
SEASONAL CHANGE IN LEAF AREA INDEX AT THREE SITES ALONG A SOUTH AMERICAN LATITUDINAL GRADIENT

- How do patterns of plant invasion change along an environmental stress gradient? - 47
Journal of Vegetation Science 17: 47-56, 2006© IAVS; Opulus Press Uppsala.
Abstract.Question: Do stressful environments facilitate plant invasionby providing refuges from intense above-ground competitionassociated with productive areas, or prevent it by favouringlocally adapted native species?Location: An invaded and fragmented oak savanna ecosys-tem structured along a landscape-level stress gradient associ-ated with soil depth, elevation, and canopy openness.Methods: Vegetation and environmental data were collectedfrom 184 plots in seven savanna remnants along the gradient.Using multivariate (CCA) and post-hoc regression analyses,we determined the relationship between environment and therichness and abundance of invasives.Results: 46 of 119 species were naturalized exotics. CCAindicated the importance of environmental variation (mostlysoil depth) for community structure but not for invasion;invasive species richness was similar in all areas. However,the abundance of invasives and their impacts on native diver-sity appear to increase significantly in less stressful habitats.Deeper soils had lower evenness and significantly fewer na-tive species. This result was associated with dominance byexotic perennial grasses and large increases in vegetationheight, suggesting strong above-ground competition.Conclusions: Low-stress environments were not moreinvasible per se but appear to be more susceptible to invasionby species with strong competitive impacts. The causes ofdecreasing exotic impact with decreasing soil depth may re-flect shifts in competitive intensity or an increased importanceof stress tolerance, both of which may favour natives. Alterna-tively, this ecosystem may simply lack high-impact invaderscapable of dominating shallow soils. Conservation challengesare twofold for this endangered plant community: controllinginvasives that currently dominate deeper-soils and accountingfor a diverse pool of invaders that proliferate when the currentdominants are removed.
Keywords: Biodiversity; British Columbia; Competition;Conservation; Facilitation; Oak savanna; Quercus garryana;Refuge; Soil depth.
Nomenclature: Douglas et al. 2001.
Patterns of plant invasion along an environmental stress gradient
MacDougall, A.S.1,2*; Boucher, J.1,3; Turkington, R.1,4 & Bradfield G.E.5
1Department of Botany, University of British Columbia, Vancouver, BC, Canada V6T 1Z4; 2Currentaddress: Department of Biology, University of Regina, Regina, SK, Canada S4S 0A2;
3E-mail [email protected]; 4E-mail [email protected]; 5E-mail [email protected];*Corresponding author; Fax +1 3063372410; E-mail [email protected]
Introduction
Invasive species are recognized as a major threat tobiological diversity worldwide, but our understandingof the dynamics of invasion remains limited (Gaston2000; Sala et al. 2000). It is particularly unclear whatmakes a community invasible (Rejmánek 1989;Williamson 1996; Myers & Bazely 2003). Elton (1958)hypothesized that increasing native diversity shouldconfer increasing resistance to invasion, but support forthis has been equivocal (Cornell & Lawton 1992;Stohlgren et al. 1999; Kennedy et al. 2002; Fargione etal. 2003). Disturbance is often considered an importantprecursor to invasion (D’Antonio 1993; Lozon &MacIsaac 1997; Davis et al. 2000) but many undis-turbed native communities host exotic flora. Environ-mental stressors that limit the invader, but not the lo-cally adapted native species, may be important (Harrison1999), although physically stressful habitats are also notfree from the impact of invasive species. Generalitieshave thus emerged on invasibility but the empiricalevidence has yet to definitively support any view inparticular.
A limiting factor for untangling the causes ofinvasibility has been the focus on the most heavilyinvaded areas and the most successful invaders. How-ever, understanding where invasions do not occur is asimportant as where they do, as not all communities areinvaded equally or affected by the same species. Onlyby expanding the examination of invasion beyond highlyaffected areas can we determine if the principles ofinvasion are contingent on particular conditions. Thisnecessitates broadening our investigations both spa-tially and structurally, including the examination ofcommunity-level patterns of invasion along broad envi-ronmental gradients (e.g. Kolb et al. 2002).
We take such an approach in a heavily invaded oaksavanna ecosystem in western Canada, examining howpatterns of plant invasion change in relation to landscape-level environmental heterogeneity. This ecosystem is

48 MacDougall, A.S. et al.
organized along a physical stress gradient of soil depth,elevation, and canopy openness, with shallower soils ofhigher elevation being more moisture-limited and sub-ject to greater solar exposure due to reduced canopycover. Although decreasing soil depth is known to re-duce above-ground biomass production for both nativeand exotic species in this system (MacDougall &Turkington 2005), it is unknown whether the composi-tion and abundance of invasives changes with soil depth,whether such factors also impact native plant diversity,and how soil depth interacts with other environmentalvariables (e.g. elevation, canopy) to shape communitystructure generally.
We test two hypotheses on the environment-inva-sion relationship, asking if areas with harsher physicalconditions are more or less susceptible to invasion.Areas of higher stress may promote an increase in therichness and abundance of invasives by serving as arefuge from the intense above-ground competition thattends to characterize mesic habitats (e.g. Tilman 1988;Keddy 2001; Grime 2001). Alternatively, such environ-ments may limit invasion by requiring highly localizedadaptations that only native species possess (Harrison1999).
Methods
Study system
The Quercus garryana ecosystem occurs from north-ern California to southern British Columbia (Fig. 1A),mostly in the lee of the Coastal Mountain range ofwestern North America (Dunn & Ewing 1997; Maret &Wilson 2000; Fuchs 2001). In British Columbia, theclimate is sub-Mediterranean, and soils are moderatelyinfertile and of post-glacial origin (Roemer 1972). Soildepths range from more than 1 m to less than a few cm(Erickson 2002). Soils tend to become shallower withincreasing elevation, although depth to bedrock can bewidely variable in lower elevation areas. Ground floradiversity is high compared to most other plant commu-nities in the region, supporting 454 vascular plant taxa,of which 144 are naturalized exotic tree, shrub, andherbaceous species. Since the early 19th century thisecosystem has become highly fragmented and is repre-sented today by remnant savanna of varying size sur-rounded by settlements or farms. There are no remnantareas completely free of exotic species. Because manynative plant taxa are now uncommon or rare (>10% oftotal native flora), the oak savanna of southwesternBritish Columbia is a hot spot for rare floral diversityand one of Canada’s most endangered terrestrial ecosys-tems (Anon. 2002).
