
Optical Detection Methods for High-Throughput Fluorescent ...
Upload
independentCategory
view
1download
0
System. Appl. Microbiol. 20,137-149 (1997) © Gustav Fischer Verlag
Phenotypically Based Taxonomy of Fluorescent Pseudomonas Strains Isolated from Four Natural Mineral Waters
S. VERHILLE, M. ELOMARI, L. COROLER, D. IZARD, and H. LECLERC
Service de bacteriologie-hygiene, Faculte de Medecine, Lille, Cedex, France
Received August 1, 1996
Summary
A numerical analysis based on 231 phenotypic characteristics was performed with 100 isolates from 4 natural mineral waters so as with forty-nine type or collection strains belonging or related to fluorescent Pseudomonas strains. Since this study has been the continuation of a previously published work (ELOMARI et aI., 1995) seven strains from the two former phenotypic groups Ia or IIb were included among the type or collection strains which have been used before in order to make easier the link with the two studies. The data were processed by the Dice similarity coefficient and unweighted pair group algorithm with arithmetic averages as well. Eight clusters (IX to XV, III) were defined at the 58% similarity level. Clusters XII, XIII, XIV and XV were further divided into 18 subclusters. Type and collection strains clustered into eight groups which correlated closely with those generated by the previous numerical analysis (la, Ie, IIb, III, IVa, IVb IVc, VII). The three phenotypically characterized groups of mineral water strains Ib, IIa and V were also defined again. Furthermore most of the isolates of the present study, fell into four new groups which did not include any type or collection strains: IX (10 strains), XIIIb (12 strains), XIIIc (9 strains), XVa (9 strains). Later on DNA/DNA hybridization studies would be necessary to specify the taxonomic status of these four groups within the genus Pseudomonas.
Key words: Taxonomy - Phenotypic analysis - Pseudomonas - Water
Introduction
Their great metabolic versatility and pathogenic properties make Pseudomonas spp. strains increasingly important for more and more fields. This large and heterogeneous group of Gram-negative bacteria are common inhabitants of most natural environments. The phenetic study of STANIER et al. (1966) provided a modern framework for the phenotypical study of Pseudomonas and became the basis of numerical analysis (SNEATH et aI., 1981) and genotypic studies (SANDS et aI., 1970; PALLERONI et aI., 1972 and 1973; CHAMPION et aI., 1980), which have led to the division of the genus into five rRNA homology groups (PALLERONI, 1984). However those groups have been reclassified. The genus Pseudomonas "sensu stricto" has been restricted to rRNA group I (DE Vos and DE LEY, 1983). In the last years, few species have been added within this rRNA group I: P. luteola and P. oryzihabitans (KODAMA et ai., 1985) which were newly reclassified respectively in the genus Chryseomonas (C. luteola) and in the new genus Flavimonas (F. oryzihabitans; HOLMES, B., 1987), P. lundensis (MOLIN et aI., 1986a), P. fragi (MOLIN and TERNSTROM,
1982 and 1986b), P. savastanoi (GARDAN et aI., 1992) and P. flavescens (HILDEBRAND et aI., 1994). Whereas recent works have clarified the taxonomic status of the other 4 rRNA groups and have led to the creation of new genera such as Burkholderia (YABUUCHI et aI., 1992), Comamonas (DE VOS et aI., 1985; TAMAoKA et aI., 1987), Acidovorax (WILLEMS et aI., 1990), Hydrogenophaga (WILLEMS et aI., 1989); Brevundimonas (SEGERS et aI., 1994), Strenotrophomonas (PALLERONI and BRADBURY, 1993), Sphingomonas (YABUUCHI et aI., 1990), and Telluria (BOWMAN et aI., 1993). Most of the works performed on these groups have pointed out the genomic heterogeneity of the genus (DE Vos and DE LEY, 1983; PALLERONI, 1981; WOESE et ai., 1984).
Pseudomonas strains are widespread in nature and particularly abundant in soil and water. However, few studies have been carried out on bacterial flora which colonizes mineral waters where the predominant occurrence of fluorescent Pseudomonas strains has already been shown (LECLERC, 1990; GUILLOT and LECLERC, 1993; AZNAR et aI., 1992).
138 S. VERHILLE et al.
Thus, specific taxonomic studies are needed to learn more about those strains, and consequently the present work deals with a numerical phenotypic analysis on a sample of fluorescent Pseudomonas strains twice as large as the one of the previous work (ELOMARI et ai., 1995), isolated from 4 natural mineral waters. The main purpose of this study is not only to try to enlarge the three former clusters lb, lIb and V (ELOMARI et ai., 19955) but also to isolate other new phenotypic groups of mineral water strains which would not include a type or collection strain.
Materials and Methods Bacterial strains: This phenotypic study relied on 149 strains
which consisted of 100 wild strains, isolated from 4 French natural mineral waters designed A, B, C, D and identified as fluorescent pseudomonads according to the criteria of PALLERONI (1984). Forty-nine type, collection and wild strains representative of the fluorescent pseudomonads were also added in for control purposes (P. aeruginosa, P. fluorescens biovars I to V, P. chlororaphis, P. aureofaciens, P. marginalis, P. putida biovars A and B, P. lundensis, P. syringae, P. cichorii, P. tolaasii and P. asplenii). Further details about the strains, including reference numbers and origins (when known), are given in Table 1.
Table 1. List of organisms studied and their classification in phenotypic analysis.
Cluster
IX (N = 10)
X(N=3)
XI (N = 5)
XII (N = 38)
Subcluster Culture collection or other reference N°
XIIa (n =2)
IIa n=4
Ib n=3
XIInc n=l
Ie n = 13
CFML 95-322 CFML 95-309 CFML 95-305 CFML 95-304 CFML 95-303 CFML 95-306, -311 CFML 95-307, -312 CFML 95-308
CFML 95-280 CFML 95-281 CFML 95-285
CFML 95-340 CFML 95-319, -325 CFML 95-329 CFML 95-270
ATCC 17482 CFML 95-287
CIP 104664T
CFML 92-113 CFML 92-102 CFML 92-104
CFML 92-123 CFML 92-133 CIP 104 663T
DSM50106 (=ATCC 17826)
ATCC 15916 ATCC 17563
ATCC 13525T
NCPPB 1616 (= PDDCC 2839) ATCC 17518 NCPPB 2192T
(= ATCC 33618T = PDDCC 2290T )
CIP 73-25 (= ATCC 17397) CFML 95-247 CFML 95-299, -301 CFML 95-297 CFML 95-298, -300
Name as received
P. fluorescens biovar II
P. rhodesiae(1)
P. veronii(ll
P. fluorescens biovar II
P. fluorescens biovar V P. fluorescens biovar I
P. fluorescens biovar I P. tolaasii P. fluorescens biovar V P. tolaasii
P. fluorescens biovar I
Isolated from"
B(l) 3 A(2) 2 A(2) 3 A(2) 5 A(2) 6 A(3) 3 A(3) 6 A(4)2
D(l) 1 D(l) 2 D(l) 3
B(l) 2 B(l) 4 B(l) 6 D(l) 1
Naphthalene-enriched medium D(3)2
A(3) 3 A(4) 3 A(3) 1 A(4) 1
B(3) 1 D(l) 2 D(l) 1
Sea water
Sea water Sputum Agaricus bisporus pre-filter tanks Cultivated mushroom Soil Cultivated mushroom Agaricus bisporus Tap water C(3) 4 D(l) 3 D(4)2 D(4) 3
Phenotypically Based Taxonomy of Fluorescent Pseudomonas Strains 139
Table 1. (Continued).
