
Heparan sulfates are critical regulators of the inhibitory ...
Upload
independentCategory
view
2download
0
Pair-wise interactions of polymerization inhibitory contact sitemutations of hemoglobin-S
Sonati Srinivasulu,1 Krishnaveni Perumalsamy,1 Rajendra Upadhya,2 Belur N. Manjula,2
Steven Feiring,4Raouf Alami,
1Eric Bouhassira,
1,3Mary E. Fabry,
1Ronald L. Nagel,
1,2
and A. Seetharama Acharya1,2,5
The linkage of pair-wise interactions of contact site mutations of HbS has been studied using
Le Lamentin [His-20 (a) fi Gln], Hoshida [Glu-43 (b) fi Gln] and a2b2T87Q mutations as the
prototype of three distinct classes of contact sites of deoxy HbS fiber. Binary mixtureexperiments established that bA-chain with the Thr-87 (b) fi Gln mutation is as potent as thec-chain of HbF (a2c2) in inhibiting polymerization. On combining the influence of Le
Lamentin mutation with that of b2T87Q mutations; the net influence is only partial additivity.
On the other hand, in binary mixture studies, combined influence of Hoshida mutation withthat of b2
T87Q mutations is synergistic. Besides, a significant level of synergistic complemen-
tation is also seen when the Le Lamentin and Hoshida mutations are combined in HbS(symmetrical tetramers). Le Lamentin and Hoshida mutation introduced into the cis-dimer ofthe asymmetric hybrid tetramer completely neutralizes the Val-6 (b) dependent polymeriza-
tion. Accordingly, we propose that combining the perturbation of intra-double strand contactsite with that of an inter-double strand contact site exhibit synergy when they are present intwo different chains of the ab dimer. A comparison of the present results with that of theearlier studies suggest that when the two contact site perturbations are from the same sub-unit
of the ab dimer only partial additivity is observed. The map of interaction linkage of thecontact site mutations exposes new strategies in the design of novel anti-sickling Hbs for thegene therapy of sickle cell disease.
KEY WORDS: Sickle cell disease; Linkage of mutations; Synergy; Neutralization gene therapy.
1. INTRODUCTION
The complex clinical manifestations of sickle cellanemia (SCA) are primarily due to the mutation of
the sixth amino acid of the b-chain of hemoglobin(Hb) (Ingram, 1956). The replacement involves thenon-conservative substitution of a charged residue(Glu) with a hydrophobic residue (Val) on the sur-face of the Hb. This change drastically reduces thesolubility of deoxy HbS leading to its polymeriza-tion into long helical fibers. The study of binarymixture of mutant Hbs and HbS has establishedthat the Wishner-Love double strands [WL-DS,found originally in HbS crystals] (Padlan and
1 Division of Hematology Department of Medicine, Albert Ein-
stein College of Medicine, 1300 Morris Park Avenue, Bronx, New
York, 10461, USA.2 Department of Physiology and Biophysics, Albert Einstein Col-
lege of Medicine, 1300 Morris Park Avenue, Bronx, New York,
10461, USA.3 Department of Cell Biology, Albert Einstein College of Medicine,
1300 Morris Park Avenue, Bronx, New York, 10461, USA.4 Dartmouth Medical Center, Medical School, Albert Einstein
College of Medicine, Lebanon, New Hampshire, USA.5 To whom correspondence should be addressed. E-mail: acharya@
aecom.yu.edu
Abbreviations: HbA, Adult human hemoglobin; HbS, Sickle Cell
hemoglobin; HbF, Fetal hemoglobin; SCA, Sickle cell anemia;
WL-DS, Wishner Love double strand; TFA, Trifluro acetic acid;
HPLC, High performance liquid chromatography.
5031572-3887/06/1200-0503/0 � 2006 Springer Science+Business Media, LLC
The Protein Journal, Vol. 25, Nos. 7–8, December 2006 (� 2006)DOI: 10.1007/s10930-006-9034-3Published Online: November 28, 2006

Love, 1985) were the assembly units of the poly-mer (Nagel and Bookchin, 1975; Nagel et al.,1973). Electron microscopy and crystallographicstudies have suggested that both deoxy HbS crystaland the fiber are constructed of 14 strands arrangedin seven double strands (Cretegny and Edelstein,1993; Dykes et al., 1978; Watowich et al., 1993). Aco-operative interaction linkage of the intra and in-ter-double strand contact sites appear to be criticalfor the polymerization reaction (Benesch et al.,1976, 1977; Manning, 1991; Nagel et al., 1980).
Mutations of the known intermolecular contactsites result in small increase in solubility and noneof the known mutations completely neutralize Val-6(b) dependent polymerization. The multiplicity ofthe inter-molecular contact regions in the deoxyHbS polymer and the non-equivalence of the impli-cated contact sites from one tetramer to the other,suggest that non-covalent interactions could resultin synergistic perturbation.
Complete neutralization of Val-6 (b) dependentpolymerization was achieved by pig a-chain as wellas human–pig chimeric a-chain (Rao et al., 2000).The inter-species hybrids of these a-chains withbS-chains carry multiple differences of intra and in-ter-double strand contact sites of the a-chain.Accordingly, the scenario of the complementation ofpolymerization inhibitory potential of the multiplecontact site sequence differences has been proposedas the molecular basis of this neutralization. Thepresence of pig a-chain both in the cis and trans-di-mer is also critical for the complete neutralization i.e.an inter-dimer synergistic complementation shouldalso be invoked to explain the inhibitory potential ofthese super-inhibitory anti-sickling a-chains. Thestrong inhibition of polymerization by mouse a-chain(Roy et al., 1993), horse a-chains and mouse–horsechimeric a-chain (Nacharaju et al., 1997) as well asthe complete neutralization by pig a-chain can beattributed to the different degrees of linkage interactionbetween the sequence differences of inter and intra-double strand contact sites of the a-chain (Nacharajuet al., 1997; Rao et al., 2000; Roy et al., 1993; Srini-vasulu et al., 1999). Mapping the sequence differencesof contact sites to completely neutralize the Val-6 (b)dependent polymerization could be potentially usefulin designing the anti-sickling Hbs for gene therapy ofsickle cell disease.
The concept that the interaction of contact siteperturbations could be used as a design strategy toneutralize the Val-6 (b) dependent polymerizationhas been invoked previously (Acharya et al., 1985;
McCune et al., 1994). Acharya et al. (1985) investi-gated the combined influence of two contact sitemodifications of HbS, both of which are inhibitoryto the polymerization. These studies suggested thepotential for full additivity of two polymerizationinhibitory contact sites by chemical modification i.e.amidation of Glu-43 (b) and reductive alkylation ofa-amino groups of HbS.
The c-chain of fetal hemoglobin (HbF) is themost potent non a-chain in the inhibition of poly-merization through the formation of asymmetrichybrids. Consistent with this, increasing the expres-sion level of c-chain by hydroxy-urea treatmenthas been shown to be beneficial in patients withsickle cell disease (Goldberg et al., 1977; Steinberg,2003).
Binary mixture experiments represent the truein vivo situation of the gene therapy of SCA. Thepresent gene therapy protocol for SCA has beenvery successful in animal models by introducing anintra-double strand lateral contact site perturbationa2b2
T87Q that imitates HbF and represent 90% ofthe HbF effect (Pawliuk et al., 2001). Further in-crease in the inhibitory potential of the mutantbA-chain by additional polymerization inhibitorymutations or by the co-presence of polymerizationinhibitory a-chains, can increase the efficiency of thegene therapy protocol.
