
Preproinsulin specific CD8+ T cells in subjects with latent autoimmune diabetes show lower frequency...
14
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Preproinsulin specific CD8+ T cells in subjects with latent autoimmune diabetes show lower frequency...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
Preproinsulin specific CD8+ T cells in subjectswith latent autoimmune diabetes show lowerfrequency and different pathophysiologicalcharacteristics than those withtype 1 diabetesNaresh Sachdevaa,⁎, Mahinder Paul a,1, Darshan Badal a, Rajendra Kumarb,Neenu Jacoba, Devi Dayal c, Anil Bhansali a,Sunil Kumar Arorab, Sanjay Kumar Bhadadaa
a Department of Endocrinology, Post Graduate Institute of Medical Education and Research (PGIMER), Chandigarh, Indiab Department of Immunopathology, Post Graduate Institute of Medical Education and Research (PGIMER), Chandigarh, Indiac Department of Pediatrics, Post Graduate Institute of Medical Education and Research (PGIMER), Chandigarh, India
Received 16 July 2014; accepted with revision 10 January 2015
KEYWORDSType 1 diabetesLatent autoimmunediabetes in adultsCD8+ T cellsDextramersPreproinsulin
Abstract Latent autoimmune diabetes in adults (LADA) resembles type 1 diabetes (T1D) in diseasepresentation except that its onset is slow.We compared pathophysiological characteristics of CD8+ Tcells recognizing preproinsulin (PPI) derived epitopes in both disease groups using MHC-I dextramers(DMRs) in peripheral blood and after in-vitro stimulation with PPI. Subjects with T1D harbored higherfrequency of DMR+ CD8+ T cells with relatively higher frequency of effector T cell subsets. Followingstimulation with PPI, an increase in DMR+ CD8+ T cells, particularly the central-memory subset wasobserved in T1D group, whereas no significant change in DMR+ CD8+ T cell subsets was observedin LADA group. Intracellular expression of Granzyme-B and Perforin in DMR+ CD8+ T cells was
Abbreviations: APC, Allophycocyanin; AO-T1D, Adult onset type 1 diabetes; BIMAS, Bioinformatics and Molecular Analysis Section; CD,Cluster of differentiation; DMR, Dextramer; ELISpot, Enzyme-linked immunospot; FITC, Fluorescein isothiocyanate; FMO, Fluorescence minusone; GAD65, Glutamic acid decarboxylase; GZM, Granzyme; HbA1C, Glycated hemoglobin; HC, Healthy controls; HIV, Humanimmunodeficiency virus; HLA, Human leukocyte antigen; IA-2, Islet antigen 2; ICA, Islet cell antibody; IEDB, Immune Epitope Database;LADA, Latent autoimmune diabetes in adults; MFI, Mean fluorescence intensity; MHC, Major histocompatibility complex; NK, Natural killer;NOD, Non obese diabetic; PBMCs, Peripheral blood mononuclear cells; PE, Phycoerythrin; PerCP, Peridinin-chlorophyll; PECy7,Phycoerythrin-Cyanine7; PFN, Perforin; PPI, Preproinsulin; PHA, Phytohaemagglutinin; SEM, Standard error of mean; SSP, Sequence-specificprimer; T1D, Type 1 diabetes; TCR, T cell receptor; TCM, Central memory T cells; TEFF, Effector T cells; TEM, Effector memory T cells; TN, Naive Tcells; Treg, Regulatory T cells; TMR, Tetramer; 7-AAD, 7-Aminoactinomycin D.⁎ Corresponding author at: Department of Endocrinology, Post Graduate Institute of Medical Education and Research (PGIMER), Chandigarh
160012, India. Fax: +91 172 2744401.E-mail address: [email protected] (N. Sachdeva).
1 Co-first author.
http://dx.doi.org/10.1016/j.clim.2015.01.0051521-6616/© 2015 Elsevier Inc. All rights reserved.
ava i l ab l e a t www.sc i enced i r ec t . com
C l i n i ca l Immuno logy
www.e l sev i e r . com / l oca te / yc l im
Clinical Immunology (2015) 157, 78–90

Author's personal copy
comparable in both the groups. In conclusion, lower frequency and inferior proliferative potential onaccount of a relatively restrained central-memory subset of PPI specific CD8+ T cells are associatedwith slow rate of disease progression in LADA.© 2015 Elsevier Inc. All rights reserved.
1. Introduction
Type 1 diabetes (T1D) is predominantly associated withactivation, differentiation and expansion of autoreactive CD8+T cells that recognize and destroy pancreatic beta-cells [1].Amongst the beta-cell associated antigens, preproinsulin (PPI)has been recognized as a key autoantigen in initiation andpathogenesis of T1D in humans as well as non-obese diabetic(NOD) mouse models [2–8]. Most of the PPI derived epitopesare reported from subjects with HLA-A*02 genotype and havebeen located in the leader sequence, B-chain and adjacentC-peptide cluster [9–11]. Autoreactive CD8+ T cells recognizesuch epitopes expressed on the surface of beta-cells inassociation with MHC-I molecules and destroy them mainlyby degranulation, releasing perforin (PFN) containing cyto-toxic granules which facilitate entry of granzymes (GZM) withserine protease activity [4].
Following antigenic recognition, autoreactive CD8+ T cellsundergo clonal expansion and differentiate into differentsubsets, including central-memory (TCM), effector-memory(TEM) and effector (TEFF) cells characterized by changesin extracellular receptors such as, the isoform of commonlymphocyte antigen, CD45RA, chemokine receptor CCR7(CD197) and intracellular expression of effector molecules[12,13]. The pattern of differentiation is mainly determined bythe strength and duration of antigenic signal and requires aid ofCD4+ T cells [14–17]. Hence, besides frequency, determinationof phenotype of autoreactive CD8+ T cells also becomesimportant in monitoring disease progression in T1D subjects orindividuals at risk and assessing efficacy of therapeutic ortransplant approaches [18,19]. However, detection of auto-reactive CD8+ T cells in peripheral blood has been challenging,particularly in populations with diverse HLA alleles. To assessautoreactivity, earlier studies used Enzyme-linked Immunospot(ELISpot) or T-cell proliferative assays which mainly reflect onCD4+ T cell responses [9]. Later, peptide MHC-I multimers,particularly tetramers (TMRs) have been used to determine thefrequency of CD8+ T cells recognizing different beta-cellepitopes with variable success rates in individuals with differentdurations of disease [10,11,20,21]. Gradual validation effortscoupled with designing of improved peptide MHC-I multimershave now overcome many technical challenges in accurateassessment of epitope specific CD8+ T cells in the background ofnumerous CD8+ T cells [16,22,23].
Beta-cell destruction is quite variable, rapid in someindividuals, mainly children, presenting with T1D, and slow inthose with latent autoimmune diabetes in adults (LADA). LADAresembles T1D and shares common autoimmune featuresand physiological characteristics except that its onset is slowand seen in adults usually misdiagnosed as type 2 diabetes,generally accounting for 10% of total diabetes cases [24,25].The underlyingmechanisms that delay the onset of diabetes in
LADA are not completely understood. Furthermore, thereis very little information on the immunophenotype of auto-reactive CD8+ T cells in autoimmune diabetes in subjects withdifferent HLA alleles, particularly their differentiation patternsfollowing exposure to beta-cell epitopes. Therefore, in thepresent study we compared and characterized PPI specificperipheral autoreactive CD8+ T cells in subjects with T1D andLADA using MHC-I dextramers (DMRs) loaded with PPI derivedpeptide epitopes. Further, we also compared the proliferativepotential of DMR+ CD8+ T cells following in-vitro stimulation ofperipheral blood mononuclear cells (PBMCs) with PPI andassessed their immunophenotypic characteristics.
2. Research design and methods
2.1. Subjects
Peripheral blood samples were obtained from 75 subjectsattending departments of Endocrinology and Pediatrics at thePost Graduate Institute of Medical Education and Research,Chandigarh, India. The study was approved by the institutionalethics committee. After obtaining informed consent subjectswere screened for autoantibodies to GAD65, insulin, IA-2 andICA and divided into 3 groups. Group 1 comprised of 39 subjectswith recently diagnosed T1D (within 1 month of diagnosis),Group 2 included 18 LADA subjects (aged N 25 years, positivefor at least one of the autoantibodies and insulin independentfor at least 6 months after diagnosis of diabetes and duringrecruitment). Group 3 comprised of 18 age and sex matchednon-diabetic healthy controls (HC). Later, additional 15subjects with adult onset T1D (AO-T1D) (aged N 25 years,antibody positive and insulin dependent) were also recruitedand compared with LADA subjects.