Cluster
XIII (N =34)
XIV (N = 11)
VII (N =3)
Subcluster Culture collection or other reference N°
XIIe n=7
Ia n=8
XIIIa n=4
XIIIb n = 12
XIIId n=2
XIIIc n=9
lIb n=3
IVb n=3
XIIInc n=l
IVc n=6
IVa n=2
CFML 95-330 CFML 95-275, -334 to -336 CFML 95-337, -339
ATCC 14150 DSM 50276 NCPPB 667T (= ATCC 10844T) ATCC 17815 ATCC 17816 DSM 50275 CFML 95-286 CFML 95-277
CFML 95-313 CFML 95-312 CFML 95-310 CFML 95-302
CFML 95-341 CFML 95-323, -324, -326, -327 CFML 95-316, -31 7, -318 CFML 95-320 CFML 95-321, -333 CFML 95-263
CFML 95-282 CFML 95-271
CFML 95-315 CFML95-260 CFML 95-246 CFML 95-262 CFML 95.259, -261 CFML 95-243 CFML 95-249, -257
DSM 50148 (= ATCC 17533) ATCC 12983 ATCC 17573
CCUG 1317 ATCC 17484 LMG 1246 (= ATCC 17430 = NCIB 10534)
DSM 50415
ATCC 17415 CCEB 518T (= ATCC 13985T) ATCC 9447 DSM 50083T (= ATCC 9446T) ATCC1 7414 ATCC 17386
DSM 50208 (= ATCC 17485) CIP 51-191T (= ATCC 12633T)
CIP 68-52T(= ATCC10145T) ATCC 15692 CIP 76-110 (= ATCC 27853)
Name as received
P. fluorescens biovar V P. marginaiis P. marginalis P. fluores cens biovar II P. fluorescens biovar II P. marginalis
P. fluorescens biovar V P. fluores cens biovar IV P. fluorescens biovar V
P. putida biovar B P. putida biovar B P. putida biovar B
P. fluorescens biovar IV
P. aureofaciens P. aureofaciens P. chlororaphis P. chlororaphis P. chlororaphis P. fluorescens biovar V
P. putida biovar A P. putida
P. aeruginosa P. aeruginosa P. aeruginosa
Isolated from':-
B(4) 3 B(4) 4 B(4) 6
? Soft rot of potato tuber Endive Lettuce Dahlia Plant D(2) 3 D(l) 1
A(3) 4 A(3) 6 A(4) 3 A(4) 5
B(l) 2 B(l ) 3 B(l) 4 B(l) 5 B(l) 6 C(4) 3
D(l ) 1 D(l ) 2
A(2) 1 C(2) 3 C(2) 4 C(2) 5 C(2) 6 C(4)4 C(4) 6
Soil Soil Polluted sea water
Soil Naphthalene-enriched medium ?
Soil Mass river clay in kerosene ? Plate contaminant ? Tryptophan-enriched water
Soil Soil
? infectd wound Blood culture

140 S. VERHILLE et al.
Table 1. (Continued).
Cluster Subcluster Culture collection or other reference N°
Name as received
Isolated from*
XV (N = 41)
III (N = 2)
NC(N=5)
XVa n=9
V n = 26
XVc n=4
XVnc n=2
CFML 95-258 CFML 95-244 CFML 95-250, -251 CFML 95-256, -264 CFML 95-255 CFML 95-245, -254
ATCC 17559 ATCC 17400 CFML 95-252 CFML 95-248, -253 CFML 95-296 CFML 95-274, -276, -279 CFML 95-265 to -268, -273 CFML 95-283, -293 CFML 95-292 CFML 95-278 CFML 95-269, -272 CFML 95-288, -289 CFML 95-290 CFML 95-291 CFML 95-294, -295
CFML 95-331 CFML 95-332, -338, -342
CFML 95-328 ATCC 17571
CCUG 18757T (= CCM 573T )
CCUG 18758
CFML 90-57 ATCC 19310T
ATCC 23835T
DSM 50259T
CFML 95-284
P. fluorescens biovar III P. fluorescens biovar III
P. fluorescens biovar III
P. lundensis P. lundensis
P. putida biovar A P. syringae P. asplenii P. cichorii
C(3) 3 C(3) 4 C(3) 6 C(4) 2 C(4) 3 C(4) 4
Plant Egg C(1) 2 C(1) 4 C(3) 4 D(I) 1 D(I) 2 D(I) 4 D(I) 6 D(2) 1 D(2) 2 D(3)2 D(3) 3 D(3) 6 D(4)4
B(4)4 B(4) 6
B(4) 6 Polluted sea water
Beef meat Beef meat
placenta Plant Plant Endive B(1) 5
* All Strains were isolated from four French natural mineral waters, called A, B, C and D (Evian, Cachat spring; Volvic, Clairvic spring; Vittel, Grande source; Contrexeville, Pavilion spring) at the following sites: at the point of emergence (1), in pipeline (2), before bottling (3) and after bottling (4). For each brand, samples were examined once in a year at different times 1,2,3,4,5 and 6. (I) in press ATCC CCEB
CCM CCUG CIP CFML DSM NCIB NCPPB
PDDCC LMG nc NC
American Type Culture Collection, Rockville, Maryland, USA. Culture Collection of Entomogenous Bacteria, Departement of Insect Pathology, Institute of Entomology, Prague, Czechoslovakia. Czechoslovak Collection of Microorganism, J. E. Purkyne, University, Brno, Czechoslovakia. Culture Collection, University of Goteborg, Goteborg, Sweden. Collection de l'institut Pasteur, Paris, France. Collection de la faculte de Medecine, Lille, France. Deutsche Sammlung von Mikroorganismen, Gottingen, Federal Republic of Germany. National Collection of Industrial Bacteria, Aberdeen, Scotland, UK. National Collection of Plant Phothgenic Bacteria, Ministry of Agriculture, Fisheries and Food, Plant Pathology Laboratory, Harpeden, Hertfordshire, UK. Plant Diseases Division Culture Collection, Auckland, New Zealand. Laboratoium voor Microbiologie en microbiele Genetica, Rijksuniversiteit Gent, Belgium Unclustered strains of the subcluster considered. Strains not belonging to any cluster.
Phenotypically Based Taxonomy of Fluorescent Pseudomonas Strains 141
Phenotypic characterization: Forty-three conventional tests were carried out as described by GAVINI et aI., 1989. The ability of each strain to assimilate a range of sole carbon source was assessed by auxanographic biotype 100 system (BioMerieux, Marcy-L'Etoile, France) containing a total of 99 carbohydrates, amino-acids and organic acids. Strips were incubated for 5 days at 30 0c. Eighty-nine enzymatic tests were performed on experimental enzymatic strips. The enzymatic
Table 2. Phenotypical characteristics of clusters.
Clusters
IX X n = 10 n=3
Phenotypic tests Conventional tests Denitrification 50" 100 Nitrate reduction 100 100 Growth on NaCI (3%) 50 a Growth on TTC':' " a a Tributyrin-esterase 80 67 Tween-esterase 70 67 Lecithinase a a Malonate 90 33 TTR':'** 30 a Carbon sources D(+) Galactose 100 100 a-D(+) Melibiose 100 a Sucrose 100 100 D(+) Raffinose 100 0 Maltotriose 100 0 Maltose 100 100 a-Lactose 100 a Lactulose 100 0 1-0Methyl-f3-galactopyranoside 100 0 1-0Methyl-a-galactopyranoside 100 a D(+) Cellobiose 100 33 f3-Gentiobiose 100 0 1-0-Methyl-f3-D-Glucopyranoside 90 0 Esculin 100 0 D(-) Ribose 100 100 L(+) Arabinose 100 100 D(+) Xylose 100 a Palatinose 100 0 a-L-Rhamnose 100 a a-L(-) Fucose 100 a D(+) Arabitol 100 100 L(-) Arabitol 100 a Xylitol 100 a D-Tagatose 90 a D-Mannitol 100 100 Maltitol 100 a D(+) Turanose 100 100 D-Sorbitol 100 67 Adonitol 100 a D-Lyxose 90 a i-Erythritol a a 3-0Methyl D Glucopyranose 100 a D-Saccharate 100 33 Mucate 100 100 L(+) Tartrate 90 a Meso-tartrate 100 0
activities tested included 59 peptidases, 10 esterases, and 20 osidases (GAVINI et aI., 1991). They were also studied with the API systems which were incubated at 30 °C for 4 h. The tests were scored as positive according to the manufacturer's instructions. In all, 231 characters were investigated. Sixty-one of these which were either positive or negative for all the strains, were not included in the numerical analysis, all of them are listed in Table 2. Other data were codified and sub-
XI XII XIII XIV XV III n=5 n = 38 n = 34 n = 11 n = 41 n=2
100 37 41 27 95 a 100 74 74 45 95 a
80 95 32 100 93 100 a 11 a 100 2 100
60 50 74 91 68 a a 26 18 73 95 a a 42 6 55 90 a
100 79 65 100 93 a 80 24 3 55 95 50
100 100 100 36 83 a a a a a a a
20 68 15 45 0 0 0 a 0 0 0 0 0 3 0 0 a 0 0 3 0 a 0 0 0 0 0 0 0 a 0 0 0 0 0 0 a 0 a a 0 0 0 0 a 0 0 0 0 0 0 0 0 0 0 a 0 0 0 0 0 0 0 0 0 0 a 18 0 0 0 0
100 100 100 91 100 a 100 92 100 18 a 100
80 87 88 a a a 0 0 a a a a a 24 3 a 66 a a a a a a 100
100 100 97 82 100 a 0 61 3 a 100 a a 61 0 a 100 a a a a a 0 a
100 100 97 82 100 a a a a a a a a a a a a a
80 100 41 a 5 a 20 61 3 a 100 a 20 68 3 a a a 20 87 3 9 100 a
0 a a 0 a 0 100 97 100 64 a a 100 100 100 64 66 a
a a 9 45 a a a 45 76 36 a a
142 S. VERHILLE et al.
Table 2. (Continud).
