Article history: Received 1 November 2007 Received in revised form 3 March 2008 Accepted 6 March 2008 Keywords: Yellow fever 17DD virus High pressure Virus inactivation abstract The successful Yellow Fever (YF) vaccine consists of the live attenuated 17D-204 or 17DD viruses. Despite its excellent record of efficacy and safety, serious adverse events have been recorded and influenced extensive vaccination in endemic areas. Therefore, alternative strategies should be considered, which may include inactivated whole virus. High hydrostatic pressure has been described as a method for viral inactivation and vaccine development. The present study evaluated whether high hydrostatic pressure would inactivate the YF 17DD virus. YF 17DD virus was grown in Vero cells in roller bottle cultures and subjected to 310 MPa for 3 h at 4 ◦ C. This treatment abolished YF infectivity and eliminated the ability of the virus to cause disease in mice. Pressure-inactivated virus elicited low level of neutralizing antibody titers although exhibited complete protection against an otherwise lethal challenge with 17DD virus in the murine model. The data warrant further development of pressure-inactivated vaccine against YF. © 2008 Elsevier B.V. All rights reserved. 1. Introduction Yellow Fever (YF) virus belongs to the Flavivirus genus of the Flaviviridae family and is an arthropod-borne agent transmitted mainly by mosquitoes of the genera Aedes and Haemagogus. YF is an illness that causes hemorrhagic fever in tropical South America and sub-Saharan Africa, and it is a serious threat to those where it remains endemic. There is no antiviral therapy and vaccination is the major strategy for YF control. A live attenuated human vaccine has been available since the late 1930s based on two live attenuated viruses, the French viscerotropic virus (YF-FVV) and the YF-17D. The latter was isolated after 176 passages in modified chick embryo culture (Lloyd et al., 1936; Theiller and Smith, 1937a; Theiller and Smith, 1937b). Subsequently, two additional substrains of 17D were developed resulting in 17D-204 and 17DD. These two substrains are the basis of the present YF vaccine available in the international market. The existing attenuated YF vaccine has proved to be the safest and the most effective arboviral vaccine ever developed (Pugachev et al., 2005). However, the presence of live virus in this vaccine may ∗ Corresponding author. Tel.: +55 21 3882 9317; fax: +55 21 2260 4727. E-mail address: lgaspar@bio.fiocruz.br (L.P. Gaspar). 1 Present address: Centro de Pesquisa em Ciˆ encia e Tecnologia do Leite, Universi- dade Norte do Paran ´ a, Fazenda Experimental da UNOPAR, 86125-000 Tamarana, PR, Brazil. represent a risk for immunocompromised persons. This hypothesis is based on the occurrence of serious adverse events associated with 17D and 17DD virus replication after vaccination (Galler et al., 2001; Martin et al., 2001; Vasconcelos et al., 2001; Struchiner et al., 2004; Martins et al., 2007). An inactivated vaccine could be used as alternative for a person who will stay in the endemic area for a short period or as a prime-boost vaccination strategy. It is noteworthy that the only other certified flavivirus vaccines such as tick-borne encephalitis virus (TBEV) for human use and Japanese encephalitis virus (JEV) for human and veterinary uses are based on inactivated viruses (Pugachev et al., 2005). The success of these inactivated vaccines suggests that a satisfactory product could also be developed for YF virus. Killed antiviral vaccines have disadvantages such as risk of incomplete inactivation, toxicity of the inactivant agent and change in immunogenic properties of the virus. Hydrostatic pressure has been indicated as an approach for viral inactivation and vaccine development (Masson et al., 2001; Ishimaru et al., 2004; Murchie et al., 2005). As high pressure does not introduce exogenous substances into the vaccine and it is often selective in its action on macromolecular structures, its use results usually in highly immunogenic preparations (Silva et al., 1992; Tian et al., 1999; Ishimaru et al., 2004). In addition, it is practical for large-scale immunization. High hydrostatic pressure can therefore be considered as a possibility for the development of an inactivated YF virus vaccine. 0166-0934/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.jviromet.2008.03.002
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Pressure-inactivated yellow fever 17DD virus: Implications for vaccine development

Journal of Virological Methods 150 (2008) 57–62
Contents lists available at ScienceDirect
Journal of Virological Methods
journa l homepage: www.e lsev ier .com/ locate / jv i romet
Pressure-inactivated yellow fever 17DD virus: Implicationsfor vaccine development
Luciane P. Gaspara,∗, Ygara S. Mendesb, Anna M.Y. Yamamuraa, Luiz F.C. Almeidaa, Elena Caridea,live
waldode Res
er (YFcacy
endemholeevel
7DD vh atin m
compta wa
Rafael B. Goncalvesb,1, Jerson L. Silvab, Andrea C. Oa Programa de Vacinas Virais, Instituto de Tecnologia em Imunobiologicos, Fundacao Osb Programa de Biologia Estrutural, Instituto de Bioquımica Medica and Centro NacionalUniversidade Federal do Rio de Janeiro, Rio de Janeiro, RJ 21941-590, Brazil
a r t i c l e i n f o
Article history:Received 1 November 2007Received in revised form 3 March 2008Accepted 6 March 2008
Keywords:Yellow fever 17DD virusHigh pressureVirus inactivation
a b s t r a c t
The successful Yellow Fevits excellent record of effiextensive vaccination inmay include inactivated winactivation and vaccine dwould inactivate the YF 1subjected to 310 MPa for 3the virus to cause diseasetiters although exhibitedthe murine model. The da
1. Introduction
Yellow Fever (YF) virus belongs to the Flavivirus genus of theFlaviviridae family and is an arthropod-borne agent transmittedmainly by mosquitoes of the genera Aedes and Haemagogus. YF isan illness that causes hemorrhagic fever in tropical South Americaand sub-Saharan Africa, and it is a serious threat to those whereit remains endemic. There is no antiviral therapy and vaccinationis the major strategy for YF control. A live attenuated humanvaccine has been available since the late 1930s based on two liveattenuated viruses, the French viscerotropic virus (YF-FVV) andthe YF-17D. The latter was isolated after 176 passages in modifiedchick embryo culture (Lloyd et al., 1936; Theiller and Smith,1937a; Theiller and Smith, 1937b). Subsequently, two additionalsubstrains of 17D were developed resulting in 17D-204 and 17DD.These two substrains are the basis of the present YF vaccineavailable in the international market.
