
Preharvest applications of growth regulators and their effect on postharvest quality of table grapes...
10
Postharvest Biology and Technology 51 (2009) 183–192 Contents lists available at ScienceDirect Postharvest Biology and Technology journal homepage: www.elsevier.com/locate/postharvbio Preharvest applications of growth regulators and their effect on postharvest quality of table grapes during cold storage Juan Pablo Zoffoli ∗ , Bernardo A. Latorre, Paulina Naranjo Facultad de Agronomía e Ingeniería Forestal, Pontificia Universidad Católica de Chile, Casilla 306-22, Santiago, Chile article info Article history: Received 7 February 2008 Accepted 25 June 2008 Keywords: Grape diseases Botrytis cinerea Gray mold Vitis vinifera Cuticle Hairline abstract Over 54,600 ha of table grapes (Vitis vinifera), mainly cvs. ‘Thompson Seedless’, ‘Flame Seedless’ and ‘Red- globe’, are planted in Chile. Almost the entire production is exported to the USA, Europe, Asia, or one of several Latin American countries, which typically requires 15–40 d of maritime transportation. During this period, several physical, physiological, and pathological factors cause table grape deterioration. Because berry size is the main quality factor in international markets, farmers often overuse the growth regula- tors, gibberellic acid (GA 3 ) and forchlorfenuron (CPPU), in an effort to increase berry size. We examined the effect of preharvest growth regulators on seedless (‘Thompson Seedless’, and ‘Ruby Seedless’) and seeded (‘Redglobe’) table grape cultivars during cold (0 ◦ C) storage plus a shelf life period of 3 d at 20 ◦ C. The overuse of GA 3 , eight instead of two GA 3 applications on Thompson Seedless, and the use of one GA 3 application on Redglobe and ‘Ruby Seedless’, increased berry pedicel thickness and lowered cuticle content but induced shatter and predisposed grapes to gray mold caused by Botrytis cinerea. In contrast, CPPU increased berry pedicel thickness and cuticle content but did not increase shatter or gray mold incidence. Clusters that were subjected to overuse of combined GA 3 and CPPU were highly sensitive to shatter, had the thickest pedicel, and developed a high gray mold incidence during cold storage. Hairline, a fine cracking developed during cold storage, was induced on ‘Thompson Seedless’ and ‘Ruby Seedless’ by growth regulators, but no hairline occurred on ‘Redglobe’ table grapes. Therefore, berry quality during cold storage is greatly influenced by growth regulator management in the vineyard. © 2008 Elsevier B.V. All rights reserved. 1. Introduction Table grapes (Vitis vinifera L.) are the main fruit crop produced in Chile with over 54,600 ha. In 2005, over 823,000 t were produced, mainly consisting of ‘Thompson Seedless’, ‘Flame Seedless’, and ‘Redglobe’ table grapes, which were exported to the USA, Europe, Asia, or one of several Latin American countries. High yields of high quality grapes are the main objectives of the Chilean table grape industry. Postharvest grape deterioration can be due to physical, physiological, or pathological factors that may occur in the vine- yard (preharvest) or after harvest. For example, rachis dehydration is a physical deterioration associated with high vapor pressure deficit between the rachis and the environment during pre- and postharvest periods (Nelson, 1985). Skin browning is the main physiological problem associated with an excessively mature Princess table grape cultivar (Vial et al., 2005). Ammonium toxicity, because of excessive nitrogen fertilization, has been associated with ∗ Corresponding author. Tel.: +56 26864159; fax: +56 25534130. E-mail address: [email protected] (J.P. Zoffoli). physiological waterberry disorder on ‘Thompson Seedless’ berries (Christensen and Boggero, 1985). Hairline cracking and bleaching develop under high SO 2 postharvest management (Gao et al., 2003; Zoffoli et al., 2008). Botrytis cinerea Pers. ex Fr. can infect during berry development, remaining latent until harvest, or appearing during storage (Holz et al., 2003; Latorre, 1986). Therefore, the inte- gration of canopy management and fungicide treatments before harvest with the use of SO 2 and cold storage (−0.5 ◦ C) after harvest are the commercial strategies implemented to control this disease (Gubler et al., 1987; Luvisi et al., 1992; Marois et al., 1992; Esterio et al., 1996; Latorre et al., 2001). Micropore density on the surface of the berry has been highly correlated with B. cinerea infection on table grapes (Blaich et al., 1984; Mlikota Gabler et al., 2003). Similarly, decay incidence increases as cuticle content decreases in the contact zone of touch- ing berries (Marois et al., 1986; Percival et al., 1993). Preharvest grape management that enhances shading (Thomas et al., 1988; English et al., 1989) and cluster tightness (Vail and Marois, 1991) increases decay susceptibility in berries of table grapes. Therefore, grape berry resistance to decay is related to the morphological fea- tures of the berries, which are genetically determined and affected by preharvest crop management in the vineyard. 0925-5214/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.postharvbio.2008.06.013
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Preharvest applications of growth regulators and their effect on postharvest quality of table grapes...

Pq
JF
a
ARA
KGBGVCH
1
Cm‘Aqi
pyidamPb
0d
Postharvest Biology and Technology 51 (2009) 183–192
Contents lists available at ScienceDirect
Postharvest Biology and Technology
journa l homepage: www.e lsev ier .com/ locate /postharvbio
reharvest applications of growth regulators and their effect on postharvestuality of table grapes during cold storage
uan Pablo Zoffoli ∗, Bernardo A. Latorre, Paulina Naranjoacultad de Agronomía e Ingeniería Forestal, Pontificia Universidad Católica de Chile, Casilla 306-22, Santiago, Chile
r t i c l e i n f o
rticle history:eceived 7 February 2008ccepted 25 June 2008
eywords:rape diseasesotrytis cinerearay molditis viniferauticleairline
a b s t r a c t
Over 54,600 ha of table grapes (Vitis vinifera), mainly cvs. ‘Thompson Seedless’, ‘Flame Seedless’ and ‘Red-globe’, are planted in Chile. Almost the entire production is exported to the USA, Europe, Asia, or one ofseveral Latin American countries, which typically requires 15–40 d of maritime transportation. During thisperiod, several physical, physiological, and pathological factors cause table grape deterioration. Becauseberry size is the main quality factor in international markets, farmers often overuse the growth regula-tors, gibberellic acid (GA3) and forchlorfenuron (CPPU), in an effort to increase berry size. We examinedthe effect of preharvest growth regulators on seedless (‘Thompson Seedless’, and ‘Ruby Seedless’) andseeded (‘Redglobe’) table grape cultivars during cold (0 ◦C) storage plus a shelf life period of 3 d at 20 ◦C.The overuse of GA3, eight instead of two GA3 applications on Thompson Seedless, and the use of oneGA3 application on Redglobe and ‘Ruby Seedless’, increased berry pedicel thickness and lowered cuticle
content but induced shatter and predisposed grapes to gray mold caused by Botrytis cinerea. In contrast,CPPU increased berry pedicel thickness and cuticle content but did not increase shatter or gray moldincidence. Clusters that were subjected to overuse of combined GA3 and CPPU were highly sensitive toshatter, had the thickest pedicel, and developed a high gray mold incidence during cold storage. Hairline,a fine cracking developed during cold storage, was induced on ‘Thompson Seedless’ and ‘Ruby Seedless’by growth regulators, but no hairline occurred on ‘Redglobe’ table grapes. Therefore, berry quality duringuence
p(dZbdgha(e
ca
cold storage is greatly infl
. Introduction
Table grapes (Vitis vinifera L.) are the main fruit crop produced inhile with over 54,600 ha. In 2005, over 823,000 t were produced,ainly consisting of ‘Thompson Seedless’, ‘Flame Seedless’, and
Redglobe’ table grapes, which were exported to the USA, Europe,sia, or one of several Latin American countries. High yields of highuality grapes are the main objectives of the Chilean table grapendustry.
Postharvest grape deterioration can be due to physical,hysiological, or pathological factors that may occur in the vine-ard (preharvest) or after harvest. For example, rachis dehydrations a physical deterioration associated with high vapor pressureeficit between the rachis and the environment during pre-
nd postharvest periods (Nelson, 1985). Skin browning is theain physiological problem associated with an excessively maturerincess table grape cultivar (Vial et al., 2005). Ammonium toxicity,ecause of excessive nitrogen fertilization, has been associated with
∗ Corresponding author. Tel.: +56 26864159; fax: +56 25534130.E-mail address: [email protected] (J.P. Zoffoli).
iigEigtb
925-5214/$ – see front matter © 2008 Elsevier B.V. All rights reserved.oi:10.1016/j.postharvbio.2008.06.013
d by growth regulator management in the vineyard.© 2008 Elsevier B.V. All rights reserved.
hysiological waterberry disorder on ‘Thompson Seedless’ berriesChristensen and Boggero, 1985). Hairline cracking and bleachingevelop under high SO2 postharvest management (Gao et al., 2003;offoli et al., 2008). Botrytis cinerea Pers. ex Fr. can infect duringerry development, remaining latent until harvest, or appearinguring storage (Holz et al., 2003; Latorre, 1986). Therefore, the inte-ration of canopy management and fungicide treatments beforearvest with the use of SO2 and cold storage (−0.5 ◦C) after harvestre the commercial strategies implemented to control this diseaseGubler et al., 1987; Luvisi et al., 1992; Marois et al., 1992; Esteriot al., 1996; Latorre et al., 2001).