2.2. Plasma C-peptide, HbA1C determination andHLA-I typing
At recruitment, fasting plasma C-peptide in all subjectswas determined by Electrochemiluminescence Immunoassay(Roche Diagnostics, Germany). Glycosylated hemoglobin(HbA1C) was estimated by cation-exchange chromatography(Bio-Rad, USA). Genomic DNA was extracted from the wholeblood (Real Biotech Corporation, Taiwan) before HLA-I typingof the subjects by PCR-SSP in 96-well typing plates (Inno-train,Germany). PCR products were resolved by electrophoresis on a2% agarose gel, followed by determination of HLA alleles.
2.3. Dextramer synthesis and validation
Eight different Allophycocyanin (APC) labeled MHC-I DMRsloaded with 9-mer PPI derived epitopes, having highest
79Preproinsulin specific CD8+ T cells in subjects with latent autoimmune diabetes

Author's personal copy
cumulative affinity for respective MHC-I molecules as deter-mined by MHC-I peptide binding prediction algorithms, BIMAS(Bioinformatics and Molecular Analysis Section, Centre forInformation Technology, NIH, USA), SYFPEITHI (BiomedicalInformatics, Heidelberg, Germany) and T cell Epitope Predic-tion Tool of Immune Epitope Database (IEDB) (La Jolla, CA)werecustom synthesized (Immudex, Denmark) (Table 2). IFN-γELISpot was performed to validate the DMRs as previouslydescribed [10,26].
2.4. Immunophenotyping and analysis of DMR+CD8+ T cells
After determining HLA-I type, fresh sample of peripheralblood was obtained from 39 T1D, 14 LADA and 15 HC subjectsfor immunophenotyping. PBMCs were isolated by densitygradient centrifugation with ficoll (Sigma, USA). After checkingcell viability by 7-aminoactinomycin D (7-AAD) exclusion,PPI specific CD8+ T cells were stained by incubating 2 × 106
PBMCs with 10 μl of respective DMRs at room temperaturein dark for 10 min. Anti-CD3 Peridinin-chlorophyll (PerCP),anti-CD8 Phycoerythrin-Cyanine7 (PECy7), anti-CD45RAAPC-H7,anti-CD197 Phycoerythrin (PE) and cocktail of Alexa-Fluor 700labeled anti-CD4, anti-CD14, anti-CD16 and anti-CD19 werethen added for 15 min followed by washing with FACSTM
buffer (BD Biosciences, USA). Cells were then permeabilizedwith CytoPerm-CytoFixTM solution and incubated with anti-Granzyme-B violet450 (V450) and Perforin-Fluoresceinisothiocyanate (FITC) for 30 min at 4 °C. After washing,cells were acquired on a flowcytometer (FACS Aria II) andanalyzed using FACS DivaTM software (6.01) (BD Biosciences,USA).
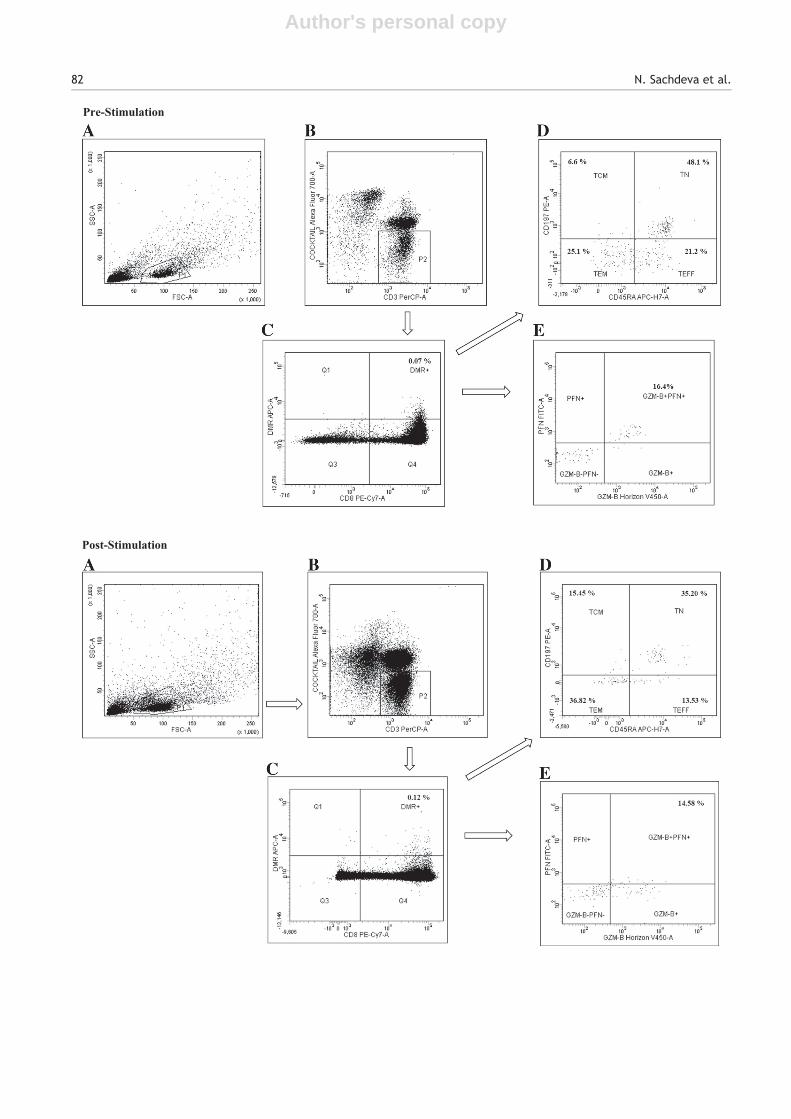
At least one million total events were acquired and aminimum of 500,000 T lymphocytes were analyzed. Gating wasfirst performed on lymphocytes followed by gating of CD3+CD4− CD14− CD16− CD19− cells to exclude CD4+ T cells,monocytes, NK cells and B cells. CD8+ T cells recognizing theDMRs were then gated and analyzed for markers of differ-entiation stages including, naïve (TN) (CD45RA+ CD197+),central-memory (TCM) (CD45RA− CD197+), effector-memory(TEM) (CD45RA− CD197−), effector (TEFF) (CD45RA+ CD197−)along with intracellular expression of GZM-B and PFN (Fig. 2).Relevant isotype controls and fluorescence minus one (FMO)tubes were used to set gates. DMRs loaded with Flu specific
peptides were used as positive control, while DMRs carryingHIV-1 specific peptides were used as negative controls(Supplemental Table 1).
2.5. In-vitro stimulation with PPI
A total of 106 PBMCs/ml were plated/well in triplicates inflat bottom 24-well tissue culture plates (BD-Falcon, USA) inRPMI-1640 medium supplemented with 0.1% penicillin, strepto-mycin, 10% fetal calf serum and stimulated with optimizedconcentration of PPI (10 μg/ml) (Link Biotech, India) or 5 μg/mlPHA (positive control) or without any stimulant (negativecontrol) for 72 h at 37 °C in 5% CO2. Following incubation,cells were harvested, stained and reanalyzed for DMR+ CD8+ Tcell subsets as described above.
2.6. Statistical analysis
Frequencies of DMR+ CD8+ T cells between different groupswere compared using unpaired t-test. Paired t-test was usedto compare the frequency and subsets of DMR+ CD8+ T cellsbefore and after stimulation with PPI. Mann–Whitney U testwas used wherever data distribution was not normal. Theanalyses were two tailed and p b 0.05 was considered assignificant.
3. Results
3.1. Clinical characteristics and HLA heterogeneityof subjects
Mean (±SEM) age at presentation of diabetes was 8.6 ±0.6 years in T1D subjects, significantly lower than the LADAsubjects (29.8 ± 1.7 years) (p b 0.001). Mean (±SEM) fastingplasma C-peptide levels were significantly higher in LADA(1.65 ± 0.35 ng/ml) than T1D subjects (0.47 ± 0.08 ng/ml)(p = 0.0001), while HbA1C was higher in T1D (12.29 ± 0.52%)(110 ± 3.3 mmol/mol) compared to LADA subjects (8.67 ±0.53%) (70 ± 3.4 mmol/mol) (p = 0.001). The T1D subjects(BMI, 16.34 ± 0.69) had a lean phenotype compared to LADAsubjects (20.27 ± 0.74) (p = 0.04) (Table 1). The HC group,majority of which included siblings of recruited subjects,
Table 1 Clinical characteristics of recruited subjects. Data is presented as mean (±SEM).
