
Bubble-free oxygenation of a bi-enzymatic system: effect on biocatalyst stability

Progress in Oceanography xxx (2008) xxx–xxx
ARTICLE IN PRESS
Contents lists available at ScienceDirect
Progress in Oceanography
journal homepage: www.elsevier .com/locate /pocean
Oxygenation episodes on the continental shelf of central Peru: Remoteforcing and benthic ecosystem response
Dimitri Gutiérrez *, E. Enríquez, S. Purca, L. Quipúzcoa, R. Marquina, G. Flores, M. GracoDirección de Investigaciones Oceanográficas, Instituto del Mar del Perú, P.O. Box 22 Callao, Peru
a r t i c l e i n f o a b s t r a c t
Article history:Accepted 14 October 2008Available online xxxx
Keywords:Peru marginBenthic communitiesThioplocaOxygen minimum zoneENSORemote forcing
0079-6611/$ - see front matter � 2008 Elsevier Ltd. Adoi:10.1016/j.pocean.2008.10.025
* Corresponding author. Tel./fax: +51 1 6250827.E-mail addresses: [email protected] (D.
mail.com (D. Gutiérrez).
Please cite this article in press as: Gutiérrenogr. (2008), doi:10.1016/j.pocean.2008.1
The interplay between the oxygen minimum zone and remotely-forced oxygenation episodes determinesthe fate of the benthic subsystem off the Central Peruvian coast. We analyzed a 12 year monthly time-series of oceanographic and benthic parameters at 94 m depth off Callao, Central Peru (12�S), to analyze:(i) near-bottom oxygen level on the continental shelf in relation to dynamic height on the equator(095�W); and (ii) benthic ecosystem responses to oxygen change (macrobiotic infauna, meiofauna, andsulphide-oxidizing bacteria, Thioploca spp.). Shelf oxygenation episodes occurred after equatorialdynamic height increases one month before, consistent with the propagation of coastal trapped waves.Several but not all of these episodes occurred during El Niños. The benthic biota responded to oxygena-tion episodes by undergoing succession through three major ecological states. Under strong oxygen defi-ciency or anoxia, the sediments were nearly defaunated of macro-invertebrates and Thioploca was scarce,such that nematode biomass dominated the macro- and meiobiotas. When frequency of oxygenationevents reduced the periods of anoxia, but the prevailing oxygen range was 10–20 lmol L�1, mats of Thi-oploca formed and dominated the biomass. Finally, with frequent and intense (>40 lmol L�1) oxygena-tion, the sediments were colonized by macrofauna, which then dominated biomass. The Thioploca stateevolved during the 2002–2003 weak EN, while the macrofauna state was developed during the onsetof the strong1997–1998 EN. Repeated episodes of strong oxygen deficiency during the summer of2004, in parallel with the occurrence of red tides in surface waters, resulted in the collapse of Thioplocamats and development of the Nematode state. Ecological interactions may affect persistence or thetransition between benthic ecosystem states.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
The upwelling zone off the Central Peruvian coast (8�S–14�S) ischaracterized by a higher biological production and a thicker andmore intense oxygen minimum zone (OMZ) than other latitudinalcoastal areas off Peru. In this zone, the main sources of the subsur-face dissolved oxygen are the Peru–Chile undercurrent (PUC),which is formed off northern Peru from a southern branch of theEquatorial Undercurrent, and the turbulent diffusion from surfacewaters; while the main sink is oxygen consumption by the highrespiratory demand of the settling and sedimentary organic mat-ter, especially near the coast (Rowe and Howarth, 1985; Codispotiet al., 1989; Fossing, 1990; Strub et al., 1998; Pennington et al.,2006). The combination of these processes causes intense oxygendepletion over the continental shelf (Gutiérrez et al., 2006a). Theoxygen regime though is subjected to significant temporal variabil-ity, especially by the poleward propagation of coastal trapped
ll rights reserved.
Gutiérrez), dim.gutierrez@g-
z, D. et al., Oxygenation episo0.025
waves (CTWs) with periods between 30 and 50 days. These waveshave their origin in the Equatorial region and occur more fre-quently during warm El Niño Southern Oscillation (ENSO) phaseperiods (Brink, 1982; Strub et al., 1998; Pizarro et al., 2001; Ca-mayo and Campos, 2006). The CTWs can be generated upon Kelvinwaves (KW) travelling the equatorial line reach the western coastof South America, but also from the propagation of free TropicalWaves, or even by anomalous activity of the northerly Panamawind jet, crossing the equatorial line (Brink, 1982; Hormazabal,2001; Camayo and Campos, 2006; Mosquera, 2006). The passageof the CTWs is associated with deepening of the nutricline, oxy-cline and the OMZ off the Peru–Chile coast (Brink et al., 1981; Mor-ales et al., 1999; Ulloa et al., 2001). The deepened nutriclinereduces rates of primary production at the surface and oxygen con-sumption in subsurface waters. Passage of equatorial waves thusincreases oxygen levels on the continental shelves.
