
Potentiation of the antitumor effect of 11-keto- β-boswellic acid by its 3-α-hexanoyloxy...
11
Molecular and Cellular Pharmacology Potentiation of the antitumor effect of 11-keto-β-boswellic acid by its 3-α-hexanoyloxy derivative Gousia Chashoo a , Shashank K. Singh a, ⁎, Dilip M. Mondhe a , Parduman R. Sharma a , Samar S. Andotra b , Bhahwal.A. Shah b , Subhash C. Taneja b , Ajit K. Saxena a a Pharmacology Division, Indian Institute of Integrative Medicine, canal road, Jammu, India b Bio-organic chemistry Division, Indian Institute of Integrative Medicine, canal road, Jammu, India abstract article info Article history: Received 4 February 2011 Received in revised form 27 June 2011 Accepted 7 July 2011 Available online 30 July 2011 Keywords: Topoisomerse I and II Hexanoyloxy-11-keto-β-boswellic acid Apoptosis HL-60 cells We recently discovered that a propionyloxy derivative of 11-keto-β-boswellic acid (PKBA) showed better anticancer potential than other boswellic acids including AKBA, encompassing the importance of acyl group at the 3-α-hydroxy position of KBA. In continuation of our previous work, other higher derivatives (with increasing alkoxy chain length at 3-α-hydroxy position) including butyryloxy (BKBA) and hexanoyloxy (HKBA) derivatives of KBA were synthesized. The respective IC 50 values of BKBA and HKBA in HL-60 cells were found to be 7.7 and 4.5 μg/ml. IC 50 value of HKBA was comparatively lower than that of BKBA, and further lower than that of the previously reported derivative (PKBA, IC 50 8.7 μg/ml). In order to compare the anticancer potential of HKBA with PKBA, detailed in vitro pro-apoptotic and in vivo anticancer studies were carried out. The induction of apoptosis by HKBA was measured using various parameters including fluorescence and scanning electron microscopy, DNA fragmentation and Annexin V-FITC binding. The extent of DNA damage was measured using neutral comet assay. HKBA was further evaluated for its effect on DNA cell cycle and mitochondria where it was found to arrest cells in G 2 /M phase and also induced loss of mitochondrial membrane potential. These events were associated with increased expression of cytosolic cytochrome c and cleavage of PARP. Target based studies showed that HKBA inhibited the enzymatic activity of topoisomerases I and II at low doses than that of PKBA. In vivo studies also revealed a low dose inhibitory effect of HKBA on ascitic and solid murine tumor models. © 2011 Elsevier B.V. All rights reserved. 1. Introduction Based on their anti-inflammatory effects boswellic acids have been used for the treatment of many diseases including Crohn's disease, ulcerative colitis, bronchial asthma, endotoxin induced hepatitis and arthritis (Shah et al., 1997). In the recent years, boswellic acids started to draw attention for their anticancer potential (Shah et al., 2007). Although, there are no direct reports of the antitumor effect of boswellic acids in clinical trials, however, results from in vitro studies have shown that boswellic acids inhibit the cellular proliferation and induce apoptosis in brain tumor (Glaser et al., 1999; Park et al., 2002; Winking et al., 2000), leukemia (Han, 1994; Hoernlein et al., 1999; Jing et al., 1999; Shao et al., 1998), melanoma, hepatoma and prostate cancer cell lines (Liu et al., 2002). Moreover, the clear mechanism by which boswellic acids inhibit the cell growth is not clear, however it has been reported that boswellic acids inhibit the enzymatic activity of topoisomerases, therefore influencing the DNA synthesis and replication in cancer cells. As has been observed by Hoernlein et al. (1999) that acetyl-11-keto-β-boswellic acid (AKBA) induced apopto- sis in HL-60 cells by inhibiting topoisomerase I, we recently observed that propionyloxy-11-keto-β-boswellic acid (PKBA) induced apopto- sis in HL-60 cells by inhibiting topoisomerase I and II at lower concentration than AKBA (Chashoo et al., 2011). This study raised a point towards the potentiating role of acyl group at 3-α-hydroxy position on the anticancer potential of KBA. Considering the increment in its anticancer potential by PKBA, we synthesized two higher derivatives of KBA including butyryloxy (BKBA) and hexanoy- loxy (HKBA) derivatives and evaluated their anti-proliferative potential vis-a-vis PKBA (Chashoo et al., 2011), in order to develop a novel compound with improved anticancer properties. The in vitro antiproliferative potential of these newly synthesized analogs was evaluated in human leukemia HL-60 and Molt-4 cell lines. The IC 50 value of HKBA (4.5 μg/ml) in HL-60 cell line was found to be lower than BKBA (7.7 μg/ml) which was comparatively even lower than the previously reported acyl derivative, PKBA (IC 50 8.7 μg/ml). Encour- aged by the results, detailed studies were carried out to evaluate the anticancer potential of this newly synthesized analog. Our study in murine tumor models demonstrated that HKBA is able to exert antitumor effects in vivo at low doses. Owing to its ability to induce cancer cells to death at low doses, mechanism based studies were European Journal of Pharmacology 668 (2011) 390–400 ⁎ Corresponding author. E-mail address: [email protected] (S.K. Singh). 0014-2999/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.ejphar.2011.07.024 Contents lists available at ScienceDirect European Journal of Pharmacology journal homepage: www.elsevier.com/locate/ejphar
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Potentiation of the antitumor effect of 11-keto- β-boswellic acid by its 3-α-hexanoyloxy...

European Journal of Pharmacology 668 (2011) 390–400
Contents lists available at ScienceDirect
European Journal of Pharmacology
j ourna l homepage: www.e lsev ie r.com/ locate /e jphar
Molecular and Cellular Pharmacology
Potentiation of the antitumor effect of 11-keto-β-boswellic acid by its3-α-hexanoyloxy derivative
Gousia Chashoo a, Shashank K. Singh a,⁎, Dilip M. Mondhe a, Parduman R. Sharma a, Samar S. Andotra b,Bhahwal.A. Shah b, Subhash C. Taneja b, Ajit K. Saxena a
a Pharmacology Division, Indian Institute of Integrative Medicine, canal road, Jammu, Indiab Bio-organic chemistry Division, Indian Institute of Integrative Medicine, canal road, Jammu, India
⁎ Corresponding author.E-mail address: [email protected] (S.K. Sin
0014-2999/$ – see front matter © 2011 Elsevier B.V. Adoi:10.1016/j.ejphar.2011.07.024
a b s t r a c t
a r t i c l e i n f oArticle history:Received 4 February 2011Received in revised form 27 June 2011Accepted 7 July 2011Available online 30 July 2011
Keywords:Topoisomerse I and IIHexanoyloxy-11-keto-β-boswellic acidApoptosisHL-60 cells
We recently discovered that a propionyloxy derivative of 11-keto-β-boswellic acid (PKBA) showed betteranticancer potential than other boswellic acids including AKBA, encompassing the importance of acyl group atthe 3-α-hydroxy position of KBA. In continuation of our previous work, other higher derivatives (withincreasing alkoxy chain length at 3-α-hydroxy position) including butyryloxy (BKBA) and hexanoyloxy(HKBA) derivatives of KBAwere synthesized. The respective IC50 values of BKBA and HKBA in HL-60 cells werefound to be 7.7 and 4.5 μg/ml. IC50 value of HKBA was comparatively lower than that of BKBA, and furtherlower than that of the previously reported derivative (PKBA, IC50 8.7 μg/ml). In order to compare theanticancer potential of HKBA with PKBA, detailed in vitro pro-apoptotic and in vivo anticancer studies werecarried out. The induction of apoptosis by HKBA was measured using various parameters includingfluorescence and scanning electron microscopy, DNA fragmentation and Annexin V-FITC binding. The extentof DNA damage was measured using neutral comet assay. HKBA was further evaluated for its effect on DNAcell cycle and mitochondria where it was found to arrest cells in G2/M phase and also induced loss ofmitochondrial membrane potential. These events were associated with increased expression of cytosoliccytochrome c and cleavage of PARP. Target based studies showed that HKBA inhibited the enzymatic activityof topoisomerases I and II at low doses than that of PKBA. In vivo studies also revealed a low dose inhibitoryeffect of HKBA on ascitic and solid murine tumor models.
gh).
ll rights reserved.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
Based on their anti-inflammatory effects boswellic acids have beenused for the treatment of many diseases including Crohn's disease,ulcerative colitis, bronchial asthma, endotoxin induced hepatitis andarthritis (Shah et al., 1997). In the recent years, boswellic acids startedto draw attention for their anticancer potential (Shah et al., 2007).Although, there are no direct reports of the antitumor effect ofboswellic acids in clinical trials, however, results from in vitro studieshave shown that boswellic acids inhibit the cellular proliferation andinduce apoptosis in brain tumor (Glaser et al., 1999; Park et al., 2002;Winking et al., 2000), leukemia (Han, 1994; Hoernlein et al., 1999;Jing et al., 1999; Shao et al., 1998), melanoma, hepatoma and prostatecancer cell lines (Liu et al., 2002). Moreover, the clear mechanism bywhich boswellic acids inhibit the cell growth is not clear, however ithas been reported that boswellic acids inhibit the enzymatic activityof topoisomerases, therefore influencing the DNA synthesis andreplication in cancer cells. As has been observed by Hoernlein et al.