The existing attenuated YF vaccine has proved to be the safestand the most effective arboviral vaccine ever developed (Pugachevet al., 2005). However, the presence of live virus in this vaccine may
∗ Corresponding author. Tel.: +55 21 3882 9317; fax: +55 21 2260 4727.E-mail address: [email protected] (L.P. Gaspar).
1 Present address: Centro de Pesquisa em Ciencia e Tecnologia do Leite, Universi-dade Norte do Parana, Fazenda Experimental da UNOPAR, 86125-000 Tamarana, PR,Brazil.
0166-0934/$ – see front matter © 2008 Elsevier B.V. All rights reserved.doi:10.1016/j.jviromet.2008.03.002
irab, Ricardo Gallera, Marcos S. Freirea
Cruz, Rio de Janeiro, RJ 21045-900, Brazilsonancia Magnetica Nuclear de Macromoleculas Jiri Jonas,
) vaccine consists of the live attenuated 17D-204 or 17DD viruses. Despiteand safety, serious adverse events have been recorded and influencedic areas. Therefore, alternative strategies should be considered, which
virus. High hydrostatic pressure has been described as a method for viralopment. The present study evaluated whether high hydrostatic pressureirus. YF 17DD virus was grown in Vero cells in roller bottle cultures and4 ◦C. This treatment abolished YF infectivity and eliminated the ability ofice. Pressure-inactivated virus elicited low level of neutralizing antibodylete protection against an otherwise lethal challenge with 17DD virus inrrant further development of pressure-inactivated vaccine against YF.
© 2008 Elsevier B.V. All rights reserved.
represent a risk for immunocompromised persons. This hypothesisis based on the occurrence of serious adverse events associatedwith 17D and 17DD virus replication after vaccination (Galler etal., 2001; Martin et al., 2001; Vasconcelos et al., 2001; Struchineret al., 2004; Martins et al., 2007). An inactivated vaccine could be
used as alternative for a person who will stay in the endemic areafor a short period or as a prime-boost vaccination strategy. It isnoteworthy that the only other certified flavivirus vaccines such astick-borne encephalitis virus (TBEV) for human use and Japaneseencephalitis virus (JEV) for human and veterinary uses are basedon inactivated viruses (Pugachev et al., 2005). The success of theseinactivated vaccines suggests that a satisfactory product could alsobe developed for YF virus.Killed antiviral vaccines have disadvantages such as risk ofincomplete inactivation, toxicity of the inactivant agent andchange in immunogenic properties of the virus. Hydrostaticpressure has been indicated as an approach for viral inactivationand vaccine development (Masson et al., 2001; Ishimaru et al.,2004; Murchie et al., 2005). As high pressure does not introduceexogenous substances into the vaccine and it is often selective inits action on macromolecular structures, its use results usuallyin highly immunogenic preparations (Silva et al., 1992; Tian etal., 1999; Ishimaru et al., 2004). In addition, it is practical forlarge-scale immunization. High hydrostatic pressure can thereforebe considered as a possibility for the development of an inactivatedYF virus vaccine.

rologic
58 L.P. Gaspar et al. / Journal of ViThe main goal of the present study was to evaluate inactivationby hydrostatic pressure and immunogenicity of 17DD virus in SwissWebster mice. The study indicates that pressure-inactivated virusinduced protective immunity in mice, although with lower neu-tralizing antibody titers when compared to titers elicited by liveattenuated virus (virus from Bio-Manguinhos commercial vaccineand virus grown in Vero cell).
2. Material and methods
2.1. Chemicals
All reagents were of analytical grade. Distilled water was filteredand deionized through a Millipore water purification system.
2.2. Cells and virus
Vero (African green monkey kidney) cells (CCL 81) and Aedesalbopictus C6/36 cells were obtained from the American Type Cul-ture Collection, Manassas, VA.
Vero cells were maintained in 199 Medium with Earle’s salts(M199; Gibco, Invitrogen Co., CA, USA), buffered with sodium bicar-bonate and supplemented with 5% fetal bovine serum (Gibco,Invitrogen Co., CA, USA), penicillin (100 U/mL) and streptomycin(100 �g/mL) (Sigma–Aldrich Co., MO, USA). C6/36 cells were culti-vated in L15 medium supplemented with 10% fetal bovine serum,0.02 mM of glutamine (Gibco, Invitrogen Co., CA, USA) and penicillin(100 U/mL) and streptomycin (100 �g/mL).