Survey procedures
The study was done in seven oak savanna remnantsin the Cowichan Valley of southeastern Vancouver Is-land (Fig. 1B). These areas vary in size, soil depth,elevation, and aspect (Table 1). All contain relativelydiverse assemblages of native species. Highly alteredremnants were avoided. Remnant size ranged from 0.3to 18 ha. Five of the sites occur at higher elevations(100-300 m), where shallower soils are generally moreprevalent. Most high-elevation sites have aspects rang-ing from south to west, which maximizes exposure tosun and wind. Less exposed areas (e.g., north-facinghillsides) are dominated by Pseudotsuga menziesii for-est that does not support oak savanna species. Six ofthe sites occur in the eastern end of the CowichanValley (Fig. 1B). The seventh site is 15 km to the west
Fig. 1. A. Main distribution of the Quercus garryana savannaecosystem in British Columbia. B. Location of six remnantoak savanna sites in the Cowichan Valley of SE VancouverIsland. Site 7, not shown, is ca. 15 km to the west of the areadepicted. Shaded areas depict the pre-settlement 1858 andcurrent 2004 distributions.
- How do patterns of plant invasion change along an environmental stress gradient? - 49
on a rock outcrop formation along the Cowichan River.It has become isolated by long-term infilling ofPseudotsuga menziesii in surrounding areas with deepersoils.
Ground flora abundance was determined at eachsite using 1-m2 plots located in savanna openings, oakunderstorey, and transition areas dominated by mix-tures of oak and Pseudotsuga menziesii. The numberof plots per site was proportional to the size of theremnant (range: 20-38 plots/site, n = 184). Plots wereselectively placed to represent opening, understorey,and transition areas; randomly placed plots were notused because they consistently missed rarer micro-habitats and, thus, over-sampled highly abundant andwide-ranging species. In each plot, we visually esti-mated percent cover of all species using a 1-m2 griddivided into 20 cm × 20 cm cells (Armesto & Pickett1985). For each species, cover was summed one cell ata time until the entire grid was covered. Six environ-mental variables were measured for each plot. Aspectand elevation were determined with a GPS unit, accu-rate to 15 m and calibrated at sea level before elevationreadings were taken. Canopy openness was measuredat 150 cm from ground level with a hemispherical lens(Solar Pathfinder), with readings ranging from 0 (fullcanopy cover) to 1 (no canopy). Slope angle wasdetermined with a clinometer. Soil depth to bedrockwas the average of four locations per plot, determinedusing a steel rod driven into the soil. Maximum vegeta-tion height (ground stratum) was measured in each plotas a surrogate for above-ground competitive intensity,recognizing that small differences in stature can havesubstantial impacts on the abundance and reproductionof subordinate species (Givnish 1982; Gaudet & Keddy1988; Grime 2001). Species richness was the numberof species per plot, and species evenness was calcu-lated using Evar (Smith & Wilson 1996).
Data analysis
To determine the relationship between environ-mental variation and the distribution of all native andexotic species, we performed a direct ordination withCanonical Correspondence Analysis (CCA) (PC-ORD- McCune & Mefford 1999). CCA was preferred overother ordination methods (PCA, NMS) because ourmain interest was in determining relationships amongthe environmental variables and plant community pat-tern (ter Braak 1986, Palmer 1993). All environmentalvariables were transformed to improve normality andhomogeneity of variance before analysis. Aspect wascosine transformed following Beers et al. (1966);canopy openness was arc-sin-square-root transformed.Elevation, slope, maximum vegetation height, and soildepth were log-transformed. CCA calculates two scoresets: weighted averaging (WA) scores and linear com-bination (LC) scores. We used WA scores becausethey are less sensitive to variations in community-leveldata that can obscure species-environment relation-ships (McCune & Grace 2002). We identified andtested the significance of potential outliers in the multi-variate space using PC-ORD’s outlier analysis. Eightoutliers were detected and removed. To test the nullhypothesis of no relationship between the species andenvironmental data matrices, we conducted a 999-runMonte Carlo randomization procedure.
To test the impact of environmental variation onvarious compositional and functional indices (e.g. even-ness [Evar], %annuals), we performed regression analy-ses using the environmental variable or variables iden-tified by the CCA as most influential on species distri-butions. Linear fits were used except when a quadraticbetter captured the relationship. Regressions were con-ducted with JMPIN (Anon. 2001).
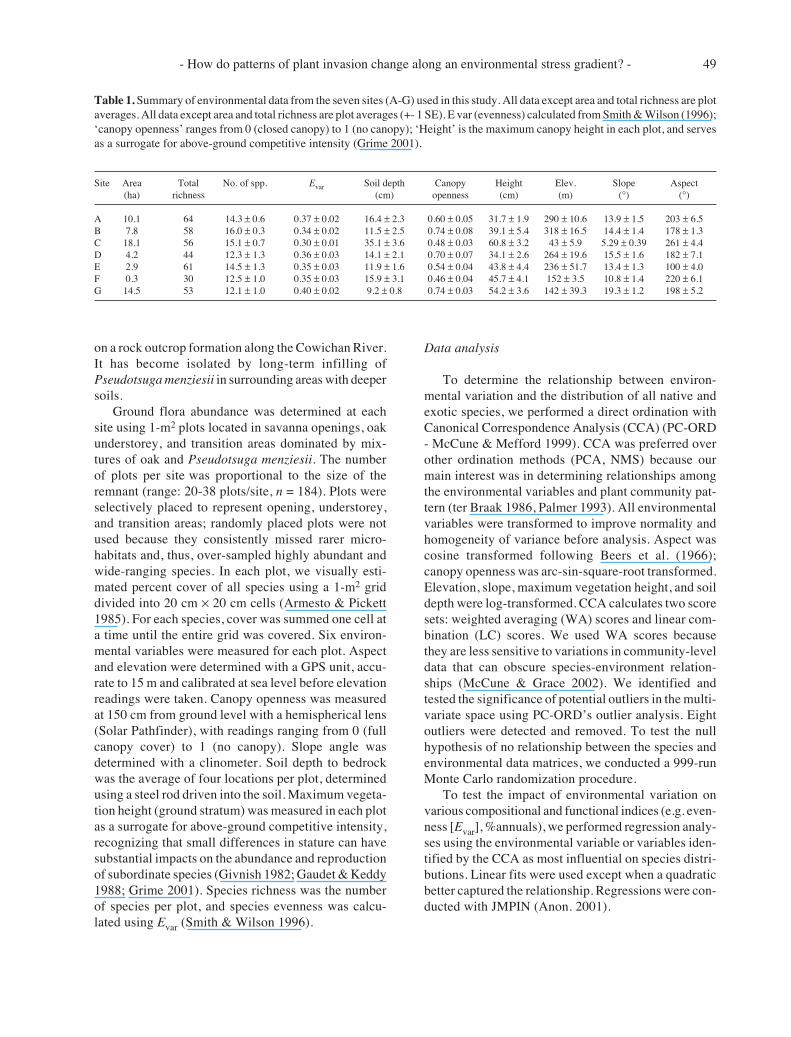
Table 1. Summary of environmental data from the seven sites (A-G) used in this study. All data except area and total richness are plotaverages. All data except area and total richness are plot averages (+- 1 SE). E var (evenness) calculated from Smith & Wilson (1996);‘canopy openness’ ranges from 0 (closed canopy) to 1 (no canopy); ‘Height’ is the maximum canopy height in each plot, and servesas a surrogate for above-ground competitive intensity (Grime 2001).