(1999) that acetyl-11-keto-β-boswellic acid (AKBA) induced apopto-sis in HL-60 cells by inhibiting topoisomerase I, we recently observedthat propionyloxy-11-keto-β-boswellic acid (PKBA) induced apopto-sis in HL-60 cells by inhibiting topoisomerase I and II at lowerconcentration than AKBA (Chashoo et al., 2011). This study raised apoint towards the potentiating role of acyl group at 3-α-hydroxyposition on the anticancer potential of KBA. Considering theincrement in its anticancer potential by PKBA, we synthesized twohigher derivatives of KBA including butyryloxy (BKBA) and hexanoy-loxy (HKBA) derivatives and evaluated their anti-proliferativepotential vis-a-vis PKBA (Chashoo et al., 2011), in order to develop anovel compound with improved anticancer properties. The in vitroantiproliferative potential of these newly synthesized analogs wasevaluated in human leukemia HL-60 and Molt-4 cell lines. The IC50value of HKBA (4.5 μg/ml) in HL-60 cell line was found to be lowerthan BKBA (7.7 μg/ml) which was comparatively even lower than thepreviously reported acyl derivative, PKBA (IC50 8.7 μg/ml). Encour-aged by the results, detailed studies were carried out to evaluate theanticancer potential of this newly synthesized analog. Our study inmurine tumor models demonstrated that HKBA is able to exertantitumor effects in vivo at low doses. Owing to its ability to inducecancer cells to death at low doses, mechanism based studies were

391G. Chashoo et al. / European Journal of Pharmacology 668 (2011) 390–400
carried out, to evaluate the pro-apoptotic potential of this newlysynthesized analog. At low concentrations, HKBA was found to induceapoptosis in HL-60 cells as studied viamicroscopy, gel electrophoresisand Annexin V-FITC binding of the cells. The DNA damage induced byHKBA was studied by neutral comet assay. Further, HKBA was foundto arrest cells in the G2/M phase of cell cycle. The induction ofapoptosis by HKBA was found to involve mitochondrial pathway,resulting in the loss of mitochondrial membrane potential, release ofproapoptotic factors from mitochondrion and activation of upstreamand downstream caspases. Furthermore, HKBA was found to inhibitthe catalytic activity of topoisomerases I and II at low doses. Overall,these established HKBA to be potentially better anticancer compoundthan other previously known boswellic acids including PKBA.
2. Materials and methods
2.1. Chemicals
3-(4,5-Dimethylthiazole-2-yl)-2,5-diphenyltetrazolium bromide(MTT), phosphate buffer saline, dimethyl sulphoxide (DMSO), RPMI-1640, ethidium bromide, propidium iodide (PI), DNase-free RNase,Proteinase K, camptothecin, 5-fluorouracil, Rh-123, penicillin, strepto-mycin, Adriamycin, mitomycin-C, fetal bovine serum, electrophoresisreagents, protein estimation kit and protein markers were purchasedfrom Sigma Chemical Co., USA.Mouse anti-human antibodies to PARP-1(#SC8007), β-actin (#SC-47778) and goat anti-mouse IgG-HRP(#SC2031) were from Santa Cruz Biotechnology, USA. Annexin-V-FITCapoptosis detection kit was fromApoalert, BD Biosciences, USA. Caspaseassay kits were from R&D systems, USA. Topoisomerase detection kitswere purchased from TopoGEN Inc., Port Orange, USA.
2.2. Cell lines, cell culture, growth conditions and treatment
Human promyelocytic leukemia HL-60 and human acute lympho-blastic leukemia (Molt-4) cell lines were purchased from NationalCenter for Cell Science, Pune. Cell lines were grown andmaintained inRPMI-1640 medium, pH 7.4. The media was supplemented withFCS (10%), penicillin (100 units/ml), streptomycin (100 μg/ml) andglutamine (2 mM). Cells were grown in CO2 incubator (Heraeus,GmbH, Germany) at 37 °C with 98% humidity and 5% CO2. Cells weretreated with BKBA and HKBA dissolved in DMSO while the untreatedcontrol cultures received only the vehicle. The concentration of DMSOadded to the cultures was maintained tob1%.
2.3. Synthesis of 3-α-hexanoyloxy-11-keto-β-boswellic acid (HKBA) and3-α-butyryloxy-11-keto-β-boswellic acid (BKBA)
11-Keto-β-boswellic acid (KBA, 2 mmol)was isolated fromamixtureof boswellic acids by method reported in literature (Shah et al., 2007).KBA was treated with hexanoic anhydride (1.2 eq) in presence ofdimethyl amino pyridine (DMAP) as a catalyst in dry DCM (10 ml)
Scheme 1. Synthesis o
(Scheme 1). After completion, the reaction mixture was treated withsaturated sodiumbicarbonate solution and extractedwith DCM (3×100ml). The crude product thus obtained after purification on a silica gelcolumn gave pure HKBA as a colorless semisolid in 95% yield. BKBA wasalso prepared following similar method as for HKBA, using butyricanhydride in 95% yield.
2.4. Spectral data of HKBA
Colorless semisolid, [α]D25 +37 (c 1.0 CHCl3). 1H NMR (200 MHz,CDCl3): δ 0.83, 0.89, 1.12, 1.15, 1.20, 1.34 (24H, 23, 25, 26, 27, 28, 29and 30 ―CH3 and ―CH2CH3), 1.34–1.89 (m, nH), 2.35 (t, J=7.3 Hz,2H, ―CH2CO), 5.31 (bs, 1H, H-3), 5.56 (bs, 1H, H-12). 13C NMR(50 MHz, CDCl3): δ 12.1, 12.9, 16.4, 17.4, 19.4, 20.1, 21.3, 22.5, 22.9,23.6, 23.9, 27.3, 27.8, 28.9, 30.2, 33.6, 34.0, 36.4, 37.3, 37.8, 38.2, 39.2,39.3, 40.9, 42.7, 44.1, 49.5, 56.5, 58.0, 59.4, 71.8, 129.4, 164.0, 171.8,177.7, 197.4; ESI-MS (m/z): 567 [M+1]+. Anal. Calc. for C36H56O5: C,76.01; H, 9.92. Found: C, 76.13; H, 10.03.
2.5. Spectral data of BKBA
White solid, [α]D25 +51.8 (c 1.0 CHCl3), mp 264–266 °C. 1H NMR(200 MHz, CDCl3): δ 0.83, 0.93, 1.0, 1.12, 1.15, 1.24 (24H, 23, 25, 26,27, 28, 29 and 30 ―CH3 and ―CH2CH3), 1.40–2.39 (m, nH), 2.33 (t,J=7.3 Hz, 2H, ―CH2CO), 5.31 (bs, 1H, H-3), 5.56 (bs, 1H, H-12). 13CNMR (50 MHz, CDCl3): δ 12.9, 13.4, 17.5, 18.4, 18.9, 20.6, 21.1, 21.3,23.6, 23.8, 27.3, 27.5, 28.3, 31.7, 33.2, 34.3, 36.8, 37.2, 37.7, 39.2, 39.3,40.9, 44.1, 44.9, 47.7, 49.1, 57.5, 59.9, 73.8, 130.2, 163.1, 172.5, 180.5,199.3; ESI-MS (m/z): 563 [M+Na]+. Anal. Calc. for C34H52O5: C,75.51; H, 9.69. Found: C, 75.64; H, 9.81.
2.6. Animal care and housing
Non-inbred Swiss albinomice from an in-house colonywere used inthe present study. The animals were housed under standard husbandryconditions as per guide for the care anduse of laboratory animals (1996).Animals were provided pelleted mice feed (M/s Ashirwad Industries,Chandigarh, India) and autoclaved water ad libitum. The study and thenumber of animals used were approved by the Institutional AnimalEthics Committee, Indian Institute of Integrative Medicine, Jammu.
2.7. Cell proliferation assay
Cells (1×104/200 μl medium)were delivered into 96-well plates andexposed to indicated concentrations of HKBA for 48 h. The cells wereincubated with 20 μl of MTT solution (2.5 mg/ml PBS) for 4 h and theMTT-formazon crystals formedweredissolved inDMSO;ODmeasuredatλ 570 nmwith a referencewavelength of 620 nm (Bhushan et al., 2006).
f HKBA and BKBA.