YF 17DD virus used in this study was originally derivedfrom a vaccine batch (035VFA035P) produced by Bio-Manguinhos(FIOCRUZ, Rio de Janeiro, Brazil). For inactivation studies, 17DDvirus was grown in Vero cells at a Multiplicity of Infection (MOI) of0.02 PFU/cell and cultured on roller bottles at 37 ◦C. Usually a cellculture supernatant was harvested at 7 days post infection (p.i.).Supernatants from infected cultures were clarified of cell debris at1000 × g for 10 min in a Beckman centrifuge using a JA-10 rotor.Protein concentration at virus preparations was determined usingBioRad Microtiter Plate Assay, essentially as described by Bradford(1976). Bovine serum albumin was used as a standard.
2.3. Virus inactivation
The high-pressure reactor has been described (Paladini andWeber, 1981) and was purchased from ISS (Champaign, IL). The sys-
tem was kept at 4 ◦C with the aid of a water circulator bath. Pressurewas increased by steps of 689.46 bar (1 MPa = 10 bar = 0.01450 kpsi).Virus inactivation was performed using different preparations ofclarified 17DD virus. Unless otherwise noted, inactivated prepara-tions of virus (310 MPa for 3 h at 4 ◦C) were administered directlyafter high pressure treatment. After virus inactivation, antigenswere quantified by the Bradford protocol (Bradford, 1976) and anELISA using the purified monoclonal antibody for E glycoproteinfrom viral envelope (ATCC, CRL-1689) modified from Monathand Nystrom (1984a) and Monath et al. (1984b). These methodswere used to compare the infectious virus with the samples afterinactivation.2.4. Assay for detection of residual virus infectivity in cell culture
Inactivated YF virus was assayed for residual live virus by inoc-ulation of 1 mL of virus onto C6/36 and Vero cells. Culture bottles of25 cm2 for both cells were inoculated in triplicate with samples ofpressure-inactivated virus. After 1 h adsorption period, cells werefed with L15 (Sigma–Aldrich Co., MO, USA) and 199 media, respec-tively. After 7 days, the culture supernatants were harvested and
al Methods 150 (2008) 57–62
infectious virus was assayed by plaque titration on Vero cell mono-layers, as described previously (Caufour et al., 2001). The resultsrepresent values from three individual experiments.
2.5. Mice studies
All studies were carried out using a protocol approved bythe Institutional Committee of Animal Care and Experimentation(CEUA – FIOCRUZ: P0152/02).
2.5.1. Detection of residual YF 17DD virus infectivity in miceFor analysis of residual live virus after YF 17DD virus inac-
tivation, 2-day old Swiss Webster mice (Mus musculus) wereinoculated by the intracerebral (i.c.) route with YF 17DD virusfrom Bio-Manguinhos vaccine (lot number 035VFA035P), YF 17DDvirus grown in Vero cell, YF 17DD pressure-inactivated virusand mock (199 medium). Viruses were diluted in complete 199culture medium and inoculum was back-titered immediately afterinoculation procedure. The amount of YF 17DD virus in PlaqueForming Unit (PFU) required to kill 50% of the mice was establishedas described (Caufour et al., 2001). Animals were monitored for 21days and deaths recorded. Moribund animals were euthanized byexposure to CO2.
2.5.2. ImmunizationThe immunogenicity study was undertaken with YF 17DD virus
produced in Vero cells. In the assay, groups of female, 3–7-week oldSwiss Webster mice were immunized by the subcutaneous (s.c.)route with a 22 gauge with doses of 0.1 mL. The schedule con-sisted of three doses with 2-week intervals. YF 17DD virus dilutedin complete 199 culture medium was inoculated with approxi-mately 104 PFU/dose. YF 17DD virus diluted in complete 199 culturemedium with approximately 104 PFU/dose was also inactivated asdescribed in Section 2.3 (310 MPa for 3 h at 4 ◦C) and inoculatedin mice. The negative control group received only a complete 199culture medium (mock group). After the immunization procedure,part of the samples that were not inoculated (YF 17DD virus, YF17DD virus inactivated and mock) were titered immediately. Micewere bled from the retro-orbital plexus using sterile, glass capillarypipettes, before the first dose and 2 weeks after each immunization.
2.5.3. Protection assayFor the protection assay, challenge was carried out 45 days after
immunization assay. 9-week old Swiss Webster mice, immunized
previously, were inoculated by the intracerebral (i.c.) with 30 �L ofa lethal dose of 5.0 log10 PFU of YF 17DD vaccine virus (lot number035VFA035P). Mice were monitored daily for mortality up to 21days. The MLD50 was calculated taking into account the cumulativedeath rates for each dilution that together with the titers in PFUoriginated from back titration to establish the amount of virus inPFU required to kill 50% of the mice as described elsewere (Freireet al., 2005).2.6. Plaque reduction neutralization assay
Antibodies titers were determined by the 50% Plaque ReductionNeutralization Test (PRNT) on Vero cells. PRNT was conducted inserial dilutions starting at 1:10 in 96-well tissue culture plates, asdescribed elsewhere (Stefano et al., 1999). Neutralizing antibodytiters were determined in pooled sera. YF neutralizing antibodytiters were expressed in international units per milliliter (IU/mL),using an “in house” serum preparation for YF 17DD contain-ing 111.5 IU/mL. This serum was standartizated according to aWHO international reference of monkey YF serum containing14,300 mIU/mL (Copenhagen Serum Institute) (Freire et al., 2005).
rological Methods 150 (2008) 57–62 59
0
2
4
6
8
6420
Time of pressurization (hours)
Tit
er lo
g 10
(PF
U/m
L)
(A)
(B)
8
10
L.P. Gaspar et al. / Journal of Vi
Table 1Analysis of residual live virus after pressure inactivationa
Sample Log10 (PFU/mL) Citophatic effectin C6/36 cell
Citophatic effectin Vero cell
Mock (199 medium) NDb − −17DD virusc 5.0 + +17DD virusd 5.0 + +17DD virusd (inactivated) NDb − −
a Inactivated YF virus (310 MPa for 3 h at 4 ◦C) was assayed for residual live virus byinoculation of 1 mL of virus onto 25 cm2 cell culture bottles of a (C6/36) cells and Verocells. Infectivity titers were determined by plaque assay on Vero cell as described inSection 2. Each assay was performed three times to guarantee the reproducibility ofthe results.
b No residual infectivity detected by plaque assay.c Virus from Bio-Manguinhos vaccine.d Virus grown in Vero cell.