Towns and his colleagues (McCune et al.,1994) designed a bA-chain based on the implicatedsequence differences contributing to the polymeriza-tion inhibitory activity of the c-chain of fetal Hb.They have expressed a b-chain with two contact sitemutations. They found only a partial additivitybetween the mutations, b(Glu-22 fi Gln) andb(Thr-87 fi Gln). These mutations represent a com-bination of a intra-double stand axial with a intra-double strand lateral contact site perturbation.
In the present study, we have combined thepolymerization inhibitory influence of inter and in-tra-double strand contact site mutations in a pair-wise fashion contributed by two different chains, a-chains and/or b-chains. The, Thr-87 (b) fi Glnmutation has been chosen as one of the intra-dou-ble strand lateral contact site, a trans-dimer contactsite of HbS polymer. The mutation His-20(a) fi Gln (Le Lamentin), the second site chosen isa intra-double strand axial contact domain site andis active in the cis-dimer of HbS. The third muta-tion chosen is Glu-43 (b) fi Gln (Hoshida muta-tion) a inter-double strand contact domain region.Solution studies have implicated this as a cis-acting
504 Srinivasulu et al.

contact site but the molecular modeling studies haveimplicated it as a trans-dimer contact site. Theinfluence of these combinations has been evaluatedwhen they are present as two copies each contactsite per tetramer or when only one copy is presentper tetramer. The results are presented here.
2. MATERIALS AND METHODS
2.1. Material and Chemicals
All buffer salts used were from Sigma chemicalcompany or Fisher Scientific. Column chromato-graphic resin fast flow Hi-trap Q-Sepharose wasfrom Pharmacia. Normal and sickle human redblood cell (RBC) hemolysates were dialyzed exten-sively against phosphate buffered saline (pH 7.4).The lysates were purified by DE-52 followed byCM-52 cellulose chromatography. The mouse RBClysate expressing the HbS–Le Lamentin mutationwas first dialyzed against PBS and further purifiedusing Hi-trap Q-Sepharose column chromatography
2.2. Generation of Transgenic Mice
Transgenic mice (C57BL6/C3HF2) were createdby co-injection of gel purified DNA fragments. Twolines of mice were generated: 1) Line 1 had a muta-tions in the a-globin gene [Codon 20: CAC (His)CAA (Gln) to generate the Le Lamentin a-chain andwas co-injected with LCR-bS and 2) Line 2 resultedfrom co-injection of LCR-a and the LCR-b mutatedat the 87 position (Thr fi Gln). Mutations wereintroduced by recombinant PCR. Both of these plas-mids contain the mini LCR as described by Forresteret al. (1989) linked to either a bS-globin or a-globingenomic fragment. The genbank coordinates of thevarious fragments are as follows: HS4 fragmentHUMHBB (accession U01317) 951-2199, HS3 fragmentHUMHBB 4274-5122, HS2 fragment HUMHBB7764-9213, HS1 fragment HUMHBB 10947-15180.b-globin gene fragment HUMHBB 61571-65473 anda-globin gene fragment HUMHBA4 (accessionJ00153) 3898-5569.
2.3. Characterization of Transgenic Mouse Lines
Mice with the Le Lamentin-bS (a2H20Qb2
S) trans-gene in the hemizygous state expressed 29.5±0.9%aH20Q and 23.2±1.5% bS as determined by denatur-
ing RP-HPLC as described in analytical methods.Blood from these mice was used for preparative iso-lation of the HbS–Le Lamentin (a2
H20Qb2S).
Mice with the a2b2T87Q (human a, bT87Q) trans-
gene were mated with mice bearing the mouse a-glo-bin knockout (Paszty et al., 1995) and those bearingthe mouse b-globin knockout (Yang et al., 1995), un-til mice homozygous for the a- and b-globin knock-outs were produced that expressed exclusively humanHbs. A minimal number of matings were required toreach homozygosity for the mouse globin knockouts.Knockout mice expressing exclusively human Hbs(full knockout mice with HbA or HbC) were used tofacilitate introduction of a and b-globin knockouts.Lysates from these mice did not require further puri-fication of the Hb component.
2.4. Chemical Modification of HbS
Preparation and chemical characterization ofHbS-Hoshida (a2b2
E6V,E43Q), HbA-Hoshida (a2b2E43Q),
double mutant HbA-Hoshida (a2b2E43Q;T87Q) and
HbS–Le Lamentin–Hoshida (a2H20Qb2
E6V;E43Q) wasachieved by the chemical modification of the b-chain glutamic acid residue at the 43 position withammonia as described earlier (Rao et al., 1995).Chemical characterization of these chemically modi-fied hemoglobins was as previously described (Raoand Acharya, 1994).
2.5. Preparative Anion-Exchange Chromatography
Mutant Hb expressed in transgenic mice waspurified by preparative anion exchange chromatog-raphy on a Hi-trap Q-Sepharose column. Therespective mouse Hb lysates were dialyzed against50 mM tris acetate buffer (pH 8.1) and loaded on acolumn (1.5� 30 cm) that was pre-equilibrated withthe same buffer. The column was eluted using a lin-ear pH gradient of 50 mM Tris acetate buffer, pH8.1–50 mM Tris acetate buffer pH 7.1 with a flowrate of 1 ml/min. About 10 ml fractions are col-lected. A total of 2 L of gradient was employed toelute the proteins from the column. The elutionprofile was monitored at 540 nm.
2.6. Analytical Procedures
Purified mutant Hb molecules from mouse lysateswere analyzed by reverse phase high performance
505Pair-wise interactions of polymerization

liquid chromatography (RP-HPLC) on Vydac C4protein column (250� 4.6 mm) using a 35–50% lin-ear gradient of solvent B (acetonitrile containing0.1% trifluoroacetic acid) in 100 min at a flow rateof 1 ml/min.
2.7. Identification of Site of Mutation and/or
Modification
Mutant a- and b-globin chains were purifiedusing semi-preparative C4 protein (Vydac) column.The mutant Hb a2b2
T87Q was modified with Ma-leidophenylcarbamate PEG-5K. This reagent spe-cifically modified the Cys-93 of b-chain, whichallowed easy isolation of the modified tryptic pep-tide (T10). Purified mutant a-globin and PEGylat-ed b mutant were digested with trypsin [1 mg/mlin 100 mM ammonium bi-carbonate (pH8.0)] at asubstrate to enzyme ratio of 1:200 (W/W) for 3 hat 37�C (Manjula et al., 2003). The reaction wasarrested by adding sufficient amount of TFA tobring the final concentration to 0.3% of TFA inthe reaction mixture. The peptide maps were devel-oped on C18 analytical (Vydac) column with agradient of 5–50% in 100 min at a flow rate of1 ml/min. Modified or mutant peptides were iden-tified by the disappearance of the normal trypticpeptide in the maps, and the presence of new pep-tides, which was isolated and subjected to aminoacid sequence analysis.
2.8. Amino Acid Sequencing
Purified tryptic peptide of a- and b-globin weresubjected to sequence by Edman degradation (Ed-man and Begg, 1967), using an Applied BiosystemProcise 494 sequencer using pulse-liquid chemistrycycles. Eight cycles of Edman degradation was runfor the modified mutant peptide.
