
p38 MAPK is a critical regulator of the constitutive and theβ4 integrin-regulated expression of...
11
p38 MAPK is a critical regulator of the constitutive and the 4 integrin-regulated expression of IL-6 in human normal thymic epithelial cells Fabrizio Mainiero 1 , Michaela Colombara 3 , Valeria Antonini 3 , Raffaele Strippoli 1 , Marcello Merola 4 , Ornella Poffe 3 , Giuseppe Tridente 2,3 and Dunia Ramarli 2 1 Department of Experimental Medicine and Pathology, Institute Pasteur-Fondazione Cenci Bolognetti, University of Rome “La Sapienza”, Rome, Italy 2 Clinical Immunology, G. B. Rossi Hospital, Verona, Italy 3 Section of Immunology, Department of Pathology, University of Verona, Verona, Italy 4 Section of Biochemistry, Department of Neurological and Vision Science, University of Verona, Verona, Italy Cytokines and adhesion receptors are key mediators in the dialog occurring between thymic epithelial cells (TEC) and thymocytes and regulating T cell maturation and epithelial embry- onic differentiation. Among cytokines, IL-6 can be critical in the thymus, fostering prolifera- tion, differentiation and/or survival of both TEC and thymocytes. We have previously reported in human normal TEC that clustering of the laminin receptor 6 4 integrin induced by thymocyte contact or monoclonal antibody-mediated cross-linking regulates IL-6 gene expression via activation of NF- B and NF-IL6 transactivators. Here we show that 64 integrin activates p38 mitogen-activated protein kinase (MAPK) and that p38 is essential for IL-6 gene expression. In fact, 4 cross-linking activated p38 and extracellular signal- regulated kinase (ERK) MAPK, Rac1, p21-activated protein kinase 1 (PAK1) and MAPK kinases (MKK) 3/MKK6. However, pharmacological blockade of p38 or ERK demonstrated that p38 inhibition abrogated both basal and 4 integrin-induced production of IL-6 prevent- ing NF- B and NF-IL6 activation, whereas ERK inhibition reduced IL-6 production, hamper- ing only NF- B activation. Overall, our results indicate that p38 MAPK and 6 4 integrin, expressed by TEC throughout their life, are critical regulators of the intrathymic availability of a cytokine controlling fate and functions of cells governing development and maintenance of thymic architecture and immune responses. Key words: MAPK / Integrin / IL-6 / Thymus / Transactivators Received 11/2/03 Revised 7/8/03 Accepted 29/8/03 [DOI 10.1002/eji.200323931] Abbreviations: TEC: Thymic epithelial cells CREB: cAMP- responsive element-binding protein ERK: Extracellular signal-regulated kinase MAPK: Mitogen-activated protein kinase PAK: p21-activated protein kinase MKK: MAPK kinase SB: SB 202190 PD: PD 98059 GST: Glutathione S- transferase 1 Introduction Thymus is a specialized microenvironment in which hematopoietic precursors commit to T cell lineage and maturate while trafficking in the subcortical, cortical and medullary areas. Animal models and organ cultures have shown that thymic epithelial cells (TEC) and thymocytes influence each other while building the thymic architec- ture during embryogenesis and then remodel it during postnatal life. Key elements of their dialog are adhesion receptors interacting at the single-cell level (i.e. mem- bers of the Ig superfamily, integrins and cadherins) and cytokines, growth factors or neuropeptides secreted in the microenvironment [1–4]. Mainly produced by TEC, IL-6 has been implicated in development and aging of human or mouse thymus. In vitro observations have indicated that IL-6 fosters prolif- eration and differentiation of TEC and thymocytes, and protects thymocytes from apoptosis. Animal models have shown that signals from members of the gp130 family, including IL-6, are essential for the postnatal maintenance of thymic architecture. In vivo evidence in humans or mice has correlated the local increase of IL-6, leukemia inhibitory factor (LIF) and oncostatin with thy- mic atrophy during aging [3–5]. IL-6 gene expression in epithelial cells is mainly regu- lated at a transcriptional level by a major transactivator, 3038 F. Mainiero et al. Eur. J. Immunol. 2003. 33: 3038–3048 © 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
Transcript of p38 MAPK is a critical regulator of the constitutive and theβ4 integrin-regulated expression of...

p38 MAPK is a critical regulator of the constitutiveand the I 4 integrin-regulated expression of IL-6 inhuman normal thymic epithelial cells
Fabrizio Mainiero1, Michaela Colombara3, Valeria Antonini3, Raffaele Strippoli1,Marcello Merola4, Ornella Poffe3, Giuseppe Tridente2,3 and Dunia Ramarli2
1 Department of Experimental Medicine and Pathology, Institute Pasteur-Fondazione CenciBolognetti, University of Rome “La Sapienza”, Rome, Italy
2 Clinical Immunology, G. B. Rossi Hospital, Verona, Italy3 Section of Immunology, Department of Pathology, University of Verona, Verona, Italy4 Section of Biochemistry, Department of Neurological and Vision Science, University of Verona,
Verona, Italy
Cytokines and adhesion receptors are key mediators in the dialog occurring between thymicepithelial cells (TEC) and thymocytes and regulating T cell maturation and epithelial embry-onic differentiation. Among cytokines, IL-6 can be critical in the thymus, fostering prolifera-tion, differentiation and/or survival of both TEC and thymocytes. We have previouslyreported in human normal TEC that clustering of the laminin receptor § 6 g 4 integrin inducedby thymocyte contact or monoclonal antibody-mediated cross-linking regulates IL-6 geneexpression via activation of NF- ‹ B and NF-IL6 transactivators. Here we show that § 6 g 4integrin activates p38 mitogen-activated protein kinase (MAPK) and that p38 is essential forIL-6 gene expression. In fact, g 4 cross-linking activated p38 and extracellular signal-regulated kinase (ERK) MAPK, Rac1, p21-activated protein kinase 1 (PAK1) and MAPKkinases (MKK) 3/MKK6. However, pharmacological blockade of p38 or ERK demonstratedthat p38 inhibition abrogated both basal and g 4 integrin-induced production of IL-6 prevent-ing NF- ‹ B and NF-IL6 activation, whereas ERK inhibition reduced IL-6 production, hamper-ing only NF- ‹ B activation. Overall, our results indicate that p38 MAPK and § 6 g 4 integrin,expressed by TEC throughout their life, are critical regulators of the intrathymic availability ofa cytokine controlling fate and functions of cells governing development and maintenance ofthymic architecture and immune responses.
Key words: MAPK / Integrin / IL-6 / Thymus / Transactivators
Received 11/2/03Revised 7/8/03Accepted 29/8/03
[DOI 10.1002/eji.200323931]
Abbreviations: TEC: Thymic epithelial cells CREB: cAMP-responsive element-binding protein ERK: Extracellularsignal-regulated kinase MAPK: Mitogen-activated proteinkinase PAK: p21-activated protein kinase MKK: MAPKkinase SB: SB 202190 PD: PD 98059 GST: Glutathione S-transferase
1 Introduction
Thymus is a specialized microenvironment in whichhematopoietic precursors commit to T cell lineage andmaturate while trafficking in the subcortical, cortical andmedullary areas. Animal models and organ cultures haveshown that thymic epithelial cells (TEC) and thymocytesinfluence each other while building the thymic architec-ture during embryogenesis and then remodel it duringpostnatal life. Key elements of their dialog are adhesion
receptors interacting at the single-cell level (i.e. mem-bers of the Ig superfamily, integrins and cadherins) andcytokines, growth factors or neuropeptides secreted inthe microenvironment [1–4].
