
Photoprotective role of NADPH:protochlorophyllide oxidoreductase A
6
Photoprotective role of NADPH:protochlorophyllide oxidoreductase A Frank Buhr* † , Majida El Bakkouri †‡ , Oscar Valdez*, Stephan Pollmann § , Nikolai Lebedev ¶ , Steffen Reinbothe ‡ , and Christiane Reinbothe* ‡ *Lehrstuhl fu ¨ r Pflanzenphysiologie, Universita ¨ t Bayreuth, Universita ¨ tsstrasse 30, D-95447 Bayreuth, Germany; ‡ Centre d’Etudes et de Recherches sur les Macromole ´ cules Organiques, Universite ´ Joseph Fourier, Centre National de la Recherche Scientifique, Fédération de Recherche en Evolution 3017, B.P. 53, F-38041 Grenoble Cedex 9, France; § Lehrstuhl fu ¨ r Pflanzenphysiologie, Ruhr-Universita ¨ t Bochum, Universita ¨ tsstrasse 150, D-44801 Bochum, Germany; and ¶ Center for BioMolecular Sciences and Engineering, U.S. Naval Research Laboratory, 4555 Overlook Avenue, Washington, DC 20375-5348 Communicated by Diter von Wettstein, Washington State University, Pullman, WA, April 24, 2008 (received for review February 1, 2008) A homology model of NADPH:protochlorophyllide (Pchlide) oxi- doreductase A (POR; E.C. 1.3.33.1) of barley is developed and verified by site-directed mutagenesis. PORA is considered a glob- ular protein consisting of nine -helices and seven -strands. The model predicts the presence of two functionally distinctive Pchlide binding sites where the pigment is coordinated by cystein residues. The pigment bound to the first, high-affinity Pchlide binding site is used for the formation of the photoactive state of the enzyme. The pigment bound to the second, low-affinity Pchlide binding site is involved in the PORA:PORB interaction, allowing for resonance energy transfer between the neighboring PORs in the complex. In the in vitro reconstituted light-harvesting POR:Pchlide complex (LHPP), light absorbed by PORA-bound Pchlide b is transferred to PORB-bound Pchlide a. That induces the conversion of Pchlide a to chlorophyllide (Chlide) a. This energy transfer eliminates the pos- sibility of Pchlide b photoreduction and prevents that excited triplet states of either Pchlides a or b accumulate and provoke singlet oxygen production. Together, our results provide a photo- protective role of PORA during greening. chlorophyll biosynthesis chloroplast development greening reactive oxygen species A key enzyme in the light-induced greening of higher plants is NADPH:protochlorophyllide (Pchlide) oxidoreductase (POR). In angiosperms such as barley, Arabidopsis (1, 2), tobacco (3) and Amaranthus tricolor (4), and gymnosperms such as pine species (5–7), several por gene families were identified that encode highly conserved POR polypeptides. PORA represents the nega- tively light-regulated POR enzyme whose level drops as a result of the concerted effect of light at the levels of transcription, mRNA stability, plastid import, and protein degradation after light-induced catalysis (8–11). PORB, the second POR protein identified in barley and Arabidopsis (1, 12), is constitutively expressed in dark- grown, illuminated, and light-adapted plants (see ref. 11 for review). A third light-induced por gene was discovered in Arabidopsis thaliana that complements PORB in green plants (13, 14). PORA and PORB constitute larger light-harvesting POR:Pch- lide complexes dubbed LHPP in the prolamellar body of etioplasts (15–18). These complexes additionally interact with galacto- and sulfolipids and establish two spectral forms of Pchlide: Pchlide-650/ 657 and Pchlide-628/632 (the first number indicates the absorption maximum, the second shows the respective fluorescence emission maximum). Pchlide-650/657 is also called photoactive Pchlide be- cause it can be converted to Chlide-684/690 with a single, 1-ms f lash of white light. Pchlide-628/632, by contrast, is nonphotoconvertible and named photoinactive Pchlide. Energy transfer takes place from photoinactive Pchlide to photoactive Pchlide and from photoinac- tive Pchlide to Chlide (summarized in ref. 19). Similar spectral pigment species and interconversions have been described for reconstituted PORA:PORB-lipid supracomplexes (15–18). All in- formation available at the moment suggests that the native and reconstituted complexes are structurally and functionally identical. The two major roles attributed to LHPP are light trapping and the dissipation of excess light, both of which are needed when dark- grown seedlings de-etiolate (15). In the present work, we provide direct evidence for a photoprotective role of PORA during greening. Results Homology Modeling of Barley PORA and PORB. The barley PORA and PORB are related in their primary amino acid sequences to short-chain dehydrogenases (SCDs) (refs. 20 –22 and see Fig. 1A for sequence comparison). For these enzymes, x-ray structures have been obtained (23–25), allowing homology models to be con- structed with MODELLER 9v3 software and 7--hydroxysteroid dehydrogenase (AHI) from Escherichia coli as a template. The obtained PORA and PORB structural models are shown in Fig. 1 B and C. Accordingly, the PORA and PORB of barley are predicted to be globular proteins consisting of nine -helices and seven -strands. Similar -helices and -strands are present in POR of Synechocystis (synPOR) and are marked -(A-I) and -(1–7), respectively, in Fig. 1. Amino acid conservation in synPOR and the barley PORA and PORB is almost perfect in the so-called Ross- mann fold representing the adenine binding cleft in the SCDs. Amino acid residues already established as being necessary for structural and functional reasons in the SCDs are also conserved in synPOR and the barley PORA and PORB (Fig. 1). It is assumed that the highly conserved Tyr and Lys residues in the YxxxK motif, in conjunction with the GASSGK/LG motif, establish the enzyme catalytic pocket and participate in the proper coordination of NADPH. The -amino group of the Lys side chain hereby is proposed to interact with the hydroxyl group of Tyr and to lower its pK a such that a proton can be donated during the catalytic mechanism. In addition, both amino acid residues are implicated in the formation of the POR photoactive state with Pchlide (22, 26 –28). The participation of one or two Cys residues in Pchlide binding has been proposed (22, 29). SynPOR contains six Cys residues of which Cys-33, Cys-85, Cys-195, and Cys-222, henceforth referred to as Cys-1, Cys-2, Cys-3, and Cys-4, respectively, have counterparts in the PORA and PORB of barley (Fig. 1 A and Table S1). Evidence for the involvement of Cys residues in pigment binding and catalysis has come from chemical modification of Cys residues with N-phenylmaleimide and other sulfhydryl group-modifying Author contributions: C.R. designed research; F.B., M.E.B., O.V., S.P., S.R., and C.R. per- formed research; F.B., M.E.B., O.V., S.P., N.L., and S.R. analyzed data; and C.R. wrote the paper. The authors declare no conflict of interest. † F.B. and M.E.B. contributed equally to this work. To whom correspondence should be addressed. E-mail: christiane.reinbothe@uni- bayreuth.de or [email protected]. This article contains supporting information online at www.pnas.org/cgi/content/full/ 0803950105/DCSupplemental. © 2008 by The National Academy of Sciences of the USA www.pnas.orgcgidoi10.1073pnas.0803950105 PNAS August 26, 2008 vol. 105 no. 34 12629 –12634 PLANT BIOLOGY
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Photoprotective role of NADPH:protochlorophyllide oxidoreductase A

Photoprotective role of NADPH:protochlorophyllideoxidoreductase AFrank Buhr*†, Majida El Bakkouri†‡, Oscar Valdez*, Stephan Pollmann§, Nikolai Lebedev¶, Steffen Reinbothe‡,and Christiane Reinbothe*‡�
*Lehrstuhl fur Pflanzenphysiologie, Universitat Bayreuth, Universitatsstrasse 30, D-95447 Bayreuth, Germany; ‡Centre d’Etudes et de Recherches sur lesMacromolecules Organiques, Universite Joseph Fourier, Centre National de la Recherche Scientifique, Fédération de Recherche en Evolution 3017, B.P. 53,F-38041 Grenoble Cedex 9, France; §Lehrstuhl fur Pflanzenphysiologie, Ruhr-Universitat Bochum, Universitatsstrasse 150, D-44801 Bochum, Germany; and¶Center for BioMolecular Sciences and Engineering, U.S. Naval Research Laboratory, 4555 Overlook Avenue, Washington, DC 20375-5348
Communicated by Diter von Wettstein, Washington State University, Pullman, WA, April 24, 2008 (received for review February 1, 2008)
A homology model of NADPH:protochlorophyllide (Pchlide) oxi-doreductase A (POR; E.C. 1.3.33.1) of barley is developed andverified by site-directed mutagenesis. PORA is considered a glob-ular protein consisting of nine �-helices and seven �-strands. Themodel predicts the presence of two functionally distinctive Pchlidebinding sites where the pigment is coordinated by cystein residues.The pigment bound to the first, high-affinity Pchlide binding site isused for the formation of the photoactive state of the enzyme. Thepigment bound to the second, low-affinity Pchlide binding site isinvolved in the PORA:PORB interaction, allowing for resonanceenergy transfer between the neighboring PORs in the complex. Inthe in vitro reconstituted light-harvesting POR:Pchlide complex(LHPP), light absorbed by PORA-bound Pchlide b is transferred toPORB-bound Pchlide a. That induces the conversion of Pchlide a tochlorophyllide (Chlide) a. This energy transfer eliminates the pos-sibility of Pchlide b photoreduction and prevents that excitedtriplet states of either Pchlides a or b accumulate and provokesinglet oxygen production. Together, our results provide a photo-protective role of PORA during greening.