Site Area Total No. of spp. Evar Soil depth Canopy Height Elev. Slope Aspect(ha) richness (cm) openness (cm) (m) (°) (°)
A 10.1 64 14.3 ± 0.6 0.37 ± 0.02 16.4 ± 2.3 0.60 ± 0.05 31.7 ± 1.9 290 ± 10.6 13.9 ± 1.5 203 ± 6.5B 7.8 58 16.0 ± 0.3 0.34 ± 0.02 11.5 ± 2.5 0.74 ± 0.08 39.1 ± 5.4 318 ± 16.5 14.4 ± 1.4 178 ± 1.3C 18.1 56 15.1 ± 0.7 0.30 ± 0.01 35.1 ± 3.6 0.48 ± 0.03 60.8 ± 3.2 43 ± 5.9 5.29 ± 0.39 261 ± 4.4D 4.2 44 12.3 ± 1.3 0.36 ± 0.03 14.1 ± 2.1 0.70 ± 0.07 34.1 ± 2.6 264 ± 19.6 15.5 ± 1.6 182 ± 7.1E 2.9 61 14.5 ± 1.3 0.35 ± 0.03 11.9 ± 1.6 0.54 ± 0.04 43.8 ± 4.4 236 ± 51.7 13.4 ± 1.3 100 ± 4.0F 0.3 30 12.5 ± 1.0 0.35 ± 0.03 15.9 ± 3.1 0.46 ± 0.04 45.7 ± 4.1 152 ± 3.5 10.8 ± 1.4 220 ± 6.1G 14.5 53 12.1 ± 1.0 0.40 ± 0.02 9.2 ± 0.8 0.74 ± 0.03 54.2 ± 3.6 142 ± 39.3 19.3 ± 1.2 198 ± 5.2
50 MacDougall, A.S. et al.
Results
Composition and species richness
We observed 119 plant species from the seven sites,46 of which were naturalized exotics. Species richnessaveraged 53 species per site (Table 1). There was norelationship between site area and species richness persite (site richness derived by pooling all plots in eachsite; r2 = 0.25, F1,6 = 1.6, p = 0.26). The largest site (SiteC ; Table 1), which has the deepest soil on average, hadthe smallest number of species per unit area reflectingthe absence of a number of native and exotic taxa thatonly occur in sites with shallower soils.
The most abundant exotic functional group was an-nual forbs (38% of all non-native species). Perennialand annual forbs were the most abundant natives (41%and 24% of all native species, respectively). There wereno native annual grasses observed, although eight nativespecies from this functional group occur elsewhere insouthwestern British Columbia (Fuchs 2001). Overall,the most frequently occurring species were the nativeperennial forbs Camassia quamash (50% of all plots),Lomatium utriculatum (46%), and Triteleia hyacinthina(45%). The annual forb Galium aparine was the mostwidely occurring non-native species (56% of all plots)and Cytisus scoparius was the most common exoticwoody species (41%); 74 of the 119 observed specieswere rare, with a plot frequency of < 5%.