Characteristic T1D LADA HC
Patients, n 39 18 18Age (years) 8.6 (±0.6) 29.8 (±1.7) 18.3 (±2.1)Sex (M/F) 22/17 11/7 10/8Fasting plasma C-peptide (ng/mL) 0.47 (±0.08) 1.65 (±0.35) a 2.96 (±0.61)HbA1C (%,mmol/mol) 12.29 (±0.52), 110 (±3.3) 8.67 (±0.53) b, 70 (±3.4) 5.31 (±0.09), 34 (±0.75)BMI (kg/m2) 16.34 (±0.69) 20.27 (±0.74) c 23.2 (±1.42)Anti-Insulin antibodies (% subjects) 15/39 (38.5%) 4/18 (21.05%) –Anti GAD65/IA2 25/39 (64.1%) 9/18 (50%) –
T1D: Type 1 diabetes, LADA: latent autoimmune diabetes in adults.a Mean (±SEM) plasma C-peptide levels were significantly higher in LADA than TID subjects (p = 0.0001).b HbA1C was higher in TID compared to LADA subjects (p = 0.001).c BMI of LADA subjects was significantly higher than T1D group (p = 0.04).
80 N. Sachdeva et al.
Author's personal copy
had higher plasma C-peptide levels and lower HbA1C than thediabetic subjects.
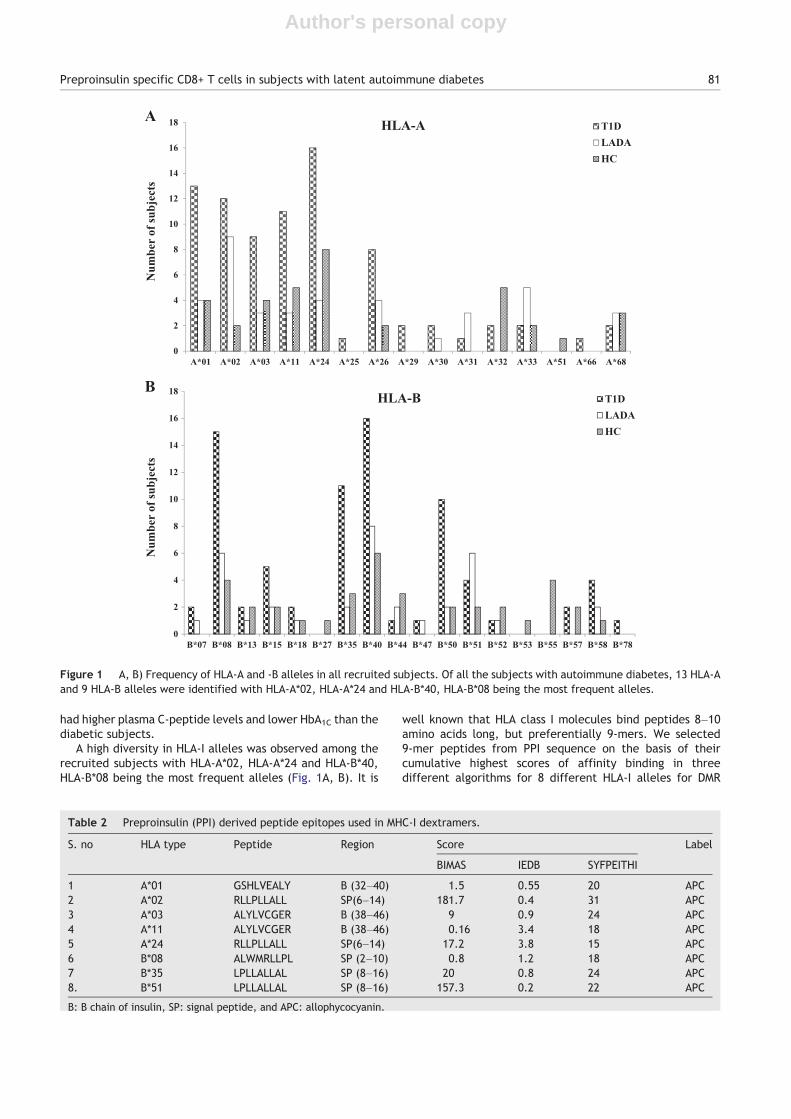
A high diversity in HLA-I alleles was observed among therecruited subjects with HLA-A*02, HLA-A*24 and HLA-B*40,HLA-B*08 being the most frequent alleles (Fig. 1A, B). It is
well known that HLA class I molecules bind peptides 8–10amino acids long, but preferentially 9-mers. We selected9-mer peptides from PPI sequence on the basis of theircumulative highest scores of affinity binding in threedifferent algorithms for 8 different HLA-I alleles for DMR
A
0
2
4
6
8
10
12
14
16
18
A*01 A*02 A*03 A*11 A*24 A*25 A*26 A*29 A*30 A*31 A*32 A*33 A*51 A*66 A*68
Num
ber
of su
bjec
ts
T1DLADAHC
HLA-A
B
0
2
4
6
8
10
12
14
16
18
B*07 B*08 B*13 B*15 B*18 B*27 B*35 B*40 B*44 B*47 B*50 B*51 B*52 B*53 B*55 B*57 B*58 B*78
Num
ber
of su
bjec
ts
T1DLADAHC
HLA-B
Figure 1 A, B) Frequency of HLA-A and -B alleles in all recruited subjects. Of all the subjects with autoimmune diabetes, 13 HLA-Aand 9 HLA-B alleles were identified with HLA-A*02, HLA-A*24 and HLA-B*40, HLA-B*08 being the most frequent alleles.
Table 2 Preproinsulin (PPI) derived peptide epitopes used in MHC-I dextramers.
S. no HLA type Peptide Region Score Label
BIMAS IEDB SYFPEITHI
1 A*01 GSHLVEALY B (32–40) 1.5 0.55 20 APC2 A*02 RLLPLLALL SP(6–14) 181.7 0.4 31 APC3 A*03 ALYLVCGER B (38–46) 9 0.9 24 APC4 A*11 ALYLVCGER B (38–46) 0.16 3.4 18 APC5 A*24 RLLPLLALL SP(6–14) 17.2 3.8 15 APC6 B*08 ALWMRLLPL SP (2–10) 0.8 1.2 18 APC7 B*35 LPLLALLAL SP (8–16) 20 0.8 24 APC8. B*51 LPLLALLAL SP (8–16) 157.3 0.2 22 APC
B: B chain of insulin, SP: signal peptide, and APC: allophycocyanin.
81Preproinsulin specific CD8+ T cells in subjects with latent autoimmune diabetes
Author's personal copy
82 N. Sachdeva et al.
Author's personal copy
analysis (Table 2). The DMR peptide complexes were furthervalidated by IFN-γ ELIspot using PHA as control (data notshown). For all HLA-I alleles, the immunodominant epitopes
were located in the signal peptide or B-chain (Table 2), someof which have been previously reported [10,11,27].
