
Population differences in the expression of nucleolus organizer regions in the grasshopper...
6
Protoplasma (2001) 217: 185-190 PROTOPLASMA @ Springer-Verlag 2001 Printed in Austria Population differences in the expression of nucleolus organizer regions in the grasshopper Eyprepocnemis plorans . M. Bakkali*, J. Cabrero, M. D. López-León, F. Perfectti, and J. P. M. Camacho ~ Departamento de Genética, Facultad de Ciencias, Universidad de Granada, Granada Received July 31, 2000 Accepted February 9, 2001 Summary. Fluorescence in situ hybridization revealed the presence of ribosomal RNA genes in paracentromeric regions of al! A chro- mosomes and in the distal half of B chromosomes in embryonic cel!s from Moroccan specimens of the grasshopper Eyprepocnemis plorans. The expression of these genes was monitored by the pres- ence of nucleoli attached to each chromosome bivalent in diplotene cel!s from males col!ected from two different Moroccan populations and was compared to previous datagiSpanish populations. Whereas only the nucleolus organizer regiéms (NORs) on S9-Sll and X chro- mosomes were active in the Spanish specimens, Moroccan individ- uals showed NOR activity in al! chromosomes. The rRNA gene s on the B chromosome were inactive in both populations. The Sgand SlO NORs were less active in Moroccan specimens than in Spanish specimen, which might be partly explained by the negative inter- dependence for expression of the SlONOR with respect to those on ~ and X chromosomes. On the other hand, the X NOR was more active in Moroccan specimens than in Spanish specimens, and this might be partly due to the positive effect that the presence of B chromosomes has on the expression of this NOR. The implications of these observations on current models of NOR activity regulation are discussed. Keywords: rDNA; Nucleolus organizer regían; B chromosome; Grasshopper; Eyprepocnemis plorans. Abbreviation: NOR nucleolus organizer regían. Introduction In species with nucleolus organizer regions (NORs) in several chromosomes a regulatory mechanism is ex- pected to exist in arder that each cell gets an appro- priate amount of rRNA. The elucidation of such a mechanism is still far from being completely under- stood, but recent research has provided some inter- esting insights. Schubert and Künzel (1990) have * Correspondence and reprints: Departamento de Genética, Facul- tad de Ciencias, Universidad de Granada, 18071 Granada, Spain. suggested that the number of rDNA cistrons is corre- lated to the expression level of a NOR. Forty years earlier, the size of the nucleolus was related to the amount of rRNA synthesized (Caspersson 1950). RecentIy, Zurita et al. (1998) have suggested that, because of their high competitiveness fm transcrip- tional factors, NORs with the greatest amount of rDNA are preferentially activated. Interindividual differences in the amount of rDNA have be en reported in Bufo marinus (Miller and Brown 1969), Homo sapiens (Evans et al. 1974), Ambystoma mexicanus (Sinclair et al. 1974), Salmo irideus (Schmidtke 1976), and Rattus norvegicus (Sasaki et al. 1986), and the variation observed by fiu- orescence in situ hybridization (FISH) reveals differ- ences in the number of cistrons (WachtIer et al. 1986, Suzuki et al. 1990, King et al. 1990, Leitch and Heslop- Harrison 1992). López-León et al. (1994) reported the presence of rDNA in all standard chromosomes from Spanish specimens of the grasshopper Eyprepocnemis plorans and distinguished between large (located on the chro- mosomes Sg,SlO,Su, and X) and small (located on the chromosomes Ll to Ms) rDNA clusters. Fm simplicity, we will llame primary NORs the large rDNA clusters and secondary NORs the small ones. With the excep- tion of a single male found in Salobreña (Granada, Spain) by 1. S. Rufas (Universidad Autónoma de Madrid, Spain, pers. commun.), only primary NORs are active in Spanish specimens of E. plorans (Cabrero et al. 1987; López-León et al. 1991,1995). In addition to the standard (A) chromosomes, almost all populations of E. plorans harbor a poly-
Transcript of Population differences in the expression of nucleolus organizer regions in the grasshopper...