Under neutral or cold ENSO phases, oxygen deficiency (<20 lM)occurs over the continental shelf bottoms while surface sedimentstend to be anoxic, e.g. with no oxygen and with free hydrogen sul-phide (Tyson and Pearson, 1991). Under these conditions benthicmacrofauna do not inhabit shelf sediments; such populations only
des on the continental shelf of central Peru: Remote ..., Prog. Ocea-
2 D. Gutiérrez et al. / Progress in Oceanography xxx (2008) xxx–xxx
ARTICLE IN PRESS
develop if sustained oxygenation occurs, allowing successfulrecruitment and growth (Tarazona et al., 1988a,b, 2001). Mats ofthe large sulphide-oxidizing/nitrate-reducing bacteria Thioplocaspp. develop under some conditions (Tarazona and Arntz, 1986;Zafra et al., 1988; Tarazona, 1990; Gutiérrez et al., 2006a). Thesebacteria glide up and down in their sheaths, obtaining energy byoxidizing hydrogen sulfide from below the sediment surface withnitrate from the water column. The mats can develop associatedto suboxic conditions in the surface sediments and in the overlyingwater, e.g. very low-oxygen levels but no hydrogen sulphide (Ty-son and Pearson, 1991; Bruland, 2006). Thus, oxic conditions or an-oxia in the sediment surface are detrimental for Thioploca growth(Gallardo, 1977; Jørgensen and Gallardo, 1999; Schulz et al.,1996, 2000). On the other hand, though information on continentalshelf meiofauna are scarce, recent work has found dense upper-slope meiofaunal populations within the OMZ off Central Peru,mostly composed of nematodes (Levin et al., 2002; Neira et al.,2001b).
Tarazona et al. (1988a,b, 2001) have described, for coastalembayments of Central Peru, the positive responses of macrofaunalcommunities to periods of oxygenation associated with El Niño’s(EN’s). Multi-year observations in the bays of Ancon (11�450S)and Independencia (14�200S) indicate that, following oxygenation,a recurrent pattern of colonization by benthic macrofauna occurs –observable as increases in diversity, abundance and biomass – andextending well beyond the termination of the EN (Tarazona et al.,1988a,b, 2001; Arntz et al., 2006). Nevertheless, local processessuch as upwelling, phytoplankton blooms followed by sinking,and sediment resuspension due to wave action or erosion mayadditionally be important in the benthic ecosystem responses toEN or oxygenation episodes (Gutiérrez et al., 2006b). The responseof deeper, continental shelf communities to EN off central Peru isless well known (Salzwedel et al., 1988; Arntz et al., 1991;Gutiérrez et al., 2002, 2006a,b). Off Callao (12�S) at 94 m depth, in-creases of macrofaunal species richness, biomass and bioturbationpotential occurred during the 1997–98 EN and lasted a year after
200200
00
400400
600600
800800
10001000
12001200
14001400
DepthDepth
W Lon
S La
titud
e 200200
00
400400
600600
800800
10001000
12001200
14001400
200200
00
400400
600600
800800
10001000
12001200
14001400
200200
00
400400
600600
800800
10001000
12001200
14001400
DepthDepth
Huach
11°
12°
13°79° 78°
78°79°
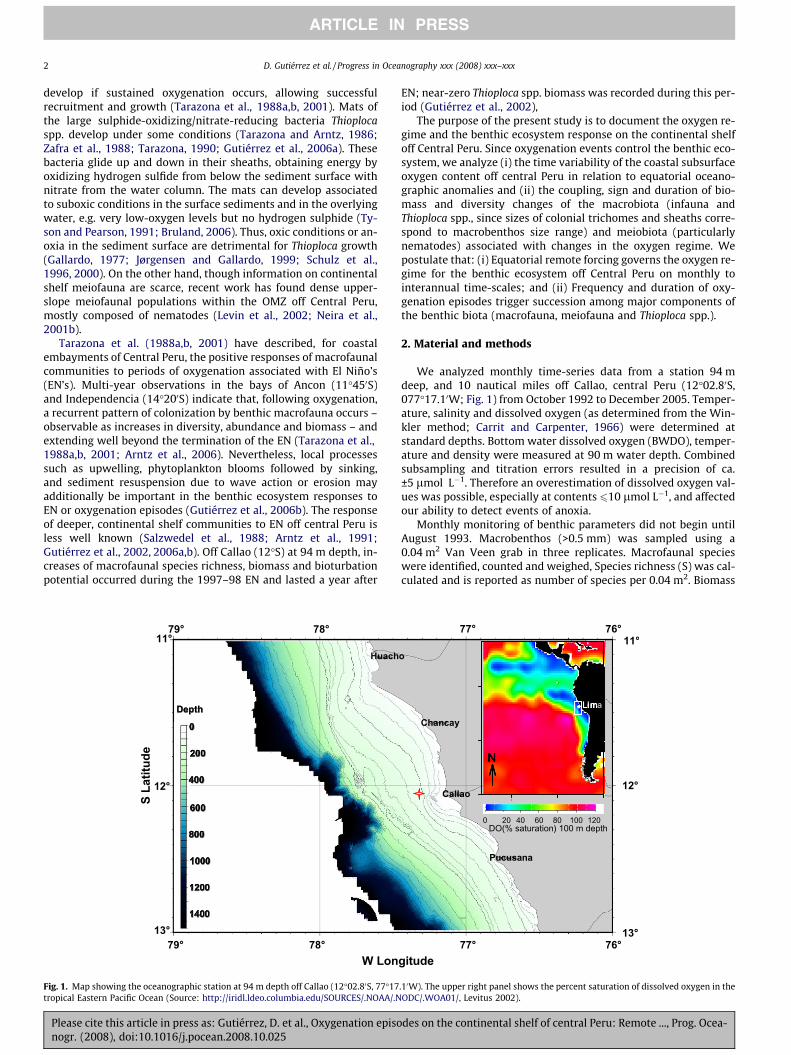
Fig. 1. Map showing the oceanographic station at 94 m depth off Callao (12�02.80S, 77�17tropical Eastern Pacific Ocean (Source: http://iridl.ldeo.columbia.edu/SOURCES/.NOAA/.N
Please cite this article in press as: Gutiérrez, D. et al., Oxygenation episonogr. (2008), doi:10.1016/j.pocean.2008.10.025
EN; near-zero Thioploca spp. biomass was recorded during this per-iod (Gutiérrez et al., 2002),
