
Pain and purinergic signaling 3 ( 2 0 1 0 ) 2 2 2 – 2 3 2
11
Review Pain and purinergic signaling Makoto Tsuda, Hidetoshi Tozaki-Saitoh, Kazuhide Inoue ⁎ Department of Molecular and System Pharmacology, Graduate School of Pharmaceutical Sciences, Kyushu University, 3-1-1 Maidashi, Higashi, Fukuoka 812-8582, Japan ARTICLE INFO ABSTRACT Article history: Accepted 11 November 2009 Available online 18 November 2009 A growing body of evidence indicates that extracellular nucleotides play important roles in the regulation of neuronal and glial functions in the nervous system through P2 purinoceptors. P2 purinoceptors are divided into two families, ionotropic receptors (P2X) and metabotropic receptors (P2Y). P2X receptors (seven types; P2X1–P2X7) contain intrinsic pores that open by binding with ATP, and P2Y receptors (eight types; P2Y1, 2, 4, 6, 11, 12, 13 and 14) are activated by nucleotides and couple to intracellular second-messenger systems through heterotrimeric G-proteins. Nucleotides are released or leaked from non-excitable cells as well as neurons in physiological and pathophysiological conditions. Studies have shown that microglia, a type of glial cells known as resident macrophages in the CNS, express several subtypes of P2X and P2Y receptors, and these receptors play a key role in pain signaling in the spinal cord under pathological conditions such as by peripheral nerve injury (called neuropathic pain). Within the spinal dorsal horn, peripheral nerve injury leads to a progressive series of changes in microglia including morphological hypertrophy of the cell body and proliferation, which are considered indicative of activation. These activated microglia upregulate expression of P2X/Y receptors (e.g., P2X4 and P2Y12). Importantly, pharmacological, molecular and genetic manipulations of the function or expression of these microglial molecules strongly suppress neuropathic pain. We expect that further investigation to determine how ATP signaling via P2X receptors participates in the pathogenesis of chronic pain will lead to a better understanding of the molecular mechanisms of pathological pain and provide clues for the development of new therapeutic drugs. © 2009 Elsevier B.V. All rights reserved. Keywords: Purinoceptor Neuropathic pain Microglia Spinal cord Contents 1. Introduction .......................................................... 223 2. Molecular structure and pharmacological profiles of purinergic receptors ........................ 224 2.1. P2X receptors ..................................................... 224 2.2. P2Y receptors ..................................................... 224 3. Purinergic signaling and neuropathic pain ......................................... 225 3.1. P2X4 receptor ..................................................... 225 3.2. P2X7 receptor ..................................................... 227 BRAIN RESEARCH REVIEWS 63 (2010) 222 – 232 ⁎ Corresponding author. Fax: +81 92 642 4729. E-mail address: [email protected] (K. Inoue). 0165-0173/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.brainresrev.2009.11.003 available at www.sciencedirect.com www.elsevier.com/locate/brainresrev
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Pain and purinergic signaling 3 ( 2 0 1 0 ) 2 2 2 – 2 3 2

B R A I N R E S E A R C H R E V I E W S 6 3 ( 2 0 1 0 ) 2 2 2 – 2 3 2
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra in res rev
Review
Pain and purinergic signaling
Makoto Tsuda, Hidetoshi Tozaki-Saitoh, Kazuhide Inoue⁎
Department of Molecular and System Pharmacology, Graduate School of Pharmaceutical Sciences, Kyushu University, 3-1-1 Maidashi,Higashi, Fukuoka 812-8582, Japan
A R T I C L E I N F O
⁎ Corresponding author. Fax: +81 92 642 4729.E-mail address: [email protected].
0165-0173/$ – see front matter © 2009 Elsevidoi:10.1016/j.brainresrev.2009.11.003
A B S T R A C T
Article history:Accepted 11 November 2009Available online 18 November 2009
A growing body of evidence indicates that extracellular nucleotides play important roles in theregulation of neuronal and glial functions in the nervous system through P2 purinoceptors. P2purinoceptors are divided into two families, ionotropic receptors (P2X) and metabotropicreceptors (P2Y). P2X receptors (seven types; P2X1–P2X7) contain intrinsic pores that open bybinding with ATP, and P2Y receptors (eight types; P2Y1, 2, 4, 6, 11, 12, 13 and 14) are activatedby nucleotides and couple to intracellular second-messenger systems through heterotrimericG-proteins. Nucleotides are released or leaked from non-excitable cells as well as neurons inphysiological and pathophysiological conditions. Studies have shown thatmicroglia, a type ofglial cells known as residentmacrophages in the CNS, express several subtypes of P2X and P2Yreceptors, and these receptors play a key role in pain signaling in the spinal cord underpathological conditions such as by peripheral nerve injury (called neuropathic pain). Withinthe spinal dorsal horn, peripheral nerve injury leads to a progressive series of changes inmicroglia including morphological hypertrophy of the cell body and proliferation, which areconsidered indicative of activation. These activated microglia upregulate expression of P2X/Yreceptors (e.g., P2X4 and P2Y12). Importantly, pharmacological, molecular and geneticmanipulations of the function or expression of these microglial molecules stronglysuppress neuropathic pain. We expect that further investigation to determine how ATPsignaling via P2X receptors participates in the pathogenesis of chronic painwill lead to a betterunderstanding of the molecular mechanisms of pathological pain and provide clues for thedevelopment of new therapeutic drugs.
© 2009 Elsevier B.V. All rights reserved.
Keywords:PurinoceptorNeuropathic painMicrogliaSpinal cord
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2232. Molecular structure and pharmacological profiles of purinergic receptors . . . . . . . . . . . . . . . . . . . . . . . . 224
2.1. P2X receptors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2242.2. P2Y receptors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 224
3. Purinergic signaling and neuropathic pain . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2253.1. P2X4 receptor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2253.2. P2X7 receptor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 227
jp (K. Inoue).
er B.V. All rights reserved.

223B R A I N R E S E A R C H R E V I E W S 6 3 ( 2 0 1 0 ) 2 2 2 – 2 3 2
3.3. P2Y6 receptor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2283.4. P2Y12 receptor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 228
4. Concluding remarks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 228Acknowledgments. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 229References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 229
1. Introduction
In 1972, Burnstock proposed new roles of nucleotides asneurotransmitters (Burnstock, 1972) even though it wasprimarily recognized that intracellular ATP is the source offree energy to maintain life and nucleotides are keymoleculeswithin cells. In 1993, the first receptors for nucleotides, calledP2 purinoceptors, were cloned (Lustig et al., 1993; Webb et al.,1993). Afterwards, numerous subtypes of these receptors werealso cloned, and subsequently scientists began to graduallyaccept the “purinergic nervous system”. Now P2 purinoceptorsare divided into two families, ionotropic receptors (P2X) andmetabotropic receptors (P2Y) (Abbracchio et al., 2006; Khakh etal., 2001; Ralevic and Burnstock, 1998). Accumulating evidenceindicates that nucleotides are released and leaked from non-excitable cells as well as neurons and are involved in cell-to-cell communication in physiological and pathophysiologicalconditions (Burnstock, 2008). We finally have learned that theplayground of nucleotides may expand throughout the entirebody. One of the most exciting findings is that glia cells makeup over 70% of the total cell population in the central nervoussystem (CNS). Glial cells are classified into astrocytes,oligodendrocytes and microglia. Astrocytes express manytypes of P2X/Y receptors and release ATP in response tovarious stimuli or even spontaneously, and communicatewith neurons at synapses, microglial cells and vascular wallsat capillaries (Abbracchio et al., 2006; Inoue et al., 2007).Microglia, known as resident macrophages of the CNS, alsoexpress many types of P2 receptors (Farber and Kettenmann,2006; Inoue et al., 2007; Inoue, 2008). Although it was longbelieved that glial cells in the nervous system were onlyphysical and nutrient supports for neurons, a growing body ofevidence has dramatically changed this classical view andindicates that neuron–glia interactions are a key componentto understand CNS functions. In addition to crucial roles innormal physiological conditions, glial cells also play impor-tant roles in pathophysiological conditions of the CNSincluding psychiatric disorders, physical trauma, and chronicpain (Miller, 2005).
Pain in response to noxious stimuli can act as an earlywarning device that alerts us to the presence of damagingstimuli. This normal ‘good’ pain usually goes away soon afterthe nociceptive stimulus is removed. In contrast, sometimespain persists for a long time. In general, chronic pain may becategorized into inflammatory pain and neuropathic pain.Inflammatory pain is the result from inflammatory responsesto trauma in peripheral tissues. This type of pain, however,still has a physiological significance in that it may assistwound repair since contact with the damaged area isminimized. Also, inflammatory pain usually goes away afterthe tissue damage is repaired and can be generally managedby treatment with known analgesics. However, neuropathic
pain does not go away even though the initial damage hashealed. It typically occurs after nerve damage that can beinduced by bone compression in cancer, diabetes, infection,autoimmune disease or physical injury (Baron, 2006). Inaddition to spontaneous pain and hyperalgesia (the increasedpain perception of noxious stimuli), a troublesome symptomof neuropathic pain is pain hypersensitivity to normallyinnocuous stimuli, a phenomenon known as tactile allodynia.This type of pain is refractory to currently available treat-ments, such as non-steroidal anti-inflammatory drugs andopioids (Scholz and Woolf, 2002; Woolf and Mannion, 1999).We are now beginning to understand that neuropathic pain isnot just a symptom of disease but is a consequence ofdisordered functioning of the nervous system (Scholz andWoolf, 2002; Woolf and Mannion, 1999; Woolf and Salter,2000). Unraveling the mechanisms of pain hypersensitivitycaused by nerve damage is therefore essential for thedevelopment of new therapeutic drugs for neuropathic pain.
