
Patterns of mangrove forest disturbance and biomass removal ...
17
ORIGINAL PAPER Patterns of mangrove forest disturbance and biomass removal due to small-scale harvesting in southwestern Madagascar Ivan R. Scales . Daniel A. Friess Received: 25 November 2018 / Accepted: 8 July 2019 / Published online: 18 July 2019 Ó The Author(s) 2019 Abstract Informal small-scale mangrove wood har- vesting has received limited attention, though it is a widespread threat to mangroves in many parts of the tropics. We investigated wood use and the impacts of harvesting on mangrove forests in the Bay of Assas- sins in southwest Madagascar. We measured forest structure, composition, and harvesting across 60 vegetation plots and investigated human uses of the mangroves through Rapid Rural Appraisal techniques. We found that unlike other mangroves in the region, those in the Bay of Assassins are dominated by Ceriops tagal. Tree harvesting rates are high, with a mean of 28.7% (SD 19.4) of trees harvested per plot. This is similar to heavily harvested mangroves in other parts of the tropics. A comparison of tree versus sapling importance of the different mangrove tree species indicates that the composition of the mangrove forest is changing, with C. tagal becoming more important. Livelihood activities drive the harvesting of certain species and size classes. Mangrove wood is used mainly for the construction of traditional housing and fencing. There are also emerging uses of man- grove wood, including seaweed (Kappaphycus alvarezii) aquaculture and the production of ‘sokay’, a lime render made by burning sea shells in mangrove wood kilns and used to improve the durability of houses. Small-scale selective harvesting of mangrove wood is important for local livelihoods but may have wide-ranging impacts on forest composition and structure. Demand for mangrove wood has grown in relation to new commodity chains for marine products, demonstrating the need for integrated landscape management that considers wetland, terrestrial and marine resources together. Keywords Anthropogenic disturbance Deforestation Degradation Forest–poverty linkages Provisioning ecosystem services Introduction Intertidal mangrove forests are globally threatened (Polidoro et al. 2010; Hamilton and Casey 2016) by a range of pressures such as aquaculture and agriculture, urban development, harvesting for paper pulp, fuel- wood, charcoal and construction materials, changes in hydrology and sediment budgets, and sea-level rise (UNEP 2014). There is considerable geographical variation in the processes that lead to mangrove deforestation and degradation, with threats differing in both scale and type. Aquaculture has been the most I. R. Scales (&) St Catharine’s College, Trumpington Street, Cambridge CB2 1RL, UK e-mail: [email protected] D. A. Friess Department of Geography, National University of Singapore, 1 Arts Link, Singapore 117570, Singapore 123 Wetlands Ecol Manage (2019) 27:609–625 https://doi.org/10.1007/s11273-019-09680-5
-
Upload
khangminh22 -
Category
Documents
-
view
3 -
download
0
Transcript of Patterns of mangrove forest disturbance and biomass removal ...

ORIGINAL PAPER
Patterns of mangrove forest disturbance and biomassremoval due to small-scale harvesting in southwesternMadagascar
Ivan R. Scales . Daniel A. Friess
Received: 25 November 2018 / Accepted: 8 July 2019 / Published online: 18 July 2019
� The Author(s) 2019
Abstract Informal small-scale mangrove wood har-
vesting has received limited attention, though it is a
widespread threat to mangroves in many parts of the
tropics. We investigated wood use and the impacts of
harvesting on mangrove forests in the Bay of Assas-
sins in southwest Madagascar. We measured forest
structure, composition, and harvesting across 60
vegetation plots and investigated human uses of the
mangroves through Rapid Rural Appraisal techniques.
We found that unlike other mangroves in the region,
those in the Bay of Assassins are dominated by
Ceriops tagal. Tree harvesting rates are high, with a
mean of 28.7% (SD 19.4) of trees harvested per plot.
This is similar to heavily harvested mangroves in other
parts of the tropics. A comparison of tree versus
sapling importance of the different mangrove tree
species indicates that the composition of the mangrove
forest is changing, with C. tagal becoming more
important. Livelihood activities drive the harvesting of
certain species and size classes. Mangrove wood is
used mainly for the construction of traditional housing
and fencing. There are also emerging uses of man-
grove wood, including seaweed (Kappaphycus
alvarezii) aquaculture and the production of ‘sokay’,
a lime render made by burning sea shells in mangrove
wood kilns and used to improve the durability of
houses. Small-scale selective harvesting of mangrove
wood is important for local livelihoods but may have
wide-ranging impacts on forest composition and
structure. Demand for mangrove wood has grown in
relation to new commodity chains for marine products,
demonstrating the need for integrated landscape
management that considers wetland, terrestrial and
marine resources together.
Keywords Anthropogenic disturbance �Deforestation � Degradation � Forest–poverty
linkages � Provisioning ecosystem services
Introduction
Intertidal mangrove forests are globally threatened
(Polidoro et al. 2010; Hamilton and Casey 2016) by a
range of pressures such as aquaculture and agriculture,
urban development, harvesting for paper pulp, fuel-
wood, charcoal and construction materials, changes in
hydrology and sediment budgets, and sea-level rise
(UNEP 2014). There is considerable geographical
variation in the processes that lead to mangrove
deforestation and degradation, with threats differing in
both scale and type. Aquaculture has been the most
I. R. Scales (&)
St Catharine’s College, Trumpington Street,
Cambridge CB2 1RL, UK
e-mail: [email protected]
D. A. Friess
Department of Geography, National University of
Singapore, 1 Arts Link, Singapore 117570, Singapore
123
Wetlands Ecol Manage (2019) 27:609–625
https://doi.org/10.1007/s11273-019-09680-5(0123456789().,-volV)( 0123456789().,-volV)

important human activity leading to mangrove loss
globally (Thomas et al. 2017), particularly driving
mangrove change in Southeast Asia and Latin Amer-
ica (Walters et al. 2008). In sub-Saharan Africa,
harvesting for fuelwood and charcoal production are
growing threats (Feka and Ajonina 2011; UNEP 2014;
Feka 2015).
Mangrove deforestation and degradation in low-
income nations is of particular concern, as poor rural
communities rely heavily on a wide range of ecosys-
tem services provided by mangrove forests. These
include the direct provisioning of building materials,
fuelwood, charcoal, animal fodder, (shell)fish and
non-timber forest products (Bandaranayake 1998;
Dahdouh-Guebas et al. 2000; Walters 2005a; Barbier
et al. 2011; Lau and Scales 2016), alongside the
benefits that these communities gain from regulating
services (e.g., coastal protection) and cultural services
(e.g., recreation, aesthetic and spiritual values).
While communities benefit from ecosystem ser-
vices, the extraction of provisioning services can have
negative impacts on the system providing them.
Informal small-scale wood harvesting is one of the
most widespread forms of resource use in mangrove
forests (e.g. Rajkaran et al. 2004; Walters 2005a, b;
Walters et al. 2008). This is especially the case in sub-
Saharan Africa (Feka and Ajonina 2011). However,
few studies of informal mangrove harvesting have
been published. Furthermore, research on the impacts
of mangrove harvesting has mostly focused on mea-
suring mangrove cover loss through forest clearance
rather than looking at other forms of human use and
disturbance. This is in part because, unlike forest
clearance, more cryptic forms of degradation caused
by harvesting and other small-scale forest disturbances
can be difficult to detect using remote sensing methods
(Dahdouh-Guebas et al. 2005). Inaccessibility and
poor infrastructure make mangrove monitoring
through field surveys difficult and time-consuming
(Feka and Morrison 2017).
Madagascar has 2% of the total global mangrove
forest cover by area (Giri and Mulhausen 2008). With
a mean per capita daily income of $1.50, it is one of the
poorest countries in the world (World Bank 2016). As
a result, rural communities are heavily reliant on the
provisioning services of ecosystems, including man-
groves, for their basic needs (Harris 2011). This
potentially has implications for the long-term
sustainability of these ecosystems, and highlights the
importance of sustainable forest management.
We take a multidisciplinary approach to investigate
the patterns and use of small-scale harvesting on
mangrove forests in the Bay of Assassins in south-
western Madagascar, a remote region where man-
groves are used by largely subsistence-based, artisanal
fishing communities (Scales et al. 2018). First, we
characterize mangrove forest composition and struc-
ture throughout the Bay. Second, we quantify patterns
of harvesting pressure on the mangrove system and the
volume of biomass removed. Third, we investigate
household use of mangrove wood. We finish by
considering the management implications of current
mangrove uses and their impacts in the Bay of
Assassins.
