
PALYNOLOGICAL RECORDS OF CLIMATE AND OCEANIC CONDITIONS IN THE LATE PLEISTOCENE AND HOLOCENE OF THE...
24
PALYNOLOGICAL RECORDS OF CLIMATE AND OCEANIC CONDITIONS IN THE LATE PLEISTOCENE AND HOLOCENE OF THE NILE CONE, SOUTHEASTERN MEDITERRANEAN, EGYPT SUZAN E.A. KHOLEIF National Institute of Oceanography and Fisheries (NIOF) Qayed Bay Alexandria Egypt e-mail: [email protected] PETA J. MUDIE Natural Resources Canada Geological Survey Canada Atlantic Dartmouth Nova Scotia B2Y 4A2 Canada Corresponding author, e-mail: [email protected] Abstract Variations in palynomorph concentrations and relative abundances, including pollen and spores, organic-walled dinoflagellate cysts, and amorphous organic matter (AOM), were studied in sediment core NC Core 2 from the upper slope (1,030 m water depth) of the Nile Cone, southeastern Mediterranean, Egypt. Each sample represents palynomorph deposition for intervals of around 500–1,000 years, and this study provides the first detailed palynological record for the Nile Cone. The dinoflagellate cyst assemblages and AOM production reflect changes in surface water temperature and river discharge in response to climate forcing as marked by variations in pollen and spore indicators of semi-desert vegetation and tropical Monsoon intensity. The basal sediments (approximately 14,600–9,500 yr before present [BP]) contain high concentrations and relative abundances of Polysphaeridium zoharyi, Polysphaeridium spp., Spiniferites elongatus, and Spiniferites spp. during the transition from postglacial to interglacial conditions. Variations in these taxa and total Impagidinium spp. suggest a gradual temperature increase, interrupted by a pre-Holocene cool interval around 11,000 yr BP. The high African monsoon index for this pre- Holocene time corresponds with pollen evidence of relatively high Nile discharge. The organic-rich S1 sapropel sediments, dated as around 9,500–6,270 yr BP, were deposited several thousand years later, during a time of lower Monsoon index, warmer surface water, and increased dinoflagellate cyst production. This was accompanied by moderate Nile flooding and oxygen-deficient or anoxic bottom water on the upper Nile Cone, mainly supporting the increased production and carbon import hypothesis for eastern Mediterranean S1 formation. Dinoflagellate cyst assemblages in the Nile Cone S1 differ from those of the deeper, more northern Levantine and Cretan basins in the near absence of heterotrophic protoperidinioid cysts, despite the uniformly high organic S1 carbon content. The prevailing climate during the formation of sapropelic sediments in the southeastern Mediterranean was tropical to subtropical, but was interrupted by an early cooler interval with reduced humidity, as marked by re-appearance of Spiniferites elongatus and increased Ephedra pollen. The carbonate-rich surface layers were probably deposited under warm dry climatic conditions with minimum Nile input, but evidence of oxidation prevents detailed interpretation. Key words: dinoflagellate cysts; pollen; spores; amorphous organic matter; sapropel S1; Nile Cone; Late Pleistocene; Holocene. Palynology, 33 (2009): 1–24 © 2009 by AASP Foundation ISSN 0191-6122 INTRODUCTION The Nile River, which drains almost 10% of Africa, is a major feature of the southeastern Mediterranean region, and its sediments cover about half of the Levantine Basin in the eastern Mediterranean Sea (Text-Figure 1). The Nile Cone is the largest of the Mediterranean deep-sea fans. This sedimentary feature differs from most other large subma-
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of PALYNOLOGICAL RECORDS OF CLIMATE AND OCEANIC CONDITIONS IN THE LATE PLEISTOCENE AND HOLOCENE OF THE...

S.E.A. Kholeif, and P.J. Mudie: Palynological records of Pleistocene and Holocene, conditions, Nile Cone, Egypt 1PALYNOLOGICAL RECORDS OF CLIMATEAND OCEANIC CONDITIONS IN THE LATEPLEISTOCENE AND HOLOCENE OF THE NILECONE, SOUTHEASTERN MEDITERRANEAN, EGYPT
SUZAN E.A. KHOLEIFNational Institute of Oceanography and Fisheries (NIOF)Qayed BayAlexandriaEgypte-mail: [email protected]
PETA J. MUDIENatural Resources CanadaGeological Survey Canada AtlanticDartmouthNova Scotia B2Y 4A2CanadaCorresponding author, e-mail: [email protected]
Abstract
Variations in palynomorph concentrations and relative abundances, including pollen and spores, organic-walled dinoflagellate cysts, andamorphous organic matter (AOM), were studied in sediment core NC Core 2 from the upper slope (1,030 m water depth) of the Nile Cone,southeastern Mediterranean, Egypt. Each sample represents palynomorph deposition for intervals of around 500–1,000 years, and this studyprovides the first detailed palynological record for the Nile Cone. The dinoflagellate cyst assemblages and AOM production reflect changesin surface water temperature and river discharge in response to climate forcing as marked by variations in pollen and spore indicators ofsemi-desert vegetation and tropical Monsoon intensity. The basal sediments (approximately 14,600–9,500 yr before present [BP]) containhigh concentrations and relative abundances of Polysphaeridium zoharyi, Polysphaeridium spp., Spiniferites elongatus, and Spiniferitesspp. during the transition from postglacial to interglacial conditions. Variations in these taxa and total Impagidinium spp. suggest a gradualtemperature increase, interrupted by a pre-Holocene cool interval around 11,000 yr BP. The high African monsoon index for this pre-Holocene time corresponds with pollen evidence of relatively high Nile discharge. The organic-rich S1 sapropel sediments, dated as around9,500–6,270 yr BP, were deposited several thousand years later, during a time of lower Monsoon index, warmer surface water, and increaseddinoflagellate cyst production. This was accompanied by moderate Nile flooding and oxygen-deficient or anoxic bottom water on the upperNile Cone, mainly supporting the increased production and carbon import hypothesis for eastern Mediterranean S1 formation.Dinoflagellate cyst assemblages in the Nile Cone S1 differ from those of the deeper, more northern Levantine and Cretan basins in the nearabsence of heterotrophic protoperidinioid cysts, despite the uniformly high organic S1 carbon content. The prevailing climate during theformation of sapropelic sediments in the southeastern Mediterranean was tropical to subtropical, but was interrupted by an early coolerinterval with reduced humidity, as marked by re-appearance of Spiniferites elongatus and increased Ephedra pollen. The carbonate-richsurface layers were probably deposited under warm dry climatic conditions with minimum Nile input, but evidence of oxidation preventsdetailed interpretation.
Key words: dinoflagellate cysts; pollen; spores; amorphous organic matter; sapropel S1; Nile Cone; Late Pleistocene;Holocene.
Palynology, 33 (2009): 1–24© 2009 by AASP Foundation ISSN 0191-6122
INTRODUCTION
The Nile River, which drains almost 10% of Africa, is amajor feature of the southeastern Mediterranean region,
and its sediments cover about half of the Levantine Basinin the eastern Mediterranean Sea (Text-Figure 1). The NileCone is the largest of the Mediterranean deep-sea fans. Thissedimentary feature differs from most other large subma-

2 PALYNOLOGY, VOLUME 33 — 2009
rine fans because it developed in a relatively small, en-closed sea, and sedimentation rates are high (Stanley andMaldonado, 1977). The proximal sediments of the NileCone are therefore expected to provide a sensitive record ofeastern Mediterranean climate change, Nile River flow,and corresponding changes in ocean circulation and strati-fication.
The southeastern Mediterranean region lies at the eastend of the transition between the humid, temperate climatesof Europe to the north and west, and the African–Asian hot,arid climate zones to the south and east. The climate historyof the region largely reflects the interaction between thejetstream-dominated European trade-wind system, and themonsoon-influenced North African–Asian climate sys-tems (Rohling and Hilgen, 1991; Scrivner et al., 2004). Theprevailing airmasses that control the Mediterranean cli-mate (Text-Figure 2A) are maritime polar or Arctic sys-tems sweeping south over Europe in winter (60%), andcontinental tropical airmasses (14%) from the Sahara and
northwest Africa in summer. Periods of intense wintercyclogenesis (“meteorological bombs”) also characterizethe central and eastern Mediterranean (Lionello et al.,2006). The present Mediterranean region receives onlyaround 450–500 mm annual rainfall between October andApril (Riley and Spolton, 1974). In southern Egypt, Sudan,and the headwaters of the Nile River, rainfall is controlledby the African and South Asian Monsoon systems (Alpertet al., 2006; Hassan, 2007), and annual rain ranges from lessthan 300 mm a-1 in Sudan to more than 750 mm a-1 inEthiopia (Ritchie, 1987; Hassan, 2007). Fresh water inEgypt thus depends heavily on the Nile flood volume,which is determined mainly by rainfall in the EthiopianHighlands (summer flood maxima) and to a lesser extent bythe flow in the Equatorial African tributaries (Williams etal., 2006).
Sapropelic sediment layers with extremely high or-ganic matter content (2–13% dry weight) are a distinctivefeature of the Pliocene–Pleistocene sediments from the
Text-Figure 1. A) Map of the Mediterranean Sea, with the square marking the location of the area studied off the delta of the NileRiver, in the Levantine Basin. B) Bathymetry (in m) and core locations in the area studied. P6150-4 and NC Core 2 are the studycores; the other letters indicate 14 of the 65 cores studied by Stanley and Maldonado (1977). The map is modified from Stanleyand Maldonado (1977) with the permission of these authors.

S.E.A. Kholeif, and P.J. Mudie: Palynological records of Pleistocene and Holocene, conditions, Nile Cone, Egypt 3
Text-Figure 2. Maps of the Mediterranean Sea showing weather systems and the circulation of water masses. A) Areas ofcyclogenesis, cyclone depression tracks, air mass sources, and their annual frequency in the Mediterranean Seas (adapted fromRiley and Spolton, 1974); c = continental, m = maritime, A =Arctic, P =Polar, T =Tropical. B) Simplified diagram of watercirculation before the Eastern Mediterranean Transient (EMT), when Atlantic Water (AW) formed a third of Levantine BasinIntermediate Water (LIW), and the Adriatic was the main source of Eastern Mediterranean Deep Water (EMDW). Black arrows= surface water; gray arrows = deep water. C) Simplified circulation during the EMT, showing AW is deflected northwards andLIW is deflected into the Cretan Basin forming EMDW (from Tsimplis et al., 2006). LSC = Levantine Shelf Current. Black arrows= surface water; light gray arrows = deep water, dark gray arrows = shelf current.

4 PALYNOLOGY, VOLUME 33 — 2009
Levantine Basin. They have been interpreted as indicat-ing large Nile flood intervals during the Late Pleistoceneto Holocene (Rossignol-Strick, 1983; 1985). However,the exact causes of Mediterranean sapropel formation arestill controversial (Murat and Got, 2000; Bianchi et al.,2006). The sapropel layers, and the flood intervals thatapparently led to their formation, have often been corre-lated with orbitally-driven climate cycles and periodicAfrican monsoon intensification (Rossignol-Strick, 1985;Gasse, 2000; Scrivner et al., 2004). Ducassou et al. (2007)also reported that turbidite deposits in the Nile Fan werecontrolled by monsoon variability in the Nile catchmentand sea level changes. Neodymium studies of the Niletributaries and of foraminifera in northern LevantineBasin cores support the theory that the Holocene sapropellayer S1 partly reflects increased Nile River discharge(Scrivner et al., 2004). However there is also evidence ofwesterly freshwater influences. In contrast, strontiumisotopes and titanium/aluminium ratios in bulk core sedi-ments indicate reduced flow to the Nile Delta during S1deposition (Krom et al., 2002).
Rossignol-Strick (1983) analyzed low diversity pollenassemblages from one to four samples in each of ninesapropels in a deep sea core from west of the Nile Cone. Itwas concluded that these assemblages, dominated by Arte-misia and Pinus pollen, recorded northward shifts of theinter-tropical convergence zone (ITCZ) and increased Nilerunoff during intensified monsoon events. Cheddadi andRossignol-Strick (1995), however, stated that the occur-rence of pollen in the sapropels of cores from shallowerwater (745–1,260 m) of the eastern Nile Cone mostlyreflects the enhanced preservation of organic matter ratherthan climatic change. Consequently, details of the relation-ship between climate, sapropel formation, and Nile runoffare uncertain, although the low resolution model ofRossignol-Strick (1983) is often cited to support the chro-nology of monsoon rain history and sapropels (Lamb et al.,2007).
This study used NC Core 2 from the upper eastern NileCone (Text-Figure 1B) to examine the integrated paleo-ecological signals from terrigenous palynomorphs (pol-len and spores) and marine palynomorphs (dinoflagellatecysts and amorphous organic matter [AOM]). These dataare then used to interpret the main events associated withchanges in paleoclimate, paleooceanography, and NileRiver discharge during the past 14,600 years. This is thefirst report on latest Pleistocene and Holocene dinoflagel-late cysts of the Nile Cone, and it is the first millennial-scale study of variations in pollen–spore concentrationsand AOM in marine sediments off the Nile Delta. Thelocation of NC Core 2 at 1,030 m water depth, proximalto the Nile Delta but below the direct effects of postglacial
sea level and pycnocline (water density boundary) oscil-lations, makes it an important target for paleoceanographicstudies of the influence of the Nile River flood dischargeto the eastern Mediterranean Sea.
ENVIRONMENTAL CONDITIONSAND SAPROPEL FORMATION
The modern Nile River has an average discharge ofapproximately 3 x 109 m3 per day (Ministry of WaterResources and Irrigation, 1993). The discharge curve isunimodal, with summer floods being linked to heavy mon-soon rainfall over the Ethiopian highlands and the Equato-rial Plateau, including the Central African Republic, Congo,Kenya, Rwanda, and Uganda (Williams et al., 2006; Hassan,2007). Before the construction of the Aswan High Dam in1964, the discharge was 4–8 x 1010 m3 per year (Wadie,1984), but this range has decreased to 2.5–4.0 x 109 m3 peryear. Also, before the Aswan High Dam, the river periodi-cally discharged a large load of dissolved and silt-adsorbednutrients through the Damietta and Rosetta channels (Text-Figure 1B) into the hyper-oligotrophic Levantine Basinduring the flood season from July to October. Throughoutthe Quaternary, the Nile River has contributed huge vol-umes of sediment to both the Egyptian continental shelf,and to the Nile Cone (Summerhayes et al., 1978; Stanleyand Maldonado, 1979).
Oceanographic factors, including temperature, salinity,and current circulation (Text-Figures 2B, C), also affect thedistribution and deposition of the Nile Cone and deep seafan sediments. The annual temperature changes of thesurface waters in the Mediterranean Sea are large andcontrol the density and other physical characteristics of thewater masses. Interannual variability of water mass pro-duction results from atmospheric forcing, including ElNiño events, and is particularly intense in the LevantineBasin (Pinardi and Mosetti, 2000). Sea surface temperature(SST) in the eastern Mediterranean Sea is 12–32°C, withminima during winter in the northern basins, and maximaduring summer in the southeastern Levantine Basin(Bartzokas et al., 1991). The Mediterranean Sea has anegative hydrological balance, with evaporative loss greatlyexceeding runoff and precipitation. Low salinity Atlanticwater (AW) entering in the upper layer of the GibraltarStrait is transformed to saline Mediterranean water thatsubsequently exits into the Atlantic via the lower layer ofLevantine intermediate water (LIW) and other bottomwater masses (Lascaratos et al., 1999). During its eastwardtravel, the salinity of AW rises from 36.15 practical salinityunits (psu) at the Straits of Gibraltar to either a maximumof 38.6 psu in the Levantine Basin (pre-1989 conditionsaccording to Ozsoy et al., 1989), or to less than 38.0 psu at