Relationship between environmental variation and thedistribution of plots and species
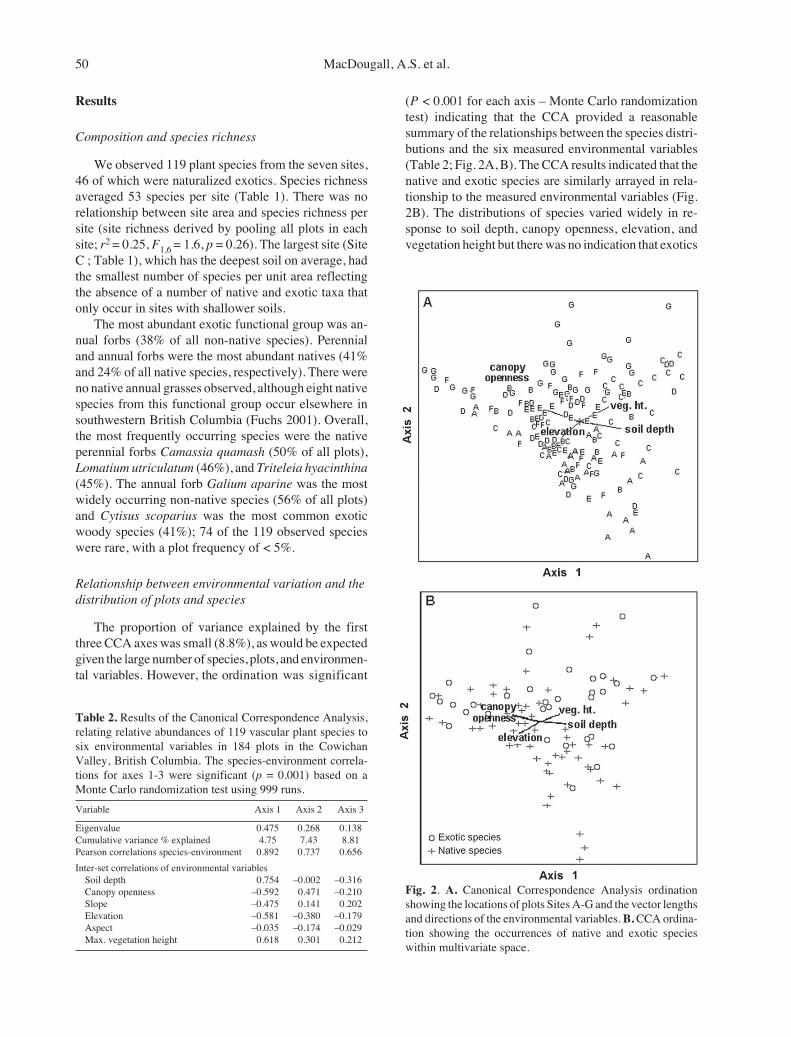
The proportion of variance explained by the firstthree CCA axes was small (8.8%), as would be expectedgiven the large number of species, plots, and environmen-tal variables. However, the ordination was significant
(P < 0.001 for each axis – Monte Carlo randomizationtest) indicating that the CCA provided a reasonablesummary of the relationships between the species distri-butions and the six measured environmental variables(Table 2; Fig. 2A, B). The CCA results indicated that thenative and exotic species are similarly arrayed in rela-tionship to the measured environmental variables (Fig.2B). The distributions of species varied widely in re-sponse to soil depth, canopy openness, elevation, andvegetation height but there was no indication that exotics
Fig. 2. A. Canonical Correspondence Analysis ordinationshowing the locations of plots Sites A-G and the vector lengthsand directions of the environmental variables. B. CCA ordina-tion showing the occurrences of native and exotic specieswithin multivariate space.
Table 2. Results of the Canonical Correspondence Analysis,relating relative abundances of 119 vascular plant species tosix environmental variables in 184 plots in the CowichanValley, British Columbia. The species-environment correla-tions for axes 1-3 were significant (p = 0.001) based on aMonte Carlo randomization test using 999 runs.
Variable Axis 1 Axis 2 Axis 3
Eigenvalue 0.475 0.268 0.138Cumulative variance % explained 4.75 7.43 8.81Pearson correlations species-environment 0.892 0.737 0.656
Inter-set correlations of environmental variablesSoil depth 0.754 –0.002 –0.316Canopy openness –0.592 0.471 –0.210Slope –0.475 0.141 0.202Elevation –0.581 –0.380 –0.179Aspect –0.035 –0.174 –0.029Max. vegetation height 0.618 0.301 0.212
- How do patterns of plant invasion change along an environmental stress gradient? - 51
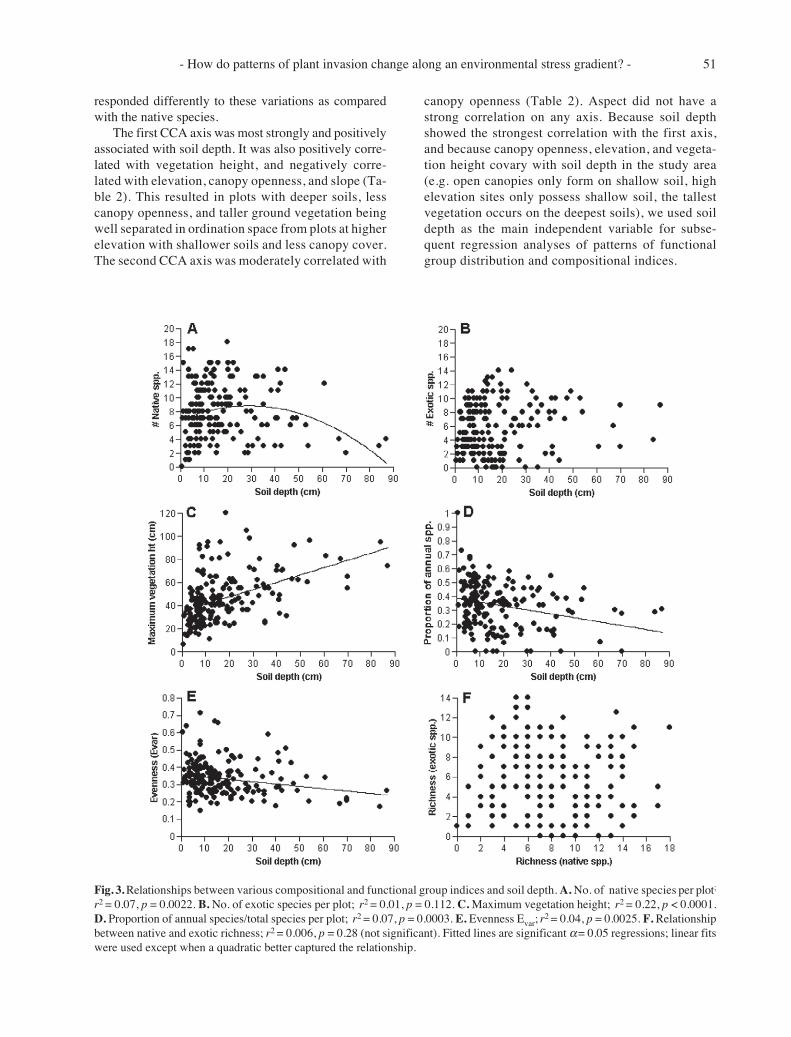
Fig. 3. Relationships between various compositional and functional group indices and soil depth. A. No. of native species per plot;
r2 = 0.07, p = 0.0022. B. No. of exotic species per plot; r2 = 0.01, p = 0.112. C. Maximum vegetation height; r2 = 0.22, p < 0.0001.D. Proportion of annual species/total species per plot; r2 = 0.07, p = 0.0003. E. Evenness Evar;
r2 = 0.04, p = 0.0025. F. Relationshipbetween native and exotic richness; r2 = 0.006, p = 0.28 (not significant). Fitted lines are significant α = 0.05 regressions; linear fitswere used except when a quadratic better captured the relationship.
responded differently to these variations as comparedwith the native species.
