
Population dynamics of Rattus argentiventer , Rattus losea , and Rattus rattus inhabiting a...
10
ORIGINAL ARTICLE Peter R. Brown Nguyen Phu Tuan Grant R. Singleton Dao Thi Hue Phung Thi Hoa Phi Thi Thu Ha Tran Quang Tan Nguyen Van Tuat Population dynamics of Rattus argentiventer, Rattus losea, and Rattus rattus inhabiting a mixed-farming system in the Red River Delta, Vietnam Received: 20 April 2005 / Accepted: 25 July 2005 / Published online: 15 September 2005 ȑ The Society of Population Ecology and Springer-Verlag Tokyo 2005 Abstract Rodent pests cause significant damage to lowland irrigated rice crops in the Red River Delta of northern Vietnam. Data from a 4-year study were examined to look at the population dynamics of the ricefield rat, Rattus argentiventer (representing 50% of captures), the lesser ricefield rat, Rattus losea (30% of captures), and the black rat, Rattus rattus complex (9% of captures) that inhabit the irrigated mixed-cropping system. We tested the hypothesis that these rodent spe- cies were breeding in response to the availability of high- quality food provided by crops rather than in response to rainfall. The abundance of rodents fluctuated annu- ally, with a main peak following the spring rice crop, and a secondary peak following the summer rice crop. There was a strong relationship between the monthly abundance of rats and rainfall, but a weak relationship between monthly rates of increase and rainfall. There were distinct peaks in breeding activity during the reproductive stages of the rice crops suggesting that changes in crop stages were more important than rainfall in this seasonal, but irrigated agroecosystem. The modal litter size for R. argentiventer was 8 (mean of 8.67±0.20 SE, range 2–16), where the mode for R. losea, was 7 (mean of 7.32±0.15 SE, range 3–14). Management of these species needs to be conducted prior to the onset of the main breeding seasons. Keywords Black rat Breeding Interspecific competition Lesser ricefield rat Lowland irrigated rice Ricefield rat Introduction Rodents are a significant problem for agriculture in Vietnam, particularly for lowland irrigated rice in the two main rice-production areas in Vietnam—the Mekong and Red River Deltas (Singleton and Petch 1994; Brown et al. 1999; Singleton 2003). Rodents damage 20–30% of the annual pre-harvest rice crop. It has been argued that if the damage can be reduced to 5% there would be enough rice to feed an additional 7.8 million Vietnamese people/an- num (Singleton 2003). It is therefore desirable to reduce the impact of rodents on rice crops, to increase food security, and improve the economic stability for the country through increased exports. The ricefield rat, Rattus argentiventer (Robinson and Kloss) is recognized as the main rodent pest in the lowland irrigated rice-growing areas of Vietnam (Brown et al. 1999; Lan et al. 2003), but other sympatric species include the lesser rice field rat (Rattus losea, Swinhoe), the black rat (Rattus rattus complex; for discussion on taxonomy see Aplin et al. 2003b), the brown rat (Rattus norvegicus, Berkenhout), the Pacific rat (Rattus exulans, Peale), bandicoot rats (Bandicota indica, Bechstein and Bandicota savilei, Thomas), and Mus species. These species also are likely to damage crops. Populations of R. argentiventer have been studied in Malaysia and Indonesia (Lam 1982; Tristiani et al. 1998; Leung et al. 1999; Jacob et al. 2003) where they comprise >95% of rodents inhabiting rice fields. In Vietnam, another population of R. argentiventer has been studied (Brown et al. 1999), and little is known about the population dynamics of R. losea and R. rattus in intensive rice cropping systems, especially in terms of the factors that drive changes in abundance and breeding. P. R. Brown (&) G. R. Singleton CSIRO Sustainable Ecosystems, GPO Box 284, 2601 Canberra, ACT, Australia E-mail: [email protected] Tel.: +61-2-62421562 Fax: +61-2-62421505 N. P. Tuan D. T. Hue P. T. Hoa P. T. T. Ha T. Q. Tan N. Van Tuat National Institute of Plant Protection, Hanoi, Vietnam Present address: G. R. Singleton International Rice Research Institute, Metro Manila, Philippines Popul Ecol (2005) 47:247–256 DOI 10.1007/s10144-005-0228-x
Transcript of Population dynamics of Rattus argentiventer , Rattus losea , and Rattus rattus inhabiting a...

ORIGINAL ARTICLE
Peter R. Brown Æ Nguyen Phu Tuan
Grant R. Singleton Æ Dao Thi Hue Æ Phung Thi Hoa
Phi Thi Thu Ha Æ Tran Quang Tan Æ Nguyen Van Tuat
Population dynamics of Rattus argentiventer, Rattus losea,and Rattus rattus inhabiting a mixed-farming systemin the Red River Delta, Vietnam
Received: 20 April 2005 / Accepted: 25 July 2005 / Published online: 15 September 2005� The Society of Population Ecology and Springer-Verlag Tokyo 2005
Abstract Rodent pests cause significant damage tolowland irrigated rice crops in the Red River Delta ofnorthern Vietnam. Data from a 4-year study wereexamined to look at the population dynamics of thericefield rat, Rattus argentiventer (representing 50% ofcaptures), the lesser ricefield rat, Rattus losea (30% ofcaptures), and the black rat, Rattus rattus complex (9%of captures) that inhabit the irrigated mixed-croppingsystem. We tested the hypothesis that these rodent spe-cies were breeding in response to the availability of high-quality food provided by crops rather than in responseto rainfall. The abundance of rodents fluctuated annu-ally, with a main peak following the spring rice crop,and a secondary peak following the summer rice crop.There was a strong relationship between the monthlyabundance of rats and rainfall, but a weak relationshipbetween monthly rates of increase and rainfall. Therewere distinct peaks in breeding activity during thereproductive stages of the rice crops suggesting thatchanges in crop stages were more important than rainfallin this seasonal, but irrigated agroecosystem. The modallitter size for R. argentiventer was 8 (mean of 8.67±0.20SE, range 2–16), where the mode for R. losea, was 7(mean of 7.32±0.15 SE, range 3–14). Management ofthese species needs to be conducted prior to the onset ofthe main breeding seasons.
Keywords Black rat Æ Breeding Æ Interspecificcompetition Æ Lesser ricefield rat ÆLowland irrigated rice Æ Ricefield rat
Introduction
Rodents are a significant problem for agriculture inVietnam, particularly for lowland irrigated rice in the twomain rice-production areas in Vietnam—theMekong andRed River Deltas (Singleton and Petch 1994; Brown et al.1999; Singleton 2003). Rodents damage 20–30% of theannual pre-harvest rice crop. It has been argued that if thedamage can be reduced to 5% there would be enough riceto feed an additional 7.8 million Vietnamese people/an-num (Singleton 2003). It is therefore desirable to reducethe impact of rodents on rice crops, to increase foodsecurity, and improve the economic stability for thecountry through increased exports.