Micropore density on the surface of the berry has been highlyorrelated with B. cinerea infection on table grapes (Blaich etl., 1984; Mlikota Gabler et al., 2003). Similarly, decay incidencencreases as cuticle content decreases in the contact zone of touch-ng berries (Marois et al., 1986; Percival et al., 1993). Preharvestrape management that enhances shading (Thomas et al., 1988;
nglish et al., 1989) and cluster tightness (Vail and Marois, 1991)ncreases decay susceptibility in berries of table grapes. Therefore,rape berry resistance to decay is related to the morphological fea-ures of the berries, which are genetically determined and affectedy preharvest crop management in the vineyard.
1 gy an
gtaHeet‘gsG‘1bdld3‘dtbCaSot
ibGbraeltb
2
2
‘m‘hcgvcfll‘c
2
tsP
aatstw1wNwa
ydwwtafgita
2
eso‘acw2D(t2t8fm‘pawauHaedt
2
c46‘
84 J.P. Zoffoli et al. / Postharvest Biolo
Berry size is the main quality factor affecting sales of tablerapes in international markets. Berry size is genetically prede-ermined among cultivars, but it can be considerably increased bydjusting the crop load (Kliewer and Weaver, 1971; Dokoozlian andirchfelt, 1985; Kington and Van Epenhuijsen, 1989; Dokoozliant al., 1994a), by employing cluster and berry thinning (Sharplest al., 1955), trunk girdling (Dokoozlian et al., 1994b), and withhe use of growth regulators. Gibberellic acid (GA3) is applied toThompson Seedless’ table grapes and other stenospermocarpicrape cultivars during the fruit set stage in order to increase berryize and achieve commercially acceptable fruit quality. At least twoA3 applications (40 �L L−1) are commercially recommended on
Thompson Seedless’ table grapes under Chilean conditions (Perez,994). However, growers often overuse GA3 in order to achieve theerry size required by international markets. These applicationselay fruit ripening and increase berry shatter in ‘Thompson Seed-
ess’ (Retamales and Cooper, 1993; Ben-Tal, 1990). Early work hasemonstrated that rates of GA3 application during berry set (from9.5 to 152.2 g ha−1) resulted in a 40% increase the berry weight of
Flame Seedless’ table grapes but reduced harvestable fruit by 30%ue to inadequate color development (Bianchi et al., 1991). Produc-ion of ‘Thompson Seedless’ with over applications of GA3 increaseud mortality the following growing season (Jawanda et al., 1974;ollins and Rawnsley, 2005). As a result, no more than three GA3pplications to increase berry size, have been recommended underouth African conditions (Mariette, 2007). Although overexposuref GA3 affects fruit quality and productivity, there are few reportshat have demonstrated detrimental effects during storage.
Alternatively, forchlorfenuron (CPPU) was introduced toncrease berry size. CPPU stimulates periclinal cell division in theerry, leading to more round or oval shaped berries compared withA3 application alone. This compound has strong cytokinin activity,ut it also delays maturity and red color development, and increasesachis size and pedicel thickness (Reynolds et al., 1992; Retamales etl., 1995). CPPU also increases berry shatter during storage (Navarrot al., 2001). In an effort to understand the significance the morpho-ogical changes produced in the berry, the purpose of this study waso examine the effect of preharvest growth regulators on decay anderry deterioration during cold storage.
. Materials and methods
.1. Table grapes
This study was conducted on 5-year-old table grapes, cvs.Redglobe’ (seeded) and ‘Thompson Seedless’ (seedless), in a com-
ercial vineyard located in the Central Valley near Santiago, Chile.Redglobe’ table grapes were supported on an overhead arbor 2 migh (pergola), spaced at 3.5 m × 3.5 m (816 plant ha−1) with 35anes per vine and three buds per cane. ‘Thompson Seedless’ tablerapes were spaced at 4 m × 4 m (625 plant ha−1) with 27 canes perine and 8–10 buds per cane. One cluster per shoot was left afterluster thinning just before flowering when inflorescences wereully developed, and a total of 30 and 40 clusters per plant wereeft as the final crop loads with ‘Redglobe’ and ‘Thompson Seed-ess’, respectively. This experiment was repeated in ‘Redglobe’ andRuby Seedless’ table grapes in Rancagua, Central Valley. The latterultivar was included because of its high susceptibility to decay.
.2. Growth regulator treatments
Clusters of ‘Thompson Seedless’ table grapes were subjectedo the following growth regulator treatments: (1) clusters wereprayed until run-off twice with 40 �L L−1 gibberellic acid (GA3,ro-Gibb 20% Valent Biosciences Chile S.A., Santiago, Chile) (2xGA3)
ioobT
d Technology 51 (2009) 183–192
t fruit set and 7 d later when berries were 4 and 5 mm respectively,s it is customarily done for berry growth on ‘Thompson Seedless’able grapes (Jensen et al., 1994); (2) in addition to the above 2xGA3pray applications, clusters were over-treated with GA3 by dippinghem in 20 �L L−1 GA3 for 30 s (8xGA3). These dipping treatmentsere applied on six occasions, on berries 5.5, 7.2, 11.5, 12.6, 13.2 and
4.1 mm in width, until 55 d after full bloom (DAFB). (3) Clustersere dipped in 6 �L L−1 forchlorfenuron (N-(2-chloro-4-pyridyl)--phenylurea (CPPU) (Sitofex KT-30 EC, 0.1% BASF, Santiago, Chile)hen berries were 4.7 mm in diameter and (4) clusters were treated
s described for treatments 2 and 3 (8xGA3 + CPPU), respectively.Clusters of ‘Redglobe’ table grapes from a commercial vine-
ard in Santiago were subjected to the following treatments: (1)ip application in 30 �L L−1 GA3 (1xGA3) for 30 s when berriesere 15 mm in diameter; (2) a single dip in 8 �L L−1 CPPU for 30 shen berries were 8 mm in diameter; (3) a single dip applica-
ion of GA3 and CPPU as indicated above (1xGA3 + CPPU), and (4)n equal number of non-treated berries, dipped in distilled wateror 30 s, were left as control. This protocol was repeated on ‘Red-lobe’ in Rancagua. In addition, ‘Ruby Seedless’ table grapes werencluded and treated as above. GA3 application was not included inhe control treatment. Therefore, clusters received a single dip GA3pplication.
.2.1. Effect on berry size and maturityBerry growth development was monitored on attached berries
very 10–20 d. With this purpose, three clusters per vine wereelected, and berry width and berry length were determinedn 10 berries per cluster starting at 29 DAFB until 123 DAFB.
Thompson Seedless’ table grapes in Santiago were harvestedt 103 DAFB, when the ratio between total soluble solid (TSS)ontent and titratable acidity (TA) was at least 20 or other-ise the TSS ≥ 16%. In Santiago, ‘Redglobe’ was monitored from
2 DAFB until 135 DAFB, and grapes were harvested at 111AFB. Firmness was determined using Durofel with a 3-mm tip
Copatechnology, France,). Durofel values were converted to New-ons (N = 9.8{exp[(Durofel value − 59.32)/14.89]}) (Polenta et al.,005). Firmness, berry weight, TSS (thermo-compensated refrac-ometer, Atago, Italy), TA (titrated with 4 g L−1 NaOH until pH.2), and the ratio between width and length were determinedor a 10-berry sample in each of five replicates. These measure-
ents were performed every 1–2 weeks from 50 or 60 DAFB forThompson Seedless’ or ‘Redglobe’, respectively. At harvest, theedicel thickness was determined at the insertion of the berry,nd the diameter of the main and lateral stems of the rachisas determined. Red color intensity of the berries was measured
ccording to the CIE scale (Commision Internale de l’Eclairage)sing a Minolta colorimeter (Minolta, CR-200 Japan) with 1: (red,◦: 71.6; C: 7.3; L: 34%), 2: (purple, H◦: 194.3; C: 5.1; L: 31.9%),nd 3: (dark red, H◦: 328.9; C: 3.1; L: 31%). In Rancagua, theffect of growth regulators on ‘Redglobe’ and ‘Ruby Seedless’ wasetermined only at harvest, 121 DAFB, and 108 DAFB, respec-ively.