The first CCA axis was most strongly and positivelyassociated with soil depth. It was also positively corre-lated with vegetation height, and negatively corre-lated with elevation, canopy openness, and slope (Ta-ble 2). This resulted in plots with deeper soils, lesscanopy openness, and taller ground vegetation beingwell separated in ordination space from plots at higherelevation with shallower soils and less canopy cover.The second CCA axis was moderately correlated with
canopy openness (Table 2). Aspect did not have astrong correlation on any axis. Because soil depthshowed the strongest correlation with the first axis,and because canopy openness, elevation, and vegeta-tion height covary with soil depth in the study area(e.g. open canopies only form on shallow soil, highelevation sites only possess shallow soil, the tallestvegetation occurs on the deepest soils), we used soildepth as the main independent variable for subse-quent regression analyses of patterns of functionalgroup distribution and compositional indices.
52 MacDougall, A.S. et al.
Relationship between soil depth and plant communityparameters
Total species richness per plot (native and exotic)was not associated with soil depth (F1,183 = 0.17, p =0.68). By contrast, there was a significant curvilinearrelationship (F2,183 = 4.59, p = 0.013) (Fig. 3A) betweennative richness and soil depth, with richness declining inthe shallowest and deepest soils. This was explained byextremely shallow soil (< 3 cm) plots having mostlybryophytes and invasive annual grasses (Aira spp.),while deeper soil (> 50 cm) plots were dominated byinvasive perennial grasses (Poa pratensis, Dactylisglomerata, Alopecurus pratensis). On deeper soils, totalcover by the invasive grasses averaged > 65% per plot.The few native species that occurred on deeper soilswere functionally identical to the dominant invasiveperennials, and included the tall-statured (>100 cm
height) native perennial grasses Elymus glaucus, Bromuscarinatus, Bromus sitchensis, Melica subulata, andFestuca rubra. Total cover by these native grasses aver-aged > 10% per plot on deeper soils. There was norelationship between exotic species richness and soildepth (F1,183 = 2.55, p = 0.112) (Fig. 3B).
Declines in native species richness with soil depthwere correlated with significant increases in maximumvegetation height (Fig. 3C) and the occurrence of exoticperennial grasses (Fig. 4). There was a significant de-cline in the percentage of annual species per plot (nativeand exotic) with soil depth (F1,183 = 13.52, p = 0.0003)(Fig. 3D). The increasing dominance by the perennialgrasses with increasing soil depth was indicated by thesignificant decline of evenness as the soils becamedeeper (F1,183 = 9.39, p = 0.0025) (Fig. 3E). No relation-ship was detected between native species richness andthe number of exotic species (F1,183 = 1.18, p = 0.279)
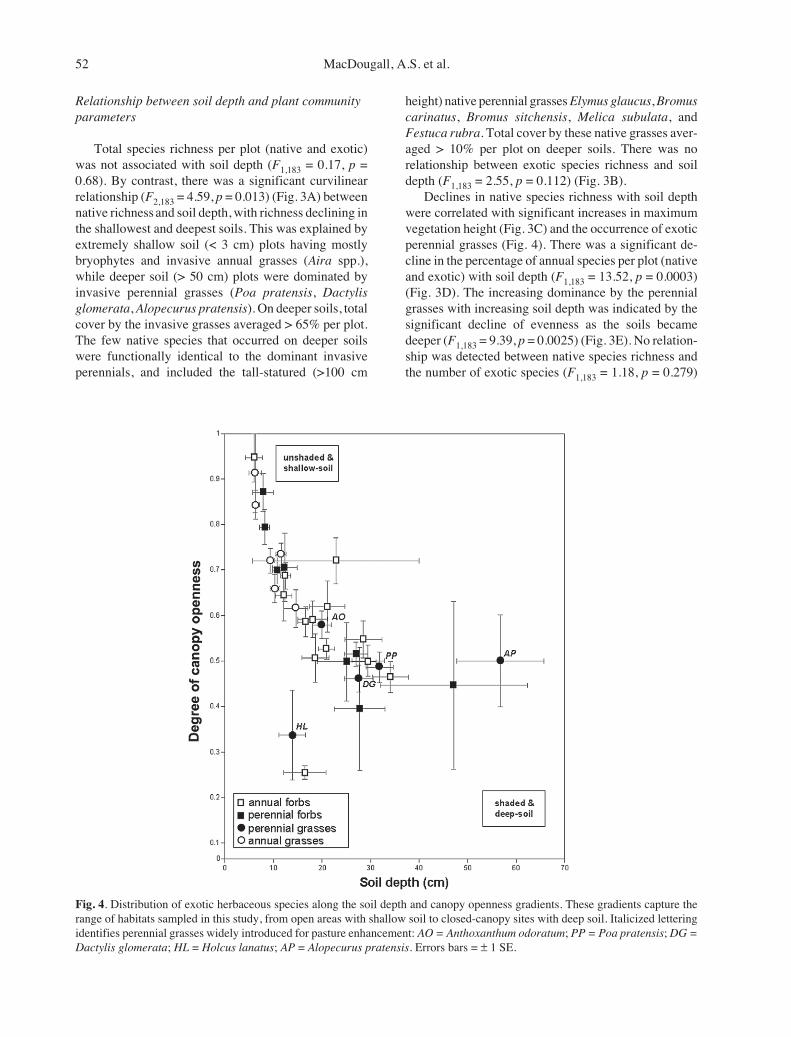
Fig. 4. Distribution of exotic herbaceous species along the soil depth and canopy openness gradients. These gradients capture therange of habitats sampled in this study, from open areas with shallow soil to closed-canopy sites with deep soil. Italicized letteringidentifies perennial grasses widely introduced for pasture enhancement: AO = Anthoxanthum odoratum; PP = Poa pratensis; DG =Dactylis glomerata; HL = Holcus lanatus; AP = Alopecurus pratensis. Errors bars = ± 1 SE.

- How do patterns of plant invasion change along an environmental stress gradient? - 53
(Fig. 3F). We also detected no relationship between soildepth and the cover of leguminous species (F1,183 =0.004, p = 0.95), lessening the possibility of an effectdue to N-fixation and soil fertility.
