
PCR and patch-clamp analysis of single neurons
6
Neuron, Vol. 14, 1095-1100, June, 1995, Copyright© 1995 by Cell Press PCR and Patch-Clamp Analysis of Single Neurons Neurotechnique Nikolaus J. Sucher* and David L. Deitchert *Laboratory of Cellular and Molecular Neuroscience Department of Neurology Children's Hospital Harvard Medical School Boston, Massachusetts 02115 tDepartment of Molecular and Cellular Physiology Stanford University Stanford, California 94305 Summary The combination of patch-clamp and molecular biol- ogy techniques has made it possible to characterize the pharmacological and biophysical properties of ion channels in single neurons and to screen for expres- sion of specific mRNAs in the same cell. Following whole-cell recording, the cytoplasm of the cell is har- vested, and RNA is reverse transcribed into cDNA and amplified in PCR with primers specific for individual ion channel subunits. Additional experiments can then be designed to relate structure and function at the pro- tein level more directly, since cells appear to regulate the composition of ion channels at least partly at the posttranscriptional stage. Introduction Ion channels are of critical importance to the normal func- tioning of nerve cells. They mediate rapid cellular signaling between neurons and are important for processes underly- ing neuronal plasticity, outgrowth, and survival. The inven- tion of the patch-clamp recording technique has made it possible to characterize the biophysical and pharmacolog- ical properties of single channels. Expression cloning, low stringency screening of cDNA libraries, and PCR/cDNA library screening has led to the cloning of genes encoding voltage- and ligand-gated ion channels. Expression stud- ies suggest that the subunit composition of a channel de- fines its pharmacological and biophysical properties, as exemplified by nicotinic acetylcholine and glutamate recep- tors (Luetje et al., 1990; Nakanishi, 1992). Consequently, the biophysical and pharmacological properties of ligand- gated ion channels in single centrat neurons should corre- late with the expression of specific receptor subunit gene mRNAs. Of particular interest is the question of which sub- unit mRNAs are coexpressed in single cells. Recently, strategies have been developed in an attempt to answer these questions directly. In one method, a whole-cell patch-clamp recording is performed on a cell in a culture or a slice preparation to characterize its biophysical and pharmacological properties, and then its cytoplasmic con- tents are harvested into the patch pipette. The RNA is reverse transcribed into cDNA, which can then be directly amplified by the PCR (Sucher et al., 1991; Lambolez et al., 1992; Bochet et al., 1994; Grigorenko and Yeh, 1994; Jonas et al., 1994). The PCR products can be analyzed by gel electrophoresis and DNA sequencing. Alternatively, the cDNA can be used to obtain amplified RNA (Van Gelder et al., 1990; Eberwine et al., 1992a, 1992b) that, in turn, can be used for expression profiling or a second round of reverse transcription (RT) and subsequent PCR (Mackler and Eberwine, 1993). General Principles of the Experimental Design and Methods Patch-clamp and PCR analysis of single neurons requires the combination of electrophysiological and molecular bio- logical techniques (Figure 1). The following discussion of some considerations that are important for successful patch-clamp and single-cell PCR experiments is based on the first published accounts of this experimental approach (Eberwine et al., 1992b; Lambolez et al., 1992; Mackler and Eberwine, 1993; Bochet et al., 1994; Jonas et al., 1994) but emphasizes many points derived from our own experience (Deitcher, 1992). Many of the details regarding the general experimental design and laboratory practice are important for the establishment of patch-clamp and PCR analysis of single cells in the laboratory but are not commonly communicated in the published research re- ports. Isolation of Total RNA from Single Cells Because of strong capillary forces, it is difficult to harvest a single intact cell with a micropipette, even in cell culture. We have found that harvesting the cytoplasmic contents of a single cell is technically much more feasible. During whole-cell recording, the cell interior is dialyzed with the internal solution contained in the patch-pipette. After a sufficiently long recording, the electrode solution should contain RNA; to obtain the remaining cytoplasmic RNA, suction is applied to the interior of the pipette before termi- nation of the recording (Figure 2). The combination of patch-clamp recording and PCR analysis requires only a few modifications to the standard procedures for whole-cell recording. The modifications in- clude the use of"silanized" glass capillaries for patch elec- trodes, measures aimed at protecting the RNA from the deleterious action of RNases, and the avoidance of con- tamination of the PCR tubes with previously amplified cDNAs. Owing to the ubiquitous presence of RNases, spe- cial care has to be taken to create an RNase-free environ- ment for patch-clamp recording. Several simple but im- portant precautions must be taken by the experimenter. It is essential to wear latex gloves at all times during the preparation of solutions and execution of the experiments to avoid contamination with RNases. In general, the usual KCI patch-pipette internal solutions do not interfere with the subsequent RT or PCR (CsCI solutions can inhibit the RT), but great care should be taken to prepare this solution under RNase-free conditions. Dedicated stocks of chemi- cals and ultrafiltrated, double-distilled water of the highest purity are used. To reduce adhesion of the RNA to the
Transcript of PCR and patch-clamp analysis of single neurons

Neuron, Vol. 14, 1095-1100, June, 1995, Copyright © 1995 by Cell Press
PCR and Patch-Clamp Analysis of Single Neurons
Neurotechnique
Nikolaus J. Sucher* and David L. Deitchert *Laboratory of Cellular and Molecular Neuroscience Department of Neurology Children's Hospital Harvard Medical School Boston, Massachusetts 02115 tDepartment of Molecular and Cellular Physiology Stanford University Stanford, California 94305
Summary
The combination of patch-clamp and molecular biol- ogy techniques has made it possible to characterize the pharmacological and biophysical properties of ion channels in single neurons and to screen for expres- sion of specific mRNAs in the same cell. Following whole-cell recording, the cytoplasm of the cell is har- vested, and RNA is reverse transcribed into cDNA and amplified in PCR with primers specific for individual ion channel subunits. Additional experiments can then be designed to relate structure and function at the pro- tein level more directly, since cells appear to regulate the composition of ion channels at least partly at the posttranscriptional stage.