Protoplasma (2001) 217: 185-190 PROTOPLASMA@ Springer-Verlag 2001Printed in Austria
Population differences in the expression of nucleolus organizer regions inthe grasshopper Eyprepocnemis plorans.M. Bakkali*, J. Cabrero, M. D. López-León, F. Perfectti, and J. P. M. Camacho
~Departamento de Genética, Facultad de Ciencias, Universidad de Granada, Granada
Received July 31, 2000
Accepted February 9, 2001
Summary. Fluorescence in situ hybridization revealed the presenceof ribosomal RNA genes in paracentromeric regions of al! A chro-mosomes and in the distal half of B chromosomes in embryoniccel!s from Moroccan specimens of the grasshopper Eyprepocnemisplorans. The expression of these genes was monitored by the pres-ence of nucleoli attached to each chromosome bivalent in diplotenecel!s from males col!ected from two different Moroccan populationsand was compared to previous datagiSpanish populations. Whereasonly the nucleolus organizer regiéms (NORs) on S9-Sll and X chro-mosomes were active in the Spanish specimens, Moroccan individ-uals showed NOR activity in al! chromosomes. The rRNA gene s onthe B chromosome were inactive in both populations. The Sgand SlONORs were less active in Moroccan specimens than in Spanishspecimen, which might be partly explained by the negative inter-dependence for expression of the SlONOR with respect to those on~ and X chromosomes. On the other hand, the X NOR was moreactive in Moroccan specimens than in Spanish specimens, and thismight be partly due to the positive effect that the presence of Bchromosomes has on the expression of this NOR. The implicationsof these observations on current models of NOR activity regulationare discussed.
Keywords: rDNA; Nucleolus organizer regían; B chromosome;Grasshopper; Eyprepocnemis plorans.
Abbreviation: NOR nucleolus organizer regían.
Introduction
In species with nucleolus organizer regions (NORs) inseveral chromosomes a regulatory mechanism is ex-pected to exist in arder that each cell gets an appro-priate amount of rRNA. The elucidation of such amechanism is still far from being completely under-stood, but recent research has provided some inter-esting insights. Schubert and Künzel (1990) have
* Correspondence and reprints: Departamento de Genética, Facul-tad de Ciencias, Universidad de Granada, 18071 Granada, Spain.
suggested that the number of rDNA cistrons is corre-lated to the expression level of a NOR. Forty yearsearlier, the size of the nucleolus was related to theamount of rRNA synthesized (Caspersson 1950).RecentIy, Zurita et al. (1998) have suggested that,because of their high competitiveness fm transcrip-tional factors, NORs with the greatest amount ofrDNA are preferentially activated.
Interindividual differences in the amount of rDNA
have be en reported in Bufo marinus (Miller andBrown 1969), Homo sapiens (Evans et al. 1974),Ambystoma mexicanus (Sinclair et al. 1974), Salmoirideus (Schmidtke 1976), and Rattus norvegicus(Sasaki et al. 1986), and the variation observed by fiu-orescence in situ hybridization (FISH) reveals differ-ences in the number of cistrons (WachtIer et al. 1986,Suzuki et al. 1990, King et al. 1990, Leitch and Heslop-Harrison 1992).
López-León et al. (1994) reported the presence ofrDNA in all standard chromosomes from Spanishspecimens of the grasshopper Eyprepocnemis ploransand distinguished between large (located on the chro-mosomes Sg,SlO,Su, and X) and small (located on thechromosomes Ll to Ms) rDNA clusters. Fm simplicity,we will llame primary NORs the large rDNA clustersand secondary NORs the small ones. With the excep-tion of a single male found in Salobreña (Granada,Spain) by 1. S. Rufas (Universidad Autónoma deMadrid, Spain, pers. commun.), only primary NORsare active in Spanish specimens of E. plorans (Cabreroet al. 1987; López-León et al. 1991,1995).
In addition to the standard (A) chromosomes,almost all populations of E. plorans harbor a poly-

186
morphism for supernumerary (B) chromosomes (for areview, see Camacho et al. 2000). These B chromo-somes contain the greatest amount of rDNA in thegenome (López-León et al. 1994, Cabrero et al. 1999),but with the exception of a single male carrying acentric fusion between the B and the longest autosome(Cabrero et al. 1987), the rDNA contained in the Bchromosome has never been found to be active.