The inverse correlation between plant height andnative species richness could conceivably be an artifactof plant size – plots on deeper soils have larger plants sothere should be fewer individuals per plot and, by impli-cation, fewer species (Oksanen 1996). To test this, werandomly pooled the 1-m2 plot data into larger samplesof 2 m2, 4 m2, and 8 m2 for every 10 cm of soil depth (e.g.0-10 cm, 11-20 cm) (Rapson et al. 1997). If plant sizeexplains diversity in the 1-m2 plots, then diversity shouldincrease on deeper soils as plot area increases. This didnot occur. Native species richness declined significantlywith soil depth in all cases (2 m2: r2 = 0.88, F2,6 = 11.20,p = 0.035; 4 m2: r2 = 0.81, F2,6 = 9.7, p = 0.047; 8 m2: r2
= 0.86, F2,6 = 12.7, p = 0.018).There was an almost complete change in the compo-
sition of exotic species from open sites with shallow soilto closed canopy sites with deeper soil (Fig. 4). Al-though exotic perennial grasses dominate many oaksavanna remnants in British Columbia, their distribu-tions were skewed significantly towards deeper soils(Fig. 4) (F1,183 = 46.4, p < 0.0001; Tukey’s HSD test).Shallow soils were mostly invaded by annual grasses(Aira spp., Vulpia bromoides, Bromus spp.), annualforbs (Galium aparine, Trifolium dubium, and Myosotisdiscolor), and a small number of perennial forbs mostlyfrom the Asteraceae family (e.g. Crepis capillaris,Hypochaeris radicata) (Fig. 4).
Discussion
General trends
The environmental stress gradient of this savannaecosystem strongly influenced patterns of plant inva-sion but not in ways that were predicted. There was nochange in the number of invasive species with environ-mental variation or with the richness of native species.However, the identity of the invaders changed withincreasing soil depth, as did their apparent impact onnative diversity. There was an almost complete turnoverin exotic species composition from shallow to deepsoils. Annual grasses, annual forbs, and a few perennialforbs mostly dominated shallow soils, while perennialgrasses dominated deeper soils. This pattern of turnover(i.e. ß-diversity) generally matched changes to nativespecies composition along the gradient (for instancefewer native annuals, more native tall-statured peren-nial grasses in deeper soils). There also appeared to besubstantial differences in the impacts of the invaders on
native plant diversity. As soil depth increased, the numberand abundance of native species declined in associationwith increasing dominance by a small number of inva-sive perennial grasses (Poa, Dactylis, Alopecurus). Thus,the hypothesis that invasibility is determined by envi-ronmental stress was not supported. Although the soildepth gradient affected the identity of the invaders andtheir likely impacts on native diversity, all areas weresusceptible to establishment by exotic taxa.
The most likely mechanism underlying these pat-terns is increasing above-ground competition with in-creasing soil depth. Plots with soils > 50 cm deep arecharacterized by tall and dense grass swards, with lightlevels < 2% full light at ground level, and by signifi-cantly higher above-ground biomass (MacDougall &Turkington 2005). Experimental disturbance treatmentsand seed additions have demonstrated that many exoticand native species that are currently limited to shallowersoils are fully capable of establishing and regeneratingin the absence of these grasses (MacDougall 2005). Itappears, therefore, that the segregation of species be-tween deep and shallow soils is driven in part by intensecompetition in more mesic areas, similar to observa-tions from other systems (Terborgh 1973; Gurevitch1986; Baskin & Baskin 1988; Keddy 2001). Theserelatively mesic areas are not more invasible per se butmore susceptible to invasion by species with strongcompetitive impacts on community structure, as hasbeen observed elsewhere (D’Antonio 1993; Milberg etal. 1999; Larson et al. 2001; Meekins & McCarthy2001; Kolb et al. 2002). Although more xeric habitats(shallow soils) were equally invaded, it does not appearthat exotic species in these areas are limiting the distri-bution or abundance of co-occurring native species.
These results beg the question of why the impact ofexotic species appears to diminish in shallower soils.Does the competitive effect of the exotics decrease withincreasing stress, possibly due to a home-site advantageby the locally adapted natives? Or does competitionbecome less relevant due to the highly limiting physicalconditions (extreme summer moisture deficits and solarexposure) that characterize these areas? The shallowsoils do appear to restrict or prevent the growth of theexotic perennial grasses, likely because these speciesflower in mid-summer when moisture availability canbe close to nil (except Anthoxanthum, which flowersMarch-May). Many native species are adapted pheno-logically and physiologically to these conditions, finish-ing their annual reproductive cycle by late spring (May)or by possessing tolerance mechanisms to low moistureand high temperatures (e.g. Opuntia, Sedum).
However, local adaptations are unlikely to be theonly cause of reduced exotic impact. Numerous invad-ers in west-coast oak savanna ecosystems of North

54 MacDougall, A.S. et al.
America are known to proliferate in conditions similarto the shallow soils of British Columbia (e.g. Bromustectorum, Centaurea spp.). This suggests that recruit-ment barriers (geographic or fortuitous), climatic thresh-olds (too far north), or the variety or subspecies thathappened to be introduced could partially explain present-day invasion patterns. Invasiveness and the impacts ofinvasives on native species, therefore, may be unpre-dictable based solely on factors such as native diversityor local environmental conditions. The traits and com-petitive strategies possessed by the arriving pool ofinvaders, and the historical circumstances underlyingtheir arrival, may also be relevant for determining whereinvasion occurs and its level of ecological impact.
Trait similarities between exotic and native species
Our results suggest that the functional characteris-tics of native species in any given habitat (e.g. tallperennial grasses in deep soils) tend to predict the traitsof the invaders of those same areas. In deep soils, exoticperennial grasses currently dominate areas where simi-larly robust native grasses (e.g. Elymus glaucus, Bromuscarinatus) most commonly occur. Native and exoticannual forbs tend to occupy shallow soils where exoticperennial grasses are absent. Levels of invasion by theexotic shrub Cytisus scoparius are matched by expan-sion of the native shrub Symphoricarpos albus in somelocations. This result is contrary to recent experimentalwork demonstrating that invasion success is likely tofavour species that are functionally distinct from thosein the recipient community (Fargione et al. 2003; Godfreeet al. 2004).