Introduction
Ion channels are of critical importance to the normal func- tioning of nerve cells. They mediate rapid cellular signaling between neurons and are important for processes underly- ing neuronal plasticity, outgrowth, and survival. The inven- tion of the patch-clamp recording technique has made it possible to characterize the biophysical and pharmacolog- ical properties of single channels. Expression cloning, low stringency screening of cDNA libraries, and PCR/cDNA library screening has led to the cloning of genes encoding voltage- and ligand-gated ion channels. Expression stud- ies suggest that the subunit composition of a channel de- fines its pharmacological and biophysical properties, as exemplified by nicotinic acetylcholine and glutamate recep- tors (Luetje et al., 1990; Nakanishi, 1992). Consequently, the biophysical and pharmacological properties of ligand- gated ion channels in single centrat neurons should corre- late with the expression of specific receptor subunit gene mRNAs. Of particular interest is the question of which sub- unit mRNAs are coexpressed in single cells. Recently, strategies have been developed in an attempt to answer these questions directly. In one method, a whole-cell patch-clamp recording is performed on a cell in a culture or a slice preparation to characterize its biophysical and pharmacological properties, and then its cytoplasmic con- tents are harvested into the patch pipette. The RNA is reverse transcribed into cDNA, which can then be directly amplified by the PCR (Sucher et al., 1991; Lambolez et al., 1992; Bochet et al., 1994; Grigorenko and Yeh, 1994;
Jonas et al., 1994). The PCR products can be analyzed by gel electrophoresis and DNA sequencing. Alternatively, the cDNA can be used to obtain amplified RNA (Van Gelder et al., 1990; Eberwine et al., 1992a, 1992b) that, in turn, can be used for expression profiling or a second round of reverse transcription (RT) and subsequent PCR (Mackler and Eberwine, 1993).
General Principles of the Experimental Design and Methods Patch-clamp and PCR analysis of single neurons requires the combination of electrophysiological and molecular bio- logical techniques (Figure 1). The following discussion of some considerations that are important for successful patch-clamp and single-cell PCR experiments is based on the first published accounts of this experimental approach (Eberwine et al., 1992b; Lambolez et al., 1992; Mackler and Eberwine, 1993; Bochet et al., 1994; Jonas et al., 1994) but emphasizes many points derived from our own experience (Deitcher, 1992). Many of the details regarding the general experimental design and laboratory practice are important for the establishment of patch-clamp and PCR analysis of single cells in the laboratory but are not commonly communicated in the published research re- ports.