The ricefield rat, Rattus argentiventer (Robinson andKloss) is recognized as the main rodent pest in thelowland irrigated rice-growing areas of Vietnam (Brownet al. 1999; Lan et al. 2003), but other sympatric speciesinclude the lesser rice field rat (Rattus losea, Swinhoe),the black rat (Rattus rattus complex; for discussion ontaxonomy see Aplin et al. 2003b), the brown rat (Rattusnorvegicus, Berkenhout), the Pacific rat (Rattus exulans,Peale), bandicoot rats (Bandicota indica, Bechstein andBandicota savilei, Thomas), and Mus species. Thesespecies also are likely to damage crops. Populations ofR. argentiventer have been studied in Malaysia andIndonesia (Lam 1982; Tristiani et al. 1998; Leung et al.1999; Jacob et al. 2003) where they comprise >95% ofrodents inhabiting rice fields. In Vietnam, anotherpopulation of R. argentiventer has been studied (Brownet al. 1999), and little is known about the populationdynamics of R. losea and R. rattus in intensive ricecropping systems, especially in terms of the factors thatdrive changes in abundance and breeding.
P. R. Brown (&) Æ G. R. SingletonCSIRO Sustainable Ecosystems,GPO Box 284, 2601 Canberra, ACT, AustraliaE-mail: [email protected].: +61-2-62421562Fax: +61-2-62421505
N. P. Tuan Æ D. T. Hue Æ P. T. Hoa Æ P. T. T. HaT. Q. Tan Æ N. Van TuatNational Institute of Plant Protection, Hanoi, Vietnam
Present address: G. R. SingletonInternational Rice Research Institute, Metro Manila, Philippines
Popul Ecol (2005) 47:247–256DOI 10.1007/s10144-005-0228-x

The distribution of R. argentiventer includes lowlandcropping areas from northern Vietnam to southernVietnam, through the lower reaches of the MekongRiver Valley into Cambodia, lowland areas of Thailand,all of Malaysia, most islands of Indonesia and someislands of the Philippines (Marshall 1977; Aplin et al.2003a). R. losea generally is found only in northern andsouthern Vietnam, Cambodia, Thailand, southern partsof China and in Taiwan. The geographic distribution ofthe various genetic forms of the R. rattus complex is stillto be resolved (Aplin et al. 2003b). The main economicimpact of these species is damage to crops, but they alsorepresent a source of disease for humans. For example,R. losea can carry ticks infected with Lyme disease (Shihet al. 1998) and harbours hanta virus (Nitatpattana et al.2002). In the central highlands of Vietnam, R. rattus is areservoir of plague (Yersinia pestis; Suntsov et al. 1997).Therefore, understanding population dynamics of thesespecies may assist in reducing damage to crops, andlessening the chance of disease transmission to humans.
Farmers use a range of control techniques to reducerodent damage (Tuan et al. 2003). There is a growingrecognition that these conventional methods are notmaintaining rodent populations below an economicthreshold (when costs of control outweigh yield gains),and so ecologically based rodent management practicesare being developed to reduce the impact of these spe-cies on rice crops (Singleton 1997; Singleton and Brown1999). This strategy requires a good understanding ofthe population dynamics of the principal pest species.The objective of the current study was to develop abetter understanding of the population dynamics of thethree main pest species of rodents in a lowland mixedrice agro-ecosystem in the Red River Delta, Vietnam, asa basis for developing ecologically based managementpractices. We tested the hypothesis that these rodentspecies were breeding in response to the availability ofhigh-quality food provided by crops rather than inresponse to rainfall. We examine population dynamicsof R. argentiventer, R. losea and to a lesser extent,R. rattus, over 4 years in Vinh Phuc Province. We re-port on temporal changes in population abundance,rate of population increase in relation to rainfall,breeding dynamics and percentage of juveniles in thepopulation.
Materials and methods
Study site
The study was located near Tien Phong village, Me LinhDistrict, Vinh Phuc Province, 40 kmnorth ofHanoi in theRed River Delta of northern Vietnam (21�08¢N,105�45¢E). Two sites were used; these were untreatedcontrol sites of a larger study assessing methods for con-trolling rodents. Each site was approximately 100 ha andthe distance between each sitewas 1 km.This locationwaschosen because an outbreak of rodents in the area in 1997/
1998 substantially damaged rice crops (N. P. Tuan,unpublished data). The principal crop grown in the area isrice. There are two main rice-growing seasons each year,spring (transplanted late February and harvested midJune) and summer (transplanted mid July and harvestedlate September). Other crops, grown throughout the year,are vegetables (broccoli, cabbage, chilli, kohlrabi, onion,pumpkin, and tomato) and flower crops (chrysanthe-mum, rose). In March 2002, 51% of the land was undervegetable crops, 20% was under rice crops, 17% wasunder flower crops, 10% was channel banks and road-sides, and 2% was fallow. In June 2002, 40% was underrice crops, 35%of the landwas under vegetables, 11%wasfallow, 10% was channel banks and roadsides, and 4%was under flower crops. At other times, particularlyduring the summer rice crop, up to 100% of the land wasplanted to rice.
Summers are hot and wet, and winters are cool anddry. Maximum temperatures in Hanoi are 15–25�C inspring, 30–36�C in summer, 25–30�C in autumn, and 10–15�C in winter. The annual average rainfall wasapproximately 1,675 mm, most falling between May andSeptember (summer). Rainfall was below average in1998, 1999, 2000, and 2002 (80, 93, 76, and 87% of thelong-term mean, respectively) and was above average in2001 (135% of the long-term mean). Farmers irrigatedtheir crops using water supplied by channels originatingfrom large storage dams in nearby hills.
Live-trapping
Two replicate lines of 20 live cage traps were set in eachof three main situations denoting: (1) big channel banks,>2 m high and >2 m wide carrying irrigation water; (2)medium banks, about 1 m high and 1 m wide, smallerchannels or paths between fields; and (3) small banks,<0.3 m high and <0.3 m wide separating rice fields.Trapping occurred for 4 consecutive nights for a total of480 trap nights per site per month. Traps were con-structed from open wire mesh (150 · 150 · 400 mm)with a hinged, sprung door. Traps were baited withroasted vegetables. Trapping occurred each month fromApril 1999 to November 2002. One site was trappedmonthly, the other less frequently depending on avail-ability of labour. Trapping results are presented as theabundance of rats captured each month. Upon capture,each animal was identified to species, marked with anear punch, weighed, and lengths of tail, hind foot, andear were measured to assist with identification to species.Breeding condition was assessed for females (lactating ifteats were raised and free of hair at their base; pregnancyby palpation).
Monthly records of rainfall (mm) were obtained forHanoi from monthly weather reports for Vietnam. TheHanoi weather station was approximately 30 km fromthe study sites. The relationship between rainfall andrats was assessed using two methods. Firstly, a simplelinear regression was performed on the abundance of
248

rodents and monthly rainfall. Secondly, the exponentialrate of increase per month (r; rate of change) wascalculated as r=ln(rat abundance for month t/ratabundance for month t�1). This relationship effectivelyremoves the effect of density and looks at how much thepopulation increases or decreases in response to rainfall.To ascertain whether rainfall, as a surrogate for foodsupply, was an important factor in driving the rodentpopulations, exponential rates of increase were corre-lated against monthly rainfall that was lagged by 0–6months, accumulated rainfall of 1–6 months and 6-monthaccumulated rainfall but lagged by 0–6 months.A similar approach has been used for other rodents(Pseudomys hermannsburgensis, Notomys alexis, andMus domesticus) in arid central Australia (Southgateand Masters 1996), and M. domesticus in wheat fields inAustralia (Brown and Singleton 1999). The periods oftime when populations increased and decreased werethen defined (spring increase, summer decrease, summerincrease, and winter decrease), and the rates of increaseover these periods were analysed by a one-way ANOVA.