.2.2. Cuticle content, microcracks and micropore numberTo determine the possible effect of growth regulators on cuticle
ontent, microcracks, and micropore number, fruit were assessed8, 60, 76, 88, 103 and 123 DAFB for ‘Thompson Seedless’ and 49,4, 83, 111, 135 DAFB for ‘Redglobe’ in Santiago. In ‘Redglobe’ and
Ruby Seedless’, from Rancagua, these parameters were character-
zed only at harvest (121 and 108 DAFB, respectively). With thisbjective, 70 disks (50 mm2) of berry skin were removed from theuter layer of 35 berries in each of five replicates (two disks pererry). Disk samples were taken in the middle portion of the berry.he cuticle proper/epicuticular wax layer was then separated from
gy an
tH1t
itm(li6ribascwp2
2
miwpgop
ts(fcatw(rwlbdb
cf
2
rpiwf(Pwge
3
3
3
mtiP(sta(
(c82ewCw
fiba18
ottlb8rt1
e(arorom(
(p(nHm
3
J.P. Zoffoli et al. / Postharvest Biolo
he underlying epidermal cell tissue in 0.22 mol L−1 ZnCl2 in 36%Cl for 24 h at room temperature (20–22 ◦C) (Holloway and Baker,968). Disks were then rinsed three times with distilled water andhe cuticles were dried at 30 ◦C for 2 h and their weight determined.
Microcracks were determined as the total number of cracksn 10 berries per replication from 50 mm2 disks of berry skinaken at the middle portion of each of 50 berries per treat-
ent. Disk tissue was examined in a UV epifluorescent microscopeNikon Optihop II, Nikon filter B-2A, 450–490 nm excitation wave-ength, 520 nm emission wavelength) after incubation for 4 minn 0.1% Acridine Orange in 0.05 mol L−1 citric acid buffer, pH.5, containing 0.5% Tween 20 (polyethylenesorbitan monolau-ate) (Peschel and Knoche, 2005). The number of microporesn the berries skin was determined after dipping each of 10erries in 10% fluorescein, sodium salt (Sigma, USA) for 5 minnd rinsing them in deionized water for 30 s. Ten disc (50 mm2)amples per replicate were examined under a UV epifluores-ent microscope lens (Nikon Optihop II, 330–385 nm excitationavelength, ×40 magnification) and the number of microporeser 50 mm2 of berry skin was recorded (Mlikota Gabler et al.,003).
.2.3. Effect on postharvest quality after storageIndependent of grape cultivars, clusters were harvested at com-
ercial maturity (>16% TSS) and individual clusters were packagedn perforated bags with 5% vented area. Then five bagged clusters
ere distributed inside a 0.3% perforated polyethylene liner andackaged in a 0.40 m × 0.30 m × 0.12 m, 5 kg carton box. One SO2enerator pad per box, provided with fast and slow release phasesf 1.2 g kg−1 sodium metabisulfite (Uvas Quality, Santiago, Chile)er box was used to prevent decay (Franck et al., 2005).
Grapes were subjected to a fast cooling for 12 h in forced airunnels at 0 ◦C to lower berry temperature to 1–4 ◦C and were thentored for 60 d (‘Thompson Seedless’ and ‘Ruby Seedless’) and 90 d‘Redglobe’) at 0 ◦C. After cold storage, grapes were maintainedor 3 d at 20 ◦C, without a generator pad, to simulate a commer-ialization period before the incidences of shatter, hairline, split,nd decay were determined. Shatter incidence was the propor-ion (by weight) of detached berries left in the box after clustersere removed. Decay incidence was expressed as the proportion
by weight) of berry cheek rot or berry rot at the base of the berryelative to total grape weight (Holz et al., 2003). Hairline incidenceas determined after staining 100 berries with 0.1 mol L−1 methy-
ene blue (Merck, Darmstadt, Germany) solution for 30 s followedy a brief rinse with water (Zoffoli et al., 2008). Split berries wereetermined by visual inspection of the total berries within eachox.
On ‘Redglobe’, bleaching incidence was assessed by visuallylassifying each berry according to the proportion of bleached sur-ace as low (<10%), moderate (11–30%) or high (>30%) incidence.
.3. Statistical analysis
Growth regulator treatments were distributed according to aandomized complete block design with five replications of twolants as experimental units in ‘Thompson Seedless’ and ‘Redglobe’
n Santiago and ‘Redglobe’ and ‘Ruby Seedless’ in Rancagua. Dataere subjected to an analysis of variance, and means were dif-
erentiated according to Fisher’s least significant difference (LSD)P ≤ 0.05) using Statgraphic (Systat Software, Inc., Maryland, USA).ercent values were transformed to arcsin sqrt(x) before analysis,here x = TSS, TA, shatter, hairline, split, hairline + split, bleaching,
ray mold incidence. Postharvest evaluation included one box ofach replication.
5eac1
d Technology 51 (2009) 183–192 185
. Results
.1. Effect on berry size and maturity
.1.1. ‘Thompson Seedless’Based on the evolution of TSS content, three groups of treat-
ents were significantly differentiated (P < 0.05) among berriesreated with growth regulators from 50 DAFB until 83 DAFB. Dur-ng berry development, a second order polynomial curve (R2 > 0.97,< 0.0001) best described the relation between TSS and time
Fig. 1A). At harvest (103 DAFB), TSS varied from 15.4 to 17.6%, andignificant differences in TSS concentration were obtained amongreatments with lowest in berries treated with 8xGA3 + CPPUnd the highest in berries treated with 2xGA3 or 2xGA3 + CPPUTable 1).
At harvest, the effect of growth regulator was significantP < 0.01) on TA, and mean differences in TA were also signifi-ant (P = 0.05) and varied from 0.81 to 0.99% with 8xGA3 andxGA3 + CPPU, respectively (Table 1). Similarly, the TSS:TA ratio was0 at harvest for 2xGA3 treated berries and for berries subjected toight GA3 applications or 2xGA3 + CPPU. However, this same ratioas reached only 123 DAFB in berries that received both 8xGA3 andPPU (data not shown). No significant differences in berry firmnessere obtained at harvest (Table 1).
Independent of growth regulator treatments, berry width wastted to a second order polynomial (R2 > 0.97, P < 0.0001). Theerry width increased rapidly from 27 to 80 DAFB and there-fter remained almost constant, reaching 15.9, 16.7, 17.1, and7.8 mm in berries treated with 2xGA3, 8xGA3, 2xGA3 + CPPU andxGA3 + CPPU, respectively, at harvest (103 DAFB) (Fig. 1E).
The evolution of the length of the berries was fitted to a sec-nd order polynomial (R2 > 0.94, P < 0.0001). Berries from 2xGA3reatment grew slowly after 103 DAFB. However, berries from otherreatments continuously grew until 123 DAFB (Fig. 1G). The berryength at harvest was 25.5, 26.3, 26.7, and 27.6 mm for controlerries (2xGA3), and berries subjected to 8xGA3, 2xGA3 + CPPU orxGA3 + CPPU, respectively (Table 1). The final length:width (L:W)atio for control (2xGA3) berries was significantly larger. Therefore,he L:W ratio obtained for control berries was 1.6 compared with.56, the mean of L:W ratio of all other treatments (data not shown).
The evolution of berry weight from 50 to 123 DAFB was bestxplained by a second order polynomial (R2 > 0.92, P < 0.0001)Fig. 1C). Berries from 2xGA3 treated berries or 8xGA3 reached
final constant weight of 4.6 g and 5.4 g at harvest 103 DAFB,espectively. In contrast, mean berry weights of 6.2 and 7 g werebtained for berries treated with 2xGA3 + CPPU and 8xGA3 + CPPU,espectively. Berries from these treatments reached a final weightf 6.8 and 7.4 g at 123 DAFB. A significant second order polyno-ial described the relation between weight development and time
R2 > 0.97, P < 0.0001).The effect of growth regulator treatments was significant
P < 0.001) on pedicel thickness and lateral rachis thickness. Meanedicel thickness significantly (P = 0.05) increased from 1.6 mmcontrol) to 2.9 mm (8xGA3 + CPPU). Similarly, lateral rachis thick-ess varied from 3.5 mm (control) to 4.6 mm (8xGA3 + CPPU).owever, growth regulator treatment had no significant effect onain rachis thickness (Table 1).