2.9. Functional Studies
O2 affinity was measured using a Hem-O-Scan(TCS medical products, Huntington Valley, PA,USA). All the O2 affinities were done at least induplicate. All O2 affinity measurements were carriedout at 37�C, in 50 mM Bis–tris and 50 mM Trisacetate buffer (pH 7.4). Accuracy of the p50 (mmof Hg) and the Hill co-efficient was ±5%.
2.10. Polymerization Studies
Polymerization of HbS samples was evaluatedby the O2 affinity method of Benesch et al. (1978).Oxygenation curves of HbS samples of six differentincreasing Hb concentrations were recorded with aHem-O-Scan at 37�C in 0.1 M phosphate buffer (pH6.8) and measured in duplicate. The solubility con-centration value (CSAT) of the sample is calculatedfrom the intercept of the bi-phasic transition of thep50 as a function of increasing Hb concentration.
2.11. Polymerization Assay in High Phosphate
Buffer
The polymerization of HbS and other semisyn-thetic Hbs was carried out in 2.0 M phosphate buf-fer (pH 7.4) at 30�C. The kinetics of polymerizationwas performed by the method of Adachi and Asak-ura (1979).
3. RESULTS
3.1. Preparation of Mutant and/or Modified Forms
of HbA and HbS
3.1.1. Isolation and Characterization of HbA[Thr-87 (b) fi Gln]
The hemolysates of (RBC) of transgenic mouseexpressing a2b2
T87Q were analyzed on RP-HPLC(Fig. 1A). The RP-HPLC map shows two globinchains; the elution position of one of these twochains corresponds to that of human a-globin chain.The other peak elutes slightly ahead of the bA-globin.
The mass spectral analysis of the globin thatelutes at the position of human a-globin in the RP-HPLC has identified as human a-globin (Table 1).The tryptic peptide mapping of this globin chainconfirmed the identification. The mass of the otherglobin component present in the RP-HPLC was15,894 Da, which is 27 mass units higher than themass of the bA-globin (Table 1) and is consistentwith the mutation of Thr fi Gln residue.
The peptide bT10 corresponds to the aminoacid sequence 83–95 of the b-chain carries the mostreactive Cys residue of Hb. We have previouslyshown that modification of HbA with maleido phe-nyl PEG5K, the Cys-93 (b) modified bA is not di-gested at it�s Lys-95, and the Mal-PEG5K modified
506 Srinivasulu et al.

tryptic peptide bT10 + 11 (composing the residues83–104 of b-chain) elutes significantly later than therest of the tryptic peptides in homogeneous formand easy to isolate (26). This procedure has beenadopted to isolate the mutant peptide since the
mutation (Thr-87 fi Gln) is conservative substitu-tion and identify the modified amino acid residueby amino acid sequencing. This PEGylatedbT10 + 11 elutes at 72.0 min, where as the unmod-ified bT10 elutes at 66.0 min. The PEGylated pep-tide from bA and the mutant human b-globin elutedat the same position. Both peptides were then sub-jected to eight cycles of Edman degradation and theresults are presented in Table 2. The amino acid inthe fifth cycle (position of b-87) from bA-globin isThr and while that from the mutant b-globin isGln. This confirms that the human b-globin ex-pressed in this transgenic mouse is bT87Q. The trans-genic mice expressing a2b2
T87Q is devoid of mouseHb and contained only human Hb as determinedby RP-HPLC. Thus, this transgenic mouse is homo-zygous for a2b
T87Q.
3.1.2. Isolation and Characterization of HbS–LeLamentin
The panel 1B depicts the RP-HPLC profile ofthe transgenic mice RBC lysate expressing HbS–LeLamentin. The profile shows the presence of fiveglobin components, three of which corresponds tothat of mouse Hb. One of these components ismouse a-globin and the other two correspond tomouse b-globin. Presumably the two other globinscorrespond to that of HbS–Le Lamentin expressedin the transgenic mice. The elution profiles of theseare close to that of human a-globin and bS-globinchains.
Fig. 1. The RP-HPLC analysis of the transgenic mouse blood ly-
sate. The transgenic mouse RBC lysate, expressing mutant hemo-
globins, are analyzed on Vydac C4 RP-HPLC analytical column
using a 35–50% acetonitrile gradient containing 0.1% TFA in
100 min. The flow rate was maintained at 1 ml/min. Panel A
shows the elution profile of the mouse RBC lysate expressing
a2b2T87Q and panel B shows the elution profile of the HbS–Le
Lamentin. The respective a- and b-chains are marked in the fig-
ure. The elution profile of purified HbA and mouse Hb are
shown in Inset.
Table 1. Identification of Mutant Globin Chains by Mass Spec-
tral Analysis
Sample Mutation
Molecular Mass (Daltons)
Calculated Experimental
aA-Globin – 15,126.4 15,127.0
aA-Globin H20Q 15,117.4 15,117.0
bA-Globin – 15,867.2 15,867.0
bA-Globin E43Q 15,866.2 15,867.0
bA-Globin T87Q 15,894.2 15,894.0
bA-Globin T87Q;E43Q 15,893.2 15,895.0
bS-Globin E6V 15,837.3 15836.0
bS-Globin E6V;E43Q 15,836.3 15,837.0
Table 2. Confirmation of the Engineered Mutations in HbS [His-
20 (a) fi Gln] and HbA [Thr-87 (b) fi Gln]
Tryptic peptide (a) T4aPEGylated tryptic peptide
(b T10 + 11b
Cycle From HbA From mutant From HbA From mutant
1 Val Val (846.6) Gly Gly (121.6)
2 Glu Glu (610.6) Thr Thr (100.1)
3 Ala Ala (758.4) Phe Phe (141.1)
4 His Gln (673.7) Ala Ala (131.8)
5 Ala Ala (668.7) Thr Gln (116.0)
6 Gly Gly (504.2) Leu Leu (164.3)
7 Glu Glu (471.3) Ser Ser (43.6)
8 Tyr Tyr (501.0) Glu Glu (83.9)
Tryptic peptides were subjected to automated Edman degradation.
Identified mutations in the tryptic peptide are shown in bold. The
recovery of amino acid is shown in parenthesis. aFrom HbS–Le
Lamentin. bFrom a2b2T87Q.
507Pair-wise interactions of polymerization

The molecular mass of the globin chain elutingat the position of b-globin is 15,836 Da and corre-sponds to that of bS. The tryptic peptide map ofthis globin has confirmed to be bS-globin chain. Onthe other hand the mass of the globin eluting at theposition of the human a-chain is 15,117 Da, lowerthan that of human a-globin and this is consistentwith the His fi Gln mutation. The tryptic peptidemap of this a-globin corresponds well with the tryp-tic peptide map of control a-globin except for theposition of aT4 (residue 17–31 of a-chain). In nor-mal a-globin this peptide elutes around 29.4 min onRP-HPLC, but this peptide was absent in the hu-man a-globin from transgenic mice. A new peakeluting around 30.8 min was present. There are noother noticeable differences in the tryptic peptidemap. The normal aT4 eluting at the 29.4 min aswell as this new peptide eluting around 30.8 minhas been subjected to eight cycles of Edman degra-dation (Table 2). The amino acid sequence of aT4from control a-globin corresponded to the aminoacid residues of 17–24 of a-globin. In the new tryp-tic peptide eluting at 30.8 min, first three residuescorresponded to human a-globin and the fourth res-idue was Gln instead of His of normal aT4. Thenext four residues also corresponded to the normal
a-globin. Thus, this study identifies the mutation inthis a-globin as His-20 (a) fi Gln.