The abundance of the two most common species,R. argentiventer and R. losea, was analysed using athree-way ANOVA using ln-transformed abundance ofrats with month, year, and species as factors. Theoccurrence of adult females breeding (lactating and/orpregnant as determined by palpation) was comparedover time using generalized linear models (analysis ofdeviance) for binomially distributed data (breeding yesor no) using the logit link. Probabilities were determinedusing the F statistic using df/residual df. The effect ofmonth, year, and species (R. argentiventer and R. losea)and all interactions were examined. Linear regressionanalyses were conducted to determine whether the pro-portion of adults in breeding condition could be pre-dicted by rainfall or crop stage. Crop stage was scoredon a scale of 1–10 (sowing/transplanting through toharvest) and analysed separately for spring and summerrice crops. This scaling of crop stage is a rough estimateof food supply and assumes that it increases linearlyfrom sowing to harvest (Tristiani and Murakami 2003).The presence of juveniles trapped in the population wascompared over time with generalized linear models for
binomially distributed data (juveniles yes or no). Thesame analysis was conducted for juveniles as for adultfemales breeding, with the addition of sex in the model.
Kill-trapping
Kill-trapping was conducted each month to collect adultfemale R. argentiventer and R. losea to assess breedingcondition in more detail. Kill-traps and live-traps werelocated at least 200 m away from the regular trappinglines. The aim was to collect at least 15 adult femaleR. argentiventer and R. losea each month where capturerates permitted. After capture, rats were weighed andmeasured and assessed for breeding condition as for live-trapped animals, then autopsied to ascertain conditionof uterus, number of embryos, trimester of embryodevelopment, and number and sets of uterine scars.
The occurrence of adult females breeding (presence ofembryos in uterus) was compared over time using thesame analysis as used for live-captured animals above.Litter sizes were compared over time using generalizedlinear models for Gaussian distributed data (ln-trans-formed counts of embryo number per pregnant female).Probabilities were determined using the F statistic usingdf/residual df. The effect of month, year, and species(R. argentiventer and R. losea) and all interactions wereexamined.
Results
Abundance of rats
The overall abundance of rats during this study was low,with the majority of trapping sessions yielding <40animals/480 trap nights per month. A total of 411 ratswere captured from 20,340 trap nights from site 1 (44trapping sessions) and 458 rats were captured from12,420 trap nights from site 2 (27 trapping sessions).Abundance peaked in July 1999 at 52 and 82 rats forsites 1 and 2, respectively. The abundance of rats wasstrongly related to the cycle of cropping and was similar
Month
A J A O D F A J A O D F A J A O D F A J A O D
Mon
thly
rai
nfal
l (m
m)
0
200
400
600
800
Abu
ndan
ce o
f rat
s
0
20
40
60
80
100
Winter crop Winter crop Winter crop
1999 2000 2001 2002
M J S N J M M J S N J M M J S N J M M J S N
Fig. 1 Average abundance(±SE) of rats from both sitesand monthly rainfall (mm,vertical bars) from Hanoi(30 km from study site). Theshaded horizontal bars at thebottom of the graph representthe approximate timing of thespring and summer rice cropsand winter vegetable crops
249
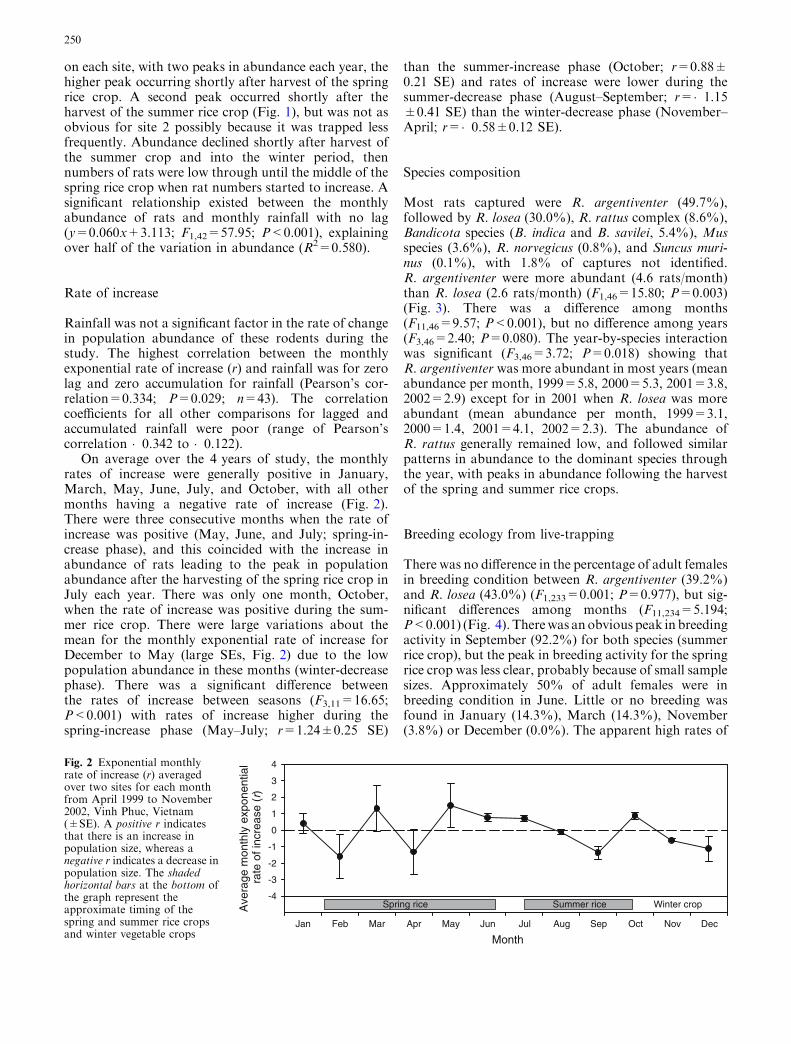
on each site, with two peaks in abundance each year, thehigher peak occurring shortly after harvest of the springrice crop. A second peak occurred shortly after theharvest of the summer rice crop (Fig. 1), but was not asobvious for site 2 possibly because it was trapped lessfrequently. Abundance declined shortly after harvest ofthe summer crop and into the winter period, thennumbers of rats were low through until the middle of thespring rice crop when rat numbers started to increase. Asignificant relationship existed between the monthlyabundance of rats and monthly rainfall with no lag(y=0.060x+3.113; F1,42=57.95; P<0.001), explainingover half of the variation in abundance (R2=0.580).
Rate of increase
Rainfall was not a significant factor in the rate of changein population abundance of these rodents during thestudy. The highest correlation between the monthlyexponential rate of increase (r) and rainfall was for zerolag and zero accumulation for rainfall (Pearson’s cor-relation=0.334; P=0.029; n=43). The correlationcoefficients for all other comparisons for lagged andaccumulated rainfall were poor (range of Pearson’scorrelation �0.342 to �0.122).