Accumulating evidence from diverse animal models ofneuropathic pain suggests that neuropathic pain mightinvolve aberrant excitability of the nervous system. Notably,in primary sensory ganglia and in the dorsal horn of the spinalcord multiple functional and anatomical alterations followperipheral nerve injury (Costigan et al., 2009; Scholz andWoolf, 2002; Woolf and Salter, 2000). While it has long beenconsidered that there are relevant changes in neurons,emerging lines of evidence have revealed that they alsooccur in glial cells, especially microglia (Marchand et al.,2005; Scholz and Woolf, 2007; Suter et al., 2007; Tsuda et al.,2005; Watkins et al., 2001). After peripheral nerve injury,microglia in the normal state (traditionally called ‘resting’microglia) in the spinal dorsal horn are converted to anactivated state through a series of cellular and molecularchanges (Echeverry et al., 2008; Suter et al., 2007; Tsuda et al.,2005). Microglia change their phenotype to the activated formfollowing altered expression of multiple molecules, includingcell surface receptors, intracellular signaling molecules, anddiffusible factors. By responding to extracellular ligands,activated microglia can evoke various cellular responses,such as migration toward afflicted sites and secretion ofproinflammatory factors. Importantly, pharmacological, mo-lecular and genetic manipulations of the function or expres-sion of these microglial molecules strongly modulateneuropathic pain (Clark et al., 2007; Griffin et al., 2007; Jin etal., 2003; Obata et al., 2007; Tanga et al., 2005; Tsuda et al., 2003,2004; Zhuang et al., 2005) and hyperexcitability of dorsal hornneurons (Coull et al., 2005; Keller et al., 2007). A growing bodyof evidence indicates that purinergic receptors are expressedin activated microglia under such pain conditions and thatP2X and P2Y receptors-mediated signaling critically contri-butes to the development and maintenance of neuropathicpain. In this article, we focus on recent developments that

224 B R A I N R E S E A R C H R E V I E W S 6 3 ( 2 0 1 0 ) 2 2 2 – 2 3 2
further the understanding of mechanisms by which P2X andP2Y receptors onmicroglia in the spinal cord participate in thepathogenesis of neuropathic pain. In addition, we summarizethe accumulated evidence from immunohistochemical, bio-chemical, molecular and behavioral studies, and we alsoprovide a framework to address the major questions thatcurrently remain unanswered.
2. Molecular structure and pharmacologicalprofiles of purinergic receptors
2.1. P2X receptors
P2X receptors belong to a family of ligand-gated ion-channelsand are cation-selective channels with almost equal perme-ability to Na+ and K+, and significant permeability to Ca2+
(Khakh et al., 2001; North, 2002; Ralevic and Burnstock, 1998).Molecular cloning has so far identified seven genes encodingP2X receptor subunits (P2X1–P2X7). All P2X subunits possesstwo transmembrane regions and have intracellular N- and C-terminals and a long extracellular loop between transmem-brane domains (Khakh et al., 2001; North, 2002; Vial et al.,2004). One third of the amino acids in the extracellular loop areconserved in at least six P2X subunits, suggesting they areinvolved in ATP binding (Vial et al., 2004). While theintracellular N-terminal regions have relatively similarlengths of amino acids, the length of the C-terminals divergesconsiderably from 30 residues in P2X6 to 240 in P2X7 (North,2002; Vial et al., 2004). In addition, there is a putative motif forphosphorylation by protein kinase C and A in the N- and C-terminals, respectively (North, 2002), and by ecto-proteinkinase C at the ectodomain (Wirkner et al., 2005).
One functional P2X receptor channel is presumably formedby a number of P2X subunits, as is considered to be the case inother ligand-gated ion channels. The number of subunits of aP2X receptor has been proposed to be three (Jiang et al., 2003;Nicke et al., 1998, 2003; Stoop et al., 1999) or four (Ding andSachs,2000; Kim et al., 1997). Recently, we analyzed the architectureand ATP-induced structural changes in P2X4 receptors usingfast-scanning atomic force microscopy (AFM) and revealed atrimer structure (Shinozaki et al., 2009). The trimer structure ofP2X receptors is also demonstrated in P2X2 receptors (Barrera etal., 2005). The trimer structure of P2X channels is stronglysupported by a recent study revealing the three-dimensionalcrystal structure of P2X4 receptor at a high resolution (Kawate etal., 2009). It appears that all three subunits are either identical,making P2X4 receptors homomeric, rather than identical, orheteromeric receptors. Biochemical studies have shown the co-assembly of P2X1/5 (Torres et al., 1998), P2X2/3 (Lewis et al.,1995), P2X2/6 (King et al., 2000), P2X4/6heteromeric receptors (Leet al., 1998). In addition, there is a report that P2X2/3 receptorsare likely to contain oneP2X2and twoP2X3subunits (Jianget al.,2003; Wilkinson et al., 2006).
Each of the seven homomeric P2X receptors and at least fourheteromeric receptors show different (but partly overlapping)electrophysiological and pharmacological properties in terms ofcurrent kinetics, desensitization rates and sensitivities toagonists and antagonists (Jarvis and Khakh, 2009; Khakh et al.,2001; North, 2002; Ralevic and Burnstock, 1998). When each one
or twoof themareexpressedheterologously in cells, ATPevokesa rapid- or slow-inactivating inward current. The former is seenincells expressingeither P2X1or P2X3 receptors, and the latter isseen in those expressing all other receptors including the fourheteromeric receptors (Khakh et al., 2001; North, 2002). Further-more, repetitive activation of P2X1 and P2X3 receptors by ATPcausesamarkeddesensitizationofATP-induced responses.α,β-Methylene ATP (αβmeATP), an analogue of ATP, is a usefulagonist for basic identification of P2X receptors containing P2X1or P2X3 (i.e., P2X1, P2X3, P2X1/5 and P2X2/3), although αβmeATPalso activatesP2X4/6 (Khakhetal., 2001;North, 2002; Ralevic andBurnstock, 1998).As for antagonists, suraminandpyridoxalpho-sphate-6-azophenyl-2′,4′-disulfonic acid (PPADS) are antago-nistic to almost all P2X receptors except rat P2X4 receptors (Buellet al., 1996). However, it should be noted that suramin alsoblocks several neurotransmitter receptors (e.g., NMDA andGABAA receptors) (Nakazawa et al., 1995; Peoples and Li, 1998).2′,3′-O-(2,4,6-trinitrophenyl)-ATP (TNP-ATP) at a nanomolarrange selectively blocks P2X1, P2X3 and P2X2/3 receptors(Virginio et al., 1998) and at high concentration also blocksP2X4 receptors (Virginio et al., 1998). Furthermore, diinosinepentaphosphate (Ip5I) is a very potent antagonist to P2X1 (Kinget al., 1999) and P2X3 receptors but, interestingly, not to P2X2and P2X2/3 receptors (Dunn et al., 2000). In 2002, the firstselective antagonist for P2X3 (and P2X2/3), A-317491, wasdeveloped (Jarvis et al., 2002). A-317491 blocks the responsesmediated by P2X3 or P2X2/3 receptors in a competitive fashionwithout any effect on other receptors, enzymes or ion-channels(Jarvis et al., 2002). Furthermore, several selective antagonistsfor P2X7 receptors, A-740003 (Honore et al., 2006), A-438079(McGaraughty et al., 2007), A-804598 (Donnelly-Roberts et al.,2009) and AZ11645373 (Stokes et al., 2006), have been developedrecently. A-317491 and A-438079 are commercially available.
