
Precision of Thermoregulation and Its Relation to Environmental Factors in the Desert Iguana,...
19
Precision of Thermoregulation and Its Relation to Environmental Factors in the Desert Iguana, Dipsosaurus dorsalis Author(s): Calvin B. DeWitt Source: Physiological Zoology, Vol. 40, No. 1 (Jan., 1967), pp. 49-66 Published by: The University of Chicago Press Stable URL: http://www.jstor.org/stable/30152438 Accessed: 21/12/2009 13:15 Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at http://www.jstor.org/action/showPublisher?publisherCode=ucpress. Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission. JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. The University of Chicago Press is collaborating with JSTOR to digitize, preserve and extend access to Physiological Zoology. http://www.jstor.org
Transcript of Precision of Thermoregulation and Its Relation to Environmental Factors in the Desert Iguana,...

Precision of Thermoregulation and Its Relation to Environmental Factors in the DesertIguana, Dipsosaurus dorsalisAuthor(s): Calvin B. DeWittSource: Physiological Zoology, Vol. 40, No. 1 (Jan., 1967), pp. 49-66Published by: The University of Chicago PressStable URL: http://www.jstor.org/stable/30152438Accessed: 21/12/2009 13:15
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available athttp://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unlessyou have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and youmay use content in the JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained athttp://www.jstor.org/action/showPublisher?publisherCode=ucpress.
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
The University of Chicago Press is collaborating with JSTOR to digitize, preserve and extend access toPhysiological Zoology.
http://www.jstor.org

PRECISION OF THERMOREGULATION AND ITS RELATION TO
ENVIRONMENTAL FACTORS IN THE DESERT IGUANA, DIPSOSAURUS DORSALIS1
CALVIN B. DEWITT2
Department of Zoblogy, University of Michigan, Ann Arbor
INTRODUCTION
T HE control of body temperature in lizards is largely dependent upon their appropriate exploitation of
their thermal environment. This behav- ioral process has been documented in a number of papers including those of Mosauer (1936), Cowles and Bogert (1944), and Saint-Girons and Saint- Girons (1956). This paper is a contribu- tion to a further understanding of be- havioral thermoregulation through an analysis of the precision of regulation and the effects of various environmental factors upon this precision. For this ini- tial study, a lizard which normally ex- periences high environmental tempera- tures and intense solar radiation, the
1I am indebted to Dr. W. R. Dawson for his guidance and suggestions during these investiga- tions. I am also grateful for the assistance given by Professor G. C. Gill, Dr. H. B. Tordoff, and Dr. W. H. Wagner, Jr. Drs. F. H. Test, N. Hartweg, D. J. Klingener, R. C. Lasiewski, P. Licht, V. Shoemaker, and Mr. W. Moberly also have assisted in many ways. Special thanks are due the fine group at the Philip L. Boyd Desert Research Center, Uni- versity of California, Riverside, particularly Dr. W. W. Mayhew, Mr. P. L. Boyd, and Mr. Lloyd Tevis, for very generously furnishing research and living facilities. I am also very grateful to my wife, Ruth Ann DeWitt, for her assistance in recording and analyzing data at the desert study site. This re- search was supported by graduate fellowships 20490, 21365, and 32231 from the National Science Founda- tion. This paper is part of a dissertation submitted in partial fulfilment of the requirements for the de- gree of Doctor of Philosophy at the University of Michigan.
2 Present address: Section of Mathematics and Sciences, University of Michigan, Dearborn.
desert iguana (Dipsosaurus dorsalis Baird and Girard), has been selected. The desert iguana is also suitable for study because many aspects of its biology have already been investigated including its ecology (Norris, 1953), social behavior (Carpenter, 1962), metabolic and cardiac response to temperature at rest (Dawson and Bartholomew, 1958), cardiac re- sponse to temperature during activity (Licht, 1965), evaporative water loss (Templeton, 1960), physiology during hibernation (Moberly, 1963), and repro- ductive physiology (Mayhew, unpub- lished). The nature and effectiveness of various behavioral thermoregulatory mechanisms of this species and the re- lation of rate of heat transfer to wind speed and body size have also been ana- lyzed (DeWitt, 1963).
MATERIAL AND METHODS
Behavioral thermoregulation of the desert iguana was studied from 1960 to 1962 under laboratory conditions at the University of Michigan and under field and laboratory conditions at the Deep Canyon Mobile Desert Laboratory lo- cated in Palm Desert, Riverside County, California. Studies at the Mobile Labora-
tory which is maintained by the Univer- sity of California at Riverside, were con- ducted in the summer of 1961. Desert
iguanas used in all laboratory studies were collected in the vicinity of Palm Springs and Palm Desert, Riverside
49
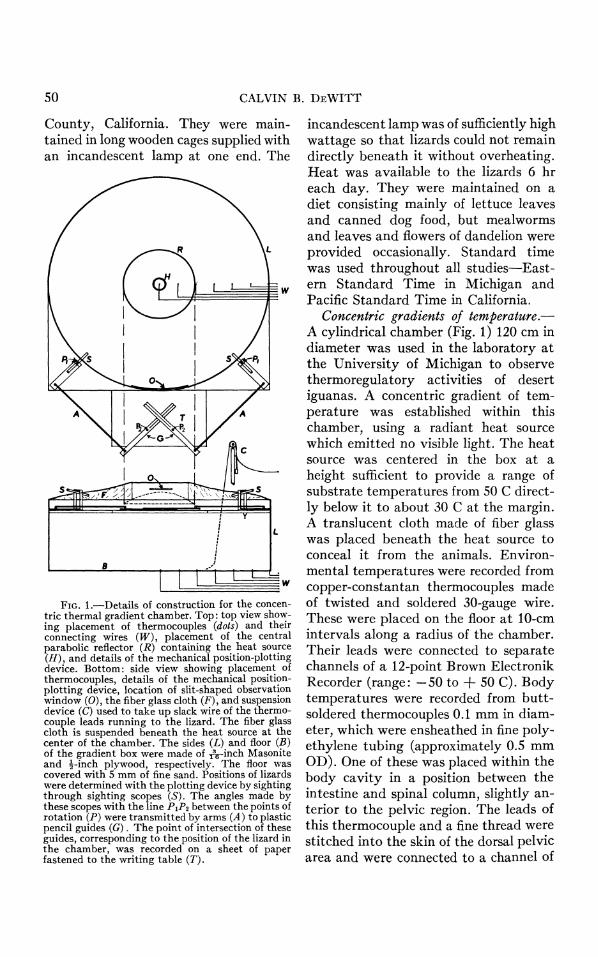
50 CALVIN B. DEWITT
County, California. They were main- tained in long wooden cages supplied with an incandescent lamp at one end. The
_
_---:-
w
o I
A oA
BW
,?-- I 1 sw , - . ,
; i
, t.
FIG. 1.-Details of construction for the concen- tric thermal gradient chamber. Top: top view show- ing placement of thermocouples (dots) and their connecting wires (W), placement of the central parabolic reflector (R) containing the heat source (H), and details of the mechanical position-plotting device. Bottom: side view showing placement of thermocouples, details of the mechanical position- plotting device, location of slit-shaped observation window (0), the fiber glass cloth (F), and suspension device (C) used to take up slack wire of the thermo- couple leads running to the lizard. The fiber glass cloth is suspended beneath the heat source at the center of the chamber. The sides (L) and floor (B) of the gradient box were made of A-inch Masonite and -inch plywood, respectively. The floor was covered with 5 mm of fine sand. Positions of lizards were determined with the plotting device by sighting through sighting scopes (S). The angles made by these scopes with the line PiP2 between the points of rotation (P) were transmitted by arms (A) to plastic pencil guides (G). The point of intersection of these guides, corresponding to the position of the lizard in the chamber, was recorded on a sheet of paper fastened to the writing table (T).
incandescent lamp was of sufficiently high wattage so that lizards could not remain directly beneath it without overheating. Heat was available to the lizards 6 hr each day. They were maintained on a diet consisting mainly of lettuce leaves and canned dog food, but mealworms and leaves and flowers of dandelion were provided occasionally. Standard time was used throughout all studies-East- ern Standard Time in Michigan and Pacific Standard Time in California.
Concentric gradients of temperature.- A cylindrical chamber (Fig. 1) 120 cm in diameter was used in the laboratory at the University of Michigan to observe thermoregulatory activities of desert iguanas. A concentric gradient of tem- perature was established within this chamber, using a radiant heat source which emitted no visible light. The heat source was centered in the box at a height sufficient to provide a range of substrate temperatures from 50 C direct- ly below it to about 30 C at the margin. A translucent cloth made of fiber glass was placed beneath the heat source to conceal it from the animals. Environ- mental temperatures were recorded from copper-constantan thermocouples made of twisted and soldered 30-gauge wire. These were placed on the floor at 10-cm intervals along a radius of the chamber. Their leads were connected to separate channels of a 12-point Brown Electronik Recorder (range: -50 to + 50 C). Body temperatures were recorded from butt- soldered thermocouples 0.1 mm in diam- eter, which were ensheathed in fine poly- ethylene tubing (approximately 0.5 mm OD). One of these was placed within the body cavity in a position between the intestine and spinal column, slightly an- terior to the pelvic region. The leads of this thermocouple and a fine thread were stitched into the skin of the dorsal pelvic area and were connected to a channel of