2.7. Statistical analysis
Means and standard deviations were calculated for residual livevirus test and immunogenicity of YF virus in mice. The Student ttest was used for comparing means. Differences were considered asstatistically significant if P value was 0.05 or less. Statistical analyseswere done using Statistica 6.0 software (Stata Corporation, CollegeStation, TX. 1999).
3. Results
3.1. Viral inactivation and tests for residual live YF virus in cellculture
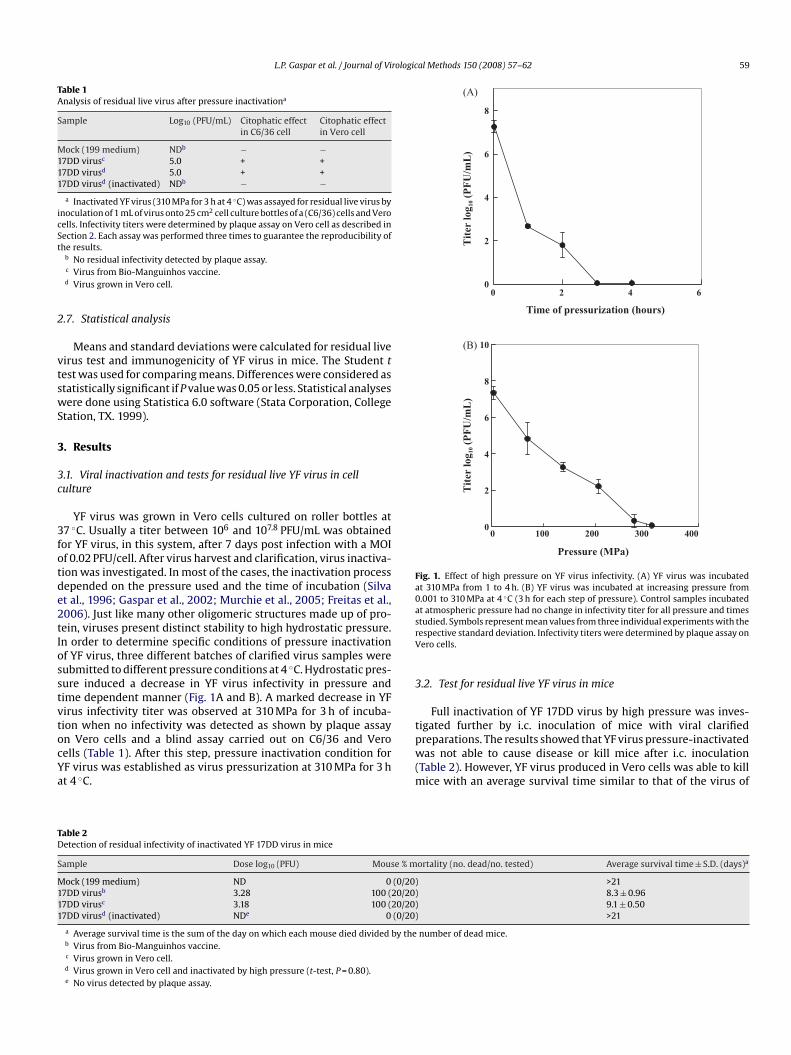
YF virus was grown in Vero cells cultured on roller bottles at37 ◦C. Usually a titer between 106 and 107.8 PFU/mL was obtainedfor YF virus, in this system, after 7 days post infection with a MOIof 0.02 PFU/cell. After virus harvest and clarification, virus inactiva-tion was investigated. In most of the cases, the inactivation processdepended on the pressure used and the time of incubation (Silvaet al., 1996; Gaspar et al., 2002; Murchie et al., 2005; Freitas et al.,2006). Just like many other oligomeric structures made up of pro-tein, viruses present distinct stability to high hydrostatic pressure.In order to determine specific conditions of pressure inactivationof YF virus, three different batches of clarified virus samples weresubmitted to different pressure conditions at 4 ◦C. Hydrostatic pres-sure induced a decrease in YF virus infectivity in pressure andtime dependent manner (Fig. 1A and B). A marked decrease in YFvirus infectivity titer was observed at 310 MPa for 3 h of incuba-tion when no infectivity was detected as shown by plaque assayon Vero cells and a blind assay carried out on C6/36 and Verocells (Table 1). After this step, pressure inactivation condition forYF virus was established as virus pressurization at 310 MPa for 3 hat 4 ◦C.
Table 2Detection of residual infectivity of inactivated YF 17DD virus in mice
Sample Dose log10 (PFU) Mouse % m
Mock (199 medium) ND 0 (0/2017DD virusb 3.28 100 (20/2017DD virusc 3.18 100 (20/2017DD virusd (inactivated) NDe 0 (0/20)
a Average survival time is the sum of the day on which each mouse died divided by theb Virus from Bio-Manguinhos vaccine.c Virus grown in Vero cell.d Virus grown in Vero cell and inactivated by high pressure (t-test, P = 0.80).e No virus detected by plaque assay.