2.2. P2Y receptors
The first G protein-coupled P2Y receptors were cloned in 1993(Lustig et al., 1993; Webb et al., 1993), and there are currentlyeight accepted P2Y receptors: P2Y1, P2Y2, P2Y4, P2Y6, P2Y11,P2Y12, P2Y13, and P2Y14. The missing numbers represent eithernonmammalian orthologs or receptors that have some sequencehomology to P2Y receptors but for which there is no functionalevidence of responsiveness to nucleotides (Abbracchio et al.,2006). P2Y receptors are 308–377 amino acid proteins with amass of 41–53 kDa after glycosylation. The seven transmem-brane (TM) domain tertiary structure of P2Y receptors iscommon to other G protein-coupled receptors (Fig. 1). P2Y areactivated by purine or pyrimidine nucleotides or sugar-nucleo-tides with subtype-dependent heterotrimeric G-proteins(Abbracchio et al., 2006). Site-directed mutagenesis studieshave shown that some positively charged residues in TM3, 6and 7 are crucial for receptor activation by nucleotides (Jiang etal., 1997). Each P2Y receptor binds to a single heterotrimeric Gprotein (Gαq/11 for P2Y1, 2, 4, 6), although P2Y11 can couple toboth Gαq/11, and Gαs, whereas P2Y12 and P2Y13 couple to Gαi
and P2Y14 couples to Gαi/o (Abbracchio et al., 2006). In responseto nucleotide activation, recombinant P2Y receptors eitheractivate the phospholipase C (PLC)-IP3 pathway through Gαq/11,which in turn releases intracellular calcium and activatesprotein kinase C, or affect adenylyl cyclase and alter cAMP
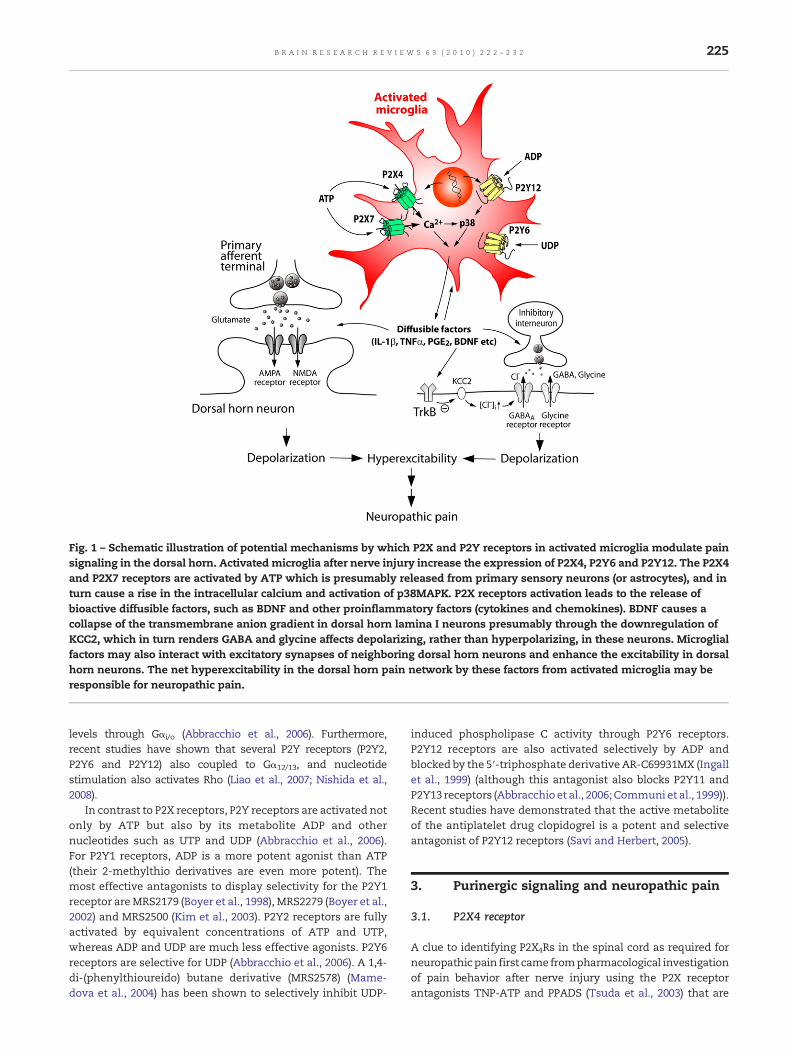
Fig. 1 – Schematic illustration of potential mechanisms by which P2X and P2Y receptors in activated microglia modulate painsignaling in the dorsal horn. Activated microglia after nerve injury increase the expression of P2X4, P2Y6 and P2Y12. The P2X4and P2X7 receptors are activated by ATP which is presumably released from primary sensory neurons (or astrocytes), and inturn cause a rise in the intracellular calcium and activation of p38MAPK. P2X receptors activation leads to the release ofbioactive diffusible factors, such as BDNF and other proinflammatory factors (cytokines and chemokines). BDNF causes acollapse of the transmembrane anion gradient in dorsal horn lamina I neurons presumably through the downregulation ofKCC2, which in turn renders GABA and glycine affects depolarizing, rather than hyperpolarizing, in these neurons. Microglialfactors may also interact with excitatory synapses of neighboring dorsal horn neurons and enhance the excitability in dorsalhorn neurons. The net hyperexcitability in the dorsal horn pain network by these factors from activated microglia may beresponsible for neuropathic pain.
225B R A I N R E S E A R C H R E V I E W S 6 3 ( 2 0 1 0 ) 2 2 2 – 2 3 2
levels through Gαi/o (Abbracchio et al., 2006). Furthermore,recent studies have shown that several P2Y receptors (P2Y2,P2Y6 and P2Y12) also coupled to Gα12/13, and nucleotidestimulation also activates Rho (Liao et al., 2007; Nishida et al.,2008).
In contrast to P2X receptors, P2Y receptors are activated notonly by ATP but also by its metabolite ADP and othernucleotides such as UTP and UDP (Abbracchio et al., 2006).For P2Y1 receptors, ADP is a more potent agonist than ATP(their 2-methylthio derivatives are even more potent). Themost effective antagonists to display selectivity for the P2Y1receptor areMRS2179 (Boyer et al., 1998), MRS2279 (Boyer et al.,2002) and MRS2500 (Kim et al., 2003). P2Y2 receptors are fullyactivated by equivalent concentrations of ATP and UTP,whereas ADP and UDP are much less effective agonists. P2Y6receptors are selective for UDP (Abbracchio et al., 2006). A 1,4-di-(phenylthioureido) butane derivative (MRS2578) (Mame-dova et al., 2004) has been shown to selectively inhibit UDP-
induced phospholipase C activity through P2Y6 receptors.P2Y12 receptors are also activated selectively by ADP andblocked by the 5′-triphosphate derivative AR-C69931MX (Ingallet al., 1999) (although this antagonist also blocks P2Y11 andP2Y13 receptors (Abbracchio et al., 2006; Communi et al., 1999)).Recent studies have demonstrated that the active metaboliteof the antiplatelet drug clopidogrel is a potent and selectiveantagonist of P2Y12 receptors (Savi and Herbert, 2005).
3. Purinergic signaling and neuropathic pain
3.1. P2X4 receptor
A clue to identifying P2X4Rs in the spinal cord as required forneuropathic pain first came frompharmacological investigationof pain behavior after nerve injury using the P2X receptorantagonists TNP-ATP and PPADS (Tsuda et al., 2003) that are

226 B R A I N R E S E A R C H R E V I E W S 6 3 ( 2 0 1 0 ) 2 2 2 – 2 3 2
known to be sensitive and insensitive to P2X4, respectively(Jarvis and Khakh, 2009; Khakh et al., 2001). Because nerveinjury-induced allodynia was effectively suppressed by TNP-ATP but not by PPADS, it was inferred that tactile allodyniadependsuponP2X4Rs in the spinal cord (Tsuda et al., 2003). Thishypothesis is substantially supported by recent findings show-ing that both mice treated spinally with a P2X4 antisenseoligonucleotide and mice lacking P2X4 show attenuated tactileallodynia after nerve injury (Tsuda et al., 2003, 2009; Ulmann etal., 2008). Expression of P2X4 receptors in the spinal cord, afternerve injury, markedly increased specifically in microglia,indicating that tonic activation of P2X4 receptors in microgliais necessary for sustaining allodynia (Tsuda et al., 2003).Moreover, it was found that spinal administration of P2X4-stimulated microglia caused otherwise normal rats to developallodynia. Therefore, P2X4 receptor activation inmicroglia is notonly necessary but is also sufficient to cause tactile allodynia(Tsuda et al., 2003).
The mechanisms by which microglia are crucial forproducing neuropathic pain must involve signaling fromactivated microglia to dorsal horn neurons, but how? Coullet al. (2005) demonstrated that P2X4-stimulated microgliainduced the hyperexcitability of dorsal horn neurons. In spinalcord slices of rats that had displayed allodynia after intrathe-cal administration of ATP-stimulated microglia, a positiveshift of the anion reversal potential (Eanion) in spinal lamina Ineurons was found, which caused GABA-evoked depolariza-tion, rather than hyperpolarization, in these neurons (Coull etal., 2005). Moreover, TNP-ATP acutely reversed the depolariz-ing shift in Eanion in lamina I neurons after nerve injury (Coullet al., 2005). These results imply that spinal microgliastimulated by P2X4 receptors cause neuropathic pain througha rise in intracellular Cl− in spinal lamina I neurons.Furthermore, Keller et al. (2007) have shown that local spinaladministration of ATP-stimulated microglia changed thephenotype of in vivo spinal lamina I output neurons, suchthat they relay innocuous mechanical input (Keller et al.,2007), which may account for allodynia. Recently, Coull et al.(2005) determined the role of brain-derived neurotrophicfactor (BDNF) as a signaling factor between microglia anddorsal horn lamina I neurons. It was found that intrathecalapplication of BDNF mimicked tactile allodynia and thedepolarizing shift in Eanion in lamina I neurons by peripheralnerve injury or by intrathecal administration of P2X4-stimu-lated microglia. Interference of signaling between BDNF andits receptor (TrkB) prevented tactile allodynia caused byperipheral nerve injury or by intrathecal administration ofP2X4-stimulated microglia. Activating P2X4 onmicroglial cellscaused the release of BDNF(Coull et al., 2005; Trang et al., 2009),an effect that is dependent on activation of p38 (Trang et al.,2009), a member of mitogen-activated protein kinase that hasbeen reported to be implicated in neuropathic pain (Jin et al.,2003; Tsuda et al., 2004). Interestingly, P2X4-mediated BDNFrelease was abolished by inhibiting SNARE (soluble N-ethylmaleimide-sensitive factor attachment protein receptor)-mediated exocytosis. Thus, these results indicate that P2X4-stimulated microglia release BDNF as a crucial factor to signalto lamina I neurons, causing a collapse of their transmem-brane anion gradient and subsequent neuronal hyperexcit-ability (Fig. 1). The γ-aminobutyric acidA (GABAA) receptor-
mediated depolarization might also produce excitationthrough voltage-sensitive Ca2+ channels and N-methyl-D-aspartate (NMDA) receptors. There is evidence that severalproinflammatory cytokines that are known to be released frommicroglia (Inoue, 2002; Inoue, 2006) modulate excitatorysynaptic transmission. Spinal injection of interleukin-1β (IL-1β) enhanced C-fiber-evoked responses and windup in wide-dynamic-range dorsal horn neurons (Reeve et al., 2000).Exogenous application of IL-1β enhanced NMDA receptor-mediated Ca2+ responses via activation of the tyrosine proteinkinase Src (Viviani et al., 2003). IL-1β was also reported todecrease GABAA receptor-mediated currents (Wang et al.,2000). A recent study has clearly demonstrated a powerfulrole for proinflammatory cytokines in excitatory or inhibitorysynaptic transmission and on neuronal activity in thesuperficial dorsal horn neurons (Ikeda et al., 2007; Kawasakiet al., 2008b). Also, inhibition of microglial ERK decreasedhyper-responsiveness of dorsal horn neurons in spinal cordinjured rats through a reduction of prostaglandin E2 produc-tion (Zhao et al., 2007). Thus, the net enhanced transmission inthe dorsal horn pain network by these factors might beresponsible for nerve injury-induced neuropathic pain.