Methods
Study site description
Madagascar’s mangroves covered an area of
2797 km2 in 2005 (Giri and Mulhausen 2008),
distributed primarily in bays and inlets along the
sheltered west coast. Madagascar is home to nine
species of mangrove tree (Spalding et al. 2010). A
substantial rainfall gradient exists along the western
coast, with the arid southwest of Madagascar receiving
approximately 400 mm of rainfall per year, and the
wetter northwest coast receiving approximately
2000 mm per year. This influences mangrove forest
structure along the coast, with trees of larger stature
found in mangroves in the northwest bays of Ambaro,
Ambanja and Mahajamba compared to the southwest
(Hutchison et al. 2014; Jones et al. 2016). Mangrove
biomass carbon averages 146.8 Mg C ha-1 in the
northwest (Jones et al. 2014), but only 46.2–73.9 Mg
C ha-1 in the southwest (Benson et al. 2017).
This study was conducted in the Bay of Assassins
(Helodrano Fagnemotse in Malagasy), a coastal inlet
in southwestern Madagascar (22�120S, 43�160N),
180 km north of the regional capital of Toliara
(Fig. 1). The climate is semi-arid, with approximately
400 mm rainfall per year. The inlet is fringed by c.
1300 ha of mangrove forest (Jones et al. 2016). There
are 10 villages around the bay, with a total population
of approximately 3000 (Peabody and Jones 2013).
With an average daily per capita income of less than
123
610 Wetlands Ecol Manage (2019) 27:609–625

US$1.50, fishing communities in southwestern Mada-
gascar are some of the country’s most isolated and
marginalized (Harris 2011). Small-scale fishing gen-
erates over 80% of household income in these
communities and provides the majority of dietary
protein (Barnes-Mauthe et al. 2013).
Field measurements of forest characteristics
and harvested status
Our vegetation survey was based on a modified
version of a mangrove forest structure and biomass
assessment protocol set out by Kauffman and Donato
(2012). We conducted measurements of mangrove
canopy cover, species composition, forest structure,
and harvesting in 60 circular plots with a seven metre
radius (covering an area of 153.94 m2). Plot locations
were randomly selected around the Bay of Assassins
to minimize sampling biases due to edge effects or
distance from human habitation. A random number
generator was used to select distances and angles
between sampling points. This sampling design also
ensured that we randomly covered all representative
forest types across the bay.
We recorded the latitude and longitude of each plot
using a handheld Global Positioning System. The
Euclidean distance of each plot to the forest edge and
the nearest settlement were calculated in ArcGIS. We
estimated canopy cover in four cardinal directions at
the centre of each plot using a concave spherical
canopy densitometer (Lemmon 1956). In each plot we
counted and identified all trees (non-harvested and
harvested) to species level, as well as measured
diameter at breast height (DBH). We defined trees as
woody mangrove vegetation with a DBH of at least
5 cm and defined saplings as woody mangrove
vegetation with a diameter greater than 1 cm but less
than 5 cm. We counted and identified to species level
every sapling in the plot. We also counted all seedlings
within a nested circular plot of 2 m radius around each
plot centre. The DBH of live trees was measured at a
height of 1.3 m above the ground surface, or 30 cm
above the highest root if the root was higher than
1.3 m (e.g., for Rhizophora mucronata). For multi-
stemmed trees, the DBHs of all stems were measured,
though this accounted for a small percentage of the
total number of measured trees. The DBH of harvested
trees was measured either at a height of 1.3 m, or
immediately below the cut location if mangroves were
harvested at a height lower than 1.3 m. We assessed
harvested status by recording the way trees had been
harvested. Trees were recorded as either harvested at
the ground surface (1); harvested below 1.3 m height
(2); harvested above 1.3 m (3); or harvested by branch
only (4).
Calculation of statistics, forest composition
and structure indices
We calculated the tree basal area (TBA) for each tree
using the following equation: TBA (m2) = (DBH/
200)2 9 3.142 m2. For each species, we calculated:
(i) the absolute frequency (the percentage of plots in
which a species was recorded); ii) the absolute density
Fig. 1 The Bay of Assassins, southwestern Madagascar show-
ing mangrove forest cover (dark grey)
123
Wetlands Ecol Manage (2019) 27:609–625 611

(the number of trees per hectare); and (iii) the absolute
cover (the total basal area of all trees per hectare). We
converted absolute measures into relative measures by
expressing the absolute measure for each species as a
percentage of the total of the absolute measures for all
trees across all species. We then calculated the
importance value for each species by adding together
relative frequency, relative density and relative cover
(Muller-Dombois et al. 1974). We calculated relative
importance values by dividing the importance values
of each species by the total importance values of all
species and multiplying by 100. We repeated this for
saplings. We calculated the regeneration rate for each
species by dividing the number of saplings by the
number of trees and multiplying by 100.
Calculations of biomass
The living aboveground biomass (AGB) and below-
ground biomass (BGB) of mangroves trees was
estimated using published, species-specific allometric
equations, in line with previous biomass studies and
accepted international protocols (e.g., Kauffman and
Donato 2012). In contrast to previous studies that
utilize a standard mangrove allometric equation (e.g.,
Abino et al. 2014; Rahman et al. 2015), we used
species-specific allometric equations (where avail-
able) to increase the robustness of our biomass
estimates. Where an equation for a particular species
was not available, it was substituted for a similar
species, e.g., Rhizophora apiculata (Putz and Chan
1986) in place of R. mucronata. In place of an equation
for Ceriops tagal we used an equation for C. australis
(Comley and McGuiness 2005); while C. australis and
C. tagal are genetically distinct species they are
botanically very similar, and until recently were
considered the same species. We also chose published
equations from locations that most closely matched
the forest characteristics and local geomorphology of
our study site where possible (Table 1). We could not
easily control for biogeographic region because of a
lack of suitable robust studies of allometry for African
mangroves. However, regional differences in allom-
etry are not generally considered significant (Chave
et al. 2014).
Surveys of mangrove wood use
To investigate mangrove wood use by local commu-
nities we used Rapid Rural Appraisal (RRA) tech-
niques (McCracken et al. 1988; Chambers 1992; Pratt
and Loizos 1992; Newing 2011). RRA covers a broad
range of methods devised to identify the problems and
strategies of households, groups and communities in a
limited time span that precludes in-depth ethnographic
fieldwork or quantitative household surveys (Pratt and
Loizos 1992; Newing 2011). The core principle of
RRA is triangulation, where data from different
techniques and informants are compared against each
other to reduce various forms of individual bias in
responses (e.g. according to age, gender or socio-
economic class) and arrive at a rigorous understanding
of major similarities and differences in resource use.
Our RRA was based on a literature review (includ-
ing data from household surveys carried out by an
environmental non-governmental organization); inter-
views with local experts working for environmental
non-environmental organizations; focus group discus-
sions in five villages; and semi-structured interviews
and transect walks with 15 key informants. We
selected these informants purposively to provide
further information on specific aspects of mangrove
use. Our informants included fishermen, individuals
involved in gleaning, and individuals involved in
mangrove wood harvesting for various purposes (e.g.
house construction and lime kiln construction). We
interviewed both men and women to ensure informa-
tion on the widest possible range of household uses
and (often gender-specific) livelihood activities relat-
ing to mangroves. Interviews lasted between 30 min
and an hour. Transect walks involved walking with
informants along a path covering major aspects of the
landscape (villages, livestock pens, farms, mangrove
forests, and dry forests) and resource uses (e.g. fishing,
gleaning, and aquaculture).
Results
The composition and structure of live mangroves
in the Bay of Assassins
While six species of mangrove tree have been
recorded in the Bay of Assassins (Benson et al.
2017), we recorded only five species in our vegetation
123
612 Wetlands Ecol Manage (2019) 27:609–625
surveys: Avicennia marina (afiafy in the local Mala-
gasy dialect), Bruguiera gymnorhiza (tangampoly), C.
tagal (tangambavy), R. mucronata (tangandahy), and
a single individual of Xylocarpus granatum. We also
came across a small number of Sonneratia alba trees
outside our plots. X. granatum is rarely found in
Malagasy mangroves (Gaudian et al. 1995). We
measured a total of 2667 live trees, 4861 live saplings
and 1058 seedlings across the 60 plots.
The mean tree basal area for live trees across the 60
plots was 19.37 m2 ha-1 (SD = 9.24). Our plots had a
mean canopy cover of 73.5% (SD = 23.07). Table 2
shows that the most important species in the man-
groves of the Bay of Assassins are C. tagal and R.
mucronata. C. tagal dominates, with the highest
frequency (100%), relative frequency (39.47%), rel-
ative density (59.1%), relative basal area cover
(52.17%), and relative importance value (50.25%).