chlorophyll biosynthesis � chloroplast development � greening �reactive oxygen species
A key enzyme in the light-induced greening of higher plants isNADPH:protochlorophyllide (Pchlide) oxidoreductase
(POR). In angiosperms such as barley, Arabidopsis (1, 2), tobacco(3) and Amaranthus tricolor (4), and gymnosperms such as pinespecies (5–7), several por gene families were identified that encodehighly conserved POR polypeptides. PORA represents the nega-tively light-regulated POR enzyme whose level drops as a result ofthe concerted effect of light at the levels of transcription, mRNAstability, plastid import, and protein degradation after light-inducedcatalysis (8–11). PORB, the second POR protein identified inbarley and Arabidopsis (1, 12), is constitutively expressed in dark-grown, illuminated, and light-adapted plants (see ref. 11 for review).A third light-induced por gene was discovered in Arabidopsisthaliana that complements PORB in green plants (13, 14).
PORA and PORB constitute larger light-harvesting POR:Pch-lide complexes dubbed LHPP in the prolamellar body of etioplasts(15–18). These complexes additionally interact with galacto- andsulfolipids and establish two spectral forms of Pchlide: Pchlide-650/657 and Pchlide-628/632 (the first number indicates the absorptionmaximum, the second shows the respective fluorescence emissionmaximum). Pchlide-650/657 is also called photoactive Pchlide be-cause it can be converted to Chlide-684/690 with a single, 1-ms flashof white light. Pchlide-628/632, by contrast, is nonphotoconvertibleand named photoinactive Pchlide. Energy transfer takes place fromphotoinactive Pchlide to photoactive Pchlide and from photoinac-tive Pchlide to Chlide (summarized in ref. 19). Similar spectralpigment species and interconversions have been described forreconstituted PORA:PORB-lipid supracomplexes (15–18). All in-formation available at the moment suggests that the native andreconstituted complexes are structurally and functionally identical.
The two major roles attributed to LHPP are light trapping and thedissipation of excess light, both of which are needed when dark-grown seedlings de-etiolate (15). In the present work, we providedirect evidence for a photoprotective role of PORA duringgreening.
ResultsHomology Modeling of Barley PORA and PORB. The barley PORAand PORB are related in their primary amino acid sequences toshort-chain dehydrogenases (SCDs) (refs. 20–22 and see Fig. 1A forsequence comparison). For these enzymes, x-ray structures havebeen obtained (23–25), allowing homology models to be con-structed with MODELLER 9v3 software and 7-�-hydroxysteroiddehydrogenase (AHI) from Escherichia coli as a template.
The obtained PORA and PORB structural models are shown inFig. 1 B and C. Accordingly, the PORA and PORB of barley arepredicted to be globular proteins consisting of nine �-helices andseven �-strands. Similar �-helices and �-strands are present in PORof Synechocystis (synPOR) and are marked �-(A-I) and �-(1–7),respectively, in Fig. 1. Amino acid conservation in synPOR and thebarley PORA and PORB is almost perfect in the so-called Ross-mann fold representing the adenine binding cleft in the SCDs.Amino acid residues already established as being necessary forstructural and functional reasons in the SCDs are also conserved insynPOR and the barley PORA and PORB (Fig. 1). It is assumedthat the highly conserved Tyr and Lys residues in the YxxxK motif,in conjunction with the GASSGK/LG motif, establish the enzymecatalytic pocket and participate in the proper coordination ofNADPH. The �-amino group of the Lys side chain hereby isproposed to interact with the hydroxyl group of Tyr and to lowerits pKa such that a proton can be donated during the catalyticmechanism. In addition, both amino acid residues are implicated inthe formation of the POR photoactive state with Pchlide (22,26–28).
The participation of one or two Cys residues in Pchlide bindinghas been proposed (22, 29). SynPOR contains six Cys residues ofwhich Cys-33, Cys-85, Cys-195, and Cys-222, henceforth referred toas Cys-1, Cys-2, Cys-3, and Cys-4, respectively, have counterparts inthe PORA and PORB of barley (Fig. 1A and Table S1).
Evidence for the involvement of Cys residues in pigment bindingand catalysis has come from chemical modification of Cys residueswith N-phenylmaleimide and other sulfhydryl group-modifying
Author contributions: C.R. designed research; F.B., M.E.B., O.V., S.P., S.R., and C.R. per-formed research; F.B., M.E.B., O.V., S.P., N.L., and S.R. analyzed data; and C.R. wrote thepaper.
The authors declare no conflict of interest.
†F.B. and M.E.B. contributed equally to this work.
�To whom correspondence should be addressed. E-mail: [email protected] or [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0803950105/DCSupplemental.