.1.2. ‘Redglobe’In Santiago, low change in TSS content was observed between
9 DAFB and 79 DAFB, increasing rapidly after this period. Differ-nces in TSS obtained were significant (P ≤ 0.05) among treatmentst harvest (111 DAFB). The lowest (16%) and highest (17.3%) con-entrations of TSS were obtained when berries were treated withxGA3 + CPPU and control berries, respectively. The evolution of
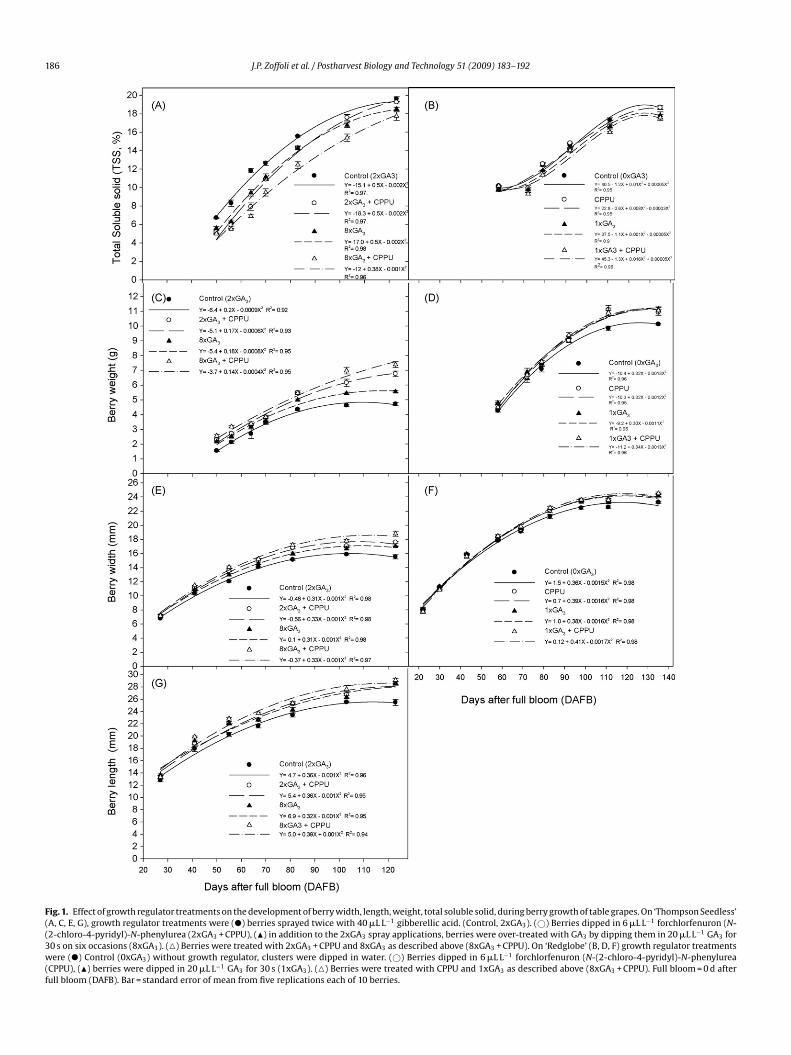
186 J.P. Zoffoli et al. / Postharvest Biology and Technology 51 (2009) 183–192
Fig. 1. Effect of growth regulator treatments on the development of berry width, length, weight, total soluble solid, during berry growth of table grapes. On ‘Thompson Seedless’(A, C, E, G), growth regulator treatments were (�) berries sprayed twice with 40 �L L−1 gibberellic acid. (Control, 2xGA3). (©) Berries dipped in 6 �L L−1 forchlorfenuron (N-(2-chloro-4-pyridyl)-N-phenylurea (2xGA3 + CPPU), (�) in addition to the 2xGA3 spray applications, berries were over-treated with GA3 by dipping them in 20 �L L−1 GA3 for30 s on six occasions (8xGA3). (�) Berries were treated with 2xGA3 + CPPU and 8xGA3 as described above (8xGA3 + CPPU). On ‘Redglobe’ (B, D, F) growth regulator treatmentswere (�) Control (0xGA3) without growth regulator, clusters were dipped in water. (©) Berries dipped in 6 �L L−1 forchlorfenuron (N-(2-chloro-4-pyridyl)-N-phenylurea(CPPU), (�) berries were dipped in 20 �L L−1 GA3 for 30 s (1xGA3). (�) Berries were treated with CPPU and 1xGA3 as described above (8xGA3 + CPPU). Full bloom = 0 d afterfull bloom (DAFB). Bar = standard error of mean from five replications each of 10 berries.

J.P.Zoffolietal./Postharvest
Biologyand
Technology51
(2009)183–192
187
Table 1Effect of growth regulator treatments on the quality of berries of ‘Thompson Seedless’, ‘Redglobe’ and ‘Ruby Seedless’ table grapes (Vitis vinifera) at harvest
Treatmentsa Firmness Newtons (N)b TSSc (%) TAc (%) Weight (g) Color (1–3)e Size (mm) Rachis thicknessc (mm) Microcrack (no. per50 mm2 disk)
Micropore (no. per50 mm2 disk)
Cuticle (g m−2)
Width Length P M L
‘Thompson Seedless’, SantiagoControl (2xGA3) 11.5ns 17.6 bd 0.83 abd 4.6 ad nd 15.9 ad 25.5 ad 1.6 ad 5.9ns 3.5 ad 0.48 ad 24.4 bd 2.78 abd
2xGA3 + CPPU 13.2 17.6 b 0.90 b 6.2 c nd 17.1 b 26.7 ab 1.7 b 6.3 3.5 a 1.08 b 25.2 b 3.11 b8xGA3 12.5 16.8 b 0.81 a 5.4 b nd 16.7 b 26.3 a 2.5 c 5.8 3.9 b 0.38 a 12.9 a 2.58 a8xGA3 + CPPU 14.0 15.4 a 0.99 c 7.0 d nd 17.8 c 27.6 b 2.9 d 6.7 4.6 c 1.00 b 16.3 a 3.03 b
‘Redglobe’, SantiagoControl (0xGA3) 11.3 b 17.3 c 0.61 b 9.8 a 2.27 b 22.5 a nd 1.7 a 4.8 a 1.9 a 0.00ns 12.9ns 3.40 bCPPU 9.0 a 16.7 b 0.60 b 11.1 b 2.47 b 23.6 b nd 2.3 b 4.6 a 2.4 b 0.00 12.3 3.58 b1xGA3 12.5 bc 16.6 b 0.52 a 10.9 b 2.13 b 23.3 b nd 2.5 b 4.6 a 2.3 b 0.00 14.8 2.98 a1xGA3 + CPPU 13.9 c 16.0 a 0.57 ab 10.9 b 1.70 a 23.5 b nd 3.3 c 5.4 b 3.1 c 0.00 14.1 3.30 ab
‘Redglobe’, RancaguaControl (0xGA3) 7.4ns 16.1 b 0.54ns 11.5ns 2.31ns 25.3 a nd 1.6 a 4.0ns 2.1 a 0.00ns 6.9 a 3.28 bCPPU 9.4 16.1 b 0.51 13.3 2.03 26.4 bc nd 2.2 b 3.2 2.3 a 0.00 7.2 a 3.41 b1xGA3 8.2 15.4 ab 0.51 11.9 2.10 25.6 ab nd 2.3 b 3.6 2.4 a 0.06 10.3 a 2.90 a1xGA3 + CPPU 8.1 14.4 a 0.52 13.1 2.17 27.0 c nd 4.8 c 3.5 3.0 b 0.02 15.1 b 3.27 b
‘Ruby Seedless’, RancaguaControl (0xGA3) 7.5 a 17.7 b 0.74 b 4.4 a 2.52ns 16.6 a 21.1 a 1.4 a 5.5ns 2.9ns 0.22 a 17.2 a 1.83 bCPPU 9.4 a 16.8 b 0.81 c 6.2 b 2.31 19.1 b 22.7 b 1.5 a 6.2 3.4 0.06 a 19.6 ab 2.24 c1xGA3 9.3 a 17.2 b 0.73 b 6.2 b 2.64 19.9 c 24.1 c 1.5 a 6.3 3.4 1.98 b 28.4 bc 1.69 ab1xGA3 + CPPU 12.8 b 15.7 a 0.66 a 6.5 b 2.34 19.6 c 23.4 bc 2.4 b 5.7 3.5 1.86 b 31.1 c 1.57 a
a On ‘Thompson Seedless’, control consisted in two spray applications of 40 �L L−1 gibberellic acid (GA3) (Pro-Gibb 20% Valent Biosciences Chile S.A.); on ‘Redglobe’ and ‘Ruby Seedless’, control were treated only with water.CPPU = dip application in 6 �L L−1 forchlorfenuron (N-(2-chloro-4-pyridyl)-N-phenylurea (Sitofex KT-30 EC, 0.1% BASF, Santiago, Chile).
b Firmness was determined using Durofel (Copatechology, France) with 3 mm tip in diameter. Durofel values were converted to Newtons (N = 9.8{exp[(Durofel value − 59.32)/14.89]}.c TSS = total soluble solid, TA = titratable acidity, P = pedicel at the insertion of the berry to the rachis, M = main rachis of the cluster, L = lateral rachis of the cluster.d Means of five replicates followed by the same letters within each table grape cultivar were not statistically significant according to Fisher’s least significant difference (LSD) (P ≤ 0.05). Percent values were transformed to
arcsin sqrt(x), where x = TSS, TA, before analysis but, untransformed values are presented. ns = non-significant. nd = not determined.e Every berry was assessed in term of color note: 1 = red, 2 = purple, 3 = dark red.