The purpose of the present study is to document the oxygen re-gime and the benthic ecosystem response on the continental shelfoff Central Peru. Since oxygenation events control the benthic eco-system, we analyze (i) the time variability of the coastal subsurfaceoxygen content off central Peru in relation to equatorial oceano-graphic anomalies and (ii) the coupling, sign and duration of bio-mass and diversity changes of the macrobiota (infauna andThioploca spp., since sizes of colonial trichomes and sheaths corre-spond to macrobenthos size range) and meiobiota (particularlynematodes) associated with changes in the oxygen regime. Wepostulate that: (i) Equatorial remote forcing governs the oxygen re-gime for the benthic ecosystem off Central Peru on monthly tointerannual time-scales; and (ii) Frequency and duration of oxy-genation episodes trigger succession among major components ofthe benthic biota (macrofauna, meiofauna and Thioploca spp.).
2. Material and methods
We analyzed monthly time-series data from a station 94 mdeep, and 10 nautical miles off Callao, central Peru (12�02.80S,077�17.10W; Fig. 1) from October 1992 to December 2005. Temper-ature, salinity and dissolved oxygen (as determined from the Win-kler method; Carrit and Carpenter, 1966) were determined atstandard depths. Bottom water dissolved oxygen (BWDO), temper-ature and density were measured at 90 m water depth. Combinedsubsampling and titration errors resulted in a precision of ca.±5 lmol L�1. Therefore an overestimation of dissolved oxygen val-ues was possible, especially at contents 610 lmol L�1, and affectedour ability to detect events of anoxia.
Monthly monitoring of benthic parameters did not begin untilAugust 1993. Macrobenthos (>0.5 mm) was sampled using a0.04 m2 Van Veen grab in three replicates. Macrofaunal specieswere identified, counted and weighed, Species richness (S) was cal-culated and is reported as number of species per 0.04 m2. Biomass
ma
N
m
N
m
N
m
N
gitude
0 20 40 60 80 100 120DO(% saturation) 100 m depth
o
Chancay
Pucusana
Callao
77° 76°13°
12°
11°76°77°
* Li* Li* Li* Li
.10W). The upper right panel shows the percent saturation of dissolved oxygen in theODC/.WOA01/, Levitus 2002).
des on the continental shelf of central Peru: Remote ..., Prog. Ocea-

D. Gutiérrez et al. / Progress in Oceanography xxx (2008) xxx–xxx 3
ARTICLE IN PRESS
of Thioploca spp. was quantified as wet weight of sheaths contain-ing cell trichomes. Percent of sheaths with trichomes was alsodetermined for each sample. Macrofaunal biomass and abundance,and Thioploca spp. biomass is reported as grams per m2.
Meiofauna was sampled from October 2002 to December 2005,every second month, using a short gravity corer. Three cores werecollected by survey and each core was subsampled with a tube of3.6 cm�2 internal diameter, corresponding to a surface area of10 cm2; only the top one centimeter was analyzed. Major meiofaun-al taxa (excluding foraminifera) were sorted, identified and counted,and the densities were expressed as number of individuals per10 cm2 (Neira et al., 2001b). The Simpson dominance index (Simp-son, 1949) was calculated on the abundances per major groups.
To further compare biotic components, wet weights of macrofa-unal invertebrates were converted to ash-free dry weights (AFDW)using taxon-specific conversions (Brey et al., 1988, and referencestherein) and then to organic carbon using a factor of 0.518 (Salonenet al., 1976). Biomass of Thioploca spp. sheaths + trichomes wereconverted to carbon by assuming dry weight is 9% of wet weight(Huettel et al., 1996), and carbon is 12.5% of dry weight (Thamdrupand Canfield, 1996). Finally, nematode wet weight was first esti-mated from biovolume measurements with microscope followingAndrassy, 1956), then converted to carbon (12.4%, following Wie-ser, 1960). Carbon is expressed as g Cm�2.