The transgenic mouse line expressing aA LeLamentin (mutant a-chain) and bS-chain also con-tains mouse globin chains. Thus, one can anticipateat least four different species of of Hb tetramers inthe mouse RBC lysate. These include HbS–Le Lam-entin, mouse Hb, and the two possible inter-specieshybrids formed by the exchange of either a- or b-sub-units of two parent molecules: HbS–Le Lamen-tin and mouse Hb. Accordingly, the purification ofHbS–Le Lamentin (a2
H20Qb2S) from the transgenic
mouse RBC lysate was accomplished using Hi-trapQ-Sepharose column (1.5� 30 cm) chromatography.
The chromatogram of the mouse RBC lysateexhibits well resolved into four species of Hb is gi-ven in Fig. 2A. The identification of these fourpeaks has been established by RP-HPLC (Fig. 2B).The four Hbs in the order of their elution fromQ-Sepharose column are HbS–Le Lamentin, mousea- and human bS-inter species hybrid (a2
Mb2S),
Human a-Le Lamentin and mouse b inter specieshybrid (a2
H20Qb2M) and mouse Hb (a2
Mb2M) respec-
tively. The fractions corresponding to HbS–LeLamentin and others were pooled separately andconcentrated and processed further.
Fig. 2. Purification of the HbS–Le Lamentin on high trap Q-Sepharose column. The chromatographic separation of the HbS–Le Lamen-
tin from the transgenic mouse RBC lysate (panel A) was achieved by hi-trap Q-Sepharose column (1.5� 30 cm) at 4�C. The column was
eluted with a gradient formed between 50 mM Tris acetate (pH 8.1) and 50 mM Tris acetate (pH 7.1) with 1000 ml each of buffer. About
10 ml fractions are collected. The absorbance of the samples is measured at 540 nm. The identification of the peaks was done by RP-
HPLC analysis as in Fig. 1 and the profile of the each peak is given in panel 3B.
508 Srinivasulu et al.

3.1.3. Preparation of HbS-Hoshida, HbS–Le-Lamen-tin–Hoshida, HbA-Hoshida and a2b2
T87Q–Hoshida
The Hoshida mutation was introduced into theparent Hbs by the amidation reaction as discussedunder methods and each of the amidated Hb waspurified from the unmodified parent material byion-exchange chromatography as described undermethods (Rao and Acharya, 1994; Rao et al., 1995).The identity of these various forms established byRP-HPLC, molecular mass (Table 1) and trypticpeptide mapping respectively.
3.1.4. Homogeneity of Mutant HbS Expressed inTransgenic Mice and their Amidated(Hoshida) Forms
The homogeneity of the various Hb sampleswas established by iso electric focusing (IEF). TheIEF pattern is given in Fig. 3. The IEF pattern ofHbS–Le Lamentin (aH20Q) was distinct from that ofHbS and it is focused between the HbS and HbA.The loss of one positive charge per dimer of HbScontributed to the difference in the IEF pattern.The electrophoretic mobility of a2b2
T87Q, was iden-tical to that of normal human HbA, consistent withthe fact that the substitution of Thr by Gln does
not contribute to the total net charge of the mole-cule.
In the case of HbS-Hoshida, the loss of thenegative charge of c-carboxyl group of Glu-43 (b) isseen clearly in the IEF pattern. Similarly, the IEFpattern of amidated mutant a2b2
T87Q,E43Q was dis-tinct from that of a2b2
T87Q. The amidation of HbS–Le Lamentin alters it�s IEF pattern much the sameway as that resulting from the amidation of HbS toHbS-Hoshida. Thus, these studies clearly establishthe homogeneity of the samples used in the presentstudy.
3.2. Functional Properties of Mutant/Modified HbS
The O2 affinity of the Hb samples was mea-sured in 50 mM Bis–Tris and 50 mM Tris-acetate,pH 7.4 at 37�C using a Hem-O-Scan (Table.3). Thep50 of HbS–Le Lamentin (7.8) is very close to thatof native HbS (7.4). It binds O2 co operatively withan n- value (Hill co-efficient) of 2.3. However, theO2 affinity of a2b2
T87Q was lower than that of HbS.The p50 value for the a2b2
T87Q mutation was 9.2.The Hill�s co-efficient for this mutant HbA was 2.3.The O2 affinity of HbS-Hoshida is slightly higherthan that of HbS; but has very limited influence onthe cooperativity. Thus, all the three mutations,HbS–Le Lamentin, a2b2
T87Q and HbS-Hoshidahave only marginal influence on the O2 affinity ofHb. Amidation of HbS–Le Lamentin and a2b2
T87Q
resulted in slight increase in the O2 affinity.The modulation of the O2 affinity of Hb by
DPG in the presence of mutant Hbs is also pre-
Table 3. Oxygen Affinity of the Mutant Hemoglobins
Sample
50 mM Bis–Tris/50 mM Tris acetate (pH
7.4)
Without DPG With DPG p50 with
DPG/p50
without DPG
p50 (n) p50 (n)
HbS 7.4 (2.3) 24.0 (2.0) 3.3
HbS–Le Lamentin 7.8 (2.2) 17.0 (2.0) 2.0 flHbS-Hoshida 6.0 (2.2) 23.0 (2.0) 3.9 ›HbS–Le
Lamentin–Hoshida
6.5 (2.2) 18.0 (2.0) 2.8 fl
a2bT87Q 9.2 (2.2) 25.0 (2.0) 2.6 fl
a2bE43Q;T87Q 6.2 (2.2) 22.0 (2.0) 3.5 ›
O2 affinity was measured using Hem-O-Scan at 37�C.p50: Partial pressure of O2 at 50% of Hb saturation.
n = Hill’s co-efficient (co operativity).
Fig. 3. The Isoelectric focusing pattern of mutant hemoglobins.
The measurements were done on a pre cast IEF gels with a gra-
dient between pH 6 and 8. The focusing was carried out at 100 A
and 1200 V for about 50 min. After the run the protein bands
were fixed with 10% trichloro acetic acid for about 3–5 min and
washed with distilled water for 1 h and dried. The corresponding
samples are labeled in the figure.
509Pair-wise interactions of polymerization

sented in Table 3. Overall influence of DPG on theO2 affinity of HbS-Hoshida and a2b2
T87Q is compa-rable to that of HbS. However, the O2 affinity ofHbS–Le Lamentin is less sensitive to the presence ofDPG than HbS-Hoshida and a2b2
T87Q. The slightlyincreased O2 affinity of HbS–Le Lamentin noted inthe absence of allosteric effector was magnified inthe presence of DPG. The consequences of a muta-tion at the AB-corner of a-chain, is communicatedto the bb-cleft, a result that was apparent from ourrecent studies with Hb-Einstein (Srinivasulu et al.,2004). The mutant a2b2
T87Q exhibits higher O2 affin-ity in the presence of DPG. Thr-87 (b) fi Glnmutation lowers the sensitivity of Hb to DPG alinkage of acceptor pocket of Hb, with bb-cleft.
3.3. Combined Influence of the Two-Polymerization
Inhibitory Mutations
The combined influence of two polymerizationinhibitory mutations could be studied by targetingthe two inhibitory mutations to both dimers of HbStetramer (polymerization behavior of symmetricalmolecule). The influence of chosen mutation on thesolubility can also be studied by binary mixturestudies wherein one mutation is targeted into eitherthe a- or b-chains of the dimer that carries the Val-6 (b) and referred to as the cis-dimer and the sec-ond mutation on to the chains of the other dimerthat provides the acceptor pocket (trans-dimer) oralternatively both mutations targeted to either cis ortrans dimer. It should be noted that the binary mix-ture studies represent the real in vivo situation ofinhibiting the polymerization through gene therapy.