PRECISION OF THERMOREGULATION IN DESERT IGUANA 51
the Brown Recorder through a device which took up slack wire inside the chamber (Fig. 1). The thread was cut slightly shorter than the wires to elimi- nate tension on the leads themselves. The temperature of each thermocouple was recorded every 3 min. Position and orientation of lizards were recorded using a mechanical plotting device (Fig. 1).
Eleven lizards were used in this series of experiments. They were maintained on a 6-hr thermal period both during measurement and for at least 3 months
prior to measurement. During the ther- mal period, general illumination was sup- plied to the chamber by four 40-w fluo- rescent tubes and to the wooden cages by incandescent lamps. Outside of the thermal period, the cages were often dimly illuminated by room light. Each lizard was studied individually in the concentric gradient on 3 consecutive days. Records of body temperature for each day were placed in class intervals of 0.5 C. For a particular day, the num- ber of records in each class interval was expressed as a percentage of the total number of records analyzed. The per- centages in each class interval were then averaged for all days in which records covered a period of at least 90 min. These final averages for each class interval were then plotted to show the frequency dis- tribution of temperatures maintained by the group of 11 lizards. The data were also analyzed so that they would justify the assumption of randomness in statis- tical tests. Selection of data employed the conservative method of using from each day's records only two randomly drawn samples separated from each other by at least 30 min.
At the Mobile Laboratory in Palm Desert, thermal preferences of 18 lizards adjusted to mid-August field conditions were determined using a simplified con- centric gradient. The cylindrical chamber
containing this gradient was constructed of corrugated cardboard, and its dimen- sions were the same as those of the chamber containing the concentric gra- dient used in the laboratory at the Uni- versity of Michigan. The chamber was fitted with a 250-w lamp which emitted radiation in both the visible and infrared wavelengths. The lamp was placed over the center of the chamber in view of the lizards. The height of the lamp was ad- justed to produce substrate temperatures ranging from about 50 C at the center of the chamber to about 30 C at its margins. During experiments, lizards in the chamber were quickly noosed and the body temperature measured with a Schultheis mercury-in-glass thermome- ter (0-50 C graduated in 0.2 C) de- scribed by Bogert (1949).
Linear gradients of temperature.-Body temperatures of desert iguanas were further studied in Michigan in a series of linear gradients of temperature main- tained by heat sources within the sub- strate. Originally these experiments were designed to permit additional compari- son of the respective thermal preferences of lizards adjusted to field and laboratory conditions. Although it turned out that these comparisons could not be made be- cause of certain inadequacies of the thermal gradients, the data were useful in illustrating maximum differences in temperature between the head and the trunk of lizards.
The linear gradients were contained in rectangular boxes of corrugated card- board 120 cm long by 13 cm wide by 40 cm high. The heat source, situated at one end of each box, consisted of a con- tainer 35 cm long by 13 cm wide by 1.2 cm high made of copper screen of ap- proximately 1-mm mesh. This container was filled with fine sand through which a heating element of nichrome wire passed in a zigzag pattern, with the