© 2008 by The National Academy of Sciences of the USA
www.pnas.org�cgi�doi�10.1073�pnas.0803950105 PNAS � August 26, 2008 � vol. 105 � no. 34 � 12629–12634
PLA
NT
BIO
LOG
Y
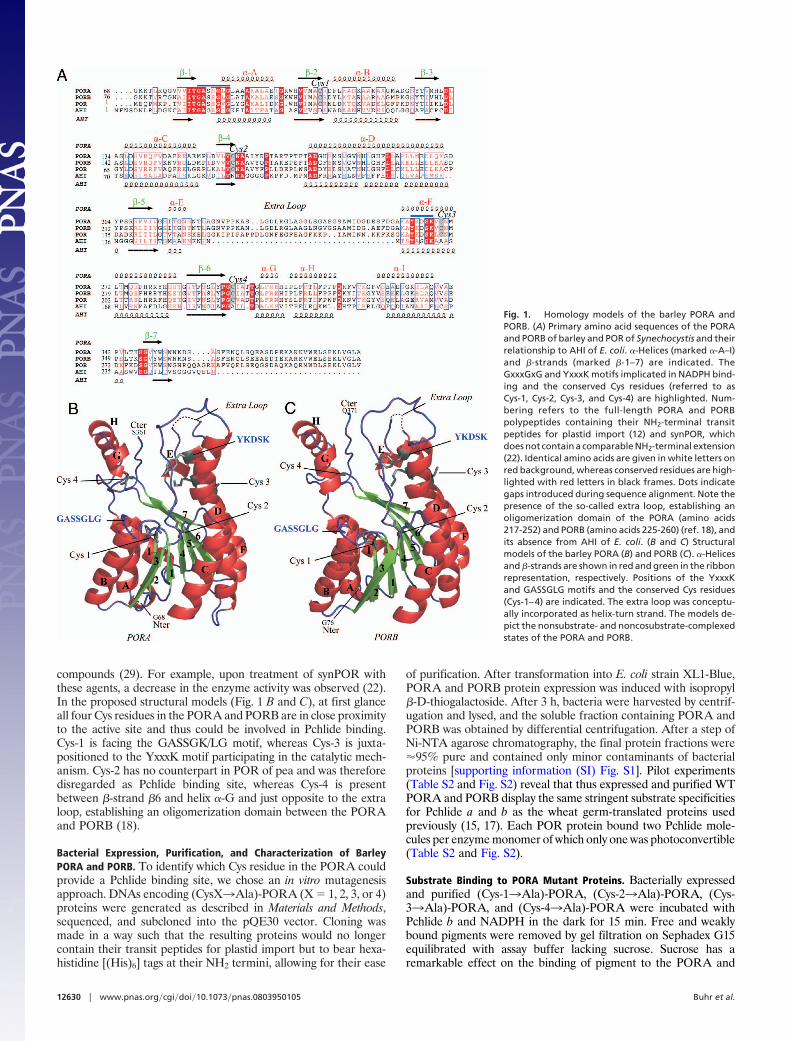
compounds (29). For example, upon treatment of synPOR withthese agents, a decrease in the enzyme activity was observed (22).In the proposed structural models (Fig. 1 B and C), at first glanceall four Cys residues in the PORA and PORB are in close proximityto the active site and thus could be involved in Pchlide binding.Cys-1 is facing the GASSGK/LG motif, whereas Cys-3 is juxta-positioned to the YxxxK motif participating in the catalytic mech-anism. Cys-2 has no counterpart in POR of pea and was thereforedisregarded as Pchlide binding site, whereas Cys-4 is presentbetween �-strand �6 and helix �-G and just opposite to the extraloop, establishing an oligomerization domain between the PORAand PORB (18).
Bacterial Expression, Purification, and Characterization of BarleyPORA and PORB. To identify which Cys residue in the PORA couldprovide a Pchlide binding site, we chose an in vitro mutagenesisapproach. DNAs encoding (CysX3Ala)-PORA (X � 1, 2, 3, or 4)proteins were generated as described in Materials and Methods,sequenced, and subcloned into the pQE30 vector. Cloning wasmade in a way such that the resulting proteins would no longercontain their transit peptides for plastid import but to bear hexa-histidine [(His)6] tags at their NH2 termini, allowing for their ease
of purification. After transformation into E. coli strain XL1-Blue,PORA and PORB protein expression was induced with isopropyl�-D-thiogalactoside. After 3 h, bacteria were harvested by centrif-ugation and lysed, and the soluble fraction containing PORA andPORB was obtained by differential centrifugation. After a step ofNi-NTA agarose chromatography, the final protein fractions were�95% pure and contained only minor contaminants of bacterialproteins [supporting information (SI) Fig. S1]. Pilot experiments(Table S2 and Fig. S2) reveal that thus expressed and purified WTPORA and PORB display the same stringent substrate specificitiesfor Pchlide a and b as the wheat germ-translated proteins usedpreviously (15, 17). Each POR protein bound two Pchlide mole-cules per enzyme monomer of which only one was photoconvertible(Table S2 and Fig. S2).
Substrate Binding to PORA Mutant Proteins. Bacterially expressedand purified (Cys-13Ala)-PORA, (Cys-23Ala)-PORA, (Cys-33Ala)-PORA, and (Cys-43Ala)-PORA were incubated withPchlide b and NADPH in the dark for 15 min. Free and weaklybound pigments were removed by gel filtration on Sephadex G15equilibrated with assay buffer lacking sucrose. Sucrose has aremarkable effect on the binding of pigment to the PORA and
Fig. 1. Homology models of the barley PORA andPORB. (A) Primary amino acid sequences of the PORAand PORB of barley and POR of Synechocystis and theirrelationship to AHI of E. coli. �-Helices (marked �-A–I)and �-strands (marked �-1–7) are indicated. TheGxxxGxG and YxxxK motifs implicated in NADPH bind-ing and the conserved Cys residues (referred to asCys-1, Cys-2, Cys-3, and Cys-4) are highlighted. Num-bering refers to the full-length PORA and PORBpolypeptides containing their NH2-terminal transitpeptides for plastid import (12) and synPOR, whichdoes not contain a comparable NH2-terminal extension(22). Identical amino acids are given in white letters onred background, whereas conserved residues are high-lighted with red letters in black frames. Dots indicategaps introduced during sequence alignment. Note thepresence of the so-called extra loop, establishing anoligomerization domain of the PORA (amino acids217-252) and PORB (amino acids 225-260) (ref. 18), andits absence from AHI of E. coli. (B and C) Structuralmodels of the barley PORA (B) and PORB (C). �-Helicesand �-strands are shown in red and green in the ribbonrepresentation, respectively. Positions of the YxxxKand GASSGLG motifs and the conserved Cys residues(Cys-1–4) are indicated. The extra loop was conceptu-ally incorporated as helix-turn strand. The models de-pict the nonsubstrate- and noncosubstrate-complexedstates of the PORA and PORB.
12630 � www.pnas.org�cgi�doi�10.1073�pnas.0803950105 Buhr et al.
PORB and stabilizes weakly bound pigments (Table S2 and ref. 30).(CysX3Ala)-PORA-pigment-NADPH ternary complexes elutedwith the flow-through in turn were extracted with acetone. Thepigments were identified and quantified by HPLC, room temper-ature absorption, and fluorescence spectroscopy (17).
Table 1 shows that, similar to the WT PORA (Table S2), the(Cys-13Ala)-PORA and (Cys-23Ala)-PORA each bound twoPchlide b molecules per enzyme monomer, only one of which wasphotoconvertible and transformed to Chlide b (see Fig. S3 A and B).Meanwhile, (Cys-33Ala)-PORA and (Cys-43Ala)-PORA wereable to bind only one Pchlide b molecule per enzyme monomer(Table 1 and Fig. S3 C and D). While Pchlide b attached to(Cys-33Ala)-PORA was nonphotoconvertible and only weaklybound to the enzyme, Pchlide b was tightly bound to (Cys-43Ala)-PORA and readily converted to Chlide b upon illumination (Table1 and Fig. S3 C and D).