In our study area, there is a strong negative associa-tion between the occurrence of exotic perennial grassesand native forbs, while the association between theoccurrence of exotic and native perennial grasses isstrongly positive. Although the native grasses are lessabundant than the invaders, they possess a range of lifehistory strategies (e.g., larger seed mass, rapid seedlinggrowth) that allows them to co-exist (MacDougall &Turkington 2004). The experimental removal of thedominant exotic grasses confirms the importance offunctional dissimilarity for invasion in this ecosystem –the species that respond greatest to the removals arethose most functionally distinct (annuals, forbs, shrubs)(MacDougall & Turkington 2005). The causes of thesepatterns are unclear but we suggest they further high-light the significance of environmental filters for deter-mining invasibility. It appears that invasion is a two-tierprocess in this system, where environmental stressorsfirst determine community membership (exotic and na-tive), and competitive interactions then determine rela-tive abundance among species that pass through these
filters. What remains unclear is why exotics emerge asthe most abundant species within any given habitat, towhich numerous hypotheses have been proposed(Williamson 1996; Callaway & Aschehoug 2000; Macket al. 2000; Myers & Bazely 2003).
Conservation implications
Our results indicate that the effectiveness and ur-gency of invasive species management are likely to varyalong environmental gradients, highlighting the needfor ecosystem-based approaches to invasion control. Inthis oak savanna ecosystem, controlling invasives ondeeper soils is a priority due to their apparent competi-tive impacts on native flora. Of the 59 plant specieslisted as provincially or nationally endangered in thisecosystem, 27 are associated with deeper soils wherethe effects of invasive species are undoubtedly exacer-bated by habitat loss and long-term fire suppression(MacDougall et al. 2004).
Complicating the control of deep-soil exotics, how-ever, is the large and functionally diverse pool of non-native species that occurs in the region. Many exoticscurrently found on shallow soils proliferate followingperennial grass removal in areas with deeper soils(MacDougall & Turkington 2005). Exotics such asBromus sterilis and Trifolium dubium, for example,dominate the deep-soil seed bank and increase threefoldor more following disturbance. The shrub Cytisusscoparius similarly proliferates with the removal of theinvasive grasses. Although some native taxa also re-spond positively to these changes (Cardamine oligo-spora), exotic species typically dominate due to higherpropagule pressure, longer seed viability, or faster growthrates in post-disturbance contexts (e.g., MacDougall &Turkington 2004). It seems, therefore, that there is nosimple strategy for controlling exotics in this system.For every management action, there appears to be one ormore species capable of aggressively responding to itseffects. Managers must thus target the most problematicspecies and also account for the likely after-effects oftheir removal, including the possible emergence of dif-ferent but similarly pernicious invaders that are pres-ently confined to areas of higher environmental stress.
Acknowledgements. Funding was provided by the NaturalSciences and Engineering Research Council of Canada, WorldWildlife Fund, Environment Canada, and MEC. We thankBrenda Costanzo and Hans Roemer for species identifica-tions, and Kate Edwards for field assistance. Significant con-tributions were provided by The Nature Conservancy of Canadaand the Garry Oak Ecosystem Recovery Team (GOERT).Thanks also to Tim Ennis, Irvin Banman, Matt Fairbarns,Wayne Erickson and Wayne Hallstrom.

- How do patterns of plant invasion change along an environmental stress gradient? - 55
Givnish, T.J. 1982. On the adaptive significance of leaf heightin forest herbs. Am. Nat. 120: 353-381.
Godfree, R., Lepschi, B. & Mallinson, D. 2004. Ecologicalfiltering of exotic plants in an Australian sub-alpine envi-ronment. J. Veg. Sci. 15: 227-236.
Grime, J.P. 2001. Plant strategies, vegetation processes, andecosystem properties. 2nd. ed. John Wiley, Chichester,UK.
Gurevitch, J. 1986. Competition and the local distribution ofthe grass Stipa neomexicana. Ecology 67: 46-57.
Harrison, S. 1999. Native and alien species diversity at thelocal and regional scales in a grazed California grassland.Oecologia 121: 99-106.
Keddy, P. 2001. Competition. Kluwer Academic Press, Bos-ton, MA, US.
Kennedy, T.A., Naeem, S., Howe, K.M., Knops, J.M.H.,Tilman, D. & Reich, P. 2002. Biodiversity as a barrier toecological invasion. Nature 417: 636-638.
Kolb, A., Alpert, P., Enters, D. & Holzapfel, C. 2002. Patternsof invasion within a grassland community. J. Ecol. 90:871-881.
Larson, D.L., Anderson, P.J. & Newton, W. 2001. Alien plantinvasion in mixed-grass prairie: effects of vegetation typeand anthropogenic disturbance. Ecol. Appl. 11: 128-141.
Lozon, J.D. & MacIssac, H.J. 1997. Biological invasions: arethey dependent on disturbance? Environ. Rev. 5: 131-144.
MacDougall, A.S. 2005. Responses of diversity and invasibilityto burning in a northern oak savanna. Ecology 86: 3354-3363.
MacDougall, A.S. & Turkington, R. 2004. Relative impor-tance of suppression-based and tolerance-based competi-tion in an invaded oak savanna. J. Ecol. 92: 422-434.
MacDougall, A.S. & Turkington, R. 2005. Are exotic speciesthe drivers or passengers of ecological change in highlydisturbed plant communities? Ecology 86: 42-55.
MacDougall, A.S., Beckwith, B.R. & Maslovat, C. 2004.Defining conservation strategies with historical perspec-tives: a case study from a degraded oak grassland ecosys-tem. Conserv. Biol. 18: 455-465.
Mack, R.N., Simberloff, D., Lonsdale, W.M., Evans, H., Clout,M. & Bazazz, F.A. 2000. Biotic invasions: causes, epide-miology, global consequences, and control. Ecol. Appl.10: 689-710.
Maret, M.P. & Wilson, M.V. 2000. Fire and seedling popula-tion dynamics in western Oregon prairies. J. Veg. Sci. 11:307-314.
McCune, B. & Grace, J.B. 2002. Analysis of ecological com-munities. MjM, Gleneden Beach, OR, US.
McCune, B. & Medford, M.J. 1999. PC-ORD: multivariateanalysis of ecological data. Version 4.17. MjM SoftwareDesign, Gleneden Beach, OR, US.