In this paper we analyze the localization and expres-sion of the NORs in Moroccan specimens of E.plorans. The frequency of NOR expression was ana-lyzed and compared with data reported by López-León et al. (1995) for Spanish specimens. Finally, theinterdependence for expression between the differentNORs, including the possible effects of B chromosomewas also analyzed.
This work was orally presented at the VIII Congresode la Sociedad Española de Biología Celular held inGranada (December 17-21, 1999).
Material and methods
Adult males and females of the grasshopper Eyprepocnemis ploranswere sampled from the Moroccan populations of SO. DE. A. andFrain (for geographicallocations see the map in Bakkali et al. 1999).Testes were immediately fixed in 3 ; 1 ethanol-acetic acid, in arderto analyze NOR express ion by means of the silver nitrate impreg-nation technique (Rufas et al. 1982). A total of 11 males from SO.DE. A. and 9 from Frain were used for this purpose, from which 128first spermatocytes at diplotene were analyzed, a meiotic stagewhere previous activity of each NOR may be evaluated by the pres-ence of a nucleolus attached to it.
Females were kept alive in the labmatmy by placing them in acage with grass and humid vermiculite to facilitate egg laying. Eggpods were incubated at 28 DCfor ten days and each egg was dis-sected in insect saline solution. The embryos were then fixed fmcytological analysis as described in Camacho et al. (1991). Chromo-Borne localization of rDNA was perfmmed on embryo cells by theFISH technique described in López-León et al. (1994) and Cabreroet al. (1999).
To compare the expression frequency of the different NORsbetween Moroccan and Spanish specimens, we used data from Jete(Granada, Spain) repmted by López-León et al. (1995). The fre-quency of activity per cell fm each NOR was compared betweenspecimens of the two geographical miles by means of the Mann-Whitney test. To test the interdependence fm activity between thedifferent NORs in the same chromosome complement, a series oflogistic regressiQn tests were done with each NOR as the dependentvariable and the remaining NORs as independent ones. This analy-Bisis very appropriate for binary variables such as NOR expression,where two states where recorded for each one, i.e., active or inac-tive. In addition, we calculated the odds ratio, a useful index fm theinterpretation Qf the results of logistic regression. It is computedfrom a 2 x 2 dassification table, which displays the predicted andobserved cla~sification of cases for a binary dependent variable.An odds ratio lower than 1, with 95% confidence limits not includ-ing the value 1, indicates negative associatio n between the twovariables involved. If the odds ratio is higher than 1 and the 95%
M. Bakkali et al.: Population differences in NOR expression
confidence limits exclude the value 1, the association is positive.Finally, the Mann~Whitney test was used to investigate possibleeffects of B chromosomes on the expression of the NORs on the Achromosomes. When multiple univariant tests were performed, weapplied the sequential Bonferroni test to avoid type I errms (Rice1989).
ResuIts
The karyotype of E. plorans consists of 22 plus XO(d' d') or xx «( «) telocentricchromosomeswhichare conventionally classified into three size groups:three long (L¡-L3), five m'edium (McMs), and threeshort (S9-S11)chromosomes; the size of the X chro-mosome being intermediate between those of L2 andL3 (John and Lewis 1965). In addition, specimens ofthis species can carry B chromosomes, which are foundin almost all natural populations (Camacho et al.2000).
