
Posttranslational regulation of NF-YA modulates NF-Y transcriptional activity
11
Molecular Biology of the Cell Vol. 19, 5203–5213, December 2008 Posttranslational Regulation of NF-YA Modulates NF-Y Transcriptional Activity Isabella Manni,* Giuseppina Caretti, † Simona Artuso,* Aymone Gurtner,* Velia Emiliozzi,* Ada Sacchi,* Roberto Mantovani, † and Giulia Piaggio* ‡ *Molecular Oncogenesis Laboratory and ‡ Rome Oncogenomic Center, Experimental Oncology Department, Regina Elena Cancer Institute, 00158 Rome, Italy; and † Biomolecular and Biotechnology Science Department, University of Milano, 20133 Milano, Italy Submitted April 6, 2008; Revised September 4, 2008; Accepted September 17, 2008 Monitoring Editor: Richard K. Assoian NF-Y binds to CCAAT motifs in the promoter region of a variety of genes involved in cell cycle progression. The NF-Y complex comprises three subunits, NF-YA, -YB, and -YC, all required for DNA binding. Expression of NF-YA fluctuates during the cell cycle and is down-regulated in postmitotic cells, indicating its role as the regulatory subunit of the complex. Control of NF-YA accumulation is posttranscriptional, NF-YA mRNA being relatively constant. Here we show that the levels of NF-YA protein are regulated posttranslationally by ubiquitylation and acetylation. A NF-YA protein carrying four mutated lysines in the C-terminal domain is more stable than the wild-type form, indicating that these lysines are ubiquitylated Two of the lysines are acetylated in vitro by p300, suggesting a competition between ubiqui- tylation and acetylation of overlapping residues. Interestingly, overexpression of a degradation-resistant NF-YA protein leads to sustained expression of mitotic cyclin complexes and increased cell proliferation, indicating that a tight regulation of NF-YA levels contributes to regulate NF-Y activity. INTRODUCTION The CCAAT-binding transcription factor NF-Y is a heteromeric protein composed of three subunits, NF-YA, -YB, and -YC, all necessary for CCAAT binding (Mantovani, 1999). The NF-Y complex supports the basal transcription of a class of regula- tory genes responsible for cell cycle progression, among which are mitotic cyclin complexes (Zwicker et al., 1995a,b; Bolognese et al., 1999; Farina et al., 1999; Korner et al., 2001; Gurtner et al., 2003, 2008; Di Agostino et al., 2006). Knock-out mice clearly demonstrates that NF-Y dependent transcription is essential during early mouse development (Bhattacharya et al., 2003). The NF-Y function in proliferation is also demonstrated by the inhibition of DNA binding by endogenous NF-Y, result- ing in retardation of fibroblast growth, which is brought about by overexpression of a dominant negative mutant of the NF-YA subunit (Hu and Maity, 2000). The differential expression of NF-YA, altering NF-Y CCAAT-binding activ- ity, has been observed in several cell lines and tissues both during cell cycle progression and under specific conditions. NF-YA expression is modulated during the cell cycle, being high in G1, further increasing in S, and then decreasing in the G2/M phase (Bolognese et al., 1999). Reduction of NF-YA expression has been reported in IMR-90 fibroblasts after serum deprivation (Chang et al., 1994) and in human monocytes (Marziali et al., 1997). The NF-YA protein is not expressed in terminally differentiated C2C12 muscle cells and adult skeletal muscle tissues (Farina et al., 1999; Gurtner et al., 2003, 2008). These observations show that levels of NF-YA dictate the function of the NF-Y complex. Interest- ingly, control of NF-YA accumulation is posttranscriptional, because NF-YA mRNA is relatively constant in growing and differentiated cells (Bolognese et al., 1999; Farina et al., 1999). Yet, the mechanisms regulating the stability of the NF-YA protein remain unknown. The covalent addition of ubiquitin, a 76-amino acid protein highly conserved among eukaryotes, is an increasingly com- mon posttranslation modification that controls both the expres- sion and the activity of numerous proteins in the eukaryotic cell (Hershko and Ciechanover, 1998; Ciechanover et al., 2000). The ubiquitin–proteasome pathway is composed of the ubiq- uitin-conjugating system and the 26S proteasome; the latter contains the multicatalytic protease complex (Hochstrasser, 1996; Hershko and Ciechanover, 1998; Thrower et al., 2000). Frequent targets of the ubiquitin modification machinery are transcription factors (Shcherbik and Haines, 2004). Indeed, many short-lived transcription factors such as E2F-1 (Hofmann et al., 1996; Hateboer et al., 1996), IkB (Chen et al., 1996), p53 (Scheffner et al., 1990), SMAD2 (Lo and Massague ´, 1999; Zhu et al., 1999), c-Jun (Treier et al., 1994), and -catenin (Aberle et al., 1997; Orford et al., 1997; Puca et al., 2008) are regulated by ubiquitylation. Most of them are unstable proteins whose rate of destruction mirrors their ability to activate transcription. The exact nature of how activation and destruction are linked is not yet clear (Collins and Tansey, 2006; Kodadek et al., 2006). Acetylation is a dynamic posttranslational modification of lysine residues. Proteins with intrinsic histone acetyltrans- ferase (HAT) activity act as transcriptional coactivators by acetylating histones and thereby induce an open chromatin conformation, which allows the transcriptional machinery to access promoters. In addition to histones, some coactivators, such as p300/CBP and the P300/CBP–associated factor (PCAF), also targets transcription factors, influencing differ- This article was published online ahead of print in MBC in Press (http://www.molbiolcell.org/cgi/doi/10.1091/mbc.E08 – 03– 0295) on September 24, 2008. Address correspondence to: Giulia Piaggio ([email protected]). © 2008 by The American Society for Cell Biology 5203
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Posttranslational regulation of NF-YA modulates NF-Y transcriptional activity

Molecular Biology of the CellVol. 19, 5203–5213, December 2008
Posttranslational Regulation of NF-YA Modulates NF-YTranscriptional ActivityIsabella Manni,* Giuseppina Caretti,† Simona Artuso,* Aymone Gurtner,*Velia Emiliozzi,* Ada Sacchi,* Roberto Mantovani,† and Giulia Piaggio*‡
*Molecular Oncogenesis Laboratory and ‡Rome Oncogenomic Center, Experimental Oncology Department,Regina Elena Cancer Institute, 00158 Rome, Italy; and †Biomolecular and Biotechnology Science Department,University of Milano, 20133 Milano, Italy
Submitted April 6, 2008; Revised September 4, 2008; Accepted September 17, 2008Monitoring Editor: Richard K. Assoian
NF-Y binds to CCAAT motifs in the promoter region of a variety of genes involved in cell cycle progression. The NF-Ycomplex comprises three subunits, NF-YA, -YB, and -YC, all required for DNA binding. Expression of NF-YA fluctuatesduring the cell cycle and is down-regulated in postmitotic cells, indicating its role as the regulatory subunit of thecomplex. Control of NF-YA accumulation is posttranscriptional, NF-YA mRNA being relatively constant. Here we showthat the levels of NF-YA protein are regulated posttranslationally by ubiquitylation and acetylation. A NF-YA proteincarrying four mutated lysines in the C-terminal domain is more stable than the wild-type form, indicating that theselysines are ubiquitylated Two of the lysines are acetylated in vitro by p300, suggesting a competition between ubiqui-tylation and acetylation of overlapping residues. Interestingly, overexpression of a degradation-resistant NF-YA proteinleads to sustained expression of mitotic cyclin complexes and increased cell proliferation, indicating that a tight regulationof NF-YA levels contributes to regulate NF-Y activity.
INTRODUCTION
The CCAAT-binding transcription factor NF-Y is a heteromericprotein composed of three subunits, NF-YA, -YB, and -YC, allnecessary for CCAAT binding (Mantovani, 1999). The NF-Ycomplex supports the basal transcription of a class of regula-tory genes responsible for cell cycle progression, among whichare mitotic cyclin complexes (Zwicker et al., 1995a,b; Bologneseet al., 1999; Farina et al., 1999; Korner et al., 2001; Gurtner et al.,2003, 2008; Di Agostino et al., 2006). Knock-out mice clearlydemonstrates that NF-Y dependent transcription is essentialduring early mouse development (Bhattacharya et al., 2003).The NF-Y function in proliferation is also demonstrated bythe inhibition of DNA binding by endogenous NF-Y, result-ing in retardation of fibroblast growth, which is broughtabout by overexpression of a dominant negative mutant ofthe NF-YA subunit (Hu and Maity, 2000). The differentialexpression of NF-YA, altering NF-Y CCAAT-binding activ-ity, has been observed in several cell lines and tissues bothduring cell cycle progression and under specific conditions.NF-YA expression is modulated during the cell cycle, beinghigh in G1, further increasing in S, and then decreasing inthe G2/M phase (Bolognese et al., 1999). Reduction ofNF-YA expression has been reported in IMR-90 fibroblastsafter serum deprivation (Chang et al., 1994) and in humanmonocytes (Marziali et al., 1997). The NF-YA protein is notexpressed in terminally differentiated C2C12 muscle cellsand adult skeletal muscle tissues (Farina et al., 1999; Gurtneret al., 2003, 2008). These observations show that levels of
NF-YA dictate the function of the NF-Y complex. Interest-ingly, control of NF-YA accumulation is posttranscriptional,because NF-YA mRNA is relatively constant in growing anddifferentiated cells (Bolognese et al., 1999; Farina et al., 1999).Yet, the mechanisms regulating the stability of the NF-YAprotein remain unknown.