Tit
er lo
g 10
(PF
U/m
L)
2
4
6
04003002001000
Pressure (MPa)
Fig. 1. Effect of high pressure on YF virus infectivity. (A) YF virus was incubatedat 310 MPa from 1 to 4 h. (B) YF virus was incubated at increasing pressure from0.001 to 310 MPa at 4 ◦C (3 h for each step of pressure). Control samples incubatedat atmospheric pressure had no change in infectivity titer for all pressure and timesstudied. Symbols represent mean values from three individual experiments with therespective standard deviation. Infectivity titers were determined by plaque assay onVero cells.
3.2. Test for residual live YF virus in mice
Full inactivation of YF 17DD virus by high pressure was inves-tigated further by i.c. inoculation of mice with viral clarifiedpreparations. The results showed that YF virus pressure-inactivatedwas not able to cause disease or kill mice after i.c. inoculation(Table 2). However, YF virus produced in Vero cells was able to killmice with an average survival time similar to that of the virus of
ortality (no. dead/no. tested) Average survival time ± S.D. (days)a
) >21) 8.3 ± 0.96) 9.1 ± 0.50
>21
number of dead mice.

rological Methods 150 (2008) 57–62
16
18
60 L.P. Gaspar et al. / Journal of Vi
Table 3Immunogenicity of 17DD virus in micea
Sample PRNT50 (mIU/mL) (days post-inoculation)
No. tested First Second (15 days) Third (30 days)
Mock (199 medium) 32 115 157 12917DD virusb 32 1549 4786 19,05517DD virusc 32 1380 3311 12,882Inactivated 17DD virusd,e 32 129 1096 2,129
a Groups of 32 mice received three doses of virus preparation 4.0 log10 PFU for17DD (from Bio-Manguinhos vaccine or grown in Vero cell) and were bled beforeeach inoculation with interval of 15 days (0–15–30 days). A third group received 199medium only (mock group). Neutralization titers were derived on pooled serum foreach group of animals. The pre-immune titer was 129 (mIU/mL).
b Virus from Bio-Manguinhos vaccine.c Virus grown in Vero cell.d Virus grown in Vero cell and inactivated by high pressure (same dilution of live
virus grown in Vero cell).e The values shown are representative of three separate immunizations with dif-
ferent preparations of inactivated YF 17DD virus.
the Bio-Manguinhos commercial vaccine (FIOCRUZ, Rio de Janeiro,Brazil). The difference between the values of average survival timefor both groups failed to reveal any statistically significant differ-ences (p = 0.80) suggesting the viruses replicate to the same extentin the mouse central nervous system.
3.3. Immunogenicity of YF virus in mice
The immunogenicity of 17DD virus inactivated by hydrostaticpressure was investigated by immunization measuring neutralizingantibodies and challenging with live YF 17DD virus by i.c. inocu-lation. For attenuated viruses, the titer of neutralizing antibodiesincreased after second and third inoculations. The commercial vac-cine was more immunogenic than virus from Vero cell-culture(Table 3). In contrast, mice immunized with inactivated virus pro-duced low neutralizing antibody response after the first inoculationwith a slight increase after the second and third booster inocula-tions and were lower than those observed for live YF 17DD viruses(P < 0.05). For all groups neutralizing antibody titers depended onthe number of doses inoculated.
3.4. Protection against inactivated YF virus
To determine if immunization of mice with inactivated YF viruspreparations would confer protection, mice were challenged i.c.
with live 17DD vaccine virus. Negative control mice were mockimmunized with complete 199 culture medium (as described inMaterials and Methods section) and positive one was immunizedwith YF 17DD Bio-Manguinhos vaccine, as shown in Table 3. Fig. 2shows that all mice immunized with inactivated YF virus resistedchallenge. In addition, it was observed that this group exhibited nosigns of illness over an observation period of 21 days post-challenge(data not shown). In contrast, mock immunized mice succumbed tochallenge with an average survival time of 9.3 days. The differencebetween the mortality rates of immunized and mock-immunizedmice was significant (p = 0.03). None of the mock-immunized micesurvived the challenge (Fig. 2). Pre-challenge sera from the 3-dosehigh-pressure vaccinated animals did not show any reactivity to YF17DD E glycoprotein. This protein was detected by Western blot-ting with 1:50 dilution of monoclonal antibody directed to the viralenvelope E glycoprotein (ATCC, CRL-1689) (data not shown).4. Discussion
Although the incidence and geographical distribution of fla-vivirus diseases have increased in recent years, human vaccines are
0
2
4
6
8
10
12
14
181614121086420
Days post-challenge
No.
mic
e su
rviv
ing
Fig. 2. Protection of mice immunized with 17DD virus inactivated by high pressure.The immunized mice were challenged with the lethal dose of 5.0 log10 PFU inocu-lated by i.c. route. The percentage of mice succumbing to infection was compared tothat of mock-infected mice. Circles represent groups immunized with live YF 17DDvaccine control, triangles stands for mice immunized with 17DD inactivated virusand squares represent the mock-immunized group.
available only for YF, Japanese encephalitis, tick-borne encephalitisand Kyasanur forest disease. The commercial YF 17DD vaccine isbased on live attenuated virus and it has proved to be very effec-tive. However, the occurrence of adverse events associated with17D and 17DD viruses warrants the development of an alternativevaccine, mainly, for immunocompromised persons. Another possi-bility, once future development permits, is the use of an inactivatedvaccine as prime-booster vaccination conjugated with a seconddose of the live attenuated virus vaccine. This latter possibilitymight avoid risks of adverse events due to replication of YF virusesand an effective and long lasting immunogenicity, characteristicthat is most difficult to achieve with inactivated viral vaccines alone.