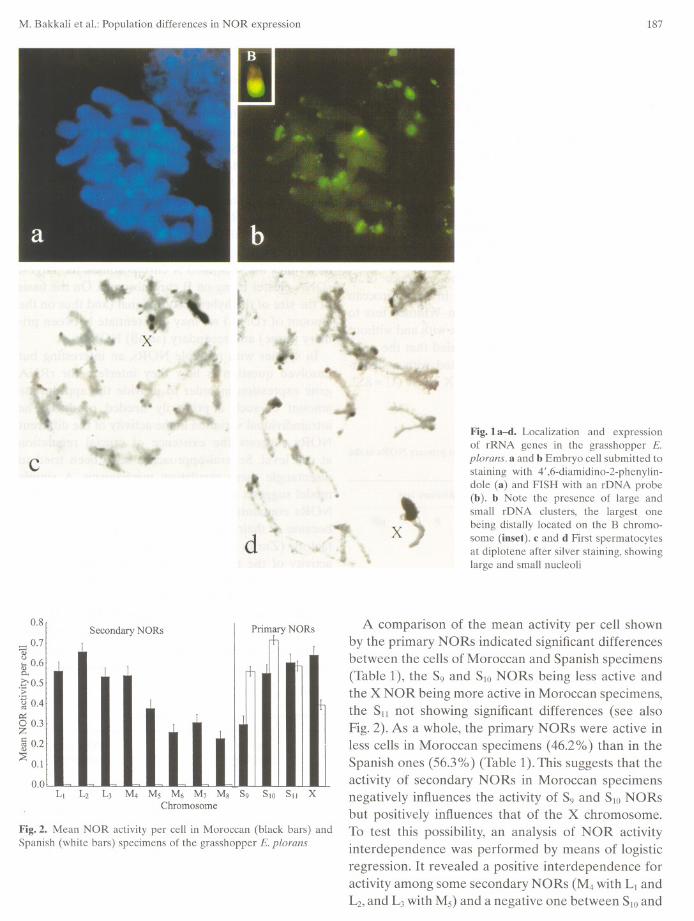
In Moroccan specimens, the FISH techniqueshowed the presence of rDNA in the paracentromericregion of all A chromosomes and the distal half of Bchromosomes (Fig. 1). The largest cluster of rDNAwas found on the B chromosome, followed by those onthe X, S9,S1O,and S11chromosomes, i.e., those A chro-mosomes harboring the primary NORs. The remain-ing autosomes (L¡ to Ms) showed minute amounts ofrDNA (see Fig. 1b). Similar results have been previ-ously reported in specimens from Spanish populationsof this grasshopper (López-León et al. 1994).
A score of 128 diplotene cells from males collectedat both Moroccan populations showed nonsignificantdifferences in the activity per cell of any of the NORsbetween So. DE. A (61 cells) and Frain (67 cells)(results not shown). This permits using the whole col-lection of cells (128) as a single sample for compari-sons with previous data obtained in Jete (Spain) byLópez-León et al. (1995). On average, 5.54 NORs outof the 12 existing on the A genome were active inMoroccan specimens (Fig. 1c, d), which was signifi-cantly higher than the 2.25 observed in Spanishspecimens (Mann-Whitney: U = 4443, P < 0.001). Thishigher number of active NORs per cell was due to theactivity on the L¡-Ms chromosomes, which never showNOR activity in Spanish populations (Cabrero et al.1987, López-León et al. 1995) (Fig.2). The rRNAgenes located on B chromosomes in Morrocan speci-mens were always inactive.
Whereas primary NORs usually yielded large nucle-oli, secondary NORs in Moroccan specimens alwaysyielded small nucleoli (see Fig. 1c, d).
M. Bakkali et al.: Population differences in NOR expression
, '"..,., iIt~.
r..... .
'i¡ t'" .
~
....
e1
d...
0.8
- 0.7v~0.6O)c..~0.5.:;:
.~ 0.4~00.3Z§ 0.2O)
::;;; 0.1
SecondaryNORs Primary NORs
r-
+
0.0L, L3 SIO Sil XM4 Ms M6 M¡ Ms S9
ChromosomeL2
Fig.2. Mean NOR activity per cel! in Moroccan (black bars) andSpanish (white bars) specimens of the grasshopper E. plorans
187
..
..
"
Fig. la-d. Localization and expressionof rRNA genes in the grasshopper E.plorans. a and b Embryo cel! submitted tostaining with 4',6-diamidino-2-phenylin-dote (a) and FISH with an rDNA probe(b). b Note the presence of large andsmal! rDNA clusters, the largest onebeing distal!y located on the B chromo-some (iuset). e and d First spermatocytesat diplotene after silver staining, showinglarge and smal! nucleoli
.. ,X)
..'"