52 CALVIN B. DEWITT
strands placed progressively farther apart toward the cooler end of the gradient. This heat source produced a very sharp horizontal gradient of temperature from about 70 to 25 C in approximately 35 cm. The cool end of the gradient was sup- plied with a cardboard shelter 20 cm long by 13 cm wide by 10 cm high, with an entrance 3 cm in diameter facing the heat source. Position of lizards, relative to marks at regular 10-cm intervals along the length of the rectangular box, and the body temperature were recorded every 5 min throughout the thermal period. Body temperature was recorded using a 48-point Brown Electronik Re- corder (0-150 F) connected to iron-con- stantan thermocouples implanted in the body cavity as described previously. The leads from these thermocouples were sus- pended from a point 1.2 m above each gradient.
Three experiments were conducted with this series of gradients. In the first experiment, desert iguanas adjusted to conditions of the Michigan laboratory were placed singly in each of six gradient boxes for one day. They were given the 6-hr thermal period under which they had been maintained for 6 months in the laboratory. Essentially continuous records of body temperature were taken simultaneously from the six lizards.
The second experiment employed simi- lar techniques using lizards caught at Palm Springs, California, on April 25, 1961, and shipped to the laboratory in Michigan. Simultaneous records of body temperature for the 16 lizards were re- corded on April 28 and 29, 1961. These animals, which were adjusted to field conditions, were maintained on a ther- mal period of 12 hr.
The third experiment was done on August 1 and 2, 1961, in Palm Desert, California, using eight desert iguanas maintained on a daily thermal period of
6 hr for the 2 days. In both the second and third experiments, the animals were placed on the heat source if they did not find it by themselves. All records were analyzed from the end of the initial warming period to the time at which the lizard retreated from the heat source and did not return.
Implantation of thermocouples in the head.-Simultaneous measurements of both the head and body temperatures were carried out on all lizards studied in the series of thermal gradients in Palm Desert, and also on a few individuals in the field. Head temperatures were meas- ured with butt-welded iron-constantan thermocouples 0.1 mm in diameter. The junction was insulated with Formvar. The cut end of the iron wire and the head end of a fine insect pin were filed until smooth and perpendicular to their main axes and then butt-welded. The pin was pushed into the side of the head, passing between the jugal and the tym- panic crest, dorsal to the pterygoid and ventral to the postorbital bones. The pin and wire were drawn through the head until the thermocouple was in the mid- line of the body. This placed it in the optic chiasma immediately ventral to the hypothalamus. The leads then were run to the pelvic area and stitched to the skin. The pin was cut off, and the ther- mocouple leads were soldered to con- necting leads running to the recorder.
Methods employed under field condi- tions.-A site in Palm Desert was select- ed for the study of desert iguanas under natural conditions because of its unusu- ally dense population of this species and because of their great calmness in the presence of persons in this area. This un- usual calmness made it possible to ap- proach carefully and noose each lizard without prior chasing. Immediately after the lizards were noosed, their body tem- peratures were registered on a Schultheis
PRECISION OF THERMOREGULATION IN DESERT IGUANA 53
thermometer. The thermometer bulb was inserted through the cloaca into the in- testine to a depth of 1-2 cm, and body temperature was read to the nearest 0.1 C by interpolation between the 0.2 C scale divisions of the thermometer. Con- tinuous recordings of both freely moving and tethered lizards were made on a few occasions using iron-constantan thermo- couples and the 48-point Brown Electron- ik Recorder. Records of various envi- ronmental temperatures at and near the point of capture were made both with mercury-in-glass thermometers shielded from direct solar radiation and with the thermocouples and Brown Recorder. A more extensive presentation of micro- climatological data from the Palm Desert study site is presented elsewhere (DeWitt, 1963).
Observations on feeding, courting, and territoriality and the effects of these ac- tivities on thermoregulation were made from the Mobile Laboratory which was located at the Palm Desert study site.
RESULTS
The level and precision of thermoregu- lation were found to be influenced by both physical and biological parameters of the environment. When observations from the laboratory and the field were compared, it soon became apparent that the distribution of environmental tem- perature was the most prominent physi- cal factor influencing this level and pre- cision. Fights between individuals and response to possible predators appeared to be the most prominent biological fac- tors influencing the level and precision of thermoregulation.
THE LEVEL AND PRECISION OF THERMO-
REGULATION IN EXPERIMENTAL
TEMPERATURE GRADIENTS
Desert iguanas in the concentric gradient of environmental temperature
maintained body temperatures near 38.5 C. They accomplished this by ap- propriate adjustment of their position, either by changing location in the gra- dient, posture, or orientation to the heat source. When body temperatures dropped to some temperature below 38.5 C, liz- ards usually moved to a position where they could become warmer. On the other
48 46 44 42
34 38- 36
0 n 32
bi 30
o 42 40-
( 38-
I1 38-
02 345
T 32- I- 30 i , I I .
o 46
44
42 -
38-
36- /
I :
30
0 I 2 3 4 5 6
TIME IN HOURS
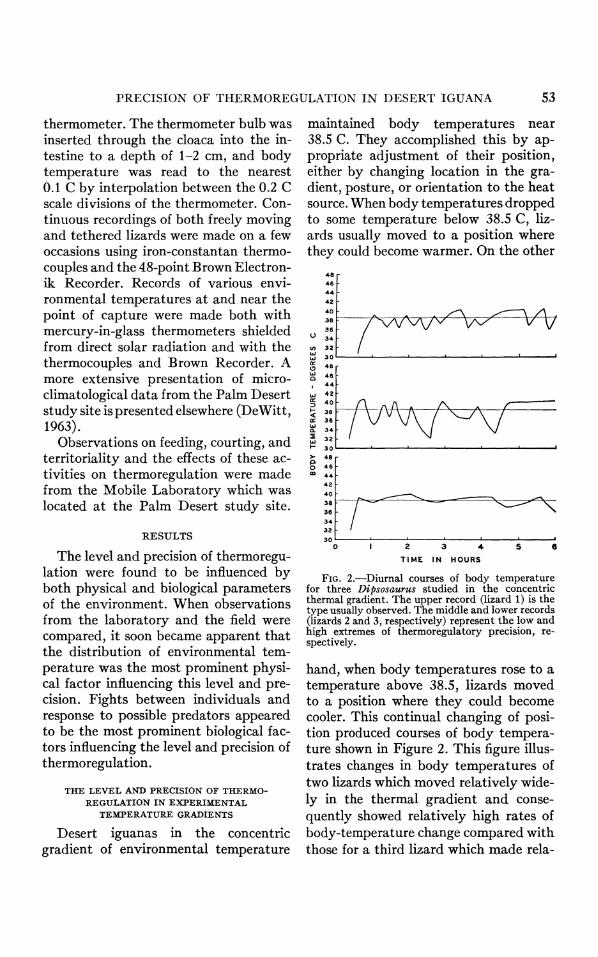
FIG. 2.-Diurnal courses of body temperature for three Dipsosaurus studied in the concentric thermal gradient. The upper record (lizard 1) is the type usually observed. The middle and lower records (lizards 2 and 3, respectively) represent the low and high extremes of thermoregulatory precision, re- spectively.
hand, when body temperatures rose to a temperature above 38.5, lizards moved to a position where they could become cooler. This continual changing of posi- tion produced courses of body tempera- ture shown in Figure 2. This figure illus- trates changes in body temperatures of two lizards which moved relatively wide- ly in the thermal gradient and conse- quently showed relatively high rates of body-temperature change compared with those for a third lizard which made rela-