Mainly produced by TEC, IL-6 has been implicated indevelopment and aging of human or mouse thymus. Invitro observations have indicated that IL-6 fosters prolif-eration and differentiation of TEC and thymocytes, andprotects thymocytes from apoptosis. Animal modelshave shown that signals from members of the gp130family, including IL-6, are essential for the postnatalmaintenance of thymic architecture. In vivo evidence inhumans or mice has correlated the local increase of IL-6,leukemia inhibitory factor (LIF) and oncostatin with thy-mic atrophy during aging [3–5].
IL-6 gene expression in epithelial cells is mainly regu-lated at a transcriptional level by a major transactivator,
3038 F. Mainiero et al. Eur. J. Immunol. 2003. 33: 3038–3048
© 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim

NF- ‹ B, cooperating with transactivators of four otherfamilies [cAMP-responsive element-binding proteins(CREB), activator protein-1 (AP-1), NF-IL6, Sp1] [6–8].Ubiquitously expressed, NF- ‹ B heterodimer is com-posed of Rel family proteins (commonly p50/p65 sub-units) and holds inactive in the cytosol bound to inhibitorproteins (I ‹ B) masking its nuclear localization sequence.Activation occurs at different cell locations. Cytosolicphosphorylation and degradation of I ‹ B disrupts the NF-‹ B-I ‹ B triplex allowing NF- ‹ B to enter the nucleus [7].
Nuclear phosphorylation of p65 subunits increases NF-‹ B transactivation adding to interactions with the basal
transcription complex [7, 8]. Structural/functional associ-ations of NF- ‹ B with other transactivators such as NF-IL6 (also named C/EBP g , LAP, NF-M, IL-6-DBP, AGP/EBP) are critical in epithelial cells for full gene expression[7]. Like NF- ‹ B, NF-IL6 is ubiquitously expressed butfunctionally regulated at translational and post-translational levels. Alternative use of AUG codonswithin the same reading frame yields translation of iso-forms progressively truncated at the N terminus andhence impaired in transactivation. Site-specific serinephosphorylation in the proteodomains conditionsnuclear localization, DNA binding activity, and functionalinteraction of the various isoforms [7–9].
Previous and ours results have demonstrated that (1)TEC derived from normal thymus and grown to orga-nized monolayers, constitutively produce extracellularmatrix proteins (laminins, fibronectin and collagentype IV); (2) these cells display two sets of integrinspolarized at discrete locations ( § 2 g 1 and § 3 g 1 heterodi-mers, mostly located at the TEC-TEC boundaries, and§ 6 g 4, colocalized with its ligand laminin at the base of
the monolayer); and (3) thymocyte or lymphocyte adhe-sion remodels this pattern, so that § 6 g 4 and § 3 g 1, butnot § 2 g 1 integrins, cluster also around individual lym-phoid cells, recognizing yet unknown cellular ligandsexpressed by T cells [10–13].
We have shown that in these TEC § 6 g 4 or § 3 g 1 integrinsregulate IL-6 production through signals that, elicited bythymocyte/lymphocyte adhesion or mAb cross-linking,induce NF- ‹ B nuclear translocation and serine phos-phorylation of active NF-IL6 isoforms (43 and 36 kDa)[11–13]. As § 6 g 4 integrin evoked the greatest responses,we further investigated some of the molecular mecha-nisms involved in its activity.
§ 6 g 4, a well-defined adhesion receptor for various lami-nin isoforms, is a special integrin. Its g 4 subunit containsan unusually long cytodomain intersecting with path-ways regulated by growth factors, phosphoinositide 3-kinase or adaptor proteins of the insulin receptorsubstrate family [14, 15]. Moreover, it can recruit the
adaptor protein Shc, thus coupling § 6 g 4 to Ras andthereby to both Rac-c-Jun N-terminal kinase (JNK) andRaf-extracellular signal-regulated kinase (ERK) signalingcascades [16, 17]. To our knowledge no data are yetavailable on g 4 integrin ability to activate p38, the thirdfamily of mitogen-activated protein kinases (MAPK).
Here we show that in normal TEC g 4 integrin recruitmentby laminin or cross-linking with specific Ab activates p38,a component with ERK of a signaling pathway includingendogenous Rac1, p21-activated protein kinase (PAK) 1and MAPK kinases (MKK) 3/MKK6, leading to activationof NF- ‹ B and NF-IL6 transactivators and regulating IL-6production. Previous reports have shown that p38 orERK MAPK exclude each other in the control of tran-scription factors activation or of IL-6 gene expression[18–20]. We found that both p38 and ERK participate tothe transcriptional regulation of the basal and the g 4-induced expression of IL-6. However, p38 dominatedover ERK, partly regulating the DNA-binding activity ofNF- ‹ B along with the serine phosphorylation of NF-IL6.
2 Results
2.1 Experimental system
Experiments were performed with TEC derived fromindependent donors, at the same passage of culture(third to fourth) and deprived of epidermal growth factorand medium complements for 48 h. This treatment didnot change g 4 expression or TEC morphology (D.Ramarli, unpublished).