0
0.2
0.4
0.6
0.8
1
1.2
0 µg/ml 10µg/ml 30 µg/ml 50 µg/ml 70 µg/ml 100µg/mlCel
l Via
bilit
y, O
D a
t 57
0 nm
HKBA
….HL-60 , IC504.5 µg/ml ----Molt-4, IC50 6 µg/ml
0
0.2
0.4
0.6
0.8
1
1.2
0 µg/ml 10µg/ml 30 µg/ml 50 µg/ml 70 µg/ml 100µg/ml
Cel
l Via
bilit
y, O
D a
t 57
0 nm
BKBA
….HL-60 , IC507.7 µg/ml ----Molt-4, IC50 9µg/ml
Fig. 1. The in vitro cell proliferation assay of BKBA and HKBAwas done using MTT dye inHL-60 and Molt-4 cells. Both BKBA and HKBA showed a dose dependent growthinhibition of HL-60 and Molt-4 cells after 48 h of incubation. Data are mean ±S.D. fromthree similar experiments.
392 G. Chashoo et al. / European Journal of Pharmacology 668 (2011) 390–400
2.8. DNA Fragmentation
Exponentially growing 2×106 cells/ml in a 6well culture plate weretreated with different concentrations of test compound for 24 h. Cellswere lysed, genomic DNA extracted and electrophoresed as describedearlier (Zhuang et al., 2000).
2.9. In vivo antitumor studies
The studies for in vivo anticancer activity in mouse models wereconducted as per the guidelines of the National Cancer Institute (NCI),USA. (Geran et al., 1972). Ehrlich ascites carcinoma (EAC) cells werecollected from the peritoneal cavity of the Swiss mice harboring 8–10 days old ascitic tumor. 1×107 EAC cells were injected intraperito-neally in Swissmice selected for the experiment on day 0. The next day,animals were randomized and divided into different groups. Thetreatment groups contained7 animals each and control groupcontained10 animals. Two treatment groups were treated with 50 mg/kg and60 mg/kg of HKBA intraperitoneally from day 1 to 9. One more of thetreatmentgroups received5-fluorouracil (20 mg/kg, i.p) and it served aspositive control. The tumor bearing control group was similarlyadministered normal saline (0.2 ml, i.p.). On day 12, animals weresacrificed and ascitic fluid was collected from the peritoneal cavity ofeach mouse for the evaluation of tumor growth. Percent tumor growthinhibition was calculated based on the total number of tumor cellspresent in the peritoneal cavity as onday 12of the experiment using thefollowing formula.
Percent tumour growth inhibition
=Av: no: of cells in control group–Av: no: of cells in treated group
Av: no: of cells in control group×100
The doses of HKBA were prepared in gum acacia (0.5% v/v) so thatthe required dose to be administered to each animal was contained in0.2 ml. For solid tumor (EAT and S-180) studies, Ehrlich asciticcarcinoma (EAC) cells were collected from the peritoneal cavity of theSwiss mice harboring 8–10 days old ascitic tumor. 1×107 EAC cellswere injected intramuscularly in right thigh of 31 Swiss male miceselected for the experiment on day 0. The next day, animals wererandomized and divided into four groups. Three treatment groupscontained 7 animals each and one control group contained 10 animals.The first and the second treatment groups received 50 and 60 mg/kgi.p. HKBA respectively from day 1 to 9. The third treatment group wastreated with 5-fluorouracil (22 mg/kg, i.p) from day 1 to 9 and itserved as positive control. The control group was similarly adminis-tered normal saline (0.2 ml, i.p.) from day 1 to 9. On day 9 and 13, thetumor bearing thigh of each animal was shaved and the longest andshortest diameters of the tumor were measured with the help of avernier caliper. Tumor weight of each animal was calculated using thefollowing formula.
Tumour weight mgð Þ = Length mmð Þ × width mmð Þ½ �22
The percent tumor growth inhibition was calculated on day 13 bycomparing the average values of treated groups with that of controlgroup. Tumor growth in saline treated control animals was taken to be100%.
2.10. Nuclear morphology
The presence of apoptotic cells were shown by staining cells withDAPI (4′-6-Diamidino-2-phenylindole) and observing them usingfluorescence microscope as per the method described earlier (Relloet al., 2005).
2.11. Scanning electron microscopy studies
For SEM, HL-60 cellswere incubatedwith 5 μg/mlHKBA for 24 h andprocessed further. (Rello et al., 2005; Waxman and Schwartz, 2003).Briefly, cells on a cover slip were fixed in glutaraldehyde, osmicated,dehydrated, dried in a critical point drier using CO2 (Balzer's Union) andcoated with gold using a sputter coater (Polaron). The specimens wereexaminedwith a JEOL-100CXII electronmicroscopewith ASID at 40 KV.
2.12. Detection of apoptosis
As described by Bhushan et al. (2006) previously, Annexin-V/propidium iodide (PI) binding assay was employed to determine theviable, early and late apoptotic cells. According to the instructiongiven by the manufacturer, 1×106 cells were treated with differentconcentrations of HKBA for 24 h and stained with Annexin V/PI. Thecells were scanned for fluorescence intensity in FL-1 (FITC) and FL-2(PI) channels. The fraction of cell population in different quadrantswas analyzed using quadrant statistics. Cells in the lower rightquadrant represented apoptosis and in the upper right quadrantrepresented necrosis or post apoptotic necrosis.
2.13. Neutral comet assay for detection ofDNAdouble strandbreaks (DSBs)
Theextent ofDNAdamage causedby single strand anddouble strandbreaks was determined by single-cell gel electrophoresis. HL-60 cells(2×106 cells/ml/well) after treatment with 5 and 10 μg/ml of HKBA for24 h were harvested and subjected to single cell gel electrophoresis asper the method described previously (Chashoo et al., 2011).
2.14. Measurement of cell cycle/DNA content
Cell cycle and sub-G1 distribution were determined by FACSanalysis. 2×106 cells were seeded in a 6 well plate and treated withdifferent concentrations of HKBA for 24 h. The cells were collected andfixed for 30 min in 70% ethanol at −20 °C. After fixation, the cellswere incubated with a staining solution [RNase A (0.1 mg/ml) and
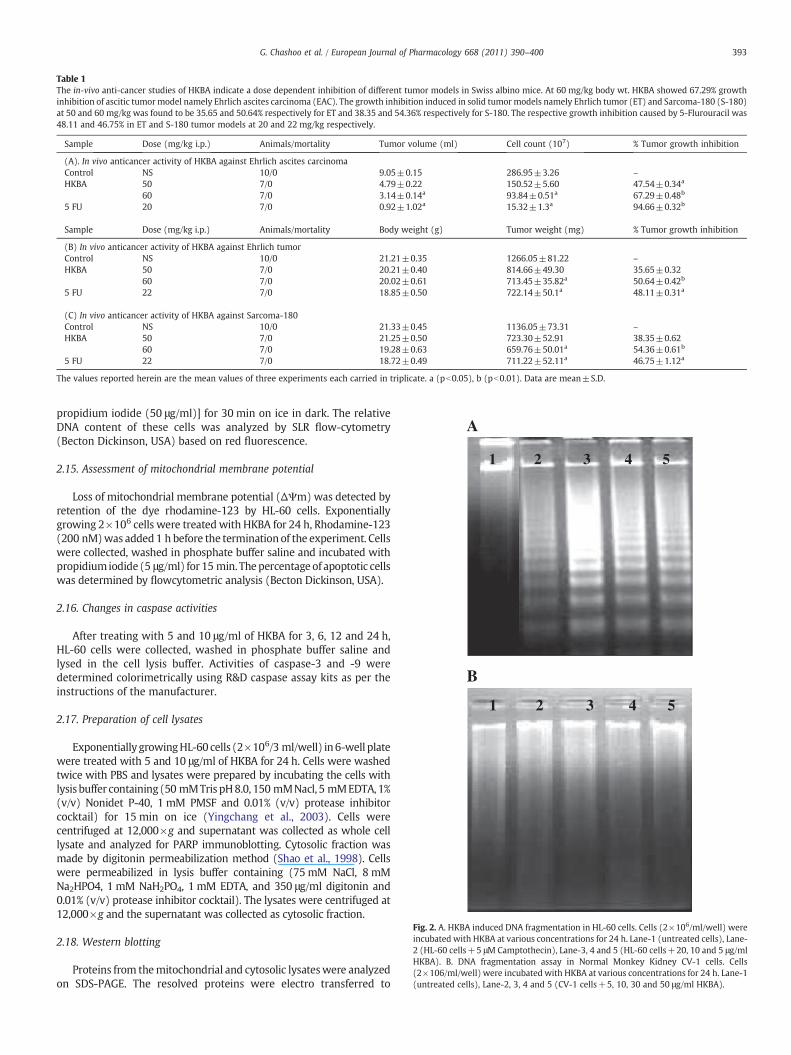
Table 1The in-vivo anti-cancer studies of HKBA indicate a dose dependent inhibition of different tumor models in Swiss albino mice. At 60 mg/kg body wt. HKBA showed 67.29% growthinhibition of ascitic tumormodel namely Ehrlich ascites carcinoma (EAC). The growth inhibition induced in solid tumor models namely Ehrlich tumor (ET) and Sarcoma-180 (S-180)at 50 and 60 mg/kg was found to be 35.65 and 50.64% respectively for ET and 38.35 and 54.36% respectively for S-180. The respective growth inhibition caused by 5-Flurouracil was48.11 and 46.75% in ET and S-180 tumor models at 20 and 22 mg/kg respectively.