54 CALVIN B. DEWITT
tively small changes in position in the gradient and consequently showed rela- tively low rates of body-temperature change. In this last animal, the small changes in position were sometimes only a few millimeters.
S50 i D
I. % II
5 40
.i.. i i i c . .
I
" I i o La 30
o - I w A "
" 0 2 3 3 38 40 424
U D I DE GRE I
PC20
I-I
I-..
I. 0
z
LaJ
30 32 34 36 38 40 42 44
MID-CLASS OF TEMPERATURE INTERVAL DEGREES C
FIG. 3.-Frequency distribution of body tem- peratures for three different Dipsosaurus showing variation among them. The central distribution (lizard 3) is representative of the level of thermo- regulation most frequently observed. The distribu- tions on the left (lizard 4) and right (lizard 5) show the low and high extremes, respectively. Each distri- bution polygon is for a single day. Class intervals of 1 C.
The precision of thermoregulation most commonly observed is represented in Figure 2 by the record of lizard 1, with the records of lizards 2 and 3 repre- senting the lower and upper extremes, respectively. Although the precision var- ied from one lizard to another, it usually was similar in the same individual on successive days.
Most lizards maintained body tem- peratures at the 38.5 C level (Fig. 3, lizard 3), but some regulated at other levels (Fig. 3, lizards 4 and 5). The level of thermoregulation for a particular in- dividual generally was similar on suc- cessive days (Fig. 4, days 1 and 2), but this level was abandoned when the lizard occasionally remained in or near the shelter (Fig. 4, day 3).
The over-all precision and level of thermoregulation in the group of 11
o 50 .I
D II U
ar 40
I I L&.
=30 I
0 .I
U
O I I "30 ii
r,-
o I
"'. I
w i I
0.
o I
Z 0
z IO\I wI i
30 32 34 36 38 40 42 44
MID*CLASS OF TEMPERATURE INTERVAL DEGREES C
FIG. 4.-Frequency distribution of body tem- peratures on 3 consecutive days for an individual Dipsosaurus (lizard 3) showing day-to-day variation. Distributions recorded on the first, second, and third days are indicated by the solid, regularly dotted, and broken lines, respectively. Class intervals of 1 C.
lizards were essentially constant on suc- cessive days (Fig. 5). The median of the distributions, which is taken as the pre- ferred level of body temperature, re- mained the same from day to day at

PRECISION OF THERMOREGULATION IN DESERT IGUANA 55
about 38.5 C. However, there was a tendency for regulation to be somewhat less precise on the first day.
The distribution of body temperatures for all individuals on all 3 days is nega- tively skewed (Fig. 6, distribution A). This skewness remains even when body
tively, as determined from a replotting of these data in a cumulative frequency distribution. The distribution of the 57 randomly selected body temperatures, which remains after records of lizards on the initial warming curve and records of lizards on the final cooling curve are
24
O~ is
.0 I
U)~ = -
I
V"A
I: ". 1
U . 16 -DAY I- - --
rr
"
S2DAY 2 : I
: t'
S1I
0 I A
* U)
-11 DAVI5 36
...
8
DL.T
I- L
o 1
rC
I-. I.
z I".
" /
31 38 33 34 35 36 37 38 39 40 41 42 43 44
MID-CLASS OF TEMPERATURE: INTERVAL - DEGREES C
FIG. 5.-Frequency distribution of body temperatures of 11 Dipsosaurus observed on each of 3 consecu- tive days in the concentric thermal gradient. All records were taken between completion of the initial period of warming and the time at which individuals went into shelter at the end of their activity periods and did not return. Class intervals of 1 C.
temperatures recorded after a lizard makes its final retreat to shelter are omitted (Fig. 6, distribution B). Distri- bution B of Figure 6, which I think characterizes control of body tempera- ture in desert iguanas, has a median of 38.5 C, with the central 50, 68, and 95%- of the distribution falling between 37.0- 39.5, 36.1-40.1, and 33.2-41.8 C, respec-
omitted, is negatively skewed (P < .01) with gl equal to -0.904 (see Snedecor, 1946, p. 174, for methods of calculating gi).
Differences in temperatures of head and body.-Body temperatures of desert igua- nas adjusted to conditions in the labora- tory or to conditions of the desert in spring and summer were measured in

56 CALVIN B. DEWITT
the rectangular gradient boxes, with the original intention of determining whether any differences in temperature preferen- da existed between these groups. How- ever, the gradients used were quite un- natural, since the source of heat was ex- clusively from the substrate. This was
six lizards adjusted to laboratory condi- tions and 12 lizards adjusted to natural conditions during spring. When the data were evaluated in light of concurrent be- havioral observations, it was possible to separate the frequency distribution into two components: a lower one associated
92 ..
(f A 0
-v..
0
n-j
I.-
I-- .
U.- _
0 :
u,
9-.
29 30 39 32 33 34 35 36 37 38 39 40 49 42 43 44 45
MID-CLASS OF TEMPERATURE INTERVAL - DEGREES C
FIG. 6.-Frequency distribution of body temperatures of 11 Dipsosaurus during occupancy of a con- centric thermal gradient on 3 consecutive days. Curve A represents all records taken after the initial period of warming to the end of the 6-hr period. Curve B represents all records taken after the initial warming period to the time at which individuals retreated to shelter the last time on the particular day. Class intervals of 0.5 C.
in contrast to the characteristics of the concentric gradient and open desert areas, where heat was supplied both by the substrate and by a source of radiant heat from above. Frequency distribu- tions of body temperature for lizards maintained in the rectangular gradient boxes were found to have the relatively high medians of 39-40 C, even for the distributions of body temperatures of
with records for lizards having the head over the warmer part of the thermal gradient than that occupied by the body, and an upper one associated with records for lizards having the head over the cooler part of the thermal gradient (Fig. 7). This suggested the existence of differences in temperature between head and body in these animals. To test whether this was the case, simultaneous