Meekins, J.F. & McCarthy, B.C. 2001. Effect of environmen-tal variation on the invasive success of a non-indigenousforest herb. Ecol. Appl. 11: 1336-1348.
Milberg, P., Lamont, B.B. & Perez-Fernandez, M.A. 1999.Survival and growth of native and exotic composites inresponse to a nutrient gradient. Plant Ecol. 145: 125-132.
Myers, J.H. & Bazely, D.R. 2003. Ecology and control ofintroduced plants. Cambridge University Press, Cam-
References
Anon. 2001. JMP start statistics. 2nd. ed. SAS Duxbury,Pacific Grove, CA, US.
Anon. 2002. Recovery strategy for Garry Oak and associatedecosystems and their associated species at risk in Canada2001-2006. GOERT (Garry Oak Ecosystem RecoveryTeam), Victoria, BC, CA.
Armesto, J.J. & Pickett, S.T.A. 1985. Experiments on distur-bance in old-field plant communities: impact on speciesrichness and abundance. Ecology 66: 230-240.
Baskin, J.M. & Baskin, C. 1988. Endemism in rock outcropplant communities of the unglaciated eastern United States:an evaluation of the roles of edaphic, genetic and lightfactors. J. Biogeogr. 15: 829-840.
Beers, T.W., Dress, P.E. & Wensel, L.C. 1966. Aspect trans-formation in site productivity research. J. For. 64: 691:692.
Callaway, R.M. & Aschehoug, E.T. 2000. Invasive plantsversus their new and old neighbors: a mechanism forexotic invasion. Science 290: 521-523.
Cornell, H.V. & Lawton, J.H. 1992. Species interactions, localand regional processes, and limits to the richness of eco-logical communities: a theoretical perspective. J. Anim.Ecol. 61: 1-12.
D’Antonio, C.M. 1993. Mechanisms controlling invasion ofcoastal plant communities by the alien succulent Carpo-brotus edulis. Ecology 74: 83-95.
Davis, M.A., Grime, J.P. & Thompson, K. 2000. Fluctuatingresources in plant communities: a general theory ofinvasibility. J. Ecol. 88: 528-534.
Douglas, G.W., Straley, G.B., Meidinger, D. & Pojar, J. 2001.Illustrated flora of British Columbia. Vols. 1-8. Ministryof Environment, Lands and Parks and Ministry of Forests.Victoria, BC, CA.
Dunn, P. & Ewing, K. 1997. Ecology and conservation of theSouth Puget Sound Prairie landscape. The Nature Con-servancy of Washington, Seattle, WA, US.
Elton, C.S. 1958. The ecology of invasions by animals andplants. University of Chicago Press, Chicago, IL, US.
Erickson, W.R. 2002. Environmental relationships of nativeGarry Oak (Quercus garryana) communities at their north-ern margin. In: Standiford, R.B., McCreary, D. & Purcell,K.L. (eds). Proceedings on the fifth symposium on oakwoodlands: oaks in California’s changing landscape, pp.179-190 USDA, Albany, CA, US.
Fargione, J., Brown, C.S. & Tilman, D. 2003. Communityassembly and invasion: an experimental test of neutralversus niche processes. Proc. Natl. Acad. Sci. USA 100:8916-8920.
Fuchs, M. 2001. Towards a recovery strategy for Garry oakand associated ecosystems in Canada: ecological assess-ment and literature review. Technical report GBEI/EC-00-030. Environment Canada, Canadian Wildlife Service,Ladner, BC, CA.
Gaston, K.J. 2000. Global patterns in biodiversity. Nature405: 220-227.
Gaudet, C.L. & Keddy, P.A. 1988. A comparative approach topredicting competitive ability from plant traits. Nature334: 242-243.

56 MacDougall, A.S. et al.
bridge, UK.Oksanen, J. 1996. Is the humped relationship between species
richness and biomass an artifact due to plot size? J. Ecol.84: 293-295.
Palmer, M.W. 1993. Putting things in even better order: theadvantages of canonical correspondence analysis. Ecol-ogy 74: 2215-2230.
Rapson, G.L., Thompson, K. & Hodgson, J.G. 1997. Thehumped relationship between species richness and bio-mass – testing its sensitivity to sample quadrat size. J.Ecol. 85: 99-100.
Rejmánek, M. 1989. Invasibility of plant communities. InDrake, J.A., DiCastri, F., Groves, R.H., Kruger, F.J.,Mooney, H.A., Rejmánek, M. & Williamson, M.H. (eds.)Ecology of biological invasion: a global perspective. Wiley,New York, NY, US.
Roemer, H.L. 1972. Forest vegetation and environments onthe Saanich Peninsula. Ph.D. Thesis, University of Victo-ria, Victoria, BC, CA.
Sala, O.E., Chapin, F.S., Armesto, J.J. et al. 2000. Global
biodiversity scenarios for the year 2100. Science 287:1770-1774.
Smith, B. & Wilson, J.B. 1996. A consumer’s guide to even-ness indices. Oikos 76: 70-82.
Stohlgren, T.J., Binkley, D., Chong, G.W., Kalkhan, M.A.,Schell, L.D., Bull, K.A., Otsuki, Y., Newman, G., Bashkin,M. & Son, Y.1999. Exotic plant species invade hot spotsof native plant diversity. Ecol. Monogr. 69: 25-46.
ter Braak, C.J.F. 1986. Canonical correspondence analysis: anew eigenvector technique for multivariate direct gradientanalysis. Ecology 67: 1167-1179.
Terborgh, J. 1973. On the notion of favourableness in plantecology. Am. Nat. 107: 481-501.
Tilman, D. 1988. Plant strategies and the dynamics andstructure of plant communities. Princeton University Press,Princeton, NJ, US.
Wedin, D.A & Tilman, D. 1990. Species effects on nitrogencycling: a test with perennial grasses. Oecologia 84: 433-441.
Williamson, M. 1996. Biological invasions. Chapman & Hall,London, UK.
Received 2 February 2005;Accepted 2 November 2005.
Co-ordinating Editor: V.D. Pillar.
Copyright © 2022 FDOKUMEN




