The goal of this study was to evaluate the immunogenicity andprotective efficacy of pressure-inactivated YF 17DD virus in themouse model.
Virus inactivation methods in cell, culture media or specificbuffers includes the use of �-propyl-lactone, binary ethylenimine,formol, irradiation, iodine, ozone, ultraviolet (UV) and photoactivecompounds. Attempts to develop an inactivated YF vaccine havebeen made since 1928. The first vaccine was prepared by phe-
nol or formaldehyde treatment of infected monkey liver and/orspleen (Hindle, 1928). Later in 1936, a study of inactivated YF vac-cine using heat or UV was performed for monkeys (Gordon andHughes, 1936). The problems described with these vaccines werethe presence of residual infectivity in viral preparations or the lossof immunogenicity.The results presented in this study describe the use of highhydrostatic pressure, a relatively new method for YF virus inac-tivation. The kinetics of YF virus was time and pressure-dependentsimilarly to other animal viruses (Jurkiewicz et al., 1995; Tian etal., 2000; Ishimaru et al., 2004; Freitas et al., 2006). Inactivationby hydrostatic pressure was rapid and irreversible, as compared toother enveloped viruses investigated previously (Silva et al., 1992;Gaspar et al., 2002). In addition, the process was shown to yieldsafe preparations demonstrated by infectivity assays, inoculation ofinactivated virus samples onto permissive cell lines (blind assay),test for residual live virus in mice and plaque assay on Vero cells.In fact, after submitting YF 17DD virus to inactivation (310 MPa for3 h at 4 ◦C) we were not able to detect any infectious particle, whatwas important to confirm the absence of residual infectivity. Mostof previous studies on YF inactivated vaccine were not controlled

rologic
L.P. Gaspar et al. / Journal of Vifor residual live virus (Hindle, 1928; Gordon and Hughes, 1936).Such safety profile is important to guarantee the elimination of theserious adverse events caused by infectious virus present in thecommercial attenuated YF vaccine.
Animal models have been used extensively for preclinical test-ing of vaccines to obtain the “proof of concept”. The results of thepresent study showed that immunization of Swiss Webster micewith up to three doses of pressure-inactivated YF 17DD virus wereeffective in eliciting a neutralizing antibody response regarding theprotection against a lethal dose of YF virus. The titers were lowerthan those obtained from mice that were immunized with live virusvaccine at any of first, second or third doses. Considering that lowneutralizing antibody titers correlate with protection (Mason et al.,1973), the immunization with pressure-inactivated 17DD virus didelicit a protective humoral response. It will be of interest to exam-ine the duration of this antibody response. Moreover, it remainsunclear the role of CD8 T cell response after YF immunization asepitopes have been identified after vaccination of mice and manwith live 17D virus (Co et al., 2002; van der Most et al., 2002). Weare presently comparing the CD8 T cell response in mice immu-nized with pressure-inactivated preparations of YF 17DD virus andits live attenuated form. Data supporting the immunogenicity ofpressure-inactivated viruses have been previously published (Silvaet al., 1992; Jurkiewicz et al., 1995; Tian et al., 1999; Tian et al., 2000;Freitas et al., 2006; Grove et al., 2006). Finally, we are currently test-ing the immunogenicity of priming with pressure-inactivated 17DDvirus and boosting with the live virus vaccine.
Taken together, the results show that high hydrostatic pressureinactivates YF virus particles and produces immunogenic particles.Pressure treatment seems to preserve important epitopes on thevirus structure, enabling the pressurized inactivated particles tostimulate a protective neutralizing antibody against YF in mouse. Itis assumed that YF virus overall structure is preserved and pressureshould only affect the flavivirus E protein dimer-based arrange-ments. The loss of YF virus infectivity after pressure inactivationis probably related to this condition as described previously forother viruses (Gaspar et al., 2002; Freitas et al., 2006; Grove et al.,2006). Detailed structural information on inactivated virus enve-lope protein will be determined to establish correlation with thepreservation of important epitopes in virus structure and with thecapacity to stimulate the immune system.
Methodological problems during the development of inacti-vated vaccines are related to antigen production, virus inactivationand lack of antigenic potency. Inactivated vaccines are believed to
be impractical because of the difficulty of obtaining high titers ofvirus in a suitable cell substrate (Putnak et al., 2005). However, thiscan be challenged by new approaches to cultivated cells in serum-free medium in micro-carriers as shown for vaccines against rabiesvirus, Japanese encephalitis virus, enterovirus type 71 and influenzavirus (Merten et al., 1999; Frazatti-Gallina et al., 2004; Wu et al.,2004; Liu et al., 2007). The next steps will be to try to improvevirus yields using micro-carriers as well as to improve the immuneresponse by formulating inactivated 17DD virus preparations withadjuvants and perform dose response studies in mice having asend-point the neutralizing antibody response.Acknowledgements
We thank the Instituto de Tecnologia em Imunobiologicos forthe interest and support to this work. We also thank the technicalassistance of Idevaldo I. Ferreira and Joel Majerowicz for the use ofanimal facilities. This work was supported in part by the Programade Desenvolvimento Tecnologico em Insumos para Saude Publica(PDTSP) from FIOCRUZ, Conselho Nacional de DesenvolvimentoCientıfico e Tecnologico (CNPq), Fundacao Carlos Chagas Filho de
al Methods 150 (2008) 57–62 61
Amparo a Pesquisa no Estado do Rio de Janeiro (FAPERJ) and Insti-tuto Milenio de Biologia Estrutural em Biomedicina e Biotecnologia(IMBEBB) of Brazil.