On average over the 4 years of study, the monthlyrates of increase were generally positive in January,March, May, June, July, and October, with all othermonths having a negative rate of increase (Fig. 2).There were three consecutive months when the rate ofincrease was positive (May, June, and July; spring-in-crease phase), and this coincided with the increase inabundance of rats leading to the peak in populationabundance after the harvesting of the spring rice crop inJuly each year. There was only one month, October,when the rate of increase was positive during the sum-mer rice crop. There were large variations about themean for the monthly exponential rate of increase forDecember to May (large SEs, Fig. 2) due to the lowpopulation abundance in these months (winter-decreasephase). There was a significant difference betweenthe rates of increase between seasons (F3,11=16.65;P<0.001) with rates of increase higher during thespring-increase phase (May–July; r=1.24±0.25 SE)
than the summer-increase phase (October; r=0.88±0.21 SE) and rates of increase were lower during thesummer-decrease phase (August–September; r=�1.15±0.41 SE) than the winter-decrease phase (November–April; r=�0.58±0.12 SE).
Species composition
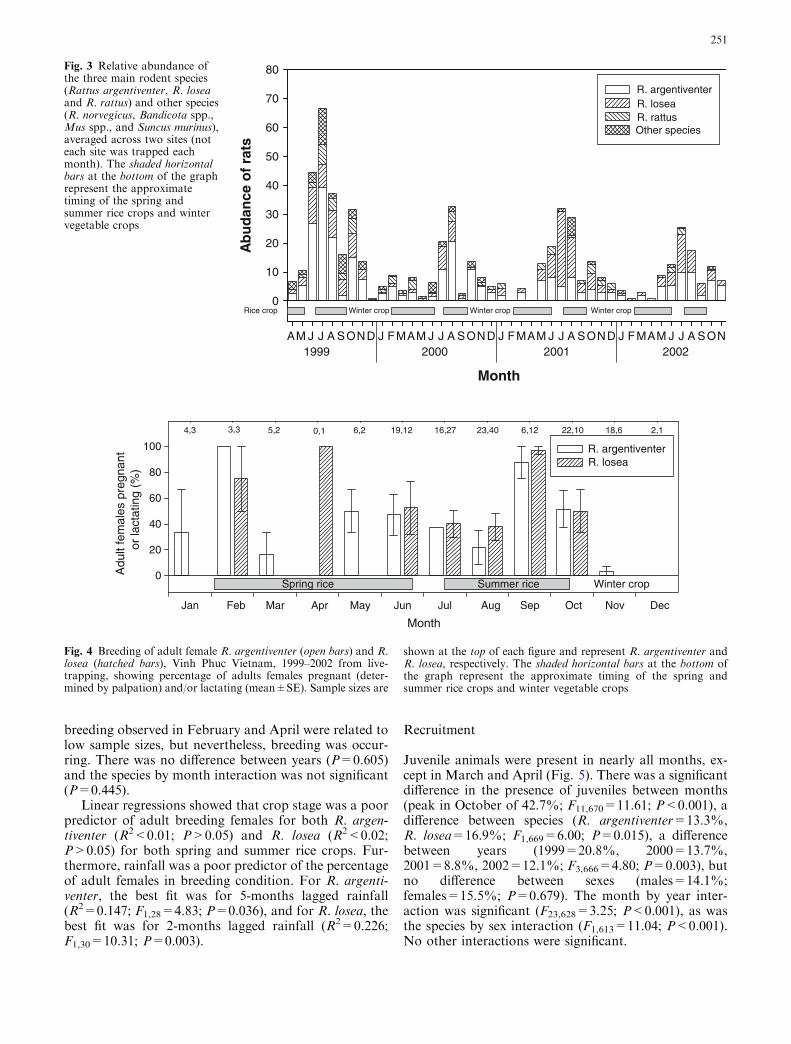
Most rats captured were R. argentiventer (49.7%),followed by R. losea (30.0%), R. rattus complex (8.6%),Bandicota species (B. indica and B. savilei, 5.4%), Musspecies (3.6%), R. norvegicus (0.8%), and Suncus muri-nus (0.1%), with 1.8% of captures not identified.R. argentiventer were more abundant (4.6 rats/month)than R. losea (2.6 rats/month) (F1,46=15.80; P=0.003)(Fig. 3). There was a difference among months(F11,46=9.57; P<0.001), but no difference among years(F3,46=2.40; P=0.080). The year-by-species interactionwas significant (F3,46=3.72; P=0.018) showing thatR. argentiventer was more abundant in most years (meanabundance per month, 1999=5.8, 2000=5.3, 2001=3.8,2002=2.9) except for in 2001 when R. losea was moreabundant (mean abundance per month, 1999=3.1,2000=1.4, 2001=4.1, 2002=2.3). The abundance ofR. rattus generally remained low, and followed similarpatterns in abundance to the dominant species throughthe year, with peaks in abundance following the harvestof the spring and summer rice crops.
Breeding ecology from live-trapping
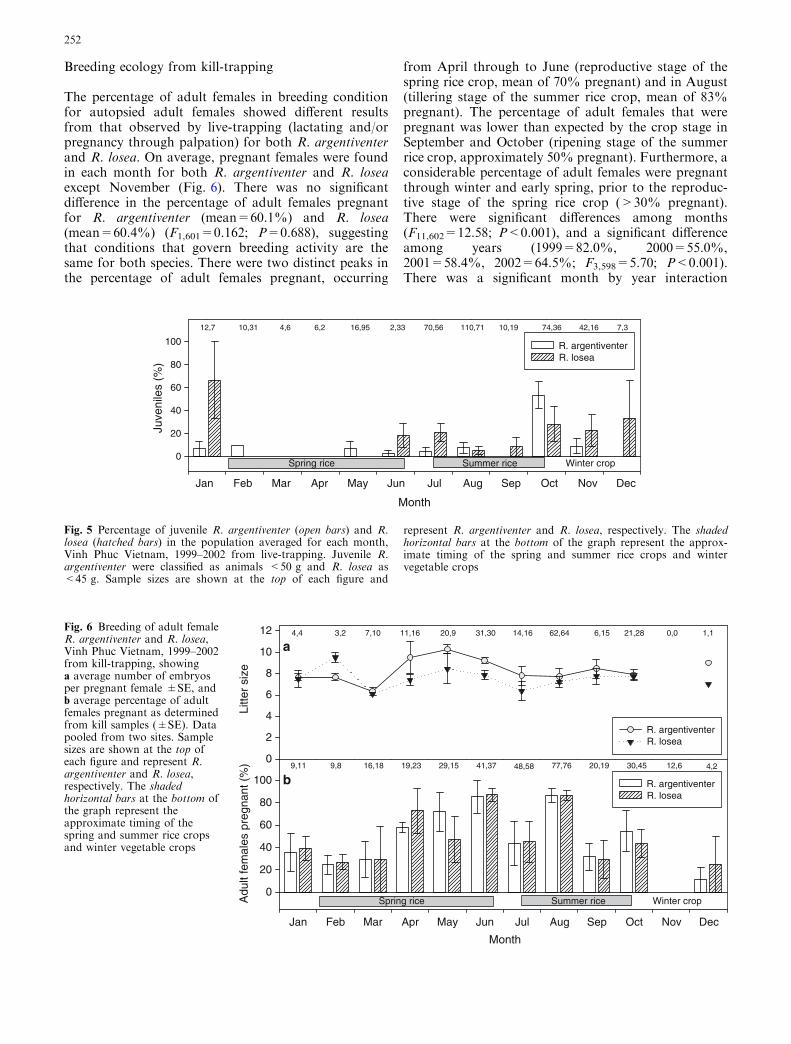
There was no difference in the percentage of adult femalesin breeding condition between R. argentiventer (39.2%)and R. losea (43.0%) (F1,233=0.001; P=0.977), but sig-nificant differences among months (F11,234=5.194;P<0.001) (Fig. 4). Therewas anobvious peak in breedingactivity in September (92.2%) for both species (summerrice crop), but the peak in breeding activity for the springrice crop was less clear, probably because of small samplesizes. Approximately 50% of adult females were inbreeding condition in June. Little or no breeding wasfound in January (14.3%), March (14.3%), November(3.8%) or December (0.0%). The apparent high rates of
Month
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Ave
rage
mon
thly
exp
onen
tial
rate
of i
ncre
ase
(r)
-4
-3
-2
-1
0
1
2
3
4
Winter cropSpring rice Summer rice
Fig. 2 Exponential monthlyrate of increase (r) averagedover two sites for each monthfrom April 1999 to November2002, Vinh Phuc, Vietnam(±SE). A positive r indicatesthat there is an increase inpopulation size, whereas anegative r indicates a decrease inpopulation size. The shadedhorizontal bars at the bottom ofthe graph represent theapproximate timing of thespring and summer rice cropsand winter vegetable crops
250
breeding observed in February and April were related tolow sample sizes, but nevertheless, breeding was occur-ring. There was no difference between years (P=0.605)and the species by month interaction was not significant(P=0.445).