Sample Dose (mg/kg i.p.) Animals/mortality Tumor volume (ml) Cell count (107) % Tumor growth inhibition
(A). In vivo anticancer activity of HKBA against Ehrlich ascites carcinomaControl NS 10/0 9.05±0.15 286.95±3.26 –
HKBA 50 7/0 4.79±0.22 150.52±5.60 47.54±0.34a
60 7/0 3.14±0.14a 93.84±0.51a 67.29±0.48b
5 FU 20 7/0 0.92±1.02a 15.32±1.3a 94.66±0.32b
Sample Dose (mg/kg i.p.) Animals/mortality Body weight (g) Tumor weight (mg) % Tumor growth inhibition
(B) In vivo anticancer activity of HKBA against Ehrlich tumorControl NS 10/0 21.21±0.35 1266.05±81.22 –
HKBA 50 7/0 20.21±0.40 814.66±49.30 35.65±0.3260 7/0 20.02±0.61 713.45±35.82a 50.64±0.42b
5 FU 22 7/0 18.85±0.50 722.14±50.1a 48.11±0.31a
(C) In vivo anticancer activity of HKBA against Sarcoma-180Control NS 10/0 21.33±0.45 1136.05±73.31 –
HKBA 50 7/0 21.25±0.50 723.30±52.91 38.35±0.6260 7/0 19.28±0.63 659.76±50.01a 54.36±0.61b
5 FU 22 7/0 18.72±0.49 711.22±52.11a 46.75±1.12a
The values reported herein are the mean values of three experiments each carried in triplicate. a (pb0.05), b (pb0.01). Data are mean±S.D.
1 2 3 4 5
1 2 3 4 5
B
A
Fig. 2. A. HKBA induced DNA fragmentation in HL-60 cells. Cells (2×106/ml/well) wereincubated with HKBA at various concentrations for 24 h. Lane-1 (untreated cells), Lane-2 (HL-60 cells+5 μM Camptothecin), Lane-3, 4 and 5 (HL-60 cells+20, 10 and 5 μg/mlHKBA). B. DNA fragmentation assay in Normal Monkey Kidney CV-1 cells. Cells(2×106/ml/well) were incubated with HKBA at various concentrations for 24 h. Lane-1(untreated cells), Lane-2, 3, 4 and 5 (CV-1 cells+5, 10, 30 and 50 μg/ml HKBA).
393G. Chashoo et al. / European Journal of Pharmacology 668 (2011) 390–400
propidium iodide (50 μg/ml)] for 30 min on ice in dark. The relativeDNA content of these cells was analyzed by SLR flow-cytometry(Becton Dickinson, USA) based on red fluorescence.
2.15. Assessment of mitochondrial membrane potential
Loss of mitochondrial membrane potential (ΔΨm) was detected byretention of the dye rhodamine-123 by HL-60 cells. Exponentiallygrowing 2×106 cells were treated with HKBA for 24 h, Rhodamine-123(200 nM)was added 1 h before the termination of the experiment. Cellswere collected, washed in phosphate buffer saline and incubated withpropidiumiodide (5 μg/ml) for 15 min. Thepercentage of apoptotic cellswas determined by flowcytometric analysis (Becton Dickinson, USA).
2.16. Changes in caspase activities
After treating with 5 and 10 μg/ml of HKBA for 3, 6, 12 and 24 h,HL-60 cells were collected, washed in phosphate buffer saline andlysed in the cell lysis buffer. Activities of caspase-3 and -9 weredetermined colorimetrically using R&D caspase assay kits as per theinstructions of the manufacturer.
2.17. Preparation of cell lysates
Exponentially growingHL-60 cells (2×106/3 ml/well) in 6-well platewere treated with 5 and 10 μg/ml of HKBA for 24 h. Cells were washedtwice with PBS and lysates were prepared by incubating the cells withlysis buffer containing(50 mMTris pH8.0, 150 mMNacl, 5 mMEDTA, 1%(v/v) Nonidet P-40, 1 mM PMSF and 0.01% (v/v) protease inhibitorcocktail) for 15 min on ice (Yingchang et al., 2003). Cells werecentrifuged at 12,000×g and supernatant was collected as whole celllysate and analyzed for PARP immunoblotting. Cytosolic fraction wasmade by digitonin permeabilization method (Shao et al., 1998). Cellswere permeabilized in lysis buffer containing (75 mM NaCl, 8 mMNa2HPO4, 1 mM NaH2PO4, 1 mM EDTA, and 350 μg/ml digitonin and0.01% (v/v) protease inhibitor cocktail). The lysates were centrifuged at12,000×g and the supernatant was collected as cytosolic fraction.
2.18. Western blotting
Proteins from themitochondrial and cytosolic lysateswere analyzedon SDS-PAGE. The resolved proteins were electro transferred to

C) HKBA 1 µg/ml, 24 h D) HKBA 5 µg/ml, 24 h
A) Untreated cells B) Camptothecin 5 µM
µM
Fig. 3.Nuclear morphology analysis of HKBA treated HL-60 cells (2×105/2 ml/6 well plate) using DAPI. (A) Untreated control cells (B) Cells treated with 5 μMCamptothecin indicatecondensation both in cytoplasm and nuclei (↑). (C) Cells treated with 1 μg/ml HKBA for 18 h indicate apoptosis (↑). (D) Cells treated with 5 μg/ml of HKBA for 18 h showing increasein apoptotic population (↑). Magnification 400×, Bar (―) 20 μM.
394 G. Chashoo et al. / European Journal of Pharmacology 668 (2011) 390–400
polyvinylidene difluoride (PVDF) membranes (Bio-RAD) over night at30 V, 4 °C. The membranes were blocked in blocking buffer (10 mMTris–HCl, 150 mMNaCl, 0.1% Tween-20) containing 5% non-fat milk for1 h and blotted with respective mouse anti-human primary antibodiesfor 2 h. Blots were washed three times in Tris Buffered Saline andTween-20 (TBST) andwere then incubatedwithhorseradishperoxidaseconjugatedmouse or rabbit secondary antibodies for 1 h, followed againby washing three times with TBST and signals detected using enhancedchemiluminescence (ECL) plus chemiluminescence's kit on X-ray film.
2.19. DNA topoisomerase I and II activity
Topoisomerse-I and II activity of HKBA was determined by HumanTopoisomerase-I and II kits from TopoGen (USA) as per the methoddescribed previously (Chashoo et al., 2011; Syrovets et al., 2007).
2.20. Statistical analysis
Data expressed as mean±S.D. unless otherwise indicated. Com-parisons were made between control and treated groups by ANOVA andp valuesb0.01 was considered significant
3. Results
3.1. In vitro anti proliferation activity in human leukemia cell lines
The cell growth inhibition of test compound in human leukemia celllines was tested by MTT assay as described earlier. HKBA inhibited thecell growth in dose dependant pattern and the IC50 value of 4.5 μg/ml inHL-60 and6 μg/ml inMolt-4 cancer cell lines after 48 h of treatmentwasobserved (Fig. 1). The IC50 values of HKBA were found to be lower thanthat of BKBA (HL-60, 7.7 μg/ml and Molt-4, 9 μg/ml) and previouslyreported PKBA (HL-60, 8.7 μg/ml and Molt-4, 9.5 μg/ml).