A comparison of the IDean activity per cell shownby the primary NORs indicated significant differencesbetween the cells of Moroccan and Spanish specimens(Table 1), the S9 and SlONORs being less active andthe X NOR being more active in Moroccan specimens,the Su not showing significant differences (see alsoFig. 2). As a whole, the primary NORs were active inless cells in Moroccan specimens (46.2%) than in theSpanish ones (56.3%) (Table 1). This suggests that theactivity of secondary NORs in Moroccan specimensnegatively influences the activity of S9 and SlONORsbut positively influences that of the X chromosome.To test this possibility, an analysis of NOR activityinterdependence was performed by means of logisticregression. It revealed a positive interdependence foractivity among some secondary NORs (M4with Ll andLz, and L3with Ms) and a negative one between SlOand

188
L:;and between SlOand X activities (Table 2).A similaranalysis of the Spanish data revealed a significantIynegative interdependence between the X NOR andthe three remaining primary NORs (Table 2). It istherefore reasonable to conclude that the activity ofsecondary NORs (e.g., L:;) negatively influences theactivity of some primary NORs (e.g., SlO)but is notresponsible for the higher activity of the X NOR in thecells of the Moroccan specimens.
Logistic regression applied to the Spanish data fromJete (López-León et al. 1995) showed a negative inter-dependence between the X chromosome and the threeremaining primary NORs (Table 2).
To detect possible effects of the presence of Bchromosomes on NOR expression in the Moroccanspecimens, we performed the Mann- Whitney test tocompare NOR activity between cells with and withoutB chromosomes. This analysis revealed that the pres-ence of B chromosomes is associated with a signifi-cant increase in the activity of the X NOR (U = 852,P = 0.017).
Table 1. Mean activity per cel! oí the diííerent primary NORs in the
genome oí E. plorans males
M. Bakkali et al.: Population differences in NOR expression
Discussion
The FISH technique with an rDNA probe provides thechromosome localization of both active and inactive
rRNA genes. It has also been suggested that the sizeof the hybridization signal may indicate the amount ofrDNA present (Suzuki et al. 1990, Leitch and Heslop-Harrison 1992, Mellink et al. 1994). Our present resultshave shown that Moroccan specimens of the grasshop-per E. plorans show a chromosomal localization anddistribution of rRNA genes similar to that reported byLópez-León et al. (1994) for Spanish specimens, withsmall rDNA clusters on Ll to Ms chromosomes andlarge ones on S9-S11and X chromosomes, the largestrDNA cluster being on B chro¡:nosomes. On the basisof the size of the hybridization signal (and thus on theamount of rDNA) we may differentiate between pri-mary (large) and secondary (small) NORs.
In species with multiple NORs, an interesting butunsolved question is how they interfere for rRNAgene expression in arder to provide the appropriateamount of such a primarily needed product. Theintraindividual variation in the activity of the differentNORs suggests the existence of crucial regulationat cell level. Several approaches have been tried todisentangle these regulatory mechanisms. A simplemodel suggests a preference for expression for thoseNORs containing a higher number of rRNA gene sbecause of their higher ability to recruit transcriptionfactors (Zurita et al. 1998). Whereas the exclusiveactivity of the primary NORs in Spanish specimens(Cabrero et al. 1987; López-León et al. 1991, 1995) isin agreement with this hypothesis, the high frequencyof activity shown by the secondary NORs in Moroc-can specimens, comparable to that of the primaryNORs (see Fig. 2), seems to be an exception. The inac-tivity of rDNA cistrons in some B chromosomes har-
Chromo- Mean NOR activity Mann-Whitney testsome per cel! oíNOR population: U P pE'
Morocco Spain(n = 128) (n=417)
S9 0.297 0.559 19699.0 7.3 x 10-6 <0.001
S1O 0.547 0.715 22211.0 4.1 x 10-3 0.0082Sl1 0.602 0.585 26249.5 7.8 x 10-1 0.7784X 0.641 0.393 20087.0 2.3 x 10-5 <0.001Mean 0.462 0.563 19581.0 5.1 x 10-6 <0.001