We used linear interpolation computed from monthly surveyprofiles to estimate the depth of (1) the 45 lmol L�1 oxygen level,taken as the upper boundary of the oxycline (�1 mL L�1), (2) the22 lmol L�1oxygen level, taken as the upper boundary of theOMZ (�0.5 mL L�1), thereafter ‘vertical position of the OMZ’, and(3) the isopycnals rt = 26 kg m�3 and rt = 26.5 kg m�3. Since theseoxygen levels are well above 610 lmol L�1, methodological diffi-culties associated with low-oxygen estimation using Winkler titra-tion minimally affected oxycline and OMZ depth determination.
The daily, 5-day averaged and monthly time-series of the Equa-torial dynamic height at 95�W (TAO project buoy, http://www.pmel.noaa.gov/tao) were employed to track the propagationof sea level anomalies (associated with equatorial Kelvin waves,KW) and their impact on the local water column conditions (ascoastal trapped waves). Callao sea surface height was obtained dai-ly from Callao (Dirección de Hidrografía Naval del Perú, DHN).Monthly average wind speed from Callao (Chucuito) was also pro-vided by the DHN. These time-series were filtered with five dayaverages and interpolated linearly to fill gaps. Anomaly valueswere computed as standard units, in order to compare variancesof the time-series, and also were analyzed with discrete wavelets(Torrence and Compo, 1998) to represent temporal changes inspectra. Pearson cross correlations were calculated to investigateconnections between the local and remote time-series with 0, +1or >1 month lags (Emery and Thomson, 1998).
3. Results
3.1. Oceanographic variability, oxygen and remote forcing
3.1.1. Kelvin waves and the water columnThe oceanographic time-series off Callao (sea surface height,
temperature, density and oxygen) off Callao record (Fig. 2) theinfluence of KWs (Table 1), which are more frequent and strongduring EN. At least seven episodes during which the Callao thermo-cline and pycnocline were deepened while sea level in Callao rosecan be identified after an increase in the Equatorial dynamic height(arrows over Fig. 2). Five of these episodes occurred during ENs(February–March and June–July 1993; April–May and December1997; and April 2002). The other two episodes occurred in non-EN years (December 1994 and July 2000). Since 2003, at least
Please cite this article in press as: Gutiérrez, D. et al., Oxygenation episonogr. (2008), doi:10.1016/j.pocean.2008.10.025
two deepening episodes occurred and the relationship with theequatorial dynamic height is clear (Fig. 2). Table 1 links the Callaoepisodes with equatorial conditions, by comparing the equatorialin situ observations with Kelvin wave signals reported in the bibli-ography (Cravatte et al., 2003; Kessler and McPhaden, 1995a,b;Kessler and Kleeman, 2000; Dewitte et al., in press).
3.1.2. Oxygen and remote forcingIn comparison with the record of temperature and density, the
time-series of dissolved oxygen exhibited more pronounced varia-tions (Fig. 2c). The position of the OMZ oscillated from 10 to morethan 90 m depth, and its overall mean depth (±one standard devi-ation) was 52.3 m (±33.2 m). Seasonal differences in the verticalposition of the OMZ are not obvious due to the high interannualvariability. All seven pycnocline depressions (above) were also re-corded as water column oxygenation episodes, as the OMZ waspushed downwards more than one standard deviation, typicallyreaching the bottom at the study site. However the oxygen time-series reveal at least four more oxygenation episodes with similaramplitude as the above mentioned events (April 1994, April 1995,December 1995/January 1996 and September 2003). These corre-spond to moderate increases in dynamic height on the equatorand sea surface heights at Callao, with weak changes of the localthermal structure. In addition, several minor oxygenation episodesalso occurred, in which the OMZ was depressed to 60–90 m, so thatBWDO was 10–20 lmol L�1. These minor oxygenation episodes oc-curred during the weak 2002–2003 EN. In contrast, from August1998 to April 2000 (e.g., during the 1998–99 La Niña; LN), therewere no oxygenation episodes, nor were there disturbance ofEquatorial dynamic height or deepening event within the watercolumn.
3.1.3. Equatorial dynamic height and Callao sea surface heightCross-correlations yielded significant relationships between the
Equatorial dynamic height and the vertical position of the OMZ,and between the Equatorial dynamic height and the 26 kg m�3 iso-pycnal depth with a one month lag (P < 0.05; Fig. 3a and b). Similarresults were obtained with subsurface (P50 m) temperature, den-sity and dissolved oxygen with a one-month lag (Table 2). Thewavelet analysis on the difference between the standardizedanomalies of the Callao sea surface height and the Equatorial dy-namic height revealed two signals, one of them at 30–50 days, inrough agreement with the one-month lag explained above, andthe other one at 5–15 days, which was more active during 2002–2003 (Fig. 3c). The linear relationships between Callao oceano-graphic variables and the Equatorial dynamic height had higherresidual variances on subsurface dissolved oxygen (52% and 56%respectively, Table 2) than did density or temperature (28% and34%, respectively, also Table 2). Similarly, the portion of unex-plained variance was higher (�57%) for the iso-oxygen depths thanfor the isotherm or isopycnal depths (�40%). The portion of vari-ance not explained is higher when only the time-series from the2003–2005 period is analyzed, reaching more than 60% for the sub-surface temperature and density, as well as for the isotherm andisopycnal depths, and about 80% for dissolved oxygen and theiso-oxygen depths (data not shown).