2.2 p38 and ERK are activated in TEC byI 4 integrin recruitment with laminin orspecific antibodies
To test whether signals from g 4 integrin induced p38and/or ERK activation, TEC were plated on dishescoated with laminin 5 or, as monolayers, subjected to g 4cross-linking. Phosphorylation of p38 and ERK wasexamined in TEC lysates by immunoblotting using Abagainst the active, phosphorylated form of both kinases.As shown in Fig. 1A, B, p38 and ERK were both basallyactivated at low levels in untreated TEC. Laminin 5 or g 4cross-linking strongly increased p38 and ERK phosphor-ylation. Upon g 4 cross-linking, p38 phosphorylationstarted at 1 min, peaked at 5–10 min and returned tobasal levels at 20 min after stimulation, whereas ERKphosphorylation was slightly delayed (Fig. 1B). Casein ormAb-mediated cross-linking of IFN § / g receptors(c mAb gam) used as controls failed to function (Fig. 1A,B). TEC secreted laminin in culture (Fig. 1C). This secre-
Eur. J. Immunol. 2003. 33: 3038–3048 p38 MAPK regulates IL-6 in human thymic epithelium 3039
© 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
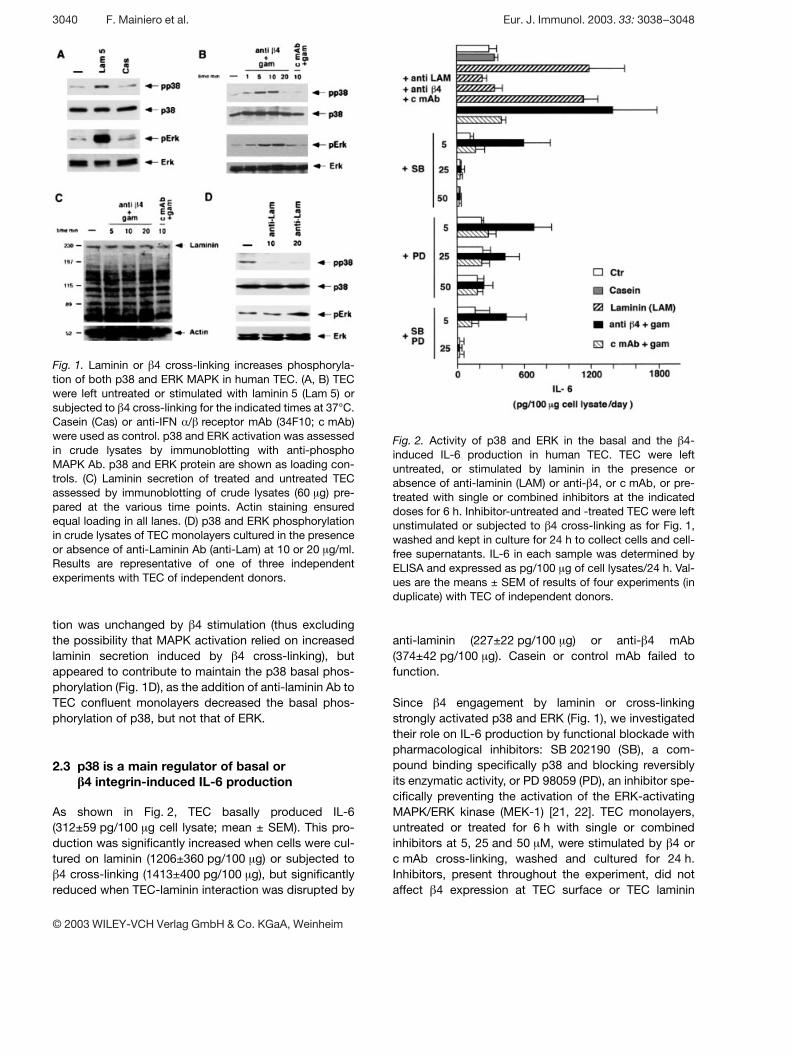
Fig. 1. Laminin or g 4 cross-linking increases phosphoryla-tion of both p38 and ERK MAPK in human TEC. (A, B) TECwere left untreated or stimulated with laminin 5 (Lam 5) orsubjected to g 4 cross-linking for the indicated times at 37°C.Casein (Cas) or anti-IFN § / g receptor mAb (34F10; c mAb)were used as control. p38 and ERK activation was assessedin crude lysates by immunoblotting with anti-phosphoMAPK Ab. p38 and ERK protein are shown as loading con-trols. (C) Laminin secretion of treated and untreated TECassessed by immunoblotting of crude lysates (60 ? g) pre-pared at the various time points. Actin staining ensuredequal loading in all lanes. (D) p38 and ERK phosphorylationin crude lysates of TEC monolayers cultured in the presenceor absence of anti-Laminin Ab (anti-Lam) at 10 or 20 ? g/ml.Results are representative of one of three independentexperiments with TEC of independent donors.
Fig. 2. Activity of p38 and ERK in the basal and the g 4-induced IL-6 production in human TEC. TEC were leftuntreated, or stimulated by laminin in the presence orabsence of anti-laminin (LAM) or anti- g 4, or c mAb, or pre-treated with single or combined inhibitors at the indicateddoses for 6 h. Inhibitor-untreated and -treated TEC were leftunstimulated or subjected to g 4 cross-linking as for Fig. 1,washed and kept in culture for 24 h to collect cells and cell-free supernatants. IL-6 in each sample was determined byELISA and expressed as pg/100 ? g of cell lysates/24 h. Val-ues are the means ± SEM of results of four experiments (induplicate) with TEC of independent donors.
tion was unchanged by g 4 stimulation (thus excludingthe possibility that MAPK activation relied on increasedlaminin secretion induced by g 4 cross-linking), butappeared to contribute to maintain the p38 basal phos-phorylation (Fig. 1D), as the addition of anti-laminin Ab toTEC confluent monolayers decreased the basal phos-phorylation of p38, but not that of ERK.
2.3 p38 is a main regulator of basal orI 4 integrin-induced IL-6 production
As shown in Fig. 2, TEC basally produced IL-6(312±59 pg/100 ? g cell lysate; mean ± SEM). This pro-duction was significantly increased when cells were cul-tured on laminin (1206±360 pg/100 ? g) or subjected tog 4 cross-linking (1413±400 pg/100 ? g), but significantly
reduced when TEC-laminin interaction was disrupted by
anti-laminin (227±22 pg/100 ? g) or anti- g 4 mAb(374±42 pg/100 ? g). Casein or control mAb failed tofunction.
Since g 4 engagement by laminin or cross-linkingstrongly activated p38 and ERK (Fig. 1), we investigatedtheir role on IL-6 production by functional blockade withpharmacological inhibitors: SB 202190 (SB), a com-pound binding specifically p38 and blocking reversiblyits enzymatic activity, or PD 98059 (PD), an inhibitor spe-cifically preventing the activation of the ERK-activatingMAPK/ERK kinase (MEK-1) [21, 22]. TEC monolayers,untreated or treated for 6 h with single or combinedinhibitors at 5, 25 and 50 ? M, were stimulated by g 4 orc mAb cross-linking, washed and cultured for 24 h.Inhibitors, present throughout the experiment, did notaffect g 4 expression at TEC surface or TEC laminin
3040 F. Mainiero et al. Eur. J. Immunol. 2003. 33: 3038–3048
© 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
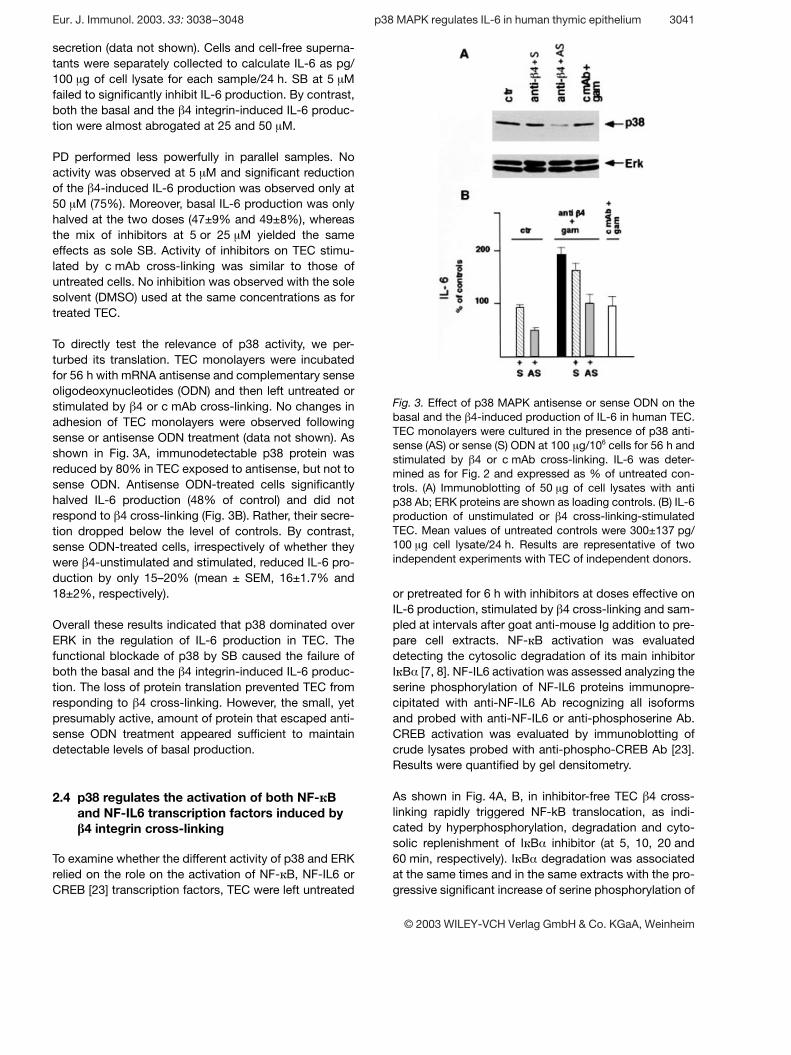
Fig. 3. Effect of p38 MAPK antisense or sense ODN on thebasal and the g 4-induced production of IL-6 in human TEC.TEC monolayers were cultured in the presence of p38 anti-sense (AS) or sense (S) ODN at 100 ? g/106 cells for 56 h andstimulated by g 4 or c mAb cross-linking. IL-6 was deter-mined as for Fig. 2 and expressed as % of untreated con-trols. (A) Immunoblotting of 50 ? g of cell lysates with antip38 Ab; ERK proteins are shown as loading controls. (B) IL-6production of unstimulated or g 4 cross-linking-stimulatedTEC. Mean values of untreated controls were 300±137 pg/100 ? g cell lysate/24 h. Results are representative of twoindependent experiments with TEC of independent donors.