The covalent addition of ubiquitin, a 76-amino acid proteinhighly conserved among eukaryotes, is an increasingly com-mon posttranslation modification that controls both the expres-sion and the activity of numerous proteins in the eukaryoticcell (Hershko and Ciechanover, 1998; Ciechanover et al., 2000).The ubiquitin–proteasome pathway is composed of the ubiq-uitin-conjugating system and the 26S proteasome; the lattercontains the multicatalytic protease complex (Hochstrasser,1996; Hershko and Ciechanover, 1998; Thrower et al., 2000).Frequent targets of the ubiquitin modification machinery aretranscription factors (Shcherbik and Haines, 2004). Indeed,many short-lived transcription factors such as E2F-1 (Hofmannet al., 1996; Hateboer et al., 1996), IkB (Chen et al., 1996), p53(Scheffner et al., 1990), SMAD2 (Lo and Massague, 1999; Zhuet al., 1999), c-Jun (Treier et al., 1994), and �-catenin (Aberleet al., 1997; Orford et al., 1997; Puca et al., 2008) are regulatedby ubiquitylation. Most of them are unstable proteinswhose rate of destruction mirrors their ability to activatetranscription. The exact nature of how activation anddestruction are linked is not yet clear (Collins and Tansey,2006; Kodadek et al., 2006).
Acetylation is a dynamic posttranslational modification oflysine residues. Proteins with intrinsic histone acetyltrans-ferase (HAT) activity act as transcriptional coactivators byacetylating histones and thereby induce an open chromatinconformation, which allows the transcriptional machinery toaccess promoters. In addition to histones, some coactivators,such as p300/CBP and the P300/CBP–associated factor(PCAF), also targets transcription factors, influencing differ-
This article was published online ahead of print in MBC in Press(http://www.molbiolcell.org/cgi/doi/10.1091/mbc.E08–03–0295)on September 24, 2008.
Address correspondence to: Giulia Piaggio ([email protected]).
© 2008 by The American Society for Cell Biology 5203

ent aspects of their function (Sterner and Berger, 2000; Chanand La Thangue, 2001). An important level of regulationlinks acetylation levels and NF-Y activity. Functionally, in-hibition of HDAC’s activity by trichostatin A (TSA) treat-ment leads to 1) activation of the MDR1 (multidrug resis-tance promoter; Jin and Scotto, 1998); 2) a dramatic increasein the activity of TGF-� II receptor promoter (Park et al.,2002); and 3) activation of the HSP70 promoter in the ab-sence of heat shock in Xenopus oocytes (Li et al., 1998). Allthese effects are strictly dependent on the presence of theCCAAT boxes. NF-Y interacts with hGCN5 (Currie, 1998), andNF-YB is acetylated in Xenopus by p300, but the consequencesof this modification have not been addressed (Li et al., 1998). Invivo studies by chromatin immunoprecipitation on cell cycle–regulated promoters highlighted the dynamic behavior ofNF-Y and HATs binding during cell cycle progression (Carettiet al., 2003; Salsi et al., 2003; Di Agostino et al., 2006).
Because of the role of ubiquitylation and acetylation inregulating the stability of several transcription factors, weinvestigated whether the NF-Y complex was subjected tothese posttranslational modifications. In this report, weshow for the first time that modulation of expression of oneNF-Y subunit, NF-YA is mediated, at least in part, by theubiquitin–proteasome degradation system. Treatment ofcells with specific proteasome inhibitors leads to a signifi-cant increase of the endogenous NF-YA protein. Mutation offour lysines in the NF-YA C-terminal domain affects ubiq-uitylation, and the resulting protein is more stable than thewild-type form, indicating that these lysines are targeted bythe ubiquitylation pathway. Two of these lysines are alsotarget of p300 acetyl transferase activity in vitro. Our resultsindicate that a posttranslational molecular mechanism reg-ulates the transcriptional activity of NF-Y controlling thestability of the regulatory subunit of the complex, NF-YA,and suggest that a competition exist between ubiquitylationand acetylation of common lysine residues of this protein.
MATERIALS AND METHODS
Cell Culture, DNA Transfections, and TreatmentsCells were cultured in DMEM containing 10% fetal bovine serum and anti-biotics. The C2C12 differentiation protocol has been already described (Farinaet al., 1999). Transient transfections in C2C12, HCT116, and NIH3T3 cells werecarried out by the BES method according to the manufacturer’s protocol. In each60-mm plate, 1.5 � 105 cells were transfected with precipitates containing suitableconcentrations of each reporter construct and of CMV �-galactosidase plasmid asan internal control for transfection efficiency. Treatments with peptide aldehydesproteasome inhibitors N-Ac-Leu-Leu-norleucinal (LLnL) were for 2, 4, or 8 h atthe concentration of 50 �M, whereas those with Z-Leu-Leu-Leu-H (MG132) andN-acetyl-leucyl-leucyl-methioninal (LLM), were for 4 or 8 h at the concentrationof 50 �M (All reagents were provided by Sigma, St. Louis, MO).
For measurement of protein half-life, cells were treated for 15 min and 2and 3 h with cycloheximide (Chx, Calbiochem, La Jolla, CA) at the concen-tration of 100 �g/ml or with Chx and LLnL together at the concentration of100 �g/ml and 50 �M, respectively. The drugs were directly added to culturemedium. Cells were harvested after treatments and lysed. The intensity of eachband was evaluated by densitometry using the NIH Image J 1.61 software(http://rsb.info.nih.gov/ij/; National Institutes of Health, Bethesda, MD).
The plasmids used in transfection experiments were: NF-YA and -YBeucaryotic expression vectors carrying the NF-YA and -YB open readingframe (ORF) under control of the SV40 promoter; the NF-YA green fluores-cent protein (GFP) carrying a fusion protein between GFP and the NF-YAORF under control of the SV40 promoter; and hemagglutinin (HA)-taggedubiquitin (UbHA) carrying a fusion protein formed by an epitope of the HAand the entire open reading frame of the ubiquitin between the CMV en-hancer and promoter and the SV40 polyadenylation signal (gift from DirkBohmann, University of Rochester, Rochester, NY). To generate NF-YA GFPmutant proteins (YA-R1, -R2, -R3, and -R2�R3), we used the Quickchangemutagenesis kit (Stratagene, La Jolla, CA) using appropriate primers onNF-YA GFP as template. Primer sequences will be available on request. Intransfection assays, pcDNA3-p300 (gift from M. Levrero, University of RomeLa Sapienza, Rome, Italy) was used. In cotransfection assays, pCCAAT-B2LUC and pmutCCAAT-B2LUC (Bolognese et al., 1999) were used. LUC
activity was assayed on whole cell extract, as described (Brasier et al., 1989).The values were normalized for �-galactosidase and protein contents.
Assessment of Efficiency of ExpressionThe expression of NF-YA and YA-R1, -R2, and -R3, as GFP fusion proteins inFigure 5C, was quantified by counting 1000 cells from five different areas(320 � 240 �m) at 0, 8, 24, and 32 h after transfection in proliferating C2C12transfected cells. The 0-h sample represents cells reseeded after an overnighttransfection and harvested immediately after attachment.
Immunolocalization of NF-YA-GFP ProteinsCells (1 � 105) were seeded on 16-mm2 coverslips overnight and fixed thenext day in 4% formaldehyde. Slides were mounted in 50% glycerol andanalyzed within 24 h. Cells were counterstained with Hoechst for DNAlabeling. Images were analyzed with a Zeiss fluorescence microscope (Axios-kop 20; Thornwood, NY).
Immunoprecipitation and Western BlotsTotal, nuclear, and cytoplasmic extracts from C2C12 cells were prepared aspreviously described (Farina et al., 1999). Proteins were resolved on 10% or
Figure 1. The proteasome pathway regulates NF-YA expression.(A) Western blot analysis of endogenous NF-YA, -YB, and -YC innuclear and cytoplasmic extracts from myoblasts and myotubestreated with 50 �M LLnL for 2, 4, and 8 h. (B) Western blot analysisof the endogenous NF-YA, -YB, -YC, and p53 in nuclear and cyto-plasmic extracts from myoblasts treated with 50 �M LLM or MG132for 4 and 8 h.
Figure 2. The half-life of NF-YA is increase by proteasome inhibi-tion. (A) Densitometric analysis of the NF-YA or -YB as a controlexpression level after normalization against the values obtained forthe control housekeeping gene tubulin under the same experimentalconditions. (B) Western blot analysis of the endogenous NF-YA and-YB, in total extracts from C2C12 cells treated with 100 �g/ml Chxfor various lengths of time and coincubated with 100 �g/ml Chxand 50 �M of LLnL. �-tubulin is used as loading control.
I. Manni et al.
Molecular Biology of the Cell5204

12% SDS-PAGE. Western blotting was performed according to the manufactur-er’s directions using the following primary antibodies: rabbit polyclonal �NF-YAand -YB (Rockland, Gilbertsville, PA), � cyclinB1, � cyclinA, � cdk1p34, �p300(Santa Cruz Biotechnology, Santa Cruz, CA), mouse monoclonal � NF-YA, aHsp70 (StessGen Biotechnologies, San Diego, CA), �-tubulin (Calbiochem), �FK2 from Affiniti Research Products (Exeter, United Kingdom) and �-acetyl-Lys(Upstate Cell Signalling, Waltham, MA), rat monoclonal � HA (Sigma).Peroxidase activity of the appropriate secondary antibodies was visualized byenhanced chemiluminescence detection system (Amersham Biosciences, Pis-cataway, NJ). For immunoprecipitations the following antibodies were used:mouse monoclonal � NF-YA (gift of R. Mantovani, University of Milano),rabbit polyclonal � NF-YB (Rockland Biosciences, Gilbertsville, PA) mousemonoclonal � FK2 (Affiniti), rat monoclonal � HA (Sigma), and mouse, rabbitor rat serum purified antibodies as control. Precleared extracts were incu-bated with protein A/G-Sepharose beads (Pierce, Rockford, IL) in lysis buffercontaining 0.05% BSA and antibodies, under constant shaking at 4°C for 2 h.After incubation, Sepharose bead–bound immunocomplexes were rinsedwith lysis buffer and eluted in 50 �l of SDS sample buffer for Western blottingand probed with � HA antibodies (Sigma). The ubiquitin ladder was quan-tified by measuring the signals of the bands by NIH Image 1.61 software.