" pB, probability corrected by the sequential Boníerroni method
Table 2. Interdependences oí the activities oí the diííerent NORs in the chromosome complement oí E. plorans
Population NORs with Type oí t degrees oí P Odds ratio (95%significant association íreedom confidence limits)interdependence
Morocco L¡-M4 + 2.64 116 0.009 3.15 (1.33-7.47)LrM4 + 3.30 116 0.001 4.77 (1.87-12.18)LrMs + 3.12 116 0.002 4.36 (1.71-11.12)LrS1O -2.51 116 0.014 0.31 (0.13-0.78)X-S1O -2.12 116 0.036 0.40 (0.17-0.94)
Spain X-S9 -3.49 413 <0.001 0.48 (0.31-0.72)X-S1O -4.46 413 <0.001 0.36 (0.23-0.56)X-Sl1 -2.56 413 0.011 0.58 (0.38-0.88)

M. Bakkali et al.: Population differences in NOR expression
boring the largest rDNA cluster in the genome, e.g.,B chromosomes in E. plorans (Cabrero et al. 1987,López-León et al. 1995, ibis paper) and Rattus rattus(Stitou et al. 2000), is another exception, although inibis case the B NOR inactivity might be a consequenceof their general heterochromatinization and geneticinactivation caused by host geno me mechanisms fordefence against intragenomic parasites (Yoder et al.1997). In such a case, the B NOR might be completelyunavailable to regulatory mechanisms for A chromo-somes. On the other hand, the activation of secondaryNORs (smaIl rDNA clusters) in Moroccan E. ploransceIls where some of the primary NORs (large rDNAclusters) were inactive suggests that more complexmodels are necessary to explain ibis case.
Although secondary NORs in Moroccan specimenswere comparatively as active as primary ones, thesize of the nucleoli yielded was rather different, theprimary NORs yielding large nucleoli and the sec-ondary NORs very smaIl ones. This is consistent withthe existence of a relationship between the number ofrRNA genes in a NOR and the amount of rRNAsynthesized and, consequentIy, the nucleolus size (DeCapoa et al. 1988, Shubert and Künze11990, Zatsepinaet al. 1996).
The activation of the secondary NORs in Moroccanspecimens might be due to some genetical and/or envi-ronmental differences determining regulatory changesin these individuals with respect to those from Spain.Since NORs are usually highly polymorphic (Hender-son et al. 1976, Schmidtke 1976, GaIleti et al. 1985,Sasaki et al. 1986, Sánchez et al. 1989), the possibilityremains that primary NORs in Moroccan specimenscould harbor less rDNA iban Spanish ones, so that theactivity of secondary ones is needed. QuantitativeFISH of primary NORs in both Moroccan and Spanishspecimens is needed to test ibis hypothesis. In addi-tion, a detailed comparison of the nucleolus sizebetween Moroccan and Spanish ceIls would help toascertain whether rRNA requirements differ betweenspecimens from the two geographical origins.
The analysis of NOR expression interdependenceindicates a negative association between the S9-SIlNORs and the X Que,in Spanish specimens. In Moroc-can specimens, the activity of the secondary NORs wasparaIleled by a decrease in the activity of the S9 andSlONORs and an increase in the activity of the XNOR. But only the SlOdecrease was directly depen-dent on the expression of the secondary NORs sinceit was negatively associated to the activity of the NOR
189
on the Lz chromosome. It is also remarkable thatthe only negative interdependence between primaryNORs in Moroccan specimens was found between theSlOand the X, those involving the S9 and Sil NORs inSpanish specimens not appearing in Morocco. Since SlOharbors the only primary NOR in Moroccan speci-mens which is not accompanied by the 180 pb DNAtandem repeat reported by López-León et al. (1994)(1 Cabrero et al. unpubl.), the possibility that ibisrepeat might playa role in NOR activity regulation,as was suggested by López-León et al. (1995), is notunlikely. The repeat could act in some unknown wayprotecting the adjacent NOR from inactivation orinterchromosomal dependence. The SlONOR, by beingdevoid of ibis repeat, would be prone to suffer depen-dencies, whereas the remaining NORs would be moreor less blind to some interchromosomal effects,perhaps depending on their amount of DNA repeat.