3.2. Subjects with T1D harbor higher frequency ofPPI specific CD8+ T cells than those with LADA
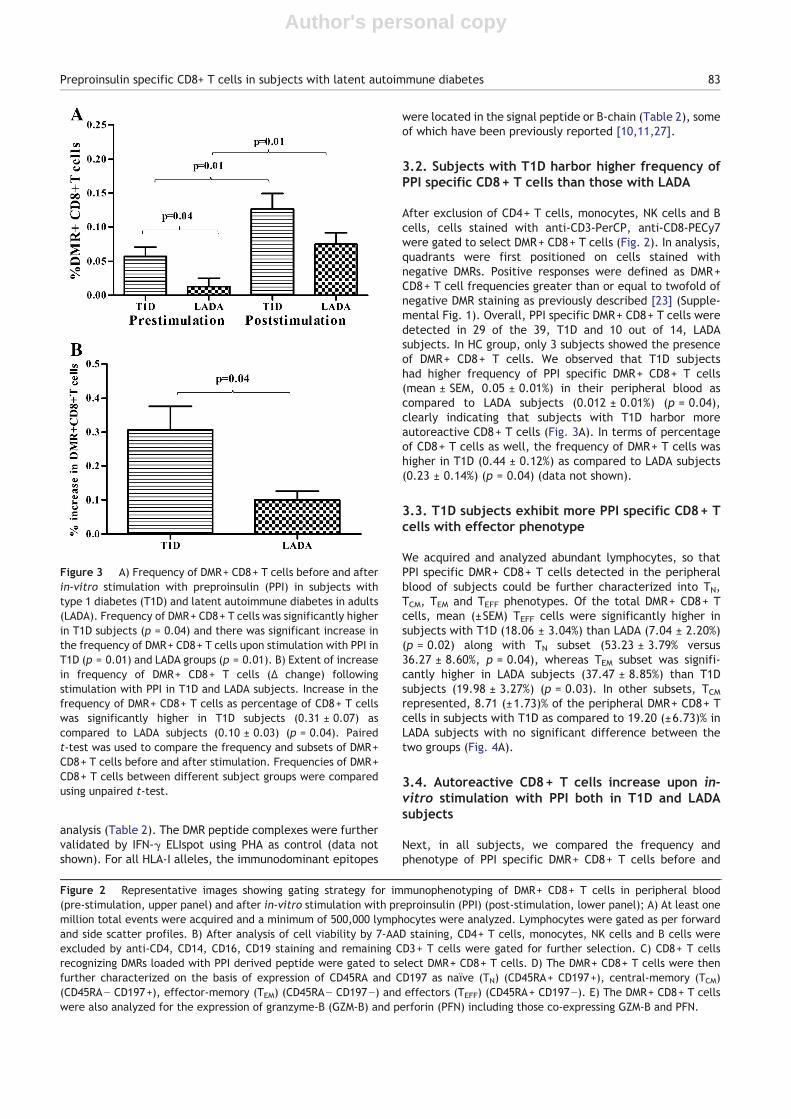
After exclusion of CD4+ T cells, monocytes, NK cells and Bcells, cells stained with anti-CD3-PerCP, anti-CD8-PECy7were gated to select DMR+ CD8+ T cells (Fig. 2). In analysis,quadrants were first positioned on cells stained withnegative DMRs. Positive responses were defined as DMR+CD8+ T cell frequencies greater than or equal to twofold ofnegative DMR staining as previously described [23] (Supple-mental Fig. 1). Overall, PPI specific DMR+ CD8+ T cells weredetected in 29 of the 39, T1D and 10 out of 14, LADAsubjects. In HC group, only 3 subjects showed the presenceof DMR+ CD8+ T cells. We observed that T1D subjectshad higher frequency of PPI specific DMR+ CD8+ T cells(mean ± SEM, 0.05 ± 0.01%) in their peripheral blood ascompared to LADA subjects (0.012 ± 0.01%) (p = 0.04),clearly indicating that subjects with T1D harbor moreautoreactive CD8+ T cells (Fig. 3A). In terms of percentageof CD8+ T cells as well, the frequency of DMR+ T cells washigher in T1D (0.44 ± 0.12%) as compared to LADA subjects(0.23 ± 0.14%) (p = 0.04) (data not shown).
3.3. T1D subjects exhibit more PPI specific CD8+ Tcells with effector phenotype
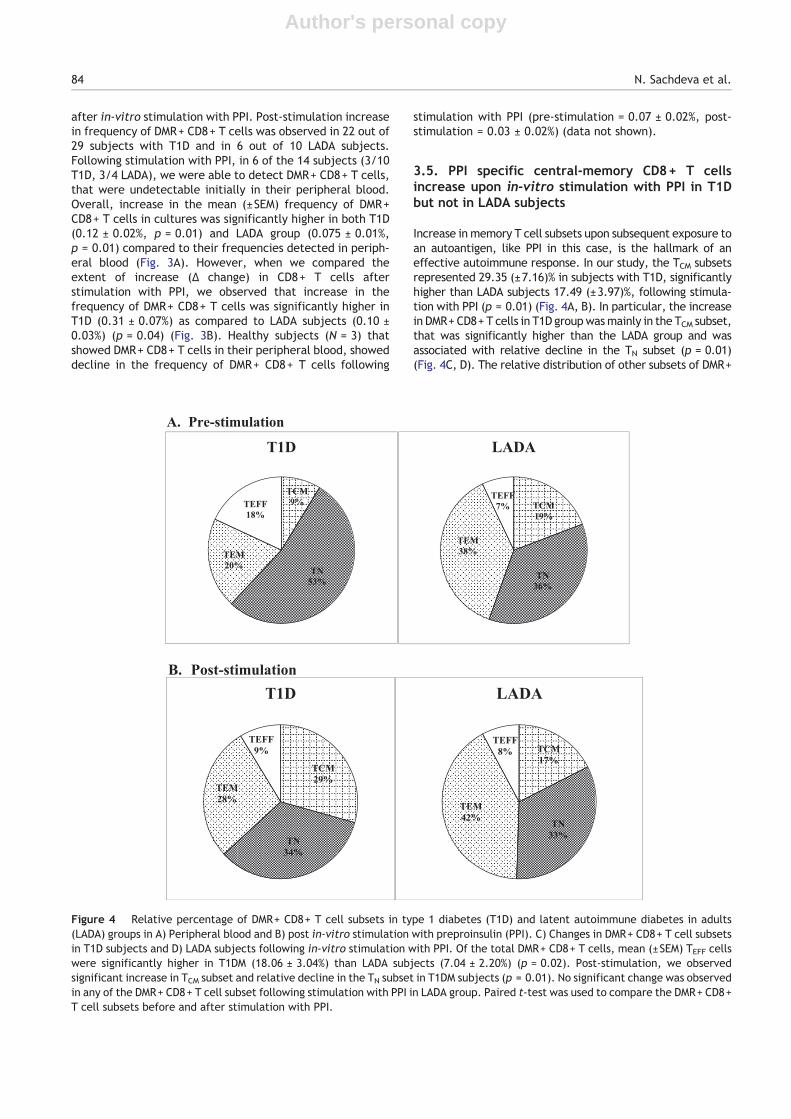
We acquired and analyzed abundant lymphocytes, so thatPPI specific DMR+ CD8+ T cells detected in the peripheralblood of subjects could be further characterized into TN,TCM, TEM and TEFF phenotypes. Of the total DMR+ CD8+ Tcells, mean (±SEM) TEFF cells were significantly higher insubjects with T1D (18.06 ± 3.04%) than LADA (7.04 ± 2.20%)(p = 0.02) along with TN subset (53.23 ± 3.79% versus36.27 ± 8.60%, p = 0.04), whereas TEM subset was signifi-cantly higher in LADA subjects (37.47 ± 8.85%) than T1Dsubjects (19.98 ± 3.27%) (p = 0.03). In other subsets, TCMrepresented, 8.71 (±1.73)% of the peripheral DMR+ CD8+ Tcells in subjects with T1D as compared to 19.20 (±6.73)% inLADA subjects with no significant difference between thetwo groups (Fig. 4A).
3.4. Autoreactive CD8+ T cells increase upon in-vitro stimulation with PPI both in T1D and LADAsubjects
Next, in all subjects, we compared the frequency andphenotype of PPI specific DMR+ CD8+ T cells before and
Figure 3 A) Frequency of DMR+ CD8+ T cells before and afterin-vitro stimulation with preproinsulin (PPI) in subjects withtype 1 diabetes (T1D) and latent autoimmune diabetes in adults(LADA). Frequency of DMR+ CD8+ T cells was significantly higherin T1D subjects (p = 0.04) and there was significant increase inthe frequency of DMR+ CD8+ T cells upon stimulation with PPI inT1D (p = 0.01) and LADA groups (p = 0.01). B) Extent of increasein frequency of DMR+ CD8+ T cells (Δ change) followingstimulation with PPI in T1D and LADA subjects. Increase in thefrequency of DMR+ CD8+ T cells as percentage of CD8+ T cellswas significantly higher in T1D subjects (0.31 ± 0.07) ascompared to LADA subjects (0.10 ± 0.03) (p = 0.04). Pairedt-test was used to compare the frequency and subsets of DMR+CD8+ T cells before and after stimulation. Frequencies of DMR+CD8+ T cells between different subject groups were comparedusing unpaired t-test.