Linear regressions showed that crop stage was a poorpredictor of adult breeding females for both R. argen-tiventer (R2<0.01; P>0.05) and R. losea (R2<0.02;P>0.05) for both spring and summer rice crops. Fur-thermore, rainfall was a poor predictor of the percentageof adult females in breeding condition. For R. argenti-venter, the best fit was for 5-months lagged rainfall(R2=0.147; F1,28=4.83; P=0.036), and for R. losea, thebest fit was for 2-months lagged rainfall (R2=0.226;F1,30=10.31; P=0.003).
Recruitment
Juvenile animals were present in nearly all months, ex-cept in March and April (Fig. 5). There was a significantdifference in the presence of juveniles between months(peak in October of 42.7%; F11,670=11.61; P<0.001), adifference between species (R. argentiventer=13.3%,R. losea=16.9%; F1,669=6.00; P=0.015), a differencebetween years (1999=20.8%, 2000=13.7%,2001=8.8%, 2002=12.1%; F3,666=4.80; P=0.003), butno difference between sexes (males=14.1%;females=15.5%; P=0.679). The month by year inter-action was significant (F23,628=3.25; P<0.001), as wasthe species by sex interaction (F1,613=11.04; P<0.001).No other interactions were significant.
Month
A J A O D F A J A O D F A J A O D F A J A O
Ab
ud
ance
of
rats
0
10
20
30
40
50
60
70
80
Other species
Rice crop Winter crop Winter crop Winter crop
1999 2000 2001 2002
R. argentiventerR. loseaR. rattus
M J S N J M M J S N J M M J S N J M M J S N
Fig. 3 Relative abundance ofthe three main rodent species(Rattus argentiventer, R. loseaand R. rattus) and other species(R. norvegicus, Bandicota spp.,Mus spp., and Suncus murinus),averaged across two sites (noteach site was trapped eachmonth). The shaded horizontalbars at the bottom of the graphrepresent the approximatetiming of the spring andsummer rice crops and wintervegetable crops
Month
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Adu
lt fe
mal
es p
regn
ant
or la
ctat
ing
(%)
0
20
40
60
80
100
Winter cropSpring rice Summer rice
4,3 3,3 5,2 0,1 6,2 19,12 16,27 23,40 6,12 22,10 18,6 2,1
R. argentiventerR. losea
Fig. 4 Breeding of adult female R. argentiventer (open bars) and R.losea (hatched bars), Vinh Phuc Vietnam, 1999–2002 from live-trapping, showing percentage of adults females pregnant (deter-mined by palpation) and/or lactating (mean±SE). Sample sizes are
shown at the top of each figure and represent R. argentiventer andR. losea, respectively. The shaded horizontal bars at the bottom ofthe graph represent the approximate timing of the spring andsummer rice crops and winter vegetable crops
251
Breeding ecology from kill-trapping
The percentage of adult females in breeding conditionfor autopsied adult females showed different resultsfrom that observed by live-trapping (lactating and/orpregnancy through palpation) for both R. argentiventerand R. losea. On average, pregnant females were foundin each month for both R. argentiventer and R. loseaexcept November (Fig. 6). There was no significantdifference in the percentage of adult females pregnantfor R. argentiventer (mean=60.1%) and R. losea(mean=60.4%) (F1,601=0.162; P=0.688), suggestingthat conditions that govern breeding activity are thesame for both species. There were two distinct peaks inthe percentage of adult females pregnant, occurring
from April through to June (reproductive stage of thespring rice crop, mean of 70% pregnant) and in August(tillering stage of the summer rice crop, mean of 83%pregnant). The percentage of adult females that werepregnant was lower than expected by the crop stage inSeptember and October (ripening stage of the summerrice crop, approximately 50% pregnant). Furthermore, aconsiderable percentage of adult females were pregnantthrough winter and early spring, prior to the reproduc-tive stage of the spring rice crop (>30% pregnant).There were significant differences among months(F11,602=12.58; P<0.001), and a significant differenceamong years (1999=82.0%, 2000=55.0%,2001=58.4%, 2002=64.5%; F3,598=5.70; P<0.001).There was a significant month by year interaction
Month
Juve
nile
s (%
)
0
20
40
60
80
100 R. argentiventer
Winter cropSpring rice Summer rice
12,7 10,31 4,6 6,2 16,95 2,33 70,56 110,71 10,19 74,36 42,16 7,3
R. losea
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Fig. 5 Percentage of juvenile R. argentiventer (open bars) and R.losea (hatched bars) in the population averaged for each month,Vinh Phuc Vietnam, 1999–2002 from live-trapping. Juvenile R.argentiventer were classified as animals <50 g and R. losea as<45 g. Sample sizes are shown at the top of each figure and
represent R. argentiventer and R. losea, respectively. The shadedhorizontal bars at the bottom of the graph represent the approx-imate timing of the spring and summer rice crops and wintervegetable crops
Month
Adu
lt fe
mal
es p
regn
ant (
%)
0
20
40
60
80
100
Litte
r si
ze
0
2
4
6
8
10
12
a
b
Winter cropSpring rice Summer rice
4,4 3,2 7,10 11,16 20,9 31,30 14,16 62,64 6,15 21,28 0,0 1,1
9,11 9,8 16,18 19,23 29,15 41,37 48,58 77,76 20,19 30,45 12,6 4,2
R. argentiventerR. losea
R. argentiventerR. losea
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Fig. 6 Breeding of adult femaleR. argentiventer and R. losea,Vinh Phuc Vietnam, 1999–2002from kill-trapping, showinga average number of embryosper pregnant female ±SE, andb average percentage of adultfemales pregnant as determinedfrom kill samples (±SE). Datapooled from two sites. Samplesizes are shown at the top ofeach figure and represent R.argentiventer and R. losea,respectively. The shadedhorizontal bars at the bottom ofthe graph represent theapproximate timing of thespring and summer rice cropsand winter vegetable crops
252

(F26,561=5.70; P<0.001), but no other interactions weresignificant (P>0.05).