S.E.A. Kholeif, and P.J. Mudie: Palynological records of Pleistocene and Holocene, conditions, Nile Cone, Egypt 5
the Straits of Sicilian during the present circulation condi-tions (Tsimplis et al., 2006).
During the summer, the surface layer of the LevantineSea is occupied by warm, salty Levantine surface water(LSW). Winter cooling increases the density of the LSW,so it sinks and mixes with underlying water of the Rhodescyclonic gyre and contributes to LIW, which is a highsalinity layer between 200 and 500 m depth within thenorthwestern Levantine Basin (Lascaratos et al., 1999;Pinardi and Mosetti, 2000).
The core used in this study of the Nile Cone, NC Core 2,is from the upper part of eastern Mediterranean deep water(EMDW) that fills the eastern basins below the base of theLIW at around 500 m. The EMDW temperature and salinityrange from 13.0–13.7°C and 38.6–38.0 psu respectively,changing gradually towards the Ionian and Levantine ba-sins by mixing of Adriatic deep water with LIW and/orCretan water (Text-Figures 2B, C). Classically, EMDWwas thought to be formed mostly in the southern AdriaticSea (Text-Figure 2B), with the input of dense NorthernAdriatic waters (Lascaratos et al., 1999; Tsimplis et al.,2006). Since 1987, however, profound circulation changeshave been observed (Tsimplis et al., 2006), including anorthward deflection of AW in the western Mediterranean,and outflow of new water from the Cretan Basin. Thiscirculation pattern is called the eastern Mediterraneantransient (EMT; Text-Figure 2C), and may characterizewarmer, drier climatic conditions in the southeastern Medi-terranean.
The eastern Mediterranean sapropel S1 has been radio-carbon-dated in numerous studies (Stanley and Maldonado,1977; Murat and Got, 2000), and has an age of approxi-mately 9,500 to 6,000 yr before present (BP) throughoutthe Levantine Basin. Rossignol-Strick (1983; 1985) usedpalynological data to show a general coincidence betweeneastern Mediterranean sapropel formation, Nile Riverfloods, orbital-scale (multimillenial) insolation changes,and high African Monsoon index (2IT
– IE, where I isinsolation, and T and E are north Tropic and Equator). Thechanges in the Monsoon index correspond to about a 0.75°latitude northerly shift in the ITCZ and greater humidity,but the timing of the insolation changes precedes that ofsapropel formation by around 3,000 years. Béthoux (1993)considered that S1 formation was preconditioned by thecessation of dense water formation in the Adriatic Seabecause of changes in sea level and/or freshwater input.Murat and Got (2000) provided a comprehensive reviewof factors that have been attributed to the formation of thisEarly Holocene sapropel, including nutrient transportdynamics, plankton production and export, marine circu-lation and stratification, and carbon preservation. Theseauthors concluded that sapropels formed during Early
Holocene basinwide stagnation below around 400 m,coupled with decreased oxidation flux at the sediment–water interface. This interpretation is supported by thecomputer model of Bianchi et al. (2006), but the role ofNile River flooding is still debated.
This paper aims primarily to reconstruct millennial-scalefluctuations in climate and Nile discharge over the upperNile Cone during the latest Pleistocene and Holocene,using dinoflagellate cysts as markers of changes in SST,salinity, and bottom water oxidation. In addition, pollen–spore concentrations and abundance of Nile indicator spe-cies are used as proxy-signals for offshore transport of fine-grained particulate organic matter to the upper Nile Coneby wind and water. The dinoflagellate cyst record for theupper Nile Cone is considered particularly important be-cause of the widespread distribution of dinoflagellate cystsfrom continental shelf to deep sea basins (Harland, 1988),and their observed correlation with SST and salinity in theAtlantic and Indian oceans (Dale, 1996; Rochon et al.,1999; Marret and Zonneveld, 2003). Furthermore, organic-walled dinoflagellate cysts are highly resistant to degrada-tion by mechanical transport and chemical dissolution, andtheir response to oxidation is known for the westernLevantine Basin (Zonneveld et al., 2001). Accordingly,dinoflagellate cysts have advantages over the previous useof calcareous and siliceous microfossils forpaleoceanographic studies of the Nile Cone, and they are avaluable data source for investigation of changes in Quater-nary paleoclimate and ocean water masses in the easternMediterranean (Morzadec-Kerfourn, 1988; Aksu et al.,1995; Zonneveld, 1996) and the adjoining inland seas(Mudie et al., 2002a).
MATERIALS AND METHODS
Introduction and Sediment Analysis
The Nile Cone core studied (NC Core 2), is about 170 cmlong, and was taken from the upper slope at a depth of 1,030m (Text-Figure 1), using a 5 cm diameter gravity corer,from the Russian Vessel R/V Academic Levrantive inDecember 1988. The core location is 32° 20' 42''N, 31° 39'0''E. Geochemical and sedimentological analyses of NCCore 2 were made soon thereafter by M. Kahil (Table 1),using large samples, of which a portion was saved for thispalynological study. The 1988 sediment studies used stan-dard grain size analysis sieving, settling tube, and weighingtechniques. Total organic carbon (TOC wt %) and calciumcarbonate (CaCO3 %) were measured by wet oxidation(Gross, 1975) and the potassium dichromate–sulfuric acidmethod (Walkley and Black, 1934), respectively. The corecontained six sediment units (Text-Figure 3). Unit I (165–
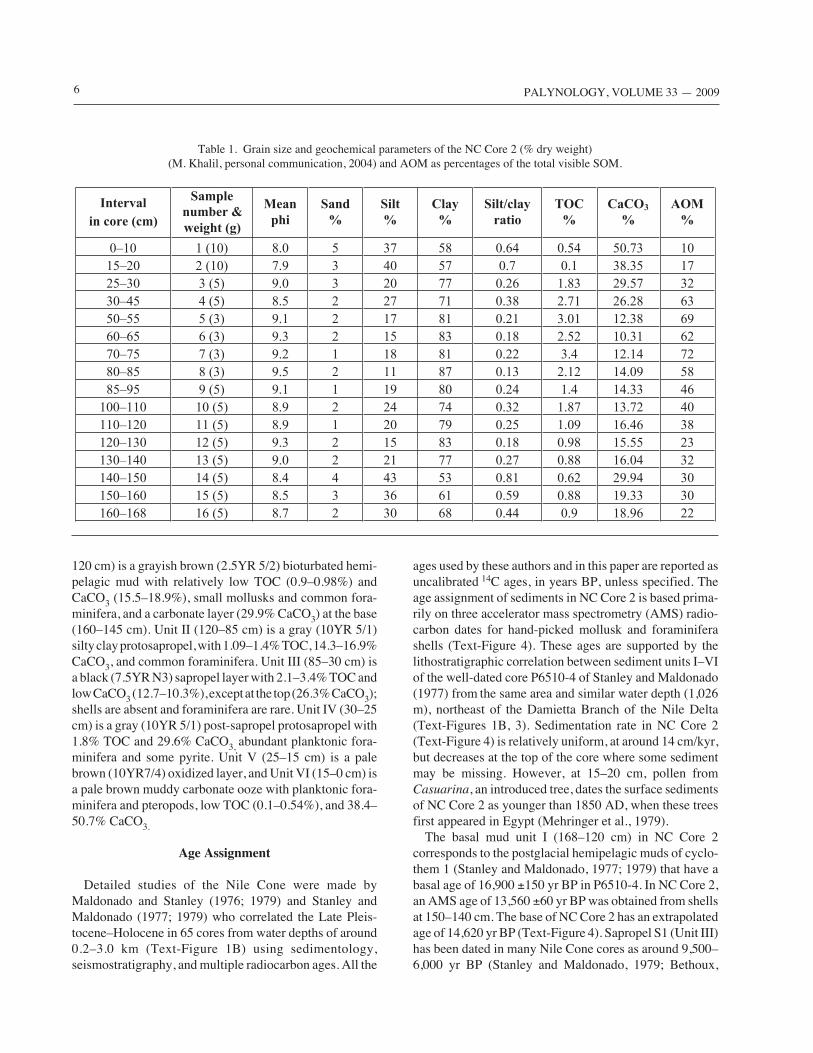
6 PALYNOLOGY, VOLUME 33 — 2009
120 cm) is a grayish brown (2.5YR 5/2) bioturbated hemi-pelagic mud with relatively low TOC (0.9–0.98%) andCaCO3 (15.5–18.9%), small mollusks and common fora-minifera, and a carbonate layer (29.9% CaCO3) at the base(160–145 cm). Unit II (120–85 cm) is a gray (10YR 5/1)silty clay protosapropel, with 1.09–1.4% TOC, 14.3–16.9%CaCO3, and common foraminifera. Unit III (85–30 cm) isa black (7.5YR N3) sapropel layer with 2.1–3.4% TOC andlow CaCO3 (12.7–10.3%), except at the top (26.3% CaCO3);shells are absent and foraminifera are rare. Unit IV (30–25cm) is a gray (10YR 5/1) post-sapropel protosapropel with1.8% TOC and 29.6% CaCO3, abundant planktonic fora-minifera and some pyrite. Unit V (25–15 cm) is a palebrown (10YR7/4) oxidized layer, and Unit VI (15–0 cm) isa pale brown muddy carbonate ooze with planktonic fora-minifera and pteropods, low TOC (0.1–0.54%), and 38.4–50.7% CaCO3.
Age Assignment
Detailed studies of the Nile Cone were made byMaldonado and Stanley (1976; 1979) and Stanley andMaldonado (1977; 1979) who correlated the Late Pleis-tocene–Holocene in 65 cores from water depths of around0.2–3.0 km (Text-Figure 1B) using sedimentology,seismostratigraphy, and multiple radiocarbon ages. All the
ages used by these authors and in this paper are reported asuncalibrated
14C ages, in years BP, unless specified. Theage assignment of sediments in NC Core 2 is based prima-rily on three accelerator mass spectrometry (AMS) radio-carbon dates for hand-picked mollusk and foraminiferashells (Text-Figure 4). These ages are supported by thelithostratigraphic correlation between sediment units I–VIof the well-dated core P6510-4 of Stanley and Maldonado(1977) from the same area and similar water depth (1,026m), northeast of the Damietta Branch of the Nile Delta(Text-Figures 1B, 3). Sedimentation rate in NC Core 2(Text-Figure 4) is relatively uniform, at around 14 cm/kyr,but decreases at the top of the core where some sedimentmay be missing. However, at 15–20 cm, pollen fromCasuarina, an introduced tree, dates the surface sedimentsof NC Core 2 as younger than 1850 AD, when these treesfirst appeared in Egypt (Mehringer et al., 1979).
The basal mud unit I (168–120 cm) in NC Core 2corresponds to the postglacial hemipelagic muds of cyclo-them 1 (Stanley and Maldonado, 1977; 1979) that have abasal age of 16,900 ±150 yr BP in P6510-4. In NC Core 2,an AMS age of 13,560 ±60 yr BP was obtained from shellsat 150–140 cm. The base of NC Core 2 has an extrapolatedage of 14,620 yr BP (Text-Figure 4). Sapropel S1 (Unit III)has been dated in many Nile Cone cores as around 9,500–6,000 yr BP (Stanley and Maldonado, 1979; Bethoux,
Intervalin core (cm)
Samplenumber &weight (g)
Meanphi
Sand%
Silt%
Clay%
Silt/clayratio
TOC%
CaCO3
%AOM
%
0–10 1 (10) 8.0 5 37 58 0.64 0.54 50.73 1015–20 2 (10) 7.9 3 40 57 0.7 0.1 38.35 1725–30 3 (5) 9.0 3 20 77 0.26 1.83 29.57 3230–45 4 (5) 8.5 2 27 71 0.38 2.71 26.28 6350–55 5 (3) 9.1 2 17 81 0.21 3.01 12.38 6960–65 6 (3) 9.3 2 15 83 0.18 2.52 10.31 6270–75 7 (3) 9.2 1 18 81 0.22 3.4 12.14 7280–85 8 (3) 9.5 2 11 87 0.13 2.12 14.09 5885–95 9 (5) 9.1 1 19 80 0.24 1.4 14.33 46
100–110 10 (5) 8.9 2 24 74 0.32 1.87 13.72 40110–120 11 (5) 8.9 1 20 79 0.25 1.09 16.46 38120–130 12 (5) 9.3 2 15 83 0.18 0.98 15.55 23130–140 13 (5) 9.0 2 21 77 0.27 0.88 16.04 32140–150 14 (5) 8.4 4 43 53 0.81 0.62 29.94 30150–160 15 (5) 8.5 3 36 61 0.59 0.88 19.33 30160–168 16 (5) 8.7 2 30 68 0.44 0.9 18.96 22
Table 1. Grain size and geochemical parameters of the NC Core 2 (% dry weight)(M. Khalil, personal communication, 2004) and AOM as percentages of the total visible SOM.