Clusters
IX X XI XII XIII XIV XV III n == 10 n==3 n==5 n == 38 n == 34 n == 11 n == 41 n==2
Tricarballylate 90 0 80 32 44 9 0 0 D-Glucuronate 100 0 100 97 88 18 0 0 D-Galacturonate 100 0 100 100 85 18 0 0 N-acetyl-D-Glucosamine 100 0 100 100 76 82 100 0 Phenyl acetate 100 0 0 3 53 45 0 0 Protocatechuate 100 100 100 100 100 100 100 0 p-Hydroxy benzoa te 100 100 100 100 100 100 90 0 (-)Quinate 100 100 100 100 100 100 100 0 Gentisate 100 0 0 0 0 36 29 0 m-Hydroxybenzoate 100 0 0 0 0 9 24 0 Benzoate 100 100 20 8 88 100 90 50 Histamine 100 0 0 5 82 100 0 0 D-Glucosamine 100 33 100 87 97 55 100 0 Itaconate 100 0 80 84 3 100 95 0 DL-~-Hydroxybutyrate 100 100 100 100 100 100 100 0 Malonate 100 33 100 100 100 100 100 0
Enzymatic tests a-D-Glucoside 0 100 0 3 0 0 0 0 N-acetyl-~-D-glucosaminidase 100 100 60 37 44 18 37 0 esterases-C 12 10 67 0 53 29 100 90 50 L-tyrosine arylamidase 60 0 20 45 53 64 22 100 L-pyrrolidone arylamidase 20 0 0 68 9 45 100 100 L-histidine arylamidase 100 100 0 79 100 100 63 100 L-aspartate arylamidase 60 3 0 74 53 91 80 100 y-glutamyl transferase 60 0 40 84 50 100 51 100 a-L-glutamate arylamidase 40 0 0 55 24 55 68 100 L-threonine aryl amidase 70 33 0 63 62 55 85 100 L-alanyl-L-phenylalanyl L- 50 0 20 74 76 91 78 100 prolyl-L-alanine arylamidase a-L-aspartyl-L-arginine 90 0 0 74 68 100 100 100 arylamidase a-L-glutamyl-L-histidine 40 0 0 53 47 73 59 100 arylamidase glycyl-L-arginine arylamidase 90 67 0 76 76 100 100 100 L-histidyl-L-serine arylamidase 20 33 0 79 12 36 87 100 L-phenylalanyl-L-proline 10 0 40 58 35 82 56 100 arylamidase N-acetyl-glycyl-L-Iysine 0 0 0 50 21 91 32 50 arylamidase L-valyl-L-tyrosyl-L-serine 50 33 0 61 38 55 90 50 arylamidase
" % of positive strains ",:. TTC: 2,3,5-Triphenyltetrazolium chlorid. ,: .• ' ,. TTR: Tetrationate reductase. All the strains in the study were King B, oxydase (except P. syringae ATCC 19310T ) and catalase positive, grew on 0% and 0.8% NaCI, hydrolysed L-tyrosine, were citrate and arginine positive and were motile. All strains were also able to utilize the following compounds as sole carbon source: glycerol, cis-aconitate, citrate, D-glucuronate, betain, D-L-a amino-n-butyrate, D L-Iactate, L-as-partate, L-glutamate, L-proline, L-alanine, proprionate. Furthermore all strains showed the following enzymatic activities: esterase-C6 and -C8, L-alanine arylamidase, methionine arylamidase, L-proline-4-methoxy arylamidase, ~-alanine arylamidase, a--L-as-partyl-L-alanine arylamidase.
Each of the strains were negative for the following tests: growth in the presence of a concentration of sodium chloride of 5% and 7% wt/vol, showed none DNase activity were not able to hydrolyse starch, did not possess any ornithine decarboxylase, lysine de-carboxylase and phenylalanine desaminase activities. All the strains were indol, acetoine, coagulase, elastase and fibrinolyse nega-tive. The strains were unable to utilize L(+) sorbose, dulcitol, hydroxyquinoline-~-glucuronide, L-tryptophan, L-histidine. None of the strains possessed the following enzyme activities: a-D-galactosidase, ~-D-galactosidase, phospho-~-D-galactosidase, a-L-arabi-nosidase, ~-D-galacturonohydrolase, ~-D-glucuronidase, a-maltosidase, ~-maltosidase, N-acetyl-a-D-glucosaminidase, ~-D-fucosi-dase, ~-L-fucosidase, ~-D-Iactosidase, a-D-mannosidase, ~-D-mannosidase, ~-D-xylosidase, a-D-xylosidase.
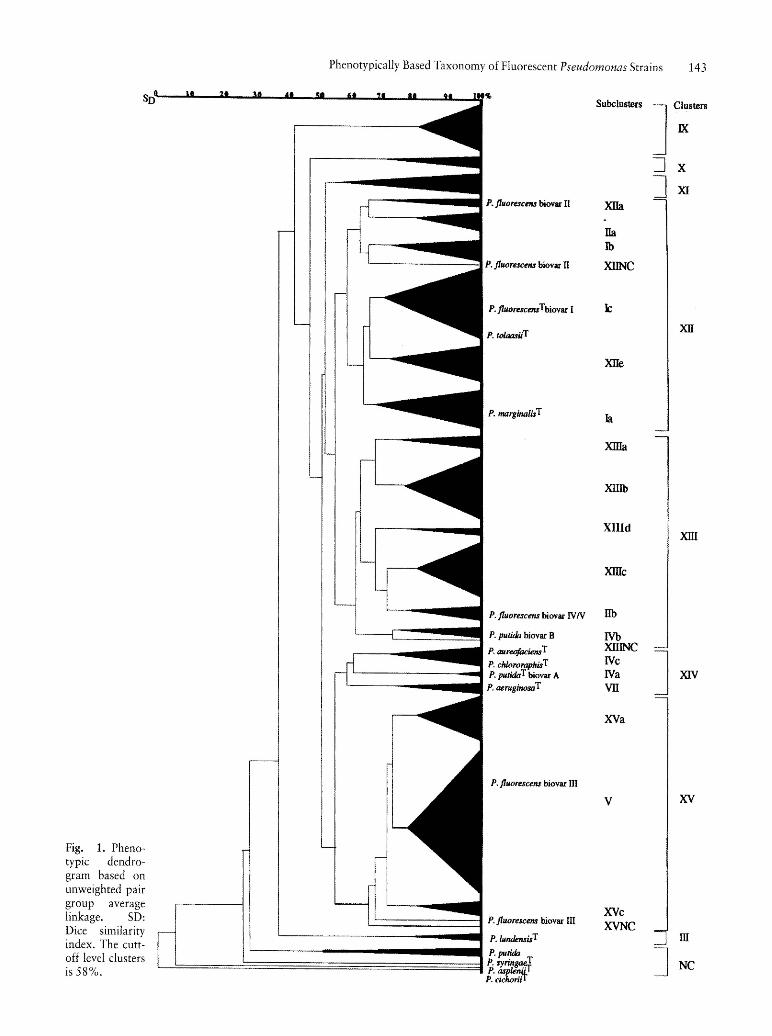
Fig. 1. Phenotypic dendrogram based on unweighted pair group average linkage. SD: Dice similarity index. The cuttoff level clusters is 58%.
Phenotypically Based Taxonomy of Fluorescent Pseudomonas Strains 143
Subclusters
r-------111111 P. jblore.rcms biovar II xna
lla Ib
'--------4P. jblorucms biovar II XIINC
P. jblore.rcms Tbiovar I Ie
P. margina/isT
P. fluore.rcens biovar IV N
P. putida biovar B
P. aureofaciens T
P. chJoro1J1Phis T P. putida T biovar A P. Ql:ruginasa T
P. fluore.rcens biovar III
L'::========I P. fluorescms biovar 1II
P. lundensis T L ____________ .. P.putida
L-1========~============================~~ryrin~ P. asp/enii. P. ciclwriiT
XIJe
Ia
XIlla
XIllb
XIIId
XIIIc
lIb
IVb XlIINC Nc Na vn
XVa
v
XVc XVNC
] ~ ]
]
Clusters
IX
x XI
XII
XIll
XN
xv
ill
] NC

144 S. VERHILLE et al.
jected to numerical analysis as described by GAVINI et al. (1989) .
Test reproducibility: All the tests were repeated on 10% of strains. The average probability of error of 2.5%, as calculated by the formula of SNEATH and JOHNSON (1972), would not produce serious distortion of taxonomic structure.