Figure 2 Representative images showing gating strategy for immunophenotyping of DMR+ CD8+ T cells in peripheral blood(pre-stimulation, upper panel) and after in-vitro stimulation with preproinsulin (PPI) (post-stimulation, lower panel); A) At least onemillion total events were acquired and a minimum of 500,000 lymphocytes were analyzed. Lymphocytes were gated as per forwardand side scatter profiles. B) After analysis of cell viability by 7-AAD staining, CD4+ T cells, monocytes, NK cells and B cells wereexcluded by anti-CD4, CD14, CD16, CD19 staining and remaining CD3+ T cells were gated for further selection. C) CD8+ T cellsrecognizing DMRs loaded with PPI derived peptide were gated to select DMR+ CD8+ T cells. D) The DMR+ CD8+ T cells were thenfurther characterized on the basis of expression of CD45RA and CD197 as naïve (TN) (CD45RA+ CD197+), central-memory (TCM)(CD45RA− CD197+), effector-memory (TEM) (CD45RA− CD197−) and effectors (TEFF) (CD45RA+ CD197−). E) The DMR+ CD8+ T cellswere also analyzed for the expression of granzyme-B (GZM-B) and perforin (PFN) including those co-expressing GZM-B and PFN.
83Preproinsulin specific CD8+ T cells in subjects with latent autoimmune diabetes
Author's personal copy
after in-vitro stimulation with PPI. Post-stimulation increasein frequency of DMR+ CD8+ T cells was observed in 22 out of29 subjects with T1D and in 6 out of 10 LADA subjects.Following stimulation with PPI, in 6 of the 14 subjects (3/10T1D, 3/4 LADA), we were able to detect DMR+ CD8+ T cells,that were undetectable initially in their peripheral blood.Overall, increase in the mean (±SEM) frequency of DMR+CD8+ T cells in cultures was significantly higher in both T1D(0.12 ± 0.02%, p = 0.01) and LADA group (0.075 ± 0.01%,p = 0.01) compared to their frequencies detected in periph-eral blood (Fig. 3A). However, when we compared theextent of increase (Δ change) in CD8+ T cells afterstimulation with PPI, we observed that increase in thefrequency of DMR+ CD8+ T cells was significantly higher inT1D (0.31 ± 0.07%) as compared to LADA subjects (0.10 ±0.03%) (p = 0.04) (Fig. 3B). Healthy subjects (N = 3) thatshowed DMR+ CD8+ T cells in their peripheral blood, showeddecline in the frequency of DMR+ CD8+ T cells following
stimulation with PPI (pre-stimulation = 0.07 ± 0.02%, post-stimulation = 0.03 ± 0.02%) (data not shown).
3.5. PPI specific central-memory CD8+ T cellsincrease upon in-vitro stimulation with PPI in T1Dbut not in LADA subjects
Increase in memory T cell subsets upon subsequent exposure toan autoantigen, like PPI in this case, is the hallmark of aneffective autoimmune response. In our study, the TCM subsetsrepresented 29.35 (±7.16)% in subjects with T1D, significantlyhigher than LADA subjects 17.49 (±3.97)%, following stimula-tion with PPI (p = 0.01) (Fig. 4A, B). In particular, the increasein DMR+ CD8+ T cells in T1D groupwasmainly in the TCM subset,that was significantly higher than the LADA group and wasassociated with relative decline in the TN subset (p = 0.01)(Fig. 4C, D). The relative distribution of other subsets of DMR+
Figure 4 Relative percentage of DMR+ CD8+ T cell subsets in type 1 diabetes (T1D) and latent autoimmune diabetes in adults(LADA) groups in A) Peripheral blood and B) post in-vitro stimulation with preproinsulin (PPI). C) Changes in DMR+ CD8+ T cell subsetsin T1D subjects and D) LADA subjects following in-vitro stimulation with PPI. Of the total DMR+ CD8+ T cells, mean (±SEM) TEFF cellswere significantly higher in T1DM (18.06 ± 3.04%) than LADA subjects (7.04 ± 2.20%) (p = 0.02). Post-stimulation, we observedsignificant increase in TCM subset and relative decline in the TN subset in T1DM subjects (p = 0.01). No significant change was observedin any of the DMR+ CD8+ T cell subset following stimulation with PPI in LADA group. Paired t-test was used to compare the DMR+ CD8+T cell subsets before and after stimulation with PPI.
84 N. Sachdeva et al.

Author's personal copy
CD8+ T cells was similar in subjects with T1D and LADA.Post-stimulation, the mean (±SEM) DMR+ CD8+ TN cellsrepresented 33.78 (±6.15)% in T1D and 32.99 (±10.85)% inLADA, TEM represented 28.17 (±5.84)% in T1D and 41.64 (±10.58)% in LADA and TEFF represented 8.69 (±2.73)% in T1D and7.85 (±2.23)% in LADA groups, respectively.
3.6. PPI specific autoreactive CD8+ T cells exhibitsimilar expression of intracellular Perforin andGranzyme-B in T1D and LADA
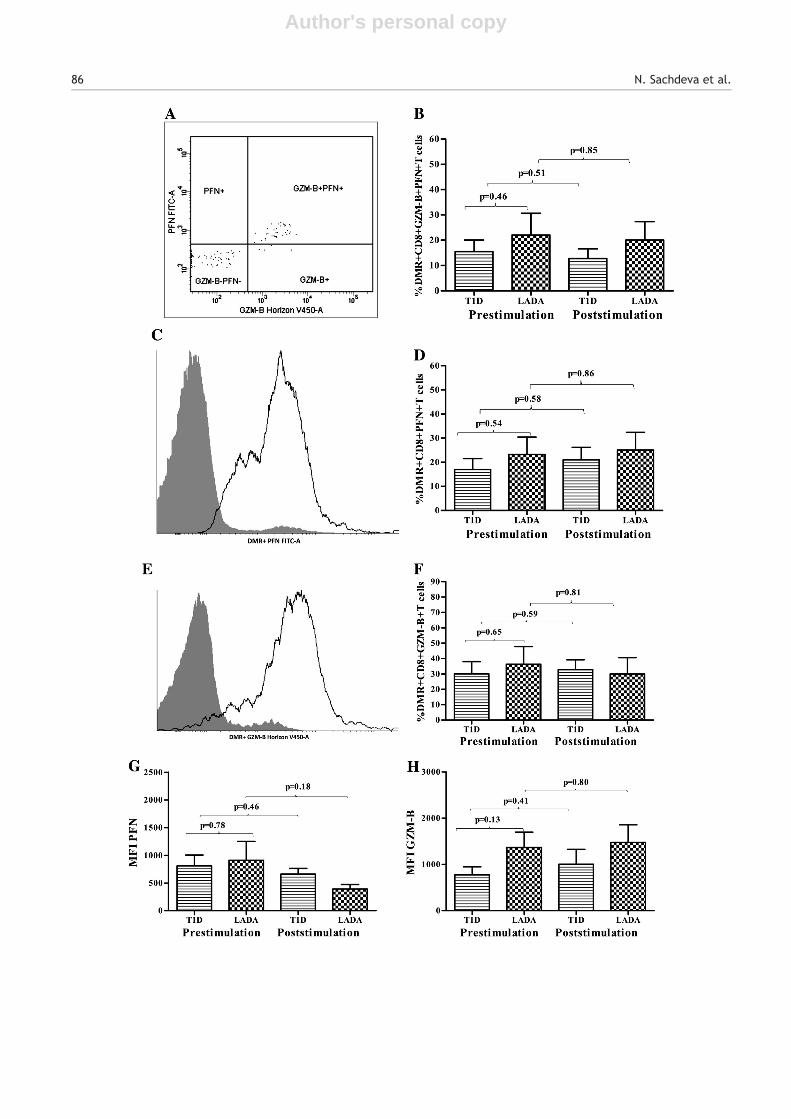
As an indicator of cytotoxic capacity, we assessed thefrequency of DMR+ CD8+ T cells expressing PFN and GZM-B.In peripheral blood we observed no significant differencein the mean (±SEM) frequency of DMR+ CD8+ T cellsco-expressing GZM-B and PFN in T1D (15.45 ± 4.55%) andLADA subjects (22.05 ± 8.56%) (p = 0.46). The frequencyof DMR+ CD8+ GZM-B+ PFN+ T cells decreased slightlyupon in-vitro stimulation with PPI in subjects with T1D
(post-stimulation, 12.71 ± 3.9%, Δ change = −2.74 ± 5.9%)though the decrease was statistically insignificant (p = 0.51).In LADA subjects as well no significant change in the frequencyof DMR+ CD8+ GZM-B+ PFN+ T cells (post-stimulation,20.03 ± 7.34%, Δ change = −2.01 ± 11.28%) (p = 0.85) wasobserved (Fig. 5A, B).