Acetyltransferase AssayAcetyltransferase assays were performed at 30°C, for 45 min using 1 �g ofsubstrates, 50 ng of p300 or PCAF in 30 �l of assay buffer containing 10%glycerol, 50 mM Tris HCl (pH 8), 1 mM DTT, 1 mM PMSF, 10 mM sodiumbutyrate and 0.1 �Ci of [3H]acetyl CoA (New England Nuclear, Boston, MA).The NF-YA, H3 and H4 proteins have been purified from inclusion bodies aspreviously described (Mantovani et al., 1992; Caretti et al., 1999). YA9 is anHis-tag protein and it has been purified as previously described (Liberati et al.,1999; Zemzoumi et al., 1999). The thioredoxin (TRX) protein has been pro-duced from pET-32b vector from Novagen (Madison, WI) according to themanufacturer’s instructions. Samples were run on 12 or 15% SDS poly-acrylamide gels, stained with Coomassie brilliant blue R-250, treated withAmplify (Amersham), dried, and autoradiographed.
Elecrophoretic Mobility Shift AssaysDNA-binding reactions of Figure 7B was performed in NDB-100/BSA with0.2–0.7 and 2 ng of NF-Y. The oligonucleotides used in electrophoretic mo-bility shift assays (EMSAs) were the cyclin B2 distal and middle CCAATboxes described in Bolognese et al. (1999). Reactions were incubated for 20min at room temperature and run in a 4.5% polyacrylamide gel (29:1 Acryl-amide/Bis ratio) in 0.5� TBE at 4°C for 3 h.
RT-PCRTotal RNA from C2C12 cells was extracted using the TRIzol RNA isolationsystem (Invitrogen, Carlsbad, CA) according to the manufacturer’s in-
structions. The first-strand cDNA was synthesized according to the man-ufacturer’s instructions (M-MLV RT kit; Invitrogen). PCR was performedwith HOT-MASTER Taq (Eppendorf, Fremont, CA) using 2 �l of cDNAreaction. PCR products were run on a 2% agarose gel and visualized withethidium bromide. The sequences of oligonucleotide primers were asfollows: NFYA: F5�atcccagcagccagtttggcag, R5�gaaaaatcgtccaccttcaccacg;p300: F5�atgccacagccccctattgg, R5�gagacactggtgcttgaccg.
The housekeeping aldolase A mRNA, used as an internal standard, wasamplified from the same cDNA reaction mixture using the following specificprimers: F5�tggatgggctgtctgaacgctgt and R5�agtgacagcagggggcactgt.
Small Interfering RNA TransfectionHuman HCT116 cells were seeded at a density of 1.6 � 106 in 100-mm culturedishes in DMEM medium supplemented with 10% FBS. The next day, cells weretransfected using Lipofectamine 2000 reagent (Invitrogen) following the manu-facturer’s instructions with 10 �l of 0.02 mM p300 small interfering RNA (siRNA;5�-AAC CCC UCC UCU UCA GCA CCA-3�; Dharmacon Research, Boulder,CO), or nontargeting siRNA scramble (Dharmacon, Lafayette, CO) as a negativecontrol.
Colony Formation AssayNIH3T3 cells were cotransfected with wild-type NF-YA or its mutants (YA-R1, YA-R2, and YA-R3) and pBABE-PURO (10:1 ratio). The cells were selectedin 2 �g/ml puromycin (Sigma) at 48 h after transfection, and the colonieswere stained and counted 2 wk later. Plates were stained with 0.5 ml of0.005% Crystal Violet for �1 h, and colonies were counted using a dissectingmicroscope.
RESULTS
The Proteasome Pathway Regulates NF-YA ExpressionTo test the hypothesis that NF-Y is a direct target of theubiquitin–proteasome pathway, NF-YA, -YB, and -YCprotein levels were measured in cells treated with differ-ent peptide aldehydes proteasome inhibitors: N-acetyl-leucyl-leucyl-norleucinal (LLnL), Z-Leu-Leu-Leu-H (MG132), andN-acetyl-leucyl-leucyl-methioninal (LLM). Murine C2C12muscle cells, myoblasts, were treated with 50 �M LLnL for2, 4 and 8 h and the levels of NF-YA, -YB, and -YC in nuclearand cytoplasmic protein extracts were analyzed by Westernblot (Figure 1A). After treatment, the NF-YA protein accu-mulated both in the cytoplasm and in the nucleus, whereas
Figure 3. Exogenous NF-YA protein is ubiquitylated invivo. C2C12 cells were transfected with plasmids encod-ing NF-YA (A–C) or -YB (D) together with UbHA. Totalextracts from these cells were immunoprecipitated withincreasing amounts of anti-NF-YA (A and B), anti-HA (C)or anti-NF-YB antibodies (D). Western blot analysis wasperformed with an anti-HA antibody in (A and D) or antiNF-YA antibody (C and B). In all immunoprecipitationexperiments an appropriate control antibody was used.Stars indicate the antibody chains used in the immunopre-cipitations (IPs).
Modifications of NF-YA Protein
Vol. 19, December 2008 5205
the protein levels of NF-YB and -YC were only slightlymodulated. We have previously demonstrated that in ter-minally differentiated C2C12 muscle cells, myotubes, NF-YAexpression is abrogated (Farina et al., 1999; Gurtner et al.,2003, 2008). Thus, we asked whether regulation of NF-YAexpression occurs at posttranslational level in this cell sys-tem. To this end, myotubes were treated with LLnL andlevels of NF-YA, -YB, and -YC were analyzed. (Figure 1A).We observed an increase of NF-YA protein prevalently inthe nucleus but also in the cytoplasm.
We tested two other compounds known to inhibit proteindegradation, MG132 and LLM (Rock et al., 1994). In myo-blasts, MG132 treatment led to an increase of NF-YA in thecytoplasm. In contrast LLM had no effect on NF-YA proteinlevels (Figure 1B). As a positive control, we analyzed p53protein level in the same experimental conditions, and weobserved that, as expected, it accumulated in the nuclearfraction after both treatments. Both LLnL and MG132 inhibitthe proteasome pathway (Wang 1990), whereas LLM is astrong inhibitor of calpain and cathepsin but a very weakinhibitor of the proteasome (Rock et al., 1994). Thus, inhibi-tion of proteasome, but not of a cysteine protease, pathwayresults in NF-YA accumulation (Figure 1B). Taken together,these results indicate that NF-YA expression is markedlyregulated at posttranslational level. In contrast, the levels ofthe YB and YC subunits are slightly affected by the protea-some inhibitors, suggesting that the modulation of expres-sion of the NF-YA subunit could dictate the activity of theNF-Y trimer.
The Half-Life of NF-YA Is Increased by ProteasomeInhibitionBecause ubiquitin-mediated protein destruction has beenmainly implicated for short-lived proteins, we sought todetermine the half-life of NF-YA. C2C12 cells were incu-bated with Chx (100 �g/ml) for 15 and 30 min and 1, 2, or3 h. As shown in Figure 2A, by densitometric analysis, theamount of NF-YA protein decreased by �50% after 2 h. Incontrast to this, the steady-state levels of NF-YB were notmodulated in the same experimental conditions, indicatingthat the half-life of these proteins is longer than 3 h.
To further support the proteasome involvement in thedegradation of the NF-YA subunit, its half-life was deter-mined in the presence of LLnL. C2C12 cells were treated for15 min or 2 or 3 h with both Chx and LLnL. As shown inFigure 2B, LLnL treatment prevented the Chx-induced de-crease of NF-YA. As expected, NF-YB protein levels werenot modulated. These results demonstrate that NF-YA deg-radation after Chx treatment is indeed mediated by theproteasome in C2C12 cells.
NF-YA Protein Is Ubiquitylated In VivoTo determine the involvement of ubiquitylation in NF-YAdegradation, a fusion protein formed by an epitope of theHA and the entire open reading frame of the UbHA (Treieret al., 1994) was cotransfected in C2C12 cells with a vectorcoding for NF-YA. Total cell extracts were immunoprecipi-tated with increasing amounts of anti-NF-YA antibody.Western blot analysis detected smeared high-molecular-weight species, characteristic of ubiquitylated proteins, bothwith anti-HA (Figure 3A) and anti-NF-YA (Figure 3B) anti-bodies. Increasing amounts of anti-NF-YA antibody immu-noprecipitated increasing amount of these protein forms.Reciprocal immunoprecipitation experiments showed thatNF-YA putative ubiquitylated forms are present in thesmeared high-molecular-weight species immunoprecipi-tated with the anti-HA antibody (Figure 3C). These species
were not detected in immunoprecipitates with control anti-body. In the same experimental conditions, low levels ofhigh-molecular-weight species were detected in extract im-munoprecipitated with anti-NF-YB antibody from C2C12cells cotransfected with vectors coding for NF-YB andUbHA, suggesting that this subunit is ubiquitylated to alesser extent than NF-YA (Figure 3D). This result is in agree-ment with the minimal effect of LLnL on NF-YB proteinlevels shown above.