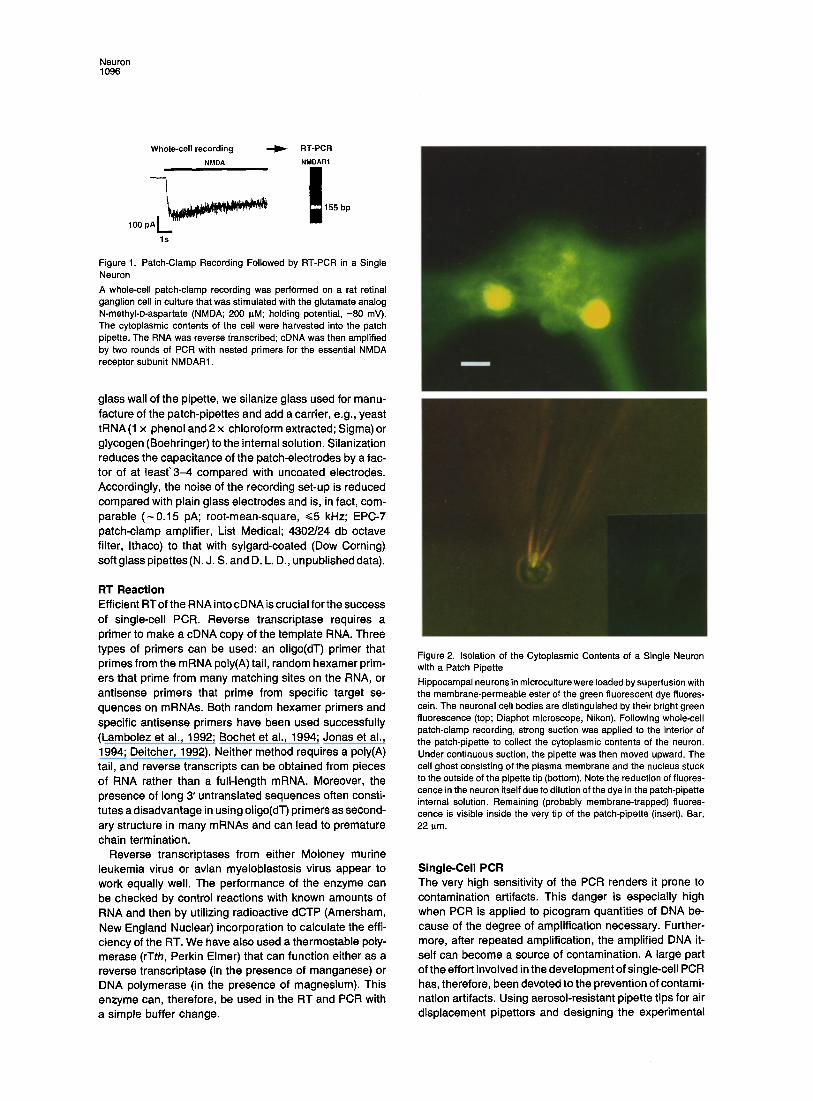
Isolation of Total RNA from Single Cells Because of strong capillary forces, it is difficult to harvest a single intact cell with a micropipette, even in cell culture. We have found that harvesting the cytoplasmic contents of a single cell is technically much more feasible. During whole-cell recording, the cell interior is dialyzed with the internal solution contained in the patch-pipette. After a sufficiently long recording, the electrode solution should contain RNA; to obtain the remaining cytoplasmic RNA, suction is applied to the interior of the pipette before termi- nation of the recording (Figure 2).
The combination of patch-clamp recording and PCR analysis requires only a few modifications to the standard procedures for whole-cell recording. The modifications in- clude the use of"silanized" glass capillaries for patch elec- trodes, measures aimed at protecting the RNA from the deleterious action of RNases, and the avoidance of con- tamination of the PCR tubes with previously amplified cDNAs. Owing to the ubiquitous presence of RNases, spe- cial care has to be taken to create an RNase-free environ- ment for patch-clamp recording. Several simple but im- portant precautions must be taken by the experimenter. It is essential to wear latex gloves at all times during the preparation of solutions and execution of the experiments to avoid contamination with RNases. In general, the usual KCI patch-pipette internal solutions do not interfere with the subsequent RT or PCR (CsCI solutions can inhibit the RT), but great care should be taken to prepare this solution under RNase-free conditions. Dedicated stocks of chemi- cals and ultrafiltrated, double-distilled water of the highest purity are used. To reduce adhesion of the RNA to the
Neuron 1096
Whole-cell recording , ~ RT-PCR NMDA NMDAR1
100 pA L lS
Figure 1. Patch-Clamp Recording Followed by RT-PCR in a Single Neuron A whole-cell patch-clamp recording was performed on a rat retinal ganglion cell in culture that was stimulated with the glutamate analog N-methyI-D-aspartate (NMDA; 200 ~M; holding potential, -80 mV). The cytoplasmic contents of the cell were harvested into the patch pipette. The RNA was reverse franscribed; cDNA was then amplified by two rounds of PCR with nested primers for the essential NMDA receptor subunit NMDARI.
glass wall of the pipette, we silanize glass used for manu- facture of the patch-pipettes and add a carrier, e.g., yeast tRNA (1 x phenol and 2 x chloroform extracted; Sigma) or glycogen (Boehringer) to the internal solution. Silanization reduces the capacitance of the patch-electrodes by a fac- tor of at least" 3-4 compared with uncoated electrodes. Accordingly, the noise of the recording set-up is reduced compared with plain glass electrodes and is, in fact, com- parable ( -0 .15 pA; root-mean-square, ~<5 kHz; EPC-7 patch-clamp amplifier, List Medical; 4302•24 db octave filter, Ithaco) to that with sylgard-coated (Dow Coming) soft glass pipettes (N. J. S. and D. L. D., unpublished data).