3.1.4. Bottom water dissolved oxygenThe missing values of BWDO from some surveys could be
extrapolated from its relationship with the OMZ depth, since about88% of the variance of the BWDO was explained by the OMZ depthvia the equation: BWDO = e(0.22 � Z�OMZ) (n = 104). The time-seriesof BWDO is shown in Fig. 4a. Values above 20 lmol L�1 were char-acteristic of the mature 1997–98 EN, and also occurred betweenJanuary and July 1993. Other peaks in BWDO were associated withthe oxygenation episodes described above. Values 610 lmol L�1
des on the continental shelf of central Peru: Remote ..., Prog. Ocea-

Fig. 2. Time-series of equatorial DH and oceanographic variables off Callao (12�S): (a) 5-day averaged equatorial dynamic height at 95�W (GAL DYNH) (black, scale reversed)and Callao SSH (blue, scale reversed); (b) Temporal variability of temperature (�C, coloured) and density – rt (kg m�3; contour lines); (c) Temporal variability of dissolvedoxygen (lmol L�1); the thick isoline correspond to the vertical position of the OMZ (oxygen content = 22 lmol L�1). Arrows depict the impact of coastal trapped waves offCallao (see Table 3). Oxygenation episodes uncorrelated with depressions of the thermocline or pycnocline are marked with black circles below the time axis.
4 D. Gutiérrez et al. / Progress in Oceanography xxx (2008) xxx–xxx
ARTICLE IN PRESS
dominated during the 1998–1999 LN, and were more frequentthan on average during late 1993, in 1996–1997 and since early
Table 1Vertical structure and oxygenation episodes off Callao related with increases of dynamic hecolumn indicates the date of the episodes. The second column indicates the warm (W) or nOceanic El Niño Index ONI (NOAA, 2003); the May 97–April 1998 warm episode corresponremote forcing wave and its reference, respectively. KW = Equatorial Kelvin wave (Niñoperiods from power spectra.
In situ event ONI episode Remote fo
February–March 1993 March–July 1993, W DecemberCTW (10–
June–July 1993 March–July 1993, W December(60 d)
December 1994–February 1995 April 1994–March 1995, W December
April–May 1997 May 1997–April 1998, W March 199CTW (50–
December 1997 May 1997–April 1998, W DecemberJuly 2000 July–September 2000, N May 2000April 2002 May 2002–March 2003, W February 2
Please cite this article in press as: Gutiérrez, D. et al., Oxygenation episonogr. (2008), doi:10.1016/j.pocean.2008.10.025
2004. Values within the range 10–20 lmol L�1 dominated thetime-series between 2002 and 2004.
ight at 95�W, Equator (Galapagos), from in situ data (arrows in Figs. 2 and 4). The firsteutral (N) conditions in the Niño 3 + 4 region in the Equatorial Pacific, according to the
ds to the 1997–98 EN. The third and fourth columns indicate the date of the closest3 + 4 region), CTW = coastal trapped wave; in brackets, significant theoretical wave
rcing Reference
1992, KW (60 d) January 1993,50 d)
Kessler and McPhaden, (1995a) andCamayo and Campos (2006)
1992–September 1993, KW Kessler and McPhaden (1995b)
1994, KW (70–120 d) Cravatte et al. (2003) and Dewitte et al.(in press)
7, KW(70–120 d) May 1997,180 d)
Cravatte et al. (2003) and Kessler andKleeman (2000)
1997, KW (70 d) Cravatte et al. (2003), KW (120 d) Dewitte et al. (in press)002, KW (120 d) Dewitte et al. (in press)
des on the continental shelf of central Peru: Remote ..., Prog. Ocea-

cross correlation function-0.4 -0.2 0.0 0.2 0.4 0.6 0.8 1.0
-15-14-13-12-11-10-9-8-7-6-5-4-3-2-10123456789
101112131415
cross correlation function-0.4 -0.2 0.0 0.2 0.4 0.6 0.8 1.0
lag
(mon
ths)
-15-14-13-12-11-10-9-8-7-6-5-4-3-2-10123456789
101112131415
a b
c
Fig. 3. (a) Cross correlation function between the 95�W equatorial DH and the OMZ depth (22 lmol L�1 iso-oxygen) off Callao; (b) Cross correlation function between the the95�W equatorial DH and the 26 kg m�3 isopycnal depth; (c) Wavelet analysis of the difference between the standardized anomalies of the daily Callao SSH and the daily 95�Wequatorial DH, showing significant signals at periods of 5–15 days and 30–50 days; the power of spectral density (/) was calculated with Morlet wavelet and is represented incolors, the significant values are enveloped by a black line and the period is represented in days (Torrence and Compo, 1998). Dotted lines in (a) and (b) indicate critical valuesat significance level a = 0.05.