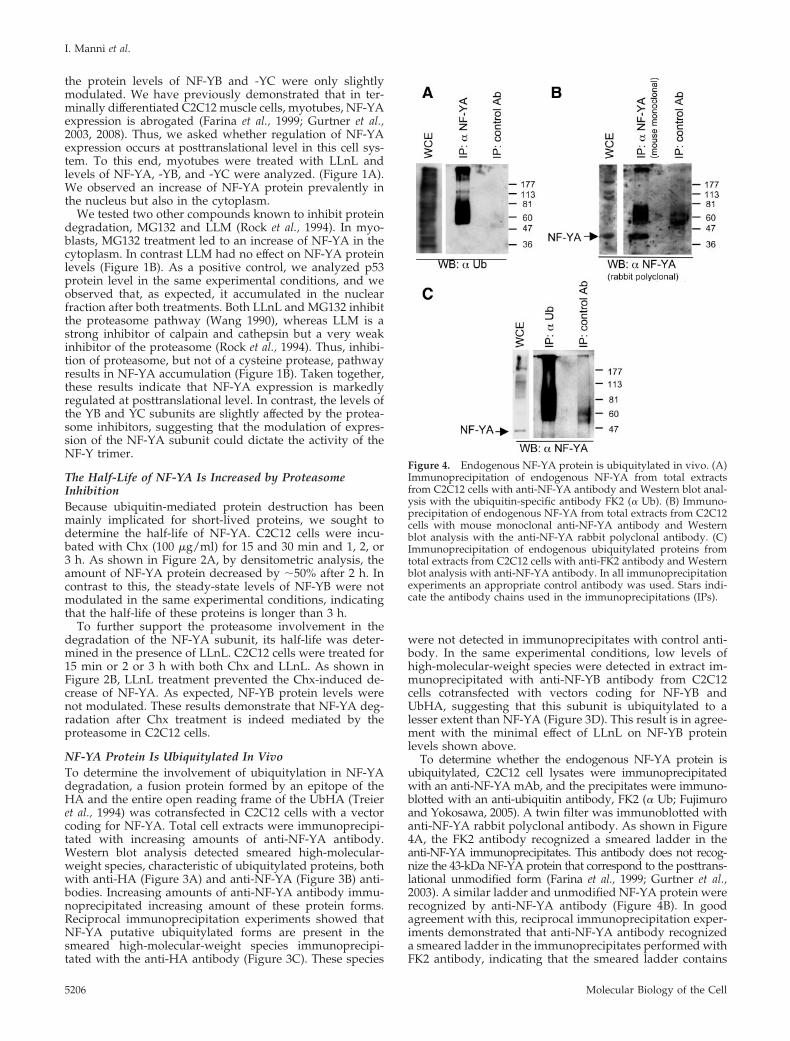
To determine whether the endogenous NF-YA protein isubiquitylated, C2C12 cell lysates were immunoprecipitatedwith an anti-NF-YA mAb, and the precipitates were immuno-blotted with an anti-ubiquitin antibody, FK2 (� Ub; Fujimuroand Yokosawa, 2005). A twin filter was immunoblotted withanti-NF-YA rabbit polyclonal antibody. As shown in Figure4A, the FK2 antibody recognized a smeared ladder in theanti-NF-YA immunoprecipitates. This antibody does not recog-nize the 43-kDa NF-YA protein that correspond to the posttrans-lational unmodified form (Farina et al., 1999; Gurtner et al.,2003). A similar ladder and unmodified NF-YA protein wererecognized by anti-NF-YA antibody (Figure 4B). In goodagreement with this, reciprocal immunoprecipitation exper-iments demonstrated that anti-NF-YA antibody recognizeda smeared ladder in the immunoprecipitates performed withFK2 antibody, indicating that the smeared ladder contains
Figure 4. Endogenous NF-YA protein is ubiquitylated in vivo. (A)Immunoprecipitation of endogenous NF-YA from total extractsfrom C2C12 cells with anti-NF-YA antibody and Western blot anal-ysis with the ubiquitin-specific antibody FK2 (� Ub). (B) Immuno-precipitation of endogenous NF-YA from total extracts from C2C12cells with mouse monoclonal anti-NF-YA antibody and Westernblot analysis with the anti-NF-YA rabbit polyclonal antibody. (C)Immunoprecipitation of endogenous ubiquitylated proteins fromtotal extracts from C2C12 cells with anti-FK2 antibody and Westernblot analysis with anti-NF-YA antibody. In all immunoprecipitationexperiments an appropriate control antibody was used. Stars indi-cate the antibody chains used in the immunoprecipitations (IPs).
I. Manni et al.
Molecular Biology of the Cell5206

ubiquitylated NF-YA protein (Figure 4C). In both experi-ments, no protein ladder was detected in immunoprecipi-tates performed with the control antibody. Taken together,these data strongly indicate that both exogenous and endog-enous NF-YA is ubiquitylated in vivo and suggest that theubiquitin–proteasome pathway degrades it.
Identification of NF-YA Lysine Residues Targets forUbiquitylationThe C-terminus domain of NF-YA protein, containing theNF-YB/-YC interaction domain and the DNA-binding do-main, is highly conserved from yeast to mammals. In par-ticular, it contains six lysine residues conserved across thespecies (Figure 5A). Lysines K269, K276, and K283 are in thesubunits interacting region, whereas lysines K289, K292, andK296 are in the linker. Because of their evolutionary conser-vation we asked whether these lysines could be target forubiquitylation. To answer this question we derived threemutants YA-R1, YA-R2, and YA-R3 in the context of thefull-length protein. In each of them two of these lysines weresubstituted with arginines (Figure 5B). Wild-type and mu-tant NF-YA were cloned in frame with GFP and overex-pressed as GFP fusion proteins in C2C12 cells. As shown inFigure 5C, the number of GFP positive cells at the end of thetime course is significantly higher in cell populations trans-fected with mutants than with wild-type NF-YA, suggestingthat mutant proteins are more stable compared with wild
type. Because GFP might influence the stability of fusionproteins we overexpressed wild-type and mutant NF-YAproteins not in frame with GFP in the same cells. As shownin Figure 5D, the stability of native mutant forms too ishigher than that of wild-type NF-YA, Taken together, theseresults indicate that the mutant proteins are more stablecompared with wild type, although the expression kineticsof the proteins is different in the two experiments (Figure 5,C and D). Moreover, the mutant proteins YA-R2 and -R3 aremore stable than -R1, suggesting that the four lysinespresent in these constructs play a major role in NF-YAprotein stability.
To investigate whether these lysines are target for ubiqui-tylation we generated a mutant protein in which all of themwere substituted with arginines (Figure 5B). We transientlytransfected C2C12 cells with either NF-YA wild-type protein(NF-YA) or mutant forms of it (YA-R1, -R2, -R3, and-R2�R3). The YA-R2�R3 mutant was more stable than wild-type NF-YA (Figure 6A) as well as the single YA-R2 and -R3mutants (Figure 5D). Consistent with the higher stability ofthese proteins, we observed that formation of ubiquitylatedNF-YA forms was partially inhibited on YA-R2, -R3, or-R2�R3 mutants (Figure 6B). Interestingly, densitometricanalysis indicated that the ubiquitin ladder of the YA-R2�R3 mutant was inhibited by �70% (Figure 6C), whereasformation of ubiquitylated forms was essentially not inhib-ited in the YA-R1 mutant, in good agreement with its re-
Figure 5. Identification of NF-YA lysine residues targets for ubiquitylation. (A) Schematic representation of the C-terminus domain of theNF-YA protein containing six lysine residues conserved across species. (B) Schematic representation of NF-YA protein with the positions ofthe lysines replaced in the different NF-YA mutants used in this work. (C) Number of GFP-positive cells in cell populations transfected withmutants (YA-R1, -R2, and YA-R3), wild-type NF-YA (NF-YA), or the empty vector as a control, at different times. (D) Time course ofwild-type and mutants NF-YA expression by Western blot analysis on total extracts from cells nontransfected (NT) or transfected withmutants (YA-R1, -R2, and -R3) or wild-type NF-YA (NF-YA).
Modifications of NF-YA Protein
Vol. 19, December 2008 5207

duced stability (Figure 5D). We conclude that lysines K283,K289, K292, and K296 are those mainly ubiquitylated in thecontest of the NF-YA protein.
Two Ubiquitylated Lysines of NF-YA Are Targets of p300Acetylation ActivityWe have previously demonstrated that p300 interacts withNF-Y. Acetylated forms of NF-YA are present in the resultingcomplex, suggesting that p300 might regulate its acetylationstatus (Di Agostino et al., 2006). To investigate whether lysinestarget for ubiquitylation undergo p300-dependent acetyla-tion, we used recombinant purified p300 and NF-YA in vitroacetylation studies (Figure 7A). As expected, p300 was ableto self-acetylate and also acetylated recombinant full lengthNF-YA. Mapping the region acetylated by p300 using dele-tion mutant, we observed that the deletion mutant YA9,which contains all the ubiquitylated lysines, was acetylatedin vitro by p300, clearly indicating that this conserved part ofthe protein is efficiently posttranslationally modified. Asexpected, the negative control, TRX, was not acetylated inthe same experimental conditions. As positive control weanalyzed the acetylation status of H3-H4 histones.
To investigate whether the lysines that undergo ubiquity-lation could be target for p300 dependent acetylation, thethree mutants YA-R1, -R2, and -R3 were used in in vitroacetylation studies. These mutants were first tested inEMSA: in dose-response experiments, all mutants were ca-
pable of binding DNA with the same efficiency of wild-typeNF-YA (Figure 7B), a clear indication of normal interactionswith the NF-YB/-YC subunits. Direct acetylations deter-mined that YA-R2, and to a lesser degree YA-R1, were notefficiently acetylated by p300 (Figure 7C), whereas the mu-tant YA-R3 still underwent acetylation. We conclude that, inthe contest of the NF-YA full-length protein, the two lysines283 and 289 are the major targets of p300 acetylation activity.Of note, these two lysines are also ubiquitylated, suggestinga possible competition between acetylation and ubiquityla-tion on these residues.
p300 Expression Impacts on NF-YA UbiquitylationTheoretically, acetylation of specific lysines could increasethe stability of a protein, because it would prevent ubiqui-tylation of the same lysine residues. To determine if p300could affect the expression of NF-YA, C2C12 cells weretransfected with p300. The amount of NF-YA was slightlyincreased in the presence of ectopic p300 (Figure 8A). Weverified the presence of ubiquitylated NF-YA forms in ly-sates of C2C12 cells cotransfected with p300 and NF-YA.Consistent with the higher stability of the protein, we ob-served that formation of ubiquitylated NF-YA forms wasinhibited in cells overexpressing p300 compared with nop300 (Figure 8B), suggesting that p300 acetylation of NF-YAcould prevent, at least in part, its ubiquitylation. Next, weassessed the function of p300 in normal physiological set-tings, employing siRNA against p300. To this end, humanHCT116 cells were transfected with an oligo pool directedagainst p300 (sip300; Gong et al., 2006) or, as control, anunrelated sequence (scramble), and incubated in the pres-ence of Chx, which blocks de novo protein synthesis. Cellu-lar extracts were prepared 2 h after Chx addition, and levelsof NF-YA and p300 proteins were determined by Westernblot analysis. As shown in Figure 8C, p300 loss did not alterNF-YA half-life in untreated cells. However, in the presenceof Chx, the antibody against NF-YA detected high-molecu-
Figure 6. Identification of NF-YA lysine residues targets for ubiq-uitylation. (A) Western blot analysis on total cell extracts fromnontransfected C2C12 cells (NT) and cells transfected with increas-ing amounts of plasmids encoding wild-type (NF-YA) or mutantNF-YA (NF-YA R2�R3; see Figure 5B). (B) Total extracts from cellsoverexpressing wild-type or YA-R1, -R2, -R3, and -R2�R3, immu-noprecipitated with anti-NF-YA antibody, or mouse serum. West-ern blot analysis was performed with an anti-ubiquitin antibodyFK2 (� Ub). In all immunoprecipitation experiments an appropriatecontrol antibody was used. Stars indicate the antibody chains usedin the immunoprecipitations (IPs). (C) Densitometric analysis ofNF-YA ubiquitin ladder.