From the above evidence, upregulation of P2X4 receptorexpression in microglia is therefore a key process in neuro-pathic pain. How peripheral nerve injury increases expressionof P2X4 receptors in microglia is not clearly understood.Activating both toll-like receptors (TLRs) and nucleotide-binding oligomerization domain 2 (NOD2, another pattern-recognition receptor) in cultured microglia increased theexpression of P2X4 at the mRNA level (Guo et al., 2006), butthe functional relevance of these receptors in vivo is stillunknown. Studies in our laboratory have recently shown thatthe extracellular matrix protein, fibronectin, increased ex-pression of mRNA and protein of P2X4 receptors in primarycultured microglial cells (Nasu-Tada et al., 2006; Tsuda et al.,2010). Cultured microglia stimulated by fibronectin alsoshowed an enhanced ATP-induced Ca2+ influx (Nasu-Tada etal., 2006). The level of fibronectin protein was elevated in thedorsal horn 3–7 days after nerve injury (Nasu-Tada et al., 2006),the time when P2X4 protein levels start to increase (Tsuda etal., 2003). Blockade of fibronectin receptors attenuated nerveinjury-induced P2X4 upregulation and allodynia (Tsuda et al.,2008a). It was also found that the upregulation of P2X4receptors in the spinal cord and allodynia after spinal nerveinjury was significantly suppressed by intrathecal adminis-tration of echistatin (Tsuda et al., 2008a). Furthermore,intrathecal delivery of fibronectin increased P2X4 expressionand produced allodynia, a behavior that was not evoked inP2X4-deficient mice. These results suggest that the fibronec-tin-integrin signaling system participates in the upregulationof P2X4 expression after nerve injury and subsequent neuro-pathic pain (Tsuda et al., 2008a).
We recently identified Lyn as a critical signalingmolecule forP2X4 upregulation in microglia (Tsuda et al., 2008b). Lyn-deficient microglial cells showed a deficit in increased P2X4expression caused by fibronectin. The level of Lyn expressionwas increased exclusively in microglia after nerve injury, andLyn-knockout mice exhibited a striking reduction in upregula-tion of P2X4 and tactile allodynia after nerve injury. Lyn tyrosinekinase distinctly activated the phosphatidylinositol 3-kinase

227B R A I N R E S E A R C H R E V I E W S 6 3 ( 2 0 1 0 ) 2 2 2 – 2 3 2
(PI3K)-Akt and mitogen-activated protein kinase kinase (MAPKkinase, MEK)-extracellular signal-regulated kinase (ERK) signal-ing cascades (Tsuda et al., 2010). Signaling through the PI3K-Aktpathway induceddegradationof p53 viamouse doubleminute 2(MDM2) in a proteasome-dependentmanner. The consequenceof an attenuated repressive effect of p53may be associatedwithenhanced P2X4 gene expression. On the other hand, activatedMEK-ERK signaling in microglia exposed to fibronectin en-hanced eukaryotic translation initiation factor 4E (eIF4E)phosphorylation status via activated MAPK-interacting proteinkinase-1 (MNK1), which may play a role in regulating P2X4expression at translational levels. Themolecular machinery forthe upregulation of P2X4 expression in microglia is nowconsidered to be as shown in Fig. 2.
There is evidence that P2X4 receptors inmicroglial cells arelocated predominantly within lysosomal compartments andare targeted there by their N- and C-terminal motifs (Qureshiet al., 2007). Notably, P2X4 receptors remain stable withinlysosomes and resist degradation by virtue of their N-linkedglycans. It is of particular interest that stimulating lysosome
Fig. 2 – Schematic representation of the hypotheticalmechanisms involved in fibronectin-induced P2X4upregulation in microglia. Extracellular fibronectin binds toα5β1 integrins on microglial cells, which leads to activationof the PI3K-Akt andMEK-ERK signaling cascades through Lyntyrosine kinase. Signaling through the PI3K-Akt pathwayinduces degradation of p53 via MDM2 in aproteasome-dependent manner. The reduction of therepressive effect of p53 may enhance P2X4 gene expression.Activated MEK-ERK signaling in microglia exposed tofibronectin enhances eIF4E phosphorylation status viaactivated MNK1, which may play a role in regulating P2X4expression at translational levels.
exocytosis, for example, using a Ca2+ ionophore, resulted inthe trafficking of P2X4 to the plasmamembrane (Qureshi et al.,2007). Given that P2X4 activity on the plasma membrane ofmicroglia is critical for microglial function, it is conceivablethat mechanisms underlying P2X4 recycling between thelysosome and the plasma membrane are important forneuropathic pain.
The above evidence indicates that P2X4 might be apotential therapeutic target to treat neuropathic pain. How-ever, there is no antagonist to strongly inhibit P2X4 receptors.We have recently demonstrated that some antidepressantsand anticonvulsants clinically used in patients with neuro-pathic pain have inhibitory effects on ATP-evoked Ca2+
response in cells expressing recombinant P2X4 receptors andprimary cultured microglial cells (Nagata et al., 2009). Amongthe drugs used, paroxetine has the most potent inhibitoryeffect. Intrathecal administration of paroxetine and fluvox-amine, but not citalopram, produced an anti-allodynic effectin an animal model of neuropathic pain, which correlatedwith the potency of inhibition of rat P2X4 receptors. Interest-ingly, it was found that the anti-allodynic effect of paroxetineis insensitive to 5-HT receptor antagonists and was alsoobserved in neuropathic pain animals with depleted 5-HT inthe spinal cord. These results suggest that the anti-allodyniceffect of antidepressants may be independent of the spinal 5-HT system and may be mediated by inhibiting P2X4 receptors(Nagata et al., 2009).
3.2. P2X7 receptor
The first evidence indicating the involvement of P2X7 receptorsin pain was provided by a study using P2X7-deficient mice.These mice exhibited reduced thermal and mechanical hyper-sensitivities after partial sciatic nerve ligation (Chessell et al.,2005). Recently, several groups have developed novel, selectiveinhibitors of P2X7 receptors and have examined their effects onneuropathic pain. Systemic (i.p.) administration of A-740003and A-438079, structurally unrelated P2X7 antagonists, reducedtactile allodynia in three different models of neuropathic painin rats (Honore et al., 2006). Furthermore, A-438079 (i.v.)reduced innocuous stimuli-evoked activity of in vivo dorsalhorn neurons in neuropathic rats, but not in sham-operatedrats (McGaraughty et al., 2007). The role of microglial P2X7receptors in neuropathic pain remains unclear because of alack of data showing the effect of intrathecal administration ofthese antagonists. Interestingly, recent evidence has indicateda structural and functional interaction between P2X7 and P2X4receptors (Casas-Pruneda et al., 2009; Guo et al., 2007). Studieshave shown co-immunoprecipitation of both receptor subtypes(Boumechache et al., 2009; Guo et al., 2007), but it might not bedue to the formation of heterotrimeric complex but to the closeassociation of neighboring P2X4 and P2X7 homotrimers(Boumechache et al., 2009; Nicke, 2008). Furthermore, incontrast to the predominant localization of P2X4 in intracellu-lar lysosome (as mentioned above), P2X7 is predominantlydistributed in the cell surface. If the trafficking of P2X4 to theplasma membrane is enhanced in activated microglia, afunctional interaction between these two receptors mightoccur. Therefore, it is of particular interest to investigate theinteraction between P2X4 and P2X7 in neuropathic pain.

228 B R A I N R E S E A R C H R E V I E W S 6 3 ( 2 0 1 0 ) 2 2 2 – 2 3 2
3.3. P2Y6 receptor
Microglial cells also express several G-protein-coupled recep-tors. In our recent study, it was found that microglial cells inculture express P2Y6 receptors (Koizumi et al., 2007) and thatafter peripheral nerve injury P2Y6 mRNA expression ismarkedly increased in the spinal cord, the time course ofwhich is parallel with that of tactile allodynia (unpublishedobservation). The function of P2Y6 in microglia remainsunknown, but we recently revealed that this receptor medi-ated microglial phagocytosis. It was found that applying UDP,an agonist for P2Y6, to primary cultured microglial cellsfacilitated phagocytosis (Koizumi et al., 2007). Neuronal injuryinduced by administration of kainic acid (KA) caused upregu-lation of P2Y6 receptors in microglia in the hippocampus. KA-evoked neuronal injury resulted in an increase in extracellularUTP, which was immediatelymetabolized into UDP in vivo andin vitro. UDP, leaked from injured neurons, caused P2Y6receptor-dependent phagocytosis. The P2Y6 receptor is upre-gulated when neurons are damaged, and could function as asensor for phagocytosis by sensing diffusible UDP signals.However, the role of upregulated P2Y6 in the spinal cord underneuropathic pain conditions remains unknown, and furtherstudies are required to determine its role.