Linear regressions showed that crop stage was a goodpredictor of pregnancy rates of adult R. argentiventer inspring (R2=0.445; F1,15=12.12; P=0.003), but not forsummer (R2=0.095; F1,14=1.48; P=0.244). Similarly,crop stage was a good predictor of pregnancy rates ofadult R. losea in spring (R2=0.516; F1,13=13.87;P=0.003), but not for summer (R2=0.025; F1,14=0.37;P=0.555). This indicates that the rats were respondingstrongly to the availability of resources provided by therice crops during the spring rice crop (increasing fromsowing through to harvesting), but the crop stage couldnot explain changes in the percentage of adult femalespregnant in the summer rice crop. Rainfall, however,was a poor predictor of the percentage of adult femalespregnant. For R. argentiventer, the best fit was for zero-lagged rainfall (R2=0.088; F1,41=3.93; P=0.054), andfor R. losea, the best fit was for 1-month lagged rainfall(R2=0.050; F1,38=2.02; P=0.164).
There was a significant difference in the litter sizeamong months (F10,339=2.90; P=0.002), with averagelitter sizes highest in May (mean=9.7±0.6 SE) andlowest in March (mean=6.3±0.3 SE) (Fig. 6a). Therewere no differences between years (P=0.098) or for themonth by species interaction (P=0.463). The litter sizefor R. argentiventer was higher (mean=8.67±0.20,mode=8, range 2–16, n=182) than for R. losea(mean=7.32±0.15, mode=7, range 3–14, n=168)(F1,338=16.09; P<0.001). The data for placental scarsshowed a similar pattern; the mode for R. argentiventerwas 9 (mean of 8.22±0.21, range 3–14, n=124), and themode for R. losea was 7 (mean of 7.30±0.19, range 2–13, n=123). The overall minimum weight of pregnantR. argentiventer was around 50 g, whereas the minimumweight of pregnant R. losea was around 45 g, andtherefore these weights represent a crude cut-off pointbetween juvenile and adult females for both species.These weights are used above to look at the percentageof juveniles in the population.
Discussion
The rodent populations inhabiting the lowland irrigatedmixed farming system in the Red River Delta ofnorthern Vietnam annually fluctuate in abundance.There was a strong relationship between abundance ofrodents and monthly rainfall, which reflected the highlyseasonal rainfall pattern, the cropping pattern used bythe farmers, and the cold temperatures during winterwhen rice could not be grown. The combination of thesefactors lowered the breeding rate of females over winterand early spring. The rodent populations in the areahave been known to reach high densities and signifi-cantly damage crops, such as the outbreak in 1997/1998(N. P. Tuan, unpublished data). Nevertheless, fromApril 1999 to November 2002, populations were rela-tively low. The peaks in abundance of rodents coincided
with harvesting of the spring and summer rice crops. Thepeak in abundance of rodents following the spring ricecrop was higher than the peak following the summer ricecrop. There were three consecutive months of positiverates of increase during the spring rice crop and only onemonth of positive rates of increase during the summerrice crop. The abundance of rodents was low during thewinter season when little or no breeding occurred.
We were unable to recapture any marked animal sowe have little understanding of whether our trap catch isa good measure of actual abundance. Our trap catch islikely to be influenced by a range of factors includingtrappability, environmental conditions and anthropo-genic activities such as ploughing and harvesting. Fur-thermore, we were unable to conduct more sophisticatedcapture–mark–recapture analyses or to examine changesin density rather than abundance alone. A more reliablemeasure of the dynamics of the rats is from pregnanciesof females from kill-trapping, and we found that therewere changes in the pregnancy rate and litter size of bothR. argentiventer and R. losea over time. There appearedto be strong associations between crop stage and rates ofpregnancy during the spring rice crops for bothR. argentiventer and for R. losea, but not during thesummer crops. Furthermore, rainfall was a poor pre-dictor of pregnancy rates for both species. Based onthese breeding dynamics, we believe that the responsesshown in changes in the abundance of rats over timewould make biological sense, and therefore we areconfident that our observations and findings areappropriate. A radio-tracking study of R. argentiventerin this area showed that most collared animals remainednear their capture location (Brown et al., in press), so itis likely that there is some behavioural response totrapping.
R. argentiventer and R. losea in Vietnam bred duringthe reproductive stages of the two main rice crops, asexpected, but they bred at other times of the year also. Therate of breeding increased through the spring rice crop,presumably as food quality improved, then remainedconstant throughout the summer rice crop. The reason forthis is not clear, but it could be due to the presence of otherhigh-quality food resources available through the summermonths such as grasses found along channel banks, rip-ening vegetable crops, or from grain spilled after harvestof the spring rice crop. In Malaysia and Indonesiabreeding by R. argentiventer is confined to the reproduc-tive stages of the wet- and dry-season rice crops with littlebreeding occurring outside these periods (Lam 1983;Tristiani et al. 1998; Leung et al. 1999). Presumably, bothR. argentiventer and R. losea in Vietnam were obtainingsufficient quality food in their diet from the vegetablecrops and other food sources to initiate or maintainbreeding at timeswhen the ricewas not in the reproductivestage. There is little known about the diet of R. argenti-venter or R. losea in the fields, but they are known to eatcrabs, snails, insects, rice and other plant material (N. P.Tuan and P. R. Brown, unpublished data). Furthermore,given thatR. argentiventer andR. losea have similar levels
253

of breeding each month they may be responding to thesame environmental cues. The breeding dynamics of ratsin the Mekong Delta of southern Vietnam were moreclosely aligned to the growth of the main rice crops, andthere was little breeding occurring outside of these periods(Brown et al. 1999).
There appeared to be a positive, although weak,relationship between the monthly exponential rate ofincrease (rate of change) and the monthly rainfall withno lag. This is probably coincidental because no strongpositive relationships existed with lagged or accumu-lated rainfall. Such relationships exist for a range ofother rodent species, including Mastomys natalensis inTanzania (Leirs et al. 1996), P. hermannsburgensis,N. alexis and M. domesticus in central Australian deserts(Southgate and Masters 1996), Peromyscus leucopus in awoodlot in the USA (Lewellen and Vessey 1998), M.domesticus in wheat fields in Australia (Brown andSingleton 1999), Phyllotis darwini in semi-arid Chile(Lima and Jaksic 1999), and for Mus musculus in beechforests in New Zealand (Choquenot and Ruscoe 2000).In general, the above species respond to accumulatedand lagged rainfall and to subsequently improved foodsupply. The relatively poor response shown here indi-cates that rainfall, although highly seasonal, was not asignificant factor in determining the populationdynamics or breeding of R. argentiventer and R. losea inthis irrigated mixed agroecosystem. Moreover, the rela-tionship between the monthly exponential rate of in-crease (rate of change) and rainfall within the samemonth does not make biological sense. If rodents wereresponding to good conditions, a lag period must occurto allow time for plants or other food sources to growand reach a stage of growth where they are highlynutritious before the rodents can respond and com-mence breeding. A delay would then occur from com-mencement of breeding to birth of the young and thenrecruitment into the trappable population. For R. ar-gentiventer and R. losea, this timing would take 3–6months. Therefore, we believe that the rodents wereresponding more to the availability of high-quality foodprovided by irrigated crops including rice and vegetables(such as kohlrabi, tomato, onion, and pumpkin). Thishypothesis was supported by the regression analysis ofincreasing pregnancy rates for both species during thespring rice crop, and then pregnancy rates remained highduring the summer rice crop.