Figure 7. p300 acetylation of NF-YA in vitro. (A) In vitro acetyla-tion by p300 of the NF-YA proteins outlined in the scheme. YA9harbors only the highly conserved part of NF-YA. (B) EMSA anal-ysis of wild-type NF-YA (NF-YA) and YA-R1, -R2, and -R3 mutantson a CCAAT-box containing oligonucleotide. A dose-response 0.1,0.3, and 1 ng of purified NF-YAs was preincubated with recombi-nant NF-YB and -YC, 5 ng, and added to labeled DNA. (C) In vitroacetylation of recombinant purified NF-YA mutants by p300. In thebottom panel, Coomassie blue staining of the SDS gel is shown.
I. Manni et al.
Molecular Biology of the Cell5208

lar-weight species in cells interfered for p300, suggesting thepresence of NF-YA putative ubiquitylated forms. Taken to-gether, these results indicate that p300 could increase thestability of NF-YA, suggesting that its acetylation mightenhance its expression, potentially by interfering with theubiquitin–proteasome pathway.
We asked whether p300-dependent increase of the NF-YAprotein was also due, at least in part, to increased transcrip-tion of the NF-YA gene. To answer this question we per-formed semiquantitative RT-PCR assays (Figure 8D). Theresults demonstrate that the NF-YA mRNA level is notchanged after p300 transfection. Therefore, we conclude thatthe stabilizing effect that p300 exerts over NF-YA is predom-inantly due to posttranslational events.
Acetylation and Ubiquitylation of NF-YA Protein inPre- and Postmitotic CellsTo begin to investigate the functional relationship betweenacetylation and ubiquitylation of NF-YA, we analyzed therelative abundance of acetylated and ubiquitylated NF-YAspecies in nuclear and cytoplasmic cell compartments. Forthis purpose, C2C12 protein extracts were immunoprecipi-tated with anti-NF-YA and subsequently blotted with FK2and anti-acetyl-lysine antibodies (Figure 9A). We observedthat the nuclear compartment contained higher levels ofacetylated NF-YA protein in comparison to the cytoplasmicone. Conversely, ubiquitylated NF-YA forms were moreabundant in the cytoplasm. Of note, a strong decrease inacetylation was observed if an artificial increase of ubiqui-tylated NF-YA forms was induced in the nucleus with LLnLtreatment (Figure 9B). These results support the hypothesisthat acetylated NF-YA protein, acting as a transcription fac-tor in the nucleus, is the active form, whereas the nonactiveubiquitylated form is prevalently stored in the cytoplasm. Ifthis hypothesis is correct, we would expect that in postmi-totic cells, where we have previously demonstrated absenceof NF-Y activity (Farina et al., 1999; Gurtner et al., 2003;Gurtner et al., 2008), only ubiquitylated NF-YA species areexpressed. As already shown in Figure 1A, myotubes ex-press NF-YA only after treatment with LLnL. Thus, nu-clear extracts from myotubes treated with LLnL wereimmunoprecipitated with anti-NF-YA and subsequentlyblotted with FK2 and anti-acetyl-lysine antibodies. Asshown in Figure 9C, only ubiquitylated NF-YA species
were present in differentiated nuclei under these experi-mental conditions (Figure 9C). Taken together, these re-sults determine that the nuclei of premitotic cells containmainly acetylated NF-YA protein, whereas in postmitoticcells, where NF-Y does not exert its activity, the protein isonly ubiquitylated.
Acetylation and Ubiquitylation of NF-YA Lysine ResiduesAre Involved in its Transcriptional ActivityWe have shown that the NF-YA mutants are capable ofbinding DNA with the same efficiency of wild-type NF-YA(Figure 7B), suggesting that they might retain efficient tran-scriptional activity. To test this hypothesis, first we deter-mined the cellular localization of the wild-type and mutantNF-YA proteins using the fluorescent properties of GFP.Analysis by fluorescence microscopy showed that GFP chi-meras of mutants and wild-type NF-YA were all targeted tothe nucleus (Figure 10A). On the basis of the ability ofmutants to bind DNA in vitro and their nuclear localizationin vivo, we investigated their ability to transactive a NF-Ytarget promoter. For this purpose, C2C12 cells were tran-siently cotransfected with YA-R1, -R2, -R3, and -R2�R3mutants or wild-type NF-YA together with the cyclin B2promoter (Figure 10B). We found that the activity of thispromoter was up-regulated in the presence of mutants com-pared with that observed when cotransfected with wild-typeNF-YA. This up-regulation was lost in the context of a cyclinB2 promoter carrying three mutated CCAAT boxes (data notshown).
To begin to address whether the increased activity ofmutant proteins is due to a lack of ubiquitylation or acety-lation, we tested the ability of p300 or ubiquitin to modulatethe transcriptional activity of wild-type NF-YA. C2C12 cellswere transiently cotransfected with wild-type NF-YA andp300 or wild-type NF-YA and UbHA together with thecyclin B2 promoter (Figure 10C). Interestingly, p300 en-hances NF-YA activity, whereas ubiquitin reduces it. Takentogether, the results indicate that a more stable NF-YA pro-tein still retains and enhances its transcriptional activity.Moreover they suggest that NF-YA transcriptional activitycould be due to a balance between acetylation and ubiqui-tylation.
Figure 8. p300 expression impacts on NF-YA ubiq-uitylation. (A) Western blot analysis of NF-YA in totalcell extracts from C2C12 cells transfected with p300.Actin was used as loading control. (B) C2C12 cellswere transfected with wild-type NF-YA and increas-ing amounts of p300. Total extracts from these cellswere immunoprecipitated with anti-NF-YA antibodyand Western blot analysis with the ubiquitin-specificantibody, FK2 (� Ub). In immunoprecipitation exper-iments an appropriate control antibody was used.Stars indicate the antibody chains used in the IPs. (C)Western blot analysis of the endogenous NF-YA andp300 in total extracts from HCT116 cells treated with100 �g/ml Chx for 2h and transfected with an oligopool directed against p300 (sip300) or an unrelatedsequence (scramble). Actin was used as loading con-trol. (D) Semiquantitative RT-PCR analysis of NF-YAand p300 expression on total RNA from C2C12 cellstransfected with increasing amounts of p300. Aldolasewas used as loading control. Ethidium bromide stain-ing of total RNA is shown. Stars indicate the antibodychains used in the immunoprecipitations (IPs).
Modifications of NF-YA Protein
Vol. 19, December 2008 5209
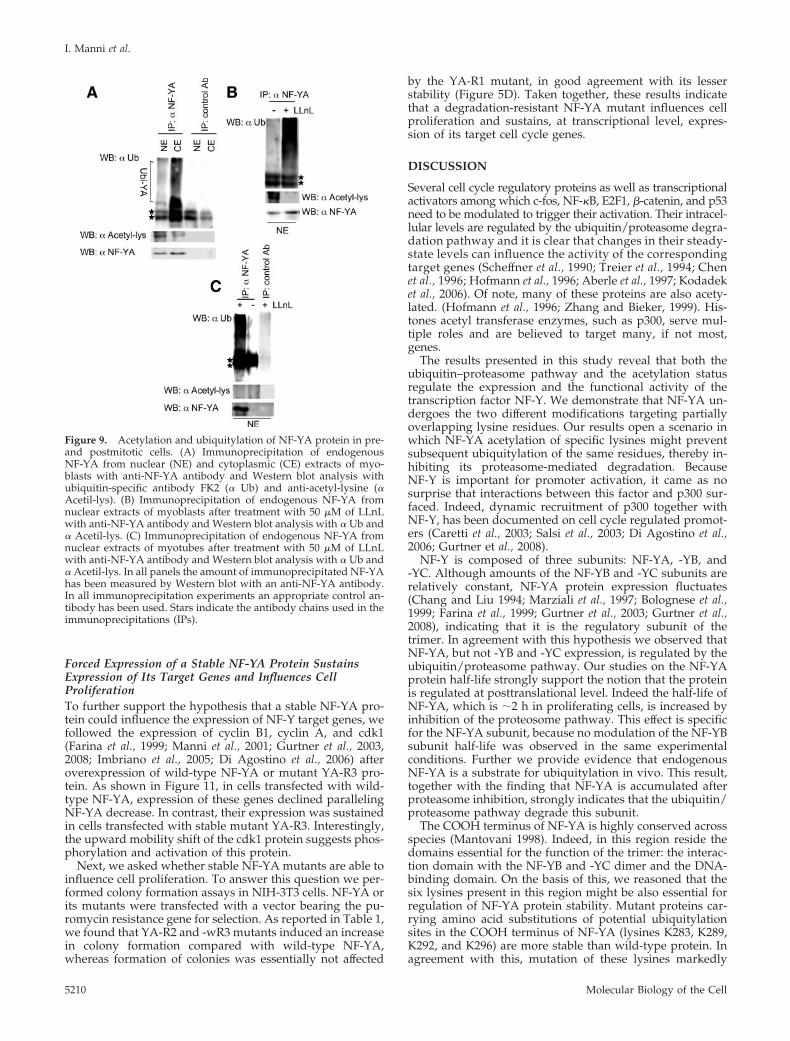
Forced Expression of a Stable NF-YA Protein SustainsExpression of Its Target Genes and Influences CellProliferationTo further support the hypothesis that a stable NF-YA pro-tein could influence the expression of NF-Y target genes, wefollowed the expression of cyclin B1, cyclin A, and cdk1(Farina et al., 1999; Manni et al., 2001; Gurtner et al., 2003,2008; Imbriano et al., 2005; Di Agostino et al., 2006) afteroverexpression of wild-type NF-YA or mutant YA-R3 pro-tein. As shown in Figure 11, in cells transfected with wild-type NF-YA, expression of these genes declined parallelingNF-YA decrease. In contrast, their expression was sustainedin cells transfected with stable mutant YA-R3. Interestingly,the upward mobility shift of the cdk1 protein suggests phos-phorylation and activation of this protein.