PRECISION OF THERMOREGULATION IN DESERT IGUANA 57
recordings of both body and head tem- peratures were made from animals sub- sequently used in the rectangular gra- dient boxes. In the next experiment, con- ducted on eight lizards adjusted to natural conditions of mid-August, head temperatures were found to be lower and showed less variation than body temper- atures (Fig. 8).
between temperature of the body and head for lizards tethered in the desert were also generally less than 1 C.
THE LEVEL AND PRECISION OF THERMO-
REGULATION UNDER NATURAL
CONDITIONS IN SUMMER
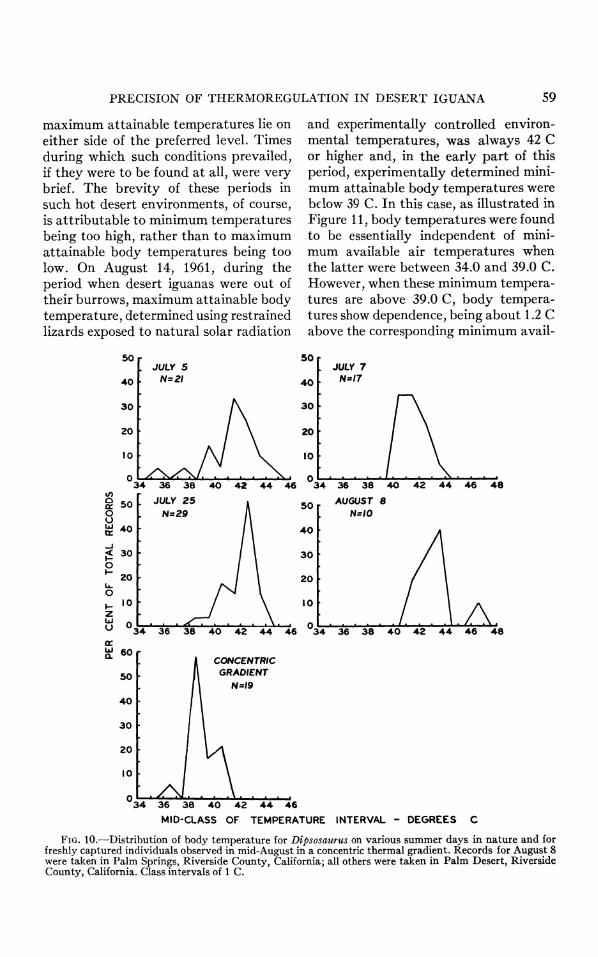
Desert iguanas in their native envi- ronment in summer often had body tem- peratures near 42 C (Fig. 10, July 5 and
14
12
o TOTAL RECORDS R ~I0
OL 0 I8
O
6 TRUNK ABOVE O HEAT SOURCE ,
4
HEAD ABOVE I-I
HEAT SOURCE --7 , "
=I \ 2 #, .,.,\ _
28 30 32 34 36 38 40 42 44 46 MID-CLASS OF TEMPERATURE INTERVAL - DEGREES C
FIG. 7.-Distribution of the body temperatures of 12 Dipsosaurus maintained in the linear thermal gradients supplied with substrate heat sources, showing the dissection of this distribution into two distribu- tions, one for lizards with their heads over the heat source for 5 min or longer, and the other for animals with their trunks over the heat source for 5 min or longer. Records are not corrected to give equal weight to the records for each individual. Class intervals of 0.5 C.
Temperature measurements of desert iguanas in nature during summer indi- cated that the head was either slightly warmer or slightly cooler than that of the body, depending upon environmental conditions and the position of the lizard in the environment (Fig. 9). The differ- ence seldom exceeded 1 C. Differences
25), a level also recorded by Norris (1953). However, freshly captured des- ert iguanas, adjusted to desert condi- tions of mid-August, maintained body temperatures at the 38.5 C level in an experimental concentric gradient (Fig. 10). Since these results suggest that the high body temperatures observed under

58 CALVIN B. DEWITT
,v - "
L4
IS
~3 I0 ~HEAD
0
8 "
0 TRUNK
06
2
0
31. 323 34 3 363 83 04 24 44
w
oW
31 32 33 34 35 36 37 38 39 40 41 42 43 44 45
MID-CLASS OF TEMPERATURE INTERVAL - DEGREES C.
FIG. 8.--Distributions of temperature of the head and trunk for eight Dipsosaurus adjusted to natural conditions. Determinations made using linear ther- mal gradients with substrate heat sources on August 2, 1961, in Palm Desert, California. Class intervals of 0.5 C.
natural conditions do not appear to be explained by acclimatization, it was worthwhile to determine the factors re- sponsible for the high body temperatures recorded under natural conditions. When the minimum environmental tempera- tures available to lizards on or above the surface of the ground are determined, it becomes apparent that these tempera- tures are often higher than 38.5 C. Con- sequently, it is obvious that when these conditions exist there is no opportunity to lower body temperature to the 38.5 C level when lizards remain on or above the surface of the ground. It is apparent then that the 42 C level of body tempera- ture observed is not necessarily repre- sentative of the preferred level. The pre- ferred level can only be reached when minimum available temperatures and
43
42 8
w~ 40
'83 41 I X ~
U pI
w
S40
gal
~38-
w
I.-/ 37
0930 0945 I000 1015 1030 1045
TIME OF DAY
FIG. 9.-Course of temperatures in the trunk and head of a freely moving Dipsosaurus on the open desert with dense shade available.
PRECISION OF THERMOREGULATION IN DESERT IGUANA 59
maximum attainable temperatures lie on either side of the preferred level. Times during which such conditions prevailed, if they were to be found at all, were very brief. The brevity of these periods in such hot desert environments, of course, is attributable to minimum temperatures being too high, rather than to maximum attainable body temperatures being too low. On August 14, 1961, during the period when desert iguanas were out of their burrows, maximum attainable body temperature, determined using restrained lizards exposed to natural solar radiation
and experimentally controlled environ- mental temperatures, was always 42 C or higher and, in the early part of this period, experimentally determined mini- mum attainable body temperatures were below 39 C. In this case, as illustrated in Figure 11, body temperatures were found to be essentially independent of mini- mum available air temperatures when the latter were between 34.0 and 39.0 C. However, when these minimum tempera- tures are above 39.0 C, body tempera- tures show dependence, being about 1.2 C above the corresponding minimum avail-
50 50 JULY 5 JULY 7
40 N=21 40 N=17
30 30
20 20
10 10
0 0 34 36 38 40 42 44 46 34 36 38 40 42 44 46 48
r)
5 JULY 25 50 AUGUST 8 S50 50 O N=29 N=IO U
" 40 40
-J S30 30 0
20 20
O IO 10
Z
U 034 36 38 40 42 44 46 34 36 38 40 42 44 46 48 (z: "
60 CONCENTRIC
50 GRADIENT N=19
40
30
20
10 I0
034 36 38 40 42 44 46
MID-CLASS OF TEMPERATURE INTERVAL - DEGREES C
FIG. 10.-Distribution of body temperature for Dipsosaurus on various summer days in nature and for freshly captured individuals observed in mid-August in a concentric thermal gradient. Records for August 8 were taken in Palm Springs, Riverside County, California; all others were taken in Palm Desert, Riverside County, California. Class intervals of 1 C.