S.E.A. Kholeif, and P.J. Mudie: Palynological records of Pleistocene and Holocene, conditions, Nile Cone, Egypt 7
1993; Emeis et al., 2000; Martinez-Ruiz et al., 2003), andthis unit in NC Core 2 has an AMS shell age of 6270 ±40yr BP at the top (30–45 cm). Unit V (Unit OL1 in northeast-ern Nile Cone cores) has an age of 5,800–2,800 yr BP inP6510-4 (Stanley and Maldonado, 1977; 1979). Unit VI inP6510-4 and the NC Core2 is a bioturbated, muddy fora-miniferal ooze, with basal ages of 2,700–2,800 yr BP inP6510-4 and 3,270 ±40 yr BP in NC Core 2.
Preparation and Analytical Methods
Sixteen samples were prepared, covering 5–15 cm inthe NC Core2 core (Table 1). The samples were either
contiguous, or separated by 5 cm intervals (Table 1). Thedry weights ranged from 3–10 g and they were processedusing standard techniques, including 10% hydrochloricacid (HCl) to dissolve carbonates, and cold 40–49%hydrofluoric acid (HF) to dissolve silicates, followed bytreatment with 34–40% HCl for up to 20 minutes toremove any fluoride precipitates. This was followed byshort ultrasonic cleaning and sieving on a 10 mm mesh.Glycerin jelly was used as a mounting medium. A Lyco-podium spike was added at the first step of preparation tocalculate the palynomorph concentrations. The concen-trations (specimens per gram) were determined using theLycopodium marker spores method of Birks and Birks
Text-Figure 3. Chronostratigraphic correlation of the NC Core 2 with the Nile Cone reference section of Stanley and Maldonado(1977), and Pillsbury core P6510-4. C1 to C4 are sedimentary cyclothems broadly corresponding to glacial-interstadial/interglacial periods. S1 to S3 are Mediterranean sapropel deposits.

8 PALYNOLOGY, VOLUME 33 — 2009
(1980). This is the palynomorphs counted multiplied bythe Lycopodium spores added, divided by the Lycopodiumspores counted multiplied by the sediment volume. Twoslides of each sample were examined using a Zeiss re-search microscope at magnifications of 400 and 1,000. Atotal of 450 palynomorphs were counted, with numbers ofpollen ranging from 77–333, and dinoflagellate cystsranging from 89–266. The dinoflagellate cyst taxonomyfollows Fensome et al. (1993), Rochon et al. (1999), andMarret and Zonneveld (2003).
Amorphous Organic Matter
The relative abundance of AOM (Text-Figure 5; Table 1)is calculated as a percentage of the total sedimentaryorganic matter (SOM) using the method of Tyson (1995).
It consists of all particulate organic components that appearstructureless under the light microscope (Plate 1, fig. 13),and includes phytoplankton- or bacterially-derived mate-rial (Tyson, 1995), plant resins, and the amorphous prod-ucts of the diagenesis of macrophyte tissues.
RESULTS
The NC Core 2 yielded rich dinoflagellate cyst assem-blages, terrestrial plant material, and AOM. Other marineor fresh/brackish water palynomorphs encountered includeacritarchs, prasinophytes, copepod eggs, and foraminiferallinings (Plate 1, fig. 9). The terrestrial sporomorphs (Kremp,1965) consist of pollen, fern and bryophyte spores, withassemblages mainly comprising bisaccate tree pollen, andherb/shrub pollen of the families Poaceae, Asteraceae, and
Text-Figure 4. Age-depth curve for the NC Core 2, showing the radiocarbon ages used to derive the sedimentation rates, theextrapolated ages cited in the text, and the line expected for a linear sedimentation rate. The inserted table and gray diamondsymbols indicate the three new AMS ages measured and used to estimate ages for the core top and base (black diamonds). Arrowsshow new 14C ages; diamonds without arrows are extrapolated ages. The triangle symbols are ages for the base of S1 in theLevantine Basin and the boundary of Units I/II as reported by Stanley and Maldonado (1979). The tabulated ages and isotope ratiosare from Beta Analytic; the conventional 14C ages are corrected for isotopic fractionation using the 13C/12C ratio. The calendarage calculations use the INTCAL04 calibration in Radicarbon 46(3), 2004.
Interval Sample Wt. Conventional Calendar Age
(cm) (g) C Age yr BP
Beta-248094Planktic & large benthic forams,
mollusks, pteropods
NILEC2-6
Beta -248093
NILEC2-10
Beta -248092Planktic & large benthic forams,
mollusks, pteropods
NILEC2-5140–150 0.4427
-0.4‰
30–45 0.0557Planktic forams, with some pyrite infill -1.52‰
Sample no. Material C/ C
3060–2750
15860–151701
6270 ± 4027 6790–6520
13560 ± 40
5–10 0.2124-0.3‰ 3270 ± 40
Core depth in cm
950010475
1316014620
31706270
0200040006000800010000120001400016000
0 20 40 60 80 100 120 140 160 180
Age
in k
yr B
P
14
1213
→
→→

S.E.A. Kholeif, and P.J. Mudie: Palynological records of Pleistocene and Holocene, conditions, Nile Cone, Egypt 9
Chenopodiaceae/Amaranthaceae (Plate 1, figs. 10, 1, and3, 4, respectively). As expected, given the offshore loca-tion of NC Core 2, the taxonomic diversity (N) of pollenand spores is low (N = 11) compared to the Egyptian coast(N = 24; Kholeif, 2007) and inland lakes (N = 46, Kholeif,2007). The AOM was produced by benthic filamentouscyanobacteria and pelagic sulfur bacteria in oxygen-defi-cient environments (Williams, 1984), and dominates sedi-ments deposited in bottom waters low in oxygen (Tyson,1995). In pre-Quaternary sediments, high AOM is associ-ated with reducing conditions, distal dysoxic–anoxic shelfenvironments, and high productivity (Batten, 1981; Tyson,1995). Total organic carbon is a direct measure of the totalamount of organic matter in sediments (Batten, 1996), andthere is normally a good correlation between AOM andTOC in dark, organic-rich (>1.0% TOC) shales depositedunder oxygen-deficient condition (Tyson, 1995; Batten,1996), and a strong positive correlation between TOC andAOM has been demonstrated in other Nile Cone studiessuch as Summerhayes et al. (1978).
In the NC Core 2, the grain sizes (Table 1) and sedimen-tation rates (Text-Figure 4) are relatively uniform throughmost of the core, with only minor increases in biogenic sandnear the base (150–140 cm) and at the top where thesedimentation rate decreases. The percentage weight TOCvalues mostly accord with the percentages of AOM (Text-
Figure 5). Both increase between 110 and 30 cm, coincid-ing with the darker pre-sapropelic and sapropel sedimentsin the intervals of less oxidizing conditions. Calcium car-bonate content is mostly relatively low (20–15%) from thebase to 55 cm, where it increases to 51% at 0–10 cm, closelymatching the relative abundances of total Impagidinium(Text-Figure 6). In contrast, the total dinoflagellate cystconcentration is mostly high (3,870–5,649 cysts g-1 sedi-ments) from the base to 85 cm, then is variable within thesapropel (around 5,000–16,000 cysts g-1), before decreas-ing to the top of the core (Table 2). Total sporomorphconcentrations (Text-Figure 5) are relatively low in UnitsI and II, but increase two- to three-fold at the top of the pre-sapropel, and remain high in Unit III and upwards into thepost-sapropel Units IV.
Ecostratigraphic assemblages
Based on palynomorph concentrations and relative abun-dances (percentages), and their relationship to the AOM,CaCO3, and TOC values, the following ecostratigraphicassemblages were distinguished.
Organic-walled dinoflagellate cysts and brackish/freshwater taxa. There are 10 genera and 32 species oforganic dinoflagellate cysts. Relative abundances of di-
Text-Figure 5. Relative abundance of major pollen groups based on total sporomorphs (pollen and spores), and AOM % and TOCwt% as percentage of total sedimentary organic matter in NC Core 2. The horizontal lines delimit the boundaries of the main pollen–spore assemblage zones described in the text.