3.2. In vivo antitumor activity
In vivo anticancer studies of HKBA revealed that tumor growthwasinhibited dose dependently in themice bearing ascitic (EAC) and solid(EAT and S-180) tumors. At intraperitoneal doses of 50 and 60 mg/kgb.wt. the growth of Ehrlich ascitic carcinoma (EAC) was inhibited by47.54% and 67.29% respectively. In case of solid tumors, HKBA showeda growth inhibition of 35.65% and 50.64% at 50 and 60 mg/kg b.wt.respectively in EAT model and 38.35% and 54.36% at the respectivedoses in S-180 model. PKBA on the other hand was found to show thesame effect in both ascitic and solid tumor models at 150 mg/kg b.wt.(Chashoo et al., 2011). Tumor growth inhibition by 5-FU used aspositive control was found to be 48.11% and 46.75% at 22 mg/kg b.wt.in ET and S-180 respectively (Table 1).
3.3. HKBA induces DNA fragmentation hallmark of apoptosis
Induction of DNA fragmentation by HKBA treatment was investi-gated by isolating genomic DNA from HL-60 cells. Cells treated withtest compound exhibited typical DNA ladder formation at 5 and10 μg/ml after 24 h (Fig. 2A). However no ladder pattern was observedinnormalmonkeykidneyCV-1 cell line treatedwithHKBA(5, 10, 30and50 μg/ml) for 24 h (Fig. 2B).
3.4. Nuclear morphologic features
HKBA influences the nuclear morphology and leads to theformation of apoptotic bodies in HL-60 cells. Cells (2×105/2 ml/6well plate) were treated with 1 and 5 μg/ml of HKBA for 18 h, stainedwith DAPI and visualized for nuclearmorphology and apoptotic bodiesusing fluorescence microscope. Nuclei of untreated cells appearedround in shape (Fig. 3A), while treatment with HKBA resulted innuclear condensation and formation of apoptotic bodies (Fig. 3C, D).Camptothecin (Fig. 3B), taken as a positive control also indicatedcondensation both in cytoplasm and nuclei. The morphological
A) Untreated cells (4000x)
B) Untreated cells (8000x)
D) HKBA 5 µg/ml (4000x)
E) HKBA 5 µg/ml (8000x)
C) Untreated cells (10,000x) F) HKBA 5 µg/ml (10,000x)
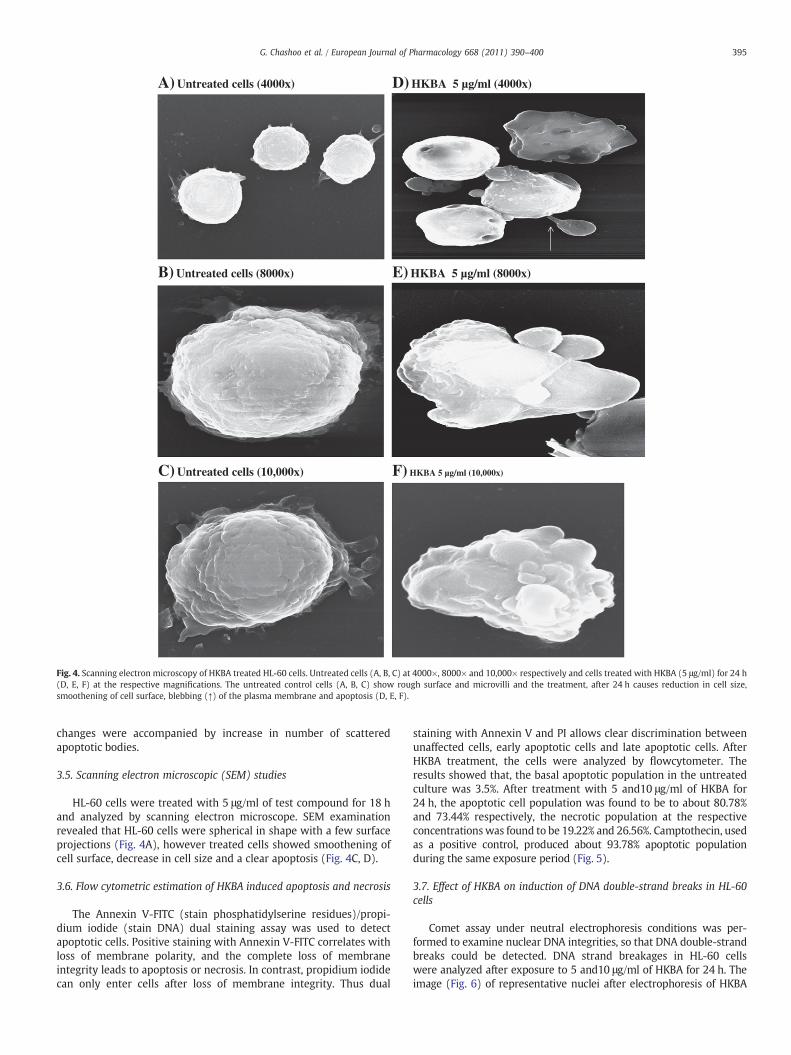
Fig. 4. Scanning electron microscopy of HKBA treated HL-60 cells. Untreated cells (A, B, C) at 4000×, 8000× and 10,000× respectively and cells treated with HKBA (5 μg/ml) for 24 h(D, E, F) at the respective magnifications. The untreated control cells (A, B, C) show rough surface and microvilli and the treatment, after 24 h causes reduction in cell size,smoothening of cell surface, blebbing (↑) of the plasma membrane and apoptosis (D, E, F).
395G. Chashoo et al. / European Journal of Pharmacology 668 (2011) 390–400
changes were accompanied by increase in number of scatteredapoptotic bodies.
3.5. Scanning electron microscopic (SEM) studies
HL-60 cells were treated with 5 μg/ml of test compound for 18 hand analyzed by scanning electron microscope. SEM examinationrevealed that HL-60 cells were spherical in shape with a few surfaceprojections (Fig. 4A), however treated cells showed smoothening ofcell surface, decrease in cell size and a clear apoptosis (Fig. 4C, D).
3.6. Flow cytometric estimation of HKBA induced apoptosis and necrosis
The Annexin V-FITC (stain phosphatidylserine residues)/propi-dium iodide (stain DNA) dual staining assay was used to detectapoptotic cells. Positive staining with Annexin V-FITC correlates withloss of membrane polarity, and the complete loss of membraneintegrity leads to apoptosis or necrosis. In contrast, propidium iodidecan only enter cells after loss of membrane integrity. Thus dual
staining with Annexin V and PI allows clear discrimination betweenunaffected cells, early apoptotic cells and late apoptotic cells. AfterHKBA treatment, the cells were analyzed by flowcytometer. Theresults showed that, the basal apoptotic population in the untreatedculture was 3.5%. After treatment with 5 and10 μg/ml of HKBA for24 h, the apoptotic cell population was found to be to about 80.78%and 73.44% respectively, the necrotic population at the respectiveconcentrationswas found to be 19.22% and 26.56%. Camptothecin, usedas a positive control, produced about 93.78% apoptotic populationduring the same exposure period (Fig. 5).
3.7. Effect of HKBA on induction of DNA double-strand breaks in HL-60cells
Comet assay under neutral electrophoresis conditions was per-formed to examine nuclear DNA integrities, so that DNA double-strandbreaks could be detected. DNA strand breakages in HL-60 cellswere analyzed after exposure to 5 and10 μg/ml of HKBA for 24 h. Theimage (Fig. 6) of representative nuclei after electrophoresis of HKBA

A) Untreated Cells B) Camptothecin 5 µM
C) HKBA 5 µg/ml D) HKBA 10 µg/ml
3.50 %
5.89 %
93.78 %
80.78 %
19.22 % 26.56 %
73.44 %
Fig. 5. Annexin V-FITC/PI staining of HKBA treated HL-60 cells. Cells were treated with indicated concentrations of HKBA for 24 h and stained with Annexin V-FITC/PI to analyzeapoptotic and necrotic cell population as described earlier. Quadrants analysis of fluorescence intensity of ungated cells in FL-1 versus FL-2 channels was from 10,000 events. Cells inthe lower right quadrant represented apoptosis while cells in the upper right quadrant represented necrosis or post apoptotic necrosis Data are representative of one of the twosimilar experiments.
396 G. Chashoo et al. / European Journal of Pharmacology 668 (2011) 390–400
(10 μg/ml) treated cells showed formation of typical comets, with HeadDNA of 68.22% and 23.42% at 5 and 10 μg/ml respectively. Tail DNAwhich reveals the actual DNA damage was found to be 51.78% and66.58% at the respective concentrations. The Olive Tail movement was6.06% and 9.56% and Tail length 18.33% and 36.21% respectively(Table 2). According to the literature (Shao et al., 1998; Yasuharaet al., 2003), a cell containing more than 50% of the total DNA in thecomet tail is defined as an apoptotic cell.