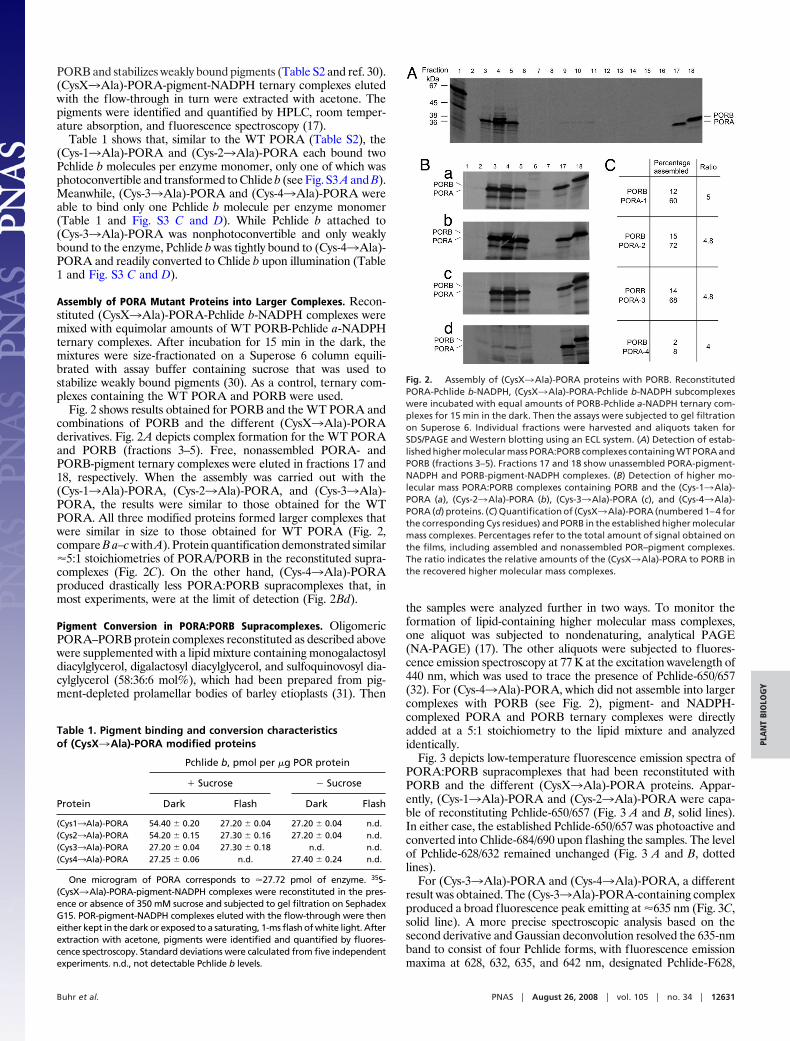
Assembly of PORA Mutant Proteins into Larger Complexes. Recon-stituted (CysX3Ala)-PORA-Pchlide b-NADPH complexes weremixed with equimolar amounts of WT PORB-Pchlide a-NADPHternary complexes. After incubation for 15 min in the dark, themixtures were size-fractionated on a Superose 6 column equili-brated with assay buffer containing sucrose that was used tostabilize weakly bound pigments (30). As a control, ternary com-plexes containing the WT PORA and PORB were used.
Fig. 2 shows results obtained for PORB and the WT PORA andcombinations of PORB and the different (CysX3Ala)-PORAderivatives. Fig. 2A depicts complex formation for the WT PORAand PORB (fractions 3–5). Free, nonassembled PORA- andPORB-pigment ternary complexes were eluted in fractions 17 and18, respectively. When the assembly was carried out with the(Cys-13Ala)-PORA, (Cys-23Ala)-PORA, and (Cys-33Ala)-PORA, the results were similar to those obtained for the WTPORA. All three modified proteins formed larger complexes thatwere similar in size to those obtained for WT PORA (Fig. 2,compare B a–c with A). Protein quantification demonstrated similar�5:1 stoichiometries of PORA/PORB in the reconstituted supra-complexes (Fig. 2C). On the other hand, (Cys-43Ala)-PORAproduced drastically less PORA:PORB supracomplexes that, inmost experiments, were at the limit of detection (Fig. 2Bd).
Pigment Conversion in PORA:PORB Supracomplexes. OligomericPORA–PORB protein complexes reconstituted as described abovewere supplemented with a lipid mixture containing monogalactosyldiacylglycerol, digalactosyl diacylglycerol, and sulfoquinovosyl dia-cylglycerol (58:36:6 mol%), which had been prepared from pig-ment-depleted prolamellar bodies of barley etioplasts (31). Then
the samples were analyzed further in two ways. To monitor theformation of lipid-containing higher molecular mass complexes,one aliquot was subjected to nondenaturing, analytical PAGE(NA-PAGE) (17). The other aliquots were subjected to fluores-cence emission spectroscopy at 77 K at the excitation wavelength of440 nm, which was used to trace the presence of Pchlide-650/657(32). For (Cys-43Ala)-PORA, which did not assemble into largercomplexes with PORB (see Fig. 2), pigment- and NADPH-complexed PORA and PORB ternary complexes were directlyadded at a 5:1 stoichiometry to the lipid mixture and analyzedidentically.
Fig. 3 depicts low-temperature fluorescence emission spectra ofPORA:PORB supracomplexes that had been reconstituted withPORB and the different (CysX3Ala)-PORA proteins. Appar-ently, (Cys-13Ala)-PORA and (Cys-23Ala)-PORA were capa-ble of reconstituting Pchlide-650/657 (Fig. 3 A and B, solid lines).In either case, the established Pchlide-650/657 was photoactive andconverted into Chlide-684/690 upon flashing the samples. The levelof Pchlide-628/632 remained unchanged (Fig. 3 A and B, dottedlines).
For (Cys-33Ala)-PORA and (Cys-43Ala)-PORA, a differentresult was obtained. The (Cys-33Ala)-PORA-containing complexproduced a broad fluorescence peak emitting at �635 nm (Fig. 3C,solid line). A more precise spectroscopic analysis based on thesecond derivative and Gaussian deconvolution resolved the 635-nmband to consist of four Pchlide forms, with fluorescence emissionmaxima at 628, 632, 635, and 642 nm, designated Pchlide-F628,
Table 1. Pigment binding and conversion characteristicsof (CysX3Ala)-PORA modified proteins
Protein
Pchlide b, pmol per �g POR protein
� Sucrose � Sucrose
Dark Flash Dark Flash
(Cys13Ala)-PORA 54.40 � 0.20 27.20 � 0.04 27.20 � 0.04 n.d.(Cys23Ala)-PORA 54.20 � 0.15 27.30 � 0.16 27.20 � 0.04 n.d.(Cys33Ala)-PORA 27.20 � 0.04 27.30 � 0.18 n.d. n.d.(Cys43Ala)-PORA 27.25 � 0.06 n.d. 27.40 � 0.24 n.d.
One microgram of PORA corresponds to �27.72 pmol of enzyme. 35S-(CysX3Ala)-PORA-pigment-NADPH complexes were reconstituted in the pres-ence or absence of 350 mM sucrose and subjected to gel filtration on SephadexG15. POR-pigment-NADPH complexes eluted with the flow-through were theneither kept in the dark or exposed to a saturating, 1-ms flash of white light. Afterextraction with acetone, pigments were identified and quantified by fluores-cence spectroscopy. Standard deviations were calculated from five independentexperiments. n.d., not detectable Pchlide b levels.