Next, we asked whether stable NF-YA mutants are able toinfluence cell proliferation. To answer this question we per-formed colony formation assays in NIH-3T3 cells. NF-YA orits mutants were transfected with a vector bearing the pu-romycin resistance gene for selection. As reported in Table 1,we found that YA-R2 and -wR3 mutants induced an increasein colony formation compared with wild-type NF-YA,whereas formation of colonies was essentially not affected
by the YA-R1 mutant, in good agreement with its lesserstability (Figure 5D). Taken together, these results indicatethat a degradation-resistant NF-YA mutant influences cellproliferation and sustains, at transcriptional level, expres-sion of its target cell cycle genes.
DISCUSSION
Several cell cycle regulatory proteins as well as transcriptionalactivators among which c-fos, NF-�B, E2F1, �-catenin, and p53need to be modulated to trigger their activation. Their intracel-lular levels are regulated by the ubiquitin/proteasome degra-dation pathway and it is clear that changes in their steady-state levels can influence the activity of the correspondingtarget genes (Scheffner et al., 1990; Treier et al., 1994; Chenet al., 1996; Hofmann et al., 1996; Aberle et al., 1997; Kodadeket al., 2006). Of note, many of these proteins are also acety-lated. (Hofmann et al., 1996; Zhang and Bieker, 1999). His-tones acetyl transferase enzymes, such as p300, serve mul-tiple roles and are believed to target many, if not most,genes.
The results presented in this study reveal that both theubiquitin–proteasome pathway and the acetylation statusregulate the expression and the functional activity of thetranscription factor NF-Y. We demonstrate that NF-YA un-dergoes the two different modifications targeting partiallyoverlapping lysine residues. Our results open a scenario inwhich NF-YA acetylation of specific lysines might preventsubsequent ubiquitylation of the same residues, thereby in-hibiting its proteasome-mediated degradation. BecauseNF-Y is important for promoter activation, it came as nosurprise that interactions between this factor and p300 sur-faced. Indeed, dynamic recruitment of p300 together withNF-Y, has been documented on cell cycle regulated promot-ers (Caretti et al., 2003; Salsi et al., 2003; Di Agostino et al.,2006; Gurtner et al., 2008).
NF-Y is composed of three subunits: NF-YA, -YB, and-YC. Although amounts of the NF-YB and -YC subunits arerelatively constant, NF-YA protein expression fluctuates(Chang and Liu 1994; Marziali et al., 1997; Bolognese et al.,1999; Farina et al., 1999; Gurtner et al., 2003; Gurtner et al.,2008), indicating that it is the regulatory subunit of thetrimer. In agreement with this hypothesis we observed thatNF-YA, but not -YB and -YC expression, is regulated by theubiquitin/proteasome pathway. Our studies on the NF-YAprotein half-life strongly support the notion that the proteinis regulated at posttranslational level. Indeed the half-life ofNF-YA, which is �2 h in proliferating cells, is increased byinhibition of the proteosome pathway. This effect is specificfor the NF-YA subunit, because no modulation of the NF-YBsubunit half-life was observed in the same experimentalconditions. Further we provide evidence that endogenousNF-YA is a substrate for ubiquitylation in vivo. This result,together with the finding that NF-YA is accumulated afterproteasome inhibition, strongly indicates that the ubiquitin/proteasome pathway degrade this subunit.
The COOH terminus of NF-YA is highly conserved acrossspecies (Mantovani 1998). Indeed, in this region reside thedomains essential for the function of the trimer: the interac-tion domain with the NF-YB and -YC dimer and the DNA-binding domain. On the basis of this, we reasoned that thesix lysines present in this region might be also essential forregulation of NF-YA protein stability. Mutant proteins car-rying amino acid substitutions of potential ubiquitylationsites in the COOH terminus of NF-YA (lysines K283, K289,K292, and K296) are more stable than wild-type protein. Inagreement with this, mutation of these lysines markedly
Figure 9. Acetylation and ubiquitylation of NF-YA protein in pre-and postmitotic cells. (A) Immunoprecipitation of endogenousNF-YA from nuclear (NE) and cytoplasmic (CE) extracts of myo-blasts with anti-NF-YA antibody and Western blot analysis withubiquitin-specific antibody FK2 (� Ub) and anti-acetyl-lysine (�Acetil-lys). (B) Immunoprecipitation of endogenous NF-YA fromnuclear extracts of myoblasts after treatment with 50 �M of LLnLwith anti-NF-YA antibody and Western blot analysis with � Ub and� Acetil-lys. (C) Immunoprecipitation of endogenous NF-YA fromnuclear extracts of myotubes after treatment with 50 �M of LLnLwith anti-NF-YA antibody and Western blot analysis with � Ub and� Acetil-lys. In all panels the amount of immunoprecipitated NF-YAhas been measured by Western blot with an anti-NF-YA antibody.In all immunoprecipitation experiments an appropriate control an-tibody has been used. Stars indicate the antibody chains used in theimmunoprecipitations (IPs).
I. Manni et al.
Molecular Biology of the Cell5210
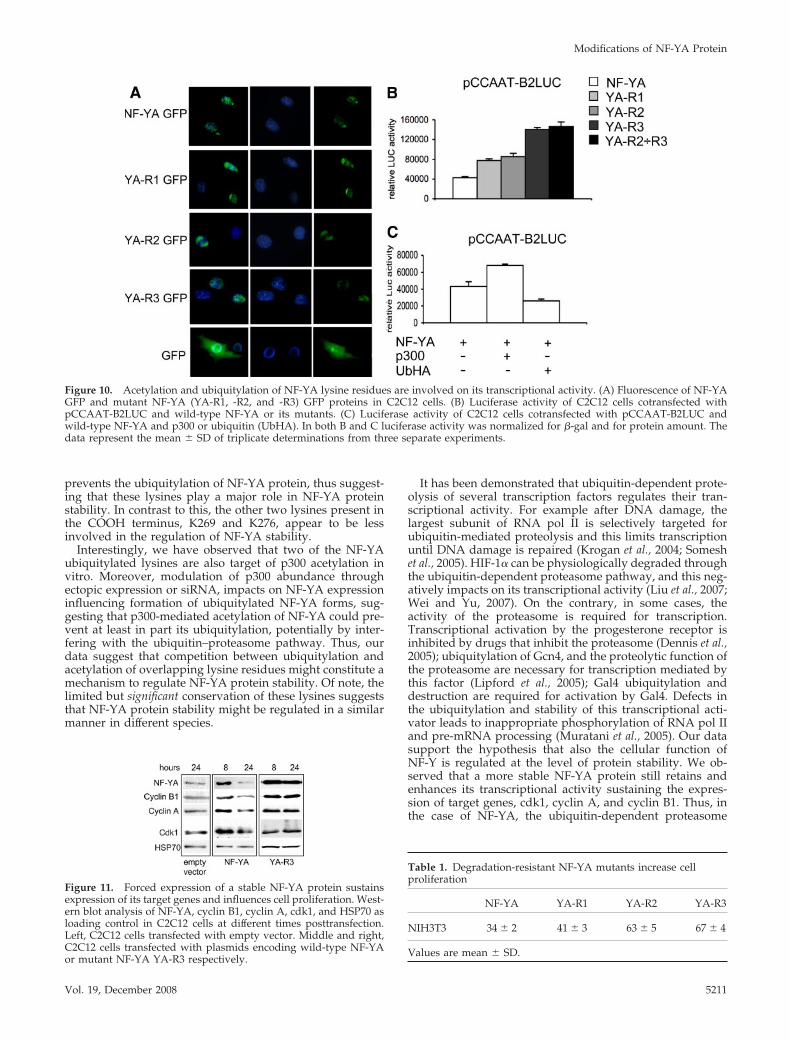
prevents the ubiquitylation of NF-YA protein, thus suggest-ing that these lysines play a major role in NF-YA proteinstability. In contrast to this, the other two lysines present inthe COOH terminus, K269 and K276, appear to be lessinvolved in the regulation of NF-YA stability.
Interestingly, we have observed that two of the NF-YAubiquitylated lysines are also target of p300 acetylation invitro. Moreover, modulation of p300 abundance throughectopic expression or siRNA, impacts on NF-YA expressioninfluencing formation of ubiquitylated NF-YA forms, sug-gesting that p300-mediated acetylation of NF-YA could pre-vent at least in part its ubiquitylation, potentially by inter-fering with the ubiquitin–proteasome pathway. Thus, ourdata suggest that competition between ubiquitylation andacetylation of overlapping lysine residues might constitute amechanism to regulate NF-YA protein stability. Of note, thelimited but significant conservation of these lysines suggeststhat NF-YA protein stability might be regulated in a similarmanner in different species.