The NOR on the X chromosome was significantIymore active in Moroccan specimens iban in Spanishones, and ibis was not due to the expression of the sec-ondary NORs since no activity interdependence wasobserved between the X and the Ll-Ms chromosomes(see Table 2). More likely is the possibility that ibishigher activity is due to the presence of B chromo-somes since the X was significantly more active in ceIlsharboring B chromosomes. However, ibis cannot bethe whole explanation since the X NOR was signifi-cantIy more active in OB cells from Morocco iban inOB ceIls from Spain (U = 8678,P = 0.0002). Increasesin the activity of the X NOR associated to the pres-ence of B chromosomes were reported by Cabreroet al. (1987) from a Spanish population of E. ploranscoIlected at Salobreña. In the Jete population analyzedby López-León et al. (1995), the X NOR also tendedto be more active in the presence of B chromosomes,but the difference did not reach significance (U =19543, P = 0.079). Changes in NOR activity patternsassociated to the presence of B chromosomes havealso been reported in the migratory locust (Salcedoet al. 1988). Whether the higher frequency of X-NORexpression in B-carrying cells has a relationship withthe presumed origin of the B chromosome from the Xchromosome (López-León et al. 1994) is an interest-ing subject for future studies.
Acknowledgments
M.E. was recipient of fellowships fram the Agencia Española
de Cooperación Internacional (Spain)/ and the Ministere de

190
l'Enseignement Supérieur, de la Formation des Cadres et de laRecherche Scientifique (Morocco). We thank Mr. SoluaimaneBakkali for his help in collecting specimens. This study was sup-ported by grants from the Spanish Dirección General de EnseñanzaSuperior (m. PB96-1433) and Plan Andaluz de Investigación, Grupono. CVI-165.
References
Bakkali M, Cabrero 1, López-León MD, Perfectti F, Camacho lPM(1999) The B chromosome polymorphism of the grasshopperEyprepocnemis plorans in north Africa I: B variants and fre-quency. Heredity 83: 428-434
Cabrero 1, Alché lD, Camacho lPM (1987) Effects of B chromo-some on the activity of nucleolus organizer regions in thegrasshopper Eyprepocnemis plorans: activation of a latent nucle-olus organizer region on a B chromosome fused to an autosome.Genome 29: 116-121
- López-León MD, Bakkali M, Camacho lPM (1999) Commonorigin of B chromosome variants in the grasshopper Eyprepoc-nemis plorans. Heredity 83: 435-439
Camacho lPM, Cabrero 1, Viseras E, López-León MD, Navas-Castillo 1, Alché lD (1991) G banding in two species of gras-shopper and its relationship to C, N, and fiuorescence bandingtechniques. Genome 34: 638-643
- Sharbel TF, Beukeboom LW (2000) B chromosome evolution.Philos Trans R Soc Lond Ser B 355: 163-178
Caspersson T (1950) Cell growth and cell function: a cytochemicalstudy. WW Norton and Co, New York
De CarDa A, Felli MP, Baldini A, Rocchi M, Archidiacono N,Aleixander C, Miller 01 (1988) Relationship between the numberand function of ribosomal genes. Human Genet 79: 301-304
Evans Hl, Buckland RA, Pardue ML (1974) Location of the genescoding for 18S and 28S ribosomal RNA in the human genome.Chromosoma 48: 405-426
Galleti PM lr, Barbosa da Silva E, Germinaro Rl (1985) A multi-pie NOR system in the fish Serrasalmus spilopleura (Serrasalmi-nae, Characidae). Rev Brasil Genet 8: 479-484
Henderson AS, Eicher EM, Yu MT, Atwood KC (1976) Variation inribosomal RNA gene number in mouse chromosomes. CytogenetCell Genet 17: 307-316
lohn B, Lewis KR (1965) Genetic speciation in the grasshopperEyprepocnemis plorans. Chromosoma 16: 548-578
King M, Coníferas N, Honeycutt RL (1990) Variation within andbetween nucleolus organizer regions in Australian hylid frogs(Anura) shown by 18S + 28S in situ hybridization. Genetica 80:17-29