Results and discussion
The phenotypic dendrogram obtained is shown figure 1. At 58% of similarity level, the numerical analysis delineated 8 main clusters (IX to XV and III to refer to Figure 1 and Table 1). Clusters XII, XIII, XIV and XV could be subdivided at higher levels of similarity, into 18 subclusters: respectively XIIa, IIa, Ib, Ie, XIIe, Ia; XIIIa to XIIId, lIb, IVb; IVc, IVa, VII; XVa, V, XVc. However 4 strains remained unclustered: P. putida biovar A CFML 90-57, P. syringae ATCC 19310\ P. asplenii ATCC 23835T, P. eiehorii DSM 50259T and the wild strain CFML 95-284 isolated from mineral water. The accurate composition of each cluster and subcluster are reported in Table 1. Phenotypic characteristics which proved useful in distinguishing between clusters and subclusters are listed respectively in Table 2 and 3. The subcluster XIIId which only included two wild strains will be neglected in this study, and consequently omitted in table 3. As often as a group could be classed as one of those of the previous study (ELOMARI et aI., 1995) the former name has been kept. Without considering strains that were not found again only because they were not included in this study, it appeared that seven groups (IIa, Ib, III, IVa, IVb, IVc, VII) were closely similar to a former one. As to subclusters la, Ie, lIb, and V minor differences could be noticed: P. fluoreseens biovar II ATCC 17816 which was unclassified before has been included in the subcluster la, P. fluroeseens biovar II ATCC 17816 which was unclassified before has been included in the subcluster la, P. tolaasii NCPPB 1616 has left the subcluster Id for the subcluster Ie, P. fluoreseens biovar III ATCC 17571 has been excluded from cluster V to remain unclassified in this study. All those differences may be attributed to the unclear taxonomic status of the considered strains. At last P. fluoreseens biovar IV DSM 50415 from the subcluster lIb remained now unclassified. As to the taxonomic status, the following groups could be identified because of being closely related to one type strain and their phenotypic characteristics were in good agreement with published data (STANIER et aI., 1966; CHAMPION et aI., 1980; PALLERONI, 1984; BARRETT et aI., 1986): subcluster Ia (n = 8) including P. marginalis ATCC 10844T gathered mostly collection strains from plant origin; subcluster IVa (n = 2) contained the type strain P. putida ATCC 12633T so as one collection strain from the same biovar; subcluster VII (n = 3) comprised the type strain P. aeruginosa ATCC 10145T associated with two P. aeruginosa strains from clinical origin; cluster III (n = 2) contained the type strain P. lundensis CCM 573T so as the collection strain P. lundensis CCUG 18758, both isolated from beef meat. The two following subclusters gathered two type strains: the subcluster IVc (P. aureofaeiensT and P.
ehlororaphisT) so as the subcluster Ie (P. fluoreseens T and P. tolaasii\ The fact that the species P. aureofaeiensT and P. ehlororaphisT were joined together in the same subcluster (IVc, n = 6) fitted in with published data of JOHNSON and PALLERONI (1989), who had shown by DNA/DNA homology experiments, that those two species differing from each other by the coloration of their pigment, formed only one genospecies. Furthermore, on the basis of data of STANIER et al. (1966), and considering only the phenotypic level, MOLIN and TERSTROM (1986b) have already included those both species (P. ehlororaphis DSM 50083T and P. aureofaeiens ATCC 13985T ) in a single taxon in their fluorescent supercluster, as also did SNEATH et al. (1981) and CHAMPION et al. (1980). As to the subcluster Ie (n = 13), it also comprised 2 type strains (P. fluoreseens biovar I ATCC 13525T and P. tolaasii NCPPB 2192T). This could be explained by the fact that P. tolaasii is a species whose relationships with well-characterized species of the genus Pseudomonas are unknown (GOOR et aI., 1986), according to the description of PALLERONI in the Bergey's manual of systematic bacteriology (PALLERONI, 1984). The following subclusters included only collection strains of species P. fluoreseens designated biovar II (subcluster XIIa: ATCC 17482) so as biovars IV and V (subclusters lIb: ATCC 12983, DSM 50148 and ATCC 17573). The subcluster IVb only contained 3 strains of P. putida biovar B (CCUG 1317, ATCC 17484, LMG 1246), which were phenotypically quite different from those of P. putida biovar A (subcluster IVa, see Table 3). At last 3 collection strains did not fall into any subcluster (P. fluoreseens biovar II DSM 50106 from the cluster XII, P. fluoreseens biovar IV DSM 50415 from the cluster XIII, and P. fluoreseens biovar III ATCC 17571 from the cluster XV) .
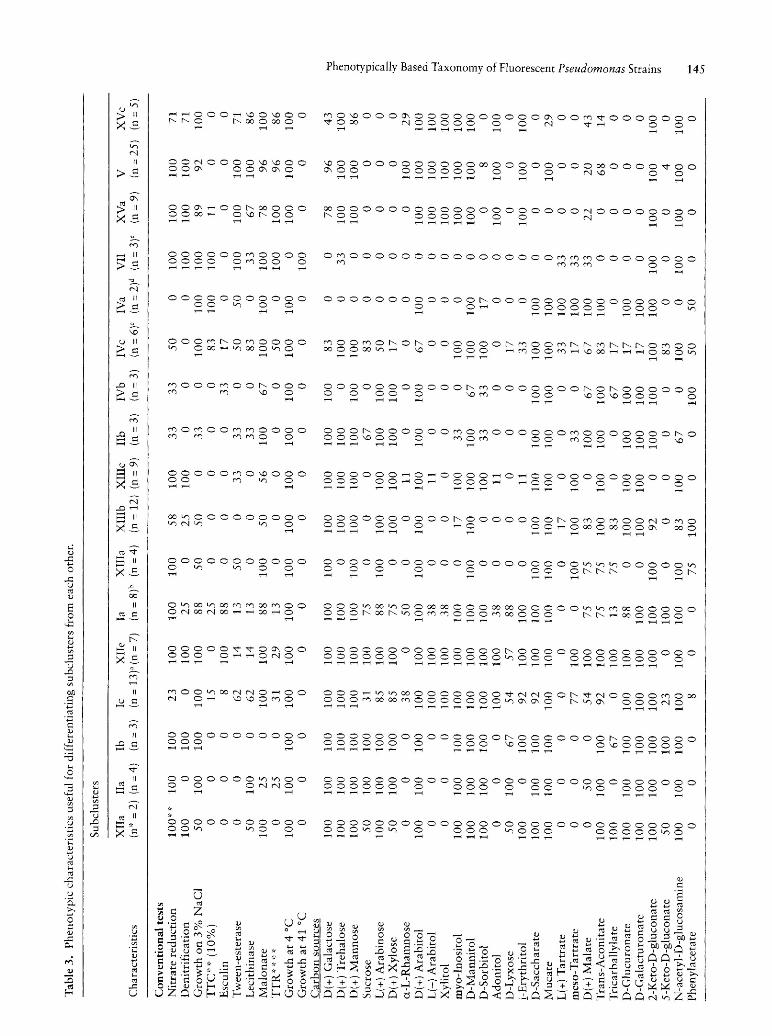
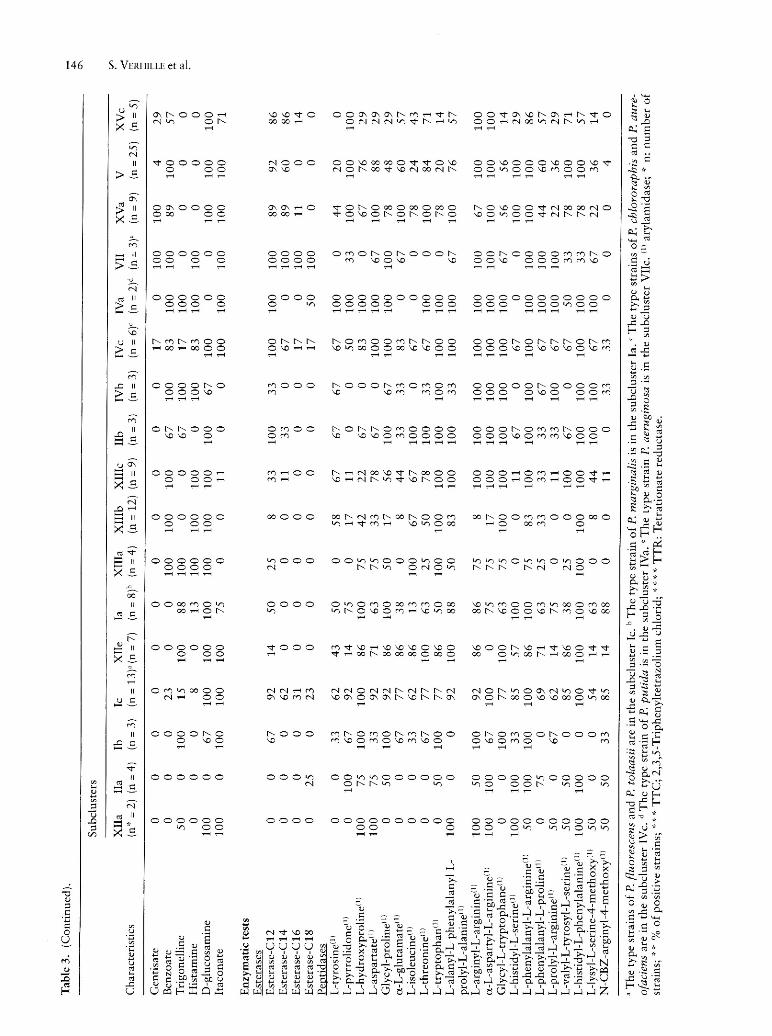
As to mineral water strains, they were mainly grouped into clusters IX, X, XI and subclusters IIa, Ib, XIIe, XIIIa, XIIIb, XIIIc, XIIId, XVa, V, and XVc. Neither the subcluster IIa nor the subcluster Ib has been enlarged with new mineral water strains. Our attention was focussed only on those of them which exhibited a numerically significant number of strains (n = 9 or more), that is to say cluster IX (n = 10) and subclusters XIIIb (n = 12), XIIIc (n = 9), XVa (n = 9) and V (n = 26). Only the last one subcluster was containing two collection strains of P. fluoreseens biovar III (ATCC 17559 and ATCC 17400). Compared to the previous work, this cluster has been considerably enlarged in mineral water strains. Phenotypic tests which proved useful for differentiating the 5 main groups of mineral waters from each other are listed in Table 4. Cluster IX was easily phenotypically wellcharacterized with respect to the other 4 subclusters. As a matter of fact only its strains were able to assimilate aD(+) melibiose, sucrose, D(+) raffinose, maltotriose, maltose, a-lactose, lactulose, l-o-methyl-p-galactopyranoside, l-o-methyl-a-galactopyranoside, D( +) cellobiose, p-gentiobiose, l-o-methyl-p-D-glucopyranoside, esculin, palatinose, a-L(-) fucose, D-tagatose, maltitol, D(+) turanose, D-Iyxose, 3-0-methyl-D-glucopyranose and L( +) tartrate. Compared to other groups only few strains of subcluster XIIIb were able to utilize myo-inositol (17
Tab
le 3
. P
heno
typi
c ch
arac
teri
stic
s us
eful
for
dif
fere
ntia
ting
sub
elus
ters
fro
m e
ach
othe
r.