3.4. P2Y12 receptor
Two recent studies have revealed that P2Y12 receptors are oneof the key regulators for neuropathic pain (Kobayashi et al.,2008; Tozaki-Saitoh et al., 2008). It was demonstrated thatintrathecally administered P2Y12 receptor antagonists, suchas MRS2395 and AR-C69931MX, or antisense oligonucleotidefor P2Y12 receptors significantly suppressed development ofneuropathic pain after spinal nerve injury (Tozaki-Saitoh etal., 2008) and partial sciatic nerve injury (Kobayashi et al.,2008). A crucial finding was established using P2Y12-knockoutmice. Genetic ablation of P2Y12 receptors failed to producetactile allodynia (Tozaki-Saitoh et al., 2008). Strategic advancesin targeting P2Y12 receptors were considered because of itsrestricted expression in the CNS. Several lines of evidencerevealed that P2Y12 receptor expression is specifically ob-served in brain and spinal cord resident microglia but is notobserved in fms- or CD11b-positive peripheralmacrophages inspleen (Haynes et al., 2006; Kobayashi et al., 2006; Pausch et al.,2004; Sasaki et al., 2003). In line with these facts, we couldconsider that P2Y12 receptor inhibition results from P2Y12receptor-gated signal modulation in the CNS. p38MAPK is asignalingmoleculewhose phosphorylation after partial sciaticnerve injury is almost completely suppressed by an antagonistor antisense oligonucleotide for P2Y12 receptors (Kobayashi etal., 2008). We have also shown that a single administration ofthe P2Y12 receptor antagonists AR-C69931MX (intrathecally)or clopidogrel (orally) to nerve-injured rats produced a strikingalleviation of established tactile allodynia (Tozaki-Saitoh etal., 2008). These effects by P2Y12 receptor antagonists are incontrast to the effects of another P2Y12 receptor antagonist,MRS2395, and a P2Y12 receptor antisense oligonucleotide,which did not reverse pain behaviors (Kobayashi et al., 2008).The exact reason for this difference remains open to question.In addition, it must be noted that P2Y12 receptors are
predominantly expressed in platelets in the periphery andinhibition of this receptor is one of the adopted mechanismsfor anti-platelet agents (Gachet, 2005).
Expression levels of P2Y12 receptors are dramaticallyincreased in microglial cells ipsilateral to peripheral nerveinjury in the spinal cord after nerve injury (Kobayashi et al.,2008; Tozaki-Saitoh et al., 2008) but down-modulated in brainmicroglia activated by slice preparation (Haynes et al., 2006). Amajor difference in each experiment was the proximal ordistal neuronal damage, which might induce different signalcascades to invoke microglial responses. Indeed, upregulationof P2Y12 receptor mRNA is also observed in a facial nerveinjury model (Sasaki et al., 2003). Detailed mechanisms ofexpressional regulation of P2Y12 receptors remain to beelucidated.
Many studies that have investigated in depth the mecha-nismof ATP-mediated chemotaxis inmicroglia have identifiedP2Y12 and P2X4 receptors as primary sensors for extracellularATP (Honda et al., 2001; Irino et al., 2008; Kanazawa et al., 2002;Ohsawa et al., 2007). Further studies have added evidenceindicating that P2Y12 receptors are implicated in the motilityof microglial cell bodies and processes in vivo or ex vivo(Davalos et al., 2005; Haynes et al., 2006; Kurpius et al., 2007).Thus, low ATP concentrations in the extracellular spaceactivate chemotaxis of microglia at the injured or inflamedsite. We already know that P2Y12 receptors are unlikely to benecessary to convert spinalmicroglia to the activated state andto increase their cell numbers because morphological andnumerical alterations of microglia were not influenced by thelack of P2Y12 receptors or by the administration of the P2Y12receptor antagonist AR-C69931MX in the spinal dorsal hornafter spinal nerve injury (Tozaki-Saitoh et al., 2008). One ofremaining possibilities regarding ATP-induced microglialchemotactic activity is that P2Y12 receptor activity inmicrogliamay influence the abilities of microglia to extend the tips oftheir branched processes toward neighboring pain transmis-sion neurons, which may in turn affect microglia–neuroncommunications.
4. Concluding remarks
We have primarily focused on the role of purinoceptorsexpressed in spinal microglia in neuropathic pain caused byperipheral nerve injury. Pharmacological, molecular andgenetic manipulations of the function or expression of thesepurinoceptors have influenced nerve injury-induced painbehaviors and hyperexcitability of the dorsal horn painpathway. Therefore, purinoceptor-mediated spinal microgliaactivity make a critical contribution to pathologically en-hanced pain processing in the dorsal horn, and microglialpurinoceptors might be promising targets for treating neuro-pathic pain. Recently, novel and established drugs that targetP2X and P2Y receptors have been reported to be effective inmodels of pain. Of note, a more recent study has revealed thethree-dimensional crystal structure of P2X4 receptor. Al-though the structure was solved in the absence of ATP,cavities between subunits thought to be the ATP binding sitewere also found. Thus, the P2X4 structure will be highlyinformative for designing new drugs targeting P2X4 receptor.

229B R A I N R E S E A R C H R E V I E W S 6 3 ( 2 0 1 0 ) 2 2 2 – 2 3 2
A predicted therapeutic benefit of interfering with microglialP2X and P2Y receptors may be that normal pain sensitivitywould be unaffected because expression or activity of most ofthese molecules are upregulated or enhanced predominantlyin activated microglia in the spinal cord where damagedsensory fibers project. In addition to microglia, recent studieshave also identified astrocyte-specific molecules and demon-strated a critical role of spinal astrocytes in neuropathic pain,particularly in the maintenance phase (Ji et al., 2006; Katsuraet al., 2008; Kawasaki et al., 2008a; Zhuang et al., 2006).Extracellular nucleotides are also a strong modulator ofastrocytes in the brain including the dorsal horn of the spinalcord (Fam et al., 2000; Salter and Hicks, 1994). How astrocyticpurinoceptors participate in neuropathic pain and interactwith microglial purinoceptors is now open for investigation. Itis expected that an increased understanding of the functionsof purinoceptors will provide us with exciting insights intopain mechanisms and clues to develop new therapeuticagents for the management of neuropathic pain.
Acknowledgments
M.T., H.T.-S. and K.I. are supported by a grant from theMinistry of Education, Culture, Sports, Science and Technol-ogy of Japan. M.T. acknowledges support from the MochidaMemorial Foundation for Medical and Pharmaceutical Re-search and the Astellas Foundation for Research on MetabolicDisorders.
R E F E R E N C E S
Abbracchio, M.P., Burnstock, G., Boeynaems, J.M., Barnard, E.A.,Boyer, J.L., Kennedy, C., Knight, G.E., Fumagalli, M., Gachet, C.,Jacobson, K.A., Weisman, G.A., 2006. International Union ofPharmacology LVIII: update on the P2Y G protein-couplednucleotide receptors: from molecular mechanisms andpathophysiology to therapy. Pharmacol. Rev. 58, 281–341.
Baron, R., 2006. Mechanisms of disease: neuropathic pain—aclinical perspective. Nat. Clin. Pract. Neurol. 2, 95–106.
Barrera, N.P., Ormond, S.J., Henderson, R.M., Murrell-Lagnado, R.D.,Edwardson, J.M., 2005. Atomic force microscopy imagingdemonstrates that P2X2 receptors are trimers but that P2X6receptor subunits do not oligomerize. J. Biol. Chem. 280,10759–10765.
Boumechache, M., Masin, M., Edwardson, J.M., Gorecki, D.C.,Murrell-Lagnado, R., 2009. Analysis of assembly and traffickingof native P2X4 and P2X7 receptor complexes in rodent immunecells. J. Biol. Chem. 284, 13446–13454.
Boyer, J.L., Mohanram, A., Camaioni, E., Jacobson, K.A., Harden,T.K., 1998. Competitive and selective antagonism of P2Y1receptors by N6-methyl 2′-deoxyadenosine 3′,5′-bisphosphate.Br. J. Pharmacol. 124, 1–3.
Boyer, J.L., Adams, M., Ravi, R.G., Jacobson, K.A., Harden, T.K., 2002.2-Chloro N(6)-methyl-(N)-methanocarba-2′-deoxyadenosine-3′,5′-bisphosphate is a selective high affinity P2Y(1) receptor antagonist.Br. J. Pharmacol. 135, 2004–2010.
Buell, G., Lewis, C., Collo, G., North, R.A., Surprenant, A., 1996. Anantagonist-insensitive P2X receptor expressed in epithelia andbrain. EMBO J. 15, 55–62.
Burnstock, G., 1972. Purinergic nerves. Pharmacol. Rev. 24,509–581.
Burnstock, G., 2008. Purinergic signalling and disorders of thecentral nervous system. Nat. Rev., Drug Discov. 7, 575–590.
Casas-Pruneda, G., Reyes, J.P., Perez-Flores, G., Perez-Cornejo, P.,Arreola, J., 2009. Functional interactions between P2X4 andP2X7 receptors from mouse salivary epithelia. J. Physiol. 587,2887–2901.