RT Reaction Efficient RT of the RNA into cD NA is crucial for the success of single-cell PCR. Reverse transcriptase requires a primer to make a cDNA copy of the template RNA. Three types of primers can be used: an oligo(dT) primer that primes from the mRNA poly(A) tail, random hexamer prim- ers that prime from many matching sites on the RNA, or antisense primers that prime from specific target se- quences on mRNAs. Both random hexamer primers and specific antisense primers have been used successfully (Lambolez et al., 1992; Bochet et al., 1994; Jonas et al., 1994; Deitcher, 1992). Neither method requires a poly(A) tail, and reverse transcripts can be obtained from pieces of RNA rather than a full-length mRNA. Moreover, the presence of long 3' untranslated sequences often consti- tutes a disadvantage in using oligo(dT) primers as second- ary structure in many mRNAs and can lead to premature chain termination.
Reverse transcriptases from either Moloney murine leukemia virus or avian myeloblastosis virus appear to work equally well. The performance of the enzyme can be checked by control reactions with known amounts of RNA and then by utilizing radioactive dCTP (Amersham, New England Nuclear) incorporation to calculate the effi- ciency of the RT. We have also used a thermostable poly- merase (rTth, Perkin Elmer) that can function either as a reverse transcriptase (in the presence of manganese) or DNA polymerase (in the presence of magnesium). This enzyme can, therefore, be used in the RT and PCR with a simple buffer change.
Figure 2. Isolation of the Cytoplasmic Contents of a Single Neuron with a Patch Pipette Hippocampal neurons in microculture were loaded by superfusion with the membrane-permeable ester of the green fluorescent dye fluores- cein. The neuronal cell bodies are distinguished by their bright green fluorescence (top; Diaphot microscope, Nikon). Following whole-cell patch-clamp recording, strong suction was applied to the interior of the patch-pipette to collect the cytoplasmic contents of the neuron. Under continuous suction, the pipette was then moved upward. The cell ghost consisting of the plasma membrane and the nucleus stuck to the outside of the pipette tip (bottom). Note the reduction of fluores- cence in the neuron itself due to dilution of the dye in the patch-pipette internal solution. Remaining (probably membrane-trapped) fluores- cence is visible inside the very tip of the patch-pipette (insert). Bar, 22 p~m.
Single-Cell PCR The very high sensitivity of the PCR renders it prone to contamination artifacts. This danger is especially high when PCR is applied to picogram quantities of DNA be- cause of the degree of amplification necessary. Further- more, after repeated amplification, the amplified DNA it- self can become a source of contamination. A large part of the effort involved in the development of si ngle-cell PCR has, therefore, been devoted to the prevention of contami- nation artifacts. Using aerosol-resistant pipette tips for air displacement pipettors and designing the experimental

Neurotechnique 1097
A bp
242 190 147
Bim ~2
,,,-- (~3 ~m ~4
Thy-1
B M 1 2 3 4 5 6 C C C 7 8 9 10 11 12 13 C
Figure 3. Multiplex PCR Analysis of Total RNA and RNA from Single Rat Retinal Ganglion Cells (A) Multiplex PCR with nested primers for nico- tinic acetylcholine receptor (nAchR) ~ sub- units. Postnatal rat retinal RNA was used for RT and multiplex PCR. Three sets of primer pairs corresponding to the nAchR a2, ct3, and ~4 subunits were used in a first-round one-tube PCR (30 cycles). This mix (0.1 v) was then used for a second PCR (25 cycles) with three pairs of nested primers. The PCR products from the second-round PCR were analyzed by agarose gel electrophoresis (right lane; molecular size standard Boehringer VIII in the left lane). PCR products were verified by Southern blotting and/or DNA sequencing. Primers for the cell surface glycoprotein Thy-1 were initially added as a positive control.
(B) Multiplex PCR applied to single retinal ganglion cells. Agarose gel electrophoresis of the PCR products that were obtained in the second-round multiplex PCR with the nAchR a subunit primers. Only the nAchR subunit ~3 was detected in 2 out of 13 ganglion cells tested (lanes labeled 1- 13). Single-cell RNA was purified before RT. Control reactions without reverse transcriptase are labeled as C. Size standard like in (A). Retinal ganglion cells were identified by the presence of the fluorescent dye granular blue following retrograde labeling in situ (Zhang et al., 1995).
protocol to reduce the chance of contamination are both extremely important. A measure that has proven most ef- fective in preventing DNA contamination is the use of DNase on buffers prior to the first strand synthesis, fol- lowed by digestion with proteinase K, which can subse- quently be heat inactivated.
Design of PCR Primers "Good" primers are essential for the success of the PCR (reviewed by Dieffenbach et al., 1993). We have found that primers of between 20 and 24 nucleotides, with guanosine and cytosine (GC) content of 5 0 o - 6 5 % work well. The two members of a primer pair to be used in the same reaction should have approximately the same GC content and similar melting temperature. Care should be taken that the 5' and 3' ends of the primers are not complemen- tary to prevent the formation of hairpins or primer dimers. It may also be desirable to add restriction sites to the 5' ends of each primer to facilitate subcloning.