References
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein-dye binding. Anal.Biochem. 72, 248–254.
Caufour, P.S., Motta, M.C., Yamamura, A.M., Vasquez, S., Ferreira, I.I., Jabour, A.V.,Bonaldo, M.C., Freire, M.S., Galler, R., 2001. Construction, characterization andimmunogenicity of recombinant yellow fever 17 D-dengue type 2 viruses. VirusRes. 79 (1–2), 1–14.
Co, M.D., Terajima, M., Cruz, J., Ennis, F.A., Rothman, A.L., 2002. Human cytotoxic Tlymphocyte responses to live attenuated 17D yellow fever vaccine: identificationof HLA-B35-restricted CTL epitopes on nonstructural proteins NS1, NS2b, NS3,and the structural protein E. Virology 293 (4), 511–517.
Frazatti-Gallina, N.M., Mourao-Fuches, R.M., Paoli, R.L., Silva, M.L., Miyaki, C., Valen-tini, E.J., Raw, I., Higashi, H.G., 2004. Vero-cell rabies vaccine produced usingserum-free medium. Vaccine 23 (4), 511–517.
Freire, M.S., Mann, G.F., Marchevsky, R.S., Yamamura, A.M., Almeida, L.F., Jabor, A.V.,Malachias, J.M., Coutinho, E.S., Galler, R., 2005. Production of yellow fever 17DDvaccine virus in primary culture of chicken embryo fibroblast: yields, thermoand genetic stability, attenuation and immunogenicity. Vaccine 23 (19), 2501–2512.
Freitas, M.S., Da Poian, A.T., Barth, M.O., Rebello, M.A., Silva, J.L., Gaspar, L.P., 2006. Thefusogenic state of Mayaro virus induced by low pH and by hydrostatic pressure.Cell Biochem. Biophys. 44 (3), 205–211.
Galler, R., Pugachev, K.V., Santos, C.L., Ocran, S.W., Jabor, A.V., Rodrigues, S.G.,Marchevsky, R.S., Freire, M.S., Almeida, L.F., Cruz, A.C., Yamamura, A.M., Rocco,I.M., da Rosa, E.S., Sousa, L.T., Vasconcelos, P.F., Guirakhoo, F., Monath, T.P., 2001.Phenotypic and molecular analyses of yellow fever 17DD vaccine viruses asso-ciated with serious adverse events in Brazil. Virology 290 (2), 309–319.
Gaspar, L.P., Silva, A.C.B., Gomes, A.M.O., Freitas, M.S., Ano Bom, A.P.D., Schwarcz,W.D., Mestecky, J., Novak, M.J., Foguel, D., Silva, J.L., 2002. Hydrostatic pressureinduces the fusion-active state of enveloped viruses. J. Biol. Chem. 277 (10),8433–8439.
Gordon, J.E., Hughes, T.P., 1936. A study of inactivated yellow fever virus as an immu-nizing agent. J. Immunol. 30, 221.
Grove, S.F., Lee, A., Lewis, T., Stewart, C.M., Chen, H., Hoover, D.G., 2006. Inactivationof foodborne viruses of significance by high pressure and other processes. J. FoodProt. 69 (4), 957–968.
Hindle, E., 1928. A yellow fever vaccine. Br. Med. J. 1, 976.Ishimaru, D., Sa-Carvalho, D., Silva, J.L., 2004. Pressure-inactivated FMDV: a potential
vaccine. Vaccine 22 (17–18), 2334–2339.Jurkiewicz, E., Villas-Boas, M., Silva, J.L., Weber, G., Hunsmann, G., Clegg, R.M., 1995.
Inactivation of simian immunodeficiency virus by hydrostatic pressure. Proc.Natl. Acad. Sci. U.S.A. 92 (15), 6935–6937.
Lloyd, W., Theiler, M., Ricci, N.I., 1936. Modification of virulence of yellow fever virusby cultivation in tissues in vitro. R. Soc. Trop. Med. Hyg. 22, 481–529.
Liu, C.C., Lian, W.C., Butler, M., Wu, Sc., 2007. High immunogenic enterovirus 71 strainand its production using serum-free microcarrier Vero. Vaccine 25 (1), 19–24.
Martin, M., Tsai, T.F., Cropp, B., Chang, G.J., Holmes, D.A., Tseng, J., Shieh, W., Zaki,S.R., Al-Sanouri, I., Cutrona, A.F., Ray, G., Weld, L.H., Cetron, M.S., 2001. Fever andmultisystem organ failure associated with 17D-204 yellow fever vaccination: a
report of four cases. Lancet 358 (9276), 98–104.Martins, R.M., Galler, R., Freire, M.S., Camacho, L.A.B., Maia, M.L.S., Homma, A., 2007.Yellow fever vaccination: some thoughts on how much is enough. Vaccine 23(2005), 3908–3914.
Mason, R.A., Tauraso, N.M., Spertzel, R.O., Ginn, R.K., 1973. Yellow fever vaccine:direct challenge of monkeys given graded doses of 17D vaccine. Appl. Microbiol.4, 539–544.