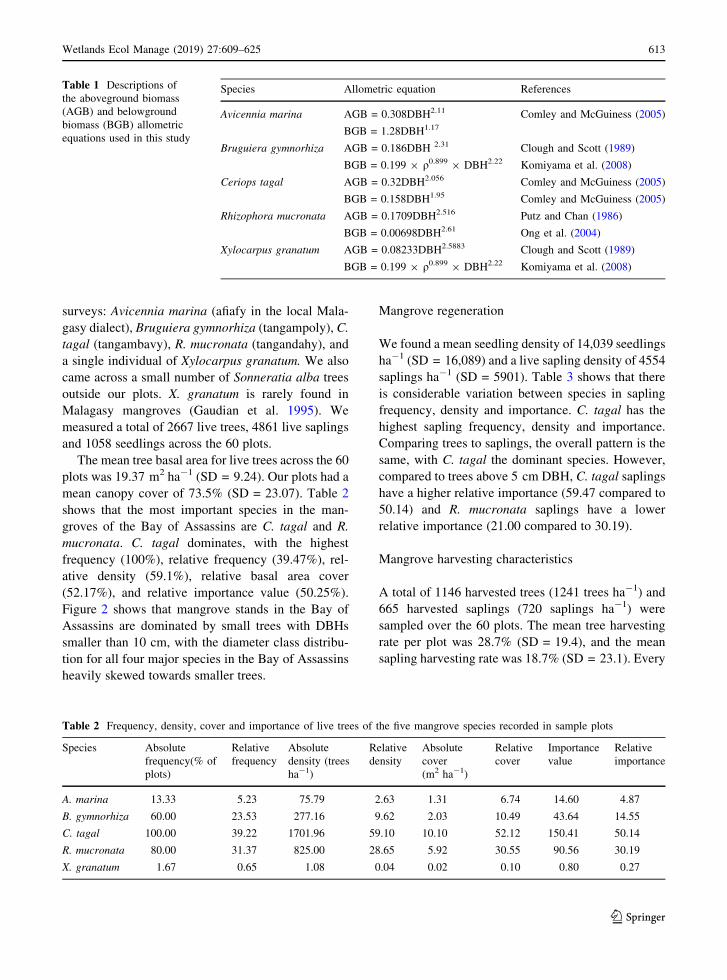
Figure 2 shows that mangrove stands in the Bay of
Assassins are dominated by small trees with DBHs
smaller than 10 cm, with the diameter class distribu-
tion for all four major species in the Bay of Assassins
heavily skewed towards smaller trees.
Mangrove regeneration
We found a mean seedling density of 14,039 seedlings
ha-1 (SD = 16,089) and a live sapling density of 4554
saplings ha-1 (SD = 5901). Table 3 shows that there
is considerable variation between species in sapling
frequency, density and importance. C. tagal has the
highest sapling frequency, density and importance.
Comparing trees to saplings, the overall pattern is the
same, with C. tagal the dominant species. However,
compared to trees above 5 cm DBH, C. tagal saplings
have a higher relative importance (59.47 compared to
50.14) and R. mucronata saplings have a lower
relative importance (21.00 compared to 30.19).
Mangrove harvesting characteristics
A total of 1146 harvested trees (1241 trees ha-1) and
665 harvested saplings (720 saplings ha-1) were
sampled over the 60 plots. The mean tree harvesting
rate per plot was 28.7% (SD = 19.4), and the mean
sapling harvesting rate was 18.7% (SD = 23.1). Every
Table 1 Descriptions of
the aboveground biomass
(AGB) and belowground
biomass (BGB) allometric
equations used in this study
Species Allometric equation References
Avicennia marina AGB = 0.308DBH2.11 Comley and McGuiness (2005)
BGB = 1.28DBH1.17
Bruguiera gymnorhiza AGB = 0.186DBH 2.31 Clough and Scott (1989)
BGB = 0.199 9 q0.899 9 DBH2.22 Komiyama et al. (2008)
Ceriops tagal AGB = 0.32DBH2.056 Comley and McGuiness (2005)
BGB = 0.158DBH1.95 Comley and McGuiness (2005)
Rhizophora mucronata AGB = 0.1709DBH2.516 Putz and Chan (1986)
BGB = 0.00698DBH2.61 Ong et al. (2004)
Xylocarpus granatum AGB = 0.08233DBH2.5883 Clough and Scott (1989)
BGB = 0.199 9 q0.899 9 DBH2.22 Komiyama et al. (2008)
Table 2 Frequency, density, cover and importance of live trees of the five mangrove species recorded in sample plots
Species Absolute
frequency(% of
plots)
Relative
frequency
Absolute
density (trees
ha-1)
Relative
density
Absolute
cover
(m2 ha-1)
Relative
cover
Importance
value
Relative
importance
A. marina 13.33 5.23 75.79 2.63 1.31 6.74 14.60 4.87
B. gymnorhiza 60.00 23.53 277.16 9.62 2.03 10.49 43.64 14.55
C. tagal 100.00 39.22 1701.96 59.10 10.10 52.12 150.41 50.14
R. mucronata 80.00 31.37 825.00 28.65 5.92 30.55 90.56 30.19
X. granatum 1.67 0.65 1.08 0.04 0.02 0.10 0.80 0.27
123
Wetlands Ecol Manage (2019) 27:609–625 613
plot sampled contained harvested trees, with plots
ranging in harvesting rate from 1.5% to 71.4%.
Harvest pressure differed between species; C. tagal
was the most harvested species, contributing 81.2% of
all harvested trees, followed by R. mucronata (13.6%),
B. gymnorhiza (4.6%) and A. marina (0.7%). Of all C.
tagal trees measured, 35.8% were harvested, com-
pared to 16.3% of R. mucronata trees, 16% of B.
gymnorhiza trees and 9.3% of A. marina trees. The
large majority of trees (92.7%) were harvested by
cutting the tree at the trunk rather than cutting
branches.
The mean DBH of harvested trees was 9.21 cm
(SD = 3.67). Looking at all species together, the most
harvested size class was 5–9 cm DBH. These
accounted for 61.59% of all harvested stems. Table 4
compares the mean DBH of non-harvested and
harvested trees for each species. It shows that for
every species apart from A. marina, the mean DBH of
harvested species was significantly greater
Fig. 2 Frequency distributions of the DBH (cm) of non-harvested and harvested trees of the four major species found in the Bay of
Assassins
Table 3 Live sapling frequency, density and regeneration rate for each species
Species Absolute
frequency (%
of plots)
Relative
frequency
Absolute
density
(saplings ha-1)
Relative
density
Importance (relative
frequency ? relative
density)
Relative
importance
Regeneration
rate
A. marina 3.33 1.52 12.99 0.29 1.81 0.90 13.95
B. gymnorhiza 65.00 29.55 350.79 7.72 37.27 18.63 102.76
C. tagal 85.00 38.64 3647.53 80.31 118.95 59.47 153.70
R. mucronata 66.70 30.30 530.51 11.68 41.98 21.00 124.76
123
614 Wetlands Ecol Manage (2019) 27:609–625

(p =\ 0.05) than the mean DBH of non-harvested
trees, indicating that harvesters are preferentially
selecting trees that are larger than the average size.
The AGB for all our plots averaged 161.9 Mg ha-1
(SD = 68.6 Mg ha-1). However, harvesting pressure
means that this average is below the potential AGB
that is available at this site. A mean AGB of 46.7
Mg ha-1 was removed through harvesting, though a
large standard deviation (42.4 Mg ha-1) is reflective
of the substantial variation in total harvested AGB
across the 60 plots, ranging from 0 to 163.3 Mg ha-1.
This means that the site AGB is 22.4% lower than the
potential AGB available at this site.
Ceriops tagal was the preferred species to be
harvested, accounting for 72.9% of the total AGB of
harvested wood (Table 5). This was followed by R.
mucronata (20.3% of all harvested wood). Size classes
were also differentially harvested. Trees with a DBH
between 5.0 cm and 9.9 cm accounted for an average
of 27.9% of all harvested trees. This is a dispropor-
tionate contribution, considering the small individual
volume of trees in this smaller size class, compared to
the larger trees harvested. Similar to the overall
volumes, C. tagal was strongly preferred as a smaller
pole, alone accounting for 21.4% of all AGB har-
vested. We discuss the uses of the different species and
size classes in ‘‘Human uses of harvested mangrove
wood’’ section.
Spatial patterns of mangrove harvesting
The plots we sampled were a mean distance of 1159 m
(SD = 606) from the nearest settlement, with a
minimum of 233 m and a maximum of 2801 m. Plots
were a mean distance of 92 m from the nearest
mangrove edge (SD = 62), with a minimum of 15 m
and a maximum of 352 m. Table 6 shows the results of
correlations conducted between distances of the plots
from the nearest human settlement; distances of the
plots from the nearest mangrove edge; and various
measures of mangrove harvesting.
The results show that there are significant inverse
correlations (p =\ 0.05) between the distance from
the nearest human settlement and the percentage of
trees harvested; the percentage of 5–9 cm and
10–14 cm DBH trees harvested; and the percentage
of B. gymnorhiza and C. tagal harvested. However,
there was no significant correlation between distance
from the nearest human settlement and the percentage
of A. marina or R. mucronata harvested. None of the
correlations between distance from the nearest man-
grove edge and tree harvesting were significant. These
results show that mangrove forests are more heavily
harvested closer to human settlements but that
distance from the mangrove edge (and thus access
from the sea or land) does not seem to influence
harvesting pressure.