S u be
l ust
ers
XII
a II
a lb
Ie
X
IIe
la
XIl
Ia
XIl
Ib
XII
Ic
IIb
lVb
lVc
IVa
VII
X
Va
V
XV
c C
hara
cter
istic
s (n
*=2)
(n=
4)
(n=
3)
(n =
13)
' (n
= 7
) (n
=8)
b (n
=4
) (n
= 1
2) (
n =
9)
(n =
3)
(n =
3)
(n =
6)'
(n =
2)d
(n =
3)'
(n =
9)
(n =
25)
(n
= 5
)
Con
vent
iona
l te
sts
Nit
rate
red
ucti
on
100"
"-10
0 10
0 23
10
0 10
0 10
0 58
10
0 33
33
50
0
100
100
100
71
Den
itri
fica
tion
10
0 0
100
0 10
0 25
0
25
100
0 0
0 0
100
100
100
71
Gro
wth
on
3 %
NaC
I 50
10
0 10
0 10
0 10
0 88
50
50
0
33
0 10
0 10
0 10
0 89
92
10
0 T
TC
"* (
10%
) 0
0 0
15
0 25
0
0 0
0 0
83
100
100
11
0 0
Esc
ulin
0
0 0
8 10
0 88
0
0 0
0 33
17
0
0 0
0 0
Tw
een-
este
rase
0
0 0
62
14
13
50
0 33
33
0
50
50
100
100
100
71
Lec
ithin
ase
50
100
0 62
14
13
0
0 0
33
0 83
0
33
67
100
86
Mal
onat
e 10
0 25
0
100
100
88
100
50
56
100
67
100
100
100
78
96
100
TTR'
:-':-'
:-':-
0 25
0
31
29
13
0 0
0 0
0 50
0
100
100
96
86
Gro
wth
at 4
°C
10
0 10
0 10
0 10
0 10
0 10
0 10
0 10
0 10
0 10
0 10
0 10
0 10
0 0
100
100
100
Gro
wth
at
41
°C
0 0
0 0
0 0
0 0
0 0
0 0
0 10
0 0
0 0
Car
bon
sour
ces
D( +
) G
alac
tose
10
0 10
0 10
0 10
0 10
0 10
0 10
0 10
0 10
0 10
0 10
0 83
0
0 78
96
43
'"
D(+
) T
reha
lose
10
0 10
0 10
0 10
0 10
0 10
0 0
100
100
100
0 10
0 0
33
100
100
100
;:r
rt>
::l
D(+
) M
anno
se
100
100
100
100
100
100
100
100
100
100
100
100
0 0
100
100
86
~
Sucr
ose
50
100
100
31
100
75
0 0
0 67
0
83
0 0
0 0
0 '-<
: '0
L(+)
Ara
bino
se
100
100
100
85
100
88
100
100
100
100
100
50
0 0
0 0
0 n- :::.
D
(+)
Xyl
ose
50
100
100
85
100
75
0 10
0 10
0 10
0 10
0 17
0
0 0
0 0
~
u-L-
Rha
mno
se
0 0
0 38
0
50
0 0
11
0 0
0 0
0 0
100
29
o:J
0>
D(+
) A
rabi
tol
100
100
100
100
100
100
100
100
100
100
100
67
100
0 10
0 10
0 10
0 '" '" CL
L
(-)
Ara
bito
l 0
0 0
100
100
38
0 0
11
0 0
0 0
0 10
0 10
0 10
0 ~
Xyl
itol
0 0
0 10
0 10
0 38
0
0 0
0 0
0 0
0 10
0 10
0 10
0 )<
myo
-Ino
sito
l 10
0 10
0 10
0 10
0 10
0 10
0 0
17
100
33
0 10
0 0
0 10
0 10
0 10
0 0 ::
l
D-M
anni
tol
100
100
100
100
100
100
100
100
100
100
67
100
100
0 10
0 10
0 10
0 0 a
D-S
orbi
tol
100
100
100
100
100
100
0 0
100
33
33
100
17
0 0
8 0
'-<: 0
Ado
nito
l 0
0 0
100
100
38
0 0
11
0 0
0 0
0 10
0 10
0 10
0 '"""
..,.,
D
-Lyx
ose
50
100
67
54
57
88
0 0
0 0
0 17
0
0 0
0 0
;::
i-E
ryth
rito
l 10
0 0
100
92
100
100
0 0
11
0 0
33
0 0
100
100
100
0 ... rt>
D-S
acch
arat
e 10
0 10
0 10
0 92
10
0 10
0 10
0 10
0 10
0 10
0 10
0 10
0 10
0 0
0 0
0 '" n
Muc
ate
100
100
100
100
100
100
100
100
100
100
100
100
100
0 0
100
29
'" g L(
+) T
artr
ate
0 0
0 0
0 0
0 17
0
0 0
33
100
33
0 0
0 "" '"
mes
o-T
artr
ate
0 0
0 7
7
100
0 10
0 10
0 10
0 33
0
17
100
33
0 0
0 '" ::=
D(+
) M
alat
e 0
50
0 54
10
0 75
75
83
0
100
67
67
100
33
22
20
43
k 0
Tra
ns-A
coni
tate
10
0 10
0 10
0 92
10
0 75
75
10
0 10
0 10
0 10
0 83
10
0 0
0 68
14
~
0 T
rica
rbal
lyla
te
100
0 67
0
100
13
75
83
0 0
67
17
0 0
0 0
0 ;:,
., D
-Glu
curo
nate
10
0 10
0 10
0 10
0 10
0 88
0
100
100
100
100
17
100
0 0
0 0
'" Vl
D-G
alac
turo
nate
10
0 10
0 10
0 10
0 10
0 10
0 0
100
100
100
100
17
100
0 0
0 0
rt ... 0>
2 -K
eto-
D-g
l uco
na te
10
0 10
0 10
0 10
0 10
0 10
0 10
0 92
0
100
100
100
100
100
100
100
100
S- '" 5 -
Ket
o-D
-glu
cona
te
50
0 10
0 23
0
100
0 0
0 0
0 83
0
0 0
4 0
N-a
cety
l-D
-glu
cosa
min
e 10
0 10
0 10
0 10
0 10
0 10
0 10
0 83
10
0 67
0
100
0 10
0 10
0 10
0 10
0 Ph
enyl
acet
ate
0 0
0 8
0 0
75
100
0 0
100
50
50
0 0
0 0
......
-+>-
v,
Tab
le 3
. (C
onti
nued
).