secretion (data not shown). Cells and cell-free superna-tants were separately collected to calculate IL-6 as pg/100 ? g of cell lysate for each sample/24 h. SB at 5 ? Mfailed to significantly inhibit IL-6 production. By contrast,both the basal and the g 4 integrin-induced IL-6 produc-tion were almost abrogated at 25 and 50 ? M.
PD performed less powerfully in parallel samples. Noactivity was observed at 5 ? M and significant reductionof the g 4-induced IL-6 production was observed only at50 ? M (75%). Moreover, basal IL-6 production was onlyhalved at the two doses (47±9% and 49±8%), whereasthe mix of inhibitors at 5 or 25 ? M yielded the sameeffects as sole SB. Activity of inhibitors on TEC stimu-lated by c mAb cross-linking was similar to those ofuntreated cells. No inhibition was observed with the solesolvent (DMSO) used at the same concentrations as fortreated TEC.
To directly test the relevance of p38 activity, we per-turbed its translation. TEC monolayers were incubatedfor 56 h with mRNA antisense and complementary senseoligodeoxynucleotides (ODN) and then left untreated orstimulated by g 4 or c mAb cross-linking. No changes inadhesion of TEC monolayers were observed followingsense or antisense ODN treatment (data not shown). Asshown in Fig. 3A, immunodetectable p38 protein wasreduced by 80% in TEC exposed to antisense, but not tosense ODN. Antisense ODN-treated cells significantlyhalved IL-6 production (48% of control) and did notrespond to g 4 cross-linking (Fig. 3B). Rather, their secre-tion dropped below the level of controls. By contrast,sense ODN-treated cells, irrespectively of whether theywere g 4-unstimulated and stimulated, reduced IL-6 pro-duction by only 15–20% (mean ± SEM, 16±1.7% and18±2%, respectively).
Overall these results indicated that p38 dominated overERK in the regulation of IL-6 production in TEC. Thefunctional blockade of p38 by SB caused the failure ofboth the basal and the g 4 integrin-induced IL-6 produc-tion. The loss of protein translation prevented TEC fromresponding to g 4 cross-linking. However, the small, yetpresumably active, amount of protein that escaped anti-sense ODN treatment appeared sufficient to maintaindetectable levels of basal production.
2.4 p38 regulates the activation of both NF- k Band NF-IL6 transcription factors induced byI 4 integrin cross-linking
To examine whether the different activity of p38 and ERKrelied on the role on the activation of NF- ‹ B, NF-IL6 orCREB [23] transcription factors, TEC were left untreated
or pretreated for 6 h with inhibitors at doses effective onIL-6 production, stimulated by g 4 cross-linking and sam-pled at intervals after goat anti-mouse Ig addition to pre-pare cell extracts. NF- ‹ B activation was evaluateddetecting the cytosolic degradation of its main inhibitorI ‹ B § [7, 8]. NF-IL6 activation was assessed analyzing theserine phosphorylation of NF-IL6 proteins immunopre-cipitated with anti-NF-IL6 Ab recognizing all isoformsand probed with anti-NF-IL6 or anti-phosphoserine Ab.CREB activation was evaluated by immunoblotting ofcrude lysates probed with anti-phospho-CREB Ab [23].Results were quantified by gel densitometry.
As shown in Fig. 4A, B, in inhibitor-free TEC g 4 cross-linking rapidly triggered NF-kB translocation, as indi-cated by hyperphosphorylation, degradation and cyto-solic replenishment of I ‹ B § inhibitor (at 5, 10, 20 and60 min, respectively). I ‹ B § degradation was associatedat the same times and in the same extracts with the pro-gressive significant increase of serine phosphorylation of
Eur. J. Immunol. 2003. 33: 3038–3048 p38 MAPK regulates IL-6 in human thymic epithelium 3041
© 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim

Fig. 4. The blockade of p38 or ERK MAPK activity differentially affects NF- ‹ B binding activity and NF-IL6 serine phosphorylationinduced by g 4 integrin. TEC monolayers were left untreated or stimulated by g 4 cross-linking as for Fig. 1 with or without a 6-hpretreatment with p38 or ERK inhibitors as for Fig. 2. Cytosolic extracts were prepared from cells harvested at the indicatedtimes. (A) Representative Western blotting of I ‹ ¥ § in untreated (–) or treated TEC (15 ? g) at the indicated times. Shown are sec-tions of the same gel. (C) Representative Western blotting of NF-IL6 proteins of the same cell samples used in (A), immunoprecip-itated (50 ? g) with anti-NF-IL6 Ab, and immunoblotted with anti-phosphoserine mAb (pSer) or anti-NF-IL6 Ab (NF-IL-6). The 5-min time point is not shown in the gel. (E) Detection of CREB and phosphorylated CREB proteins by Western blotting of crudelysates of cells stimulated as for (A). Shown are sections of immunoblots of the same experiment. (B, D, F) Gel densitometry(three independent experiments) of (A, B, E). Bars represent the mean values ± SEM of OD/mm2 of bands, re-calculated as per-cent of untreated controls. NF-IL6, pSer-NF-IL6, pCREB and CREB proteins, all of the apparent molecular mass of 43 kDa, areindicated by arrows.
the active NF-IL6 isoforms (2.5-fold the basal level at10 min; Fig. 4C, D), but not with changes in the phos-phorylation of CREB (Fig. 4E, F). SB at 25 ? M preventedI ‹ B § degradation and drastically reduced the increase ofNF-IL6 phosphorylation induced by g 4 cross-linking,whereas in parallel samples PD only prevented I ‹ B §degradation. Similar results were obtained when thedose of the drugs was raised to 50 ? M (not shown).c mAb cross-linking or the presence of the sole solventwere not effective in all assays. These results indicatedthat g 4 cross-linking induced activation of both NF- ‹ Band NF-IL6 with similar kinetics in the same TEC, andthat p38, but not ERK, mediated both processes, consis-tently with the dramatic effect of its functional blockadeon IL-6 production.