1 gy and Technology 51 (2009) 183–192
Tp
sthfi(
w(asn
3toat(
tcm
bDb(
3
epct(sit4aB(t
3
3
an1ca428
ttcaD
Fig. 2. Effect of growth regulators on cuticle content of ‘Thompson Seedless’ and‘Redglobe’ table grape cvs. during berry development. Growth regulator treatmentswere (A) ‘Thompson Seedless’ (�) Berries sprayed twice with 40 �L L−1 gibberellicacid. (Control, 2xGA3); (©) berries dipped in 6 �L L−1 forchlorfenuron (N-(2-chloro-4-pyridyl)-N-phenylurea (CPPU); (�) in addition to the 2xGA3 spray applications,berries were over-treated with GA3 by dipping them in 20 �L L−1 GA3 for 30 s onsix occasions (8xGA3); (�) berries were treated with CPPU and 8xGA3 as describedabove (8xGA3 + CPPU). (B) ‘Redglobe’ (�) Control (0xGA3) without growth regulator,clusters were dipped in water; (©) berries dipped in 6 �L L−1 forchlorfenuron (N-(2-cf(B
4m(th(
at1t
3
bta1
88 J.P. Zoffoli et al. / Postharvest Biolo
SS content during berry development was fitted to a third orderolynomial (R2 > 0.95, P < 0.0001) (Fig. 1B).
The growth regulator treatments had significant effects on berryize and berry weight at harvest. Berry width varied from 22.5 (con-rol) to 23.6 mm (CPPU or 1xGA3 or 1xGA3 + CPPU treatments) atarvest, and the relationship between DAFB and berry weight wastted to a second order polynomial (R2 > 0.95, P < 0.0001) (Table 1)Fig. 1D).
Clusters treated with 1xGA3 had the lowest TA (0.52%) comparedith all other treatments at harvest (111 DAFB) (Table 1). Significant
P = 0.05) differences in the TSS:TA ratio were obtained at harvestmong growth regulator treatments, being the lowest in clustersubjected to CPPU and the highest in clusters treated with GA3 (dataot shown).
Pedicel thickness increased from 1.7 mm (control) to 2.5 and.3 mm with 1xGA3 and 1xGA3 + CPPU, respectively. Clusters onlyreated with CPPU produced pedicels thickness similar to thosebtained on clusters treated with 1xGA3. The thickness of the lateralnd main rachis increased significantly (P = 0.05) from 1.9 (con-rol) to 3.1 mm (1xGA3 + CPPU), and from 4.8 (control) to 5.4 mm1xGA3 + CPPU), respectively (Table 1).
Development of berry color was influenced by growth regula-or treatments, 1xGA3 + CPPU treatment produced the lightest redolor berries with a 1.7 value compared with 2.27 of control treat-ent (Table 1).In Rancagua, growth regulator treatments significantly affected
erry size, TSS content and thickness of the pedicel at harvest (121AFB). However, the effects of growth regulator treatments on TA,erry weight, color and firmness at harvest were not significantTable 1).
.1.3. ‘Ruby Seedless’The growth regulator treatment had a significant (P < 0.001)
ffect on TSS, berry size (length and width), berry weight, TA,edicel thickness and firmness at harvest (108 DAFB). The TSSontent significantly (P = 0.05) varied from 15.7 (1xGA3 + CPPU)o 17.7% (control). Differences in berry length were significantP = 0.05), with 24.1 mm for the longest berries (1xGA3), and thehortest (21.1 mm) berries were obtained on control clusters. Sim-larly, berry width varied significantly (P = 0.05) from 16.6 (0xGA3)o 19.9 mm for 1xGA3. Berry weight increased significantly from.4 (control) to 6.5 g (1xGA3 + CPPU). TA values varied significantlymong treatments from 0.66% (1xGA3 + CPPU) to 0.81% (CPPU).erry firmness varied significantly from 7.5 N (control) to 12.8 N1xGA3 + CPPU). Pedicel thickness increased from 1.4 mm (control)o 2.4 mm (1xGA3 + CPPU) (Table 1).
.2. Cuticle content, microcracks and micropore number
.2.1. Cuticle contentIn ‘Thompson Seedless’ and ‘Redglobe’ in Santiago or Rancagua
nd ‘Ruby Seedless’ in Rancagua, growth regulators had a sig-ificant (P < 0.001) effect on cuticle content at harvest (103, 111,21, 108 DAFB, respectively). On ‘Thompson Seedless’, cuticleontent significantly varied from 2.58 to 3.11 g m−2 on 8xGA3nd 2xGA3 + CPPU-treated berries, respectively (Table 1). During8–123 DAFB, cuticle content decreased from 3.53 to 2.48, 3.76 to.71, 4.08 to 2.97, and 4.68 to 3.06 g m−2 for 8xGA3, 2xGA3 (control),xGA3 + CPPU, and 2xGA3 + CPPU, treatments respectively (Fig. 2A).
On ‘Redglobe’, in Santiago and Rancagua, mean differences in
he content of berry cuticle at harvest (111 and 121 DAFB, respec-ively) were significant (P = 0.05). On ‘Redglobe’ Santiago, the lowestontent was 2.98 g m−2, and the highest was 3.58 g m−2 for 1xGA3nd CPPU treatments, respectively. Cuticle content during 48–135AFB varied from 3.65 to 2.93, 3.99 to 3.10, 4.07 to 3.41 andmca08
hloro-4-pyridyl)-N-phenylurea (CPPU); (�) berries were dipped in 20 �L L−1 GA3
or 30 s (1xGA3); (�) berries were treated with CPPU and 1xGA3 as described above1xGA3 + CPPU). Mean of five replications each of 35 berries. Full bloom = 0 DAFB.ar = standard error.
.19 to 3.46 g m−2 for 1xGA3, 1xGA3 + CPPU, control, CPPU treat-ents, respectively (Fig. 2B). On ‘Redglobe’ at Rancagua, the lowest
2.9 g m−2) content of cuticle at harvest was obtained in berriesreated with 1xGA3, and it was significantly different from theighest content (3.41 g m−2) obtained on grape treated with CPPUTable 1).
On ‘Ruby Seedless’, the differences in the content of berry cuticlet harvest (108 DAFB) were significant (P = 0.05). The lowest con-ent (1.57 g m−2) of cuticle was obtained in the berries treated withxGA3 + CPPU and the highest concentration (2.24 g m−2) in berriesreated with CPPU (Table 1).
.2.2. Microcracks and microporesOn ‘Thompson Seedless’, the number of microcracks per 50 mm2
erry skin tissue increased from 0.16 to 0.43 on fruit 2xGA3reated berries and from 0.12–0.04 to 2.7–2.4 in 8xGA3 + CPPUnd 2xGA3 + CPPU, respectively, including the period from 76 to23 DAFB. In the case of 8xGA3 treated berries, the amount of
icrocracks remained relatively low (0.38) until 103 DAFB, andonsiderably increased (0.98) at 123 DAFB (Fig. 3). At harvest themount of microcracks was significantly (P = 0.05) different, with.48, 0.38, 1.0 and 1.08 for berry treated with 2xGA3, 8xGA3,xGA3 + CPPU and 2xGA3 + CPPU, respectively. Microcracks were

J.P. Zoffoli et al. / Postharvest Biology an
Fig. 3. Effect of growth regulators on microcrack incidence of ‘Thompson Seedless’table grape cv. during berry development. Growth regulator treatments were (�)berries sprayed twice with 40 �L L−1 gibberellic acid. (Control, 2xGA3). In additionto the 2xGA3 spray applications, berries were (©) dipped in 6 �L L−1 forchlorfenuron(N-(2-chloro-4-pyridyl)-N-phenylurea (CPPU); (�) over-treated with GA3 by dippingtwo
atoC‘e
o
SSi(
3
oaassor‘
4
blgbosa2
According to this study, the largest berries on ‘Thompson
TEt
T
‘
‘
‘
‘
‘0
v
wn
hem in 20 �L L−1 GA3 for 30 s on six occasions (8xGA3); (�) berries were treatedith CPPU and 8xGA3 as described above (8xGA3 + CPPU). Full bloom = 0 DAFB. Mean
f five replications each of 10 disks of 50 mm2 of berry skin.
lso detected on berries from ‘Ruby Seedless’, at harvest. In con-rol berries, where no growth regulator was applied, the amountf microcracks was 0.22 compared with 0.06, 1.86 and 1.98 withPPU, 1xGA3 and 1xGA3 + CPPU-treated berries, respectively. On
Redglobe’, microcracks were not detected at any of the vineyardsvaluated (Table 1).