......
.j:>.
'" S
ubcl
uste
rs
I1a
Ib
la
XII
Ia
XII
Ib
XII
Ic
lIb
IV
b IV
c V
II
XV
a V
X
Vc
~
XII
a Ie
X
IIe
IVa
<
Cha
ract
eris
tics
(n
"=2
) (n
=4
) (n
= 3
) (n
= 1
3)' (
n =
7)
(n=
8)b
(n
=4
) (n
= 1
2) (
n =
9)
(n=
3)
(n=
3)
(n =
6)'
(n =
2)d
(n
= 3
)' (n
= 9
) (n
= 2
5)
(n =
5)
m
::0
::I:
Gen
tisa
te
0 0
0 0
0 0
0 0
0 0
0 1
7
0 1
00
10
0 4
29
P r- m
B
enzo
ate
0 0
0 23
0
0 1
00
1
00
1
00
6
7
100
83
100
10
0
89
100
57
~
Tri
gone
llin
e 5
0
0 10
0 15
10
0 88
10
0 0
0 6
7
10
0
17
10
0 0
0 0
0 ~
His
tam
ine
0 0
0 8
0 13
1
00
1
00
1
00
0
10
0
83
10
0
100
0 0
0 D
-glu
cosa
min
e 1
00
0
67
10
0 10
0 10
0 10
0 1
00
1
00
10
0 6
7
10
0
0 0
100
100
10
0
Itac
onat
e 1
00
0
100
100
100
75
0 0
11
0 0
10
0
10
0
100
100
10
0
71
Enz
ymat
ic te
sts
Est
eras
es
Est
eras
e-C
12
0 0
67
92
14
50
25
8
33
100
33
10
0
10
0
100
89
92
86
E
ster
ase-
C14
0
0 0
62
0
0 0
0 11
33
0
67
0
100
89
60
86
E
ster
ase-
C 1
6 0
0 0
31
0 0
0 0
0 0
0 1
7
0 10
0 11
0
14
E
ster
ase-
C18
0
25
0 23
0
0 0
0 0
0 0
17
5
0
10
0
0 0
0 P
epti
dase
s L
-tyr
osin
e(1 )
0
0 33
6
2
43
50
0 58
6
7
67
6
7
67
1
00
0
44
20
0
L-p
yrro
lido
ne11
) 0
100
67
9
2
14
75
0 1
7
11
0 0
50
1
00
33
10
0 1
00
1
00
L-
h ydr
oxyp
roli
ne(1
) 1
00
75
10
0 1
00
86
10
0 75
4
2
22
6
7
0 83
1
00
0
67
7
6
29
L
-asp
arta
te(1
) 1
00
75
33
92
71
63
75
33
78
6
7
0 1
00
1
00
6
7
100
88
29
G
lycy
l-pr
olin
e(l)
0
50
10
0 9
2
86
100
50
17
5
6
100
67
1
00
10
0 10
0 78
48
2
9
a-L
-glu
tam
ate(
1)
0 0
67
7
7
86
38
0 8
44
33
33
83
0
67
10
0 6
0
57
L
-iso
leuc
ine(
1)
0 0
33
62
86
13
10
0 6
7
67
10
0 0
67
0
0 78
2
4
43
L
-thr
eoni
ne(1
) 0
0 6
7
77
10
0 63
25
5
0
78
10
0
33
67
1
00
0
100
84
71
L-t
rypt
opha
n(1
) 0
50
100
77
86
50
10
0 1
00
1
00
1
00
10
0 1
00
10
0 0
78
20
1
4
L-a
lany
l-L
-phe
nyla
lany
l L
-ID
O 0
0 92
10
0 88
50
83
10
0 1
00
33
1
00
10
0 6
7
100
76
57
pr
olyl
-L-a
lani
ne(l
) L
-arg
inyl
-L-a
rgin
ine(
1)
10
0
50
100
92
86
86
75
8 10
0 1
00
10
0 1
00
10
0 10
0 6
7
100
10
0
a-L
-asp
arty
l-L
-arg
inin
e(1
) 1
00
10
0 6
7
100
0 75
75
1
7
100
10
0
100
10
0
100
100
10
0
100
10
0
Gly
cyl-
L-t
rypt
opha
ne(1
) 0
0 10
0 77
10
0 63
75
1
00
1
00
1
00
10
0 1
00
10
0 6
7
56
56
1
4
L-h
isti
dyl-
L-s
erin
e(1
) 10
0 10
0 33
85
5
7
100
0 0
11
67
0
67
0
0 10
0 10
0 2
9
L-p
heny
lala
nyl-
L-a
rgin
ine(
1)
50
10
0 10
0 1
00
86
10
0 75
83
1
00
1
00
10
0 1
00
10
0 10
0 1
00
1
00
8
6
L-p
heny
lala
nyl-
L-p
roli
ne(1
) 0
75
0 6
9
71
63
25
33
33
33
67
6
7
10
0
100
44
60
5
7
L-p
roly
l-L
-arg
inin
e(l)
5
0
0 6
7
62
1
4
75
0 0
11
33
100
67
1
00
10
0 2
2
36
29
L
-val
yl-L
-tyr
osyl
-L-s
erin
e(1
) 5
0
50
0
85
86
38
25
0 10
0 6
7
0 6
7
50
33
78
1
00
71
L
-his
tidy
l-L
-phe
nyla
lani
ne(1
) 1
00
10
0 0
10
0
100
100
10
0
10
0
100
100
10
0
10
0
10
0
33
78
100
57
L
-lys
yl-L
-ser
ine-
4-m
eth
ox
yl 1)
5
0
0 0
54
1
4
63
0 8
44
10
0 10
0 6
7
100
67
2
2
36
14
N
-CB
Z-a
rgin
yl-4
-met
hox
yl l
) 5
0
50
33
85
1
4
88
0 0
11
0 33
33
0
0 0
4 0
a T
he t
ype
stra
ins
of
P. f
/uor
esce
ns a
nd P
. to
laas
ii a
re i
n th
e su
bclu
ster
Ie.
b T
he t
ype
stra
in o
f P.
mar
gina
lis
is i
n th
e su
bclu
ster
la.
'T
he t
ype
stra
ins
of
P. c
hlor
orap
his
and
P.
aure
-of
acie
ns a
re i
n th
e su
bclu
ster
IV
c. d
The
typ
e st
rain
of
P. p
utid
a is
in
the
subc
lust
er I
Va.
'T
he
type
str
ain
P. a
erug
inos
a is
in
the
subc
lust
er V
IIc.
(I)
ar
ylam
idas
e; "
n:
nu
mb
er o
f st
rain
s; '"
'" %
of
posi
tive
str
ains
; ,. ,
,,:. T
IC;
2,3,
5-T
riph
enyl
tetr
azol
ium
chl
orid
; ,. '
"'" ,.
TT
R:
Tet
rati
onat
e re
duct
ase.
Phenotypically Based Taxonomy of Fluorescent Pseudomonas Strains 147
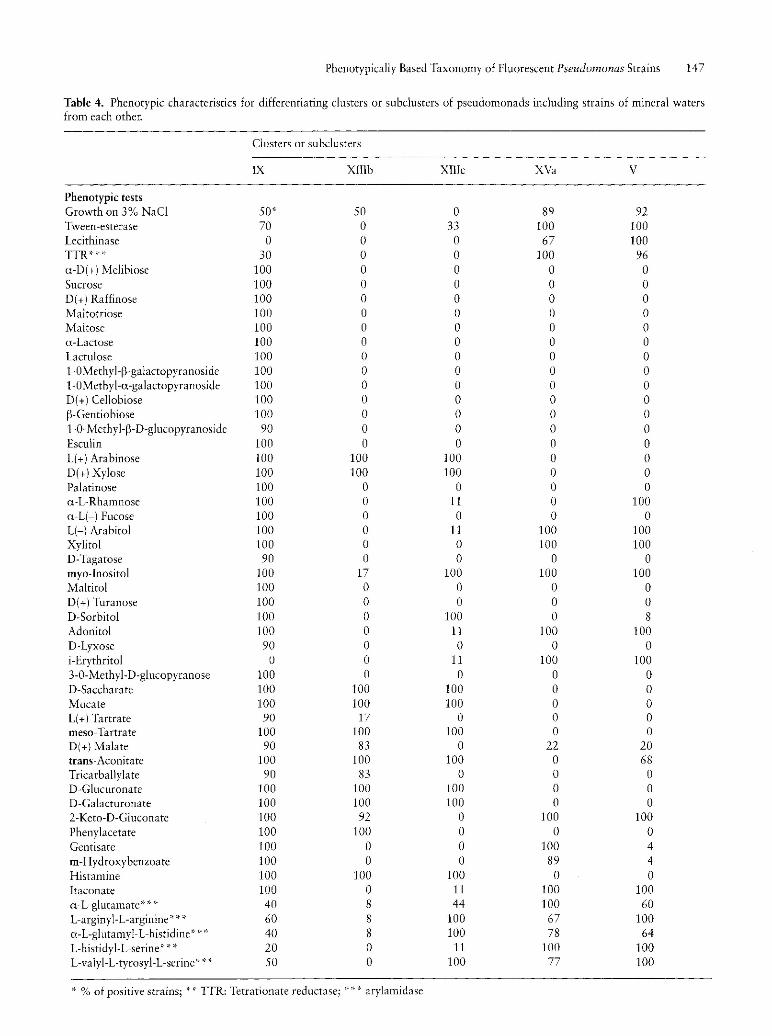
Table 4. Phenotypic characteristics for differentiating clusters or subclusters of pseudomonads including strains of mineral waters from each other.