2.5 p38 regulates the basal activation of bothNF- k B and NF-IL6 transcription factors inTEC monolayers
As p38 inhibition almost abrogated also the basal IL-6production (see Fig. 2), we investigated its role on thebasal activation of NF- ‹ B and NF-IL6 transactivators.TEC, cultured with SB or PD at 25 and 50 ? M and sam-pled at 0, 2, 6 and 18 h after drug addition were used toprepare cell extracts. NF-IL6 activation was evaluated asfor Fig. 4. For NF- ‹ B, instead of analyzing the decreaseof I ‹ B § induced by a single stimulus (such as g 4 cross-linking), we measured the nuclear binding activity result-ing from basal signaling ongoing in TEC monolayers.Results were quantified by gel densitometry.
3042 F. Mainiero et al. Eur. J. Immunol. 2003. 33: 3038–3048
© 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim

Fig. 5. Pharmacological blockade of p38 or ERK MAPK activity differentially affects the basal level of NF- ‹ B binding activity andNF-IL6 serine phosphorylation. Nuclear and cytosolic extracts were prepared from TEC monolayers, cultured in the presence ofSB or PD inhibitors at 25 and 50 ? M and harvested at 2 (not shown in the gel), 6 and 18 h. (A) Representative time course of NF-‹ B binding activity of nuclear extracts (6 ? g) probed with [ + -32P]ATP-double-stranded oligoprobe. Retarded bands of NF- ‹ B het-
erodimers, of CBF-1 proteins, and free probe are indicated. (B) Gel densitometry of three independent experiments. Bars repre-sent mean values ± SEM of relevant bands expressed as for Fig. 4. (C) Nuclear extracts of the same cell samples (50 ? g) immu-noprecipitated and immunoblotted as for Fig. 4C. NF-IL6- and pSer-NF-IL6-specific bands are indicated. (D) Gel densitometry ofthree independent experiments. Bars represent mean values ± SEM of relevant bands as for (B).
As shown in Fig. 5A, B, no significant effects wereobserved after 2 h of treatment with each inhibitor (notshown in the gel). Both caused instead a significant aver-age decrease of 40% and 60% at 6 h and 18 h, respec-tively. No earlier or greater activity was achieved whenthe dose of the inhibitor was raised to 50 ? M (notshown). CBF-1, a member of the Notch family binding asequence in the NF- ‹ B site of IL-6 promoter [24],remained substantially unmodified, thus ensuring equalprotein probing and treatment specificity.
Immunoblotting of extracts of the same cell samplesimmunoprecipitated as for Fig. 4C, D, showed that SBprogressively reduced the serine phosphorylation of theNF-IL6 43-kDa isoform to barely detectable levels,whereas PD did not exert significant effects. Lamininsecretion, detected by immunoblotting as in Fig. 1, wasunchanged by the treatment with SB or PD at the variousdoses (not shown). These results demonstrated that p38regulates the basal production of IL-6 by controlling thebasal activation of NF- ‹ B and NF-IL6 transactivators.
2.6 Rac1, PAK1 and MKK3/MKK6 arecomponents of the signaling pathwaytriggered by I 4 cross-linking and resulting inp38 activation
Due to the relevance of p38 activation, we investigatedsome of the g 4-mediated mechanisms tuning its phos-phorylation. PAK1, MKK3 and MKK6, all found to beimplicated in p38 activation, were investigated as first[25, 26]. PAK1 activation was analyzed by in vitro kinaseassays performed on anti-PAK1 immunocomplexes iso-lated from unstimulated or g 4–stimulated TEC. As visual-ized by the phosphorylation of the exogenous substrateMBP, PAK1 activation was present at 5 min and maximalat 10 min after stimulation (Fig. 6A).
To assess whether g 4 cross-linking could also induceactivation of MKK3/MKK6, two closely related dual-specificity protein kinases that activate p38 MAPK, weperformed Western blot analysis using phosphospecificAb. As shown in Fig. 6B, g 4 cross-linking caused signifi-cant activation of MKK3/MKK6, which was rapid, peak-ing at 5 min, but still persistent at 10 min. It is known thatthe serine/threonine kinase PAK1 binds GTP-bound
Eur. J. Immunol. 2003. 33: 3038–3048 p38 MAPK regulates IL-6 in human thymic epithelium 3043
© 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim

Fig. 6. Activation of PAK1, MKK3/MKK6 and Rac1 by g 4 integrin cross-linking in human TEC. TEC were left untreated or stimu-lated as in Fig. 1 for the indicated times at 37°C. (A) In vitro kinase assay performed with anti-PAK1 immunoprecipitates usingMBP as substrate. PAK1 immunoprecipitated from each sample was evaluated by immunoblotting with anti-PAK1 Ab. (B) Activa-tion of MKK3/MKK6 examined by immunoblotting of crude lysates. The amounts of MKK3 protein are shown on the bottom. (C)Rac1 activation was evaluated incubating TEC (extracted as above) with the GST-PAK fusion protein bound to glutathione-coupled Sepharose beads at 4°C for 30 min. Bound active GTP-Rac molecules were analyzed by Western blotting using an anti-Rac1 mAb. Cell lysates probed for total Rac1 are shown as loading control on the bottom. Results are representative of one ofthree independent experiments.
Rac1 [27]. Thus, to examine whether g 4 cross-linkingactivates Rac1 in TEC, we performed a Rac activationassay with a glutathione S-transferase (GST)-PAK fusionprotein binding and precipitating the active form of Rac.We found that g 4 cross-linking caused endogenousRac1 activation (Fig. 6C). c mAb cross-linking failed tofunction in all assays. As the time course of Rac1 activa-tion paralleled that of p38, PAK1 and MKK3/MKK6, theseresults overall indicated that g 4 integrin-induced p38activation was associated in TEC with a persistent acti-vation of PAK1, MKK3/MKK6 and Rac1.
3 Discussion
Animals and/or cellular models have demonstrated thatgrowth, survival, migration and adhesion of normal ortumor epithelia can be modulated by § 6 g 4 integrin,through distinct signaling pathways originating from itscytoplasmic tail and regulating the assembly/de-assembly of hemidesmosomes or the expression of cellcycle- or cell survival-related genes [16, 17]. Less atten-tion has been paid to this integrin as a regulator of thesecretion of normal and tumor epithelia. We show herefor the first time that § 6 g 4 integrin interaction with exog-
enous or TEC-secreted laminins regulates p38 phos-phorylation, and that p38 activity is an essential regulatorof IL-6 gene expression in TEC.
Results were obtained in cells maintained as close aspossible to “physiological” conditions. Signals from g 4integrin were elicited by laminins or mAb cross-linkingwith soluble secondary Ab. Under these conditions, wefound that g 4 stimulation intensely increased the low lev-els of activation of p38, ERK, Rac1, PAK1 and MKK3/MKK6 found in unstimulated cells. It has been reportedthat Rac1 controls ERK, Jnk, and p38 activation, thatPAK1 is involved in ERK and p38 activation, and thatMKK3/MKK6 lie upstream the activation of p38 [25–28].No data were available before our report on activation ofp38 signaling pathway following g 4 integrin stimulation.