To increase the specificity and sensitivity of the PCR, we routinely use nested primers in consecutive PCRs; i.e., a first round of PCR is run with one set of outer primers, and then an aliquot (5 ~1) of this reaction is used for a second round of PCR with a set of internal primers. The 5' and 3' primers are chosen from parts specific to the gene to be detected to avoid amplification of closely ho- mologous genes. Moreover, the sequences of the 5' and 3' primers of each pair should be taken from different ex- ons to avoid amplifying genomic DNA, which will yield PCR products of larger size compared with amplification of cDNA.
Primers meeting the above criteria generally work at annealing temperatures between 48°C and 60°C. The ex- act protocol for the PCR (Mg 2÷ and primer concentration, annealing temperature, and number of cycles) should be optimized. For every PCR, we include control tubes with- out cDNA to exclude contamination artifacts. Primer stocks
should be aliquoted to avoid contamination. The prepara- tory experiments should be carried out in a separate loca- tion, i.e., not in the same laboratory as the single-celt PCR experiments.
In most instances, the goal of combined patch-clamp and single-cell PCR is the detection of a pattern of expres- sion of multiple genes in a single cell, rather than the detec- tion of a single transcript. We have established conditions for a nested multiplex PCR with primers for three of the nicotinic acetylcholine receptor ~ subunits (Figures 3A and 3B).
Other methods include the use of degenerate primers (Kwok et al., 1994) and the use of primers that are homolo- gous to highly conserved regions in a family of genes. The latter approach was successfully applied by Lambolez and colleagues (1992). In their study, the first round of amplifi- cation was performed using primers that target a se- quence that is highly conserved in AMPA-type glutamate receptor genes. Primer pairs specific for individual genes and splice variants were used in the second round to differ- entiate between them. Alternatively, a second round of PCR was conducted with the same primer pair used in the first round, and the resulting PCR products were ana- lyzed by restriction enzyme analysis.
We used a degenerate primer approach to amplify N-methyI-D-aspartate (NMDA)-type glutamate receptors. The 3' primer was designed to target a sequence found in both NMDA and non-NMDA receptors. For the 5' primer, we chose a sequence motif found only in NMDA receptor subunits. The degenerate primers were tested with cDNA from retina, brain, and PC12 cells. The PCR products were subcloned, and >40 individual clones were sequenced. We obtained products corresponding to NMDAR1 and all known NR2 subunits but not the non-NMDA receptor sub- units. In addition, we identified one subclone with a sequence from a previously unknown gene with close ho- mology to NMDA receptor subunits. This subcione was sub- sequently used to isolate full-length clones by cDNA library

Neuron 1098
M 10 4 10 3 10 2
Figure 4. One-TubeRT-PCRwithStandardRNAandrTthPolymerase Agarose gel electrophoresis followed by ethidium bromide staining indicated that the FIT and PCR amplification (45 cycles) were approxi- mately quantitative for 102 to 104 control RNA molecules, The size of the PCR product is 308 bp. Size standard (M) is Boehringer VIII.
screening (Sucher et al., 1994). These primers should be useful for single-cell PCR experiments.
Optimization of the RT and PCRs The conditions for the RT reaction and PCR should be optimized with small amounts of a standard RNA sample (e.g., from brain) before single-cell experiments are at- tempted. RNA is isolated from brain, quantified, reverse transcribed, and the cDNA is then diluted to a concentra- tion of 50-500 pg/p.l. For the PCR, 1 Id cDNA (correspond- ing to 1-10 cells) is used.
The conditions developed with very small amounts of cDNA ( - 50 pg) are subsequently applied to PCR with cDNA from single neurons. A high number of amplification cycles (45 cycles for each of the two PCR rounds) creates an increased risk of contamination. Even with very high standards for cleanliness, false positive results can be obtained. In fact, designing the appropriate controls and dealing with potential contamination has been a significant part of the development of reliable single-cell PCR. We have virtually been able to abolish the contamination prob- lem by reducing the number of amplification cycles (25-35 cycles) in each set of PCR and by treating our solutions with DNase (RNase-free DNase I) before adding the cDNA.
Quantitative Single-Cell PCR? Quantitative measurement of mRNAs by PCR has been reported to detect 2-fold differences in mRNAs (Chelly et al., 1990). However, quantitation of the very low number of mRNA copies in single cells is complicated by the chaotic nature of the PCR during the initial cycles of the reaction. The problems associated with attempts at developing RT- PCR to quantitate gene expression have been discussed in detail elsewhere (Gause and Adamovicz, 1994). Using the rTth kit from Perkin Elmer, we have been able to reli- ably reverse transcribe and amplify -100 copies of the control RNA. Furthermore, ethidium bromide-stained gels indicate that the amplification was approximately quantita- tive across a range of 10 ~ to 104 control RNA molecules (Figure 4). It should be kept in mind that the reaction condi- tions were optimized for the respective template and prim- ers. The efficiency of the PCR, however, varies consider- ably depending on the specific template and primers used
(Cha and Thilly, 1993). Although these control experiments indicate that it is possible to deduce the initial number of target molecules from the final amount of PCR product, it is in practice much more realistic to determine relative differences in the number of target molecules.