Clusters or subclusters
IX XIIIb XIIIc XVa V
Phenotypic tests Growth on 3% NaCI 50" 50 0 89 92 Tween -esterase 70 0 33 100 100 Lecithinase 0 0 0 67 100 TTR""" 30 0 0 100 96 a-D(+) Melibiose 100 0 0 0 0 Sucrose 100 0 0 0 0 D ( +) Raffinose 100 0 0 0 0 Maltotriose 100 0 0 0 0 Maltose 100 0 0 0 0 a-Lactose 100 0 0 0 0 Lactulose 100 0 0 0 0 1-0Methyl-~-galactopyranoside 100 0 0 0 0 1-0Methyl-a-galactopyranoside 100 0 0 0 0 D(+) Cellobiose 100 0 0 0 0 ~-Gentiobiose 100 0 0 0 0 1-0-Methyl-~-D-glucopyranoside 90 0 0 0 0 Esculin 100 0 0 0 0 L(+) Arabinose 100 100 100 0 0 D(+) Xylose 100 100 100 0 0 Palatinose 100 0 0 0 0 a-L-Rhamnose 100 0 11 0 100 a-L(-) Fucose 100 0 0 0 0 L(-) Arabitol 100 0 11 100 100 Xylitol 100 0 0 100 100 D-Tagatose 90 0 0 0 0 myo-Inositol 100 17 100 100 100 Maltitol 100 0 0 0 0 D(+) Turanose 100 0 0 0 0 D-Sorbitol 100 0 100 0 8 Adonitol 100 0 11 100 100 D-Lyxose 90 0 0 a a i -Erythri tol a a 11 100 100 3 -0-Methyl-D-glucopyranose 100 0 a a 0 D-Saccharate 100 100 100 0 a Mucate 100 100 100 a a L(+) Tartrate 90 17 a a a meso-Tartrate 100 100 100 a a D(+) Malate 90 83 a 22 20 trans-Aconitate 100 100 100 a 68 Tricarballylate 90 83 a 0 0 D-Glucuronate 100 100 100 a a D-Galacturonate 100 100 100 a a 2-Keto-D-Gluconate 100 92 a 100 100 Phenyl acetate 100 100 a a a Gentisate 100 0 a 100 4 m-Hydroxybenzoate 100 a a 89 4 Histamine 100 100 100 a 0 Itaconate 100 a 11 100 100 a-L-glutamate':' ,. ':. 40 8 44 100 60 L-arginyl-L-arginine" ':. ,. 60 8 100 67 100 a-L-glutamyl-L-histidine" ,:." 40 8 100 78 64 L-histidyl-L-serine" ':. ,. 20 a 11 100 100 L-valyl-L-tyrosyl-L-serine':' ,. ,:. 50 a 100 77 100
':. % of positive strains; ",. TTR: Tetrationate reductase; ':. ':. ". arylamidase

148 S. VERHILLE et al.
against 100%). It is the only one which also showed an unsignificant percentage of strains which possessed the L-arginyl-L-arginine arylamidase activity (8%). As to subcluster XIIIc, it distinguished itself from the other by the inability of its strains to grow on 2-keto-D-gluconate. Compared to other groups, strains of subclusters XIIIb and XIIIc were unable to metabolize xylitol, L(-) arabitol, itaconate and adonitol whereas strains of others groups did. The ability to utilize L( +) arabinose, D( +) xylose, D-saccharate, mucate, meso-tartrate, D-glucuronate, D-galacturonate and histamine were specific characteristics of subclusters IX, XIIIb, XIIIc with regard to subclusters XVa and V. The percentage of strains of groups IX, XIIIb and XIIIc which grew on i-erythritol was unsignificant with respect to the 100% of those of the subclusters XVa and V. Compared to subcluster XIIIb, only the strains of subcluster XIIIc were able to grow on D-sorbitol and exhibited L-arginyl-L-arginine arylamidase, a-L-glutamyl-L-histidine arylamidase and L-valyl-L-tyrosyl-L-serine arylamidase activities, whereas its strains were unable to grow on phenylacetate. At last only strains of subcluster XVa utilized gentisate and a-Lrhamnose as sole carbon source. These results should make it easier the allocation of strains isolated from minerai water to one of those five groups.
The foregoing work not only resulted in groupings of control strains in agreement with the previous study (ELOMARI et aI., 1995) but also allowed us to define 5 homogeneous and numerically significant clusters of Pseudomonas strains isolated from 4 natural mineral waters, on the basis of phenotypic level. Only one of them, the subcluster V, has already been defined in previous work, but this time enlarged in mineral waters strains. However, it is clear that we can be led astray drawing conclusions exclusively on the basis of numerical taxonomy, as not only unrelated type strains may form a cluster as do mineral water isolates, but also closely related species can be placed in different clusters due to properties received vertically from ancestors (PALLERONI, 1993). That is why numerical taxonomy must be regarded as a first stage of much more elaborate investigations such as DNNDNA hybridizations, to reveal whether those 5 groups are also homogeneous on genomic levels. That would help us to point out the taxonomic status of these groups, their relationships to the genus Pseudomonas and their ecological significance.
Acknowledgments We thank C. DUMOULIN for her technical assistance.
References
AZNAR, R., ALCAlDE, E., GARAY, E.: Numerical taxonomy of pseudomonads isolated from water, sediment and eels. System Appl. Microbiol. 14,235-246 (1992).
BARRETT, E. L., SOLANES, R. E., TANG, J. S., PALLERONI, N. J.: Pseudomonas fluorescens biovar V: its resolution into distinct component groups and the relationship of these groups
to other P. fluorescens biovars, to P. putida, and to psychrotrophic pseudomonads associated with food spoilage. J. Gen. Microbiol. 132,2709-2721 (1986).
BOWMAN, J. P., SLY, L. 1., HAYWARD, A. c., SPIEGEL, Y., STACKEBRANDT, E.: Telluria mixta (Pseudomonas mixta BOWMAN, SLY, and HAYWARD 1988) gen. nov., comb. nov., and Telluria chitinolytica sp. nov. soil-dwelling organisms which actively degrade polysaccharides. Int. J. Syst. Bacteriol. 43, 120-124 (1993).
CHAMPION, A. B., BARRETT, E. L., PALLERONI, N. J., SODERBERG, K. L., KUNISAWA, R., CONTOPOULOU, R., WILSON, A. c., DOUDOROFF, M.: Evolution in Pseudomonas fluorescens. J. Gen. Microbiol. 120,485-511 (1980).
DE VOS, P., DE LEY, J.: Intra- and intergeneric similarities of Pseudomonas and Xanthomonas ribosomonal ribonucleic acid cistrons. Int. J. Syst. Bacteriol. 33,487-509 (1983).
DE VOS, P., KERSTERS, K., FALSEN, E., POT, B., GILLIS, M., SEGERS, P., DE LEY, J.: Comamonas DAVID and PARK 1962 gen. nov., nom. rev. emend., and Comamonas terrigena HUGH 1962 sp. nov., nom. rev. Int. J. Syst. Bacteriol. 35, 443-453 (1985).
ELOMARI, M., COROLER, L., IZARD, D., LECLERC, H.: A numerical taxonomic study of fluorescent pseudomonas strains isolated from natural mineral waters. J. Appl. Bacteriol. 78, 71-81 (1995).
GARDAN, L., BOLLET, c., ABU GHORRAH, M., GRIMONT, E, GRIMONT, P. A. D.: DNA relatedness among the pathovar strains of Pseudomonas syringae subsp. savastanoi JANSE (1982) and proposal of Pseudomonas savastanoi sp. nov. Int. J. Syst. Bacteriol. 42, 606-612 (1992).
GAVINI, E, HOLMES, B., IZARD, D., BEJI, A., BERNIGAUD, A., JAKUBCZAK, E.: Numerical taxonomy of Pseudomonas alcaligenes, P. pseudoalcaligenes, P. mendocina, P. stutzeri and related bacteria. Int. J. Syst. Bacteriol. 39, 135-144 (1989).