Growth regulator treatments significant affected the microp-re density (micropores 50 mm−2) of the berry skin in ‘Thompson
Sacm
able 2ffect of growth regulator treatments on firmness, shatter, hairline, splitting, hairline plushe berry base of table grape (Vitis vinifera) berries after 60 d (‘Thompson Seedless’ and ‘R
reatmentsa Firmness Newtons (N)b Shatter (%) Hairline (%) Splitting (%
Thompson Seedless’, SantiagoControl (2xGA3) 5.4ns 2.9 ac 2.9 ac 1.2 ac
2xGA3 + CPPU 5.5 2.9 a 7.8 b 6.2 b8xGA3 5.8 10.6 b 12.2 b 3.2 a8xGA3 + CPPU 5.8 9.5 b 7.0 b 7.0 b
Redglobe’, SantiagoControl (0xGA3) 7.4 b 0.4 a 0.00 0.0ns
CPPU 7.0 b 0.4 a 0.00 0.01xGA3 6.2 a 1.9 a 0.00 0.01xGA3 + CPPU 6.9 ab 15.1 b 0.10 0.1
Redglobe’, RancaguaControl (0xGA3) 6.7ns 0.1 a 0.00 0.0ns
CPPU 6.8 0.2 a 0.00 0.01xGA3 6.8 1.0 a 0.00 0.01xGA3 + CPPU 6.5 6.4 b 0.00 7.1
Ruby Seedless’, RancaguaControl (0xGA3) 7.4 a 2.6 a 1.2 a 0.0ns
CPPU 9.5 b 4.5 ab 3.7 b 0.01xGA3 7.4 a 5.1 bc 3.0 b 0.21xGA3 + CPPU 9.9 b 7.5 c 6.3 b 3.6
a On ‘Thompson Seedless’, control consisted in two spray applications of 40 �L L−1 gibRuby Seedless’, control clusters were treated only with water. CPPU = dip application in.1% BASF, Santiago, Chile).b Firmness was determined using Durofel (Copatechology, France) with 3 mm tip
alue − 59.32)/14.89]}.c Means of five replicates followed by the same letters were not statistically significanere transformed to arcsin sqrt(x) (x = Shatter, hairline, splitting, hairline + splitting, bleachs = non-significant. nd = not determined.
d Technology 51 (2009) 183–192 189
eedless’ (12.9–25.2), ‘Redglobe’, Rancagua (6.9–15.1) and ‘Rubyeedless’ (17.2–31.1). Growth regulator treatments had no signif-cant effect on micropore density on ‘Redglobe’ berries in SantiagoTable 1).
.3. Effect on postharvest quality
The incidences of shatter, hairline, splitting, and gray moldn ‘Thompson Seedless’ table grapes were significantly (P < 0.001)ffected by growth regulator treatments after 60 d at 0 ◦C plus 3 dt 20 ◦C. The effect of growth regulator was significant on firmness,hatter, bleaching, and gray mold incidence in ‘Redglobe’, and it wasignificant on firmness, shatter, hairline, and gray mold incidencen ‘Ruby Seedless’ at 90 and 60 d of storage at 0 ◦C plus 3 d at 20 ◦C,espectively. Irrespective of location, no hairline was developed onRedglobe’ (Table 2).
. Discussion
The best prices of table grapes are always obtained for largeerries in the international market. Therefore, in order to obtain the
argest possible berries, farmers often overused GA3, repeating thisrowth regulator treatment six to seven times between the smallerry (4 mm in diameter) stage until veraison. Alternatively, the usef CPPU has been suggested. CPPU is a potent growth regulator withtrong cytokinin activity (Reynolds et al., 1992). The use of CPPU isccepted by the Environmental Protection Agency in the USA (EPA,003).
eedless’ were obtained with many repeated applications of GA3lone or with CPPU applied after the traditional 2xGA3 treatment,onfirming the efficacy of these treatments as berry growth pro-oters (Reynolds et al., 1992; Jensen et al., 1994; Wolf et al.,
splitting, bleaching and gray mold (Botrytis cinerea) incidence on berry cheek or atuby Seedless’) and 90 d (‘Redglobe’) at 0 ◦C plus 3 d at 20 ◦C
) Hairline + splitting (%) Bleaching (%) Gray mold incidence (%)
Berry base Berry cheek Total
4.0 ac 0.00ns 0.6 ac 0.9 ac 1.5 ac
14.0 b 0.00 1.6 b 1.5 b 3.1 b15.4 b 0.00 7.7 d 5.1 c 12.8 d14.0 b 0.00 4.0 c 2.8 b 6.8 c
0.00ns 2.5 a 2.3 a 1.2 a 3.5 a0.00 0.5 a 3.0 a 1.2 a 4.2 a0.00 7.7 b 3.8 a 5.1 b 8.9 b0.20 nd 15.2 b 3.8 b 19.0 b
0.00ns 2.4 ab 0.6 a 0.2 a 0.8 a0.00 1.7 a 0.2 a 0.5 a 0.7 a0.00 8.5 c 0.6 a 0.8 a 1.4 a7.10 4.1 bc 2.7 b 1.9 b 4.6 b
1.2 a 5.9ns 1.0 a 4.6 a 5.6 a3.7 b 11.0 4.6 ab 4.7 ab 9.3 ab3.2 b 6.9 7.1 b 13.4 c 20.5 c9.9 b 9.1 8.3 b 10.6 bc 18.9 bc
berellic acid (GA3) (Pro-Gibb 20% Valent Biosciences Chile S.A.); on ‘Redglobe’ and6 �L L−1 forchlorfenuron (N-(2-chloro-4-pyridyl)-N-phenylurea (Sitofex KT-30 EC,
in diameter. Durofel values were converted to Newtons (N = 9.8{exp[(Durofel
t according to Fisher’s least significant difference (LSD) (P ≤ 0.05). Percent valuesing, gray mold incidence) before analysis but, untransformed values are presented.

1 gy an
1arv(Pa
iC((iaTaa
ossssp
cem
o‘rpd
gHwabgcpio
FS(fSc(R
90 J.P. Zoffoli et al. / Postharvest Biolo
994; Retamales et al., 1995). However, over application of GA3nd CPPU application (8xGA3 + CPPU) retarded TSS accumulation,eaching the minimum concentration of TSS for commercial har-est 10 d later than the control, as has been previously documentedBen-Arie et al., 1997; Retamales et al., 1995; Wolf et al., 1994).ostharvest deterioration due to hairline, SO2 bleaching, shatter,nd decay was significantly affected by growth regulators.
Hairline incidence, as previously defined (Zoffoli et al., 2008),ncreased nearly three times when clusters were treated withPPU after traditional GA3 application or with over application8xGA3 + CPPU) or when the clusters were over-treated with GA38xGA3) after 60 d at 0 ◦C plus 3 d at 20 ◦C. At the same time, shatterncidence was severe on clusters over-treated with GA3 (8xGA3)lone or when combined with CPPU treatment (8xGA3 + CPPU).otal gray mold incidence, developed at the berry base (pedicel end)nd at the cheek of the berries, considerably increased (1.5–12.8%)s hairline incidence and shatter incidence increased.
As previously reported, gray mold mainly appeared at the baser on the cheek of the berries, but it was not observed at thetylar end of the berry (Holz et al., 2003). A significant relation-
hip between gray mold incidence at the base of the berry andhatter incidence was explained by an exponential curve (Fig. 4),uggesting that infection begins at insertion of the berry to theedicel. Therefore, practices that reduce the flexibility of the pedi-lT14
ig. 4. Relationship between shatter incidence and incidence of gray mold (Botrytis cineedless’, Santiago. Growth regulator treatments were (�) berries sprayed twice with 4N-(2-chloro-4-pyridyl)-N-phenylurea (CPPU); (�) in addition to the 2xGA3 spray applor 30 s on six occasions (8xGA3); (�) berries were treated with CPPU and 8xGA3 as deedless’ Rancagua, respectively (�) Control (0xGA3) without growth regulator, clustehloro-4-pyridyl)-N-phenylurea (CPPU); (�) berries were dipped in 20 �L L−1 GA3 for 31xGA3 + CPPU). YThompson Seedless = 0.4 + 0.31X + 0.019X2, R2 = 0.64, P = 0.0002. YRedglobe Santiago2 = 0.82, P < 0.0001, YRuby Seedless Rancagua = −0.27 + 0.81X − 0.048X2, R2 = 0.83, P < 0.0001.
d Technology 51 (2009) 183–192
el, as was suggested previously (Retamales and Cooper, 1993), maynhance berry shatter and consequently increase the risk of grayold.Pedicel thickness increased by over-treatment of GA3 (8xGA3)
r with CPPU (8xGA3 + CPPU) and increased shatter incidence inThompson Seedless’, ‘Redglobe’ and ‘Ruby Seedless’ (Fig. 5). Otheresearchers have found that a sharp angle of insertion between theedicel and berry has been associated with high susceptibility toecay incidence compared with an obtuse angle (Sarig et al., 1998).