Overall, there was little effect of rainfall on the pop-ulation dynamics or breeding rates of R. argentiventer orR. losea. Rainfall would have little direct impact onthese rodents except during periods of high rainfall andin low-lying areas, where burrows could be flooded.Flooding did not occur during this study. In the MekongDelta of southern Vietnam, Brown et al. (1999) foundthat flooding reduced population abundance of rats, andpresumably affected survival directly.
Tristiani and Murakami (2003) examined the rate ofincrease of R. argentiventer in response to food supply inan enclosure in West Java, Indonesia. They assumed
that nutritional quality linearly increased from trans-planting to harvest of the rice crop. They concluded thatR. argentiventer was able to adapt to the growth stagesof the rice crop by reproducing when food supply wasoptimal (Tristiani and Murakami 2003), but reproduc-tive data were not provided. Furthermore, they did notlook at the response of the population to lagged foodsupply.
The litter sizes for R. argentiventer of eight to nineobserved in this study were generally lower than littersizes of nine to 12 reported for Malaysia and Indo-nesia (Lam 1980, 1983; Murakami et al. 1990) and inKien Giang Province in the Mekong Delta of southernVietnam, where the average litter size for R. argenti-venter was 10.8 (Brown et al. 1999). It is unclearwhether the lower litter size observed for R. argenti-venter in the Red River Delta of Vietnam was due tocompetition with R. losea and other rodent species orbecause of the availability of vegetable crops in themixed irrigated farming system in Vinh Phuc Provinceof Vietnam. Theory suggests that there should be areduction in fecundity, survivorship or growth throughinterspecific competition (Begon et al. 1996). There arefew published examples to support this theory in ro-dents. However, Eccard and Ylonen (2002) found thatreproduction of bank voles in enclosures was nega-tively affected by the presence of field voles, with fewerbank vole females breeding, although, there was nodifference in the litter size between their treatments.Another possibility is that there are latitudinal effectson litter size. While latitudinal effects on the cyclicdynamics of arvicoline rodents have been well studied(Henttonen et al. 1985; Saitoh et al. 1998; Tkadlec andStenseth 2001), there are few studies on the effects oflatitude and litter size. It is generally considered that inthe northern hemisphere there is a positive relationshipwith latitude and litter size, as found for P. leucopus(Long 1973), but Bilenca et al. (1994) found a negativerelationship in the southern hemisphere for Akodonazare. It seems that photoperiod, temperature andavailability of food influence litter size rather thanlatitude alone (Bilenca et al. 1994; Trillmich 2000). Ourdata on R. argentiventer in Vietnam compared to thoseobserved for R. argentiventer in Indonesia andMalaysia are consistent with reduced fecundity,through lower litter size, because of interspecific com-petition with R. losea, and there is a suggestion thatthere is a negative relationship with latitude and littersize.
The minimum weight of pregnant R. argentiventerand R. losea were determined as 50 and 45 g,respectively. The published figures for adult R. ar-gentiventer are slightly heavier. Murakami et al. (1990)found female R. argentiventer with open vagina from28 g body weight (estimated at only 28 days old), butpregnant females from 60 g body weight (estimated as40 days old). Lam (1983) showed that in double-cropped areas, females showed signs of sexual matu-rity with perforated vagina, placental scars and much
254

enlarged mammae starting at the 60 g mid-pointweight class (the size of the weight classes was notstated).
Because we found pregnant female R. argentiventerand R. losea above 50 and 45 g respectively, we classifiedanimals that were lighter in weight as juveniles. This wasa crude method for determining the presence of juvenilesin the population. A general observation was thatbreeding was occurring in nearly all months, and as aconsequence, juveniles were observed in nearly allmonths except for the early part of the spring rice crop.
The recapture rate of marked animals was zero for allspecies. It is well known that the recapture rates forR. argentiventer are very low (Brown et al. 1999; Leunget al. 1999; Jacob et al. 2003). Because there were norecaptures of marked animals, it was not possible todetermine the growth rates of individuals or to calculatesurvival. An apparent survival rate could be calculatedbased on the shift of body weights from one month tothe next at a population level; however, little is knownabout the growth rates of R. argentiventer, or whetherthis would be an appropriate method for estimatingsurvival. The age of R. argentiventer can be estimatedfrom eye lens weight (Vejaratpimol and Liat 1983), butthe relationship between eye lens weight and bodyweight was not provided. This requires further investi-gation.
Conclusion
R. argentiventer and R. losea show similar dynamics inthis complex farming system in the Red River Delta,Vietnam. There was a strong relationship between themonthly abundance of rats and rainfall, but a weakrelationship between monthly rates of increase andrainfall. While breeding was occurring during thereproductive stage of the rice crop as expected, somebreeding was occurring outside of these periods, pre-sumably because of the availability of high-qualityfood provided by the vegetable crops. Given thesesimilar dynamics, it is likely that these two speciescompete for resources. There are many interestingecological questions relating to the competition be-tween R. argentiventer and R. losea which can beinvestigated, such as examining diet preferences andusing radio-telemetry to study the habitat use of boththese species. The findings from this study will help torefine the management of the rodent community byfocusing management prior to the onset of the mainbreeding periods.
Acknowledgements We thank all the village heads and farmers in-volved in the project for their support and willingness to participatein the project. Staff of the National Institute for Plant Protection,Vietnam, provided excellent support while in the field and gettingthe project underway. In particular we thank Le Than Hoa, PhamThi Lien, and Pham Van Kien. We thank Drs J. Jacobs and K.Williams for critical comments on the manuscript. This research ispart of the Australian Centre for International Agricultural
Research funded project Management of Rodents in Rice-BasedFarming Systems of Southeast Asia (AS1 98/36) and was con-ducted in accordance with the Australian Code of Practice for theCare and Use of Animals for Scientific Purposes. The approval nos.were WEAEC 98/99-09 and SEAEC 01/02-09.