Fig. 2. Assembly of (CysX3Ala)-PORA proteins with PORB. ReconstitutedPORA-Pchlide b-NADPH, (CysX3Ala)-PORA-Pchlide b-NADPH subcomplexeswere incubated with equal amounts of PORB-Pchlide a-NADPH ternary com-plexes for 15 min in the dark. Then the assays were subjected to gel filtrationon Superose 6. Individual fractions were harvested and aliquots taken forSDS/PAGE and Western blotting using an ECL system. (A) Detection of estab-lished higher molecular mass PORA:PORB complexes containing WT PORA andPORB (fractions 3–5). Fractions 17 and 18 show unassembled PORA-pigment-NADPH and PORB-pigment-NADPH complexes. (B) Detection of higher mo-lecular mass PORA:PORB complexes containing PORB and the (Cys-13Ala)-PORA (a), (Cys-23Ala)-PORA (b), (Cys-33Ala)-PORA (c), and (Cys-43Ala)-PORA (d) proteins. (C) Quantification of (CysX3Ala)-PORA (numbered 1–4 forthe corresponding Cys residues) and PORB in the established higher molecularmass complexes. Percentages refer to the total amount of signal obtained onthe films, including assembled and nonassembled POR–pigment complexes.The ratio indicates the relative amounts of the (CysX3Ala)-PORA to PORB inthe recovered higher molecular mass complexes.
Buhr et al. PNAS � August 26, 2008 � vol. 105 � no. 34 � 12631
PLA
NT
BIO
LOG
Y
Pchlide-F632, Pchlide-F635, and Pchlide-F642, respectively. Uponflash light illumination, almost no visible change occurred in thelow-temperature Pchlide fluorescence emission spectrum, exceptfor a minor decrease in the Pchlide-F635 and Pchlide-F642 peaks.In addition, a new pigment fluorescence peak appeared with anemission maximum at 672 nm (Fig. 3C, dotted line). Pigmentanalysis in acetone at room temperature by fluorescence spectros-copy and HPLC on C30 RP columns showed that small amounts ofChlide a had been produced (data not shown). By contrast, noChlide a was detectable for assays containing (Cys-43Ala)-PORA(Fig. 3D and data not shown).
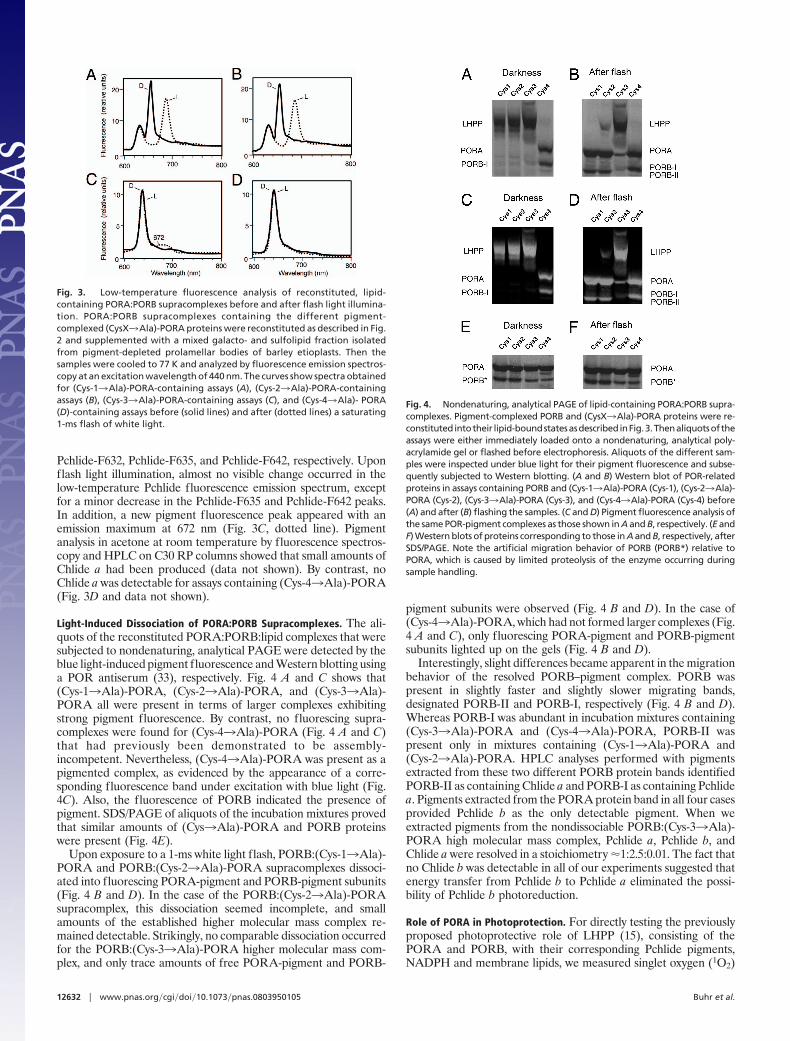
Light-Induced Dissociation of PORA:PORB Supracomplexes. The ali-quots of the reconstituted PORA:PORB:lipid complexes that weresubjected to nondenaturing, analytical PAGE were detected by theblue light-induced pigment fluorescence and Western blotting usinga POR antiserum (33), respectively. Fig. 4 A and C shows that(Cys-13Ala)-PORA, (Cys-23Ala)-PORA, and (Cys-33Ala)-PORA all were present in terms of larger complexes exhibitingstrong pigment fluorescence. By contrast, no fluorescing supra-complexes were found for (Cys-43Ala)-PORA (Fig. 4 A and C)that had previously been demonstrated to be assembly-incompetent. Nevertheless, (Cys-43Ala)-PORA was present as apigmented complex, as evidenced by the appearance of a corre-sponding fluorescence band under excitation with blue light (Fig.4C). Also, the fluorescence of PORB indicated the presence ofpigment. SDS/PAGE of aliquots of the incubation mixtures provedthat similar amounts of (Cys3Ala)-PORA and PORB proteinswere present (Fig. 4E).
Upon exposure to a 1-ms white light flash, PORB:(Cys-13Ala)-PORA and PORB:(Cys-23Ala)-PORA supracomplexes dissoci-ated into fluorescing PORA-pigment and PORB-pigment subunits(Fig. 4 B and D). In the case of the PORB:(Cys-23Ala)-PORAsupracomplex, this dissociation seemed incomplete, and smallamounts of the established higher molecular mass complex re-mained detectable. Strikingly, no comparable dissociation occurredfor the PORB:(Cys-33Ala)-PORA higher molecular mass com-plex, and only trace amounts of free PORA-pigment and PORB-
pigment subunits were observed (Fig. 4 B and D). In the case of(Cys-43Ala)-PORA, which had not formed larger complexes (Fig.4 A and C), only fluorescing PORA-pigment and PORB-pigmentsubunits lighted up on the gels (Fig. 4 B and D).
Interestingly, slight differences became apparent in the migrationbehavior of the resolved PORB–pigment complex. PORB waspresent in slightly faster and slightly slower migrating bands,designated PORB-II and PORB-I, respectively (Fig. 4 B and D).Whereas PORB-I was abundant in incubation mixtures containing(Cys-33Ala)-PORA and (Cys-43Ala)-PORA, PORB-II waspresent only in mixtures containing (Cys-13Ala)-PORA and(Cys-23Ala)-PORA. HPLC analyses performed with pigmentsextracted from these two different PORB protein bands identifiedPORB-II as containing Chlide a and PORB-I as containing Pchlidea. Pigments extracted from the PORA protein band in all four casesprovided Pchlide b as the only detectable pigment. When weextracted pigments from the nondissociable PORB:(Cys-33Ala)-PORA high molecular mass complex, Pchlide a, Pchlide b, andChlide a were resolved in a stoichiometry �1:2.5:0.01. The fact thatno Chlide b was detectable in all of our experiments suggested thatenergy transfer from Pchlide b to Pchlide a eliminated the possi-bility of Pchlide b photoreduction.