Chessell, I.P., Hatcher, J.P., Bountra, C., Michel, A.D., Hughes, J.P.,Green, P., Egerton, J., Murfin, M., Richardson, J., Peck, W.L.,Grahames, C.B., Casula, M.A., Yiangou, Y., Birch, R., Anand, P.,Buell, G.N., 2005. Disruption of the P2X7 purinoceptor geneabolishes chronic inflammatory and neuropathic pain. Pain114, 386–396.
Clark, A.K., Gentry, C., Bradbury, E.J., McMahon, S.B., Malcangio,M., 2007. Role of spinal microglia in rat models of peripheralnerve injury and inflammation. Eur. J. Pain 11, 223–230.
Communi, D., Robaye, B., Boeynaems, J.M., 1999. Pharmacologicalcharacterization of the human P2Y11 receptor. Br. J. Pharmacol.128, 1199–1206.
Costigan, M., Scholz, J., Woolf, C.J., 2009. Neuropathic pain: amaladaptive response of the nervous system to damage. Annu.Rev. Neurosci. 32, 1–32.
Coull, J.A., Beggs, S., Boudreau, D., Boivin, D., Tsuda, M., Inoue, K.,Gravel, C., Salter, M.W., De Koninck, Y., 2005. BDNF frommicroglia causes the shift in neuronal anion gradientunderlying neuropathic pain. Nature 438, 1017–1021.
Davalos, D., Grutzendler, J., Yang, G., Kim, J.V., Zuo, Y., Jung, S.,Littman, D.R., Dustin, M.L., Gan,W.B., 2005. ATPmediates rapidmicroglial response to local brain injury in vivo. Nat. Neurosci.8, 752–758.
Ding, S., Sachs, F., 2000. Inactivation of P2X2 purinoceptors bydivalent cations. J. Physiol. 522 (Pt 2), 199–214.
Donnelly-Roberts, D.L., Namovic, M.T., Surber, B., Vaidyanathan,S.X., Perez-Medrano, A., Wang, Y., Carroll, W.A., Jarvis, M.F.,2009. [3H]A-804598 ([3H]2-cyano-1-[(1S)-1-phenylethyl]-3-quinolin-5-ylguanidine) is a novel, potent, and selectiveantagonist radioligand for P2X7 receptors. Neuropharmacology56, 223–229.
Dunn, P.M., Liu, M., Zhong, Y., King, B.F., Burnstock, G., 2000.Diinosine pentaphosphate: an antagonist which discriminatesbetween recombinant P2X(3) and P2X(2/3) receptors andbetween two P2X receptors in rat sensory neurones. Br. J.Pharmacol. 130, 1378–1384.
Echeverry, S., Shi, X.Q., Zhang, J., 2008. Characterization of cellproliferation in rat spinal cord following peripheral nerveinjury and the relationship with neuropathic pain. Pain 135,37–47.
Fam, S.R., Gallagher, C.J., Salter, M.W., 2000. P2Y(1)purinoceptor-mediated Ca(2+) signaling and Ca(2+) wavepropagation in dorsal spinal cord astrocytes. J. Neurosci. 20,2800–2808.
Farber, K., Kettenmann, H., 2006. Purinergic signaling andmicroglia. Pflugers. Arch. 452, 615–621.
Gachet, C., 2005. The platelet P2 receptors as molecular targets forold and new antiplatelet drugs. Pharmacol. Ther. 108, 180–192.
Griffin, R.S., Costigan, M., Brenner, G.J., Ma, C.H., Scholz, J., Moss,A., Allchorne, A.J., Stahl, G.L., Woolf, C.J., 2007. Complementinduction in spinal cord microglia results in anaphylatoxinC5a-mediated pain hypersensitivity. J. Neurosci. 27, 8699–8708.
Guo, L.H., Guo, K.T., Wendel, H.P., Schluesener, H.J., 2006.Combinations of TLR and NOD2 ligands stimulate ratmicroglial P2X4R expression. Biochem. Biophys. Res. Commun.349, 1156–1162.
Guo, C., Masin, M., Qureshi, O.S., Murrell-Lagnado, R.D., 2007.Evidence for functional P2X4/P2X7 heteromeric receptors. Mol.Pharmacol. 72, 1447–1456.
Haynes, S.E., Hollopeter, G., Yang, G., Kurpius, D., Dailey, M.E., Gan,W.B., Julius, D., 2006. The P2Y(12) receptor regulates microglialactivation by extracellular nucleotides. Nat. Neurosci. 9,1512–1519.

230 B R A I N R E S E A R C H R E V I E W S 6 3 ( 2 0 1 0 ) 2 2 2 – 2 3 2
Honda, S., Sasaki, Y., Ohsawa, K., Imai, Y., Nakamura, Y., Inoue, K.,Kohsaka, S., 2001. Extracellular ATP or ADP induce chemotaxisof cultured microglia through Gi/o-coupled P2Y receptors. J.Neurosci. 21, 1975–1982.
Honore, P., Donnelly-Roberts, D., Namovic, M.T., Hsieh, G., Zhu,C.Z., Mikusa, J.P., Hernandez, G., Zhong, C., Gauvin, D.M.,Chandran, P., Harris, R., Medrano, A.P., Carroll, W., Marsh, K.,Sullivan, J.P., Faltynek, C.R., Jarvis, M.F., 2006. A-740003[N-(1-{[(cyanoimino)(5-quinolinylamino) methyl]amino}-2,2-dimethylpropyl)-2-(3,4-dimethoxyphenyl)acetamide], a noveland selective P2X7 receptor antagonist, dose-dependentlyreduces neuropathic pain in the rat. J. Pharmacol. Exp. Ther. 319,1376–1385.
Ikeda, H., Tsuda, M., Inoue, K., Murase, K., 2007. Long-termpotentiation of neuronal excitation by neuron–glia interactionsin the rat spinal dorsal horn. Eur. J. Neurosci. 25, 1297–1306.
Ingall, A.H., Dixon, J., Bailey, A., Coombs, M.E., Cox, D., McInally, J.I.,Hunt, S.F., Kindon, N.D., Teobald, B.J., Willis, P.A., Humphries,R.G., Leff, P., Clegg, J.A., Smith, J.A., Tomlinson, W., 1999.Antagonists of the platelet P2T receptor: a novel approach toantithrombotic therapy. J. Med. Chem. 42, 213–220.
Inoue, K., 2002. Microglial activation by purines and pyrimidines.Glia 40, 156–163.
Inoue, K., 2006. The function of microglia through purinergicreceptors: neuropathic pain and cytokine release. Pharmacol.Ther. 109, 210–226.
Inoue, K., 2008. Purinergic systems in microglia. Cell. Mol. Life Sci.65, 3074–3080.
Inoue, K., Koizumi, S., Tsuda, M., 2007. The role of nucleotides inthe neuron–glia communication responsible for the brainfunctions. J. Neurochem. 102, 1447–1458.
Irino, Y., Nakamura, Y., Inoue, K., Kohsaka, S., Ohsawa, K., 2008.Akt activation is involved in P2Y12 receptor-mediatedchemotaxis of microglia. J. Neurosci. Res. 86, 1511–1519.
Jarvis, M.F., Burgard, E.C., McGaraughty, S., Honore, P., Lynch, K.,Brennan, T.J., Subieta, A., Van Biesen, T., Cartmell, J., Bianchi,B., Niforatos, W., Kage, K., Yu, H., Mikusa, J., Wismer, C.T., Zhu,C.Z., Chu, K., Lee, C.H., Stewart, A.O., Polakowski, J., Cox, B.F.,Kowaluk, E., Williams, M., Sullivan, J., Faltynek, C., 2002.A-317491, a novel potent and selective non-nucleotide antagonistof P2X3 and P2X2/3 receptors, reduces chronic inflammatory andneuropathic pain in the rat. Proc. Natl. Acad. Sci. U. S. A. 99,17179–17184.
Jarvis, M.F., Khakh, B.S., 2009. ATP-gated P2X cation-channels.Neuropharmacology 56, 208–215.
Ji, R.R., Kawasaki, Y., Zhuang, Z.Y., Wen, Y.R., Decosterd, I., 2006.Possible role of spinal astrocytes in maintaining chronic painsensitization: review of current evidence with focus on bFGF/JNK pathway. Neuron Glia Biol. 2, 259–269.
Jiang, Q., Guo, D., Lee, B.X., Van Rhee, A.M., Kim, Y.C., Nicholas,R.A., Schachter, J.B., Harden, T.K., Jacobson, K.A., 1997. Amutational analysis of residues essential for ligand recognitionat the human P2Y1 receptor. Mol. Pharmacol. 52, 499–507.
Jiang, L.H., Kim, M., Spelta, V., Bo, X., Surprenant, A., North, R.A.,2003. Subunit arrangement in P2X receptors. J. Neurosci. 23,8903–8910.
Jin, S.X., Zhuang, Z.Y., Woolf, C.J., Ji, R.R., 2003. p38mitogen-activated protein kinase is activated after a spinalnerve ligation in spinal cord microglia and dorsal root ganglionneurons and contributes to the generation of neuropathic pain.J. Neurosci. 23, 4017–4022.
Kanazawa, H., Ohsawa, K., Sasaki, Y., Kohsaka, S., Imai, Y., 2002.Macrophage/microglia-specific protein Iba1 enhancesmembrane ruffling and Rac activation via phospholipaseC-gamma-dependent pathway. J. Biol. Chem. 277,20026–20032.
Katsura, H., Obata, K., Miyoshi, K., Kondo, T., Yamanaka, H.,Kobayashi, K., Dai, Y., Fukuoka, T., Sakagami, M., Noguchi, K.,2008. Transforming growth factor-activated kinase 1 induced
in spinal astrocytes contributes tomechanical hypersensitivityafter nerve injury. Glia 56, 723–733.