10 PALYNOLOGY, VOLUME 33 — 2009
Depth in cm TaxaBitectatodinium sp. 0 0 0 0 0 0 0 0 0 0 0 0 59 0 0 0
Echinidium sp. 0 0 0 0 0 0 46 0 42 72 28 46 20 68 0 0
Fromea sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 68 48 44
Impagidinium aculeatum 59 0 141 396 0 73 0 39 170 144 28 70 20 34 0 153
Impagidinium paradoxum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 153
Impagidinium patulum 15 15 0 0 0 0 0 0 106 48 0 93 59 237 72 175
Impagidinium sphaericum 37 0 100 99 0 73 0 39 106 168 0 0 39 135 24 87
Impagidinium spp. 7 0 141 545 206 437 0 39 404 216 110 93 176 608 312 240
Total Impagidinium 129 27 382 1041 206 583 0 116 786 575 138 256 293 1014 408 809
Lingulodinium machaerophorum 0 0 100 396 482 437 46 1084 743 575 413 604 117 439 48 547
Nemaotosphaeropsis labyrinthus 0 0 0 0 0 0 0 0 0 0 0 0 59 0 0 0
Operculodinium centrocarpum 0 0 141 248 69 364 367 465 319 144 193 232 293 237 24 87
Operculodinium israelianum 15 0 241 297 0 437 138 581 255 144 110 139 274 372 72 22
Polysphaeridium zoharyi 7 0 40 0 0 0 0 194 85 96 193 465 176 743 192 415
Polysphaeridium spp. 0 0 0 0 0 0 92 77 42 0 83 139 156 608 120 219
Total Polysphaeridium 7 0 40 0 0 0 92 194 127 96 193 604 333 743 312 634
Spiniferites belerius 0 0 20 0 0 0 0 232 212 168 220 279 215 68 48 44
Spiniferites bentorii 0 7 0 0 0 0 0 0 0 0 0 0 39 68 0 153
Spiniferites delicatus 0 0 0 297 206 146 0 232 382 96 55 139 196 169 0 44
Spiniferites elongatus 0 0 0 0 0 0 229 0 149 24 83 256 117 135 72 131
Spiniferites lazus 0 0 0 0 0 0 92 0 170 0 138 163 0 68 72 197
Spiniferites mirabilis 0 0 120 248 0 0 0 232 191 144 193 976 274 642 0 416
Spiniferites ramosus 7 0 362 347 344 437 92 465 934 456 1349 767 587 541 72 131
Spiniferites spp. 7 0 141 149 344 437 46 232 871 528 440 1045 626 946 144 481
Total Spiniferites 15 0 663 1041 895 1020 459 1394 2910 1415 2478 3624 2034 2635 408 1596
Stelladinium sp. 0 0 0 0 0 0 92 0 0 0 0 0 0 0 0 0
Unidentified dinocysts 35 10 261 248 482 510 413 232 467 360 303 139 156 38 48 131
Total dinocysts 191 32 1808 3271 2134 3352 1652 4142 5649 3381 3937 5645 3658 6221 1367 3870
Acritarchs 37 0 201 149 551 146 92 232 340 360 138 0 0 0 120 109
Crustacean eggs 0 0 60 347 895 1822 964 465 276 168 165 70 0 0 24 131
Microforaminiferal Lining 0 0 201 0 0 0 0 0 0 0 0 23 0 0 24 44
Prasinophyte / Tasmanites 74 15 402 248 344 510 872 387 234 360 303 139 98 83 48 109
Total Marine Count 302 47 2672 4014 3923 5830 3579 5226 6499 4268 4543 5877 3756 6304 1583 4263
Total sporomorphs 67 119 5967 15065 18308 24267 14499 7782 3738 1545 993 1786 528 473 168 437
concentrations per gram 369 166 8639 19079 22231 30097 18078 13008 10237 5813 5535 7663 4284 6777 1750 4700
150–160 160–16885–95 100–110 110–120 120–130 130–140 140–15050–55 60–65 70–75 80–85 0–10 15–20 25–30 30–45
Depth in cm TaxaBitectatodinium sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 1.6 0.0 0.0 0.0
Echinidium sp. 0.0 0.0 0.0 0.0 0.0 0.0 2.8 0.0 0.8 2.1 0.7 0.8 0.5 1.1 0.0 0.0
Fromea sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 1.1 3.5 1.1
Impagidinium aculeatum 31.2 0.0 7.8 12.1 0.0 2.2 0.0 0.9 3.0 4.3 0.7 1.2 0.5 0.5 0.0 4.0
Impagidinium paradoxum 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 4.0
Impagidinium patulum 7.8 45.9 0.0 0.0 0.0 0.0 0.0 0.0 1.9 1.4 0.0 1.6 1.6 3.8 5.3 4.5
Impagidinium sphaericum 19.5 0.0 5.6 3.0 0.0 2.2 0.0 0.9 1.9 5.0 0.0 0.0 1.1 2.2 1.8 2.3
Impagidinium spp. 3.9 0.0 7.8 16.7 9.7 13.0 0.0 0.9 7.1 6.4 2.8 1.6 4.8 9.8 22.8 6.2
Total Impagidinium % 67.6 83.5 21.1 31.8 9.7 17.4 0.0 2.8 13.9 17.0 3.5 4.5 8.0 16.3 29.8 20.9
Lingulodinium machaerophorum 0.0 0.0 5.6 12.1 22.6 13.0 2.8 26.2 13.2 17.0 10.5 10.7 3.2 7.1 3.5 14.1
Nemaotosphaeropsis labyrinthus 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 1.6 0.0 0.0 0.0
Operculodinium centrocarpum 0.0 0.0 7.8 7.6 3.2 10.9 22.2 11.2 5.6 4.3 4.9 4.1 8.0 3.8 1.8 2.3
Operculodinium israelianum 7.8 0.0 13.3 9.1 0.0 13.0 8.3 14.0 4.5 4.3 2.8 2.5 7.5 6.0 5.3 0.6
Polysphaeridium zoharyi 3.9 0.0 2.2 0.0 0.0 0.0 0.0 4.7 1.5 2.8 4.9 8.2 4.8 11.9 14.0 10.7
Polysphaeridium spp. 0.0 0.0 0.0 0.0 0.0 0.0 5.6 1.9 0.8 0.0 2.1 2.5 4.3 9.8 8.8 5.6
Total Polysphaeridium % 3.9 0.0 2.2 0.0 0.0 0.0 5.6 4.7 2.3 2.8 4.9 10.7 9.1 11.9 22.8 16.4
Spiniferites belerius 0.0 0.0 1.1 0.0 0.0 0.0 0.0 5.6 3.8 5.0 5.6 4.9 5.9 1.1 3.5 1.1
Spiniferites bentorii 0.0 22.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 1.1 1.1 0.0 4.0
Spiniferites delicatus 0.0 0.0 0.0 9.1 9.7 4.3 0.0 5.6 6.8 2.8 1.4 2.5 5.3 2.7 0.0 1.1
Spiniferites elongatus 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 2.6 0.7 2.1 4.5 3.2 2.2 5.3 3.4
Spiniferites lazus 0.0 0.0 0.0 0.0 0.0 0.0 5.6 0.0 3.0 0.0 3.5 2.9 0.0 1.1 5.3 5.1
Spiniferites mirabilis 0.0 0.0 6.7 7.6 0.0 0.0 0.0 5.6 3.4 4.3 4.9 17.3 7.5 10.3 0.0 10.7
Spiniferites ramosus 3.9 0.0 20.0 10.6 16.1 13.0 5.6 11.2 16.5 13.5 34.3 13.6 16.0 8.7 5.3 3.4
Spiniferites spp. 3.9 0.0 7.8 4.5 16.1 13.0 2.8 5.6 15.4 15.6 11.2 18.5 17.1 15.2 10.5 12.4
Total Spiniferites 7.8 23.0 36.7 31.8 41.9 30.4 27.8 33.6 51.5 41.8 62.9 64.2 55.6 42.4 29.8 41.2
Stelladinium sp. 0.0 0.0 0.0 0.0 0.0 0.0 5.6 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Unidentified dinocysts 18.1 31.2 14.4 7.6 22.6 15.2 25.0 5.6 8.3 10.6 7.7 2.5 4.3 0.6 3.5 3.4
Total dinocysts % 63.1 68.5 67.7 81.5 54.4 57.5 46.2 79.3 86.9 79.2 86.7 96.0 97.4 98.7 86.4 90.8
Acritarchs 12.3 0.0 7.5 3.7 14.0 2.5 2.6 4.4 5.2 8.4 3.0 0.0 0.0 0.0 7.6 2.6
Crustacean eggs 0.0 0.0 2.3 8.6 22.8 31.3 26.9 8.9 4.2 3.9 3.6 1.2 0.0 0.0 1.5 3.1
Microforaminiferal Lining 0.0 0.0 7.5 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 1.5 1.0
Prasinophyte / Tasmanites 24.6 31.5 15.0 6.2 8.8 8.8 24.4 7.4 3.6 8.4 6.7 2.4 2.6 1.3 3.0 2.6
Total Marine Count % 81.8 28.4 30.9 21.0 17.6 19.4 19.8 40.2 63.5 73.4 82.1 76.7 87.7 93.0 90.4 90.7
Total sporomorphs % 18.2 71.6 69.1 79.0 82.4 80.6 80.2 59.8 36.5 26.6 17.9 23.3 12.3 7.0 9.6 9.3
85–95 100–110 110–120120–130 130–140 140–150 150–160 160–168 0–10 15–20 25–30 30–45 50–55 60–65 70–75 80–85
Table 2. The concentrations and relative abundances of marine palynomorphs in NC Core 2. 2A - Concentrations(numbers g-1) of dinoflagellate cysts and other marine or freshwater palynomorphs. 2B - Relative abundances,
with percentages of dinoflagellate cysts based on the total dinoflagellate cyst count; the percentagesof the other marine elements are based on the total marine count.
A
B

S.E.A. Kholeif, and P.J. Mudie: Palynological records of Pleistocene and Holocene, conditions, Nile Cone, Egypt 11
Table 3. The concentrations and relative abundances of sporomorphs in NC Core 2. A) Sporomorph concentrations(number g-1). B) Relative abundances of sporomorphs, based on the total count of pollen, fern, and bryophyte spores.
Depth in cm
Taxa
Artemisia 0 0 201 743 1170 1093 459 271 85 48 21 62 0 0 0 0
Asteraceae 0 0 80 991 1927 1676 688 542 488 145 21 185 59 0 0 0
Casuarina 0 7 0 0 69 0 0 0 0 0 0 0 0 0 0 0
Cheno/Amaranth 0 0 402 1734 3028 5611 4955 1936 637 72 0 103 39 0 0 0
Cyperaceae 0 0 181 892 1652 1603 780 736 446 145 42 123 0 0 0 0
Ephedra 0 0 40 248 206 437 321 77 255 72 84 0 0 0 0 0
Poaceae 0 0 241 793 482 1968 2661 968 319 97 63 267 117 0 0 0
Podocarpus 7 30 3013 3518 3097 2915 918 852 425 314 422 267 137 169 96 131
Pinus/Cedrus 30 37 804 1487 1721 1457 459 310 149 145 148 103 20 169 0 153
Other bisaccates 15 30 402 446 482 146 0 194 64 0 0 0 0 101 0 0
Plantaginaceae 0 0 0 347 138 364 229 77 42 72 0 82 0 0 0 0
Polypodiaceae 0 0 0 198 0 291 138 116 0 0 21 0 0 0 0 0
Porate pollen 0 0 0 347 344 437 275 0 0 0 0 0 0 0 0 0
Riccia 0 0 0 0 0 0 0 39 64 72 42 41 0 0 0 0
Tricolpate pollen 0 0 0 743 895 1530 826 232 149 0 42 82 0 0 0 0
Tricolporate pollen 0 0 221 842 1170 2551 918 542 276 97 42 103 0 0 0 0
Triporate 0 0 60 743 1032 1312 551 194 127 24 42 123 0 0 0 0
Unidentified pollen 0 0 100 496 619 364 138 310 149 97 0 123 98 0 0 66
Unidentified spores 15 15 221 496 275 510 184 232 0 145 0 123 59 0 0 87
Verrucate spores 0 0 0 0 0 0 0 155 64 0 0 0 0 34 72 0
Total sporomorphs 67 119 5967 15065 18308 24267 14499 7782 3738 1545 993 1786 528 473 168 437
Fungi 0 0 201 595 688 2186 2845 1355 743 772 317 267 117 0 0 219
130-140 140 -150 150-160 160-16885-95 10 -110 110-120 120-13050-55 60-65 70-75 80-85 0-10 15 20 25 30 30-45
Depth in cmTaxa
Artemisia 0.0 0.0 3.4 4.9 6.4 4.5 3.2 3.5 2.3 3.1 2.1 3.4 0.0 0.0 0.0 0.0
Asteraceae 0.0 0.0 1.3 6.6 10.5 6.9 4.7 7.0 13.1 9.4 2.1 10.3 11.1 0.0 0.0 0.0
Casuarina 0.0 6.3 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Cheno/Amaranth 0.0 0.0 6.7 11.5 16.5 23.1 34.2 24.9 17.0 4.7 0.0 5.7 7.4 0.0 0.0 0.0
Cyperaceae 0.0 0.0 3.0 5.9 9.0 6.6 5.4 9.5 11.9 9.4 4.3 6.9 0.0 0.0 0.0 0.0
Ephedra 0.0 0.0 0.7 1.6 1.1 1.8 2.2 1.0 6.8 4.7 8.5 0.0 0.0 0.0 0.0 0.0
Poaceae 0.0 0.0 4.0 5.3 2.6 8.1 18.4 12.4 8.5 6.3 6.4 14.9 22.2 0.0 0.0 0.0
Podocarpus 11.1 25.0 50.5 23.4 16.9 12.0 6.3 10.9 11.4 20.3 42.6 14.9 25.9 35.7 57.1 30.0
Pinus/Cedrus 44.4 31.3 13.5 9.9 9.4 6.0 3.2 4.0 4.0 9.4 14.9 5.7 3.7 35.7 0.0 35.0
Other bisaccates 22.2 25.0 6.7 3.0 2.6 0.6 0.0 2.5 1.7 0.0 0.0 0.0 0.0 21.4 0.0 0.0
Plantaginaceae 0.0 0.0 0.0 2.3 0.8 1.5 1.6 1.0 1.1 4.7 0.0 4.6 0.0 0.0 0.0 0.0
Polypodiaceae 0.0 0.0 0.0 1.3 0.0 1.2 0.9 1.5 0.0 0.0 2.1 0.0 0.0 0.0 0.0 0.0
Porate pollen 0.0 0.0 0.0 2.3 1.9 1.8 1.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Riccia 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.5 1.7 4.7 4.3 2.3 0.0 0.0 0.0 0.0
Tricolpate pollen 0.0 0.0 0.0 4.9 4.9 6.3 5.7 3.0 4.0 0.0 4.3 4.6 0.0 0.0 0.0 0.0
Tricolporate pollen 0.0 0.0 3.7 5.6 6.4 10.5 6.3 7.0 7.4 6.3 4.3 5.7 0.0 0.0 0.0 0.0
Triporate 0.0 0.0 1.0 4.9 5.6 5.4 3.8 2.5 3.4 1.6 4.3 6.9 0.0 0.0 0.0 0.0
Unidentified pollen 0.0 0.0 1.7 3.3 3.4 1.5 0.9 4.0 4.0 6.3 0.0 6.9 18.5 0.0 0.0 15.0
Unidentified spores 22.2 12.5 3.7 3.3 1.5 2.1 1.3 3.0 0.0 9.4 0.0 6.9 11.1 0.0 0.0 20.0
Verrucate spores 0.0 0.0 0.0 0.0 0.0 0.0 0.0 2.0 1.7 0.0 0.0 0.0 0.0 7.1 42.9 0.0
Total sporomorphs 18.2 71.6 69.1 79.0 82.4 80.6 80.2 59.8 36.5 26.6 17.9 23.3 12.3 7.0 9.6 9.3
130–140 140–150 150–160 160–16885–95 100–110 110–120 120–1300–10 15–20 25–30 30–45 50–55 60–65 70–75 80–85
noflagellate cyst species were calculated based on the totaldinoflagellate cyst count (Table 3), although the totalmarine count includes acritarchs, dinoflagellate cysts,microforaminiferal linings, and prasinophytes (Table 3).Four groups of aquatic palynomorphs were recognized, asfollows:
Group 1. These species have highest relative abundancesin temperate to tropical oceanic areas with low nutrientconcentration in the surface water. They are Impagidiniumaculeatum (Plate 2, fig. 11), Impagidinium paradoxum(Plate 2, fig. 17), Impagidinium patulum (Plate 2, fig. 13),
Impagidinium sphaericum, Impagidinium strialatum (Plate2, figs. 9, 10), and Impagidinium cf. variaseptum. Thesespecies are also distinguished by their high resistance todecay in oxidized sediments (Zonneveld et al., 2001).
Group 2. These species have high relative abundanceseither in upwelling areas associated with tropical mon-soons (Zonneveld et al., 1997), or in conditions of lowphosphate (Marret and Zonneveld, 2003). They are thegonyaulacoid Polysphaeridium zoharyi (Plate 2, fig. 6),and presumably other species of Polysphaeridium, and theprotoperidinioids Echinidinium sp. and Protoperidinium
A
B