3.8. Effect of HKBA on cell cycle distribution in HL-60 cells
An effective strategy to inhibit tumor growth is deregulated cellcycle progression in cancer cells. Effects of HKBA on cell cycle
A) Untreated cells
B) Camptothecin 5 µM
C) HKBA 10 µg/ml
Fig. 6. Comet formation in HKBA treated HL-60 cells. (A) Untreated cells (B)Camptothecin taken as a positive control. (C) HKBA treated HL-60 cells after 24 h ofincubation.
progression in HL-60 cells were then examined. Cells were treatedwith test compound at a dosage of 1, 5, and 10 μg/ml for 24 h andFluorescence Activated Cell Sorting (FACS) analysis was followed. TheDNA histogram showed that HKBA induced dose dependent increasein hypo diploid sub-G1 DNA fraction (b2n DNA) (Fig. 7). The sub-G1DNA fraction was 3.76% in untreated cells however after treatmentwith test compound (10 μg/ml) it increased to about 88.13% (Table 3).Further, HKBA was found to arrest the G2/M phase of cell cycle.
3.9. Loss of mitochondrial membrane potential by HKBA
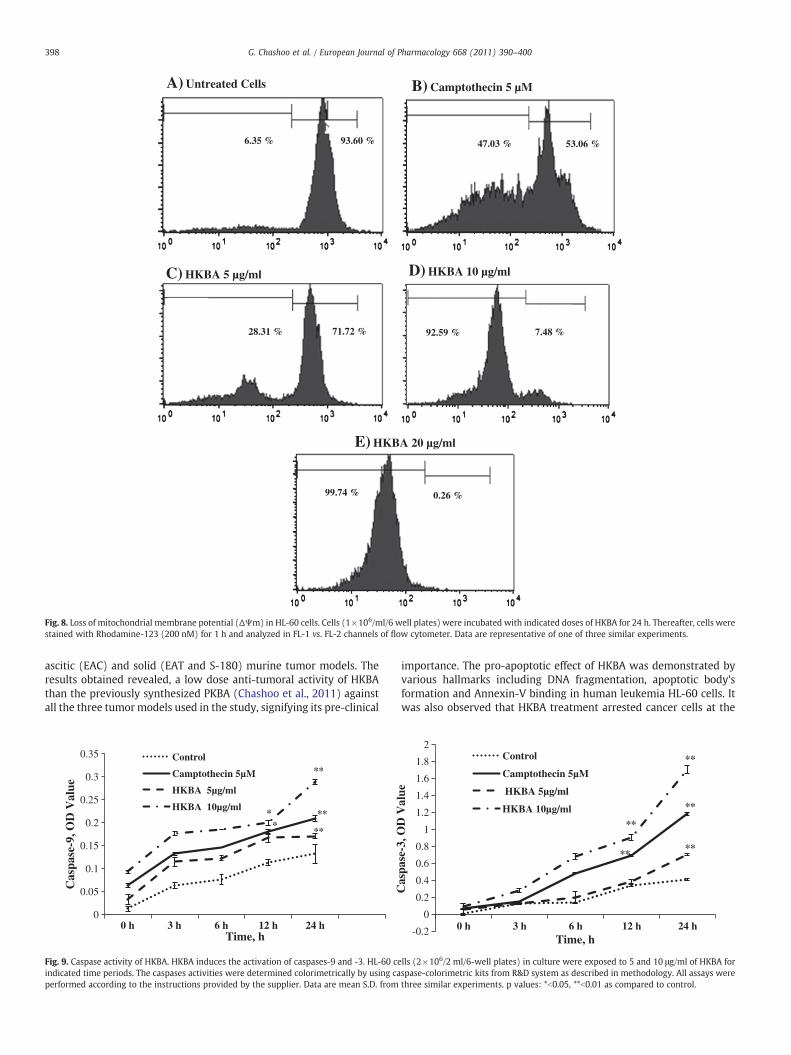
After rhodamine-123 incubation the fluorescent intensity, anindicator for mitochondrial membrane potential was reduced from93.60% to 0.26% in HKBA (10 μg/ml) treated cells for 24 h, consistentwith the results that HL-60 cell apoptosis occurred 24 h aftertreatment with HKBA at the same concentration (Fig. 8).
3.10. HKBA stimulates caspase activities in HL-60 cells
HL-60 cells were exposed to HKBA and evaluated for caspase-3 and-9 activities. The test compound predominately induced a dose and
Table 2The single cell gel electrophoresis of HKBA.
Head DNA Tail DNA Olive Tailmovement
Tail length
Untreated Cells 90.91±0.011 9.09±0.031 0.501±0.04 1.34±0.011Camptothecin 5 μM 60.06±0.02 39.94±0.015 8.84±0.022 35.13±0.042HKBA 5 μg/ml 68.22±0.02 51.78±0.031 6.06±0.011 18.33±0.021HKBA 10 μg/ml 23.42±0.04 66.58±0.033 12.33±0.042 36.21±0.036
Comet assay analysis of HKBA treated HL-60 cells after 24 h of incubation revealed ahead DNA of 68.22% and 23.42%, Tail DNA of 51.78% and 66.58% at 5 and 10 μg/mlrespectively which signifies the DNA damage. The Olive Tail movement was 6.06% and12.33% while a Tail length of 18.33% and 36.21% was observed at the respectiveconcentrations. Camptothecin taken as a positive control revealed a Head DNA of60.06% and a tail DNA of 39.94%. Data are mean S.D. from three similar experiments.

Fig. 7. Cell cycle analysis of HKBA treated HL-60 cells. HL-60 Cells (2×106 cells/ml/well) were exposed to indicated concentrations of HKBA for 24 h and stained with PI to determineDNA fluorescence and cell cycle phase distribution as described in materials and methods. Fraction of cells for sub-G1 population analyzed from FL2-A vs. cell counts is shown (%).Data are representative of one of three similar experiments.
397G. Chashoo et al. / European Journal of Pharmacology 668 (2011) 390–400
time dependant increase in caspase-3 and -9 activities in HL-60 cells,registering a ten-fold increase at the highest concentration of HKBAused (Fig. 9). In the light of HKBA induction of apoptosis these findingsamply demonstrate the role of caspase-9 activation in up regulatingdownstream events leading to caspase-3 activation (Craen et al.,1999; Slee et al., 1999).
3.11. HKBA induces release of cytochrome c from mitochondria andCleavage of PARP in HL-60 cells
HKBA induced loss of mitochondrial membrane potential in HL-60cells was found to induce the release of cytochrome c frommitochondrion into the cytosol. The increased expression of cyto-chrome c in the cytosol was observed in a concentration dependantmanner (Fig. 10). In the further consequence of events the releasedcytochrome c activated caspase-9 which in turn activates caspase-3.Caspase-3 as a matter of fact uses another DNA repair enzyme, poly(ADP ribose) polymerase (PARP) as a substrate and inhibits the repairof damaged DNA. Therefore a cleaved product of PARP was observedin our study (Fig. 10). These studies thus confirm that the induction ofapoptosis by HKBA was due to the activation of intrinsic pathway.
Table 3The cell cycle phase distribution of HKBA in HL-60 cells.
Sample Sub G1%/(apoptotic) G1/G0 S G2/M
Untreated cells 3.76% 47.42 23.44 28.61Camptothecin 5 μM 56.08 32.17 6.93 5.76HKBA 1 μg/ml 3.0% 51.42 23.34 29.21HKBA 5 μg/ml 74.33% 12.68 17.21 4.43HKBA 10 μg/ml 88.13% 9.63 2.11 0.89
3.12. Topoisomerase I and II inhibitory assay
Topoisomerases (I and II) are the essential enzymes for prolifer-ation of eukaryotic cells (Nakagawa et al., 2006; Osheroff et al., 1983).Drugs that target topoisomerases are among the most effectiveanticancer drugs. In our previous studies we found that PKBAinhibited the enzymatic activity of both the topoisomerases I and IIat 20 μg/ml. The present study showed that HKBA was also able toinhibit the enzymatic activity of topoisomerases I and II, at lowerconcentration of 10 μg/ml (Fig. 11). The inhibitory activity of HKBAwas compared with the reference molecules camptothecin andetoposide for topoisomerase I and II respectively.