In case of PPI specific CD8+ T cells expressing PFN i.e., DMR+CD8+ PFN+ T cells, we did not observe any significant change inthe mean (±SEM) frequency in T1D group (pre-stimulation18.17 ± 4.7%; post-stimulation, 21.7 ± 5.4%, Δ change =3.53 ± 7.2%) (p = 0.58) and LADA group (pre-stimulation,23.30 ± 7.13%; post-stimulation, 25.09 ± 7.34%, Δ change =1.79 ± 10.24%) (p = 0.86) following stimulation with PPI(Fig. 5C, D). Similarly we observed no significant change inPPI specific CD8+ T cells expressing GZM-B i.e., DMR+ CD8+GZM-B+ T cells following stimulation with PPI in T1D(pre-stimulation, 30.06 ± 7.96; post-stimulation, 32.76 ±6.39%, Δ change = 2.70 ± 10.21%) (p = 0.59) and LADA groups(pre-stimulation, 34.02 ± 13.77%; post-stimulation, 29.96 ±10.61%, Δ change = −4.06 ± 17.39%) (p = 0.81) (Fig. 5E, F).
D
Figure 4 (continued).
85Preproinsulin specific CD8+ T cells in subjects with latent autoimmune diabetes
Author's personal copy
86 N. Sachdeva et al.

Author's personal copy
In order to assess expression of PFN and GZM-B per cell onPPI specific CD8+ T cells, we also compared the meanfluorescence intensity (MFI) of PFN and GZM-B on DMR+CD8+ T cells. However, in peripheral blood and post culture,we observed no significant difference in expression of PFNor GZM-B on DMR+ CD8+ T cells between the two groups (Fig. 5G, H).
3.7. AO-T1D subjects also harbor more PPI specificCD8+ T cells with TEFF phenotype and exhibit higherproliferation rate of TCM cells than LADA subjects
To investigate whether, the observed differences in DMR+CD8+ T cells are influenced by differences in age andmetabolic state of T1D and LADA subjects, we also recruited15 insulin-dependent AO-T1D subjects (mean age, 31.43 ±2.6 years; HbA1c, 9.98 ± 0.49%; fasting plasma C-peptide,0.63 ± 0.27 ng/ml) and performed immunophenotyping ofDMR+ CD8+ T cells, before and after in-vitro stimulationwith PPI. DMR positive responses were observed in 11 of the15 subjects (73.3%). Compared to LADA subjects, AO-T1Dsubjects had nearly significant higher frequency of DMR+CD8+ T cells (0.03 ± 0.01% versus 0.012 ± 0.01%, p = 0.06)with significantly higher proportion of TEFF cells (19.90 ±3.63% versus 7.04 ± 2.20% (p = 0.01) (Supplemental Table2). Similar to the T1D group, we observed a significantincrease in frequency of DMR+ CD8+ T cells followingstimulation with PPI (pre-stimulation, 0.03 ± 0.01%; post-stimulation, 0.07 ± 0.01%) (p = 0.04) (Supplemental Fig. 4)that was mainly associated with increase in TCM subset(pre-stimulation, 7.86 ± 1.6%; post-stimulation, 21.17 ±0.88%) (p = 0.02) (Supplemental Fig. 5). We observed nosignificant change in other DMR+ CD8+ T cell subsets beforeand after stimulation with PPI, including intracellular expres-sion of GZM-B and PFN (Supplemental Fig. 6).
4. Discussion
Pathogenesis of autoimmune diabetes is driven largely byislet-reactive CD8+ T cells causing beta-cell destruction.Compared to T1D, destruction of beta-cells occurs at aslower pace in LADA. In an attempt to elucidate differencesin autoreactive CD8+ T cells, we compared immunophe-notypic characteristics of PPI specific CD8+ T cells isolatedfrom their peripheral blood. At first, we measured thefrequency of PPI specific CD8+ T cells; second, we measuredthe cytolytic potential of CD8+ T cells in terms of PFN andGZM-B expression; third we studied the proliferativepotential of PPI specific CD8+ T cells along with theirdifferentiation stages, and followed the changes in immuno-phenotype of these cells after in-vitro stimulation with
PPI. Our data demonstrates that LADA subjects harbor lowerfrequency of peripheral PPI specific CD8+ T cells, particularlythose with effector phenotype and possess lesser proliferativepotential than those in T1D subjects on account of theirrelatively restrained TCM subset. To our knowledge, this is afirst report on comparison of autoreactive CD8+ T cells in thetwo forms of autoimmune diabetes encompassing individualswith diverse HLA-I alleles.
In general a greater heterogeneity in HLA-I type wasseen in our subjects, comprising of 14 different HLA-A and10 HLA-B alleles, as expected, due to genetic diversityin northern-India. For DMR analysis, we selected peptideepitopes with highest affinity, so that subjects could becompared on the same scale, despite of HLA heterogeneity.We preferred DMRs over TMRs as peptide MHC-I carriers, asDMRs carry multiple MHC-peptide complexes that interactsimultaneously with multiple TCRs, allowing for a morestable interaction with the T cell with high avidity [28].Previous studies have shown that autoreactive CD8+ Tcells are not detectable in all subjects with autoimmunediabetes. Toma et al. [10] detected the same in 48% ofrecent-onset and 25% of long standing T1D subjects, while,Luce et al. [11] reported the same in 68% of recent-onset and83% of long-standing cases. Here, we were able to detect PPIspecific autoreactive CD8+ T cells in 50 of the 68 subjects(73.5%), although the frequency was low in many subjects.This was expected as we used only single epitope tocharacterize autoreactive CD8+ T cells. Though it is possibleto screen repertoire of various PPI epitopes including thosewith low affinity using combinations of MHCmultimers [16,29],we restricted our approach to single epitope specific CD8+ Tcells as we further intended to compare their characteristics indifferent groups.
A direct association of autoreactive T cells with beta-celldestruction in pancreatic islets has been demonstrated inT1D subjects [27,30]. Since LADA has a slower rate ofprogression, it was thought that these subjects might havelow frequency of autoreactive CD8+ T cells. Here indeed, weobserved that LADA subjects had lower frequency of PPIspecific DMR+ CD8+ T cells in their peripheral circulation ascompared to T1D subjects. Till date, the only data availableon T cell responses in LADA subjects is based on a cellularimmunoblotting assay demonstrating that T cell recognitionof islet proteins is similar to those with T1D with minordifferences [31]. We further observed higher frequency ofTEFF subsets in subjects with T1D including AO-T1D, alsosuggesting that TEFF subsets are detectable in periphery evenwhen selectively sequestered in pancreas. In contrast, majorproportion of autoreactive CD8+ T cells in LADA subjectscomprised of TEM type indicating that most of the PPI specificCD8+ T cells have the potential for pancreatic migration.However it remains to be seen, whether these cells are
Figure 5 A, B) Representative flowcytograms and mean (±SEM) frequency of DMR+ CD8+ T cells expressing Granzyme-B (GZM-B)and perforin (PFN); C, D) PFN only; E,F) GZM-B only, before and after in-vitro stimulation with preproinsulin (PPI) in subjects withtype 1 diabetes (T1D) and latent autoimmune diabetes in adults (LADA). No significant difference was observed in the frequency ofDMR+ CD8+ T cells expressing PFN and/or GZM-B in T1DM and LADA subjects. G) Expression of PFN and H) GZM-B per DMR+ CD8+ T cellin terms of mean fluorescence intensity (MFI) was also assessed before and after in-vitro stimulation with PPI in T1DM and LADAsubjects. Again, we observed no significant difference in mean (±SEM) of GZM-B and PFN expression in DMR+ CD8+ T cells betweenthe two groups. Paired t-test was used to compare the percentages of DMR+ CD8+ T cells expressing PFN and GZM-B before and afterstimulation with PPI. Percentages of DMR+ CD8+ T cells expressing the same, between both subject groups were compared usingunpaired t-test.