It has been demonstrated that ubiquitin-dependent prote-olysis of several transcription factors regulates their tran-scriptional activity. For example after DNA damage, thelargest subunit of RNA pol II is selectively targeted forubiquitin-mediated proteolysis and this limits transcriptionuntil DNA damage is repaired (Krogan et al., 2004; Someshet al., 2005). HIF-1� can be physiologically degraded throughthe ubiquitin-dependent proteasome pathway, and this neg-atively impacts on its transcriptional activity (Liu et al., 2007;Wei and Yu, 2007). On the contrary, in some cases, theactivity of the proteasome is required for transcription.Transcriptional activation by the progesterone receptor isinhibited by drugs that inhibit the proteasome (Dennis et al.,2005); ubiquitylation of Gcn4, and the proteolytic function ofthe proteasome are necessary for transcription mediated bythis factor (Lipford et al., 2005); Gal4 ubiquitylation anddestruction are required for activation by Gal4. Defects inthe ubiquitylation and stability of this transcriptional acti-vator leads to inappropriate phosphorylation of RNA pol IIand pre-mRNA processing (Muratani et al., 2005). Our datasupport the hypothesis that also the cellular function ofNF-Y is regulated at the level of protein stability. We ob-served that a more stable NF-YA protein still retains andenhances its transcriptional activity sustaining the expres-sion of target genes, cdk1, cyclin A, and cyclin B1. Thus, inthe case of NF-YA, the ubiquitin-dependent proteasome
Figure 11. Forced expression of a stable NF-YA protein sustainsexpression of its target genes and influences cell proliferation. West-ern blot analysis of NF-YA, cyclin B1, cyclin A, cdk1, and HSP70 asloading control in C2C12 cells at different times posttransfection.Left, C2C12 cells transfected with empty vector. Middle and right,C2C12 cells transfected with plasmids encoding wild-type NF-YAor mutant NF-YA YA-R3 respectively.
Figure 10. Acetylation and ubiquitylation of NF-YA lysine residues are involved on its transcriptional activity. (A) Fluorescence of NF-YAGFP and mutant NF-YA (YA-R1, -R2, and -R3) GFP proteins in C2C12 cells. (B) Luciferase activity of C2C12 cells cotransfected withpCCAAT-B2LUC and wild-type NF-YA or its mutants. (C) Luciferase activity of C2C12 cells cotransfected with pCCAAT-B2LUC andwild-type NF-YA and p300 or ubiquitin (UbHA). In both B and C luciferase activity was normalized for �-gal and for protein amount. Thedata represent the mean � SD of triplicate determinations from three separate experiments.
Table 1. Degradation-resistant NF-YA mutants increase cellproliferation
NF-YA YA-R1 YA-R2 YA-R3
NIH3T3 34 � 2 41 � 3 63 � 5 67 � 4
Values are mean � SD.
Modifications of NF-YA Protein
Vol. 19, December 2008 5211

pathway negatively impacts on NF-Y transcriptional activ-ity. In good agreement with this, we observed that theNF-YA fraction in the nucleus, where it is suppose to act astranscription factor, is more acetylated than the cytoplasmicone, whereas the ubiquitylated form is prevalently stored inthe cytoplasm. Moreover, we observed that the nuclei ofpremitotic cells contain largely acetylated NF-YA protein,whereas in postmitotic cells, where NF-Y does not exert itsactivity, NF-YA protein is only ubiquitylated.
Finally, we observed that degradation-resistant NF-YAmutants increase cell proliferation. As discussed above,these mutants also sustain expression of NF-Y target genes.Thus, one possibility is that these mutants increase cellproliferation through the up-regulation of NF-Y targetgenes. NF-Y could serve as a common transcription factorfor an increasing number of cell cycle control genes (Elkon etal., 2003). This suggests that the more stable NF-YA proteincould also sustain the expression of other genes involved incell cycle progression, known to be targets of NF-Y. Inagreement with this, the levels of NF-Y activity in the cellsstrongly influences cell proliferation. It has been reportedthat inhibition of NF-YA expression blocks cell cycle pro-gression in G1 and G2 (Hu and Maity, 2000) and the knockout of the NF-YA subunit in mice leads to embryo lethality(Bhattacharya et al., 2003). Thus, there may be specific mech-anisms for limiting free NF-Y levels, failure of which wouldcompromise cell survival and/or homeostasis. The destruc-tion of NF-YA by Ub-mediated proteolysis could be one ofthe mechanisms that the cells have evolved to keep theactivity of NF-Y tightly regulated.
ACKNOWLEDGMENTS
The authors thank Dirk Bohmann for generously providing ubiquitin plas-mids, Silvia Bacchetti for editing the manuscript, Maria Pia Gentileschi fortechnical advice, and Vincenzo Giusti for computing assistance. This workhas been partially supported by grants from Associazione Italiana Ricerca sulCancro (AIRC), Ministero della Sanita’ (ICS-120.4/RA00-90; R.F.02/184), andISS-ACC to G.P. and from AIRC, CARIPLO_NOBEL, and COFIN to R.M.
REFERENCES
Aberle, H., Bauer, A., Stappert, J., Kispert, A., and Kemler, R. (1997). Beta-catenin is a target for the ubiquitin-proteasome pathway. EMBO J. 16, 3797–3804.
Bhattacharya, A., Deng, J. M., Zhang, Z., Behringer, R., de Crombrugghe, B.,and Maity, S. N. (2003). The B subunit of the CCAAT box binding transcrip-tion factor complex CBF/NF-Y) is essential for early mouse development andcell proliferation. Cancer Res. 63, 8167–8172.
Bolognese, F., Wasner, F., Dohna, C. L., Gurtner, A., Ronchi, A., Muller, H.,Manni, I., Mossner, J., Piaggio, G., Mantovani, R., and Engeland, K. (1999).The cyclin B2 promoter depends on NF-Y, a trimer whose CCAAT-bindingactivity is cell cycle regulated. Oncogene 18, 1845–1853.
Brasier, A. R., Tate, J. E., and Habener, J. F. (1989). Optimized use of the fireflyluciferase assay as a reporter gene in mammalian cell lines. Biotechniques 7,1116–1122.
Caretti, G., Motta, M. C., and Mantovani, R. (1999). NF-Y associates withH3–H4 tetramers and octamers by multiple mechanisms. Mol. Cell. Biol. 19,8591–8603.
Caretti, G., Salsi, V., Vecchi, C., Imbriano, C., and Mantovani, R. (2003).Dynamic recruitment of NF-Y and histone acetyltransferases on cell-cyclepromoters. J. Biol. Chem. 278, 30435–30440.
Chan, H. M., and La Thangue, N. B. (2001). p300/CBP proteins: HATs fortranscriptional bridges and scaffolds. J. Cell Sci. 4, 2363–2373.
Chang, Z. F., and Liu, C. J. (1994). Human thymidine kinase CCAAT-bindingprotein is NF-Y, whose A subunit expression is serum-dependent in humanIMR-90 diploid fibroblasts. J. Biol. Chem. 26, 17893–17898.
Chen, Z. J., Parent, L., and Maniatis, T. (1996). Site-specific phosphorylation ofIkappaBalpha by a novel ubiquitination-dependent protein kinase activity.Cell 84, 853–862.
Ciechanover, A., Orian, A., and Schwartz, A. L. (2000). Ubiquitin-mediatedproteolysis: biological regulation via destruction. BioEssays 22, 442–451.
Collins, G. A., and Tansey, W. P. (2006). The proteasome: a utility tool fortranscription. Curr. Opin. Genet Dev. 16, 197–202.
Currie, R. A. (1998). Biochemical characterization of the NF-Y transcriptionfactor complex during B lymphocyte development. J. Biol. Chem. 273, 1430–1434.
Dennis, A. P., Lonard, D. M., Nawaz, Z., and O’Malley, B. W. (2005). Inhibi-tion of the 26S proteasome blocks progesterone receptor-dependent transcrip-tion through failed recruitment of RNA polymerase II. J. Steroid. Biochem.Mol. Biol. 94, 337–346.
Di Agostino, S., Strano, S., Emiliozzi, V., Zerbini, V., Mottolese, M., Sacchi, A.,Blandino, G., and Piaggio, G. (2006). Gain of function of mutant p53, themutant p53/NF-Y protein complex reveals an aberrant transcriptional mech-anism of cell cycle regulation. Cancer Cell 10, 191–202.
Elkon, R., Linhart, C., Sharan, R., Shamir, R., and Shiloh, Y. (2003). Genome-wide in silico identification of transcriptional regulators controlling the cellcycle in human cells. Genome Res. 13, 773–780.
Farina, A., Manni, I., Fontemaggi, G., Tiainen, M., Cenciarelli, C., Bellorini, M.,Mantovani, R., Sacchi, A., and Piaggio, G. (1999). Down-regulation of cyclinB1 gene transcription in terminally differentiated skeletal muscle cells isassociated with loss of functional CCAAT-binding NF-Y complex. Oncogene18, 2818–2827.
Fujimuro, M., and Yokosawa, H. (2005). Production of antipolyubiquitinmonoclonal antibodies and their use for characterization and isolation ofpolyubiquitinated proteins. Methods Enzymol. 399, 75–86.
Gong, J., Zhu, J., Goodman, O. B., Jr., Pestell, R. G., Schlegel, P. N., Nanus,D. M., and Shen, R. (2006). Activation of p300 histone acetyltransferaseactivity and acetylation of the androgen receptor by bombesin in prostatecancer cells. Oncogene 25, 2011–2021.
Gurtner, A., Fuschi, P., Magi, F., Colussi, C., Gaetano, C., Dobbelstein, M.,Sacchi, A., and Piaggio, G. (2008). NF-Y dependent epigenetic modificationsdiscriminate between proliferating and postmitotic tissue. PlosOne 3, e2047.