12 PALYNOLOGY, VOLUME 33 — 2009
stellatum (see Zonneveld et al., 1997). Polysphaeridiumzoharyi is however a neritic taxon, while the others aremore typical of deep water.
Group 3. These species have relatively broad ecologicalranges within temperate to tropical regions. They includeLingulodinium machaerophorum (Plate 2, figs. 14, 18, 19),Operculodinium centrocarpum (Plate 2, fig. 20),Operculodinium israelianum (Plate 2, fig. 5), Spiniferitesbelerius (Spiniferites membranaceus s.l. of Marret andZonneveld, 2003), Spiniferites bentorii (Plate 2, fig. 3),Spiniferites hyperacanthus (Plate 1, fig. 1), Spiniferitesmirabilis (Plate 2, fig.7), and Spiniferites ramosus (Plate 2,fig. 12). Spiniferites delicatus (Plate 2, fig. 8), and Spiniferiteslazus (Plate 2, fig. 2) are also associated with this group,both of which tend to be coastal species most commonlyfound at salinities above 30 psu.
Group 4. These palynomorphs are brackish and freshwa-ter taxa that are transported into the marine water column,and include reticulate spores of Spirogyra (Plate 1, fig. 14)and psilate zygospores, prasinophytes, and microforam-iniferal linings.
Pollen and spores. The relative abundances of pollenand spores are based on total pollen, and fern/bryophytespores. This total excludes fungal and other unknownspores, mostly Glomus-type (Plate 1, fig. 11) and unknownpsilate taxa, possibly including Azolla nilotica. Two cat-egories of sporomorph assemblages were recognized.
Group 1: Semidesert (Steppe) Pollen. These assem-blages comprise predominantly herbaceous or shrub pollentaxa that were transported by either wind or Nile Riverwater. They are associated with Nile Delta growth andregressive sea level conditions (Kholeif, 2004). Taxa in-clude Poaceae, Cyperaceae (Plate 1, fig. 2), Ephedra (Plate1, fig. 8), Chenopodiaceae and Asteraceae pollen, and
spores of Polypodiaceae (Plate 1, fig. 12) and Riccia (Plate1, fig. 7). Poaceae and Cyperaceae pollen in particular areconsidered major components of Nile input at times of lowsea level because of their abundance in Late Holoceneeastern Nile Delta marshes (Leroy, 1992) and their localdominance in swamps upstream (Ritchie, 1987). Pollen ofPoaceae and spores of the liverwort Riccia, which is onlyfertile when growing on wet soil, have been termed niloticindicators (Cheddadi and Rossignol-Strick, 1995; Kholeif,2004).
Group 2: Monsoon Pollen. These assemblages prima-rily comprise bisaccate pollen, represented mainly by thetropical highland tree Podocarpus (Plate 1, fig. 5), thesouthern afromontanous pollen type of Leroy (1992), andto some extent by Cedrus and Pinus. The former are alsocommon in the Atlas Mountains of northwest Africa andcould be transported by westerly winds. Within Egypt,however, softwood conifers are relatively rare, and theirpollen is sparse in Holocene lake sediments, for example amaximum of 3.5% in the Fayoum Depression (Kholeif,2007). Currently, Podocarpus is restricted to the southernhighlands, mainly the montane mist forests of Ethiopiabetween 1,100 and 1,800 m (Chedaddi and Rossignol-Strick, 1995). Other common European bisaccate treepollen, for example Abies and Picea, were not found. Inprevious studies of the Nile Delta (Saad and Sami, 1967; ElBeialy et al., 2001), it was concluded that most bisaccatepollen are part of the southern forest pollen input. Further-more, that they are allochthonous elements derived fromtrees in central Africa, Ethiopia, and south Sudan, trans-ported northwards by both Nile River water and by mon-soon winds. Usually, bisaccate pollen is not considered animportant paleoclimatic marker because of its widespreaddistribution by wind, but it can be useful if the source areaof the parent plants is geographically restricted (Hughesand Moody-Stuart, 1967). By contrast, Cheddadi andRossignol-Strick (1995) collected Cedrus and Pinus pollen
1 Asteraceae Liguliflorae pollen, #5/1, R35/3.2 Cyperaceae pollen, #7/1, F17/3.3, 4 Chenopodiaceae/Amaranthaceae pollen, #6/2, U34/3.5 Podocarpus pollen, #5/1, N40/3.6 Pediastrum simplex, #9/1, Q36/1.7 Riccia spore, #7/2, R18/4.8 Ephedra fragilis-type pollen, #7/2, P32.
PLATE 1
Transmitted light photomicrographs of selected pollen, spore, and algal taxa, and amorphous organic matter (AOM) from NC Core 2. Thescale bar represents 10 µm. The microscope slides are curated at the National Institute of Oceanography and Fisheries (NIOF), Alexandria,Egypt. The alphanumerics are sample/slide numbers and England Finder coordinates respectively.
9 Microforaminiferal lining, #3/1, C44/1.10 Poaceae pollen, #4/1, C28/3.11 Glomus-type fungal spore, #4/2, L46.12 Polypodiaceae fern spore, #6/1, V33/4.13 Amorphous organic matter (AOM), #5/1, M19/2.14 Reticulate Spirogyra (fresh water alga) zygospore, #7/
1, G33/4.

S.E.A. Kholeif, and P.J. Mudie: Palynological records of Pleistocene and Holocene, conditions, Nile Cone, Egypt 13

14 PALYNOLOGY, VOLUME 33 — 2009
in traps placed on ships in the Levantine Basin, and theyconsidered these to be part of circum-Mediterranean plantinput. These authors reported that only Podocarpus is areliable tropical pollen indicator, and together with pterido-phyte spores, indicated downstream transport of Nile waterduring intervals of heavy monsoon rains. It is also notablethat Podocarpus pollen is rare in the Aegean Sea (Aksu etal., 1995), and that the bisaccates Abies and Picea areabsent from the Nile Cone cores.
The time-depth variations in relative abundances ofselected indicator pollen and spores, dinoflagellate cystspecies, AOM, CaCO3, and TOC are illustrated in Text-Figures 5 and 6. Three main ecostratigraphic units arerecognized, as summarized below.
Units I and II. Lowermost calcareous and pre-sapro-pel layers, 168–85 cm (14,620–9,500 yr BP). In this LatePleistocene–Holocene interval, dinoflagellate cysts havetheir maximum relative abundance in NC Core 2 (Table3). Species of Spiniferites dominate (around 30–64% oftotal dinoflagellate cysts), representing dinoflagellate cystgroup 3. The total Spiniferites species increase graduallyupwards, reaching a peak (64.2 % total dinoflagellatecysts) immediately below the top of Unit I (120–130 cm).The Spiniferites species include Spiniferites ramosus (16–34% of total dinoflagellate cysts), Spiniferiteshyperacanthus (up to 10%), together with Spiniferitesbelerius (up to 6%), and Spiniferites delicatus, Spiniferites
elongatus (Plate 2, fig. 4), and Spiniferites lazus (up to5%). Small amounts of Spiniferites rubinus (Plate 2, figs.15, 16) are also present. The next most abundant speciesin Group 3 is the temperate–tropical euryhaline speciesLingulodinium machaerophorum (maximum 14%).Operculodinium israelianum and Operculodiniumcentrocarpum are present (up to 8%). Bitectatodinium sp.and Nematosphaeropsis cf. labyrinthus are rare (1.6%,only in Unit I).
Dinoflagellate cyst group 1 is also important at the baseof Unit 1, before 13,160 yr BP, where total Impagidiniumspp. represent 29.8%. This Impagidinium peak is fol-lowed by an upwards decrease to a minimum of 4.5% at120 cm, followed by a slight increase (Text-Figure 6). TheImpagidinium species are mainly Impagidiniumaculeatum, Impagidinium patulum, and Impagidiniumsphaericum, but also include Impagidinium paradoxumand rare Impagidinium strialatum, as well as otherImpagidinium species. Other common dinoflagellate cystsin Unit I are Polysphaeridium zoharyi and otherPolysphaeridium species (Dinoflagellate cyst group 2),with a peak of 22.8% before 13,160 yr BP, and a notabledecrease in Unit II.
Pollen and spore concentrations in the NC Core 2 haverelatively low values (168–1,786 grains g-1) and minimumrelative abundances in the early postglacial Unit I (Text-Figure 5), where they comprise 7% of the total palynomor-phs. However they increase after 13,000 yr BP, before the
PLATE 2
Transmitted light photomicrographs of selected dinoflagellate cysts from NC Core 2. The scale bar represents 10 µm. The microscope slidesare curated at the National Institute of Oceanography and Fisheries (NIOF), Alexandria, Egypt. The alphanumerics are sample/slidenumbers and England Finder coordinates respectively.
1 Spiniferites hyperacanthus, #15/1, R35, lateral view ofarcheopyle.
2 Spiniferites lazus, #16/1, S15/2, mid focus.3 Spiniferites bentorii, #13/2, J38/3, high focus.4 Spiniferites elongatus, #15/1, S21, high focus on
archeopyle.5 Operculodinium israelianum, #11/1, D60/3, high focus
on spines.6 Polysphaeridium zoharyi, #14/2, Q28/2, mid focus,
showing some bifurcate processes.7 Spiniferites mirabilis, #12/1, M29/3, mid focus.8 Spiniferites delicatus, #4/1, K37, showing petalloid
process terminations.9, 10 Impagidinium strialatum, #4/1, K37, 9 - high focus,
dorsal surface; 10 - mid focus.11 Impagidinium aculeatum, #15/1, 38/2, dorsal view.12 Spiniferites ramosus, #1/2, T28/4.
13 Impagidinium patulum, #16/2, D60/3, ventro-lateralview.
14, 18, Lingulodinium machaerophorum, 14, 18 - long spined19 morphotypes. 14 - #10/2, L28, cyst with compound
archeopyle; 18 - #9/1, Q36/1, cyst with single archeopyleparaplate. 19 - short spined morphotype with largearcheopyle, #12/1, R18.
15 Spiniferites cf. rubinus, #13/1, M35/1, with granulatebody and parasutural septa.
16 Spiniferites sp., #13/1, L22, ventral view of weaklydefined paratabulation, thin bi- or trifurcate spines andflange linking parasulcal processes.
17 Impagidinium paradoxum, #16/1, I22/3, ventral sur-face.
20 Operculodinium centrocarpum, short spinedmorphotype, #2/1, T11/4, high focus, dorsal view.

S.E.A. Kholeif, and P.J. Mudie: Palynological records of Pleistocene and Holocene, conditions, Nile Cone, Egypt 15

16 PALYNOLOGY, VOLUME 33 — 2009
start of pre-sapropelic Unit II. The monsoon pollen indica-tor Podocarpus is dominant (20–57%) in most of Units Iand II, while total bisaccate pollen decreases from a maxi-mum of 92.9% in Unit I to 17% in Unit II. Before about13,000 yr BP in Unit 1, the samples contain only Pinus andPodocarpus pollen, unidentified small trilete spores (pos-sibly including Azolla cf. nilotica microspores), and fernspores. After 13,000 yr, the sporomorph assemblages aredominated by the semi-desert pollen group, with Asteraceae,Chenopodiaceae, and Poaceae pollen, fern and Riccia sporesappearing between 140 and 120 cm. These are followed bythe Saharo–Mediterranean pollen indicators Artemisia (aspart of the Asteraceae) and Ephedra immediately belowUnit II (Ritchie, 1987). Cyperaceae pollen have maximumrelative abundances (9–12%) and maximum concentra-tions in Unit II.
Unit III. Sapropel layer, 85–30 cm (approximately9,500–6,000 yr BP). The S1 sapropel is distinguished bya high organic carbon content of 2.1–3.4% dry weight andthe AOM percentage ranges between 58 to 72% (Table 1).The concentrations of almost all palynomorphs are higherin the sapropel than the underlying or overlying organic-rich mud units (Table 3). The major change in dinoflagel-late cyst assemblages is the disappearance of some spe-
cies of Group 1 (Impagidinium patulum and Impagidiniumparadoxum) and of Group 3 (Spiniferites mirabilis andSpiniferites bentorii). The upwelling cyst Group 2 is seenonly in the lowermost sapropel layer (80–85 cm), whereit is sparse (4.7% of the total dinoflagellate cysts).Spiniferites belerius, Spiniferites bentorii, and Spiniferiteslazus also appear only at the base of the sapropel.Spiniferites elongatus (14%) occurs from 75 to 70 cm; thisspecies is most common in temperate to cold waters(Marret and Zonneveld, 2003) and is virtually absent inmodern sediments in the Levantine Sea (Marret et al.,2008). The group 3 species Operculodinium centrocarpum(up to 22%), Operculodinium israelianum (14%), andLingulodinium machaerophorum (28%) have their maxi-mum percentages in the sapropel. Group 4 elements,dominated by prasinophytes, have maximum concentra-tions (2,615 cysts g-1) and relative abundances (24% oftotal marine count) in the sapropel. Pediastrum (Plate 1,fig. 6) occurs only in this unit.
The sapropel is also distinguished by the maximumconcentration and relative abundance of Chenopodiaceae/Amaranthaceae pollen and the first consistent occurrencesof Polypodiaceae spores (Table 2). However, Podocarpuspollen concentration decreases in concentration to 425grains g-1 and in percentage of total palynomorphs (10.4%)
Text-Figure 6. Relative abundances of dinoflagellate cysts in NC Core 2. The dinoflagellate cyst percentages are based on the totaldinoflagellate cyst count. The horizontal lines delimit boundaries of the main pollen–spore assemblage zones described in the text.