Kawasaki, Y., Xu, Z.Z., Wang, X., Park, J.Y., Zhuang, Z.Y., Tan, P.H.,Gao, Y.J., Roy, K., Corfas, G., Lo, E.H., Ji, R.R., 2008a. Distinct rolesof matrix metalloproteases in the early- and late-phasedevelopment of neuropathic pain. Nat. Med. 14, 331–336.
Kawasaki, Y., Zhang, L., Cheng, J.K., Ji, R.R., 2008b. Cytokinemechanisms of central sensitization: distinct and overlappingrole of interleukin-1beta, interleukin-6, and tumor necrosisfactor-alpha in regulating synaptic and neuronal activity in thesuperficial spinal cord. J. Neurosci. 28, 5189–5194.
Kawate, T., Michel, J.C., Birdsong, W.T., Gouaux, E., 2009. Crystalstructure of the ATP-gated P2X(4) ion channel in the closedstate. Nature 460, 592–598.
Keller, A.F., Beggs, S., Salter, M.W., De Koninck, Y., 2007.Transformation of the output of spinal lamina I neurons afternerve injury andmicroglia stimulation underlying neuropathicpain. Mol. Pain. 3, 27.
Khakh, B.S., Burnstock, G., Kennedy, C., King, B.F., North, R.A.,Seguela, P., Voigt, M., Humphrey, P.P., 2001. InternationalUnion of Pharmacology XXIV. Current status of thenomenclature and properties of P2X receptors and theirsubunits. Pharmacol. Rev. 53, 107–118.
Kim, M., Yoo, O.J., Choe, S., 1997. Molecular assembly of theextracellular domain of P2X2, an ATP-gated ion channel.Biochem. Biophys. Res. Commun. 240, 618–622.
Kim, H.S., Ohno, M., Xu, B., Kim, H.O., Choi, Y., Ji, X.D., Maddileti, S.,Marquez, V.E., Harden, T.K., Jacobson, K.A., 2003. 2-Substitutionof adenine nucleotide analogues containing a bicyclo[3.1.0]hexane ring system locked in a northern conformation:enhanced potency as P2Y1 receptor antagonists. J. Med. Chem.46, 4974–4987.
King, B.F., Liu, M., Pintor, J., Gualix, J., Miras-Portugal, M.T.,Burnstock, G., 1999. Diinosine pentaphosphate (IP5I) is a potentantagonist at recombinant rat P2X1 receptors. Br. J. Pharmacol.128, 981–988.
King, B.F., Townsend-Nicholson, A., Wildman, S.S., Thomas, T.,Spyer, K.M., Burnstock, G., 2000. Coexpression of rat P2X2 andP2X6 subunits in Xenopus oocytes. J. Neurosci. 20, 4871–4877.
Kobayashi, K., Fukuoka, T., Yamanaka, H., Dai, Y., Obata, K.,Tokunaga, A., Noguchi, K., 2006. Neurons and glial cellsdifferentially express P2Y receptor mRNAs in the rat dorsalroot ganglion and spinal cord. J. Comp. Neurol. 498, 443–454.
Kobayashi, K., Yamanaka, H., Fukuoka, T., Dai, Y., Obata, K.,Noguchi, K., 2008. P2Y12 receptor upregulation in activatedmicroglia is a gateway of p38 signaling and neuropathic pain.J. Neurosci. 28, 2892–2902.
Koizumi, S., Shigemoto-Mogami, Y., Nasu-Tada, K., Shinozaki, Y.,Ohsawa, K., Tsuda, M., Joshi, B.V., Jacobson, K.A., Kohsaka, S.,Inoue, K., 2007. UDP acting at P2Y6 receptors is a mediator ofmicroglial phagocytosis. Nature 446, 1091–1095.
Kurpius, D., Nolley, E.P., Dailey, M.E., 2007. Purines induce directedmigration and rapid homing of microglia to injured pyramidalneurons in developing hippocampus. Glia 55, 873–884.
Le, K.T., Babinski, K., Seguela, P., 1998. Central P2X4 and P2X6channel subunits coassemble into a novel heteromeric ATPreceptor. J. Neurosci. 18, 7152–7159.
Lewis, C., Neidhart, S., Holy, C., North, R.A., Buell, G., Surprenant,A., 1995. Coexpression of P2X2 and P2X3 receptor subunits canaccount for ATP-gated currents in sensory neurons. Nature377, 432–435.
Liao, Z., Seye, C.I., Weisman, G.A., Erb, L., 2007. The P2Y2nucleotide receptor requires interaction with alpha v integrinsto access and activate G12. J. Cell. Sci. 120, 1654–1662.
Lustig, K.D., Shiau, A.K., Brake, A.J., Julius, D., 1993. Expressioncloning of an ATP receptor from mouse neuroblastoma cells.Proc. Natl. Acad. Sci. U. S. A. 90, 5113–5117.
Mamedova, L.K., Joshi, B.V., Gao, Z.G., von Kugelgen, I., Jacobson,K.A., 2004. Diisothiocyanate derivatives as potent,

231B R A I N R E S E A R C H R E V I E W S 6 3 ( 2 0 1 0 ) 2 2 2 – 2 3 2
insurmountable antagonists of P2Y6 nucleotide receptors.Biochem. Pharmacol. 67, 1763–1770.
Marchand, F., Perretti, M., McMahon, S.B., 2005. Role of theimmune system in chronic pain. Nat. Rev., Neurosci. 6,521–532.
McGaraughty, S., Chu, K.L., Namovic, M.T., Donnelly-Roberts, D.L.,Harris, R.R., Zhang, X.F., Shieh, C.C., Wismer, C.T., Zhu, C.Z.,Gauvin, D.M., Fabiyi, A.C., Honore, P., Gregg, R.J., Kort, M.E.,Nelson, D.W., Carroll, W.A., Marsh, K., Faltynek, C.R., Jarvis,M.F., 2007. P2X7-related modulation of pathological nociceptionin rats. Neuroscience 146, 1817–1828.
Miller, G., 2005. Neuroscience. The dark side of glia. Science 308,778–781.
Nagata, K., Imai, T., Yamashita, T., Tsuda, M., Tozaki-Saitoh, H.,Inoue, K., 2009. Antidepressants inhibit P2X4 receptor function:a possible involvement in neuropathic pain relief. Mol. Pain 5,20.
Nakazawa, K., Inoue, K., Ito, K., Koizumi, S., 1995. Inhibition bysuramin and reactive blue 2 of GABA and glutamate receptorchannels in rat hippocampal neurons. Naunyn SchmiedebergsArch. Pharmacol. 351, 202–208.
Nasu-Tada, K., Koizumi, S., Tsuda, M., Kunifusa, E., Inoue, K., 2006.Possible involvement of increase in spinal fibronectinfollowing peripheral nerve injury in upregulation of microglialP2X(4), a key molecule for mechanical allodynia. Glia 53,769–775.
Nicke, A., Baumert, H.G., Rettinger, J., Eichele, A., Lambrecht, G.,Mutschler, E., Schmalzing, G., 1998. P2X1 and P2X3 receptorsform stable trimers: a novel structural motif of ligand-gated ionchannels. EMBO J. 17, 3016–3028.
Nicke, A., Rettinger, J., Schmalzing, G., 2003. Monomeric anddimeric byproducts are the principal functional elements ofhigher order P2X1 concatamers. Mol. Pharmacol. 63, 243–252.
Nicke, A., 2008. Homotrimeric complexes are the dominantassembly state of native P2X7 subunits. Biochem. Biophys. Res.Commun. 377, 803–808.
Nishida, M., Sato, Y., Uemura, A., Narita, Y., Tozaki-Saitoh, H.,Nakaya, M., Ide, T., Suzuki, K., Inoue, K., Nagao, T., Kurose, H.,2008. P2Y6 receptor-Galpha12/13 signalling in cardiomyocytestriggers pressure overload-induced cardiac fibrosis. EMBO J. 27,3104–3115.
North, R.A., 2002. Molecular physiology of P2X receptors. Physiol.Rev. 82, 1013–1067.
Obata, K., Katsura, H., Mizushima, T., Sakurai, J., Kobayashi, K.,Yamanaka, H., Dai, Y., Fukuoka, T., Noguchi, K., 2007. Roles ofextracellular signal-regulated protein kinases 5 in spinalmicroglia and primary sensory neurons for neuropathic pain. J.Neurochem. 102, 1569–1584.
Ohsawa, K., Irino, Y., Nakamura, Y., Akazawa, C., Inoue, K.,Kohsaka, S., 2007. Involvement of P2X(4) and P2Y(12) receptorsin ATP-induced microglial chemotaxis. Glia 55, 604–616.
Pausch, M.H., Lai, M., Tseng, E., Paulsen, J., Bates, B., Kwak, S., 2004.Functional expression of human andmouse P2Y12 receptors inSaccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 324,171–177.
Peoples, R.W., Li, C., 1998. Inhibition of NMDA-gated ion channelsby the P2 purinoceptor antagonists suramin and reactive blue 2in mouse hippocampal neurones. Br. J. Pharmacol. 124,400–408.
Qureshi, O.S., Paramasivam, A., Yu, J.C., Murrell-Lagnado, R.D.,2007. Regulation of P2X4 receptors by lysosomal targeting,glycan protection and exocytosis. J. Cell. Sci. 120, 3838–3849.
Ralevic, V., Burnstock, G., 1998. Receptors for purines andpyrimidines. Pharmacol. Rev. 50, 413–492.