3.3.1. Pair-Wise Combination of Two PolymerizationInhibitory Mutations in the Symmetrical Molecule
The polymerization behavior of HbS, HbS-Hoshida, HbS–Le Lamentin and HbS–Le Lamen-tin–Hoshida is presented in Table 4. The solubilityof HbS–Le Lamentin is 21.5 g/dl and this represents
a 26% increase in the solubility of HbS(Table 4). On the other hand the solubility ofHbS-Hoshida is about 41% higher than HbS(CSAT value for the HbS-Hoshida is 24 g/dl), con-siderably higher than that of HbS–Le Lamentin.When Le Lamentin and Hoshida mutations com-bined together on the HbS, the solubility of HbSwith two mutations is about 33 g/dl. This repre-sents a solubility increase of 94% compared toHbS. As a result of engineering the Hoshidamutation, the equilibrium solubility of HbS–LeLamentin is increased by about 53%. Viewed theother way, the solubility of HbS-Hoshida is in-creased by about 38% when the Le Lamentinmutation is engineered. Thus, it is clear thatwhen the two mutations, one coming from inter-double strand and the other from intra-doublestrand axial contact domain, are present togetheralong with the sickle mutation, they appear todevelop a degree of synergy in inhibiting the Val-6 (b) dependent polymerization reaction.
3.3.2. Polymerization Inhibitory Potential of Thr-87(b) fi Gln Mutation in the Trans-Dimer
The solubility of equi-molar mixture of HbSand a2b2
T87Q has been compared with that of equi-molar mixture of HbS and HbA. The presence ofthe mutation Thr-87 (b) fi Gln in the trans-dimerof the asymmetric hybrid of HbS and HbA
Table 5. Polymerization Inhibitory Potential of a2b2T87Q in
Binary Mixture Studies
Sample
Position
of
mutation
Solubilitya
(g/dl)
Contact
site
Location
% Increase
over that of
(HbS + HbA)b
HbS None 17.0 – –
HbS + HbA Trans 23.5 Lateral –
HbS + a2b2T87Q Trans 29.5 Lateral �26
HbS + HbF Trans 30.0 Lateral �26
a solubility measured by oxygen affinity method.b % increase compared with that of binary mixtures.
Table 4. Pair-Wise Interaction of Le Lamentin and Hoshida Mutations in HbS
Sample Position Solubilitya (g/dl) Contact site location % Increase in solubility
HbS Cis & Trans 17.0 Lateral –
HbS–Le Lamentin Cis & Trans 21.5 Axial 26
HbS-Hoshida Cis & Trans 24.0 Inter-double strand 41
HbS–Le Lamentin–Hoshida Cis & Trans 33.0 Axial & Inter-double strand 94
a Solubility measured by oxygen affinity method as in materials and methods.
510 Srinivasulu et al.
increased the solubility of the control (equi-molarmixture of HbS and HbA) sample by nearly 26%(Table 5). The solubility of equi-molar mixtures ofHbS and HbF is also given in Table 5 for compari-son. Interestingly, this increase in the solubility ofHbS by the presence of a2b2
T87Q is nearly compara-ble to that of HbF. Therefore, the polymerizationinhibitory potential of bT87Q mutation in the trans-dimer of the asymmetric hybrid is comparable tothat afforded by the c-chain of HbF in trans-dimer.The results suggest that though the c-chain hasother sequence differences at the implicated intermolecular contact sites these do not appear to con-tribute to polymerization inhibition in the presenceof Thr-87 (b) fi Gln.
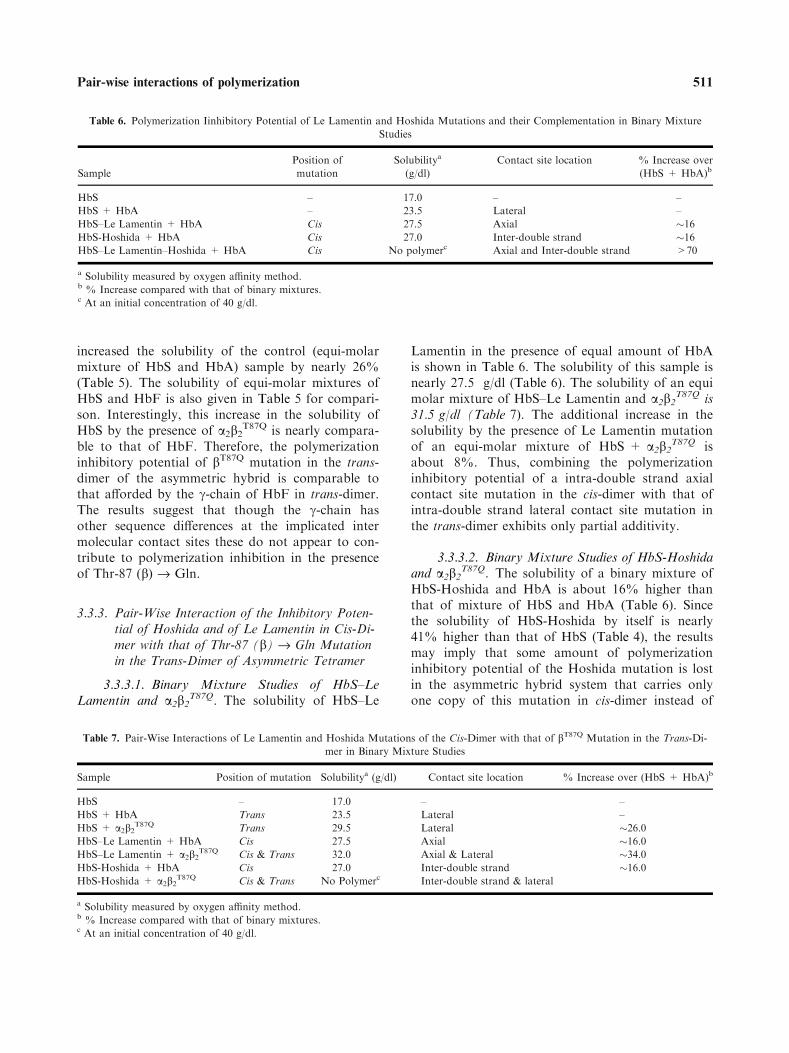
3.3.3. Pair-Wise Interaction of the Inhibitory Poten-tial of Hoshida and of Le Lamentin in Cis-Di-mer with that of Thr-87 (b) fi Gln Mutationin the Trans-Dimer of Asymmetric Tetramer
3.3.3.1. Binary Mixture Studies of HbS–LeLamentin and a2b2
T87Q. The solubility of HbS–Le
Lamentin in the presence of equal amount of HbAis shown in Table 6. The solubility of this sample isnearly 27.5 g/dl (Table 6). The solubility of an equimolar mixture of HbS–Le Lamentin and a2b2
T87Q is31.5 g/dl (Table 7). The additional increase in thesolubility by the presence of Le Lamentin mutationof an equi-molar mixture of HbS + a2b2
T87Q isabout 8%. Thus, combining the polymerizationinhibitory potential of a intra-double strand axialcontact site mutation in the cis-dimer with that ofintra-double strand lateral contact site mutation inthe trans-dimer exhibits only partial additivity.