Table 4 Comparison of the mean DBH in cm (with standard deviations) of non-harvested and harvested trees
Mean DBH (cm) of non-harvested trees Mean DBH (cm) of harvested trees t p
All trees 8.53 (3.55) 9.21 (3.67) - 5.26 \ 0.001
A. marina 12.48 (8.03) 10.66 (3.53) 1.16 0.26
B. gymnorhiza 9.20 (3.98) 11.81 (4.64) - 4.38 \ 0.001
C. tagal 8.16 (2.97) 8.96 (3.62) - 5.70 \ 0.001
R. mucronata 8.85 (3.59) 9.74 (3.21) - 3.09 0.002
Table 5 Percentage of
above-ground biomass
(AGB, Mg Ha-1) harvested
by species and size class.
Total = 100%
DBH[ 5.0\ 10.0 cm (%) DBH[ 10 cm (%)
A. marina 0.20 0.58
B. gymnorhiza 0.56 5.21
C. tagal 21.40 51.52
R. mucronata 5.77 14.53
A. marina 0.00 0.23
123
Wetlands Ecol Manage (2019) 27:609–625 615

Human uses of harvested mangrove wood
The Rapid Rural Appraisal showed that mangrove
trees in the Bay of Assassins were harvested: (i) as
fuelwood for domestic cooking; (ii) as a building
material for housing and fencing; (iii) as a building
material for seaweed aquaculture; and (iv) for the
construction of lime kilns. Respondents suggested that
the use of mangrove wood in lime kilns and in seaweed
aquaculture are recent, having only developed over the
last 10 years. Mangrove wood was also recently used
in sea cucumber (Holothuria scabra) aquaculture to
construct growing-out enclosures. However, man-
grove wood has now been replaced with stronger
and more durable steel rods for this use.
With regards to domestic fuelwood, saplings and
trees less than 10 cm DBH were sometimes used but
wood from neighbouring terrestrial dry forests was
preferentially used as a fuel as it is usually drier and
more easily combustible. The most frequent use of
mangrove wood was for house and fence construction.
These activities almost exclusively use mangrove
wood rather than wood from terrestrial forests. Larger
poles (between 10 cm and 20 cm diameter) were used
to build the frames of houses (Fig. 3a), with smaller
poles (5 to 10 cm diameter) used to support wall and
roof material. Respondents generally favoured R.
mucronata for the construction of house frames
because of its resistance to decay.
Poles of C. tagal were used in the construction of
fencing around houses (background of Fig. 3b) and to
construct livestock enclosures. Fences are particularly
important due to high incidences of banditry in the
region. Fence poles were used in large quantities due
to frequent maintenance and replacement require-
ments, with poles lasting 2 or 3 years. Our informants
told us that fence construction was the most common
reason for harvesting mangrove wood. This is sup-
ported by the vegetation survey (‘‘Mangrove harvest-
ing characteristics’’ section), which showed that a
disproportionate volume of harvested AGB was C.
tagal stems with a DBH of between 5 and 9.9 cm (the
preferred species and size class for fencing).
A more recent use of mangrove wood in the Bay of
Assassins involves seaweed (Kappaphycus alvarezii)
aquaculture. Saplings and trees with a DBH of 5 to
9 cm and a length of approximately 1 m were used as
stakes for seaweed farming. The stakes provide anchor
points for ropes on which the algae grow, but needed
replacing every 6–24 months. Mangrove poles were
also used to build tables to dry the seaweed (Fig. 3b).
Another more recent use of mangrove wood in the
Bay of Assassins involved the construction of lime
kilns (Fig. 3c). These were used to produce ‘sokay’, a
sea-shell based lime render used to improve the
durability of house walls (Fig. 3d). To construct a kiln,
layers of sea shells are sandwiched between layers of
mangrove wood. Burning the shells converts calcium
Table 6 Correlation coefficients for relationship between distance from the nearest human settlement and distance from the nearest
mangrove edge
Distance from nearest human settlement
(m)
Distance from nearest mangrove edge
(m)
r p r p
Percentage of trees harvested - 0.43 \ 0.001 - 0.21 0.10
Percentage of saplings harvested 0.08 0.53 - 0.03 0.80
Percentage of trees 5–9 cm DBH harvested - 0.42 \ 0.001 - 0.23 0.07
Percentage of trees 10–14 cm DBH harvested - 0.39 0.002 - 0.12 0.37
Percentage of trees 15–19 cm DBH harvested - 0.13 0.36 0.18 0.22
Percentage of trees C 20 cm DBH harvested - 0.09 0.71 0.36 0.10
Percentage of A. marina trees harvested - 0.40 0.37 - 0.12 0.80
Percentage of B. gymnorhiza trees harvested - 0.38 0.02 - 0.09 0.56
Percentage of C. tagal trees harvested - 0.47 \ 0.001 - 0.29 0.03
Percentage of R. mucronata trees harvested - 0.23 0.11 - 0.18 0.21
123
616 Wetlands Ecol Manage (2019) 27:609–625

carbonate (CaCO3) to calcium oxide (CaO). Water is
added to the resulting powder and the paste is then
applied to house walls. Lime render was the preferred
wall material because of its relative durability com-
pared to the mangrove saplings and reeds used in
traditional houses (Fig. 3b). Madagascar regularly
experiences tropical cyclones with strong winds and
heavy rain, and respondents reported that lime
rendered walls are more resistant to cyclone damage.
We found, based on measurements of 10 kilns
found in five villages, that a typical lime kiln measures
c. 2.6 m in length, 2.3 m in width and 1.2 m in height,
using mangrove poles of c. 10–15 cm diameter. Kiln
construction requires c. 120 poles of mangrove wood,
with a total volume of c. 2.4 m3. Respondents stated
that their preferred species of mangrove wood for kiln
construction was R. mucronata because it burns hotter
than other mangrove and terrestrial forest tree species,
Fig. 3 Examples of the use of mangrove wood: saplings and
trees used in house construction (a); saplings used as fencing
(background of b); trees used to construct seaweed drying racks
(foreground of b); and large trees used to construct kilns
(c) which produce lime render for houses (d)
123
Wetlands Ecol Manage (2019) 27:609–625 617

producing a purer lime powder. However, in our
village surveys and observations of the kilns we noted
that other species, particularly C. tagal and B.
gymnorhiza were also used, presumably due to
availability. Kiln builders stated that the ideal size
class for mangrove trees to build kilns was between
10 cm and 15 cm DBH. Trees over 20 cm DBH were
too large for an individual to carry out of the
mangroves, and as such are harvested infrequently.
Discussion
Forest structure, composition and regeneration
The mix of species in the Bay of Assassins is typical of
southwest Madagascar’s mangroves, with five species
recorded in our plots, compared to a total of six species
known to be present in the Bay (Benson et al. 2017).
However, in comparison to other sites in southwest
Madagascar, the mangroves of the Bay of Assassins
are unusual in that they are strongly dominated by C.
tagal. In our survey this species was found in all plots
and was the dominant species in 65% of plots,
accounting for 65.52% of all trees sampled. In
comparison, the mangroves of the Mangoky River
Delta, located approximately 80 km north of the Bay
of Assassins, are dominated by R. mucronata and A.
marina (Rakotomavo and Fromard 2010), as are the
mangroves of the Tulear Lagoon, 180 km to the south
of our study site (Laroche et al. 1997).
It is possible that geomorphology plays a role in the
differences between these different sites, since man-
grove species distribution is heavily influenced by
abiotic factors such as inundation frequency (Leong
et al. 2018). However, information on the geomorphic
setting of these locations is not available. The
dominance of C. tagal in the Bay of Assassins may
also be driven by historical harvesting pressure on R.
mucronata, coupled with C. tagal being the dominant
regeneration species (as suggested by its higher
importance values). Other East African case studies
have shown that the preferential harvesting of R.
mucronata (due to its high density and calorific value)
leads to a change in mangrove community dominated
by C. tagal over time (Kairo et al. 2002). However, a
lack of historical data on harvesting or vegetation
structure at this sites means we are unable to test this
hypothesis in the Bay of Assassins.
We also found that for all four mangrove tree
species commonly found in the Bay of Assassins (A.
marina, B. gymnorhiza, C. tagal, and R. mucronata),
mangrove forests were dominated by small trees that
were \ 10 cm DBH. This could be due to a combi-
nation of climatic pressures and harvesting pressure.
Mangrove growth rates and biomass accumulation are
often linked to climatic variables such as cyclone
frequency (Simard et al. 2019) and, more importantly,
long term distribution of rainfall (Krauss et al. 2007;
Sanders et al. 2016). Arid conditions (such as those in
our study site) are associated with high soil salinities,
which may restrict mangrove growth (Cintron et al.