Role of PORA in Photoprotection. For directly testing the previouslyproposed photoprotective role of LHPP (15), consisting of thePORA and PORB, with their corresponding Pchlide pigments,NADPH and membrane lipids, we measured singlet oxygen (1O2)
Fig. 3. Low-temperature fluorescence analysis of reconstituted, lipid-containing PORA:PORB supracomplexes before and after flash light illumina-tion. PORA:PORB supracomplexes containing the different pigment-complexed (CysX3Ala)-PORA proteins were reconstituted as described in Fig.2 and supplemented with a mixed galacto- and sulfolipid fraction isolatedfrom pigment-depleted prolamellar bodies of barley etioplasts. Then thesamples were cooled to 77 K and analyzed by fluorescence emission spectros-copy at an excitation wavelength of 440 nm. The curves show spectra obtainedfor (Cys-13Ala)-PORA-containing assays (A), (Cys-23Ala)-PORA-containingassays (B), (Cys-33Ala)-PORA-containing assays (C), and (Cys-43Ala)- PORA(D)-containing assays before (solid lines) and after (dotted lines) a saturating1-ms flash of white light.
Fig. 4. Nondenaturing, analytical PAGE of lipid-containing PORA:PORB supra-complexes. Pigment-complexed PORB and (CysX3Ala)-PORA proteins were re-constituted intotheir lipid-boundstatesasdescribedinFig.3.Thenaliquotsoftheassays were either immediately loaded onto a nondenaturing, analytical poly-acrylamide gel or flashed before electrophoresis. Aliquots of the different sam-ples were inspected under blue light for their pigment fluorescence and subse-quently subjected to Western blotting. (A and B) Western blot of POR-relatedproteins in assays containing PORB and (Cys-13Ala)-PORA (Cys-1), (Cys-23Ala)-PORA (Cys-2), (Cys-33Ala)-PORA (Cys-3), and (Cys-43Ala)-PORA (Cys-4) before(A) and after (B) flashing the samples. (C and D) Pigment fluorescence analysis ofthe same POR-pigment complexes as those shown in A and B, respectively. (E andF) Western blots of proteins corresponding to those in A and B, respectively, afterSDS/PAGE. Note the artificial migration behavior of PORB (PORB*) relative toPORA, which is caused by limited proteolysis of the enzyme occurring duringsample handling.
12632 � www.pnas.org�cgi�doi�10.1073�pnas.0803950105 Buhr et al.

production in vitro. Lipid-containing PORA:PORB supracom-plexes were reconstituted as described in Fig. 4, and the methodused the DanePy reagent (34, 35), which is a dansyl-based 1O2sensor undergoing quenching of its green fluorescence upon react-ing with singlet oxygen.
Fig. 5 shows time courses of 1O2 evolution for the different(CysX3Ala)-PORA-Pchlide b-NADPH:PORB-Pchlide a-NADPH:lipid supracomplexes and unassembled ternary com-plexes. For assays containing (Cys-13Ala)-PORA- and (Cys-23Ala)-PORA, no 1O2 production was detectable. For assaysconsisting of nonphotodissociable (Cys-33Ala)-PORA:PORBcomplexes, low rates of 1O2 production were measured that reacheda plateau after 10 min. By contrast, 1O2 production was linear forat least 1 h and yielded �10-fold higher final 1O2 levels in assaysconsisting of unassembled (Cys-43Ala)-PORA and PORB. Thelinear relationship between 1O2 production and time excluded thepossibility that the nitroxide radicals formed from DanePy werepartly reduced and transformed into inactive states in the sam-ples (34).
DiscussionIn the present work, a structure–function study was performed forthe PORA of barley. PORA represents the negatively light-regulated POR enzyme that rapidly disappears from etiolatedplants upon illumination. Its expression and role is confined toetiolated plants, where it operates in light harvesting, energytransfer, and photoprotection during greening (36). Both aspectsare illustrated in the present work. Using an in vitro-mutagenesisapproach, two distinctive Pchlide binding sites were identified in thePORA. Two of the four evolutionarily conserved Cys residues,namely Cys-268/200 (operationally defined as Cys-3) and Cys-295/227 (operationally defined as Cys-4) were identified as participatingin the establishment of the photoactive enzyme state and LHPPassembly, respectively. In the reconstituted LHPP complex, Pchlideb bound with NADPH to the PORA was involved in energy transferto PORB-bound Pchlide a. By virtue of this mechanism the risk waskept low that triplet states of Pchlides a and b accumulated andprovoked singlet oxygen production. Carotenoids present in theprolamellar body (37) may provide an additional mechanism toquench both excited singlet and triplet states of Pchlide and Chlidein etiolated plants undergoing greening.
Sequence alignments show that Cys-268/200 (Cys-3) is adjacentto the NADPH binding site. The overall sequence, which is definedas YKDSKVC in helix �–F, is identical in the PORA and PORBand is highly conserved between different POR enzymes, includingsynPOR (Fig. 1A) and pea POR (21). In the case of pea POR, theconserved Tyr and Lys residues have documented roles in NADPHand Pchlide binding. Lebedev et al. (28) reported that substrate andcosubstrate binding to POR occurs in two steps: an initial looseassociation of Pchlide and NADPH and a subsequent sort of‘‘induced fit’’ that gives rise to the photoactive enzyme state. In thestructural model depicted in Fig. 1, the YxxxK motif, comprising theuniversally conserved Tyr and Lys residues and the adjacent Cys-3,is readily accessible to its ligands. An as-yet-undetermined PORconformational change then is supposed to lead to the tightcoordination of all reactants (8, 9, 28).
Cys-295/227 (Cys-4) constitutes a second, low-affinity Pchlide bbinding site. We observed that Pchlide b bound to Cys-4 participatesin PORA:PORB interactions. Replacement of Cys-4 by an Alaresidue reduced by half the amount of bound pigment per PORAenzyme monomer and gave rise to assembly-incompetent PORAmolecules.
Both Cys-3 and Cys-4 are involved in energy transfer. This isapparent from the lack of Pchlide-650/657 in assays containing the(Cys-33Ala)-PORA and (Cys-43Ala)-PORA mutant proteins.In either case, no photoactive Pchlide-650/657 was detectable bylow-temperature fluorescence spectroscopy. Instead, bright pig-ment peaks appeared emitting at 630–635 nm. (Cys-33Ala)-PORA established higher molecular mass complexes similar in sizeto those formed with WT PORA and the (Cys-13Ala)-PORA and(Cys-23Ala)-PORA. But these (Cys-33Ala)-PORA-containingcomplexes were largely photoinactive and nondissociable upontreatment with flashes of white light. Trace amounts of Chlide awere produced. Spectroscopic evidence was obtained for a route ofchlorophyll a synthesis not involving Pchlide-650/657 but dependingon a Pchlide species fluorescing at 642 nm (Pchlide-F642). A similarPchlide species had been detected by Lebedev et al. (32) using thedet340/cop1 mutant of Arabidopsis thaliana that is largely depletedof PORA protein because of constitutive activation of phytochromesignal transduction depressing porA transcription. Lebedev et al.observed that etiolated det340/cop1 seedlings were especially light-sensitive (32). Similar results were reproduced in this work forPORB:(Cys-33Ala)-PORA supracomplexes reconstituted withmembrane lipids. We interpret our result as evidence for a partialprotection of PORB by (Cys-33Ala)-PORA from excess light.