Masson, P., Tonello, C., Balny, C., 2001. High pressure biotechnology in medicine andpharmaceutical science. J. Biomed. Biotech. 1 (2), 85–88.
Merten, O.W., Manuguerra, J.C., Hannoun, C., Vander Werf, S., 1999. Production ofinfluenza virus in serum-free mammalian cell cultures. Dev. Biol. Stand. 98,23–37.
Monath, T.P., Nystrom, R.R., 1984a. Detection of yellow fever virus in serum byenzyme immunoassay. Am. J. Trop. Med. Hyg. 33 (1), 151–157.
Monath, T.P., Schlesinger, J.J., Brandiss, M.W., Cropp, C.B., Prange, W.C., 1984b. Yel-low fever monoclonal antibodies: type-specific and cross-reactive determinantsidentified by immunofluorescence. Am. J. Trop. Med. Hyg. 33 (4), 695–698.
Murchie, L.W., Cruz-Romero, M., Kerry, J.P., Linton, M., Patterson, M.F., Smiddy, M.,Kelly, A.L., 2005. High pressure processing of shellfish: a review of microbio-logical and other quality aspects. Innovative Food Sci. Emerg. Technol. 6 (3),257–270.
Paladini, A.A., Weber, G., 1981. Pressure-induced reversible dissociation of enolase.Biochemistry 20 (9), 2587–2593.
Pugachev, K.V., Guirakhoo, F., Monath, T.P., 2005. New developments in flavivirusvaccines with special attention to yellow fever. Curr. Opin. Infect. Dis. 18 (5),387–394.
Putnak, R.J., Coller, B.A., Voss, G., Vaughn, D.W., Clements, D., Peters, I., Bignami, G.,Houng, H.S., Chen, R.C., Barvir, D.A., Seriwatana, J., Cayphas, S., Garcon, N., Ghey-

62 L.P. Gaspar et al. / Journal of Virologic
sen, D., Kanesa-Thasan, N., McDonell, M., Humphreys, T., Eckels, K.H., Prieels,J.P., Innis, B.L., 2005. An evaluation of dengue type-2 inactivated, recombinantsubunit, and live-attenuated vaccine candidates in the rhesus macaque model.Vaccine 23 (35), 4442–4452.
Silva, J.L., Luan, P., Glaser, M., Voss, E.W., Weber, G., 1992. Effects of hydro-static pressure on a membrane-enveloped virus: high immunogenicity of thepressure-inactivated virus. J. Virol. 66 (4), 2111–2117.
Silva, J.L., Foguel, D., Da Poian, A.T., Prevelige, P.E., 1996. The use of hydrostatic pres-sure as a tool to study viruses and other macromolecular assemblages. Curr.Opin. Struct. Biol. 6 (2), 166–175.
Stefano, I., Sato, H.K., Pannuti, C.S., Omoto, T.M., Mann, G., Freire, M.S., Yamamura,A.M., Vasconcelos, P.F., Oselka, G.W., Weckx, L.W., Salgado, M.F., Noale, L.F., Souza,V.A., 1999. Recent immunization against measles does not interfere with thesero-response to yellow fever vaccine. Vaccine 17 (9–10), 1042–1046.
Struchiner, C.J., Luz, P.M., Dourado, I., Sato, H.K., Aguiar, S.G., Ribeiro, J.G.L., Soares,R.C.R., Codeco, C.T., 2004. Risk of fatal adverse events associated with 17DDyellow fever vaccine. Epidemiol. Infect. 132 (5), 939–946.
Theiller, M., Smith, H.H., 1937a. The effect of prolonged cultivation in vitro upon thepathogenicity of yellow fever virus. J. Exp. Med. 65, 767–786.
al Methods 150 (2008) 57–62
Theiller, M., Smith, H.H., 1937b. Use of yellow fever virus modified by in vitro culti-vation for human immunization. J. Exp. Med. 65, 787–800.
Tian, S.M., Qian, J.F., Shao, G.Q., Ruan, K.C., 1999. High immunogenicity of thepressure-inactivated virus. Acta Bioch. Biophys. Sin. 31 (3), 334–336.
Tian, S.M., Ruan, K.C., Qian, J.F., Shao, G.Q., Balny, C., 2000. Effects of hydrostaticpressure on the structure and biological activity of infectious bursal diseasevirus. Eur. J. Biochem. 267 (14), 4486–4494.
van der Most, R.G., Harrington, L.E., Giuggio, V., Mahar, P.L., Ahmed, R., 2002. Yellowfever virus 17D envelope and NS3 proteins are major targets of the antiviral Tcell response in mice. Virology 296 (1), 117–124.
Vasconcelos, P.F., Luna, E.J., Galler, R., Silva, L.J., Coimbra, T.L., Barros, V.L., Monath, T.P.,Rodrigues, S.G., Laval, C., Costa, Z.G., Vilela, M.F., Santos, C.L., Papaiordanou, P.M.,Alves, V.A., Andrade, L.D., Sato, H.K., Rosa, E.S., Froguas, G.B., Lacava, E., Almeida,L.M., Cruz, A.C., Rocco, I.M., Santos, R.T., Oliva, O.F., 2001. Brazilian Yellow FeverVaccine Evaluation Group. Serious adverse events associated with yellow fever17DD vaccine in Brazil: a report of two cases. Lancet 358 (9276), 91–97.
Wu, S.C., Liu, C.C., Lian, W.C., 2004. Optimization of microcarrier cell culture pro-cess for the inactivated enterovirus type 71 vaccine development. Vaccine 22(29–30), 3858–3864.




