D. Gutiérrez et al. / Progress in Oceanography xxx (2008) xxx–xxx 5
ARTICLE IN PRESS
3.2. Benthic ecosystem
Surface sediments at this station are composed of diatomaceousand organic-rich (>4% TOC) mud, with high water content in thetop layer (>90%). Over time the sediments release free H2S or Thi-oploca spp. mats develop on. Water content decreases exponen-tially below the sediment surface and compact sediments (watercontent <60%) are present at about 10–15 cm deep.
3.2.1. MacrofaunaMeans and standard deviations of species richness (S), total
abundance (N) and total biomass (B) were 3 ± 2 spp. per0.04 m�2, 442 ± 1092 ind. m�2 and 4.3 ± 10.3 g m�2, respectively.Fig. 4b shows the temporal variabilities of S and B. Species richnessrose from August to October 1993, following the seven-month per-iod of oxygenation. The community was dominated by the amphi-
Please cite this article in press as: Gutiérrez, D. et al., Oxygenation episonogr. (2008), doi:10.1016/j.pocean.2008.10.025
pod Ampelisca araucana and the spionid polychaete Paraprionospiopinnata; other species that colonized the bottom were the poly-chaetes Nephtys ferruginea and Sigambra bassi, and the bivalve mol-lusk Pitar sp. The switch to strong oxygen deficiency was followedby a rapid decline down to zero values of the community parame-ters in February 1994. Thereafter S and B oscillated at low levelsuntil the end of 1995, but with transient increases in April 1994and February 1995, associated with oxygenation events. Near def-aunation was observed in late 1996 until February 1997, duringstrong oxygen deficiency. A rapid and early response of benthicmacrofauna to EN-associated oxygenation occurred from April toMay 1997, consisting in increases of S, N and B up to 3–10 timestheir long-term means. This early colonization was dominated byP. pinnata and A. araucana. From the oxygenated period September1997 to June 1998, S. bassi became dominant, and enteropneusts,nemerteans, several bivalves (e.g., Pitar sp.) and small decapods
des on the continental shelf of central Peru: Remote ..., Prog. Ocea-

Table 2Pearson product-moment correlation coefficients of monthly Equatorial DynamicHeight (1 month lag) at 95�W (DYNH-1), and monthly wind velocity at Callao (CAWINDVEL) with oceanographic parameters during the monthly field surveys offCallao: sea surface height (CASSH), oxygen concentration at 50 m depth (OX50),oxygen concentration at 90 m depth, near the bottom (BWDO), temperature at 50 mdepth (TEMP50), temperature near the bottom (TEMP90), sigma-t at 50 m depth(SIGMAT50), sigma-t near the bottom (SIGMAT90), OMZ 22 lmol L�1 upper boundarydepth (Z-OMZ), 45 lmol L�1 iso-oxygen depth (Z-OXYC), 15 �C isotherm depth (Z-15ISOT), 14 �C isotherm depth (Z-14ISOT), 26 kg m�3 isopycnal depth (Z-26ISOP), and26.5 kg m�3 isopycnal depth (Z-26.5ISOP). All the coefficients are significant(p < 0.001). Number of observations (n) is in brackets.
DYNH (�1) CAWINDVEL
CASSH 0.660 0.489(n = 146) (n = 112)
OX50 0.666 0.422(n = 122) (n = 87)
BWDO 0.654 0.384(n = 102) (n = 65)
TEMP50 0.849 0.460(n = 110) (n = 76)
TEMP90 0.849 0.449(n = 99) (n = 65)
SIGMAT50 �0.811 �0.507(n = 105) (n = 7)
SIGMAT90 �0.806 �0.503(n = 94) (n = 60)
Z-OMZ 0.656 0.418(n = 119) (n = 86)
Z-OXYC 0.643 0.402(n = 120) (n = 87)
Z-15ISOT 0.809 0.448(n = 109) (n = 7)
Z-14ISOT 0.657 0.334(n = 109) (n = 76)
Z-26ISOP 0.789 0.488(n = 108) (n = 75)
Z-26.5ISOP 0.752 0.426(n = 107) (n = 74)
6 D. Gutiérrez et al. / Progress in Oceanography xxx (2008) xxx–xxx
ARTICLE IN PRESS
colonized the sediments. S, N and B ranged 4–9 spp. per 0.04 m�2,336–1348 ind. m�2 and 4.2–74.5 g m�2, respectively. Larger organ-isms, mostly enteropneusts and nemerteans, accounted for the in-creases in biomass. In spite of the development of strong oxygendeficiency and even anoxia during the 1998–1999 LN, higher-than-mean values of the community parameters persisted forabout a year after the onset of the low-oxygen 1998–99 LN condi-tions. During late1999, community parameters decreased rapidlyto near-defaunation from September 2001 to November 2002. Onlyjuvenile specimens of P. pinnata or A. araucana were found duringthis period in the sediments. After December 2002, the communityparameters have fluctuated from near-zero values to small peakswithin the long-term means (e.g., early 2003, early 2004 and late2005 for biomass).