References
Aplin KP, Brown PR, Jacob J, Krebs CJ, Singleton GR (2003a)Field methods for rodent studies in Asia and the Indo-Pacific.ACIAR monograph 100. Australian Centre for InternationalAgricultural Research, Canberra
Aplin KP, Chesser T, ten Have J (2003b) Evolutionary biology ofthe genus Rattus: profile of an archetypal rodent pest. In: Sin-gleton GR, Hinds LA, Krebs CJ, Spratt D (eds) Rats, mice andpeople: rodent biology and management. ACIAR monograph96. Australian Centre for International Agricultural Research,Canberra, pp 487–498
Begon M, Harper JL, Townsend CR (1996) Ecology—individuals,populations, and communities. Blackwell, Boston
Bilenca DN, Zuleta GA, Kravetz FO (1994) Latitudinal variationin litter size and length of the breeding season in populations ofpampas mice, Akodon azare. Mammalia 58:563–568
Brown PR, Singleton GR (1999) Rate of increase as a function ofrainfall for house mouseMus domesticus populations in a cereal-growing region in southern Australia. J Appl Ecol 36:484–493
Brown PR, Hung NQ, Hung NM, van Wensveen M (1999) Pop-ulation ecology and management of rodent pests in the MekongRiver Delta, Vietnam. In: Singleton GR, Hinds LA, Leirs H,Zhang Z (eds) Ecologically based management of rodent pests.Australian Centre for International Agricultural Research,Canberra, pp 319–337
Brown PR, Tuan NP, Banks PB (in press) Movements, habitat useand response of ricefield rats to removal in an intensive crop-ping system in Vietnam. Belg J Zool
Choquenot D, Ruscoe WA (2000) Mouse population eruptions inNew Zealand forests: the role of population density and seed-fall. J Appl Ecol 69:1058–1070
Eccard JA, Ylonen H (2002) Direct interference or indirectexploitation? An experimental study of fitness costs of inter-specific competition in voles. Oikos 99:580–590
Henttonen H, McGuire AD, Hansson L (1985) Comparisons ofamplitudes and frequencies (spectral analysis) of density vari-ations in long-term data sets of Clethrionomys species. AnnZool Fenn 22:221–227
Jacob J, Sudarmaji, Singleton GR (2003) Ecologically basedmanagement of rice-field rats on a village scale in West Java:experimental approach and assessment of habitat use. In: Sin-gleton GR, Hinds LA, Krebs CJ, Spratt D (eds) Rats, mice andpeople: rodent biology and management. ACIAR monograph96. Australian Centre for International Agricultural Research,Canberra, pp 191–196
Lam YM (1980) Reproductive behaviour of the rice field rat,Rattus argentiventer and implications for its control. Proceed-ings of the National Rice Conference, Kuala Lumpur, Malay-sia, pp 243–257
Lam YM (1982) Rats as rice field pests—their importance and con-trol. In: Khoo KC (ed) Rodent pests of agricultural crops inMalaysia. Plant Protection Society,KualaLumpur,Malaysia, pp9–17
Lam YM (1983) Reproduction in the ricefield rat Rattus argenti-venter. Malay Nat J 36:249–282
Lan LP, Aplin KP, Hung NM, Quoc NV, Chien HV, Sang ND,Singleton GR (2003) Rodent communities and historical trendsin rodent damage in the Mekong Delta of Vietnam: establishingan ecological basis for effective pest management. In: SingletonGR, Hinds LA, Krebs CJ, Spratt D (eds) Rats, mice and peo-ple: rodent biology and management. ACIAR monograph 96.Australian Centre for International Agricultural Research,Canberra, pp 290–296
255

Leirs H, Verhagen R, Verheyen W, Mwanjabe P, Mbise T (1996)Forecasting rodent outbreaks in Africa—an ecological basis forMastomys control in Tanzania. J Appl Ecol 33:937–943
Leung LKP, Singleton GR, Sudarmaji Rahmini (1999) Ecologicallybased population management of the rice-field rat in Indonesia.In: SingletonGR,Hinds LA, LeirsH, ZhangZ (eds) Ecologicallybased management of rodent pests. Australian Centre for Inter-national Agricultural Research, Canberra, pp 305–318
Lewellen RH, Vessey SH (1998) The effect of density dependenceand weather on population size of a polyvoltine species. EcolMonogr 6:571–594
Lima M, Jaksic FM (1999) Population rate of change in the leaf-eared mouse: the role of density-dependence, seasonality andrainfall. Aust J Ecol 24:110–116
Long CA (1973) Reproduction, growth, and development in high-latitude and low-latitude populations of Peromyscus leucopus(Rodentia). Southwest Nat 18:11–20
Marshall JT (1977) Family Muridae: rats and mice. In: Lekagul B,McNeely JA (eds) Mammals of Thailand. Association ofConservation of Wildlife, Bangkok, pp 397–487
Murakami O, Priyono J, Tristiani H (1990) Population manage-ment of the ricefield rat in Indonesia. In: Quick GR (eds) Ro-dents and rice. International Rice Research Institute, LosBanos, Philippines, pp 49–54
Nitatpattana N, Henrich T, Palabodeewat S, Tangkanakul W,Poonsuksombat D, Chauvancy G, Barbazan P, Yoksan S,Gonzalez JP (2002) Hantaan virus antibody prevelance inpopulations of several provinces of northeastern Thailand.Trop Med Int Health 7:840–845
Saitoh T, Stenseth NC, Bjørnstad ON (1998) The populationdynamics of the vole Clethrionomys rufocanus in Hokkaido,Japan. Res Popul Ecol 40:61–76
Shih CM, Wang JC, Chao LL, Wu TN (1998) Lyme disease inTaiwan: first human patient with characteristic erythemachronicum migrans skin lesion. J Clin Microbiol 36:807–808
Singleton GR (1997) Integrated management of rodents: a south-east Asian and Australian perspective. Belg J Zool 127:157–169
Singleton GR (2003) Impacts of rodents on rice production in Asia.(IRRI discussion paper series no. 43) International Rice Re-search Institute, Los Banos, Philippines
Singleton GR, Brown PR (1999) Management of mouse plagues inAustralia: integration of population ecology, bio-control andbest farm practice. In: Cowan DP, Feare CJ (eds) Advances invertebrate pest management. Filander, Berlin, pp 189–203
Singleton GR, Petch DA (1994) A review of the biology andmanagement of rodent pests in Southeast Asia. ACIAR tech-nical report no. 30. Australian Centre for International Agri-cultural Research, Canberra
Southgate R, Masters P (1996) Fluctuations of rodent populationsin response to rainfall and fire in a Central Australian hum-mock grassland dominated by Plectrachne schinzii. Wildl Res23:289–303
Suntsov VV, Huong LTV, Suntsova NI, Gratz NG (1997) Plaguefoci in Vietnam: zoological and parasitological aspects. BullWHO 75:117–123
Tkadlec E, Stenseth NC (2001) A new geographic gradient invole population dynamics. Proc R Soc Lond B Biol Sci268:1547–1552
Trillmich F (2000) Effects of low temperature and photoperiod onreproduction in the female wild guinea pig (Cavia aperea).J Mammal 81:586–594
Tristiani H, Murakami O (2003) Rates of population increase inthe ricefield rat (Rattus argentiventer) as a function of foodsupply: an enclosure study in Jatisari, West Java. J Zool259:239–244
Tristiani H, Priyono J, Murakami O (1998) Seasonal changes in thepopulation density and reproduction of the ricefield rat, Rattusargentiventer (Rodentia: Muridae), in West Java. Mammalia62:227–239
Tuan NP, Williams SJ, Brown PR, Singleton GR, Tan TQ, HueDT, Ha PTT, Hoa PT (2003) Farmers’ perceptions and prac-tices in rat management in Vinh Phuc Province, northernVietnam. In: Singleton GR, Hinds LA, Krebs CJ, Spratt D(eds) Rats, mice and people: rodent biology and management.ACIAR monograph 96. Australian Centre for InternationalAgricultural Research, Canberra, pp 399–402
Vejaratpimol R, Liat LB (1983) Age estimation of Rattus argenti-venter from eye lens weight. J Sci Soc Thai 9:107–118
256




