A major breakthrough toward the understanding of PORA’spresumed photoprotective role during greening was provided by theexperiment described in Fig. 5. The measured rates of 1O2 produc-tion were extremely low for functional LHPP complexes. Presum-ably because of energy transfer from Pchlide b to Pchlide a andsubsequent Pchlide a to Chlide a reduction, no significant levels of1O2 were produced. By contrast, perturbation of LHPP’s normalfunction, such as assembly of (Cys-33Ala)-PORA with PORB intolarger, nonlight-dissociable complexes, led to a marked increase inthe rate of 1O2 evolution. Even more deleterious effects wereobserved for assays containing unassembled, but lipid-associated(Cys-43Ala)-PORA and PORB. In this case, probably most of theincoming photons drove singlet oxygen formation. Although theatomic details of LHPP are not known yet, our work defines aphotoprotective role of PORA during greening.
Materials and MethodsPrimers. Primers used were: Oligo-S, 5�-pCCGCGAGACCCACCCTTGGAGGCTCCA-GATTTATC-3�; M1 (Cys-1063Ala)-PORA, 5�-pCACGTCGTCATGGCGGCGCGC-GACTTCCTCAAG-3�; M2 (Cys-1583Ala)-PORA, 5�-pCTGGATGTGCTGGTCGC-GAACGCCGCCATCTAC-3�; M3 (Cys-2683Ala)-PORA, 5�-pAAGGACAGC-AAGGTGGCCACATGCTGACCATG-3�; and M4 (Cys-2953Ala)-PORA, 5�-pTCGCTCTACCCGGGCGCCATCGCCACG-GGG-3�.
Fig. 5. 1O2 production by reconstituted (CysX3Ala)-PORA:PORB:lipid com-plexes. Lipid-containing PORA:PORB supracomplexes containing the different(CysX3Ala)-PORA proteins were reconstituted as described in Fig. 3. 1O2
production was measured with the DanePy reagent after successive 1-msflashes of nonsaturating white light, each applied at 1-min intervals. E,(Cys-13Ala)-PORA; F, (Cys-23Ala)-PORA; �, (Cys-33Ala)-PORA; ■ , (Cys-43Ala)-PORA.
Buhr et al. PNAS � August 26, 2008 � vol. 105 � no. 34 � 12633
PLA
NT
BIO
LOG
Y

Production of (CysX3Ala)PORA Mutant Proteins. DNAs encoding the different(Cys3Ala)-PORA proteins lacking their transit peptides for plastid import weregenerated with clone A7 (12) as template and the Promega GeneEditor in vitrosite-directed mutagenesis kit. The following primer combinations were used:Oligo-S plus M1, Oligo-S plus M2, Oligo-S plus M3, and Oligo-S plus M4. Theresulting DNAs were subcloned into pQE30 vector (Qiagen) and transformed intoE. coli strainXL1-Blue.Detailsonproteinexpressionandpurificationareprovidedin SI Text. Alternatively, 35S methionine-labeled POR proteins were produced bycoupled transcription/translation of respective clones in wheat germ lysates (17).
Assembly Assay of LHPP. Equimolar amounts of the different reconstituted(CysX3Ala)-PORA-Pchlide b-NADPH and PORB-Pchlide a-NADPH ternary com-plexes were mixed and incubated in the dark for 15 min. One aliquot of thereaction mixtures was immediately precipitated with trichloroacetic acid andused for radioactivity measurements done with a liquid scintillation counter.Another aliquot was subjected to gel filtration on Superose 6 (Amersham Phar-maciaBiotech)equilibrated inassaybuffercontaining350mMsucrose. Individualfractions were harvested, and aliquots were withdrawn for radioactivity mea-surements. Alternatively, nonradiolabeled proteins were used and detected byWestern blotting, using POR-specific antiserum (33). In either case, appropriatefractions were pooled and protein-precipitated with trichloroacetic acid, pro-cessed with acetone, ethanol, and diethyl ether, and resolved on 10–20% poly-acrylamide gradients containing SDS (38). After electrophoresis, 35S-PORA:PORBhigher molecular weight complexes and nonassembled 35S-PORA, 35S-PORB, and35S-PORA derivatives were visualized by autoradiography or, in experimentswhere nonradiolabeled proteins had been used, by Western blotting.
Singlet Oxygen Measurements. 1O2 production was measured by using either theelectrophoretically resolved PORA:PORB:lipid complexes or nonfractionatedcomplexes and the DanePy method developed by Kalei et al. (34, 35). Fluores-cence emission spectroscopy was carried out at an excitation wavelength of 330nm and collecting fluorescence emission between 515 and 550 nm and above 650nm in a Life Sciences spectrometer LS50 (PerkinElmer).
Pigment Measurements. HPLC was performed on either C18 reverse-phase (RP)silica gel columns (Macherey-Nagel; 250 � 4.6 mm, Nucleosil ODS 5 �m) or C30 RPcolumns (YMC; 250 � 4.6 mm, 5 �m), using established procedures and a VarianProStar model 410 apparatus, ProStar model 240 pump, and ProStar 330 photo-diode array detector (16, 17). In some experiments, Chlides a and b were sepa-rated on RP18 Gromsil columns (Grom), using a gradient of 100% acetone,applied for 3 min, and 60% acetone/40% sodium acetate-supplemented water,pH 6.5, reached within 20 min (39). Low-temperature luminescence spectroscopywas performed at 77 K at excitation wavelengths of either 440 or 470 nm in theLS50 spectrometer (see above) (32).
ACKNOWLEDGMENTS. We thank Dr. E. Hideg (Institute of Plant Biology,Biological Research Center, Hungarian Academy of Sciences, Szeged, Hun-gary) for gift of the DanePy reagent and Dr. G. Tichtinsky for technicalassistance with the bacterial expression and purification of the barley PORAand PORB. This study was supported by the Chaire d’Excellence program of theFrench Ministry of Research and Deutsche Forschungsgemeinschaft ResearchProject Grants DFG-RE1465/1-1, DFG-RE1465/1--2, and DFG-RE1465/1-3(to C.R.).
1. Armstrong GA, Runge S, Frick G, Sperling U, Apel K (1995) Identification of NADPH:proto-chlorophyllideoxidoreductasesAandB:Abranchedpathwayforlight-dependentchlorophyllbiosynthesis in Arabidopsis thaliana. Plant Physiol 108:1505–1517.
2. Oosawa N, et al. (2000) Identification and light-induced expression of a novel gene ofNADPH-protochlorophyllide oxidoreductase isoform in Arabidopsis thaliana. FEBSLett 474:133–136.
3. Masuda T, et al. (2002) Identification of two differentially regulated isoforms ofprotochlorophyllide oxidoreductase (POR) from tobacco revealed a wide variety oflight- and development-dependent regulations of POR gene expression among an-giosperms. Photosynth Res 74:165–172.
4. Iwamoto K, Fukuda H, Sugiyama M (2001) Elimination of POR expression correlateswith red leaf formation in Amaranthus tricolor. Plant J 27:275–284.
5. Forreiter C, Apel K (1993) Light-independent and light-dependent protochlorophyl-lide-reducing activities and two distinct NADPH-protochlorophyllide oxidoreductasepoly-peptides in mountain pine (Pinus mugo). Planta 190:536–545.
6. Skinner JS, Timko MP (1998) Loblolly pine (Pinus taeda L.) contains multiple-expressedgenes encoding light-dependent NADPH:protochlorophyllide oxidoreductase (POR).Plant Cell Physiol 39:795–806.
7. Spano AJ, He Z, Timko MP (1992) NADPH: protochlorophyllide oxidoreductases inwhite pine (Pinus strobus) and loblolly pine (P. taeda). Mol Gen Genet 236:86–95.