60 CALVIN B. DEWITT
able air temperatures. At any particular time, minimum substrate temperatures on this day remained about 2 C below minimum air temperature, and body temperature, therefore, consistently ex- ceeded the former by about 3.2 C. Since
46
45
44
r .*
0
44
o / :.
38
/
"
34 35 36 37 38 39 40 41 4 43
MINIMUM AIR TEMPERATURE AT 4 CM - DEGREES C
39
34 35 36 37 38 39 40 41 42 43
MINIMUM AIR TEMPERATURE AT 4 CM - DEGREEs c
FIG. l.--Comparison of body temperatures of Dipsosaurus recorded August 14 at Palm Desert with minimum air temperature at 4 cm, showing no correlation (slope = -0.02; SD about the regression line = 0.77 C) prior to the time minimum air tempera- ture reaches 39 C, and showing positive correlation (slope = 1.00; so about the regression line = 0.65 C) after the time minimum air temperature reaches 39 C or higher.
minimum available temperatures rose with respect to time during the early part of the day and exceeded 39 C, the dependence of body temperature upon minimum available substrate tempera- tures was occasionally reflected by slowly rising body temperatures as shown in Figure 12. Some of the body-temperature distributions determined from the data of this figure are presented in Figure 10.
THE INFLUENCE OF BEHAVIORAL FACTORS
ON THE LEVEL AND PRECISION
OF THIIERMOREGULATION
Thermoregulation was most strongly influenced by territorial behavior and response to predators, somewhat less by feeding, and an apparently negligible amount by courtship.
Territorial behavior of desert iguanas most frequently consisted of threats which involved raising and lowering the head and anterior trunk several times in succession. In early morning, these threats were exchanged between indi- viduals in open areas. There seemed to be no great stimulus for lizards to remain
44 4 , * 44 Og 43 43
42 JULY 6
42
41 - 41
40 - * * 40
40 40
44 0 * 44
u 43 * 0 _ JULY 25 43
42 I . 42 **
wt 41 o
41
( 40 *40
oa 44 44 43 * e
A JULY 5 43
,. 42
t* 42 n- 41 Zo o, 41
4I- 40 * 40 L 39 * 39
138 38 - 37 45
>.. 36- JULY 7 -44 a
.o
35 t
o A "o -
-45 42 A . 44
41 -43
0 e
39 :o - A 4
40
*- AUGUSTI4
40
39 39 " so ' oeo '
'oo ' aoo 38 0800 0900 5000 5O00
TIME OF DAY
FIG. 12.-Diurnal courses of body temperature for Dipsosaurus under natural conditions in Palm Desert. Note the marked tendency for body tem- peratures to rise with time, particularly in the lower three plots.
in the open for long, except when push- ups comprised a portion of the sequence of behavior culminating in actual fight- ing between individuals. Two cases of

PRECISION OF THERMOREGULATION IN DESERT IGUANA 61
fighting were observed on the open sand on July 27. In one of these fights, which lasted from 8:50 to 8:55 A.M., exposure to the direct sun probably did not cause body temperatures to exceed 43 C, since neither lizard was panting-the thresh- old for panting in desert iguanas being at this temperature (Templeton, 1960). In the second encounter, which involved two other individuals and lasted from 9:18 to 9:27 A.M., panting began in both lizards about 15 sec after fighting com- menced. The body temperature of the victorious individual immediately after fighting was 45.9 C. In non-fighting desert iguanas under similar environ- mental conditions, such a high body temperature should occur much less than 1% of the time, judging by data for August 14 (Fig. 11).
Body temperatures of desert iguanas occasionally appear to rise above normal in hot weather when a person or predator is present, although this was never ob- served at Palm Desert. In Palm Springs on August 8, one desert iguana became heated to 46.5 C (see Fig. 10) while re- maining motionless from 8:59 to 9:04 A.M. while I was working within about 12 m of it. During this time, it remained in the open on a stick 4 cm above the surface, with its side perpendicular to solar radiation. It is of interest that a minimum substrate temperature of 40.0 was available beneath a nearby bush. Panting was not discernible in this lizard. Normally when heated to such an ex- tent, desert iguanas retreated to burrows or to the shade of a bush. Apparently, remaining motionless took precedence over moving to a bush in the case de- scribed. Movement to subterranean shel- ter was apparently prevented, since I must have been between the lizard and all readily accessible burrows.
Other unusually high body tempera-
tures of 46.4 C (Norris, 1953) and 46.8 (Mayhew, personal communication) may also represent cases where lizards did not retreat to shelter because of the presence of an observer. The situation may arise where a lizard is cut off from the burrow in its territory by a possible predator and from burrows in adjoining territories by pressures of territorial behavior.
Feeding apparently did not interfere with regulation of body temperature. Al- though it generally occurred in open areas, it was usually completed in the early part of the activity period when environmental temperatures were still fairly low. The diet of desert iguanas con- sisted almost exclusively of plants (pri- marily the foliage of Coldenia, Hymeno- clea, Euphorbia, Dalea, and the flowers of Eriogonum and Petalonyx), which, of course, were available at all times of day. The few occasions when insects were available occurred early in the activity period. Feeding on insects was observed on three of these occasions. The latest of these occurred at 8:45 A.M., and the body temperature of the lizard after feeding was not significantly above the level to be expected of non-feeding indi- viduals under similar conditions. Only once was extensive feeding observed under severe environmental conditions.
DISCUSSION
The negatively skewed distribution of body temperature: consequences for data analysis and possible physiological basis. -Some of the characteristics of the thermoregulatory system become evident from the analysis of the temporal courses of body temperature. First, the precision of behavioral thermoregulation as meas- ured in a thermal gradient is sufficient to restrict the central 50% of the body temperatures to the range between 37.0 and 39.5 C. Second, this analysis shows