4. Discussion
Our previous data had shown that propionyloxy-11-keto-β-boswellic acid (PKBA) induced apoptosis in HL-60 cells at lowerconcentrations than the naturally existing acetyl-11-keto-β-boswellicacid (AKBA) (Chashoo et al., 2011). Since the earlier data revealed theimportance of an acyl group at the 3-α-hydroxy position on theanticancer potential of KBA, it was therefore envisaged that novelanalogs of KBA with increasing acyl chain length at 3-α-hydroxyposition will be synthesized and subjected to anticancer studies. Inthis communication, butyryloxy (BKBA) and hexanoyloxy (HKBA)derivatives of KBA were synthesized. The antiproliferative potential ofthese analogs was evaluated in human leukemia HL-60 and MOLT-4cell lines and it was observed that HKBA showed lower IC50 valuesthan BKBA and previously reported PKBA as well. These observationsprompted us to carry out detailed anticancer studies to evaluate the invivo antitumor and pro-apoptotic potential of this newly synthesizedanalog. The in vivo anticancer studies of HKBA were performed in
A) Untreated Cells B) Camptothecin 5 µM
C) HKBA 5 µg/ml D) HKBA 10 µg/ml
6.35 % 93.60 % 47.03 % 53.06 %
28.31 % 71.72 % 92.59 % 7.48 %
E) HKBA 20 µg/ml
99.74 % 0.26 %
Fig. 8. Loss of mitochondrial membrane potential (ΔΨm) in HL-60 cells. Cells (1×106/ml/6 well plates) were incubated with indicated doses of HKBA for 24 h. Thereafter, cells werestained with Rhodamine-123 (200 nM) for 1 h and analyzed in FL-1 vs. FL-2 channels of flow cytometer. Data are representative of one of three similar experiments.
398 G. Chashoo et al. / European Journal of Pharmacology 668 (2011) 390–400
ascitic (EAC) and solid (EAT and S-180) murine tumor models. Theresults obtained revealed, a low dose anti-tumoral activity of HKBAthan the previously synthesized PKBA (Chashoo et al., 2011) againstall the three tumormodels used in the study, signifying its pre-clinical
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0 h 3 h 6 h 12 h 24 h
Cas
pase
-9, O
D V
alue
Time, h
Control
Camptothecin 5µM
HKBA 5µg/ml
HKBA 10µg/ml**
***
*
**
Fig. 9. Caspase activity of HKBA. HKBA induces the activation of caspases-9 and -3. HL-60 ceindicated time periods. The caspases activities were determined colorimetrically by using caperformed according to the instructions provided by the supplier. Data are mean S.D. from
importance. The pro-apoptotic effect of HKBA was demonstrated byvarious hallmarks including DNA fragmentation, apoptotic body'sformation and Annexin-V binding in human leukemia HL-60 cells. Itwas also observed that HKBA treatment arrested cancer cells at the
-0.2
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
0 h 3 h 6 h 12 h 24 h
Cas
pase
-3, O
D V
alue
Time, h
Control
Camptothecin 5µM
HKBA 5µg/ml
HKBA 10µg/ml
****
**
**
**
lls (2×106/2 ml/6-well plates) in culture were exposed to 5 and 10 μg/ml of HKBA forspase-colorimetric kits from R&D system as described in methodology. All assays werethree similar experiments. p values: *b0.05, **b0.01 as compared to control.

Fig. 10. HKBA influences the expression of cytochrome c and PARP in HL-60 cells. Cells (2×106/3 ml/6-well plates) were treated with 5 and 10 μg/ml of HKBA for 24 h and differentfractions were prepared and immunoblotted as described in material and methods. β-Actin used as internal control to represent the same amount of proteins applied for SDS-PAGE.Specific antibodies were used for detection of cytochrome c and PARP. Data are representative of one of three similar experiments.
399G. Chashoo et al. / European Journal of Pharmacology 668 (2011) 390–400
G2/M phase and showed significant increment in the Sub G1 phase ofthe cell cycle. The literature precedent and the kinetic data availablehave indicated that mitochondrion undergoes major changes inmembrane integrity before classical signs of apoptosis becomemanifest (Gruber et al., 2004). It is widely accepted that thealterations of mitochondrial function, such as breakdown of themitochondrial transmembrane potential (Ψm) and release of proa-poptotic cytochrome c, constitute key events of the mitochondrialapoptosis signaling cascade (Susin et al., 1998). In the current study,HKBA was found to induce changes in the mitochondrial transmem-brane potential which was found to be associated with the release ofcytochrome c from mitochondria into the cytosol. The releasedcytochrome c from the mitochondrion resulted in the upregulation ofthe initiator caspase-9 which in turn activated the executionercaspase-3. Once activated, caspase-3 induced cleavage of poly (ADP-ribose) polymerase (PARP) thus prevented any unwanted repair ofthe damaged DNA. Moreover, the cleavage of PARP is consistent withDNA fragmentation, which results in the morphologic and biochem-ical features of apoptosis. These findings therefore indicate thatmitochondrial signaling pathway participate in the HKBA inducedapoptosis. Although, boswellic acids have repeatedly been reported toinduce apoptosis in cancer cells, however, the results obtained fromthe current study demonstrated the low dose apoptotic potential of anewly synthesized analog, HKBA. Since our previous data showedthat, PKBA induced cancer cells to death by inhibiting the enzymaticactivity of topoisomerases I and II, HKBA was therefore subjected totopoisomerase inhibitory studies, where it was found to inhibit thecatalytic activity of both topoisomerases I and II at low doses. Severalreports have revealed that chemotherapeutic agents that targettopoisomerases I and II set in motion a series of biochemical changesthat culminate in cell death (Bielawski et al., 2006). These antitumordrugs interact with both topoisomerases I and II simultaneously. Thismechanism of action appears to be advantageous, because selectiveinhibition of topoisomerase I has been reported to increase topo-isomerase II enzyme activity and vice-versa, which may be importantfor the development of drug resistance. In this regard, a singlecompound able to inhibit both topoisomerase I and II may present theadvantage of improving anti topoisomerase activity, with reduced
1 2 3 4 5 6 7
Fig. 11. HKBA inhibits topoisomerase-I and II activity. Inhibitory assay of topoisomerase-Iand II was performed by using topoisomerase-I and II kits from Topogen. Lane 1 (PBR322DNA), Lane2 (PBR322DNA+Topoisomerase-II), Lane3(PBR322DNA+Topoisomerase-II+Etoposide 100 μM), Lane 4 (PBR322 DNA+Topoisomerase-II+HKBA 10 μg/ml), Lane 5(PBR322 DNA+Topoisomerase-I), Lane 6 (PBR322 DNA+Topoisomerase-I+Camptothecin 100 μM) and Lane 7 (PBR322 DNA+Topoisomerase-I+HKBA 10 μg/ml).
toxic side effects, with respect to the combination of two inhibitors(Salerno et al., 2010). Although, dual inhibitors theoretically over-come these problems, however, some of the inhibitors like Intoplicinand XR11576 have been found to be associated with liver toxicity,diarrhea, nausea, vomiting and alopecia (De Jonge et al., 2004; Salernoet al., 2010). Besides this, the chemopreventive effects of well knowntopoisomerase targeting drugs such as camptothecin and etoposidehave also been found to be associated with multiple and severe sideeffects such as myelosuppression, thrombocytopenia, anemia, bonemarrow and gastrointestinal toxicity (Hartman and Lipp, 2006).Boswellic acids in contrast to this are known to have good safetyprofile showing no severe side effects (Gupta et al., 1997;Mantle et al.,2001; Safayhi et al., 2000). In addition, we reported recently that PKBAwas non-toxic to the normal cells, the present investigation alsoshowed that in spite of the increased anticancer potency at lowerdoses, HKBA seems to be selective for tumor cells, since no apoptosiswas observed in normal cells. In summary, this paper constitutes thefirst instance for the demonstration of anticancer activity of a newlysynthesized 3-α-hexanoyloxy derivative of KBA, HKBA. The abovedata revealed that, compared to the previously reported boswellicacids, HKBA showed a low dose antiproliferative and pro-apoptoticpotential in human leukemia HL-60 cells. Besides this, a low doseantitumor potential in ascitic and solid murine tumormodels was alsoobserved. Keeping all this into consideration, whether the anticancereffects HKBA at low doses characterized here have any clinicalrelevance, remains to be determined. Further, pharmacokineticstudies should address this problem.
5. Conclusion
The current study leads us to the conclusion that HKBA has a betterantiproliferative and proapoptotic potential against HL-60 cell linethan the previously known boswellic acids. Furthermore, it has beenfound that HKBA showed a low dose anti-tumoral activity againstmice tumor models. Owing to its potent pro-apoptotic and antitumoractivity, HKBA deserves to be investigated further for its pharmaco-kinetic profile which will indeed clarify its therapeutic index.
Acknowledgment
The authors are thankful to Prof. Qazi Naveed Ahmad for hisassistance in writing this manuscript. They also express their sincerethanks to Council of Scientific and Industrial Research, India, forproviding senior research fellowship to Gousia Chashoo.
References
Bhushan, S., Singh, J., Rao, M.J., Saxena, A.K., Qazi, G.N., 2006. A novel lignan compositionfrom Cedrus deodara induces apoptosis and early nitric oxide generation in humanleukemia Molt-4 and HL-60 cells. Nitric oxide 14, 72–88.