GAVINI, E, POURCHER, A. M., NEUT, c., MONGET, D., ROMOND, c., OGER, c., IZARD, D.: Phenotypic differentiation of Bifidobacteria of human and animal origins. Int. J. Syst. Bacteriol. 41,548-557 (1991).
GOOR, M., VANTOMME, R., SWINGS, J., GILLIS, M., KERSTERS, K., DE LEY, J.: Phenotypic and genotypic diversity of Pseudomonas tolaasii and white line reacting organisms isolated from cultivated mushrooms. J. Gen. Microbiol. 132, 2249-2264 (1986).
GUILLOT, E., LECLERC, H.: Bacterial flora in natural mineral waters: characterization by ribosomal ribonucleic acid gene restriction patterns. System. Appl. Microbiol. 16, 483-493 (1993).
HOLMES, B., STEIGERWALT, A. G., WEAVER, R. E., BRENNER, DON J.: Chryseomonas luteola comb. nov. and Flavimonas oryzihabitans gen. nov., comb. nov., Pseudomonas-like species from human clinical specimens and formerly known, respectively, as groups Ve-1 and Ve-2. Int. J. Syst. Bacteriol. 37, 245-250 (1987).
HILDEBRAND, D. c., PALLERONI, N. J., HENDSON, M., TOTH, J., JOHNSON, J. L.: Pseudomonas flavescens sp. nov., isolated from walnut blight cankers. Int. J. Syst. Bacteriol. 44, 410-415 (1994).
JOHNSON, J. L., PALLERONI, N. J.: Desoxyribonucleic acid similarities among Pseudomonas species. Int. J. Syst. Bacteriol. 39,230-235 (1989).
KODAMA, K., KIMURA, N., KOMAGATA, K.: Two new species of Pseudomonas: P. oryzihabitans isolated from rice padly and clinical specimens and P. luteola isolated from clinical specimens. Int. J. Syst. Bacteriol. 4, 467-474 (1985).
LECLERC, H.: Le microbisme des eaux minerales naturelles. Hydrogeologie 4,279-285 (1990).

Phenotypically Based Taxonomy of Fluorescent Pseudomonas Strains 149
MOLIN, G., TERNSTROM, A.: Numerical taxonomy of Psychotrophic pseudomonads. J. Gen. Microbiol. 128, 1249-1264 (1982).
MOLIN, G., TERNSTROM, A., URSING, J.: Pseudomonas lundensis, a new bacterial species isolated from meat. Int. J. Syst. Bacteriol. 36, 339-342 (1986a).
MOLIN, G., TERNSTROM, A.: Phenotypically based taxonomy of psychrotrophic Pseudomonas isolated from spoiled meat, water, and soil. Int. J. Syst. Bacteriol. 36,257-274 (1986b).
PALLERONI, N. ]., BALLARD, R. W., RALSTON, E., DOUDOROFF, M.: Deoxyribonucleic acid homologies among some Pseudomonas species. J. Bacteriol. 110, 1-11 (1972).
PALLERONI, N. ]., KUNISAWA, R., CONTOPOULOU, R., DOUDOROFF, M.: Nucleic acid homologies in the genus Pseudomonas. Int. J. Syst. Bacteriol. 23, 333-339 (1973).
PALLERONI, N. J.: Introduction to the family Pseudomonadaceae, pp. 655-665. In: The Prokaryotes. A handbook on habitats, isolation and identification of bacteria (In STARR, M. P., STOLP, H., TRuPER, M. G., BALOWS, A., SCHLEGEL, H. G., ed.). Springer-Verlag; Berlin, 1981.
PALLERONI, N. J.: Pseudomonadaceae, pp. 141-199. In: Bergey's manual of systematic bacteriology, vol. 1 (KRIEG, N. R., HOLT, J. G., eds.). Baltimore, the Williams and Wilkins Co. (1984).
PALLERONI, N. J., BRADBURY, J. E: Strenotrophomonas, a new bacterial genus for Xanthomonas maltophilia (HUGH 1980) SWINGS et aI., 1983. Int. J. Syst. Bacteriol. 43, 606-609 (1993).
PALLERONI, N. J.: Pseudomonas classification. A new case history in the taxonomy of Gram-negative bacteria. Antonie van Leeuwenhoek 64, 231-251 (1993).
SANDS, D. c., SCHROTH, M. N., HILDEBRAND, D. c.: Taxonomy of phytopathogenic pseudomonads. J. Bacteriol. 101, 9-23 (1970).
SEGERS, P., VANCANNEYT, M., POT, B., TORcK, U., HOSTE, B., DEWETTINCK, D., FALSEN, E., KERSTERS, K., DE VOS, P.: Classification of Pseudomonas diminuta LEIFSON and HUGH 1954 and pseudomonas vesicularis BUSING, DOLL, and FREYTAG 1953 in Brevundimonas gen. nov. as Brevundimonas diminuta comb. nov. and Brevundimonas vescularis comb. nov., respectively. Int. J. Bacteriol. 44, 499-510 (1994).
SNEATH, P. H. A., JOHNSON, R.: The influence on numerical taxonomic similarities of errors in microbiological tests. J. Gen. Microbiol. 72, 377-392 (1972).
SNEATH, P. H. A., STEVENS, M., SACKIN, M. J.: Numerical taxonomy of Pseudomonas based on published records of substrate utilization. Antonie van Leeuwenhoek. J. Microbiol. Serol. 47, 423-448 (1981).
STANIER, R. Y., PALI.ERONI, N. ]., DOUDOROFF, N.: The aerobic pseudomonads: a taxonomic study. J. Gen. Microbiol. 43, 159-271 (1966).
TAMAOKA, ]., HA, D. M., KOMAGATA, K.: Reclassification of Pseudomonas acidovorans DEN DOOREN DE JONG 1926 and Pseudomonas testosteroni MARCUS and TALALAY 1956 as Comamonas acidovorans comb. nov. and Comamonas testosteroni comb. nov., with an emended description of the genus Comamonas. Int. J. Syst. Bacteriol. 37, 52-59 (1987).
WOESE, C. R., BLANZ, P., HAHN, C. M.: What isn't a pseudomonad: the importance of nomenclature in bacterial classification. Syst. Appl. Microbiol. 5,179-195 (1984).
WILLEMS, A., BUSSE,]., GOOR, M., POT, B., FALSEN, E., JANTZEN, E., HOSTE, B., GILLIS, M., KERSTERS, K., AULlNG, G. DE LEY, J.: Hydrogenophaga, a new genus of hydrogenoxidizing bacteria that includes Hydrogenophaga (lava comb. nov. (formerly Pseudomonas flava), Hydrogenophaga palleronii (formerly Pseudomonas palleronii), Hydrogenophaga pseudoflava (formerly Pseudomonas pseudoflava and "Pseudomonas carboxydoflava "), and Hydrogenophaga taeniospiralis (formerly Pseudomonas taeniospiralis). Int. J. Syst. Bacteriol. 39, 319-333 (1989).
WILLEMS, A., FALSEN, E., POT, B., JANTZEN, E., HOSTE, B., VANDAMME, P., GILLIS, M., KERSTERS, K., DE LEY, J.: Acidovorax, a new genus for Pseudomonas facilis, Pseudomonas delafieldii, EF group 13, EF group 16, and several clinical isolates, with the species Acidorax facilis comb. nov., Acidovorax delafieldii comb. nov., and Acidovorax temperans sp. nov. Int. J. Syst. Bacteriol. 40, 384-398 (1990).
YABUUCHI, E., YANO, I., OYAIZU, H., HASHIMOTO, Y., EZAKI, T., YAMAMOTO, H.: Proposals of Sphingomonas paucimobilis gen. nov. and comb. nov., Sphingomonas parapaucimobilis sp. nov., Sphingomonas yanoikuyae sp. nov., Sphingomonas adhaesiva sp. nov., Sphingomonas capsulata comb. nov., and two genospecies of the genus Sphingomonas. Microbiol. Immunonol. 34, 99-119 (1990).
YABUUCHI, E., KOSAKO, Y., OYAIZU, H., YANO, I., HOTTA, H., HASHIMOTO, Y, EZAKI, T., ARAKAWA, M.: Proposal of Burkholderia gen. nov. and transfer of seven species of the genus Pseudomonas homology group II to the new genus, with the type species Burkholderia cepacia (PALLERONI and HOLMES 1981) comb. nov. Microbiol. Immunol. 36, 1251-1275 (1992).
Corresponding author: H. LECLERC, Service de bacteriologie-hygiene, Faculte de Medecine, 1 Place de Verdun, F - 59045 Lille, Cedex, France
Copyright © 2022 FDOKUMEN




