Leitch IJ, Heslop-Harrison lS (1992) Physical mapping of the 18S -5.8S - 26S rRNA genes in barley by in situ hybridisation. Genome35: 1013-1018
López-León MD, Cabrero 1,Camacho lPM (1991) A nucleolus orga-nizer region in a B chromosome inactivated by DNA methylation.Chromosoma 100: 134-138
- Neves N, Schwarzacher T, Heslop-Harrison lS, Hewitt GM,Camacho lPM (1994) Possible origin of a B chromosome deducedfrom its DNA composition using double FISH technique. Chro-mosome Res 2: 87-92
M. Bakkali et al.: Population differences in NOR expression
- Cabrero 1,Camacho lPM (1995) Changes in NOR activity patternin the presence of supernumerary heterochromatin in the grass-hopper Eyprepocnemis plorans. Genome 38: 68-74
Mellink CHM, Bosma AA, De Haan NA (1994) Variation in size ofAg-NORs and fiuorescent rDNA in situ hybridization signals insix breeds of domestic pig. Hereditas 120: 141-149
Miller L, Brown DD (1969) Variation in the activity of nucleolusorganizes and their ribosomal gene contento Chromosoma 28:430-444
Rice WR (1989) Analysing tables of statistical tests. Evolution 43:223-225
Rufas lS, Iturra P,De Souza W,Esponda P (1982) Simple silver stain-ing procedure for the locahzation of nucleolus and nucleolus orga-nizes under light and electron microscopy. Arch Biol 93: 267-276
Salceda Fl, Viseras E, Camacho lPM (1988) The B chromosomes ofLocusta migratoria IIr: effects on the activity of nucleolar orga-nizer regions. Genome 30: 387-394
Sánchez A, Burgos M, Jiménez R, Díaz de la Guardia R (1989)Quantitative analysis of silver staining of the nucleolus organiz-ing region in Eliomys quercinus. Genome 32: 978-982
Sasaki M, Nishida C, Kodama Y (1986) Characterization of silverstained nucleolus organizer regions (AG-NORs) in 16 inbredstrains of the Norway rat, Rattus norvegicus. Cytogenet CellGenet 41: 83-88
Schmidtke 1, Schulte B, Kuhl P, Engel W (1976) Gene action in fishof tetraploid origin V: Cellular RNA and protein content andenzyme activities in cyprinid, clupeoid, and salmonoid species.Biochem Genet 14: 975-980
Shubert I, Künzel G (1990) Position dependent NOR activity inbarley. Chromosoma 99: 352-359
Sinclair lH, Carral CR, Humphrey RR (1974) Variation in rDNAredundance level and NOR length in normal and variant hiles ofthe Mexican axolotl. 1 Cell Sci 15: 239-257
Stitou S, Díaz de la Guardia R, liménez R, Burgos M (2000) Inac-tive ribosomal cistrons are spread throughout the B chromosomesof Rattus rattus (Rodentia, Muridae): implications for their originand evolution. Chromosome Res 8: 305-311
Suzuki H, Kurihara Y, Kanehisa T, Moriwaki K (1990) Variation inthe distribution of the silver staining nucleolus organizer regionson the chromosomes of the wild mouse, Mus musculus. Mol BiolEvol 7: 271-282
Wachtler F, Hopman AH, Wiegant 1, Schwarzacher HG (1986) Onthe position of nucleolus organizer regions (NORs) in interphasenuclei: studies with a new, non autoradiographic in situ hybridiza-tion method. Exp Cell Res 167: 227-240
Yoder lA, Walsh Cp, Bestor TH (1997) Cytosine methylation andthe ecology of intragenomic parasites. Trends Genet 13: 335-340
Zatsepina OV, Schofer C, Weipoltshammer K, Mosgoeller W,Almeder M, Stefanova VN, lordan EG, Wachtler F (1996) TheRNA-polymerase-I transcription factor UF and rDNA are 10-cated at the same majar sites in both interphase and mitotic pigembryonic kidney. Cytogenet Cell Genet 73: 274-278
Zurita F, liménez R, Burgos M, Díaz de la Guardia R (1998)Sequential silver staining and in situ hybridization reveal a directassociation between rDNA levels and the expression of homolo-gous nucleolus organizing regions: a hypothesis for NOR struc-ture and function. 1 Cell Sci 111: 1433-1439
.;




