S.E.A. Kholeif, and P.J. Mudie: Palynological records of Pleistocene and Holocene, conditions, Nile Cone, Egypt 17
except at the top, where there is a second increase to 3,518grains g-1 (23.4%).
Unit IV. Post-sapropel organic mud layer, 30–25 cm(approximately 6,000–3,000 yr BP). The post–sapropelicprotosapropel is marked by lower TOC percentages (1.8 wt%) and AOM percentages (32%), and by a slight increasein biogenic sand (3%). There is a decrease in percentages ofLingulodinium machaerophorum and Operculodiniumcentrocarpum, and an increase in Operculodiniumisraelianum, Polysphaeridium zoharyi, and Spiniferitesramosus in addition to the reappearance of Spiniferitesmirabilis. The pollen and spore concentrations and thepercentages of most species is lower than in the underlyingsediments (62% of the total palynomorphs), but bisaccatepollen increases to 70% of total pollen and spores, mostlyas Podocarpus.
Units V and VI. Oxidized layer (25–15 cm) andcalcareous ooze (155–0 cm), approximately 3,000 to <150 yr BP. The uppermost units are characterized byminimum organic carbon content (0.1–0.5%) and AOM(10–15%). Within this interval, most dinoflagellate cystgenera found in the lower sediments disappear, except forthe Group 1 species including Impagidinium aculeatum(31%), Impagidinium patulum (46%), Impagidiniumsphaericum (20%), and Impagidinium spp. (4%). Theoxidation-resistant species Impagidinium aculeatum,Impagidinium patulum, and Impagidinium sphaericumhave their highest relative abundance in this unit, but theiraverage concentration remains the same. In contrast,Operculodinium israelianum (8%) and Polysphaeridiumzoharyi (4%) have their lowest concentrations in thisinterval (Table 2), although they are resistant to aerobicdegradation (Zonneveld et al., 2001). Spiniferites species,which are common to abundant in all the lower units, aremostly absent in the surface carbonate ooze except forSpiniferites ramosus (20%) and Spiniferites bentorii (23%)that are moderately sensitive to oxidation. The terrestrialpalynomorphs have their lowest concentrations and rela-tive abundances (3.9% of total palynomorphs) in Units Vand VI. Low concentrations of the bisaccate pollen aretypical. The only other pollen is from the introduced,drought-resistant tree Casuarina that is now common in theNile Delta (Ayyad et al., 1992). Part of this low diversityprobably reflects reduced carbon preservation, but selec-tive transport and distance offshore is also involved. Forexample, air samplers on ships in the Levantine Basin(Cheddadi and Rossignol, 1995) recorded 15 wind-bornepollen taxa compared to the Nile Delta pollen rain with 71taxa (Ayyad et al., 1992). Unpublished data indicate thatthe low TOC sediments in NC Core 1 from the inner
Egyptian Shelf, younger than 2,250 yr BP, contain 12pollen–spore taxa, including common Podocarpus andother tropical tree pollen (Meliaceae, Palmaceae), herbpollen, and fern spores. Pollen deposition in Late Holocenesediments of the region thus decreases strongly with dis-tance offshore, as recorded in many other marine environ-ments (Mudie and McCarthy, 2006).
DISCUSSION
The records of organic-walled dinoflagellate cysts, pol-len, and spores, and AOM variations in the NC Core 2 canbe explained by taphonomic conditions and three mainpaleoenvironmental factors. These are climate fluctuation,the African monsoon index, and the volume of water andfine-grained sediment discharged by the Nile River. Thetaphonomic factors need to be considered first. This isbecause some thin-walled pollen (Havinga, 1964), andmost protoperidinioids and some gonyaulacoid dinoflagel-late cysts are sensitive to oxidation. Fortunately, the indexof dinoflagellate cyst oxidation sensitivity for the easternMediterranean (Zonneveld et al., 2001) and other qualita-tive data for the Aegean Sea (Aksu et al., 1999), provide abasis for evaluating the importance of changes in organiccarbon preservation in NC Core 2.
In the NC Core 2, the sedimentation rates of Units I toIII are virtually identical (Text-Figure 4), but the highestconcentrations of thin-walled Cyperaceae pollen and totaldinoflagellate cysts occur in the relative low TOC contentUnits I and II, not in the organic-rich Unit III sapropel.This result was not expected if the changes in abundanceonly reflected better preservation of organic matter, assuggested for dinoflagellate cysts in the deep LevantineBasin (Zonneveld et al., 2001), and for pollen in coresfrom water depths of 740–1,260 m on the Nile Cone(Cheddadi and Rossignol-Strick, 1995). Comparison ofthe upper Nile Cone S1 dinoflagellate cyst assemblageswith those in the S1 sapropel of the deep (2,150 m)western Levantine Basin reveals some similarities. Theseare the abundances of the oxidation-resistant gonyaulacoidgenus Impagidinium and the moderately-resistantLingulodinium machaerophorum. However, the deep basinsapropel assemblages show several important differencesfrom the Nile Cone assemblages in the abundance ofmoderately-resistant Spiniferites spp., and commonprotoperidinioids, including thin-walled forms such asBrigantedinium. Similar features are seen in the shal-lower (630 m) Central Aegean Basin, although here thereis a major shift from high Brigantedinium influxes at thebase to increased gonyaulacoid influx at the top, despitethe consistently high percentages of TOC (>2–6%) in thesapropel (Aksu et al., 1999).

18 PALYNOLOGY, VOLUME 33 — 2009
The near absence of protoperidinioid species in the NileCone core, the total absence of Protoperidinium in the S1sapropel, and the organic-rich pre- and post-sapropelicmuds of NC Core 2 all point to a change in cyst produc-tivity during the time of sapropel deposition. This is notmerely a change in preservation as a result of bottomwater stagnation. The lack of the oxidation-sensitiveprotoperidinioids in the sapropel is contrary to what isexpected from the simple model of a switch to suboxic–anoxic bottom water conditions (for example the stagna-tion hypothesis of Calvert and Fontugne, 2001), andsuggests that the surface water environmental conditionswere more important than the taphonomic history indetermining the S1 cyst assemblages. Similar results werereported by Aksu et al. (1995; 1999) for the Aegean SeaS1 sapropel, and by models predicting a combination ofenhanced surface production, increased organic matterexport, and stagnant bottom water (Murat and Got, 2000;Bianchi et al., 2006).
If it is assumed that taphonomic alteration of organic-walled dinoflagellate cysts, pollen, and spores cannot ex-plain the assemblage changes that characterize the sapropellayer S1 and the other moderately organic-rich sedimentunits I–IV (TOC >0.8%) in the Nile Cone core, possiblepaleoclimatic/paleoceanographic forcing factors may beimplicated.
Sea surface Conditions, Nile Discharge Fluctuationsand African Monsoon Intensity
Despite the coarse sampling density of the NC Core 2,where each sample represents deposition over core depthsof 5–15 cm, and time intervals of around 500 to 1,500 years,there are several notable variations in the palynomorphrecords for NC Core 2. These suggest important changes inthe climate during the time of deposition of Units I to IV.Three major changes in environmental conditions are indi-cated:
1. The Unit I and II silty clay and pre-sapropelic mud units(165–85 cm) were deposited between around 14,620and 9,500 yr BP, during the transition from postglacialto interglacial conditions. The average sedimentationrate during this time is about 13.7 cm per millennium.High concentrations and relative abundances ofImpagidinium spp., Polysphaeridium zoharyi, andPolysphaeridium spp. in Unit I suggest an initial tem-perature rise from the base to around 130 or 120 cm(Text-Figure 6), immediately before the pre-sapropelicUnit II. From studies in the Gulf of Gabes off Tunisia,Morzadec-Kerfourn (1983) concluded that the occur-rence of Polysphaeridium zoharyi in the highly saline
neritic Gulf waters mainly reflects arid environmentalconditions. However, more recent wide-scale studies(Marret and Zonneveld, 2003; Marret et al., 2007)indicate that Polysphaeridium zoharyi is a warm waterspecies that is also tolerant of low surface salinities. Theoccurrence of Spiniferites elongatus, however, sug-gests that temperate to cold surface water conditionsprevailed for at least part of the year (Zonneveld et al.,1996; Rochon et al., 1999).Low pollen production and reduced Nile discharge isindicated for the Late Pleistocene interval from around14,620 to 13,160 yr BP (165–140 cm) because of thelow total sporomorph concentrations, and the nearabsence of species other than bisaccate pollen and fernspores. Relatively low AOM values and TOC percent-ages indicate more oxidizing conditions for Units I andII, so reduced pollen preservation might be involved(Tyson, 1995). However, the presence of oxidation-sensitive dinoflagellate cysts, and the upward increasein the pollen and spore diversity of Unit I, particularlyabove 120 cm (around 12,000 yr BP), point to apaleoclimatic/hydrological change. It is therefore likelythat the sparse sporomorph assemblages at the base ofNC Core 2 correspond to the interval of lower NileRiver discharge prior to around 13,000 yr BP (i.e.14,500 calender years BP) documented for Ethiopianand Kenyan lakes (Williams et al., 2006; Lamb et al.,2007).The first appearances of Artemisia and Ephedra (part ofthe Asteraceae, see Table 3) at around 120 cm, mark thestart of a period of cold, dry conditions, as recorded inmost of the Mediterranean region (Mudie et al., 2002b).However, Cyperaceae and Poaceae pollen indicate thelocal presence of extensive wetlands either along theEgyptian coast, or in the Nile Valley, possibly similar tothose of the modern Nile Delta (Ayyad et al., 1992;Leroy, 1992). The occurrence of Riccia and freshwateralgal spores also point towards increased discharge ofthe Nile River after approximately 12,000 years (120cm). Increased input of the tropical Podocarpus pollenappears to mark the strengthening of the African Mon-soon Index and the northward movement of the ITCZ,as predicted by the orbital insolation-driven climatemodel of Rossignol-Strick (1983; 1985), although thetime notably precedes that of S1 sapropel deposition bythousands of years.
2. The Unit II (120–85 m) pre-sapropel organic mud(protosapropel) deposited around 12,000–9,500 yr BPis marked by increased percentages of the euryhalinespecies Lingulodinium machaerophorum, suggestingwarm, stratified surface water and a rise in nutrients. In