This point may be illustrated by a quantitative analysis of the results obtained by Lambolez and colleagues in their study involving the amplification of AMPA receptor subunits in single Purkinje cells. The authors determined the amount of specific product obtained from single-cell cDNA after 35 cycles of PCR. The yield per PCR varied from 10 to 100 ng of a 750 bp product as estimated from agarose gels. This corresponds to - 1,2 x 10 TM molecules. Under optimal PCR conditions (efficiency = 1; Cha and Thilly, 1993), with a doubling of PCR products starting at the fourth cycle of the reaction, only ~ 25 molecules would have had to be present initially to obtain 100 ng of product after 35 PCR cycles. Analysis of the PCR products re- vealed that they corresponded to at least five individual GluR 1-4 splice variants. Thus, only five molecules of each splice variant would have been necessary to yield these results had the PCR been conducted with absolute effi- ciency. However, the efficiency of the PCR is generally less than 1 and was so probably in these experiments, given that PCR products corresponding to several splice variants were detected in reactions with only 10-20 ng of final PCR product (corresponding to an average of -1 molecule of each splice variant at the start of the PCR). It is likely, therefore, that the actual number of molecules corresponding to each splice variant at the beginning of the PCR was higher than the apparent number. Not only is the efficiency of PCR generally less than 1, but the effi- ciency of the RT is also frequently low.
For relative quantitation of mRNAs, a standard must be amplified with the target cDNA. The standard can either be an mRNA that is present in the cells (endogenous RNA) or an exogenous RNA that is added to the single-cell RNA before RT. The use of an endogenous standard has sev- eral advantages. First, the detection of standard cDNA after RT and PCR indicates the successful isolation of single-cell RNA. Second, the amount of cDNA correspond- ing to the endogenous marker is an indicator of the degree of degradation and purity of the sample. Although some attempts at relative quantitation have been reported pre- viously, it should be noted that control experiments were either conducted with RNA levels that were up to 105- to 106-fold higher than in a single cell (Lambolez et al., 1992) or did not attempt to take the RT step into account (Jonas et al., 1994). This is important because the efficiency of the RT reaction has not been characterized in detail at very low levels of template and most likely varies with the particular primers and priming methods used. Moreover, it is impossible to control the efficiency of the RNA isolation from a single cell entirely.
Conclusion and Outlook The extreme sensitivity of the PCR poses a formidable experimental challenge because of the very low signal-to- noise ratio when low copy num her transcripts are amplified

Neurotechnique 1099
from single cells. Further study toward the end of quantita- tive single-cell PCR is required. Particularly, the efficiency of the RNA harvest and the RT must be determined. A particularly chal lenging and promising field for future in- vestigations is the detection in single cells of cell t ype- specific expression of ion channel gene splice variants and their effect on function (Sommer et al., 1990; Tingley et al., 1993). Single-cell PCR can be used to investigate the expression of mRNAs coding for other proteins, e.g., different isoforms of enzymes. However, the control of gene expression does not end at transcription. Posttran- scriptional control of RNA fate, posttranslational pro- cessing, and regulated assembly and disassembly of multisubunit proteins are emerging as important factors in neuronal function. Single-cell PCR experiments will l ikely raise the awareness for the importance of posttranscrip- tional mechanisms in the control of gene expression (Sheng et al., 1993; Sucher et a l , 1993; Brose et al., 1994; Zhang et al., 1995).
Experimental Procedures
Preparation of "Slick" Glass Pipettes Glass capillaries (Fisher Scientific, Fair Lawn, N J) used for manufactur- ing of patch-pipettes were soaked in a 5% solution of dimethyldichloro- silane (Sigma, St. Louis, MO) in CHCI~ (Fluka, Ronkonkoma, NY) for 20 rain. Excess liquid was then poured off and allowed to evaporate in a fume hood. Finally, the capillaries were autoclaved (15 rain) and steam dried (15 rain). The capillaries were stored in 50 ml tubes (Corn- ing) for up to several weeks. The electrode resistance measured in the bath was 1-4 MQ with standard solutions.
Isolation of Total RNA from Single Cells Suction was applied to the interior of the patch pipette. Inspection through the microscope showed that the cell was shrinking and its contents were moving into the pipette tip. The remnants of the cell, including the nucleus, stuck to the outside of the pipette tip. Under continuous suction, the pipette was reeved upward across the liquid- air interface. The pipette contents ( -7-10 ~1 of the internal solution) were then ejected by positive pressure (6-9 psi) into a reaction tube (0.65 ml, silanized and autoclaved) containing either the RT mix or a denaturing solution for RNA purificaticn.