62 CALVIN B. DEWITT
that the lower half of the central 50%- of the body temperatures (the 25/% lying below the 38.5 C median) is distributed over a wider range than the upper half. This is reflected in the negative skewness of the frequency distribution of body temperature (Fig. 6). Some investigators, in their analysis of thermoregulation of other species, have discarded the lower body temperatures in similarly skewed distributions on the assumption that all of these records are aberrant or have been recorded in the "basking range" (see Cowles and Bogert, 1944; Sould, 1963). Although there may be some jus- tification for eliminating records of body temperature recorded from lizards during their initial basking period, all records of low body temperature cannot simply be assumed to lie in the basking range. This is especially apparent from records of desert iguanas in the experimental thermal gradient which show that these low body temperatures occur throughout the day, after the initial period of warm- ing as well as before it. Moreover, similar records of low body temperature have also been recorded throughout the day for Sceloporus orcutti in nature (Mayhew, 1963). The only records, then, which can justly be eliminated from analysis appear to be those which are definitely known to have occurred prior to and during the initial period of warming. In addition, there may be some justification for elimi- nating all body temperatures recorded after the lizard retreats to shelter at the end of the activity period and does not return. After omitting these records from analysis, however, the distribution still is negatively skewed. Since this skewness appears to be real, it is worthwhile to speculate on why this might be so. The mechanism for regulation of temperature could provide the basis for the observed distribution if it depended upon detec-
tion of deviations from a set point of some rate process bearing a direct expo- nential relation to body temperature. As an example, the intensity of physiologi- cal processes, as reflected by oxygen con- sumption, might be regulated about a particular level according to a normal curve of variation. Since physiological activity bears a direct exponential rela- tion to temperature, the corresponding body-temperature distribution for the animals concerned would necessarily be negatively skewed. Such a system of regulation might also equally apply to other animals whose distribution of body temperatures is negatively skewed.
Expression of the level and precision of control.-Since skewness apparently can be characteristic of the distribution of body temperature for a lizard, it seems important, first of all, to determine the degree and kind of skewness which may be present, using an estimate of skewness such as g1 (Snedecor, 1946, p. 174). Only if the distribution is found to be normal can the mean and standard deviation adequately describe the level and preci- sion of control. However, if the distribu- tion is skewed, the level of body-temper- ature control can be expressed by the median of the distribution, and the pre- cision can be expressed by the range of temperature within which a certain per- centage of all observations are found. The percentages useful for this include those which correspond to plus and minus 1.96, 1.00, and 0.674 times the standard deviation of a normal distribu- tion. These are the central 95, 68, and 50% of the distribution, respectively. For both normal and skewed distribu- tions, the preferendum can be expressed by the center of the distribution plus and minus the difference between it and the limits of the central 68% of the distribu- tion. For desert iguanas in the concentric

PRECISION OF THERMOREGULATION IN DESERT IGUANA 63
thermal gradient, this preferendum is
38.5 16C. -2.4
The direction and degree of skewness are indicated by the ratio of the positive figure to the negative figure.
The central 95% of the records for desert iguanas (33.2-41.8 C) corresponds very well to the temperatures given by Cowles and Bogert (1944, p. 279) as de- limiting the "normal activity range" for this species (34-41 C). The use of the central 95% of the distribution, however, is advantageous since it is not delimited by single, unusual occurrences of body temperatures at the extremes of the dis- tribution.
The influence of physical and biologi- cal factors on the process of control now can be assessed using as a basis the analysis of the process of body-tempera- ture control.
The relation of thermoregulation to ther- mal factors of the environment.-The suc- cessful behavioral control of body tem- perature is largely dependent upon the availability of the necessary thermal con- ditions in the environment. However, these conditions may at times be inac- cessible to a lizard, due to their absence from the area or to isolation of a lizard from these conditions by barriers such as territorial boundaries. At times, thermal conditions may be insufficient to permit body temperature to rise to the preferred level, and at other times they may be insufficient to permit body temperatures below this level. The former conditions may occur often in the temperate zone, and under these conditions lizards would be expected to be very close to the maxi- mum attainable temperature. Desert iguanas very likely encounter such con- ditions only at times in the early part of the year. Extreme conditions of the
other type, namely, those insufficient to permit body temperatures below the pre- ferred level, may occur on occasion in some environments and consistently occur for desert iguanas at Palm Desert in summer (DeWitt, 1963). Under these conditions, active control of body tem- perature at the preferred level is no longer possible above ground, and lizards must either retreat to burrows or allow body temperatures to rise parallel to the rising minimum attainable temperatures of the environment. For desert iguanas, this rise is usually permitted only to the level at which panting begins.
Permitting body temperatures to rise parallel to minimum attainable tempera- tures of the environment when these temperatures are above the preferred level probably is not peculiar to desert iguanas. Mosauer (1936) found that Uma notata came to rest in a laboratory ther- mal gradient at soil temperatures vary- ing from 35 to 40 C, but in the field this same species came to rest at soil tempera- tures ranging from 37 to 43 C. Although Cowles and Bogert (1944) attributed this difference to inadequacy of the ap- paratus, pursuit of lizards in the field resulting in overheating, and other "diffi- culties inherent in thermal work in the field," it now seems likely that this is due to U. notata permitting body tem- peratures to rise when minimum attain- able temperatures exceed the preferred range. Similarly, the discrepancy report- ed by Fitch (1956) between his data for Cnemidophorus sexlineatus in Kansas and Bogert's (1949) data for this species in Florida may be due to the animals in Florida being forced above the preferred range in contrast to those in Kansas. Body temperatures of some tropical spe- cies of Anolis are lower in the earlier half of the activity period than those in the later half (Ruibal, 1961), but it is not

64 CALVIN B. DEWITT
possible to determine from the data pre- sented for this species whether this is due to the inclusion of records from lizards in the initial heating range of the morn- ing or whether the difference is due to body temperatures in the afternoon being forced to rise above the preferred range. In a tropical environment, the latter possibility seems less likely.
The advantage of permitting body temperatures to rise above the preferred range is to increase the period during which activity is possible above ground. On very warm summer days, this be- havior may increase the time suitable for activity above ground from 0.5 to about 3 hr. This lengthening of the ac- tivity period above ground has the ob- vious advantage of prolonging the period in which feeding and other necessary activities can be conducted. As such, it may well be one of the major factors con- tributing to the success of diurnal desert lizards.
The relation of panting to thermal fac- tors of the environment.-Under some cir- cumstances, lizards remain above ground after the threshold for panting is sur- passed. Although not all lizards pant when exposed to high temperatures (Stebbins, 1961), some lizards, such as Uromastix and Varanus, use it as an effective means of cooling (Langlois, 1902). For desert iguanas at body tem- peratures from 32 to 44 C, the amount of heat lost by evaporation of water is approximately compensated for by meta- bolic heat production (Templeton, 1960). Above the panting threshold of 43-44 C, however, evaporation appears to be effective in cooling, as indicated by the results of some initial experiments I have done on panting in this species. In these experiments, desert iguanas with initial body temperatures of 38.5 C were placed in a flow of air at 50 C and about 400
ft/minute. Their trunk and head temper- atures leveled off at 3 and 6 C, respec- tively, below air temperature after 35 min, and these differences were main- tained for at least another 25 min.
Diferences in temperature of head and body.-Maximum differences between temperatures of head and body for desert iguanas under certain unusual experi- mental conditions are similar to those recorded by Heath (1964) from horned lizards (Phrynosoma) prior to, during, and following emergence in an experi- mental chamber supplied with an over- head heat lamp. For desert iguanas in nature, such large differences apparently do not usually exist. During panting, however, as indicated above, the head may be maintained at a temperature 3 C less than that of the body. Also, since desert iguanas during emergence may remain for some time at the burrow en- trance with only their heads exposed to solar radiation, particularly when there is some disturbance in the vicinity (DeWitt, unpublished data), it seems likely that large differences exist under some conditions during emergence. While heating the head of desert iguanas, I have noted that the eyes appear to swell and that blinking of the eyes begins, which suggests that desert iguanas em- ploy the same mechanism for equalizing temperature of head and body as that described by Heath (1964) for horned lizards. Pressures in the cranial blood sinuses of desert iguanas occasionally rise enough under these conditions so that the sinuses burst, resulting in blood running out of the eye.
The relation of the precision and level of thermoregulation to behavioral factors.- Even when environmental conditions are known to be suitable for regulation at the preferendum, body temperatures of des- ert iguanas in nature may not be those