1978) and cause them to allocate greater biomass to
the below-ground fraction to minimize salinity gradi-
ents around the roots and aid in water uptake (Clough
et al. 1997; Alongi et al. 2005). This would lead to
lower biomass partitioning (and DBH) above ground.
Extreme aridity has thus been suggested as a reason for
low biomass and small DBH for mangroves along the
arid coast of Western Australia (Clough et al. 1997),
though other studies in Australia show different results
(Comley and McGuiness 2005). Thus, harvesting
pressure probably also plays a role in controlling size
classes. In general, harvested trees were significantly
larger than non-harvested trees (with the exception of
A. marina, which is not preferred as harvested material
to the same degree as the species of Rhizophoraceae),
with the mean DBH of remaining non-harvested trees
being an average of\ 9 cm.
Harvesting pressure and patterns in the Bay
of Assassins
Unlike many other parts of Madagascar (cf. Jones et al.
2014; Giri and Mulhausen 2008), the clear cutting of
mangroves is, at present, relatively rare in the Bay of
Assassins. The region experienced a rate of mangrove
cover loss of 0.27% per year between 2002 and 2014
(Benson et al. 2017), compared to 1.19% per year for
mangrove in northwestern Madagascar between 1990
and 2010 (Jones et al. 2014); 0.85% per year between
1951 and 2000 in the Mangoky River delta (Rako-
tomavo and Fromard 2010); and 0.52% per year for the
whole of Madagascar between 2000 and 2005 (Giri
and Mulhausen 2008). Thus, mangrove forests in the
Bay of Assassins are not, at present, facing the large-
scale deforestation threats that other mangrove forests
in Madagascar are experiencing. In other parts of
123
618 Wetlands Ecol Manage (2019) 27:609–625
Madagascar, mangrove deforestation has been driven
primarily by clearance for agriculture, large-scale
commercial shrimp aquaculture, and commercial
charcoal production (Giri and Mulhausen 2008;
Rakotomavo and Fromard 2010; Jones et al. 2016),
none of which are currently resource uses found in the
Bay of Assassins. This is possibly because the Bay of
Assassins is too arid and currently too inaccessible
from major markets for these alternative land uses to
be financially viable.
During the vegetation survey we only came across
one instance of clear cutting, which was outside our
randomly-assigned vegetation plots and was related to
a large order for lime render. Instead, the ecological
impacts of mangrove wood harvesting are more
cryptic (Dahdouh-Guebas et al. 2005). We did not
come across a single unharvested plot during our
extensive survey. Thus, the main stressor on the Bay of
Assassins system is selective harvesting for a range of
household construction activities, particularly fencing.
However, even with harvesting, mean canopy cover
was high (73.5%, SD = 23.07).
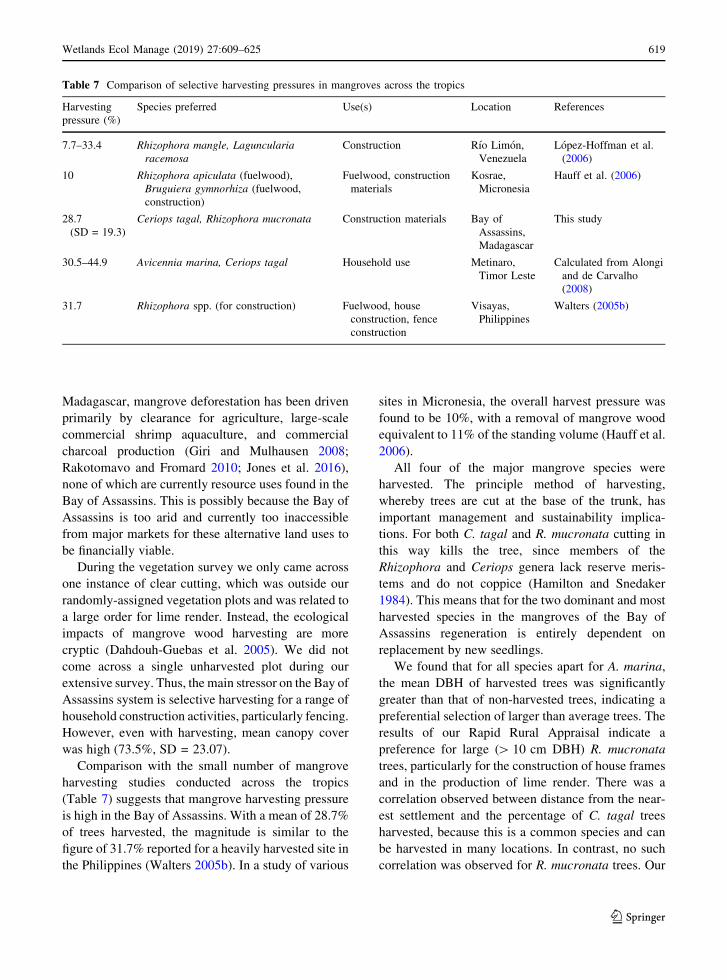
Comparison with the small number of mangrove
harvesting studies conducted across the tropics
(Table 7) suggests that mangrove harvesting pressure
is high in the Bay of Assassins. With a mean of 28.7%
of trees harvested, the magnitude is similar to the
figure of 31.7% reported for a heavily harvested site in
the Philippines (Walters 2005b). In a study of various
sites in Micronesia, the overall harvest pressure was
found to be 10%, with a removal of mangrove wood
equivalent to 11% of the standing volume (Hauff et al.
2006).
All four of the major mangrove species were
harvested. The principle method of harvesting,
whereby trees are cut at the base of the trunk, has
important management and sustainability implica-
tions. For both C. tagal and R. mucronata cutting in
this way kills the tree, since members of the
Rhizophora and Ceriops genera lack reserve meris-
tems and do not coppice (Hamilton and Snedaker
1984). This means that for the two dominant and most
harvested species in the mangroves of the Bay of
Assassins regeneration is entirely dependent on
replacement by new seedlings.
We found that for all species apart for A. marina,
the mean DBH of harvested trees was significantly
greater than that of non-harvested trees, indicating a
preferential selection of larger than average trees. The
results of our Rapid Rural Appraisal indicate a
preference for large ([ 10 cm DBH) R. mucronata
trees, particularly for the construction of house frames
and in the production of lime render. There was a
correlation observed between distance from the near-
est settlement and the percentage of C. tagal trees
harvested, because this is a common species and can
be harvested in many locations. In contrast, no such
correlation was observed for R. mucronata trees. Our
Table 7 Comparison of selective harvesting pressures in mangroves across the tropics
Harvesting
pressure (%)
Species preferred Use(s) Location References
7.7–33.4 Rhizophora mangle, Laguncularia
racemosa
Construction Rıo Limon,
Venezuela
Lopez-Hoffman et al.
(2006)
10 Rhizophora apiculata (fuelwood),
Bruguiera gymnorhiza (fuelwood,
construction)
Fuelwood, construction
materials
Kosrae,
Micronesia
Hauff et al. (2006)
28.7
(SD = 19.3)
Ceriops tagal, Rhizophora mucronata Construction materials Bay of
Assassins,
Madagascar
This study
30.5–44.9 Avicennia marina, Ceriops tagal Household use Metinaro,
Timor Leste
Calculated from Alongi
and de Carvalho
(2008)
31.7 Rhizophora spp. (for construction) Fuelwood, house
construction, fence
construction
Visayas,
Philippines
Walters (2005b)
123
Wetlands Ecol Manage (2019) 27:609–625 619

key informants informed us that as large ([ 10 cm
DBH) R. mucronata trees are particularly desirable,
resource users will travel further to extract them.
Taken together with the importance values of trees
and saplings, these results suggest a shift in commu-
nity composition and structure, with harvesting lead-
ing to a relative absence of large trees, and an increase
in the importance of younger and smaller C. tagal
trees. This is similar to other studies that have shown
that harvesting pressure on Rhizophora spp. is not
driven by distance; instead, resource users will travel
to locations depending on resource availability (Dah-
douh-Guebas et al. 2000; Palacios and Cantera 2017).
In South Africa, a study found that R. mucronata was
heavily harvested while adjacent species such as A.
marina were not (Rajkaran et al. 2004).
Rhizophora species have historically been pre-
ferred for harvest due to their resistance to insect pests
when used as construction materials (e.g. De Puydt
1868) and their high calorific value, which makes
them ideal fuelwood for domestic purposes and, in the
case of the Bay of Assassins, in the construction of
lime kilns. They are still preferred by local commu-
nities for these reasons today (e.g. Dahdouh-Guebas
et al. 2000). This fact has important implications for
the spatial planning of potential management areas
versus the spatial distribution of preferred species.
Contributions of mangrove harvesting to local
livelihoods
Mangroves are often used for fuelwood and charcoal
production in sub-Saharan Africa (Dahdouh-Guebas
et al. 2000; Rajkaran et al. 2004) and other tropical
coastal areas (e.g. Oo 2002; Sudtongkong and Webb
2008; Moriizumi et al. 2010), though this was not the
case in the Bay of Assassins. Forest users instead
prefer to use fuelwood resources collected from
terrestrial spiny forest, which is itself heavily threat-
ened (Seddon et al. 2000; Casse et al. 2004).