3.2.2. ThioplocaFollowing Schulz et al. (1996), a ‘mat’ of Thioploca is defined as
the condition wherein a network of interwoven trichomes andsheaths at sediment:water interface is dense enough to stabilizethe sediment. From parallel observations of cores and grab sam-ples, this mat condition occurs when Thioploca biomass in grabsamples is J 30 g m�2. Thioploca fluctuated from zero to131.9 g m�2. The percent of sheaths with trichomes was positivelycorrelated with the wet weight of sheath material (r = 0.62,n = 128, p < 0.001). Zero or nearly zero Thioploca biomasses wererecorded during 1993. From early 1994 to July 1994, biomass in-creased slightly as did percent of sheaths with trichromes(Fig. 4c). Over the following year, both biomass and sheaths withtrichomes increased but with large fluctuations, and from February1996 to February 1997, both parameters fell to near-zero. Duringthe 1997–98 EN and the 1998–99 LN, Thioploca trichomes almost
Please cite this article in press as: Gutiérrez, D. et al., Oxygenation episonogr. (2008), doi:10.1016/j.pocean.2008.10.025
disappeared, but empty sheaths were found during the EN. Thiop-loca was then absent until September 2000, followed by a gradualincrease of biomass occurred from October 2000 to December2001, when most of the Thioploca sheaths contained trichomes.The highest biomasses (>100 g m�2) were found in 2002, and matswith sheaths containing trichomes remained dominant (70–90%)until January 2004. Over the next six months, both parameters de-creased with large fluctuations. Since then, from July 2004 to July2005, reduced levels of both parameters have occurred (30–70%and 0–10.7 g m�2, for percent sheaths with trichomes and biomass,respectively), followed by higher but fluctuating biomass values inthe last portion of 2005.
3.2.3. MeiofaunaBecause the meiofauna consists mainly of nematodes, Simp-
son’s dominance index is correlated with nematodes abundance(r = 0.78, n = 20, p < 0.001). From September 2002 until April2003, low nematode and total meiofaunal abundances occurred(<100 ind. 10 cm�2), and as a consequence of the low nematodenumbers, diversity was relatively high (3–7 taxa 10 cm�2; notshown). In addition to nematodes (65–90% of abundance), the mei-obiota also consisted of nemerteans, rotifers, ciliates and harpacti-coid copepods. Diversity fell suddenly in July 2003, when onlynematodes and rotifers were found. From early 2004 to April2005, nematodes abundance increased to 2052 ± 404 10 cm�2. Inparallel the nematode dominance reached 90–100%. Finally, inthe second half of 2005, only nematodes were found and those atreduced abundances (5–293 ind. 10 cm�2).
3.3. Correlations with environmental variables and among biologicalparameters
Among the macrofaunal and meiofaunal community parame-ters, the macrofaunal biomass and species richness, and the meio-faunal phyletic Simpson Index exhibited significant correlationswith the BWDO and the one-month lagged Equatorial dynamicheight (Table 3). In contrast, no significant correlation was foundbetween the Thioploca’s biomass or percent sheaths with tric-homes and these abiotic factors. On the other hand, significant neg-ative correlations were observed between the macrofaunalbiomass and species richness with Thioploca’s biomass and percentsheaths with trichomes, and between the meiofaunal phyleticSimpson Index and those Thioploca parameters. However, meiofa-unal and macrofaunal community parameters were not signifi-cantly correlated (Table 3).
4. Discussion
4.1. Remote forcing and subsurface dissolved oxygen
The average 50 m depth of the OMZ upper boundary at our Cal-lao station is in agreement with previous estimates for this latitude(Wooster and Gilmartin, 1961; Zuta and Guillén, 1970). However,the temporal variability of OMZ depth is very high, even duringnon-EN years (Fig. 2). OMZ depth is essentially non-seasonal, asis thermocline depth, SST and other oceanographic parametersoff the low-latitude Peruvian coast (Bohle-Carbonell, 1989; Tarazo-na, 1990). On the other hand, the significant correlation of the OMZposition with the one-month lagged Equatorial dynamic height isalmost certainly due to poleward propagation of CTWs from theequator (Brink, 1982; Chavez et al., 1984; Hormazabal et al.,2001). As mentioned above, CTWs can develop from diversesources, so that a variety of atmospheric/oceanic processes in theEquatorial region can trigger the propagation of the anomalies inthe oceanographic conditions along the Peruvian coast.