3.3.3.2. Binary Mixture Studies of HbS-Hoshidaand a2b2
T87Q. The solubility of a binary mixture ofHbS-Hoshida and HbA is about 16% higher thanthat of mixture of HbS and HbA (Table 6). Sincethe solubility of HbS-Hoshida by itself is nearly41% higher than that of HbS (Table 4), the resultsmay imply that some amount of polymerizationinhibitory potential of the Hoshida mutation is lostin the asymmetric hybrid system that carries onlyone copy of this mutation in cis-dimer instead of
Table 6. Polymerization Iinhibitory Potential of Le Lamentin and Hoshida Mutations and their Complementation in Binary Mixture
Studies
Sample
Position of
mutation
Solubilitya
(g/dl)
Contact site location % Increase over
(HbS + HbA)b
HbS – 17.0 – –
HbS + HbA – 23.5 Lateral –
HbS–Le Lamentin + HbA Cis 27.5 Axial �16HbS-Hoshida + HbA Cis 27.0 Inter-double strand �16HbS–Le Lamentin–Hoshida + HbA Cis No polymerc Axial and Inter-double strand >70
a Solubility measured by oxygen affinity method.b % Increase compared with that of binary mixtures.c At an initial concentration of 40 g/dl.
Table 7. Pair-Wise Interactions of Le Lamentin and Hoshida Mutations of the Cis-Dimer with that of bT87Q Mutation in the Trans-Di-
mer in Binary Mixture Studies
Sample Position of mutation Solubilitya (g/dl) Contact site location % Increase over (HbS + HbA)b
HbS – 17.0 – –
HbS + HbA Trans 23.5 Lateral –
HbS + a2b2T87Q Trans 29.5 Lateral �26.0
HbS–Le Lamentin + HbA Cis 27.5 Axial �16.0HbS–Le Lamentin + a2b2
T87Q Cis & Trans 32.0 Axial & Lateral �34.0HbS-Hoshida + HbA Cis 27.0 Inter-double strand �16.0HbS-Hoshida + a2b2
T87Q Cis & Trans No Polymerc Inter-double strand & lateral
a Solubility measured by oxygen affinity method.b % Increase compared with that of binary mixtures.c At an initial concentration of 40 g/dl.
511Pair-wise interactions of polymerization

two copies per tetramer. A small but noticeable(5%) increase is seen when the Hoshida mutation isplaced in the trans-dimer. The results suggest thatsegregation of the polymerization inhibitory poten-tial of the Hoshida mutation into the cis-dimer andtrans-dimer, significant amount of the polymeriza-tion inhibitory activity of the Hoshida mutation islost. Hence, to achieve the maximum polymeriza-tion inhibition by the Hoshida mutation, the muta-tion needs to be present on both cis and trans-dimers of the molecule. This situation of Hoshidamutation acting from both cis and trans-dimer iscomparable to that of HbS-Sealy mutation (Cre-peau et al., 1981).
On the other hand, when the Hoshida muta-tion present in the cis-dimer and the Thr-87(b) fi Gln mutation is present in the trans-dimer,binary mixtures do not polymerize even at a con-centration of 40 g/dl. These two mutations result ina complete neutralization of Val-6 (b) dependentpolymerization. The combined polymerization inhi-bition of the two mutations is much stronger thaneach one of them. The Hoshida and Thr-87(b) fi Gln mutation synergistically complementswith Thr-87 (b) fi Gln to completely neutralizeVal-6 (b) dependent polymerization. The solubilityof an equi molar mixtures of HbS and a2b2
E43Q,T87Q
is comparable to that of equi molar mixtures ofHbS and a2b2
T87Q (Table 6). bE43Q mutation is si-lent when present in the trans-dimer along withThr-87 (b) fi Gln mutation.
HbS–Le Lamentin–Hoshida in the presence ofHbA did not polymerize even at 40 g/dl suggestingthat, when a second bS-chain is replaced by bA-chain, there is an additional polymerization inhibi-tion by Le Lamentin–Hoshida combination com-pare to the sum of these two contact sitesindividually (Table 6).
3.4. Polymerization Assay in High Phosphate Buffer
We have also studied the polymerization inhibi-tion of these mutant hemoglobins individually andpair-wise fashion in 2.0 M phosphate buffer and theresults are given in Fig. 4. The solubility of HbS–LeLamentin and HbS-Hoshida were higher than HbS(Fig. 4A). The relative increase in solubility ofHbS–Le Lamentin was 32% higher, while that ofHbS-Hoshida was 48% higher than that of HbS.The increase in solubility in high phosphate bufferwas slightly higher but comparable to that of
oxygen affinity method for these mutant hemoglo-bins. The solubility of mutant HbA (bT87Q) wasestablished by binary mixture studies in high phos-phate buffer (Fig. 4B). The increase in the solubilityof equi-molar HbS + HbA (bT87Q) was 28% higherthan that of HbS + HbA and comparable to thatof oxygen affinity method.
The synergistic complementation of thesemutant hemoglobins is also studied in high phos-phate buffer in a pair-wise fashion and the resultsare given in Fig. 4B. The solubility of the equi-mo-lar mixture of HbS–Le Lamentin and HbA (bT87Q)was 74% higher than that of equi-molar mixtures ofHbS + HbA, thus suggesting a additive effect ofthese mutations when combined. However, the solu-bility of the equi-molar mixture of HbS-Hosh-ida + HbA (bT87Q) was 300% higher than that of
Fig. 4. Polymerization assay of mutant hemoglobins in high
phosphate buffer. The kinetics of polymerization of mutant
hemoglobins was done in 2.0 M phosphate buffer (pH 7.4). The
solubility of the mutant hemoglobin was measured after centrifu-
gation of the sample at 555 nm. Panel A shows the solubility of
symmetric hemoglobin sample. (-s- HbS, -D- HbS–Le Lamentin
and -h- HbS-Hoshida). Panel B shows the solubility of the
asymmetric hemoglobin samples generated by binary mixture
studies. (-•- HbS + HbA, -x- HbS + HbA (bT87Q), -m-HbS- Le
Lamentin + HbA (bT87Q) and -n- HbS-Hoshida + HbA (bT87Q).
512 Srinivasulu et al.

HbS + HbA. Taken together these result suggestthat inter double strand contact site in cis-dimer(HbS-Hoshida) and intra-double strand lateral con-tact site [HbA (bT87Q)] in trans-dimer synergisticallycomplement to inhibit the Val-6 (b) dependent poly-merization.
4. DISCUSSION
The pair-wise interaction of the polymerizationinhibitory mutations of HbS (three intermolecularcontact sites) has been investigated in HbS, as wellas in binary mixture of HbS/mutated HbS and HbAand mutated HbA. The pair-wise interactions of thecontact sites studied here are schematically shown in
Fig. 5. Panel A shows the generation of Hbs withtwo mutations of contact domains acting from thecis-dimer of HbS. One mutation is active from thecis-a-chain (Le Lamentin mutation) and the other isactive from the bS-chain (Hoshida mutation). Themutations are introduced on both copies of the aand b-chains of the tetramer (symmetrical molecule).