In the Bay of Assassins, mangrove wood is used
primarily in the construction of houses and fences,
where its ability to resist decay makes it a favoured
material. Mangrove wood plays an important role in
household security, as it is used to build fences to
protect property (including livestock) from banditry. It
is also used in the reinforcement of house walls (and
thus protection from cyclones) through the production
of lime render. The harvesting of 5–9 cm trees, which
are primarily used as fencing material, accounts for the
majority of above-ground mangrove wood biomass
removed.
We have also documented emerging uses of
mangrove wood. The recent development of seaweed
and sea cucumber aquaculture has created new
demand for mangrove wood. In addition, the produc-
tion of lime render using mangrove wood kilns is also
creating demand for mangrove wood. Lime render has
only recently been identified as a significant resource
use associated with mangrove harvesting in the Bay of
Assassins (Scales et al. 2018) and, to our knowledge,
has not been recorded elsewhere in Madagascar or
sub-Saharan Africa. Rendering the walls of an average
house requires the lime produced by two kilns, which
themselves require 4.8 m3 of mangrove wood. To put
this in perspective, the frame and un-rendered walls of
a typical house in the region require 0.9 m3 of wood
(Rasolofo 1997). A rendered house uses approxi-
mately six times more mangrove wood than an un-
rendered house.
Reconciling the ecology and management
of the Bay of Assassins
This study highlights the strong dependence that local
communities have on the mangrove forests in the Bay
of Assassins, with provisioning ecosystem services
such as seafood (Aina 2010) and wood products
(Scales et al. 2018; this study) making an important
contribution to livelihoods and the local economy. It is
crucial to manage these resources sustainably, so that
they can continue to provide wood and fuel resources
into the future without adverse ecological impact.
How sustainable are current mangrove harvesting
practices? Extrapolating our data on harvested trees
from the 60 vegetation plots (0.92 ha) to the 1300 ha
of mangroves in the Bay of Assassins indicates that
there are approximately 2.76 million live trees in the
Bay of Assassins with a DBH of 5–9 cm. A household
survey carried out in seven of the ten villages in 2014
estimated that mangrove harvesting for fence poles
accounted for approximately 50 stems per person per
year (Blue Ventures 2015). This translates a total of
150,000 stems per year, or 5.43% of the trees in the
5–9 cm size class, for all the villages in the Bay of
Assassins.
Our study suggests that while mean canopy cover
(73.5%) remains high and the mangroves of the Bay of
123
620 Wetlands Ecol Manage (2019) 27:609–625

Assassins are currently regenerating, ecological
impact is apparent. Looking to the future, it is likely
that human pressures on the mangroves of the Bay of
Assassins will continue to increase. The human
population in the region is growing. Data are scarce,
but informants reported significant in-migration over
the last 5 years. This is supported by Epps (2007), who
found that\ 30% of inhabitants in Lamboara and
Ampasilava were born in those settlements. Coastal
areas in southwest Madagascar are experiencing rapid
in-migration, as poor agricultural households move to
the coast seeking more secure livelihoods (Brugge-
mann et al. 2012). A growing population means more
demand for mangrove wood, especially for housing
and fencing. The harvesting of 5–9 cm trees, which
are primarily used as fencing material, accounts for a
disproportionate percentage of above-ground man-
grove wood biomass removed, considering their small
individual volume, and those managing the mangroves
of the Bay of Assassins may need to look for
alternative fencing material if demand continues to
grow.
In addition to population growth, the region is also
experiencing economic change, which is increasing
demand for mangrove wood. Over the last 10 years the
region has become connected to global commodity
chains of sea cucumber, seaweed and octopus (Aina
2010; Barnes-Mauthe et al. 2013). This has created
new pressures on mangroves through the wood needed
for aquaculture, for example to construct sea cucum-
ber enclosures (although these are now made of steel
rods), seaweed anchors and seaweed drying tables.
Growing income from aquaculture has also led to
increased demand for lime render for houses, as
households who are able to benefit from the new
commodity chains for marine products choose to
improve the durability of their houses.
The harvesting of sea cucumber, seaweed and
octopus have all increased over the last 10 years (Aina
2010; Barnes-Mauthe et al. 2013; Blue Ventures
2015). For example, the harvest of K. alvarezii in 2014
was over 55 tonnes, three times that of 2012. This has
important implications for the regions mangroves as
three tonnes of mangrove wood are required to
produce one tonne of dried seaweed (Blue Ventures
2015). Key informants also reported an increase in the
use of lime render. This is supported by previous
household surveys. In 2006, 28% of buildings in
Lamboara village had walls covered in lime render
(Epps 2007). By 2014 this figure had gone up to 65%
(Blue Ventures 2015). Lime production is a time-
consuming process, requiring men to find, cut, and
carry heavy trees. Most households do not have the
time to render their own walls, focusing their attention
on fishing and gleaning. However, with a rise in
income from aquaculture, wealthier households are
paying others to produce lime. As a result, lime render
is considered a status symbol. Demand for lime render,
and thus for mangrove wood to use in lime kilns, is
likely to grow with a growing population and/or rising
incomes. Growing demand for large ([ 10 cm DBH)
R. mucronata trees is occurring in a mangrove forest
that is dominated by C. tagal trees below 10 cm DBH.
To date, efforts to manage natural resources in the
Bay of Assassins have tended to focus on a single
ecosystem or species, for example establishing closed
seasons for octopus fisheries, introducing new tech-
niques for sea cucumber aquaculture, and creating
protected areas for mangrove forest where wood
harvesting is forbidden (Andriamalala and Gardner
2010; Aina 2010; Benbow et al. 2014; Cripps and
Gardner 2016). The key resource management lesson
is that livelihoods in coastal communities cut across
terrestrial, wetland and aquatic ecosystems. This
means that mangroves are socio-ecologically linked
to other ecosystems and must therefore be managed as
part of broader landscape-based approaches. For
example, increases in wealth from marine resources
such as sea cucumber, algae and octopus are likely to
lead to greater demand for more durable housing
material which, unless alternatives are found, will lead
to increased demand for lime render and thus place
pressure on mangrove forests. Another potential cross-
ecosystem linkage involves domestic fuelwood. While
mangrove wood is the preferred material for construc-
tion, terrestrial dry forests are the preferred source of
fuelwood. However, a reduction in terrestrial dry
forest (for example due to forest clearance for
agriculture or over-harvesting of fuelwood), is likely
to lead to more pressure on mangroves as households
seek out alternative sources of domestic fuel. There-
fore, management policies need to carefully model
expected increases in demand for mangrove wood for
different uses and consider developing alternative
sources of domestic fuel and alternative ways to
improve the durability of houses.
123
Wetlands Ecol Manage (2019) 27:609–625 621

Conclusions
Small-scale wood harvesting by local communities
can be a significant driver of degradation in many
mangrove forests across the tropics (UNEP 2014).
Given the importance of mangroves to the livelihoods
of coastal communities in the tropics and the potential
impact of human activities on mangrove cover,
structure and composition, it is remarkable that so
few studies of small-scale mangrove harvesting have
been published. Our understanding of mangrove uses
and their ecological impacts is hindered by a narrow
focus on land cover change measured through remote
sensing, which misses more cryptic forms of ecolog-
ical change (Dahdouh-Guebas et al. 2005). There is
thus a need for field surveys, combined with studies of
human resource use, to reveal cryptic processes of
ecological change that are not easily revealed through
remote sensing.
This study, based on a combination of mangrove
vegetation surveys and RRA techniques, has shown
that poor coastal communities in southwest Madagas-
car use mangrove wood for both domestic and
commercial purposes. We have shown high harvesting
pressure. Our study also shows that harvesting is
spatially variable and can have observable ecological
impacts on multiple mangrove forest community
characteristics. It also shows that the local community
has clear intended uses, which drives the harvesting of
particular species and size classes.
Environmental policy in the tropics often focuses
on demographic change and the implications of
population growth on resource use. There is a lack
of understanding of the diverse ways in which low-
income households, particularly in remote regions,
rely on mangrove resources and a tendency to assume
all poor households use natural resources in the same
way (Angelsen and Wunder 2003; Belcher 2005). This
study found that resource extraction, poverty and
livelihoods are interlinked, with increasing livelihood
status changing the type of resource extracted (in this
case R. mucronata wood for lime kiln construction).
Thus, livelihood status is likely to impact the species
preferred and the volume harvested. Even remote
coastal regions such as the Bay of Assassins are
increasingly linked to global commodity chains,
leading to changes in resource use. More attention
needs to be paid, through household surveys, to the
quantities of mangrove wood used for different small-
scale purposes, and how these vary according to socio-
economic characteristics such as wealth and migration
status, so that adverse ecological impacts on forest
resources can be better quantified, anticipated and
managed.