PRECISION OF THERMOREGULATION IN DESERT IGUANA 65
expected. This certainly is the case for lizards which are fighting or are remain- ing quiet in the presence of a predator, but it is also true at other times as well. For example, from 8:00 to 11:00 A.M. on August 14, 1961, conditions were such that body temperatures would be expect- ed to be both above and below the pre- ferred level. Although body tempera- tures were very near this level, none were below it. This probably reflects both behavioral pressures forcing desert igua- nas to move out of the shade during the course of their normal activities and the contribution of extensive, open unshaded areas to elevating body temperatures during the normal course of movements by these lizards between shaded areas.
Location of the central control of body temperature.-Information on the cen- tral and peripheral thermoreceptors as- sumed to be involved in thermoregula- tion of lizards is not yet available. How- ever, some of the results of this study may appear to some readers to supply some information on the location of the central control. One of these results, pre- sented in Figure 8, shows that, under certain peculiar environmental condi- tions, the head temperature apparently is regulated with greater precision than body temperature. Even though one might like to implicate the hypothalamus as the site of central control, I regard this to be inadequate evidence that the central receptors are located in the head. This apparently greater precision of head temperature may simply reflect a ther- mal environment which is less variable at the level of the head than it is at the level of the body. More adequate, but still weak, evidence that the central con- trol of body temperature may be located in the head is suggested when one com- pares Figures 6 and 8 and observes that
it is the center of the distribution of head temperature in Figure 8, and not that of body temperature, which closely corre- sponds to the level of regulation indicat- ed in Figure 6.
SUMMARY
1. Desert iguanas exercise control over their body temperature by appropriate adjustment of their position in artificial and natural thermal gradients.
2. The preferred level of body tem- perature, which is defined as the median of the body-temperature distribution, is 38.5 C.
3. Thermoregulation in desert iguanas is fairly precise with the central 50, 68, and 95% of the body temperatures rang- ing between 37.0-39.5, 36.1-40.1, and 33.2-41.8 C, respectively.
4. The body-temperature distribution for desert iguanas is negatively skewed. This may reflect a characteristic of the system for temperature sensing employed by lizards. In analysis of body-tempera- ture preferences, neither the usual as- sumption of normality nor the common practice of casting off records of low body temperatures is wholly justified.
5. Under certain experimental condi- tions, temperatures of the head and trunk may differ by several degrees; but under natural conditions, these tempera- tures do not differ by more than a degree.
6. Under desert conditions during summer, desert iguanas prolong their activity period by permitting body tem- peratures to rise above the preferred level to 42 C and higher.
7. Body temperatures are permitted to rise well above the preferred level during territorial fighting behavior and in the presence of predators but generally are not permitted to do this during feed- ing and courtship.

66 CALVIN B. DEWITT
LITERATURE CITED
BOGERT, C. M. 1949. Thermoregulation in reptiles, a factor in evolution. Evolution 3:195-211.
CARPENTER, C. C. 1962. Patterns of social behavior in the desert iguana, Dipsosaurus dorsalis. Copeia 1962:396-405.
COWLES, R. B., and C. M. BOGERT. 1944. A pre- liminary study of thermal requirements of desert reptiles. Amer. Mus. Natur. Histor. Bull. 83: 261-296.
DAWSON, W. R., and G. A. BARTHOLOMEW. 1958. Metabolic and cardiac responses to temperature in the lizard Dipsosacurus dorsalis. Physiol. Zo61. 31:100-111.
DEWITT, C. B. 1963. Behavioral thermoregulation in the iguanid lizard, Dipsosaurus dorsalis. Ph.D. thesis, Univ. Michigan. 92 pp. Univ. Microfilms, Ann Arbor, Mich. (Diss. Abstr. 25: 2670).
FITCH, H. S. 1956. Temperature responses in free- living amphibians and reptiles of northeastern Kansas. Univ. Kansas Mus. Nat. Hist. Pub. 8: 417-475.
HEATH, J. E. 1964. Head-body temperature differ- ences in horned lizards. Physiol. Zoo1. 37:273- 279.
LANGLOIS, J. 1902. La rdgulation thermique chez les poikilothermes. J. Physiol. Pathol. G6n. 4:249- 256.
LICUT, P. 1965. Effects of temperature on heart
rates of lizards during rest and activity. Physiol. Zoo1. 38:129-137.
MAYHEW, W. W. 1963. Temperature preferences of Sceloporus orcutti. Herpetologica 18:217-233.
MOBERLY, W. R. 1963. Hibernation in the desert iguana, Dipsosaurus dorsalis. Physiol. Zoo1. 36: 152-160.
MOSAUER, W. 1936. The toleration of solar heat in desert reptiles. Ecology 17:56-66.
NORRIS, K. S. 1953. The ecology of the desert iguana Dipsosaurus dorsalis. Ecology 34:265-287.
RUIBAL, R. 1961. Thermal responses of five species of tropical lizards. Evolution 15:98-111.
SAINT-GIRONS, H., and M. SAINT-GIRONS. 1956. Cycle d'activit6 et thermorigulation chez les reptiles (1izards et serpents). Vie Milieu 7:133- 226.
SNEDECOR, G. W. 1946. Statistical methods applied to experiments in agriculture and biology. Iowa State Coll. Press, Ames. 485 pp.
SOULA, M. 1963. Aspects of thermoregulation in nine species of lizards from Baja, California. Copeia 1963:107-115.
STEBBINS, R. C. 1961. Body temperature studies in South African lizards, Koedoe 1961 (4):54-67.
TEMPLETON, J. R. 1960. Respiration and water loss at the higher temperatures in the desert iguana, Dipsosaurus dorsalis. Physiol. Zoil. 33:136-145.




