In ‘Thompson Seedless’, gibberellic acid (8xGA3) increased totalray mold incidence (at the cheek and at the base of the berry).owever, gray mold incidence was diminished from 12.8 to 6.8%hen berries were also treated with CPPU (8xGA3 + CPPU). This
ppears to be associated with the highest cuticle content inducedy CPPU: 3.03 g m−2 compared with 2.58 g m−2. However, the lowray mold incidence obtained on 2xGA3-treated berries at similarontent of cuticle suggests that other effects, associated to mor-hological, anatomical or antifungal chemical constituents may be
nvolved (Mlikota Gabler et al., 2003). A similar effect was alsobtained when berries of ‘Redglobe’ from Santiago or ‘Ruby Seed-
ess’ from Rancagua treated with 1xGA3 and CPPU were compared.he incidence of gray mold at the cheek of the berry was 5.1% withxGA3 compared with 1.2% on CPPU treatment and 13.4% instead of.7% on ‘Ruby Seedless’ berries, although these table grape cultivarserea) developed at the berry base on Vitis vinifera table grape cv. (A) ‘Thompson0 �L L−1 gibberellic acid. (2xGA3); (©) berries dipped in 6 �L L−1 forchlorfenuronications, berries were over-treated with GA3 by dipping them in 20 �L L−1 GA3
escribed above (8xGA3 + CPPU) (B–D). ‘Redglobe’ Santiago, Rancagua and ‘Rubyrs were dipped in water; (©) berries dipped in 6 �L L−1 forchlorfenuron (N-(2-0 s (1xGA3); (�) berries were treated with CPPU and 1xGA3 as described above= 2.05 + 1.4X − 0.03X2, R2 = 0.83, P < 0.0001, YRedglobe Rancagua = 0.55 − 0.18X + 0.066X2,

J.P. Zoffoli et al. / Postharvest Biology and Technology 51 (2009) 183–192 191
Fig. 5. Relationship between pedicel thickness induced by growth regulator treatments and shatter incidence after 60 d (‘Thompson Seedless’ and ‘Ruby Seedless’) and90 d (‘Redglobe’) storage at 0 ◦C plus 3 d at 20 ◦C. Growth regulator treatments on ‘Thompson Seedless’ were (�) berries sprayed twice with 40 �L L−1 gibberellic acid.(Control, 2xGA3). In addition to the 2xGA3 spray applications, berries were (©) dipped in 6 �L L−1 forchlorfenuron (N-(2-chloro-4-pyridyl)-N-phenylurea (CPPU); (�) over-treated with GA3 by dipping them in 20 �L L−1 GA3 for 30 s on six occasions (8xGA3); (�) berries were treated with CPPU and 8xGA3 as described above (8xGA3 + CPPU)( ) Cond �) berC − 3.5Y + 4.02
aC‘rwggoa
d‘dihBt
elgcIGtd
ments on ‘Thompson Seedless’ and ‘Ruby Seedless’ table grapes, butno hairline occurred on ‘Redglobe’ table grapes. Therefore, berryquality during storage is highly dependent on growth regulatormanagement in the vineyard. These results do not ignore the fact
B–D). ‘Redglobe’ Santiago, Rancagua and ‘Ruby Seedless’ Rancagua, respectively; (�
ipped in 6 �L L−1 forchlorfenuron (N-(2-chloro-4-pyridyl)-N-phenylurea (CPPU); (PPU and 1xGA3 as described above (1xGA3 + CPPU). YThompson Seedless = −22.0 + 21.3X
Redglobe Rancagua = 8.9 − 10.3X + 2.9X2, R2 = 0.84, P < 0.0001, YRuby Seedless Rancagua = −1.98
re physiologically and phenotypically very distant (Tables 1 and 2).PPU has also induced resistant to Rhizopus stolonifer infection on
Redglobe’ berries (Ben-Arie et al., 1997). A negative and significantelation between cuticle content and cheek gray mold incidenceas obtained on ‘Thompson Seedless’ and ‘Ruby seedless’ table
rape cultivars (Fig. 6). As previously reported, ‘Ruby Seedless’ tablerape was one of the most sensitive cultivar to gray mold devel-pment (Mlikota Gabler et al., 2003). In this experiment, this wasssociated with the lowest cuticle content.
On the basis of the results obtained, CPPU increased hairlineevelopment on elongated berries such as those produced by
Thompson Seedless’ and ‘Ruby Seedless’ cultivars. This may beue to reduced berry elasticity resulting from high cuticle content
nduced by CPPU-treated berries and the periclinar type of cell thatas been recognized to enhance cracking disorder (Considine andrown, 1981). However, further research is needed to demonstratehis phenomenon.
In conclusion, the results obtained in this study are compellingvidence that the overuse of GA3 increased pedicel thickness butowered cuticle content, induced shatter and predisposed grapes toray mold. In contrast, CPPU increased berry pedicel thickness and
uticle content but did not increase gray mold or shatter incidence.t was interesting that clusters that were subjected to overuse ofA3 and CPPU treatments were highly sensitivity to shatter, hadhe thickest pedicel, and developed a high gray mold incidenceuring storage. Hairline was induced by all growth regulator treat-
FcsoP
trol (0xGA3) without growth regulator, clusters were dipped in water; (©) berriesries were dipped in 20 �L L−1 GA3 for 30 s (1xGA3); (�) berries were treated withX2, R2 = 0.66, P < 0.0001. YRedglobe Santiago = 44.6 − 43.1X + 10.3X2, R2 = 0.88, P < 0.0001,X, R2 = 0.4, P = 0.0028.
ig. 6. Relationship between cuticle content at harvest and gray mold (Botrytisinerea) incidence developed on the cheek of the berry on table grapes cvs. ‘Thomp-on Seedless’ and ‘Ruby Seedless’ after 60 d storage at 0 ◦C plus 3 d at 20 ◦C. Meanf five replications each of one box. YThompson Seedless = 51.3 − 30.8X + 4.8X2, R2 = 0.34,= 0.03, YRuby Seedless = 27.4 − 10.6X, R2 = 0.42, P = 0.002.

1 gy an
tei
A
I1
R
B
B
B
B
C
C
C
D
D
D
E
E
E
F
G
G
H
H
J
J
K
K
L
L
L
MM
M
M
N
N
P
P
P
P
R
R
R
S
S
T
V
VPrincess table grapes. Calif. Agric. 59, 103–108.
Wolf, E.E.H., Viljoen, J.A., Nieuwenhuys, A., Loubser, J.T., 1994. The effect of forchlor-
92 J.P. Zoffoli et al. / Postharvest Biolo
hat other cultural practices that promote pedicel thickness, thatnhance shatter incidence, and lower cuticle content may also benvolved.
cknowledgement
The research was funded by Conicyt (Comisión Nacional denvestigación Científica y Tecnológica (Chile), Fondecyt grant no.050798.
eferences
en-Arie, R., Sarig, P., Cohen-Ahbut, Y., Zutkhi, Y., Sonego, L., Kapulonov, T., Lisker,N., 1997. CPPU and GA3 effects on pre- and post-harvest quality of seedless andseeded grapes. Acta Hortic. 463, 349–355.
en-Tal, Y., 1990. Effect of gibberellin treatments on ripening and berry drop from‘Thompson Seedless’ grapes. Am. J. Enol. Viticult. 41, 142–146.
ianchi, M., Jensen, F., Moriyama, M., 1991. Management of berry size and color in‘Flame Seedless’. In: Proceedings of the Coachella Valley Table Grape Seminar,Coachella, CA.
laich, R., Stein, U., Wind, R., 1984. Perforation in der cuticula von weinbeeren alsmorphologischer faktor der Botrytisresistenz. Vitis 23, 242–256.
hristensen, L.P., Boggero, J.D., 1985. A study of mineral nutrition relationships ofwaterberry in ‘Thompson Seedless’. Am. J. Enol. Viticult. 36, 57–64.
ollins, C., Rawnsley, B., 2005. Factors influencing primary bud necrosis (PBN) inAustralian vineyards. Acta Hortic. 689, 81–86.