Finally, our study suggests that mangroves must be
managed as part of broader landscape-based
approaches. In the Bay of Assassins resource man-
agement policies have tended to focus on single
species and ecosystems. For example, there have been
efforts to improve the sustainability of marine
resource use through the introduction of a closed
season for octopus fisheries and new techniques for sea
cucumber aquaculture. However, income from marine
resources such as sea cucumber, seaweed and octopus
has led to growing demand for lime render to improve
the durability of houses. Lime render is made in
mangrove wood kilns and growing demand has led to
increased pressure on mangrove forests. In turn, the
loss of mangroves has implications for natural coastal
fisheries, due to their role in the life-cycle of fish and
other marine fauna (Whitfield 2017) that also play an
important role in local livelihoods. Therefore, man-
agement plans must carefully consider how changes in
the socio-ecological dynamics of one natural resource
are likely to impact those of other resources that
households depend on. Research has shown that
governance decentralization and community man-
grove management can improve mangrove condition
if strong community institutions are present to
enforcement common rules of natural resource man-
agement (Primavera 2001; Sudtongkong and Webb
2008; Damastuti and de Groot 2017). Community-
based management approaches will therefore be of
crucial importance in any attempt to manage man-
groves as part of a broader landscape-based approach.
Acknowledgements This research was funded through an
Environment and Sustainability Grant from the Royal
Geographical Society (with IBG). We thank Blue Ventures for
their logistical support; Michael Strongoff, Jose Njara
Ranaivoson and Helen Scales for fieldwork assistance; and
Philip Stickler (Department of Geography, University of
Cambridge) for cartographic assistance. We also thank the
two anonymous reviewers for their comments and suggestions,
which helped to improve the clarity of the paper.
Open Access This article is distributed under the terms of the
Creative Commons Attribution 4.0 International License (http://
creativecommons.org/licenses/by/4.0/), which permits unre-
stricted use, distribution, and reproduction in any medium,
provided you give appropriate credit to the original
123
622 Wetlands Ecol Manage (2019) 27:609–625

author(s) and the source, provide a link to the Creative Com-
mons license, and indicate if changes were made.
References
Abino AC, Castillo JAA, Lee YJ (2014) Assessment of species
diversity, biomass and carbon sequestration potential of a
natural mangrove stand in Samar, the Philippines. For Sci
Technol 10:2–8
Aina TAN (2010) Management of octopus fishery off south west
Madagascar. United Nations University Fisheries Training
Programme, Reykjavık
Alongi DM, de Carvalho NA (2008) The effect of small-scale
logging on stand characteristics and soil biogeochemistry
in mangrove forests of Timor Leste. For Ecol Manage
255:1359–1366
Alongi DM, Clough BF, Robertson AI (2005) Nutrient-use
efficiency in arid-zone forests of the mangroves Rhi-
zophora stylosa and Avicennia marina. Aquat Bot
82:121–131
Andriamalala G, Gardner C (2010) L’utilisation du dina comme
outil de gouvernance des ressources naturelles: lecons tir-
ees de Velondriake, sud-ouest de Madagascar. Trop Con-
serv Sci 3:447–472
Angelsen A, Wunder S (2003) Exploring the forest–poverty
link: key concepts, issues and research implications.
CIFOR Occasional Paper No. 40. Center for International
Forestry Research, Jakarta
Bandaranayake WM (1998) Traditional and medicinal uses of
mangroves. Mangroves Saltmarshes 2:133–148
Barbier EB, Hacker SD, Kennedy C, Koch EW, Stier AC, Sil-
liman BR (2011) The value of estuarine and coastal
ecosystem services. Ecol Monogr 81:169–193
Barnes-Mauthe M, Oleson KL, Zafindrasilivonona B (2013)
The total economic value of small-scale fisheries with a
characterization of post-landing trends: an application in
Madagascar with global relevance. Fish Res 147:175–185
Belcher BM (2005) Forest product markets, forests and poverty
reduction. Int For Rev 7:82–89
Benbow S, Humber F, Oliver TA, Oleson KLL, Raberinary D,
Nadon M, Ratsimbazafy H, Harris A (2014) Lessons learnt
from experimental temporary octopus fishing closures in
south-west Madagascar: benefits of concurrent closures.
Afr J Mar Sci 36:31–37
Benson L, Glass L, Jones TG, Ravaoarinorotsihoarana L,
Rakotomahazo C (2017) Mangrove carbon stocks and
ecosystem cover dynamics in southwest Madagascar and
the implications for local management. Forests 8:190
Blue Ventures (2015) Mangrove use in the Bay of Assassins.
Blue Ventures Conservation, London
Bruggemann JH, Rodier M, Guillaume MMM, Andrefouet S,
Arfi R, Cinner JE, Pichon M, Ramahatratra F, Rasoama-
nendrika F, Zinke J, McClanahan TR (2012) Wicked
social-ecological problems forcing unprecedented change
on the latitudinal margins of coral reefs: the case of
southwest Madagascar. Ecol Soc 17:47
Casse T, Miloj A, Ranaivoson S, Randriamanarivo JR (2004)
Causes of deforestation in southwestern Madagascar: what
do we know? For Policy Econ 6:33–48
Chambers R (1992) Rural appraisal: rapid, relaxed and partici-
patory. IDS Discussion Paper 311. Institute of Develop-
ment Studies, Brighton
Chave J, Rejou-Mechain M, Burquez A, Chidumayo E, Colgan,
MS, Delitti WBC, Duque A, Eid T, Fearnside PM, Good-
man RC, Henry M, Martınez- Yrızar A, Mugasha WA,
Muller-Landau HC, Mencuccini M, Nelson BW, Ngo-
manda A, Nogueira EM, Ortiz-Malavassi E, Pelissier R,
Ploton P, Ryan CM, Saldarriaga JG, Vieilledent G (2014)
Improved allometric models to estimate the aboveground
biomass of tropical trees. Glob Chang Biol 20:3177–3190
Cintron G, Lugo AE, Pool DJ, Morris G (1978) Mangroves of
arid environments in puerto rico and adjacent Islands.
Biotropica 10:110–121
Clough BF, Dixon P, Dalhaus O (1997) Allometric relationships
for estimating biomass in multi-stemmed mangrove trees.
Aust J Bot 45:1023–1031
Clough BF, Scott K (1989) Allometric relationships for esti-
mating above-ground biomass in six mangrove species. For
Ecol Manag 27:117–127
Comley BW, McGuiness KA (2005) Above- and below-ground
biomass, and allometry, of four common northern Aus-
tralian mangroves. Aust J Bot 53:431–436
Cripps G, Gardner CJ (2016) Human migration and marine
protected areas: insights from Vezo fishers in Madagascar.
Geoforum 74:49–62
Dahdouh-Guebas F, Mathenge C, Kairo JG, Koedam N (2000)
Utilization of mangrove wood products around Mida Creek
(Kenya) amongst subsistence and commercial users. Econ
Bot 54:513–527
Dahdouh-Guebas F, Hettiarachchi S, Lo Seen D, Batelaan S,
Sooriyarachchi S, Jayatissa LP, Koedam N (2005) Tran-
sitions in ancient inland freshwater resource management
in Sri Lanka affect biota and human populations in and
around coastal lagoons. Curr Biol 15:579–586
Damastuti E, de Groot R (2017) Effectiveness of community-
based mangrove management for sustainable resource use
and livelihood support: a case study of four villages in
Central Java, Indonesia. J Environ Manage 203:510–521
Epps M (2007) Socioeconomic baseline assessment: Imple-
menting the socioeconomic monitoring guidelines in
southwest madagascar. Blue Ventures Conservation,
London, UK
De Puydt ML (1868) Account of scientific exploration in the
Isthmus of Darien in the years 1861 and 1865. J R Geogr
Soc Lond 38:69–110
Feka NZ (2015) Sustainable management of mangrove forests
in West Africa: a new policy perspective? Ocean Coast
Manag 116:341–352
Feka NZ, Ajonina GN (2011) Drivers causing decline of man-
grove in West-Central Africa: a review. Int J Biodivers Sci
Ecosyst Serv Manag 7:217–230
Feka NZ, Morrison I (2017) Managing mangroves for coastal
ecosystems change: a decade and beyond of conservation
experiences and lessons for and from west-central Africa.
J Ecol Nat Environ 9:99–123
Gaudian G, Koyo A, Wells S (1995) Marine Region 12: East
Africa. In: Kelleher G, Bleakley C, Wells S (eds) A global
123
Wetlands Ecol Manage (2019) 27:609–625 623

representative system of marine protected areas. Central
Indian Ocean, Arabian Seas, East Africa and East Asian
Seas, vol III. Great Barrier Reef Marine Park Authority,
The World Bank, The World Conservation Union (IUCN).