Experiments done in the presence of inhibitors demon-strated that both p38 and ERK were involved in IL-6 generegulation in TEC. However, the concentration of the p38inhibitor SB required to completely inhibit IL-6 secretionmay have affected the activity of Lck or protein kinase A(PKA), as previously indicated by “in vitro” kinase assays[29]. Although “in vivo” inhibitions may require higherconcentrations than “in vitro” kinase assays [29] and
3044 F. Mainiero et al. Eur. J. Immunol. 2003. 33: 3038–3048
© 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim

§ 6 g 4 integrin seems to lack the association with Lck[30], an involvement in the pathways elicited by g 4 cross-linking in our cells cannot be excluded. The same appliesfor PKA, although several reports have indicated thatPKA can negatively regulate g 2 integrin activity and thatg 1 integrins suppress or stimulate PKA activity [31–33],
but no data are available at our knowledge on PKAinvolvement in g 4 functions or vice versa. Overall, ourdata obtained with p38 antisense ODN showed that theyreduced p38 protein synthesis in TEC (80%), signifi-cantly halved the basal IL-6 production (48% of control),and blocked that induced by g 4 cross-linking (Fig. 3 B),thus strongly suggesting the functional relevance of p38.
When comparing p38 and ERK activity, we found that theblockade of each kinase hampered the production of IL-6, but the dramatic reduction resulting from p38 inhibi-tion best demonstrated its functional predominance.One of the mechanisms underlying p38 predominanceappears to be its dual activity on NF- ‹ B and NF-IL6. Infact, ERK regulated NF- ‹ B translocation, but its block-ade did not abrogate IL-6 production. Only when NF- ‹ Band NF-IL6 activations were both prevented in loss orloss-function experiments of p38, TEC production of IL-6 dropped to barely detectable levels. This indicated thatIL-6 gene expression in TEC needs, as in other cells, themultiple interactions of NF- ‹ B and NF-IL6 [7].
The finding that p38 phosphorylation was regulated byg 4 integrin is of particular interest in thymic epithelium.
Strongly expressed at the TEC surface throughout theirlife cycle, § 6 g 4 integrin mediates TEC-extracellularmatrix (ECM) and TEC-thymocyte adhesion at the sametime [11–13]. We here clearly demonstrate that laminin-§ 6 g 4 interaction is one of the mechanisms maintaining
the basal phosphorylation of p38 and hence the basalproduction of IL-6 as both decreased upon disruption ofTEC-TEC-secreted laminin interaction by anti-lamininAb. If so, signals delivered by g 4 integrin during ECM-ligand recognition could contribute to maintain a basallevel of IL-6 production whereas additional recruitmentsupon thymocyte tethering could push IL-6 gene intowaves of expression finally regulating the overall amountof cytokine present in the microenvironment.
It is well established that IL-6 exerts multiple activitieswithin the thymus, due to the cell sharing of its receptor.Autocrine activity is exerted directly or synergisticallywith other cytokines or epidermal growth factor (EGF)and accounts for the control of proliferation and differen-tiation of thymic epithelium [2, 5]. In this regard, the find-ing that § 6 g 4 activates Rac1 in the pathway leading toIL-6 gene expression is interesting. Recent findings haveshown that Rac1 activation decreases the expression ofcadherin thereby loosing intercellular adhesions [34]. If
so, g 4-activated Rac1 could favor TEC responses to themitogenic activity of IL-6 meanwhile participating in theinduction of its expression.
Paracrine activities of IL-6 comprise the promotion ofsurvival and proliferation of developing thymocytes and,as recently described in human cells, the regulation ofmonocyte-dendritic differentiation [35]. Thus, thedeployment of the full range of IL-6 activities may be crit-ical for development and homeostasis of both epithelialcell meshwork and lymphoid lineage, thus finally regulat-ing the maintenance of thymic architecture. Moreover, itsanti-apoptotic activities may skew towards positiveselection thymocytes that are shaping their TCR reper-toire. As undesired effect IL-6 might also support autore-active clones, thus playing a role in autoimmune dis-eases associated to thymus pathology such as myasthe-nia gravis, where IL-6 hyperproduction by medullary TEChas been counted among the pathogenetic mechanisms[36]. This considering, the regulation of IL-6 gene expres-sion by p38 elucidated in the present report may lead toa deeper understanding of physiological and pathologi-cal processes taking place in the thymus and open a wayto possible therapeutic manipulations.
4 Materials and methods
4.1 Reagents
Laminin, casein, anti-phosphoserine and anti-laminin (LAM-89) mAb, and goat anti-rabbit Ab were from SIGMA (Milan,Italy); anti- g 4 integrin mAb (3E1), PD and SB inhibitors werefrom Calbiochem-Novabiochem (San Diego, CA); anti-IFN § /g receptor mAb (34F10) was a gift of Dr. M. Tovey (Viral
Oncology, CNRS, Villejuif, France); FITC-labeled F(ab’)2-goatanti-mouse Ig was from Becton Dickinson (Mountain View,CA); anti-ERK, anti-p38, anti-NF-IL6 and anti-I ‹ B § antiserawere from Santa Cruz Biotechnology (Santa Cruz, CA). Puri-fied anti-mouse Ig was from Zymed Laboratories, Inc. (SanFrancisco, CA). Anti-CREB and anti-phospho-CREB anti-sera were from Upstate Biotechnology (Waltham, MA). Puri-fied F(ab’)2-goat anti-mouse Ig was from Cappell Laborato-ries (Cooper Biomedical, Malvern, PA). Sequences of senseand antisense ODN for p38, provided by MWG Biotech SpA(Florence, Italy), were as follows: sense 5’-AAGATGTCG-CAGGAGAGGCC-3’; antisense 5’-GGCCTCTCCTGCGA-CATCTT-3’. Laminin 5 matrices were prepared from mousemammary RAC-11P/SD cells as previously described [16,17].
Eur. J. Immunol. 2003. 33: 3038–3048 p38 MAPK regulates IL-6 in human thymic epithelium 3045
© 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim

4.2 Culture and treatment of TEC
Primary TEC were derived under informed consent from dis-carded tissue of 14 independent thymuses of patients( X 5 years of age) undergoing cardiac surgery as previouslydescribed [11–13]. Secondary cultures were grown to con-fluence at 1/3 of medium complements and then deprived ofthem for 48 h. If indicated, TEC were incubated with saturat-ing doses of the appropriate mAb (5 ? g/ml, 45 min at 4°C),washed and incubated (37°C) for different times with F(ab’)2-goat anti-mouse Ig (5 ? g/ml). Long- and short-term stimula-tions were obtained by plating 2×106 cells on laminin (30 ? g/well)-precoated (overnight, 4°C) 24-well Costar plates or onlaminin 5 matrices. TEC, deprived of growth factors andmedium complements while reaching confluence, were cul-tured for 24 h in the presence or absence of anti-laminin,anti- g 4 or control Ab. When indicated, TEC were pretreated(6 h at 37°C) with single or combined ERK or p38 inhibitorsat 5, 25 and 50 ? M in medium without complements but inthe presence of solvent (DMSO) used at 1:1,000 final dilu-tion. Inhibitors were present throughout the experiments. Ifindicated, before stimulation TEC (2.5×105/ml) were treatedwith antisense and complementary sense ODN (100 ? g/ml)in culture medium for 56 h [28].