RT cDNA was reverse transcribed from RNA using reverse transcriptase from avian myeloblastosis virus with random hexamer primers. The hexamers and RNA in diethyl pyrocarbcnate-treated H20 were heated to 70°C for 10 min and then chilled on ice before adding the additional components. RNA from single cells was reverse transcribed in a 20 ~1 mix containing 5 ~.M random hexamers, 1 mM 2'-deoxynucleoside 5'-triphosphates (dNTPs; Pharmacia), 40 U RNasin (Promega), and 32 U avian myeloblastosis virus reverse transcriptase (Molecular Genetic Resources) in 1 x buffer (10x buffer: 0.5 M Tris [pH 8.8 at 25°C], 100 mM MgCI2, 10 mM dithiothreitol, 10 mM EDTA, 100 ~g/ ml bovine serum albumin). The mix was incubated for 1 hr at 42°C. The reaction was stopped by incubating at 80°C for 5 rain, and the mix was then placed on ice for 10 rain
RNA purification appeared to be beneficial in some cases. The RNA in the patch-pipette was purified with denaturing solution (scaled down from Chomczynski and Sacchi, 1987; RNazol, Biotecx Laboratories, Houston, TX). The purified RNA pellet was rinsed three times with cold 70% ethanol and stored at -80°C in 70°/0 ethanol or resuspended in H2O for immediate use. Centrifugation steps were performed using Eppendorf and Sorvall (DuPont) centrifuges.
PCR Two rounds of PCR (30-35 and 25-30 cycles in Perkin Elmer thermocy- clers 480 or 9600) with two sets of nested primers were used. Samples
without reverse transcriptase were used to detect contamination by cDNAs. The second round was performed with 5 p,I amplified cDNA from the first PCR. It may be advantageous to purify the first-round PCR products before further use in the second round (PCR purification columns, Qiagen). The PCR was optimized for each primer pair using 50 pg to 5 ng of a standard cDNA. The Mg 2÷ concentration was varied between 0.5 and 3 raM, the temperature between 48°C and 60°C. The PCR (50-100 ill mix) contained 200 pM of each dNTP, 10 pmol 5' primer, 10 pmol 3' primer, 2.5 U thermostable DNA polymerase in 1 x reaction buffer (10 x buffer: 100 m M Tris [pH 8.3], 500 mM KCI, with or without 0.01% gelatin [w/v]). In the first-round PCR mix without primers, cDNA and Taq polymerase (Perkin Elmer) was pretreated with 5-10 U DNase I for 15 min at 37°C to destroy contaminating DNA. The DNase was then digested with 12 p.g Proteinase K (Boehringer) at 70°C for 20 min, which was heat inactivated (85°C for 20 rain and 95°C for 15 min). Primers, cDNA, and Taq polymerase were added before the PCR. PCR products were analyzed by agarose gel electro- phoresis (American Bioanalytical; Owl Scientific), restriction digests (enzymes from New England Biolabs, Boehdnger, and Promega), and DNA sequencing (cycle sequencing kit and Prism DNA sequencer, Applied Biosystems).
Acknowledgments
Part of this work was supported by National Institutes of Health grant PO1 HD29587. We would like to thank Drs. Connie L. Cepko and Stuart A. Lipton for support and encouragement, Marc Awobuluyi, Michael Yeh, and Joseph Yuan for critical reading of the manuscript, Dick Bennett for DNA sequencing, Michele Wu for technical assis- tance, and Dr. Michael M. Segal for help with the experiment for Fig- ure 2.
Received January 4, 1995.