The World Bank Environment Department, Washington
DC
Giri C, Mulhausen J (2008) Mangrove forest distribution and
dynamics in Madagascar. Sensors 8:2104–2117
Hamilton SE, Casey D (2016) Creation of a high spatio-tem-
poral resolution global database of continuous mangrove
forest cover for the 21st century (CGMFC-21). Glob Ecol
Biogeogr 25:729–738
Hamilton LS, Snedaker SC (1984) Handbook for mangrove area
management. IUCN Honolulu, Honolulu
Harris AR (2011) Out of sight but no longer out of mind: a
climate of change for marine conservation in Madagascar.
Madag Conserv Dev 6:7–14
Hauff RD, Ewel KC, Jack KC (2006) Tracking human distur-
bance in mangroves: estimating harvest rates on a
Micronesian Island. Wetl Ecol Manage 14:95–105
Hutchison J, Manica A, Swetnam R, Balmford A, Spalding M
(2014) Predicting global patterns in mangrove forest bio-
mass. Conserv Lett 7:233–240
Jones TG, Rakoto Ratsimba H, Ravaoarinorotsihoarana L,
Cripps G, Bey A (2014) Ecological variability and carbon
stock estimates of mangrove ecosystems in northwestern
Madagascar. Forests 5:177–205
Jones TG, Glass L, Gandhi S, Ravaoarinorotsihoarana L, Carro
A, Benson L, Ratsimba HR, Giri C, Randriamanatena D,
Cripps G (2016) Madagascar’s mangroves: quantifying
nation-wide and ecosystem specific dynamics, and detailed
contemporary mapping of distinct ecosystems. Remote
Sens 8:106
Kairo JG, Dahdouh-Guebas F, Gwada PO, Ochieng C, Koedam
N (2002) Regeneration status of mangrove forests in Mida
Creek, Kenya: a compromised or secured future? Ambio
31:562–568
Kauffman JB, Donato DC (2012) Protocols for the measure-
ment, monitoring and reporting of structure, biomass and
carbon stocks in mangrove forests. Working Paper 86.
CIFOR, Bogota
Komiyama A, Ong JE, Poungparnc S (2008) Allometry, bio-
mass, and productivity of mangrove forests: A review.
Aquat Bot 89:128–137
Krauss KW, Keeland BD, Allen JA, Ewel KC, Johnson DJ
(2007) Effects of season, rainfall, and hydrogeomorphic
setting on mangrove tree growth in Micronesia. Biotropica
39:161–170
Laroche J, Baran E, Rasoanandrasana NB (1997) Temporal
patterns in a fish assemblage of a semiarid mangrove zone
in Madagascar. J Fish Biol 51:3–20
Lau JD, Scales IR (2016) Identity, subjectivity and natural
resource use: how ethnicity, gender and class intersect to
influence mangrove oyster harvesting in The Gambia.
Geoforum 69:136–146
Lemmon PE (1956) A spherical densiometer for estimating
forest overstory density. For Sci 2:314–320
Leong RC, Friess DA, Crase B, Lee WK, Webb EL (2018)
High-resolution pattern of mangrove species distribution is
controlled by surface elevation. Estuar Coast Shelf Sci
202:185–192
Lopez-Hoffman L, Monroe IE, Narvaez E, Martinez-Ramos M,
Ackerly DD (2006) Sustainability of mangrove harvesting:
how do harvesters’ perceptions differ from ecological
analysis? Ecol Soc 11:14
McCracken A, Pretty W, Conway GR (1988) An introduction to
rapid rural appraisal for agricultural development. Inter-
national Institute for Environment and Development,
London
Moriizumi Y, Matsui N, Hondo H (2010) Simplified life cycle
assessment of mangrove management: a case of plantation
on wastelands in Thailand. J Clean Prod 18:1629–1638
Muller-Dombois D, Ellenberg H (1974) Aims and Methods of
Vegetation Ecology. Wiley, New York
Newing H (2011) Conducting research in conservation: a social
science perspective. Routledge, London
Oo N (2002) Present state and problems of mangrove manage-
ment in Myanmar. Trees 16:218–223
Ong JE, Gong WK, Wong CH (2004) Allometry and parti-
tioning of the mangrove, Rhizophora apiculata. For Ecol
Manag 188:395–408
Palacios ML, Cantera JR (2017) Mangrove timber use as an
ecosystem service in the Colombian Pacific. Hydrobiologia
803:345–358
Peabody S, Jones B (2013) Plan d’Amenagement et de Gestion
de la NAP Velondriake. Blue Ventures Conservation,
London
Polidoro BA, Carpenter KE, Collins L, Duke NC, Ellison AM,
Ellison JC, Farnsworth EJ, Fernando ES, Kathiresan K,
Koedam NE, Livingstone SR, Miyagi T, Moore GE, Nam
VN, Ong JE, Primavera JH, Salmo SG, Sanciangco JC,
Sukardjo S, Wang Y, Yong JWH (2010) The loss of spe-
cies: mangrove extinction risk and geographic areas of
global concern. PLoS ONE 5:e10095
Pratt B, Loizos P (1992) Choosing research methods: data col-
lection for development workers. Oxfam, Oxford
Primavera J (2001) Community-based rehabilitation of man-
groves. In: Utilizing different aquatic resources for liveli-
hoods in Asia: a resource book. The World Fish Center,
Dhaka, pp 394–402
Putz FE, Chan HT (1986) Tree growth, dynamics, and produc-
tivity in a mature mangrove forest in Malaysia. For Ecol
Manag 17:211–230
Rahman MM, Khan MNI, Hoquelmran AKF, Ahmed I (2015)
Carbon stock in the Sundarbans mangrove forest: spatial
variations in vegetation types and salinity zones. Wetl Ecol
Manag 23:269–283
Rajkaran A, Adams JB, du Preez DR (2004) A method for
monitoring mangrove harvesting at the Mngazana estuary,
South Africa. Afr J Aquat Sci 29:57–65
Rakotomavo A, Fromard F (2010) Dynamics of mangrove for-
ests in the Mangoky River delta, Madagascar, under the
influence of natural and human factors. For Ecol Manage
259:1161–1169
Rasolofo MV (1997) Use of mangroves by traditional fishermen
in Madagascar. Mangroves Saltmarshes 1:243–253
Sanders CJ, Maher DT, Tait DR, Williams D, Holloway C,
Sippo JZ, Santos IR (2016) Are global mangrove carbon
stocks driven by rainfall? J Geophys Res. https://doi.org/
10.1002/2016jg003510
Scales IR, Friess DA, Glass L, Ravaoarinorotsihoarana L (2018)
Rural livelihoods and mangrove degradation in south-west
123
624 Wetlands Ecol Manage (2019) 27:609–625

Madagascar: lime production as an emerging threat. Oryx
52:641–645
Seddon N, Tobias J, Yount JW, Ramanampamonjy JR,
Butchart S, Randrianizahana H (2000) Conservation issues
and priorities in the Mikea Forest of south-west Mada-
gascar. Oryx 34:287–304
Simard M, Fatoyinbo L, Smetanka C, Rivera-Monroy VH,
Castaneda-Moya E, Thomas N, Van der Stocken T (2019)
Mangrove canopy height globally related to precipitation,
temperature and cyclone frequency. Nat Geosci 12:40–45
Spalding M, Kainuma M, Collins L (2010) World atlas of
mangroves. Earthscan, London
Sudtongkong C, Webb EL (2008) Outcomes of state- vs. com-
munity-based mangrove management in southern Thai-
land. Ecol Soc 13:27
Thomas N, Lucas R, Bunting P, Hardy A, Rosenqvist A, Simard
M (2017) Distribution and drivers of global mangrove
forest change, 1996–2010. PLoS ONE 12:e0179302
UNEP (2014) The importance of mangroves to people: a call to
action. United Nations Environment Programme, World
Conservation Monitoring Centre, Cambridge
Walters BB (2005a) Ecological effects of small-scale cutting of
Philippine mangrove forests. For Ecol Manage
206:331–348
Walters BB (2005b) Patterns of local wood use and cutting of
Philippine mangrove forests. Econ Bot 59:66–76
Walters BB, Ronnback P, Kovacs JM, Crona B, Hussain S,
Badola R, Primavera JH, Barbier E, Dahdouh-Guebas F
(2008) Ethnobiology, socio-economics and management
of mangrove forests: a review. Aquat Bot 89:220–236
Whitfield AK (2017) The role of seagrass meadows, mangrove
forests, salt marshes and reed beds as nursery areas and
food sources for fishes in estuaries. Rev Fish Biol Fisheries
27:75–110
World Bank (2016) Development goals in an era of demo-
graphic change. World Bank, Washington DC
Publisher’s Note Springer Nature remains neutral with
regard to jurisdictional claims in published maps and
institutional affiliations.
123
Wetlands Ecol Manage (2019) 27:609–625 625




