S.E.A. Kholeif, and P.J. Mudie: Palynological records of Pleistocene and Holocene, conditions, Nile Cone, Egypt 19
modern sediments, this species ranges from cool tem-perate to tropical (Bradford and Wall, 1984; Edwardsand Andrle, 1992) but it is primarily noted as respond-ing to high nutrient input and stratified warm water(Marret and Zonneveld, 2003; Mertens et al., 2008).Spiniferites mirabilis s.l. of Marret and Zonneveld(2003), which includes Spiniferites hyperacanthus, hasa restricted distribution in the pre-sapropel sediment.This taxon is a temperate to tropical species, associatedwith a broad range of trophic conditions. Increases inthe warm water taxa Impagidinium spp. andOperculodinium israelianum may indicate higher sur-face water temperature in the Early Holocene, follow-ing the cool Younger Dryas interval (11,000–10,500 yrBP) that is marked in NC Core 2 by a peak in Spiniferiteselongatus at 130–120 cm.
3. In Unit III (85–30 cm), a period of slightly highersedimentation (around 14 cm kyr-1) and organic burialtook place during the Holocene, from approximately9,500–6,000 yr BP, with deposition of the S1 sapropel.The absence of turbidites in S1 of the NC Core 2 issuggested by the grain size homogeneity (Table 1),although the sample interval is too coarse to revealfining-upwards of silty muds. However, other evi-dence is the absence of nearshore species such asPolysphaeridium zoharyi and protoperidinioids asso-ciated with coastal waters. The sapropel depositioncoincides with the time of the humid climatic trendthat affected the Mediterranean and northwest Sudanduring the Early to Mid Holocene (Ritchie, 1987;Béthoux, 1993). High AOM values (>60%) indicatethat the visible S1 unit was deposited under anoxic–dysoxic bottom water conditions (Caratini et al., 1981;Tissot and Pelet, 1981). High AOM percentages rela-tive to the total organic debris also indicates goodpreservation of autochtonous planktonic SOM (Tyson,1995). A tropical to temperate-subtropical climate andhigh surface salinity is indicated by higher percentagesof Operculodinium israelianum. It is also notable that,unlike the Levantine and Aegean Basin S1 sapropels,the Nile Cone sapropel shows a reduction in relativeabundance of eurythermal Spiniferites species. Further-more, it contains almost no Spiniferites elongatus,which is an indicator of temperate to cold waters.Increased moisture and improved climate conditions inthe Saharan region during this period are suggested bythe higher concentrations of pollen and spores. The Nileand Sudanian swamp indicators Cyperaceae and Poaceaeremain high, at least in the lower sapropel. However, sealevel rise would have progressively decreased the areaof the marsh-covered postglacial Nile delta. There is a
conspicuous occurrence of Polypodium fern spores inthe sapropel. Ferns do not grow in the Nile Delta (Leroy,1992), and the spores are considered good markers ofNile discharge. There is a reduction of the tropicalhighland forest indicator Podocarpus, but it remainsconsistently more than two times higher than Cedrusand Pinus pollen. A short time of cooler, drier climatemay be indicated at the base (85 cm) and middle (75–70cm) of the sapropel. This is due to the reappearance ofthe cool-temperate species Spiniferites elongatus, andincreased concentrations of Ephedra pollen (Table 3)that is a good marker of arid Saharan conditions(Cheddadi and Rossignol-Strick, 1995).It is more difficult to interpret the assemblages in UnitsV and VI because only two samples were studied, andlow pollen concentration and diversity may reflect bothgreater oxidation and distance offshore from the Nilewater source. An increasingly warm climate with de-creasing moisture is suggested by the reappearance ofoxidation-sensitive Polysphaeridium, and by high con-centrations of Impagidinium (especially in the top 10cm), and by high CaCO3. There is a decline in the niloticindicators and total pollen and spores (17% total terres-trial palynomorphs). Only bisaccate pollen, Poaceae,and rare Casuarina grains are present. The absence/reduction of oxidation-resistant Artemisia and cheno-pod pollen at both slope (NC Core 2) and inner shelfsites (NC core 1; Marret et al., 2008) is remarkable andsuggests a drying up of coastal saltmarshes (Ayyad etal., 1992) and less winter rain (Mudie et al., 2002b).
The effect of Nile discharge on S1 sapropel deposition
The NC Core 2 provides the first relatively high resolu-tion palynological record for sapropels of the upper NileCone. The most detailed previous study is Cheddadi andRossignol-Strick (1995); this had only one or two samplesfor S1, and only reported total pollen concentrations. De-spite the lack of fine-scale sampling in NC Core 2, it is nowclear that the high monsoon influence coincides with theLate Pleistocene deposits in the Levantine Basin, terminat-ing after the cooling interval around 12,000 yr BP, andpreceding the time of sapropel formation by several thou-sand years. Based on the stratigraphic distribution andconcentrations of nilotic markers like Poaceae, Polypodium,and Riccia in NC Core 2, the Nile River discharge appar-ently increases after 13,160 yr BP, reaching its maximumvalue at the time of sapropel layer formation. The relativelylarge contribution of Nile water during sapropel formationfrom around 9,500 to 6,270 yr BP is evident as an increaseof Poaceae during the cooler interval at the base, and by thestrong presence of Cyperus-type pollen and Polypodiaceae

20 PALYNOLOGY, VOLUME 33 — 2009
fern spores. These are considered to be the major compo-nents of Nile water input. The peak of total sporomorphscoinciding with S1 may partly reflect the low bottom wateroxygen, but the persistence of high sporomorph concentra-tions in the post sapropelic layer indicates an increase ofterrigenous organic matter, possibly aided by the highermarine production and more rapid deposition of pollen asfecal pellets (Mudie and McCarthy, 2006).
The Nile discharge appears to reach its minimum dis-charge during the deposition of the uppermost sediments(30–0 cm), as suggested by low values of AOM, pollen,and TOC. However, because the uppermost sediments arestrongly oxidized, and the site is at the distal end of theNile river plume, the pollen signal is weak and does notallow clear paleoenvironmental interpretation. The lowvalues of Pinus and Poaceae pollen essentially provide“background noise” against which to measure the moremeaningful Late Pleistocene and Early–Mid Holocenepaleoclimatic and paleohydrological signals. It is remark-able, however, that the pollen assemblages in Core 22from the more northerly Cretan Basin of the Aegean Sea(Aksu et al., 1995a) contained a far more diverse pollenassemblage, including Abies, Picea, Pistacia, Quercus,Rhamnus, and Tilia, all of which are absent throughoutNC Core 2, regardless of the oxidation state as measuredby the percentage of TOC.
CONCLUSIONS
Millennial-scale changes in palynomorph assemblagesand AOM are evident in this expanded series of Cyclothem1 sediments from the upper Nile cone. The NC Core 2records several major changes in paleoclimate, monsoonintensity, and Nile River discharge. This is despite thesparse sample coverage available and some taphonomiclimitations related to distance offshore and oxidation of theuppermost sediment. The paleoclimatic history is summa-rized as follows:
1. The dinoflagellate cyst assemblages in all the units inthe NC Core 2 from the upper slope of the Eastern NileCone are dominated by the gonyaulacoid taxaImpagidinium and Spiniferites. This is regardless of thepercentage of TOC, including the S1 sapropel layerwith 2–3% TOC. Spiniferites spp, with the cool waterindicator Spiniferites elongatus, dominate the earlypostglacial interval, together with other neritic, warmwater indicators such as Polysphaeridium zoharyi. Thepresence of Impagidinium is positively correlated withCaCO3 content, and indicates relatively warm surfacewater in the southeastern Mediterranean throughout thepast 14,600 years.
2. The basal and pre-sapropelic interval (165–85 cm) inNC core 2 represents the transition period from glacialto interglacial conditions at approximately 14,600–9,500 yr BP. The interval is generally marked by thestrong presence of the monsoon pollen indicatorPodocarpus, and variable Nile discharge as shown bypeaks in Poaceae pollen and Riccia spores. Both di-noflagellate cyst and sporomorph assemblages indicatea gradual increase in temperature, but the presence ofArtemisia and Ephedra, and a peak in Spiniferiteselongatus around 12,000 yr BP may mark the YoungerDryas cold event, with cooler, drier conditions. Thehigh African monsoon index from about 13,000–10,000yr BP generally coincides with the occurrence of south-ern tree and Sudanian swamp pollen, as expected fromincreased Nile discharge and northward movement ofthe ITCZ.
3. The visible sapropel S1 in the upper NC core 2 isdistinguished by an absence of turbidites, and it differsfrom deep water Levantine Basin cores in the nearabsence of protoperidinioids. However, it was depos-ited in highly stratified and oxygen-deficient bottomwater. Sporomorphs and dinoflagellate cysts indicate atropical to subtropical climate, and possibly increasedNile flooding as indicated by occurrence ofPolypodiaceae fern spores, persistent presence ofCyperaceae pollen, and a peak in Poaceae near the base.Temperature and humidity may have been reducedduring a short interval of dry–cool climate near the base(75–70 cm), perhaps corresponding to the 8,200 yr BPcooling event.
4. The post-sapropelic oxidized layer and carbonate oozemay have been deposited under warm, dry climaticconditions during the past approximately 3,000 years,with relatively low Nile input. However, greater oxida-tion at the top of the core limits paleoenvironmentalinterpretations.
ACKNOWLEDGMENTS
The authors gratefully acknowledge Dr. Mona Khalil(National Institute of Oceanography and Fisheries (NIOF),Alexandria, Egypt), for providing the samples andgeochemical data. Sincere thanks go to Professor KarinZonneveld (Bremen University, Germany) and to Dr.Fabienne Marret (University of Liverpool, U.K.), for theirhelp in identification, providing references, and sugges-tions during the preparation of the manuscript. We alsothank four reviewers, particularly Drs. Rex Harland andSuzanne Leroy, for their helpful comments.

S.E.A. Kholeif, and P.J. Mudie: Palynological records of Pleistocene and Holocene, conditions, Nile Cone, Egypt 21
References Cited
AKSU, A.E., YASAR, D., MUDIE, P.J., and GILLESPIE, H.1995 Paleoclimatic and paleoceanographic circumstances
leading to the development of sapropel layer S1 inthe Aegean Sea basins. PalaeoceanographyPalaeoclimatology Palaeoecology, 116: 71–101.
AKSU, A.E., ABRAJANO, J., MUDIE, P.J., and YASAR, D.1999 Organic geochemical and palynological evidence
for terrigenous origin of the organic matter in Aegeansapropel S1. Marine Geology, 153: 303–318.
ALPERT, P., BALDI, M., ILANI, R., KRICHAK, S., PRICE, C.,RODO, R., SAARONI, H., ZIV, B., KISCHA, P., BARKAN, J.,MARIOTTI, A., and XOPLAKI, E.
2006 Relations between climate variability in the Mediter-ranean region and the Tropics: ENSO, South Asianand African Monsoons, Hurricanes and Saharan dust.In: Lionello, P., Malanotte-Rizzoli, P., and Boscolo,R, (eds.), Mediterranean Climate Variability. Devel-opments in Earth and Environmental Sciences, 4:149–177.
AYYAD, S.M., MOORE, P.D., and ZAHRAN, M.A.1992 Modern pollen rain studies of the Nile Delta, Egypt.
New Phytologist, 121(4): 663–657.BARTZOKAS, A., METAXAS, D.A., KATERI, M., andEXARCHOS, N.
1991 Sea surface temperature in the Mediterranean: statis-tical properties. Journal Rivista di MeteorologiaAeronautica, 51(1–2): 47–64.
BATTEN, D.J.1981 Palynofacies, organic maturation and source poten-
tial for petroleum. In: Brooks, J. (ed.), Organicmaturation studies and fossil fuel exploration. Aca-demic Press, London, 201–223.
1996 Chapter 26B. Palynofacies and palaeoenvironmentalinterpretation. In: Jansonius, J., and McGregor D.C.,(eds.), Palynology: Principles and Applications.American Association of Stratigraphic Palynolo-gists Foundation, Dallas, 3: 1065–1084.
BTHOUX, J.-P.1993 Mediterranean sapropel formation, dynamic and cli-
matic viewpoints. Oceanologica Acta, 16(2): 127–133.
BIANCHI, D., ZAVATARELLI, M., PINARDI, N., CAPOZZI,R., CAPOTONDI, L., CORSELLI, C., and MASINA, S.
2006 Simulations of ecosystem response during the sapro-pel S1 deposition event. PalaeoceanographyPalaeoclimatology Palaeoecology, 235: 265–287.
BIRKS, H.J.B., and BIRKS, H.H.1980 Quaternary palaeoecology. Arnold, London, 289 p.
BRADFORD, M.R., and WALL, D.A.1984 The distribution of recent organic-walled dinoflagel-
late cysts in the Persian Gulf, Gulf of Oman, andnorthwestern Arabian Sea. PalaeontographicaAbteilung B, 192: 16–84.
CALVERT, S.E., and FONTUGNE, M.R.2001 On the late Pleistocene–Holocene sapropel record of
climatic and oceanographic variability in the easternMediterranean. Paleoceanography, 16(1): 78–94.
CARATINI, C., BELLET, J., TISSOT, C., and SEURIN, M.1981 Étude microscopique de la matière organique
palynologie et palynofaciès. In: Caratini, C. (ed.),Géochimie Organique des Sédiments MarinsProfonds, ORGON IV: Golfe d’Aden, Mer d’Oman.Éditions du Centre National de la RechercheScientifique, Paris, 265–307.
CHEDDADI, R., and ROSSIGNOL-STRICK, M.1995 Eastern Mediterranean Quaternary paleoclimates from
pollen and isotope records of marine cores in the Nilecone area. Paleoceanography, 10(2): 291–300.
DALE, B.1996 Chapter 31. Dinoflagellate cyst ecology: modeling
and geological application. In: Jansonius, J., andMcGregor, D.C. (eds.), Palynology: Principles andApplications. American Association of StratigraphicPalynologists Foundation, Dallas, 3: 1249–1275.
DUCASSOU, E., CAPOTONDI, L., MURAT, A.,BERNASCONI, S.M., MULDER, T., GONTHIER, E., MIGEON,S., DUPRAT, J., GIRAUDEAU, J., and MASCLE, J.
2007 Multiproxy Late Quaternary stratigraphy of the Niledeep-sea turbidite system-Towards a chronology ofdeep-sea terrigeneous systems. Sedimentary Geol-ogy, 200: 1–13.
EDWARDS, L.E., and ANDRLE, V.A.S.1992 Distribution of selected dinoflagellate cysts in mod-
ern marine sediments. In: Head, M.J., and Wrenn,J.H. (eds.), Neogene and Quaternary dinoflagellatecysts and acritarchs. American Association of Strati-graphic Palynologists Foundation, Dallas, 259–288.
EL BEIALY, S.Y., EDWARDS, K.J., and EL-MAHMOUDI,A.S.
2001 Geophysical and palynological investigations of theTell El Dabaa archaeological site, Nile Delta, Egypt.Antiquity, 73: 735–744.
EMEIS, K.C., SAKAMOTO, T., WEHAUSEN, R., andBRUMSACK, H.J.
2000 The Sapropel record of the Eastern MediterraneanSea – Results of Ocean Drilling Program Leg-160.Palaeogeography PalaeoclimatologyPalaeoecology, 158: 371–395.
FENSOME, R.A., TAYLOR, F.J.R., NORRIS, G., SARJEANT,W.A.S., WHARTON, D.I., and WILLIAMS, G.L.
1993 A classification of living and fossil dinoflagellates.Micropaleontology Special Publication, 7, 351 p.