87Preproinsulin specific CD8+ T cells in subjects with latent autoimmune diabetes

Author's personal copy
capable of mounting robust beta-cell destructive responsesin these subjects, though reports in NOD mouse models haveassigned them a major role in islet destruction [32].Recently, encouraging results were obtained from TIDALstudy that targeted TEM cells with alefacept, an engineeredfusion protein targeting surface molecule, CD2 [33]. Takentogether, these results suggest that approaches targetingTEM subsets may have better outcome in LADA subjects,compared to those with T1D.
Antigen specific stimulation is commonly used to enrichepitope specific T cells. We observed an increase in thefrequency of DMR+ CD8+ T cells in most of the subjectsfollowing in-vitro stimulation with PPI including those withundetectable peripheral DMR+ CD8+ T cells, indicating thatautoreactive CD8+ T cells in autoimmune diabetes havethe ability to proliferate upon antigen re-exposure, even ifpresent in low numbers. Notably, frequency of DMR+ CD8+ Tcells increased significantly more in T1D group, implyingthat PPI specific CD8+ T cells in subjects with T1D possessgreater proliferative potential. Another striking differencebetween T1D and LADA is the persistence and magnitudeof autoantigen, in this case PPI, which could influence thecourse of T-cell differentiation and trafficking. In addition,formation of TCM subsets following antigenic stimulationis a hallmark of robust T cell response [12]. We attemptedto look at the same T cell dynamics to understand whetherthere are any differences between the two groups indifferentiation of autoreactive CD8+ T cells followingin-vitro stimulation with PPI. Post-stimulation, there wasan increase in proportion of TCM subsets in T1D and AO-T1Dgroups, whereas the same subset remained relativelyunchanged in LADA group. Amplification of TCM cells posesa major hurdle and at the same time presents an attractivetarget to control beta-cell autoimmunity, reinforcing the factthat immunotherapeutic approaches should target antigenspecific TCM subset, besides the TEM subset and promoteformation of antigen specific regulatory T cells [34–38].Further, we also analyzed the expression of GZM-B and PFN inDMR+ CD8+ T cells, assuming that their peripheral frequencymight reflect the extent of beta-cell damage. However, theT1D and LADA subjects did not demonstrate any difference inthe expression of these effector molecules, indicating that PPIspecific CD8+ T cells bear similar pathogenicity in both thedisease groups, though we did not compare the expression ofthese effector molecules in specific subsets like TEFF cells dueto their low numbers.
Since T1D manifests more aggressively in children thanadults, a more activated immune state could be expected indiabetic children. However, like T1D, the AO-T1D subjectsdemonstrated similar differences in characteristics of PPIspecific CD8+ T cells in comparison to LADA subjects,indicating that the observed differences in antigen specificCD8+ T cells are not greatly influenced by differences in ageand such changes can be observed at disease onset.Moreover, it must be noted that LADA is a heterogeneousentity with several immunological and metabolic variations,besides the underlying, yet unknown genetic causes and itwould be too early to draw conclusions on the basis ofcharacteristics of autoreactive CD8+ T cells specific for asingle epitope. Also, since we had enrolled most of thesubjects that were recently diagnosed, persistence of theobserved CD8+ T cell characteristics in the long term,
particularly when beta-cell mass is almost destroyed,remains to be established. Although our study was focussedon PPI specific cells, the state of autoimmunity is affectedby CD8+ T cells reactive to other autoantigens as well.Nonetheless, we have at least shown that PPI specific CD8+ Tcells are present in both the slower and aggressive forms ofautoimmune diabetes with different immunophenotypiccharacteristics. In conclusion, we have demonstrated thatslower rate of disease progression in LADA is associatedwith low frequency of PPI specific CD8+ T cells which haveinferior proliferative potential on account of a relativelyrestrained TCM subset. This further implies that beta-cellprevention approaches in autoimmune diabetes must focusnot only on blunting specific TEFF and TEM cells but alsosuppress formation of TCM cells that can be assessed by usingsensitive immunophenotyping assays. Such strategies holdhigh promise in conjunction with beta-cell regeneration orislet transplant approaches.
Conflict of interest statement
The authors have no conflict of interest.
Contribution statement
NS conceptualized the study and its design. MP, RK, DB andNJ performed the experiments and acquired the data. NS,DD, AB, SKA and SKB recruited the subjects and analyzed thedata. NS and DD supervised the study. NS and MP drafted themanuscript. All authors revised the article and approved thefinal version of the manuscript. NS is the guarantor of thiswork and, as such, had full access to all of the data in thestudy and takes responsibility for the integrity of the dataand the accuracy of the data analysis.
Acknowledgments
We thank Rakesh Kumar, Department of Pediatrics, PinakiDutta, Rama Walia and Ashu Rastogi, Department ofEndocrinology, PGIMER, Chandigarh for help in recruitmentof study subjects. We also thank Ravi Sharma, Departmentof Ophthalmology, PGIMER, Chandigarh, Raj Davinder,Dhananjay Kumar and Jyoti Shailon, Department of Endocri-nology, PGIMER, Chandigarh for help in conducting theexperiments and performing routine diagnostic investiga-tions of the study subjects. We also thank participants ingiving consent and providing blood samples wheneverrequired. This study was supported by Department ofBiotechnology (DBT), Ministry of Science and Technology,Government of India (Grant No. BT/PR13480/Med/30/273/2010). The funding sources had no involvement in the studydesign, interpretation of data, or decision to submit thearticle for publication.
Appendix A. Supplementary data
Supplementary data to this article can be found online athttp://dx.doi.org/10.1016/j.clim.2015.01.005.
88 N. Sachdeva et al.

Author's personal copy
References
[1] R. Mallone, E. Martinuzzi, P. Blancou, G. Novelli, G. Afonso, M.Dolz, et al., CD8+ T-cell responses identify β-cell autoimmu-nity in human type 1 diabetes, Diabetes 56 (3) (2007) 613–621.
[2] A. Skowera, R.J. Ellis, R. Varela-Calviño, S. Arif, G.C. Huang, C.Van-Krinks, et al., CTLs are targeted to kill β cells in patientswith type 1 diabetes through recognition of a glucose-regulatedpreproinsulin epitope, J. Clin. Invest. 118 (10) (2008) 3390–3402.
[3] D. Kronenberg, R.R. Knight, M. Estorninho, R.J. Ellis, M.G.Kester, A. De Ru, et al., Circulating preproinsulin signalpeptide-specific CD8 T cells restricted by the susceptibilitymolecule HLA-A24 are expanded at onset of type 1 diabetesand kill β-cells, Diabetes 61 (7) (2012) 1752–1759.
[4] R.R. Knight, D. Kronenberg, M. Zhao, G.C. Huang, M. Eichmann,A. Bulek, et al., Human β-cell killing by autoreactivepreproinsulin-specific CD8 T cells is predominantly granule-mediated with the potency dependent upon T-cell receptoravidity, Diabetes 62 (1) (2013) 205–213.
[5] M.P. Marron, R.T. Graser, H.D. Chapman, D.V. Serreze, Function-al evidence for the mediation of diabetogenic T cell responses byHLA-A2. 1 MHC class I molecules through transgenic expression inNOD mice, Proc. Natl. Acad. Sci. 99 (21) (2002) 13753–13758.
[6] J.D. Trudeau, C. Kelly-Smith, C.B. Verchere, J.F. Elliott, J.P.Dutz, D.T. Finegood, et al., Prediction of spontaneous autoim-mune diabetes in NOD mice by quantification of autoreactive Tcells in peripheral blood, J. Clin. Investig. 111 (2) (2003) 217–223.
[7] M. Nakayama, N. Abiru, H. Moriyama, N. Babaya, E. Liu, D. Miao,et al., Prime role for an insulin epitope in the development oftype 1 diabetes in NOD mice, Nature 435 (7039) (2005) 220–223.
[8] T. Takaki, M.P. Marron, C.E. Mathews, S.T. Guttmann, R. Bottino,M. Trucco, et al., HLA-A* 0201-restricted T cells from humanizedNOD mice recognize autoantigens of potential clinical relevanceto type 1 diabetes, J. Immunol. 176 (5) (2006) 3257–3265.