8. Reinbothe S, Runge S, Reinbothe C, van Cleve B, Apel K (1995) Substrate-dependenttransport of the NADPH:protochlorophyllide oxidoreductase into isolated plastids.Plant Cell 7:161–172.
9. Reinbothe S, Reinbothe C, Runge S, Apel K (1995) Enzymatic product formation impairsboth the chloroplast receptor binding function as well as translocation competence ofthe NADPH:protochlorophyllide oxidoreductase, a nuclear-encoded plastid protein.J Cell Biol 129:299–308.
10. Reinbothe S, Reinbothe C, Holtorf H, Apel K (1995) Two NADPH:proto-chlorophyllide oxidoreductases in barley: Evidence for the selective disappearanceof PORA during the light-induced greening of etiolated seedlings. Plant Cell7:1933–1940.
11. Reinbothe S, Reinbothe C, Lebedev N, Apel K (1996) PORA and PORB, two light-dependent protochlorophyllide-reducing enzymes of angiosperm chlorophyll biosyn-thesis. Plant Cell 8:763–769.
12. Holtorf H, Reinbothe S, Reinbothe C, Bereza B, Apel K (1995) Two routes of chloro-phyllide synthesis that are differentially regulated by light in barley. Proc Natl Acad SciUSA 92:3254–3258.
13. Su Q, Frick G, Armstrong G, Apel K (2001) PORC of Arabidopsis thaliana: A third light-and NADPH-dependent protochlorophyllide oxidoreductase that is differentially reg-ulated by light. Plant Mol Biol 47:805–813.
14. Pattanayak GK, Tripathy BC (2002) Catalytic function of a novel protein protochloro-phyllide oxidoreductase C of Arabidopsis thaliana. Biochem Biophys Res Commun291:921–924.
15. Reinbothe C, Lebedev N, Reinbothe S (1999) A protochlorophyllide light-harvestingcomplex involved in de-etiolation of higher plants. Nature 397:80–84.
16. Reinbothe S, Pollmann S, Reinbothe C (2003) In situ conversion of protochlorophyllideb to protochlorophyllide a in barley: Evidence for a novel role of 7-formyl reductase inthe prolamellar body of etioplasts. J Biol Chem 278:800–806.
17. Reinbothe C, Buhr F, Pollmann S, Reinbothe S (2003) In vitro reconstitution of LHPPwith protochlorophyllides a and b. J Biol Chem 278:807–815.
18. Reinbothe C, Lepinat A, Deckers M, Beck E, Reinbothe S (2003) The extra loop distin-guishing POR from the structurally related short-chain alcohol dehydrogenases isdispensable for pigment binding, but needed for the assembly of LHPP. J Biol Chem278:816–822.
19. Lebedev N, Timko MP (1998) Protochlorophyllide photoreduction. Photosynth Res58:5–23.
20. Baker ME (1994) Protochlorophyllide oxidoreductase is homologous to human car-bonyl reductase and pig 20-�-hydroxysteroid dehydrogenase. Biochem J 300:605–607.
21. Dahlin C, et al. (1999) The role of protein surface charge in catalytic activity andchloroplast membrane association of the pea NADPH:protochlorophyllide oxi-doreductase (POR) as revealed by alanine scanning mutagenesis. Plant Mol Biol39:309 –323.
22. Townley HE, Sessions RB, Clarke AR, Dafforn TR, Griffith TW (2001) Protochlorophyllideoxidoreductase: A homology model examined by site-directed mutagenesis. ProteinsStruct Funct Genet 44:329–335.
23. Ghosh D, et al. (1991) Three-dimensional structure of holo 3-�-, 20-�-hydroxysteroiddehydrogenase: A member of a short-chain dehydrogenase family. Proc Natl Acad SciUSA 88:10064–10068.
24. Tsigelny I, Baker ME (1995) Structures important in mammalian �- and 17-�-hydroxysteroid dehydrogenases. J Steroid Biochem Mol Biol 55:445–621.
25. Varughese KL, Skinner MM, Whitley JM, Matthews DA, Xuong NH (1992) Crystalstructure of rat liver dihydropteridine reductase. Proc Natl Acad Sci USA 89:6080–6084.
26. Wilks H, Timko MP (1995) A light-dependent complementation system for analysis ofNADPH:protochlorophyllide oxidoreductase: Identification and mutagenesis of twoconserved residues that are essential for enzyme activity. Proc Natl Acad Sci USA92:724–728.
27. Lebedev N, Timko MP (1999) Protochlorophyllide oxidoreductase B-catalyzed proto-chlorophyllide photoreduction in vitro: Insight into the mechanism of chlorophyllformation in light-adapted plants. Proc Natl Acad Sci USA 96:9954–9959.
28. Lebedev N, Karginova O, McIvor W, Timko MP (2001) Tyr-275 and Lys-279 stabilizeNADPH within the catalytic site of NADPH:protochlorophyllide oxidoreductase and areinvolved in the formation of the enzyme photoactive state. Biochemistry 40:12562–12574.
29. Oliver PR, Griffiths TW (1981) Covalent labeling of the NADPH:protochlorophyllide oxi-doreductase from etioplast membranes with [3H]N-phenylmaleimide. Biochem J 195:93–101.
30. Reinbothe C, et al. (2006) In vitro mutagenesis of PORB: Two distinct protochlorophyl-lide binding sites participate in enzyme catalysis and assembly. Mol Gen Genomics275:540–552.
31. Ryberg M, Sandelius AS, Selstam E (1983) Lipid composition of prolamellar bodies andprothylakoids of wheat etioplasts. Physiol Plant 57:555–560.
32. Lebedev N, van Cleve B, Armstrong G, Apel K (1995) Chlorophyll synthesis in ade-etiolated (det340) mutant of Arabidopsis without NADPH-protochlorophyllide(PChlide) oxidoreductase (POR) A and photoactive PChlide-F655. Plant Cell 7:2081–2090.
33. Schulz R, et al. (1989) Nucleotide sequence of a cDNA coding for the NADPH-protochlorophyllide oxidoreductase (PCR) of barley (Hordeum vulgare L.) and expres-sion in Escherichia coli. Mol Gen Genet 217:355–361.
34. Kalai T, Hankovszky O, Hideg E, Jeko J, Hideg K (2002) Synthesis and structureoptimization of double (fluorescent and spin) sensor molecules. ARKIVOC iii:112–120.
35. Kalai T, Hideg E, Vass I, Hideg K (1998) Double (fluorescent and spin) sensor fordetection of reactive oxygen species in the thylakoid membranes. Free Radical BiolMed 24:649–652.
36. Reinbothe C, et al. (2004) LHPP, the light-harvesting NADPH:protochlorophyllide(Pchlide) oxidoreductase:Pchlide complex of etiolated plants, is developmentally ex-pressed across the barley leaf gradient. Plant Sci 167:1027–1041.
37. Park H, Kreunen SS, Cuttriss AJ, DellaPenna D, Pogson BJ (2002) Identification of thecarotenoid isomerase provides insight into carotenoid biosynthesis, prolamellar bodyformation, and photomorphogenesis. Plant Cell 14:289–292.
38. Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature 227:680–685.
39. Oster U, Tanaka R, Tanaka A, Rudiger W (2000) Cloning and functional expression ofthe gene encoding the key enzyme for chlorophyll b biosynthesis (CAO) from Arabi-dopsis thaliana. Plant J 21:305–310.
12634 � www.pnas.org�cgi�doi�10.1073�pnas.0803950105 Buhr et al.




