4.3 Cell extracts and electrophoretic mobility shiftassay
Nuclear and cytoplasmic extracts were prepared from TECas previously described [11–13]. Protein amount was mea-sured by Coomassie protein assay reagent (Pierce, Rock-ford, IL). DNA binding activity was determined using a [ + -32P]ATP-labeled double-stranded probe encompassing theNF- ‹ B binding site of the IL-6 promoter [24]. Cell extracts(6 ? g) were incubated for 30 min at room temperature withthe probe (1×105 cpm/sample) in 20 ? l of binding buffer[20 mM Tris-HCl pH 7.5, 0.1 M NaCl, 1 mM dithiothreitol(DTT), 1 mM EDTA, 1 mg/ml BSA, 0.1% Nonidet-P40(NP40), 4% glycerol] containing 1 ? g of poly(dI-dC) (Phar-macia, Uppsala, Sweden). Nucleoprotein complexes wereelectrophoresed on a 5% polyacrylamide gel in 0.05 M Tris-borate/1 mM EDTA at 150 V, dried gels were exposed toAmersham Hyperfilm films (Amersham, Little Chalfont, GB),and signals were quantified with an UltroScan Densitometerand the built-in software (LKB, Bromma, Sweden).
4.4 Immunoprecipitation, Western blotting andin vitro kinase assay
Cell lysates were prepared as previously described for elec-trophoretic mobility shift assay. Immunoprecipitation wascarried out with 20 ? g of extracts precleared with 20 ? l ofprotein A-Sepharose beads (Pharmacia) in lysis buffer(20 mM Tris-HCl pH 7.2, 0.5 mM DTT, 1 mM EDTA, 0.5 mMPMSF adjusted to 0.15 M NaCl) for 1 h at room temperature,incubated with anti NF-IL6 Ab at 1:1,000 dilution for 2 h on
ice, precipitated with 20 ? l of protein A-Sepharose beads for1 h at room temperature and washed in 0.05 M Tris-HClpH 7.2, 0.15 M NaCl, 0.1% NP40. Lysates (20 ? g) or immu-noprecipitates were fractionated by 12% SDS-PAGE, elect-roblotted to nitrocellulose membrane (Bio-Rad, Milan, Italy)and probed with anti-phosphoserine mAb in TBST contain-ing 1.5% w/v low-fat dry milk. Membranes were washedwith TBST buffer and bands revealed by Enhanced Chemilu-minescence system (ECL, Amersham).
Phosphorylated forms of ERK, p38 and MKK3/MKK6 weredetected by immunoblotting using PhosphoPlus Erk andp38 and MKK3/6 Ab Kit (New England Biolabs, Hitchin, GB)according to the manufacturer’s instructions. Rac1 activa-tion was evaluated by incubating TEC extracts with the GST-PAK fusion protein (kindly provided by Dr. J. G. Collard, TheNetherlands Cancer Institute, Amsterdam, The Nether-lands), bound to glutathione-coupled Sepharose beads at4°C for 30 min. Bound active GTP-Rac molecules were ana-lyzed by immunoblotting using an anti-Rac1 mAb (UpstateBiotechnology, Lake Placid, NY). PAK1 was immunoprecipi-tated from TEC lysates with anti-PAK1 Ab (Santa Cruz Bio-technology) and subjected to in vitro kinase assay as previ-ously described [28].
4.5 IL-6
IL-6 was measured by ELISA according to manufacturer’sinstructions (Endogen, Woburn, MA). Optical density (OD)values, plotted on the standard curve, were expressed aspg/100 ? g of cell lysates/24 h for each sample.
4.6 Statistical analysis
Statistical analysis of the results of Fig. 2–5 was performedapplying the parametric unpaired t-test. Significance wasaccepted when p X 0.05.
Acknowledgements: We are indebted to Dr. M. Tovey forproviding 34F10 mAb, to Dr. J. G. Collard for the GST-PAKfusion protein and to Dr. M. Cippitelli for anti-CREB Ab. Wegratefully acknowledge Prof. P. C. Marchisio and Prof. A.Santoni for helpful suggestions and criticisms and Prof. A.Colombatti for invaluable advice regarding laminin experi-ments. This work was partially supported by Telethon(n.1178) to D. Ramarli, MURST 1999 to Marcello Merola andto Stefania Morrone, and MURST 2001 to Fabrizio Mainieroand to Giuseppe Tridente.
References
1 Boyd, R. L., Tucek, C. L., Godfrey, D. I., Izon, D. J, Wilson, T. J.,Davidson, N. J., Bean A. G., Ladyman H. M., Ritter, M. A. andHugo, P., The thymic microenvironment. Immunol. Today 1993.14: 445–459.
3046 F. Mainiero et al. Eur. J. Immunol. 2003. 33: 3038–3048
© 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim

2 Manley, N., Thymus organogenesis and molecular mechanismsof thymic epithelial cell differentiation. Semin. Immunol. 2000. 12:421–428.
3 Savino, W., Serra Villa-Verde, D. M., Alves, L. A. and Dar-denne, M., Neuroendocrine control of the thymus. Ann. N. Y.Acad. Sci. 1998. 840: 470–479.
4 Sempowsky, G. D., Hale, L. P., Sundy, J. S., Massey, J. M.,Koup R. A., Douek, D. C., Patel, D. D. and Haynes B. F., Leuke-mia inhibitory factor, oncostatin M, IL-6, and stem cell factormRNA expression in human thymus increases with age and isassociated with thymic atrophy. J. Immunol. 2000. 164:2180–2187.
5 Screpanti, I., Meco, D., Scarpa, S., Morrone, S., Frati, L.,Gulino, A. and Modesti, A., Neuromodulatory loop mediated bynerve growth factor and interleukin 6 in thymic stromal cell cul-tures. Proc. Natl. Acad. Sci. USA 1992. 89: 3209–3212.
6 Wery-Zennaro, S., Zugaza, J. L., Letourneur, M., Bertoglio, J.and Pierre, J., IL-4 regulation of IL-6 production involves Rac/Cdc42- and p38 MAPK-dependent pathways in keratinocytes.Oncogene 2000. 19: 1596–1604.
7 Akira, S. and Kishimoto, T., NF-IL6 and NF- ‹ B in cytokine generegulation. Adv. Immunol. 1997. 65: 1–46.
8 Li, Q. and Verma, I. M., NF- ‹ B regulation in the immune system.Nat. Immunol. 2002. 2: 725–734.
9 Ossipow, V., Descombes, P. and Schibler, U., CAAT/enhancerbinding protein mRNA is translated into multiple proteins with dif-ferent transcription activation potentials. Proc. Natl. Acad. Sci.USA. 1993. 90: 8219–8223.
10 Berrih, S., Savino, W. and Cohen, S., Extracellular matrix of thehuman thymus. An immunofluorescence study on frozen sec-tions and cultured thymic epithelial cells. J. Histochem. Cyto-chem. 1985. 33: 655–664.
11 Ramarli, D., Scupoli, M. T., Fiorini, E., Poffe, O., Brentegani,M., Villa, A., Cecchini, G., Tridente, G. and Marchisio, P. C.,Thymocyte contact or mAb-mediated clustering of § 3 g 1 or § 6 g 4integrins activate IL-6 transcription factors (NF- ‹ B and NF-IL6)and IL-6 production in human thymic epithelial cells. Blood 1998.92: 3745–3755.
12 Fiorini, E., Marchisio, P. C., Scupoli, M. T., Poffe, O., Tagliabue,E., Brentegani, M., Colombatti, M., Santini, F., Tridente, G. andRamarli, D., Adhesion of immature and mature T cells induces inhuman thymic epithelial cells (TEC) activation of IL-6 gene tran-scription factors (NF-kappaB and NF-IL6) and IL-6 gene expres-sion: role of alpha3beta1 and alpha6beta4 integrins. Dev. Immu-nol. 2000. 7: 195–208.