Gurtner, A., Manni, I., Fuschi, P., Mantovani, R., Guadagni, F., Sacchi, A., andPiaggio, G. (2003). Requirement for down-regulation of the CCAAT-bindingactivity of the NF-Y transcription factor during skeletal muscle differentiation.Mol. Biol. Cell 14, 2706–2715.
Hateboer, G., Kerkhoven, R. M., Shvarts, A., Bernards, R., and Beijersbergen,R. L. (1996). Degradation of E2F by the ubiquitin-proteasome pathway: reg-ulation by retinoblastoma family proteins and adenovirus transforming pro-teins. Genes Dev. 10, 2960–2970.
Hershko, A., and Ciechanover, A. (1998). The ubiquitin system. Annu. Rev.Biochem. 67, 425–749.
Hochstrasser, M. (1996). Ubiquitin-dependent protein degradation. Annu.Rev. Genet. 30, 405–439.
Hofmann, F., Martelli, F., Livingston, D. M., and Wang, Z. (1996). The reti-noblastoma gene product protects E2F-1 from degradation by the ubiquitin-proteasome pathway. Genes Dev. 10, 2949–2959.
Hu, Q., and Maity, S. N. (2000). Stable expression of a dominant negativemutant of CCAAT binding factor/NF-Y in mouse fibroblast cells resulting inretardation of cell growth and inhibition of transcription of various cellulargenes. J. Biol. Chem. 275, 4435–4444.
Imbriano, C., Gurtner, A., Cocchiarella, F., Di Agostino, S., Basile, V., Gostissa,M., Dobbelstein, M., Del Sal, G., Piaggio, G., and Mantovani, R. (2005). Directp53 transcriptional repression: in vivo analysis of CCAAT-containing G2/Mpromoters. Mol. Cell. Biol. 25, 3737–3751.
Jin, S., and Scotto, K. W. (1998). Transcriptional regulation of the MDR1 geneby histone acetyltransferase and deacetylase is mediated by NF-Y. Mol. Cell.Biol. 18, 4377–4384.
Kodadek, T., Sikder, D., and Nalley, K. (2006). Keeping transcriptional acti-vators under control. Cell 127, 261–264.
Korner, K., Jerome, V., Schmidt, T., and Muller, R. (2001). Cycle regulation ofthe murine cdc25B promoter: essential role for nuclear factor-Y and a proxi-mal repressor element. J. Biol. Chem. 276, 9662–9669.
Krogan, N. J., Lam, M. H., Fillingham, J., Keogh, M. C., Gebbia, M., Li, J.,Datta, N., Cagney, G., Buratowsky, S., Emili, S., and Greenblatt, J. F. (2004).Proteasome involvement in the repair of DNA double-strand breaks. Mol.Cell 16, 1027–1034.
Li, Q., Herrler, M., Landsberger, N., Kaludov, N., Ogryzko, V. V., Nakatani,Y., and Wolffe, A. P. (1998). Xenopus NF-Y pre-sets chromatin to potentiatep300 and acetylation-responsive transcription from the Xenopus hsp70 pro-moter in vivo. EMBO J. 17, 6300–6315.
I. Manni et al.
Molecular Biology of the Cell5212

Liberati, C., di Silvio, A., Ottolenghi, S., and Mantovani, R. (1999). NF-Ybinding to twin CCAAT boxes: role of Q-rich domains and histone foldhelices. J. Mol. Biol. 29, 1441–1455.
Lipford, J. R., Smith, G. T., Chi, Y., and Deshaies, R. J. (2005). A putativestimulatory role for turnover in gene expression. Nature 438, 113–116.
Liu, Y. V., Baek, J. H., Zhang, H., Diez, R., Cole, R. N., and Semenza, G. L.(2007). RACK1 competes with HSP90 for binding to HIF-1� and is requiredfor O2-independent and HSP90 inhibitor-induced degradation of HIF-1�.Mol. Cell 25, 207–217.
Lo, R. S., and Massague, J. (1999). Ubiquitin-dependent degradation of TGF-beta- activated Smad2. Nat. Cell Biol. 1, 472–478.
Manni, I., Mazzaro, G., Gurtner, A., Mantovani, R., Haugwitz, U., Krause, K.,Engeland, K., Sacchi, A., Soddu, S., and Piaggio, G. (2001). NF-Y mediates thetranscriptional inhibition of the cyclin B1, cyclin B2, and cdc25C promotersupon induced G2 arrest. J. Biol. Chem. 276, 5570–5576.
Mantovani, R., Pessara, U., Tronche, F., Li, X. Y., Knapp, A. M., Pasquali, J. L.,Benoist, C., and Mathis, D. (1992). Monoclonal antibodies to NF-Y define itsfunction in MHC class II and albumin gene transcription. EMBO J. 11, 3315–3322.
Mantovani, R. (1998). A survey of 178 NF-Y binding CCAAT boxes. NucleicAcids Res. 26, 1135–1143.
Mantovani, R. (1999). The molecular biology of the CCAAT boxes. NucleicAcids Res. 26, 1135–1143.
Marziali, G., Perrotti, E., Ilari, R., Testa, U., Coccia, E. M., and Battistini, A.(1997). Transcriptional regulation of the ferritin heavy-chain gene: the activityof the CCAAT binding factor NF-Y is modulated in heme-treated Friendleukemia cells and during monocyte-to-macrophage differentiation. Mol. Cell.Biol. 17, 1387–1395.
Muratani, M., Kung, C., Shokat, K. M., and Tansey, W. P. (2005). The F boxprotein Dsg1/Mdm30 is a transcriptional coactivator that stimulates Gal4turnover and cotranscriptional mRNA processing. Cell 120, 887–899.
Orford, K., Crockett, C., Jensen, J. P., Weissman, A. M., and Byers, S. W.(1997). Serine phosphorylation-regulated ubiquitination and degradation ofbeta-catenin. J. Biol. Chem. 272, 24735–24738.
Park, S. H. et al. (2002). Transcriptional regulation of the transforming growthfactor beta type II receptor gene by histone acetyltransferase and deacetylaseis mediated by NF-Y in human breast cancer cells. J. Biol. Chem. 277, 5168–5174.
Puca, R., Nardinocchi, L., and D’Orazi, G. (2008). Regulation of vascularendothelial growth factor expression by homeodomain-interacting proteinkinase-2. J. Exp. Clin. Cancer Res. 27, 22–28.
Rock, K. L., Gramm, C., Rothstein, L., Clark, K., Stein, R., Dick, L., Hwang, D.,and Goldberg, A. L. (1994). Inhibitors of the proteasome block the degrada-
tion of most cell proteins and the generation of peptides presented on MHCclass I molecules. Cell 78, 761–771.
Salsi, V., Caretti, G., Wasner, M., Reinhard, W., Haugwitz, U., Engeland, K.and Mantovani, R. (2003). Interactions between p300 and multiple NF-Ytrimers govern cyclin B2 promoter function. J. Biol. Chem. 278, 6642–6650.
Scheffner, M., Werness, B. A., Huibregtse, J. M., Levine, A. J., and Howley,P. M. (1990). The E6 oncoprotein encoded by human papillomavirus types 16and 18 promotes the degradation of p53. Cell 63, 1129–1136.
Shcherbik, N., and Haines, D. S. (2004). Ub on the move. J. Cell. Biochem. 93,11–19.
Somesh, B. P., Reid J., Liu, W.-F., Søgaard, T.M.M., Erdjument-Bromage, H.,Tempst, P. and Svejstrup, J. Q. (2005). Multiple mechanisms confining RNApolymerase II ubiquitination to polymerases undergoing transcriptional ar-rest. Cell 121, 913–923.
Sterner, D. E., and Berger, S. H. (2000). Acetylation of histones and transcrip-tion-related factors. Mol. Biol. Rev. 64, 435–459.
Thrower, J. S., Hoffman, L., Rechsteiner, M., and Pickart, C. M. (2000). Rec-ognition of the polyubiquitin proteolytic signal. EMBO J. 19, 94–102.
Treier, M., Staszewski, L. M., and Bohmann, D. (1994). Ubiquitin-dependentc-Jun degradation in vivo is mediated by the delta domain. Cell 78, 787–798.
Wang, K.K.W. (1990). Developing selective inhibitors of calpain. Trends Phar-macol. Sci. 11, 139–142.
Wei, W., and Yu, X. D. (2007). Hypoxia-inducible factors: crosstalk betweentheir protein stability and protein degradation. Cancer Lett. 257, 145–156.
Zemzoumi, K., Frontini, M., Bellorini, M., and Mantovani, R. (1999). NF-Yhistone fold alpha1 helices help impart CCAAT specificity. J. Mol. Biol. 19,327–337.
Zhang, W., and Bieker, J. (1999). Acetylation and modulation of erythroidKruppel-like factor (EKLF) activity by interaction with histone acetyltrans-ferases. Proc. Natl. Acad. Sci. USA 95, 9855–9860.
Zhu, H., Kavsak, P., Abdollah, S., Wrana, J. L., and Thomsen, G. H. (1999). ASMAD ubiquitin ligase targets the BMP pathway and affects embryonicpattern formation. Nature 400, 687–693.
Zwicker, J., Gross, C., Lucibello, F. C., Truss, M., Ehlert, F., Engeland, K., andMuller, R. (1995a). Cell cycle regulation of cdc25C transcription is mediatedby the periodic repression of the glutamine-rich activators NF-Y and Sp1.Nucleic Acids Res. 23, 3822–3830.
Zwicker, J., Lucibello, F. C., Wolfraim, L. A., Gross, C., Truss, M., Engeland,K., and Muller, R. (1995b). Cell cycle regulation of the cyclin A, cdc25C andcdc2 genes is based on a common mechanism of transcriptional repression.EMBO J. 14, 4514–4522.
Modifications of NF-YA Protein
Vol. 19, December 2008 5213




