In panel B, the scheme for mapping the conse-quences of introducing a single copy of contact sitemutation into HbS through binary mixture studiesthat facilitates the generation of an asymmetric hy-brid in situ is presented. The mutation can be selec-tively placed in the trans-dimer of the asymmetricmolecule by having the contact site mutations inHbA. Similarly, by having the desired mutation inthe HbS, the mutation can be selectively placed in
Fig. 5. Schematic representation of the pair-wise combination of polymerization inhibitory mutations: Panel A shows the strategy for the
incorporation of the intra-double strand axial contact site in the a-chain (aH20Q) and inter-double strand contact site in the b-chain(bE43Q) of the sickle hemoglobin. Panel B shows the incorporation of intra double strand contact site (bT87Q) of bA-chain into the tetra-
mer in binary mixture studies. Panel C shows the incorporation of either the intra-double strand axial or the inter-double strand contact
sites in the cis-dimer and the intra-double strand lateral contact site in the trans-dimer in binary mixture studies. Only one of the possibil-
ities is shown here.
513Pair-wise interactions of polymerization

the cis-dimer of the asymmetric molecule. Panel Cdepicts binary mixture experiments when a desireddouble mutant HbS is mixed with HbA or mutantHbA, the strategy to introduce three mutations intothe molecule in a stereo specific fashion.
HbS-Hoshida inhibits polymerization by actingfrom both the cis- and trans-dimers of the polymerand a synergistic interaction linkage exists betweenit’s inhibitory role in the cis- and trans-dimers.When the contribution of one copy of Hoshidamutation in inhibition of polymerization is consid-ered, stronger inhibition resides in the cis-dimer, butsome polymerization inhibition is also seen whenthe Hoshida mutation is placed in the trans-dimer.But when mutation was placed in both the dimersthe inhibition is very strong, reflecting the synergybetween the cis and trans inhibitory potential in thesymmetric hybrid. The solubility of deoxy doublemutant, HbS–Le Lamentin–Hoshida, demonstratesthat the Hoshida and Le Lamentin mutation exhib-its synergy in their polymerization inhibition.
The results of the present study also establishthat polymerization inhibition by a2b2
T87Q is com-parable to that HbF (a2c2) (Goldberg et al., 1977).In earlier studies, it has been suggested that c87Gln,c80 Asp and c22 Ala are the sites responsible forthe inhibition of the polymerization by the c-chain.The present studies showed that contact site se-quence differences of c-chain other than that ofThr-87 (b) fi Gln are not significant contributors tothe polymerization inhibition. i.e. the contact sitemutations Asp-80 and Glu-22 do not exhibit anyadditivity in the presence of Gln-87 (c).
The above results are consistent with the stud-ies of McCune et al. (1994) that has shown thatbE22A,T87Q inhibits polymerization comparable toHbF, implying that Glu-22 fi Ala mutation intro-duced into bchains of the trans-dimer of HbS doesnot play any role in the presence of Thr-87(b) fi Gln mutation. Similarly, the Thr-87(b) fi Lys mutation inhibits polymerization as dem-onstrated in binary mixture experiments usingHbD-Ibadan as well as by the solubility of HbS D-Ibadan (Ho et al., 1996). The influence of side chainat b(87) on polymerization was also studied byreplacing Thr with Leu, Ile, Trp, Asp and Lys bydelay time kinetics of polymerization (Adachi et al.,1994; Witkowska et al., 1991). The interaction link-age of Asp-80 (b) with that of Gln-87 (b) has notbeen mapped yet.
An equi-molar mixture of HbS-Hoshida anda2b2
T87Q does not polymerize even at a concentra-
tion of 40 g/dl of tetramer. Accordingly, we con-clude that combining a inter-double strand contactsite mutation of cis-dimer with that of a intra-dou-ble strand lateral contact site mutation, bT87Q oftrans-dimers results in a synergistic complementa-tion of their polymerization inhibition.
The pattern of synergy reported here, is differ-ent than those by Himanen et al. (1996) and byMcCune et al. (1994), where a contact site mutationis introduced in the same b-chain. The two pertur-bations targeted to the b-chain of the trans-dimer.In the latter studies, the contact site mutations areengineered in to the same b-chain.
In the present study, the two mutations areintroduced one each into the a and bS-chains ofHbS. The pair-wise interaction seen in HbS–LeLamentin–Hoshida is synergistic. On the otherhand, an equi-molar mixture of HbS–Le Lamentin–Hoshida and HbA does not polymerize. In the bin-ary mixture system, Hoshida and Le Lamentinmutation exhibits a synergistic interaction. In allcases the synergistic complementation is seen whenthe two contact site mutations are present either intwo chains (sub-units) of the same ab-dimer, or inthe case of Hoshida mutation, the perturbations areplaced in the b-chains of two different dimers.
The level of polymerization inhibition by thepair-wise Le Lamentin–Hoshida mutations in bin-ary mixture studies, implies that in the absence ofthe second Val-6 (b) in the Wishner-Love double-strand, either the stability of the double-strand itselfor assembly of the double strands into the fiber orboth are influenced much more. The current modelsof the quinary structure of deoxy HbS do not dif-ferentiate between the fibers of symmetric tetramerand asymmetric tetramer. It may be noted that theinhibition of polymerization by the Hoshida muta-tion in one copy (cis or trans) does not account forthe complete inhibition by the Hoshida mutation inHbS. The assignment of the Hoshida mutation asactive from the cis-dimer is also not consistent withthe current models of deoxy HbS fiber, whichimplies that this Hoshida mutation acts from thetrans-dimer of HbS. This reflects potential differ-ences in the polymer stability of symmetric hybridsand of asymmetric hybrids and the need for thecross correlation of the quinary structure of deoxyHbS as reflected by the symmetrical molecule andasymmetric hybrid, aAbSaAbA.
The linkage of the interaction of the polymeri-zation inhibitory contact sites of HbS that havebeen studied so far is presented in Table 8. Most of
514 Srinivasulu et al.

the double mutants in HbS system (homo tetramer)exhibited only a partial additivity. On the otherhand the contact site mutation combination of a in-tra-double strand lateral contact site mutation withthat of a intra-double strand axial contact sitemutation resulted in partial additivity. The latter re-sult is in good agreement with the earlier studies ofthe similar combination of contact sites, aP114R/bT87K (Ho et al., 1996). Similarly, the combinationof two intra-double strand axial contact sites,aK16Q/aL113H or aE23Q/aL113H is also failed to exhibitsynergistic additivity (Sivaram et al., 2001).
In the present study, the double mutantaH20Q/bE43Q (combination of intra-double strandaxial and inter-double strand) exhibited a synergyin HbS polymerization inhibition. The completeneutralization of the HbS polymerization seenhere on the combination of one intra-doublestrand lateral and one inter-double strand contactsites, bT87Q/bE43Q in binary mixture studies isinteresting and important and distinguishes fromother studies. Similarly the pair-wise combination
of the intra-double strand axial contact site andinter-double strand contact site perturbation(aH20Q/bE43Q) is also synergistic in the binary mix-ture studies. However, these results are in contrastto the earlier observation of similar set of combi-nation of contact sites from b-chain, bL88A/bK95I
that exhibited only partial additivity. These studiesestablish a synergistic complementation of poly-merization inhibitory mutation, when a intra- andinter-double strand contact sites are combined.The two contact site perturbations need to bespatially separated either by being in two differentchains of the same dimer or in two different dim-mers to achieve synergy. These results can facili-tate the design of new anti-sickling Hbs for thegene therapy of sickle cell anemia.