onsidine, J., Brown, K., 1981. Physical aspect of fruit growth: theoretical analysis ofdistribution of surface growth forces in fruits in relation to cracking and splitting.Plant Physiol. 68, 371–376.
okoozlian, N.K., Hirchfelt, D.J., 1985. The influence of cluster thinning at variousstages of fruit development on ‘Flame Seedless’ table grapes. Am. J. Enol. Viticult.46, 429–436.
okoozlian, N.K., Luvisi, D.A., Schrader, P.L., Kosareff, J.J., 1994a. Influence of vine cropload on ‘Redglobe’ table grapes. In: International Symposium on Table GrapeProduction, Anaheim, CA. Am. Soc. Enol. Viticul, pp. 221–214.
okoozlian, N.K., Luvisi, D.A., Schrader, P.L., Moriyama, M.M., 1994b. Influence oftrunk girdle timing and ethephon on the quality of Crimpson Seedless tablegrapes. In: International Symposium on Table Grape Production, Anaheim, CA.Am. Soc. Enol. Viticul, pp. 237–240.
nglish, J.T., Thomas, C.S., Marois, J.J., Gluber, W.D., 1989. Microclimates of grapevinecanopies associated with leaf removal and control of Botrytis bunch rot. Phy-topathology 79, 395–401.
PA, 2003. Registration Forchlorfenuron; Notice of Filing a Pesticide Peti-tion to Establish a Tolerance for a Certain Pesticide Chemical in or onFood. USA Environmental Protection Agency. http://www.epa.gov/fedrgstr/EPA-PEST/2003/May/Day-16/p12360.htm (Accessed: December 2007).
sterio, M., Auger, J., Drogguett, A., Arroyo, A., 1996. Effectiveness of biological inte-grated and traditional control programs of Botrytis cinerea in table grape in theCentral Valley of Chile. In: XIth International Botrytis Simposium, Wageningen,The Netherlands, p. 73.
ranck, J., Latorre, B.A., Torres, R., Zoffoli, J.P., 2005. The effect of preharvest fungicideand postharvest sulfur dioxide use on postharvest decay of table grapes causedby Penicillium expansum. Postharvest Biol. Technol. 37, 20–30.
ao, H., Hu, X., Liu, L., Zhang, H., Wang, S., 2003. Study on sensitivity of table grapesto SO2. Acta Hortic. 628, 541–545.
ubler, W.D., Marois, J.J., Bledsoe, A.M., Bettiga, L.J., 1987. Control of Botrytis bunchrot of grape with canopy management. Plant Dis. 71, 599–601.
olloway, P.J., Baker, E.A., 1968. Isolation of plant cuticles with zinc chloride-hydrochloric acid solution. Plant Physiol. 43, 1878–1879.
olz, G., Gutschow, M., Coertze, S., Calitz, F., 2003. Occurrence of Botrytis cinerea andsubsequent disease expression at different positions on leaves and bunches ofgrape. Plant Dis. 87, 351–358.
awanda, J.S., Singh, R., Pal, R.N., 1974. Effect of growth regulators on floral bud drop,fruit characteristics and quality of ‘Thompson Seedless’ grapes (Vitis vinifera L.).Vitis 13, 215–221.
Z
d Technology 51 (2009) 183–192
ensen, F.L., Bianchi, M., Moriyama, M., Kazarian, D., 1994. Influence of applicationtiming of gibberellin berry-enlarging treatments on the maturity of ‘ThompsonSeedless’ table grapes. In: International Symposium on Table Grape Production,Anaheim, CA. Am. Soc. Enol. Viticul, pp. 47–49.
ington, C.M., Van Epenhuijsen, C.W., 1989. Influence of leaf area on fruit devel-opment and quality of Italia glasshouse table grape. Am. J. Enol. Viticult. 40,130–134.
liewer, W.M., Weaver, R.J., 1971. Effect of crop level and leaf area on growth, com-position and coloration of Tokay grapes. Am. J. Enol. Viticult. 22, 172–177.
atorre, B.A., 1986. Manejo de Botrytis cinerea en uva de mesa. Revista Fruticola(Chile) 7, 75–88.
atorre, B.A., Lillo, C., Rioja, M.E., 2001. Eficacia de los tratamientos fungicidas parael control de Botrytis cinerea de la vid en función de la época de aplicación. Cien.Inv. Agr. 28, 61–66.
uvisi, D.A., Shorey, H., Smilanick, J., Thompson, J., Gump, B.H., Knutson, J., 1992.Sulfur dioxide fumigation of table grapes. Univ. Calif. Div. Agric. Nat. Resour.,Bull., 1932.
ariette, G., 2007. Guidelines for Preparing Export Table Grapes. Capespan, 77 pp.arois, J., Bledsoe, A., Bettiga, L., 1992. Bunch rots. In: Grape pest management, 2nd
ed. Univ. Calif. Div. Agric. Nat. Resour. Publ. 3343.arois, J.J., Nelson, J.K., Morrison, J.C., Lile, L.S., Bledsoe, A.M., 1986. The influence of
berry contact within grape clusters on the development of Botrytis cinerea andepicuticular wax. Am. J. Enol. Viticult. 37, 293–296.
likota Gabler, F., Smilanick, J.L., Mansour, M., Ramming, D.W., Mackey, B.E., 2003.Correlations of morphological, anatomical, and chemical features of grapeberries with resistance to Botrytis cinerea. Phytopathology 93, 1263–1273.
avarro, M., Retamales, J., Defilippi, B., 2001. Efecto del arreglo de racimo y aplicaciónde citoquinina sintética (CPPU) en la calidad de uva de mesa variedad sultaninatratada con dos fuentes de giberelinas. Agric. Tec. (Chile) 61, 15–25.
elson, K.E., 1985. Harvesting and Handling California Table Grapes for Market. Uni-versity of California, Division of Agriculture Science, Oakland, CA, USA, Pub. 1913,67 pp.
ercival, D.C., Sullivan, J.A., Fisher, K.H., 1993. Effect of cluster exposure, berry con-tact and cultivar on cuticular membrane formation and occurrence of bunch rot(Botrytis cinerea PERS. FR.) with 3 Vitis vinifera L. cultivars. Vitis 32, 87–97.
eschel, S., Knoche, M., 2005. Characterization of microcracks in the cuticle of devel-oping sweet cherry fruit. J. Am. Soc. Hortic. Sci. 103, 487–495.
erez, H., 1994. Table grape production in Chile. In: International Symposium onTable Grape Production, Anaheim, CA. Am. Soc. Enol. Viticul, pp. 1–7.
olenta, G., Buddle, C., Murray, R., 2005. Effect of different pre-storage anoxictreatments on ethanol and acetaldehyde content in peaches. Postharvest Biol.Technol. 38, 247–253.
etamales, J., Bangerth, F., Cooper, T., Callejas, R., 1995. Effect of CPPU and GA3 onfruit quality of Sultanina table grape. Acta Hortic. 394, 149–157.
etamales, J., Cooper, T., 1993. Berry drop and fruit removal forces as related withGA3 applications in table grapes. Acta Hortic. 329, 81–83.
eynolds, A.G., Wardle, D.A., Zurowski, C., Looney, N.E., 1992. Phenylureas CPPU andThidiazuron affect yield components, fruit composition, and storage potentialof four seedless grape selections. J. Am. Soc. Hortic. Sci. 117, 85–89.
arig, P., Zutkhi, Y., Lisker, N., Shkelerman, Y., Ben-Arie, R., 1998. Natural and inducedresistance of table grape to bunch rots. Acta Hortic. 464, 65–70.
harples, G.C., Hilgerman, R.H., Milne, L., 1955. The relation of cluster thinning andtrunk girdling of Cardinal grapes to yield and quality of fruit in Arizona. Proc.Am. Soc. Hortic. Sci. 65, 225–233.
homas, C.S., Marois, J.J., English, J.T., 1988. The effects of wind speed, temperature,and relative humidity on development of aerial mycelium and conidia of Botrytiscinerea on grape. Phytopathology 78, 260–265.
ail, M.E., Marois, J.J., 1991. Grape cluster architecture and the susceptibility ofberries to Botrytis cinerea. Phytopathology 81, 188–191.
ial, P., Crisosto, C., Crisosto, G., 2005. Early harvest delays berry skin browning of
fenuron on bunch quality in table grapes. In: International Symposium on TableGrape Production, Anaheim, CA. Am. Soc. Enol. Viticul, pp. 50–53.
offoli, J.P., Latorre, B.A., Naranjo, P., 2008. Hairline, a postharvest cracking disorderin table grapes induced by sulfur dioxide. Postharvest Biol. Technol. 47, 90–97.




