22 PALYNOLOGY, VOLUME 33 — 2009
GASSE, F.2000 Hydrological changes in the African tropics since the
Last Glacial Maximum. Quaternary Science Reviews,19(1–5): 189–211.
GROSS, M.G.1975 Carbon analysis. In: Carver, R.E. (ed.), Procedures in
sedimentary petrology. Wiley-Interscience, New York,573–596.
HARLAND, R.1988 Dinoflagellates, their cysts and Quaternary stratigra-
phy. New Phytologist, 108(1): 111–120.HASSAN, F.A.
2007 Extreme Nile floods and famines in Medieval Egypt(AD 930–1500) and their climatic implications. Qua-ternary International, 173/174: 101–112.
HAVINGA, A.A.1964 Investigation into differential corrosion susceptibility
of pollen and spores. Pollen et Spores, 6: 621–635.HUGHES, N.F., and MOODY-STUART, J.C.
1967. Palynological facies and correlation in the EnglishWealden. Review of Palaeobotany and Palynology, 1:259–268.
KHOLEIF, S.E.A.2004 Palynology and palaeovegetation reconstruction in
late Quaternary sediments of the southern Suez Isth-mus, Egypt. Journal of African Earth Sciences, 22(1):93–112.
2007 Palynology of mangrove sediments in the HamataArea, Red Sea Coast, Egypt: vegetation and restora-tion overview. In: Isermann, M., and Kiehl, K. (eds.),Restoration of Coastal Ecosystems. Coastline Re-ports, 7: 5–16.
KREMP, G.O.W.1965 Morphologic Encyclopedia of Palynology. University
of Arizona Press, Tucson, 186 p.KROM, M.D., STANLEY, J.D., CLIFF, R.A., and WOODWARD,
J.C.2002 Nile River sediment fluctuations over the past 7000
years and their key role in sapropel development.Geology, 30(1): 71–74.
LAMB, H.F., BATES, C.R., COOMBES, P.V., MARSHALL,M.H., UMER, M., DAVIES, S.J., and DEJEN, E.
2007 Late Pleistocene dessication of Lake Tana, source ofthe Blue Nile. Quaternary Science Reviews, 26(3–4):287–299.
LASCARATOS, A., ROETHER, W., NITTIS, K., and KLEIN, B.1999 Recent changes in deep water formation and spread-
ing in the Eastern Mediterranean Sea: a review. Jour-nal of Progress in Oceanography, 44: 5–36.
LEROY, S.A.G.1992 Palynological evidence of Azolla nilotica Dec. in
recent Holocene of the eastern Nile Delta and
palaeoenvironment. Vegetation History andArchaeobotany, 1: 43–52.
LIONELLO, P., BHEND, J., BUZZI, A., DELLA-MARTA, P.M.,KRICHAK, S.O., JANSÀ, A., MAHERAS, P., SANNA, A.,TRIGO, I.F., and TRIGO, R.
2006 Cyclones in the Mediterranean region: climatologyand effects of the environment. In: Lionello, P.,Malanotte-Rizzolli, P., and Boscolo, R. (eds.), Medi-terranean Climatic Variability. Developments in Earthand Environmental Sciences, 4: 325–372.
MALDONADO, A., and STANLEY, D.J.1976 The Nile Cone: Submarine fan development by cyclic
sedimentation. Marine Geology, 20: 27–40.1979 Depositional patterns and Late Quaternary evolution
of two Mediterranean submarine fans: a comparison.Marine Geology, 31: 215–250.
MARRET, F., and ZONNEVELD, K.A.F.2003 Atlas of modern organic-walled dinoflagellate cyst
distributions. Review of Palaeobotany and Palynol-ogy, 125: 1–200.
MARRET, F., MUDIE, P.J., AKSU, A.E., and HISCOTT, R.N.2007 A Holocene dinoflagellate cyst record of a two-step
transformation of the Neoeuxinian brackish waterlake into the Black Sea. Quaternary International,167–168, 2007, doi:10.1016/j.quaint.2007.01.010(online).
MARRET, F., MUDIE, P.J., and KHOLEIF, S.2008 Dinoflagellate cyst assemblages in the Black Sea-
Mediterranean Corridor. Eighth International Con-ference on Modern and Fossil Dinoflagellates,Montreal, Canada, May 2008, 33 (abstract).
MARTINEZ-RUIZ, F., PAYTAN, A., KASTNER, M.,GONZÁLEZ-DONOSO, J.M., LINARES, D., BERNASCONI,S.M., and JIMENEZ-ESPEJO, F.J.
2003 A comparative study of the geological and mineral-ogical characteristics of the S1 sapropel in the westernand eastern Mediterranean. PalaeogeographyPalaeoclimatology Palaeoecology, 190: 23–37.
MEHRINGER, P.J., PETERSEN, K.L., and HASSAN, F.A.1979 A pollen record from Birket Qarun and the recent
history of the Fayum, Egypt. Quaternary Research,11: 238–256.
MERTENS, K., RIBEIRO, S., BOUMETARHAN, I., CANER,H., COMBOURIEU-NEBOUT, N., DALE, B., DE VERNAL, A.,ELLEGAARD, M., FILIPOVA, M., GODHE, A., GRØSFJELD,K., HARLAND, R., HOLZWARTH, U., KOTTHOF, U., LEROY,S., LONDEIX, L., MARRET, F., MATSUOKA, K., MUDIE, P.,PEÑA-MANJARREZ, J.L., PERSSON, A., POPESCU, S.,SANGIORGI, A., and LOUWYE, S.
2008 Lingulodinium machaerophoru process length varia-tions in surface sediments: a salinity proxy? EighthInternational Conference on Modern and Fossil Di-

S.E.A. Kholeif, and P.J. Mudie: Palynological records of Pleistocene and Holocene, conditions, Nile Cone, Egypt 23
noflagellates, Montreal, Canada, May 2008, 37(abstract).
MINISTRY OF WATER RESOURCES AND IRRIGATION1993 Egyptian Ministry of Water Resources and Irriga-
tion, Annual Report. Government of Arab Republicof Egypt, Cairo, Egypt, 38 p.
MORZADEC-KERFOURN, M.T.1983 Intérêt des kystes de dinoflagellés pour
l’établissement de reconstitutionpaléogéographique: exemple du Golfe de Gabès(Tunisie). Cahiers de Micropaléontologie, 4: 15–22.
1988 Paléoclimats et paléoenvironments, du tardiglaciaireau Récent, en Méditerranée orientale, à l’est dudelta du Nil; l’apport des microfossiles à membraneorganique. Bulletin des Centres de Recherches Ex-ploration-Production Elf-Aquitaine, 12: 267–275.
MUDIE, P.J., and McCARTHY, F.M.G.2006 Marine palynology: potentials for onshore–offshore
correlation of Pleistocene–Holocene records. Trans-actions of the Royal Society of South Africa, 61(2):135–158.
MUDIE, P.J., ROCHON, A., AKSU, A.E., and GILLESPIE, H.2002a Dinoflagellate cysts and freshwater algae and fun-
gal spores as salinity indicators in Late Quaternarycores from Marmara and Black Seas. Marine Geol-ogy, 190: 203–231.
MUDIE, P.J., ROCHON, A., and AKSU, A.E.2002b Pollen stratigraphy of Late Quaternary cores from
Marmara Sea: land–sea correlation and paleocli-matic history. Marine Geology, 190: 233–260.
MURAT, A., and GOT, H.2000 Organic carbon variations of the eastern Mediterra-
nean Holocene sapropel: a key for understandingformation processes. Palaeoceanography Palaeo-climatology Palaeoecology, 158: 241–257.
OZOY, E., HECHT, A., and UNLUTA, U.1989 Circulation and hydrology of the Levantine Basin:
Result of POEM coordinate experiments 1985–1986. Journal of Progress in Oceanography, 22:125–170.
PINARDI, N., and MOSSETTI, E.2000 Variability of the large-scale General Circulation of
the Mediterranean Sea from observation and mod-eling: a review. Palaeogeography Palaeoclima-tology Palaeoecology, 158: 153–173.
RILEY, D., and SPOLTON, L.1974 World Weather and Climate. Cambridge Univer-
sity Press, 120 p.RITCHIE, J.C.
1987. A Holocene pollen record from Bir Atrun, North-west Sudan. Pollen et Spores, 24(4): 391–410.
ROCHON, A., DE VERNAL, A., TURON, J-L.,MATTHIESSEN, J., and HEAD, M.J.
1999 Distribution of recent dinoflagellate cysts in sur-face sediments from the North Atlantic Ocean andadjacent seas in relation to sea-surface parameters.American Association of Stratigraphic Palynolo-gists Contribution Series, No. 35, 146 p.
ROHLING, E.J., and HILGEN, F.J.1991 The eastern Mediterranean climate at times of sapro-
pel formation: a review. Geologie en Mijnbouw, 70:253–264.
ROSSIGNOL-STRICK, M.1983 African monsoon, an immediate climate response
to orbital insolation. Nature, 304: 46–49.1985 Mediterranean Quaternary sapropels, an immediate
response of the African monsoon to variation ofinsolation. Palaeogeography PalaeoclimatologyPalaeoecology, 49: 237–263.
SAAD, S.I., and SAMI, S.1967 Studies of pollen and spore content of Nile Delta
deposits (Berenbal region). Pollen et Spores, 9(3):467–503.
SCRIVNER, A.E., VANCE, D., and ROHLING, E.J.2004 New neodymium isotope data quantify Nile in-
volvement in Mediterranean anoxic episodes. Ge-ology, 32(7): 565–568.
STANLEY, D.J., and MALDONADO, A.1977 Nile Cone: Late Quaternary stratigraphy and sedi-
ment dispersal. Nature, 266: 129–135.1979 Levantine Sea-Nile Cone lithostratigraphic evolu-
tion: quantitative analysis and correlation with paleo-climatic and eustatic oscillations in the Late Qua-ternary. Sedimentary Geology, 23: 37–65.
SUMMERHAYES, C.P., SESTINI, G., MISDORPS, R., andMARKS, N.
1978 Nile Delta: Nature and evolution of continentalshelf sediments. Marine Geology, 27: 43–65.
TISSOT, B., and PELET, R.1981 Sources and fate of organic matter in ancient sedi-
ments. Oceanologica Acta, Special Issue, Actes26th Congrès International de Gèologie, ColloqueGèologie des Ocèans, Paris 1980, 97–103.
TSIMPLIS, M.N., ZERVALIS, V., JOSEY, S.A., PENEVA,E.I., STRUGLIA, M.V., STANEV, E.V., THEOCHARIS, A.,LIONELLO, P., MALANOTTE-RIZZOLI, P., ARTALE, V.,TRAGOU, E., and OGUUZ, T.
2006 Changes in the oceanography of the Mediterra-nean Sea and their link to climate variability. In:Lionello, P., Malanotte-Rizzoli, P., and Boscolo,R. (eds.), Mediterranean Climate Variability.Developments in Earth and Environmental Sci-ences, 4: 227–282.

24 PALYNOLOGY, VOLUME 33 — 2009
TYSON, R.V.1995 Sedimentary organic matter - organic facies and
palynofacies. Chapman and Hall, London, 615 p.VERSTEEGH, G.J.M., and ZONNEVELD, K.A.F.
2002 Use of selective degradation to separate preservationfrom productivity. Geology, 30: 615–618.
WADIE, W.F.1984 The effect of the regulation of the Nile River dis-
charge on the oceanographic conditions of the south-eastern part of the Mediterranean Sea. Acta Adriatica,25: 29–43.
WALKLEY, A., and BLACK, I.A.1934 An examination of Degtjareff method for determin-
ing soil organic matter and a proposed modificationof the chromic acid titration method. Soil Science,37: 29–37.
WILLIAMS, L.A.1984 Subtidal stromatolites in Monterey Formation and
other organic rich rocks as suggested source con-tributors to petroleum formation. American Associa-tion of Petroleum Geologists Bulletin, 68: 1879–1893.
WILLIAMS, M., TALBOT, M., AHARON, P., SALAAM, Y.A.,WILLIAMS, F., and BRENDELAND, K.I.
2006 Abrupt return of the summer monsoon 15,000 yearsago: new supporting evidence from the lower WhiteNile Valley and Lake Albert. Quaternary ScienceReviews, 25: 2651–2665.
ZONNEVELD, K.A.F.1996 Palaeoclimatic reconstruction of the last deglaciation
(18–8 Ka B.P.) in the Adriatic Sea region; a land-seacorrelation based on palynological evidence.Palaeogeography Palaeoclimatology Palaeoecol-ogy, 122: 89–106.
1997 New species of organic walled dinoflagellate cystsfrom modern sediments of the Arabian sea (IndianOcean). Review of Palaeobotany and Palynology,97: 319–337.
ZONNEVELD, K.A.F., GANSSEN, G., TROELSTRA, S.,VERSTEEGH, G.J.M., and VISSCHER, H.
1997 Mechanisms forcing abrupt fluctuations of the In-dian Ocean summer monsoon during the lastdeglaciation. Quaternary Science Reviews, 16: 187–201.
ZONNEVELD, K.A.F., VERSTEEGH, G.J.M., and DE LANGE,G.J.
2001 Palaeoproductivity and post-depositional aerobicorganic matter decay reflected by dinoflagellate cystassemblages of the Eastern Mediterranean S1 sapro-pel, Marine Geology, 172: 181–195.




