References
Bochet, P., Audinat, E., Lambolez, B., Crepel, F., Rossier, J., line, M., Tsuzuki, K., and Ozawa, S. (1994). Subunit composition at the single-cell level explains functional properties of a glutamate-gated channel. Neuron 12, 383-366. Brose, N., Huntley, G. W., Stern-Bach, Y., Sharma, G., Morrison, J. H., and Heinemann, S. F. (1994). Differential assembly of coex- pressed glutamate receptor subunits in neurons of rat cerebral cortex. J. Biol. Chem. 269, 16780-16784. Cha, R. S., and Thilly, W. G. (1993). Specificity, efficiency, and fidelity of PCR. PCR Meth. Appl. (Suppl.) 3, 18-29. Chelly, J., Montarras, D., Pinset, C., Berwald-Netter, Y., and Kaplan, J.-C. (1990). Quantitative estimation of minor mRNAs by cDNA- polymerase chain reaction: application to dystrophin mRNA in cultured myogenic and brain cells. Eur. J. Biochem. 187, 691-698. Chomczynski, P., and Sacchi, N. (1987). Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extrac- tion. Anal. Biochem. 162, 156-159. Deitcher, D. L. (1992). Isolation and characterization of a novel homeo- box gene expressed in sensory organs. PhD thesis, Harvard Medical School, Cambridge, Massachusetts. Dieffenbach, C. W., Lowe, T. M. J., and Dveksler, G. S. (1993). General concepts for PCR primer design. PCR Meth. Appl. (Suppl.) 3, 30-37. Eberwine, J., Spencer, C., Miyashiro, K., Mackler, S., and Finnell, R. (1992a). Complementary DNA synthesis in situ: methods and applica- tions. Meth. Enzymol. 216, 80-100. Eberwine, J., Yeh, H., Miyashiro, K., Cao, Y., Nair, S., Finnell, R., Zettel, M., and Coleman, P. (1992b). Analysis of gene expression in single live neurons. Proc. Natl. Acad. Sci. USA 89, 3010-3014. Gause, W. C., and Adamovicz, J. (1994). The use of the PCR to quanti- tate gene expression. PCR Meth. Appl. (Suppl.) 3, 123-135. Grigorenko, E. V., and Yeh, H. H. (1994). GABAA receptor subunit profiles in single photoreceptors and bipolar cells of the rat retina. Invest. Ophthal. Vis. Sci. 35, 1364. Jonas, P., Racca, C., Sakmann, B., Seeburg, P. H., and Monyer, H.

Neuron 1100
(1994). Differences in Ca ~- permeability of AMPA-type glutamate re- ceptor channels in neocortical neurons caused by differential GluR-B subunit expression. Neuron 12, 1281-1289. Kwok, S., Chang, S.-Y., Sninsky, J. J., and Wang, A. (1994). A guide to the design and use of mismatched and degenerate primers. PCR Meth. Appl. (Suppl.) 3, 39-47. Lambolez, B., Audinat, E., Bochet, P., Crepel, F., and Rossier, J. (1992). AMPA receptor subunits expressed by single Purkinje cells. Neuron 9, 247-258. Luetje, C. W., Patrick, J. W., and S6gu~la, P. (1990). Nicotine receptors in the mammalian brain. FASEB J. 4, 2753-2760. Mackler, S. A., and Eberwine, J. H. (1993). Diversity of glutamate receptor subunit mRNA expression within live hippocampal CA1 neu- rons. Mol. Pharmacol. 44, 308-315. Nakanishi, S. (1992). Molecular diversity of glutamate receptors and implications for brain function. Science 258, 597-603. Sheng, M., Liao, Y. J., Jan, Y. N., and Jan, L. Y. (1993). Presynaptic A-current based on heteromultimeric K + channels detected in vivo. Nature 365, 72-75. Sommer, B., Kein&nen, K., Verdoorn, T. A., Wisden, W., Burnashev, N., Herb, A., K6hler, M., Takagi, T., Sakmann, B., and Seeburg, P. H. (1990). Flip and flop: a cell-specific functional switch in glutamate- operated channels of the CNS. Science 249, 1580-1585. Sucher, N. J., Brose, N., Deitcher, D. L., Awobuluyi, M., Gasic, G. P., Bading, H., Cepko, C. L., Greenberg, M. E., Jahn, R., Heinemann, S. F., and Lipton. S. A. (1993). Expression of endogenous NMDAR1 transcripts without receptor protein suggests post-transcriptional con- trol in PC12 cells. J. Biol. Chem. 268, 22299-22304. Sucher, N. J., Chi, C. L., Awobuluyi, M., Leclerc, C. L., Deitcher, D. L., and Lipton. S. A. (1994). A novel gene in the CNS with intermedi- ate homology to NMDA and non-NMDA receptor subunits. Soc. Neu- rosci. Abstr. 20, 737. Sucher, N. J., Deitcher, D. L., Cepko, C. L., and Lipton, S. A. (1991). Patch clamp and PCR on a single rat retinal ganglion cell: neuronal nicotinic acetylcholine receptor (nAChR) subunits. Invest. Ophthal. Vis. Sci. 32, 992. Tingley, W. G., Roche, K. W., Thompson, A. K., and Huganir, R. L. (1993). Regulation of NMDA receptor phosphorylation by alternative splicing of the C-terminal domain. Nature 364, 70-73. Van Geider, R. N., von Zastrow, M. E., Yool, A., Dement, W. C., Bar- chas, J. D., and Eberwine, J. H. (1990). Amplified RNA synthesized from limited quantities of heterogeneous cDNA. Proc. Natl. Acad. Sci. USA 87, 1663-1667. Zhang, D., Sucher, N. J., and Lipton, S. A. (1995). Co-expression of AMPNkainate receptor-operated channels with high and low Ca 2÷ permeability in single rat retinal ganglion gells. Neuroscience, in press.
![Patch Antenna[1]](https://static.fdokumen.com/doc/165x107/63158e4cc32ab5e46f0d5c89/patch-antenna1.jpg)



