[9] A. Toma, S. Haddouk, J.-P. Briand, L. Camoin, H. Gahery, F.Connan, et al., Recognition of a subregion of human proinsulinby class I-restricted T cells in type 1 diabetic patients, Proc.Natl. Acad. Sci. U. S. A. 102 (30) (2005) 10581–10586.
[10] A. Toma, T. Laïka, S. Haddouk, S. Luce, J.-P. Briand, L. Camoin,et al., Recognition of human proinsulin leader sequence by classI–restricted T-cells in HLA-A* 0201 transgenic mice and in humantype 1 diabetes, Diabetes 58 (2) (2009) 394–402.
[11] S. Luce, F. Lemonnier, J.-P. Briand, J. Coste, N. Lahlou, S. Muller,et al., Single insulin-specific CD8+ T cells show characteristicgene expression profiles in human type 1 diabetes, Diabetes 60(12) (2011) 3289–3299.
[12] F. Sallusto, J. Geginat, A. Lanzavecchia, Central memory andeffector memory T cell subsets: function, generation, andmaintenance, Annu. Rev. Immunol. 22 (2004) 745–763.
[13] F. Lemaître, H.D. Moreau, L. Vedele, P. Bousso, PhenotypicCD8+ T cell diversification occurs before, during, and after thefirst T cell division, J. Immunol. 191 (4) (2013) 1578–1585.
[14] V. Appay, R.A. van Lier, F. Sallusto, M. Roederer, Phenotypeand function of human T lymphocyte subsets: consensus andissues, Cytometry Part A 73 (11) (2008) 975–983.
[15] O. Bannard, M. Kraman, D.T. Fearon, Secondary replicativefunction of CD8+ T cells that had developed an effectorphenotype, Science 323 (5913) (2009) 505–509.
[16] J.H. Velthuis, W.W. Unger, J.R. Abreu, G. Duinkerken, K.Franken, M. Peakman, et al., Simultaneous detection ofcirculating autoreactive CD8+ T-cells specific for different isletcell-associated epitopes using combinatorial MHC multimers,Diabetes 59 (7) (2010) 1721–1730.
[17] C. Kim, M.A. Williams, Nature and nurture: T-cell receptor-dependent and T-cell receptor-independent differentiationcues in the selection of the memory T-cell pool, Immunology131 (3) (2010) 310–317.
[18] C. Boitard, T-lymphocyte recognition of beta cells in type 1diabetes: clinical perspectives, Diab. Metab. 39 (6) (2013)459–466.
[19] R. Mallone, B.O. Roep, Biomarkers for immune intervention trialsin type 1 diabetes, Clin. Immunol. 149 (3) (2013) 286–296.
[20] G. Sarikonda, J. Pettus, S. Phatak, S. Sachithanantham, J.F.Miller, J.D. Wesley, et al., CD8 T-cell reactivity to isletantigens is unique to type 1 while CD4 T-cell reactivity existsin both type 1 and type 2 diabetes, J. Autoimmun. 50 (2014)77–82.
[21] S. Cernea, K.C. Herold, Monitoring of antigen-specificCD8 T cells in patients with type 1 diabetes treated withantiCD3 monoclonal antibodies, Clin. Immunol. 134 (2) (2010)121–129.
[22] R. Mallone, S. Mannering, B. Brooks‐Worrell, I. Durinovic‐Belló,C. Cilio, F. Wong, et al., Isolation and preservation of peripheralblood mononuclear cells for analysis of islet antigen‐reactive Tcell responses: position statement of the T‐Cell WorkshopCommittee of the Immunology of Diabetes Society, Clin. Exp.Immunol. 163 (1) (2011) 33–49.
[23] R. Mallone, M. Scotto, C. Janicki, E. James, L. Fitzgerald‐Miller, R. Wagner, et al., Immunology of Diabetes SocietyT‐Cell Workshop: HLA class I tetramer‐directed epitopevalidation initiative T‐Cell Workshop Report—HLA Class ITetramer Validation Initiative, Diabetes Metab. Res. Rev. 27(8) (2011) 720–726.
[24] P. Zimmet, R. Turner, D. McCarty, M. Rowley, I. Mackay,Crucial points at diagnosis. Type 2 diabetes or slow type 1diabetes, Diabetes Care 22 (Suppl. 2) (1999) B59–B64.
[25] R.G. Naik, B.M. Brooks-Worrell, J.P. Palmer, Latent autoim-mune diabetes in adults, J. Clin. Endocrinol. Metab. 94 (12)(2009) 4635–4644.
[26] G. Afonso, M. Scotto, A. Renand, J. Arvastsson, D. Vassilieff,C.M. Cilio, et al., Critical parameters in blood processing forT-cell assays: validation on ELISpot and tetramer platforms, J.Immunol. Methods 359 (1) (2010) 28–36.
[27] K.T. Coppieters, F. Dotta, N. Amirian, P.D. Campbell, T.W. Kay,M.A. Atkinson, et al., Demonstration of islet-autoreactive CD8 Tcells in insulitic lesions from recent onset and long-term type 1diabetes patients, J. Exp. Med. 209 (1) (2012) 51–60.
[28] G. Dolton, A. Lissina, A. Skowera, K. Ladell, K. Tungatt, E.Jones, et al., Comparison of peptide‐major histocompatibilitycomplex tetramers and dextramers for the identification ofantigen‐specific T‐cells, Clin. Exp. Immunol. 177 (1) (2014)47–63.
[29] W.W. Unger, J. Velthuis, J.R. Abreu, S. Laban, E. Quinten,M.G. Kester, et al., Discovery of low-affinity preproinsulinepitopes and detection of autoreactive CD8 T-cells usingcombinatorial MHC multimers, J. Autoimmun. 37 (3) (2011)151–159.
[30] K.L. Graham, B. Krishnamurthy, S. Fynch, R. Ayala-Perez, R.M.Slattery, P. Santamaria, et al., Intra-islet proliferation ofcytotoxic T lymphocytes contributes to insulitis progression,Eur. J. Immunol. 42 (7) (2012) 1717–1722.
[31] B. Brooks-Worrell, R. Juneja, A. Minokadeh, C. Greenbaum, J.Palmer, Cellular immune responses to human islet proteins inantibody-positive type 2 diabetic patients, Diabetes 48 (5) (1999)983–988.
[32] J. Chee, H.J. Ko, A. Skowera, G. Jhala, T. Catterall, K.L. Graham,et al., Effector-memory T cells develop in islets and reportislet pathology in type 1 diabetes, J. Immunol. 192 (2) (2014)572–580.
[33] M.R. Rigby, L.A. DiMeglio, M.S. Rendell, E.I. Felner, J.M.Dostou, S.E. Gitelman, et al., Targeting of memory T cellswith alefacept in new-onset type 1 diabetes (T1DAL study):12 month results of a randomised, double-blind, placebo-controlled phase 2 trial, Lancet Diabetes Endocrinol. 1 (4)(2013) 284–294.
89Preproinsulin specific CD8+ T cells in subjects with latent autoimmune diabetes

Author's personal copy
[34] H. Wulff, M. Pennington, Targeting effector memory T-cells withKv1. 3 blockers, Curr. Opin. Drug Discov. Dev. 10 (4) (2007) 438.
[35] S.A. Long, M.R. Walker, M. Rieck, E. James, W.W. Kwok, S.Sanda, et al., Functional islet‐specific Treg can be generatedfrom CD4+ CD25− T cells of healthy and type 1 diabeticsubjects, Eur. J. Immunol. 39 (2) (2009) 612–620.
[36] J.R. Abreu, B.O. Roep, Targeting proinsulin-reactive CD8+ Tcells: a new direction for type 1 diabetes treatment, Expert.Rev. Clin. Immunol. 9 (11) (2013) 1001–1003.
[37] S.E. Gitelman, P.A. Gottlieb, M.R. Rigby, E.I. Felner, S.M.Willi, L.K. Fisher, et al., Antithymocyte globulin treatment forpatients with recent-onset type 1 diabetes: 12-month resultsof a randomised, placebo-controlled, phase 2 trial, LancetDiabetes Endocrinol. 1 (4) (2013) 306–316.
[38] K.T. Coppieters, L.C. Harrison, M.G. von Herrath, Trials in type 1diabetes: antigen-specific therapies, Clin. Immunol. 149 (3) (2013)345–355.
90 N. Sachdeva et al.




