Reeve, A.J., Patel, S., Fox, A., Walker, K., Urban, L., 2000.Intrathecally administered endotoxin or cytokines produceallodynia, hyperalgesia and changes in spinal cord neuronalresponses to nociceptive stimuli in the rat. Eur. J. Pain 4,247–257.
Salter, M.W., Hicks, J.L., 1994. ATP-evoked increases inintracellular calcium in neurons and glia from the dorsal spinalcord. J. Neurosci. 14, 1563–1575.
Sasaki, Y., Hoshi, M., Akazawa, C., Nakamura, Y., Tsuzuki, H.,Inoue, K., Kohsaka, S., 2003. Selective expression ofGi/o-coupled ATP receptor P2Y12 in microglia in rat brain.Glia 44,, 242–250.
Savi, P., Herbert, J.M., 2005. Clopidogrel and ticlopidine: P2Y12adenosine diphosphate-receptor antagonists for theprevention of atherothrombosis. Semin. Thromb. Hemost. 31,174–183.
Scholz, J., Woolf, C.J., 2002. Can we conquer pain? Nat. Neurosci. 5Suppl, 1062–1067.
Scholz, J., Woolf, C.J., 2007. The neuropathic pain triad: neurons,immune cells and glia. Nat. Neurosci. 10, 1361–1368.
Shinozaki, Y., Sumitomo, K., Tsuda, M., Koizumi, S., Inoue, K.,Torimitsu, K., 2009. Direct observation of ATP-inducedconformational changes in single P2X4 receptors. PLoS Biol.e103, 7.
Stokes, L., Jiang, L.H., Alcaraz, L., Bent, J., Bowers, K., Fagura, M.,Furber, M., Mortimore, M., Lawson, M., Theaker, J., Laurent, C.,Braddock, M., Surprenant, A., 2006. Characterization of aselective and potent antagonist of human P2X(7) receptors,AZ11645373. Br. J. Pharmacol. 149, 880–887.
Stoop, R., Thomas, S., Rassendren, F., Kawashima, E., Buell, G.,Surprenant, A., North, R.A., 1999. Contribution of individualsubunits to the multimeric P2X(2) receptor: estimates based onmethanethiosulfonate block at T336C. Mol. Pharmacol. 56,973–981.
Suter, M.R., Wen, Y.R., Decosterd, I., Ji, R.R., 2007. Do glial cellscontrol pain? Neuron Glia Biol. 3, 255–268.
Tanga, F.Y., Nutile-McMenemy, N., DeLeo, J.A., 2005. The CNS roleof Toll-like receptor 4 in innate neuroimmunity and painfulneuropathy. Proc. Natl. Acad. Sci. U. S. A. 102, 5856–5861.
Torres, G.E., Haines, W.R., Egan, T.M., Voigt, M.M., 1998.Co-expression of P2X1 and P2X5 receptor subunits reveals anovel ATP-gated ion channel. Mol. Pharmacol. 54, 989–993.
Tozaki-Saitoh, H., Tsuda, M., Miyata, H., Ueda, K., Kohsaka, S.,Inoue, K., 2008. P2Y12 receptors in spinalmicroglia are requiredfor neuropathic pain after peripheral nerve injury. J. Neurosci.28, 4949–4956.
Trang, T., Beggs, S., Wan, X., Salter, M.W., 2009.P2X4-receptor-mediated synthesis and release ofbrain-derived neurotrophic factor in microglia is dependent oncalcium and p38-mitogen-activated protein kinase activation.J. Neurosci. 29, 3518–3528.
Tsuda, M., Shigemoto-Mogami, Y., Koizumi, S., Mizokoshi, A.,Kohsaka, S., Salter, M.W., Inoue, K., 2003. P2X4 receptorsinduced in spinal microglia gate tactile allodynia after nerveinjury. Nature 424, 778–783.
Tsuda, M., Mizokoshi, A., Shigemoto-Mogami, Y., Koizumi, S.,Inoue, K., 2004. Activation of p38 mitogen-activated proteinkinase in spinal hyperactive microglia contributes to painhypersensitivity following peripheral nerve injury. Glia 45,89–95.
Tsuda, M., Inoue, K., Salter, M.W., 2005. Neuropathic pain andspinal microglia: a big problem from molecules in qsmallq glia.Trends Neurosci. 28, 101–107.
Tsuda, M., Toyomitsu, E., Komatsu, T., Masuda, T., Kunifusa, E.,Nasu-Tada, K., Koizumi, S., Yamamoto, K., Ando, J., Inoue, K.,2008a. Fibronectin/integrin system is involved in P2X(4)receptor upregulation in the spinal cord and neuropathic painafter nerve injury. Glia 56, 579–585.
Tsuda, M., Tozaki-Saitoh, H., Masuda, T., Toyomitsu, E., Tezuka, T.,Yamamoto, T., Inoue, K., 2008b. Lyn tyrosine kinase is requiredfor P2X(4) receptor upregulation and neuropathic pain afterperipheral nerve injury. Glia 56, 50–58.
Tsuda, M., Kuboyama, K., Inoue, T., Nagata, K., Tozaki-Saitoh, H.,Inoue, K., 2009. Behavioral phenotypes of mice lacking

232 B R A I N R E S E A R C H R E V I E W S 6 3 ( 2 0 1 0 ) 2 2 2 – 2 3 2
purinergic P2X4 receptors in acute and chronic pain assays.Mol. Pain 5, 28.
Tsuda, M., Toyomitsu, E., Kometani, M., Tozaki-Saitoh, H., Inoue,K., 2010. Mechanisms underlying fibronectin-inducedupregulation of P2X4R expression in microglia: distinct roles ofPI3K-Akt and MEK-ERK signaling pathways. J. Cell. Mol. Med.13, 3251–3259.
Ulmann, L., Hatcher, J.P., Hughes, J.P., Chaumont, S., Green, P.J.,Conquet, F., Buell, G.N., Reeve, A.J., Chessell, I.P., Rassendren,F., 2008. Up-regulation of P2X4 receptors in spinal microgliaafter peripheral nerve injury mediates BDNF release andneuropathic pain. J. Neurosci. 28, 11263–11268.
Vial, C., Roberts, J.A., Evans, R.J., 2004. Molecular properties ofATP-gated P2X receptor ion channels. Trends Pharmacol. Sci.25, 487–493.
Virginio, C., Robertson, G., Surprenant, A., North, R.A., 1998.Trinitrophenyl-substituted nucleotides are potent antagonistsselective for P2X1, P2X3, and heteromeric P2X2/3 receptors.Mol. Pharmacol. 53, 969–973.
Viviani, B., Bartesaghi, S., Gardoni, F., Vezzani, A., Behrens, M.M.,Bartfai, T., Binaglia, M., Corsini, E., Di Luca, M., Galli, C.L.,Marinovich, M., 2003. Interleukin-1beta enhances NMDAreceptor-mediated intracellular calcium increase throughactivation of the Src family of kinases. J. Neurosci. 23,8692–8700.
Wang, S., Cheng, Q., Malik, S., Yang, J., 2000. Interleukin-1betainhibits gamma-aminobutyric acid type A (GABA(A)) receptorcurrent in cultured hippocampal neurons. J. Pharmacol. Exp.Ther. 292, 497–504.
Watkins, L.R., Milligan, E.D., Maier, S.F., 2001. Glial activation: adriving force for pathological pain. Trends Neurosci. 24,450–455.
Webb, T.E., Simon, J., Krishek, B.J., Bateson, A.N., Smart, T.G., King,B.F., Burnstock, G., Barnard, E.A., 1993. Cloning and functionalexpression of a brain G-protein-coupled ATP receptor. FEBSLett. 324, 219–225.
Wilkinson, W.J., Jiang, L.H., Surprenant, A., North, R.A., 2006. Roleof ectodomain lysines in the subunits of the heteromeric P2X2/3 receptor. Mol. Pharmacol. 70, 1159–1163.
Wirkner, K., Stanchev, D., Koles, L., Klebingat, M., Dihazi, H.,Flehmig, G., Vial, C., Evans, R.J., Furst, S., Mager, P.P., Eschrich,K., Illes, P., 2005. Regulation of human recombinant P2X3receptors by ecto-protein kinase C. J. Neurosci. 25,7734–7742.
Woolf, C.J., Mannion, R.J., 1999. Neuropathic pain: aetiology,symptoms, mechanisms, and management. Lancet 353,1959–1964.
Woolf, C.J., Salter, M.W., 2000. Neuronal plasticity: increasing thegain in pain. Science 288, 1765–1769.
Zhao, P., Waxman, S.G., Hains, B.C., 2007. Extracellularsignal-regulated kinase-regulated microglia–neuron signalingby prostaglandin E2 contributes to pain after spinal cord injury.J. Neurosci. 27, 2357–2368.
Zhuang, Z.Y., Gerner, P., Woolf, C.J., Ji, R.R., 2005. ERK issequentially activated in neurons, microglia, and astrocytes byspinal nerve ligation and contributes to mechanical allodyniain this neuropathic pain model. Pain 114, 149–159.
Zhuang, Z.Y., Wen, Y.R., Zhang, D.R., Borsello, T., Bonny, C.,Strichartz, G.R., Decosterd, I., Ji, R.R., 2006. A peptide c-JunN-terminal kinase (JNK) inhibitor blocks mechanical allodyniaafter spinal nerve ligation: respective roles of JNK activationin primary sensory neurons and spinal astrocytes forneuropathic pain development and maintenance. J. Neurosci.26, 3551–3560.




