13 Scupoli, M. T., Fiorini, E., Marchisio, P. C., Poffe, O., Tagliabue,E., Brentegani, M., Tridente, G. and Ramarli, D., Lymphoidadhesion promotes human thymic epithelial cell survival via NF-(kappa)B activation. J. Cell. Sci. 2000. 113: 169–177.
14 Falcioni, R., Perrotti, N., Piaggio, G., Kennel, S. K. and Sacchi,A., Insulin-induced phosphorylation of the beta-4 integrin subunitexpressed on murine metastatic carcinoma cells. Mol. Carcinog.1989. 2: 361–368.
15 Shaw, L. M., Identification of insulin receptor substrate 1 (IRS-1)and IRS-2 as signaling intermediates in the § 6 g 4 integrin-dependent activation of phosphoinositide 3-OH kinase and pro-motion of invasion. Mol. Cell. Biol. 2001. 21: 5083–5093.
16 Mainiero, F., Pepe, A., Wary, K. K., Spinardi, L., Mohammadi,M., Schlessinger, J. and Giancotti, F. G., Signal transduction bythe integrin § 6 g 4 distinct g 4 subunit sites mediate recruitment ofShc/Grb2 and association with the cytoskeleton of hemidesmo-somes. EMBO J. 1995. 14: 4470–4481.
17 Mainiero, F., Murgia, C., Wary, K. K., Curatola, A. M., Pepe, A.,Blumemberg, M., Westwick, J. K., Der, C. J. and Giancotti, F.G., The coupling of § 6 g 4 integrin to Ras-MAP kinase pathwaysmediated by Shc controls keratinocyte proliferation. EMBO J.1997. 16: 2365–2375.
18 Jefferies, C. A. and O’Neil, L. A., Rac1 regulates interleukin 1-induced nuclear factor ‹ B activation in an inhibitory protein I ‹ B § -independent manner by enhancing the ability of the p65 subunitto transactivate gene expression. J. Biol. Chem. 2000. 275:3114–3120.
19 Vanden Berghe, W., Plaisance, S., Boone E., De Bosscher, K.,Schmitz, M.-L., Fiers, W. and Haegeman, G., p38 and extracel-lular signal-regulated kinase mitogen-activated protein kinasepathways are required for nuclear factor- ‹ B p65 transactivationmediated by tumor necrosis factor. J. Biol. Chem. 1998. 273:3285–3290.
20 Craig, R., Larkin, A., Mingo, A. M., Thuerauf, D. J., Andrews,C., McDonough, P. M. and Glembotski, C. C., p38MAPK andNF- ‹ B collaborate to induce interleukin-6 gene expression andrelease. J. Biol. Chem. 2000. 275: 23814–23824.
21 Lee, J. C., Laydon, J. T., McDonnel, P. C., Gallagher, T. F.,Kumar, S., Gree, D., McNulty, D., Blumenthal, M. J., Heys, J.R. and Landvatter, S. W., A protein kinase involved in the regula-tion of inflammatory cytokine biosynthesis. Nature 1994. 372:739–746.
22 Alessi, D. R., Cuenda, A., Cohen, P., Dudley D. T. and Saltiel,A. R., PD 98059 is a specific inhibitor of the mitogen-activatedprotein kinase kinase in vitro and in vivo. J. Biol. Chem. 1995.270: 27489–27494.
23 Tokunou, T., Ichiki, T., Takeda, K., Funakoshi, Y, Iino, N., Shi-mokawa, H., Egashira, K. and Takeshita, A., Trombin inducesinterleukin-6 expression through the cAMP1 response element invascular smooth muscle cells. Arterioscler. Thromb. Vasc. Biol.2001. 21: 1759–1763.
24 Palmieri, M., Sasso, M. P., Monese, R., Merola, M., Faggioli,L., Tovey, M. and Furia, A., Interaction of the nuclear proteinCBF1 with the ‹ B site of the IL-6 gene promoter. Nucleic AcidRes. 1999. 13: 2785–2791.
25 Bagrodia, S., Derijard, B., Davis, R. J. and Cerione, R. A.,Cdc42 and PAK-mediated signaling leads to Jun kinase and p38mitogen-activated protein kinase activation. J. Biol. Chem. 1995.270: 27995–27998.
26 Raingeaud, J., Whitmarsh, A. J., Barrett, T., Derijard, B. andDavis, R. J., MKK3- and MKK6-regulated gene expression ismediated by the p38 mitogen-activated protein kinase signaltransduction pathway. Mol. Cell. Biol. 1996. 16: 1247–1255.
27 Martin, G. A., Bollag, G., McCormick, F. and Abo, A., A novelserine kinase activated by Rac1/Cdc42Hs-dependent autophos-phorylation is related to PAK65 and STE20. EMBO J. 1995. 14:1970–1978.
28 Mainiero, F., Soriani, A., Strippoli, R., Jacobelli, J., Gismondi,A., Piccoli, M., Frati, L. and Santoni, A., Rac1/p38 MAPK sig-naling pathway controls g 1 integrin-induced interleukin-8 (IL-8)production in human natural killer (NK) cells. Immunity 2001. 2:7–16.
29 Davies, S. P., Reddy, H., Caivano, M. and Cohen, P., Specificityand mechanism of action of some commonly used protein kinaseinhibitors. Biochem. J. 2000. 351: 95–105.
30 Mariotti, A., Kedeshian, P. A., Dans, M., Curatola, A. M.,Gagnoux-Palacios, L. and Giancotti, F. G., EGF-R signalingthrough Fyn kinase disrupts the function of integrin § 6 g 4 at hemi-desmosomes: role in epithelial cell migration and carcinoma inva-sion. J. Cell Biol. 2001. 155: 447–457.
Eur. J. Immunol. 2003. 33: 3038–3048 p38 MAPK regulates IL-6 in human thymic epithelium 3047
© 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim

31 Jones, S. L., Protein kinase A regulates beta 2 integrin avidity inneutrophils. J. Leuk. Biol. 2002. 71: 1042–1048.
32 Kim, S., Bakre, M., Yin, H. and Varner, J. A., Inhibition of endo-thelial cell survival and angiogenesis by protein kinase A. J. Clin.Invest. 2002. 110: 913–914.
33 O’Connor, K. L. and Mercurio, A. M., Protein kinase A regulatesRac and is required for the growth factor-stimulated migration ofcarcinoma cells. J. Biol. Chem. 2001. 276: 47895–47900.
34 Braga, V. M. M., Betson, M., Li, X. and Lamarche-Vane, N.,Activation of the small GTPase Rac is sufficient to disruptcadherins-dependent cell-cell adhesion in normal human kerati-nocytes. Mol. Biol. Cell. 2000. 11: 3703–3721.
35 Chomarat, P., Banchereau, J., Davoust, J. and Palicha, A. K.,IL-6 switches the differentiation of monocytes from dendritic cellsto macrophages. Nat. Immunol. 2000. 1: 510–514.
36 Vincent, A., Unravelling the pathogenesis of myasthenia gravis.Nat. Rev. Immunol. 2002. 2: 797–804.
Correspondence: Dunia Ramarli, Serv. Imm. Clinica, Policli-nico G. B. Rossi, V. le L. A. Scuro n. 1, I-37134 Verona, ItalyFax: +39-045-8202459e-mail: dunia.ramarli — mail.univr.it
3048 F. Mainiero et al. Eur. J. Immunol. 2003. 33: 3038–3048
© 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim




