Bielawski, K., Winnicka, K., Bielawska, A., 2006. Inhibition of DNA topoisomerases I andII, and growth inhibition of breast cancer MCF-7 cells by ouabain, digoxin andproscillaridin A. Biol. Pharm. Bull. 29, 1493–1497.
Chashoo, G., Singh, S.K., Sharma, P.R., Mondhe, D.M., Hamid, A., Andotra, S.S., Shah, B.A.,Qazi, N.A., Taneja, S.C., Saxena, A.K., 2011. A Propionyloxy derivative of 11-keto-β-

400 G. Chashoo et al. / European Journal of Pharmacology 668 (2011) 390–400
boswellic acid induces apoptosis in HL-60 cells mediated through topoisomerase I& II inhibition. Chem. Biol. Interact. 189, 60–71.
Craen, V.D., Declercq, W.M., Brande, I.V.D., Fiers, W., Vandenabeele, P., 1999. Theproteolytic procaspase activation network, an in vitro analysis. Cell Death Differ. 6,1117–1124.
De Jonge, M.J.A., Kaye, S., Verweij, J., Brock, C., Reade, S., Scurr, M., van Doorn, L., Verheij,C., Loos, W., Brindley, C., Mistry, P., Cooper, M., Judson, I., 2004. Phase I andpharmacokinetic study of XR11576, an oral topoisomerase I and II inhibitor,administered on days 1–5 of a 3-weekly cycle in patients with advanced solidtumours. Br. J. Cancer 91, 1459–1465.
Geran, R.I., Greenberg, N.H., MacDonald, M.M., Schumacher, A.M., Abbott, B.J., 1972.Protocols for screening chemical agents and natural products against animaltumors and other biological systems. Cancer Chemother. Rep. 3, 1–103.
Glaser, T., Winter, S., Groscurth, P., Safayhi, H., Sailer, E.R., Ammon, H.P., Schabet, M.,Weller, M., 1999. Boswellic acids and malignant glioma: induction of apoptosis butno modulation of drug sensitivity. Br. J. Cancer 80, 756–765.
Gruber, C., Henkel, M., Budach, W., Belka, C., Jendrossek, V., 2004. Involvement oftyrosine kinase p56/Lck in apoptosis induction by anticancer drugs. Biochem.Pharmacol. 67, 1859–1872.
Gupta, I., Parihar, A., Malhotra, P., Singh, G.B., Luettke, R., Safayhi, H., Ammon, H.P., 1997.Effect of Boswellia serrata gum resin in patients with ulcerative colitis. Eur. J. Med.Res. 2, 1–7.
Han, R., 1994. Highlight on the studies of anticancer drugs derived from plants in China.Stem Cells 12, 53–63.
Hartman, J.T., Lipp, H.P., 2006. Camptothecin and podophyllotoxin derivatives:inhibitors of topoisomerase I and II — mechanisms of action, pharmacokineticsand toxicity profile. Drug Saf. 29, 209–230.
Hoernlein, R.F., Orlikowsky, T.H., Zehrer, C., 1999. Acetyl-11-keto-beta-boswellicacid induce apoptosis in HL-60 and CCRFCEM cells and inhibit topoisomerase-I.J. Pharmacol. Exp. Ther. 288, 613–619.
Jing,Y.,Nakajo, S., Xia, L., Nakaya,K., Fang,Q.,Waxman, S.,Han,R., 1999.Boswellic acidacetateinduces differentiation and apoptosis in leukemia cell lines. Leuk. Res. 23, 43–50.
Liu, J.J., Nilsson, A., Oredsson, S., Badmaev, V., Duan, R.D., 2002. Keto- and acetyl-keto-boswellic acids inhibit proliferation and induce apoptosis in Hep G2 cells via acaspase-8 dependent pathway. Int. J. Mol. Med. 10, 501–505.
Mantle, D., Gok, M.A., Lennard, T.W., 2001. Adverse and beneficial effect of plantextracts on skin and skin disorders. Adverse Drug React. Toxicol. Rev. 20, 89–103.
Nakagawa, H., Saito, H., Ikegami, Y., Aida-Hyugaji, S., Sawada, S., Ishikawa, T., 2006.Molecular modeling of new camptothecin analogues to circumvent ABCG2-mediated drug resistance in cancer. Cancer Lett. 234, 81–89.
Osheroff, N., Shelton, E.R., Brutlag, D.L., 1983. DNA topoisomerase II from Drosophillamelanogaster: relaxation of supercoiled DNA. J. Biol. Chem. 258, 9536–9543.
Park, Y.S., Lee, J.H., Harwalkar, J.A., Bondar, J., Safayhi, H., Golubic, M., 2002. Acetyl-11-keto-beta-boswellic acid (AKBA) is cytotoxic for meningioma cells and inhibits
phosphorylation of the extracellular-signal regulated kinase 1 and 2. Adv. Exp.Med. Biol. 507, 387–393.
Rello, S., Stockert, J.C., Moreno, V., Gámez, A., Pacheco, M., Juarranz, A., Cañete, M.,Villanueva, A., 2005. Morphological criteria to distinguish cell death induced byapoptotic and necrotic treatments. Apoptosis 10, 201–208.
Safayhi, H., Boden, S.E., Schweizer, S., Ammon, H.P.T., 2000. Concentration dependentpotentiating and inhibitory effects of Boswellia extract on 5- LOX productformation in stimulated PMNLs. Planta Med. 66, 110–113.
Salerno, S., Da Settimo, F., Taliani, S., Simorini, F., La Motta, C., Fornaciari, G., Marini,A.M., 2010. Recent advances in the development of dual topoisomerase I and IIinhibitors as anticancer drugs. Curr. Med. Chem. 17, 4270–4290.
Shah, B.A., Qazi, G.N., Taneja, S.C., 1997. Boswellic acids: a group of medicinallyimportant compounds. Nat. Prod. Rep. 26, 72–89 (and the references therein).
Shah, B.A., Kumar, A., Gupta, P., Sharma, M., Sethi, V.K., Saxena, A.K., Singh, J., Qazi, G.N.,Taneja, S.C., 2007. Cytotoxic and apoptotic activities of novel amino analogues ofboswellic acids. BMCL 17, 6411–6416.
Shao, Y., Ho, C.T., Chin, C.K., Badmaev, V., Ma, W., Huang, M.T., 1998. Inhibitory activityof boswellic acids from Boswellia serrata against human leukemia HL-60 cells inculture. Planta Med. 64, 328–331.
Slee, E.A., Harte, M.T., Kluck, R.M., Wolf, B.B., Casiano, C.A., Newmeyer, D.D., Wang, H.G.,Reed, J.C., Nicholson, D.W., Alnemri, A.S., Green, D.R., Martin, S.J., 1999. Ordering thecytochrome c-initiated caspase cascade, Hierarchical activation of caspases-2, -3,-6, -7, -8 and -10 in a caspase-9 dependent manner. J. Cell Biol. 144, 281–292.
Susin, S.A., Zamzami, N., Kroemer, G., 1998. Mitochondria as regulators of apoptosis:doubt no more. Biophys. Acta 1366, 151–165.
Syrovets, T., Büchele, B., Gedig, E., Slupsky, J.R., Simmet, T., 2007. Acetyl-Boswellic acidsare novel catalytic inhibitors of human topoisomerases I and II. Mol. Pharmacol. 58,71–81.
Waxman, D.J., Schwartz, P.S., 2003. Harnessing apoptosis for improved anti-cancer genetherapy. Cancer Res. 63, 8563–8572.
Winking, M., Sarikaya, S., Rahmanian, A., Jodicke, A., Boker, D.K., 2000. Boswellic acidsinhibit glioma growth: a new treatment option. J. Neurooncol. 46, 97–103.
Yasuhara, S., Zhu, Y., Matsui, T., Tipirneni, N., Yasuhara, Y., Kaneki, M., Rosenzweig, A.,Martyn, J.A.J., 2003. Comparison of comet assay, electron microscopy, and flowcytometry for detection of apoptosis. J. Histochem. Cytochem. 51, 873–885.
Yingchang, M., Thomas, S.D., Xiaohua, X., Casson, L.K., Miller, D.M., Bates, P.J., 2003.Apoptosis in leukemia cell is accompanied by alterations in the level andlocalization of nucleolin. J. Biol. Chem. 278, 8572–8579.
Zhuang, S., Demirs, J.T., Kochevar, I.E., 2000. p38 mitogen-activated protein kinasemediates bid cleavage, mitochondrial dysfunction, and caspase-3 activation duringapoptosis induced by singlet oxygen but not by hydrogen peroxide. J. Biol. Chem.275, 25939–25948.




