ACKNOWLEDGMENTS
This work was supported by following NationalInstitute of Health grants; NIH/NHLBI 1M01 RR
Table 8. Compilation of all the Known Pair-Wise Interaction of Contact Site Mutations of HbS
Contact site mutations Solubility Interaction linkage
Pattern of interactionSingle Double HbS Binary mixture HbS Binary mixture
aH20Q(IDA) 21.5a
bE43Q(ID) 24.0a
bT87Q(IDL) 29.5a
aH20Q/bE43Q) 32.0a Synergy Synergy A&D
aH20Q/bT87Q 31.5a Partial additive A&L
bE43Q/bT87Q >40.0a Synergy L&D
aK16Q(IDA) 39.0b
aE23Q(IDA) 44.0b
aL113H(IDA) 53.0b
aK16Q/aL113H 60.0b Partial additive A&A
aE23Q/aL113H 55.0b Partial additive A&A
aP114R(IDA) 33.0c
bT87K(IDL) 27.0c
aP114R/bT87 K >33.0c Partial additive A&L
bL88A(IDL) 67.0b
bK95I(ID) 90.0b
bL88A/bK95I 91.0b Partial additive L&D
bE22A(IDA)
bT87Q(IDL)
bE22A/bT87Q No Additivity A&L
The molecular contact sites are identified as; A: Intra-double strand axial, L: intra-double strand lateral and D: inter-double strand.a determined by oxygen affinity method.b determined by dextran method.c determined by equilibrium gelation method.
515Pair-wise interactions of polymerization

12248, NIH/NHLBI 5P60HL-38655, NIH/NHLBIP01HL55435, and PO1 HL58512.
REFERENCES
Acharya, A. S., Sussman, L. G., and Seetharam, R. (1985).J. Protein Chem. 4: 215–225.
Adachi, K., and Asakura, T. (1979). J. Biol. Chem. 254: 7765–7771.Adachi, K., Konitzer, P., and Surrey, S. (1994). J. Biol. Chem. 269:
9562–9567.Benesch, R. E., Kwong, S., Benesch, R., and Edalji, R. (1977).
Nature 269: 772–775.Benesch, R. E., Yung, S., Benesch, R., Mack, J., and Schneider, R.
G. (1976). Nature 260: 219–221.Benesch, R. E., Edalji, R., Kwong, S., and Benesch, R. (1978). Anal
Biochem. 89: 162–173.Crepeau, R. H., Edelstein, S. J., Szalay, M., Benesch, R. E.,
Benesch, R., Kwong, S., and Edalji, R. (1981). Proc. Natl.Acad. Sci. 78: 1406–1410.
Cretegny, I., and Edelstein, S. J. (1993). J. Mol. Biol. 230: 733–738.Dykes, G., Crepeau, R. H., and Edelstein, S. J. (1978). Nature 272:
506–510.Edman, P., and Begg, G. A. (1967). Eur. J. Biochem. 1: 80–91.Forrester, W. C., Novak, V., Gelinas, R., and Groudine, M. (1989).
Proc. Natl. Acad. Sci. (USA) 86: 5439–5443.Goldberg, M. A., Husson, M. A., and Bunn, H. F. (1977). J. Biol.
Chem. 252: 3414–3421.Himanen, J. P., Mirza, U. A., Chait, B. T., Bookchin, R. M., and
Manning, J. M. (1996). J. Biol. Chem. 271: 25152–25156.Ho, C., Willis, B. F., Shen, T. J., Dazhen, N. T., Sun, D. P., Tam,
M. F., Suzuka, S. M., Fabry, M. E., and Nagel, R. L. (1996).J. Mol. Biol. 263: 475–485.
Ingram, V. M. (1956) Nature 178: 792–794.Manjula, B. N., Tsai, A., Upadhya, R., Perumalsamy, K., Smith,
P. K., Malavalli, A., Vandegriff, K., Winslow, R. M.,Intaglietta, M., Prabhakaran, M., Friedman, J. M., andAcharya, A. S. (2003). Bioconjug. Chem. 14: 464–472.
Manning, J. M. (1991) Adv. Enzymol. Relat. Areas Mol. Biol. 64:55–91.
McCune, S. L., Reilly, M. P., Chomo, M. J., Asakura, T., andTowens, T. M. (1994). Proc. Natl. Acad. Sci. 91: 9852–9856.
Nacharaju, P., Roy, R. P., White, S. P., Nagel, R. L, and Acharya,A. S. (1997). J. Biol. Chem. 272: 27869–27876.
Nagel, R. L., and Bookchin, R. M. (1975). In: Levere R. D. (ed.),Sickle Cell Anemia and Other Hemoglobino-Pathies. Aca-demic Press, New York.
Nagel, R. L., Rieder, R. F., Bookchin, R. M., and James, G. W.(1973). Biochem. Biophys. Res. Comm. 53: 1240–1245.
Nagel, R. L., Johnson, J., Bookchin, R. M., Garel, M. C., Rosa, J.,Schiliro, G., Wajcman, H., Labie, D., Moo-Penn, W., andCastro, O. (1980). Nature 283: 832–834.
Padlan, E. A., and Love, W. E. (1985). J. Biol. Chem. 260: 8280–8291.
Paszty, C., Mohandas, N., Stevens, M. E., Loring, J. F., Liebhaber,S. A., Brion, C. M., and Rubin, E. M. (1995). Nature Genetics11: 33–39.
Pawliuk, R., Westerman, K. A., Fabry, M. E., Payen, E., Tighe, R.,Bouhassira, E. E., Acharya, S. A., Ellis, J., London, I. M.,Eaves, C. J., Humphries, R. K., Beuzard, Y., Nagel, R. L., andLeboulch, P. (2001). Science 294: 2368–2371.
Rao, M. J., Malavalli, A., Manjula, B. N., Kumar, R., Prabhak-aran, M., Sun, D. P., Ho, N. T., Ho, C., Nagel, R. L., andAcharya, A. S. (2000). J. Mol. Biol. 300: 1389–1406.
Rao, M. J., Iyer, K. S., and Acharya, A. S. (1995). J. Biol. Chem.270: 19250–19255.
Rao, M. J., and Acharya, A. S. (1994).Methods Enzymol. 231: 246–267.
Roy, R. P., Nagel, R. L., and Acharya, A. S. (1993). J. Biol. Chem.268: 16406–16412.
Sivaram, M. V., Sudha, R., and Roy, R. P. (2001). J. Biol. Chem.276: 18209–18215.
Srinivasulu, S., Malavalli, A., Prabhakaran, M., Nagel, R. L, andAcharya, A. S. (1999). Protein Eng. 12: 1105–1111.
Srinivasulu, S., Manjula, B. N., Nagel, R. L., Tsai, C. H., Ho, C.,Prabhakaran, M., and Acharya, S. A. (2004). Protein. Sci. 13:1266–1275.
Steinberg, M. H. (2003) Curr. Hematol. Rep. 2: 95–101.Witkowska, H. E., Lubin, B. H., Beuzard, Y., Baruchel, S.,
Esseltine, D.W., Vichinsky, E. P., Kleman, K. M., Bardakdj-ian-Michau, J., Pinkoski, L., and Cahn, S. (1991).New Engl. J.Med. 325: 1150–1154.
Watowich, S. J., Gross, L. J., and Josephs, R. (1993). J. Struct.Biol. 111: 161–179.
Yang, B., Kirby, S., Lewis, J., Detloff, P. J., Maeda, N., andSmithies, O. (1995). Proc. Natl. Acad. Sci. (USA) 92: 11608–11612.
516 Srinivasulu et al.
Copyright © 2022 FDOKUMEN




