des on the continental shelf of central Peru: Remote ..., Prog. Ocea-

1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006
BW
DO
(µm
ol L
-1)
0
10
20
30
40
80
120
OM
Z d
epth
(m
)
0
20
40
60
80M
acro
fau
nal
Bio
mas
s (g
m-2
)
0
10
20
30
50
100
150
200
spec
ies
rich
nes
s(N
° sp
p 0
.04
m-2
)
0
2
4
6
8
10
12
al biohnes
Th
iop
loca
Bio
mas
s (g
m-2
)
0
10
20
30
50
100
150
200
%sh
eath
s w
ith
tri
cho
mes
0
20
40
60
80
100
Thi ass% s ric
Year
1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 Mei
ofa
un
al d
ensi
ty (
N° 1
0 cm
-2)
0100200300400500
1000
1500
2000
2500
Ph
ylet
ic S
imp
son
Ind
ex
0.0
0.2
0.4
0.6
0.8
1.0
1.2
M nsS x
ioploca biomasheaths with t
Meiofaunal denSimpson Index
macrofaunspecies ric
schomes
sity
omassss
a
b
d
c
Fig. 4. Time-series of benthic community parameters related to subsurface and bottom dissolved oxygen variability: (a) bottom water dissolved oxygen content (blackcircles) and Z-OMZ (dotted line); the short – dash lines correspond to the 10 and 20 lmol L�1 content levels near the bottom; (b) macrofaunal biomass (black circles) andspecies richness (mean and ± 1 S.D., open circles); (c) biomass of Thioploca spp. (black circles) and percent of Thioploca sheath material with trichomes (dotted line); and (d)meiofaunal density and phyletic dominance Simpson Index. Arrows depict the impact of coastal trapped waves off Callao (see Table 3). Shaded red, blue-sky and green boxesenvelope the periods of the benthic ecosystem states: macrofauna state, Nematoda state and Thioploca state, respectively.
D. Gutiérrez et al. / Progress in Oceanography xxx (2008) xxx–xxx 7
ARTICLE IN PRESS
The variance not explained by the relationship between thesubsurface dissolved oxygen or the oxycline/OMZ depth with the
Please cite this article in press as: Gutiérrez, D. et al., Oxygenation episonogr. (2008), doi:10.1016/j.pocean.2008.10.025
Equatorial dynamic height is higher than those from the subsurfacetemperature and density. Some oxygenation episodes were not
des on the continental shelf of central Peru: Remote ..., Prog. Ocea-

Tabl
e3
Pear
son
prod
uct-
mom
ent
corr
elat
ion
coef
fici
ents
betw
een
abio
tic
fact
ors
BWD
O,
CAW
IND
VEL
and
DYN
H(�
1)an
dbe
nthi
cco
mm
unit
ypa
ram
eter
s,an
dam
ong
bent
hic
com
mun
ity
para
met
ers:
mac
rofa
unal
biom
ass
(MA
CRO
FB)
,m
acro
faun
alde
nsit
y(M
ACR
OF
N),
mac
rofa
unal
spec
ies
rich
ness
(MA
CRO
FS)
,mei
ofau
nald
ensi
ty(M
EIO
FN
),m
eiof
auna
lphy
leti
cgr
oups
rich
ness
(MEI
OF
PHYL
S),m
eiof
auna
lSim
pson
Inde
x(S
IMPS
ON
IND
EX),
perc
ent
Thio
ploc
ash
eath
sw
ith
tric
hom
es(%
LSH
EATH
S),T
hiop
loca
biom
ass
(TH
IOP
B),a
ndN
emat
oda
num
eric
aldo
min
ance
(NEM
AT
D).
Abi
otic
fact
ors
abbr
evia
tion
sar
eex
plai
ned
inTa
ble
2.Th
eco
rrel
atio
nsth
atar
est
atis
tica
llysi
gnifi
cant
ata
=0.
05(*
),a
=0.
01(*
*)an
da
=0.
001
(***
)ar
ein
dica
ted.
MA
CR
OF
BM
AC
RO
FN
MA
CR
OF
SM
EIO
FN
MEI
OF
PHY
LSSI
MPS
ON
IND
EX%
LSH
EATH
STH
IOP
BN
EMA
TD
CA
WIN
DV
EL0.
196
(n=
77)
0.19
0(n
=77
)0.
174
(n=
77)
––
–�
0.09
8(n
=77
)0.
004
(n=
77)
BW
DO
0.19
4*(n
=11
0)0.
032
(n=
110)
.240
*(n
=11
0)�
.069
0(n
=20
)0.
205
(n=
20)
�0.
435
(p=
0.05
5)(n
=20
)�
0.14
4(n
=11
0)�
0.08
4(n
=11
0)�
.398
(n=
20)
DY
NH
(�1)
0.29
2**(n
=10
8)0.
149
(n=
108)
0.26
3**
(n=
108)
�0.
308
(n=
20)
0.23
1(n
=20
)�
0.52
4*(n
=20
)�
0.07
2(n
=10
8).0
185
(n=
108)
�0.
363
(n=
20)
MA
CR
OF
B.2
34*
(n=
114)
.624
2***
(n=
114)
�0.
157
(n=
20)
0.28
3(n
=20
)�
0.36
5(n
=20
)�
.366
***
(n=
114)
�.2
00*
(n=
114)
�0.
199
(n=
20)
MA
CR
OF
N.4
17**
*(n
=11
4)�
0.03
4(n
=20
)0.
118
(n=
20)
0.02
4(n
=20
)0.
051
(n=
114)
�0.
040
(n=
114)
0.01
4(n
=20
)M
AC
RO
FS
�0.
170
(n=
20)
�0.
044
(n=
20)
�0.
030
(n=
20)
�.4
74**
*(n
=11
4)�
.311
**(n
=11
4)0.
061
(n=
20)
MEI
OF
N0.
288
(n=
20)
0.23
7(n
=20
)�
0.34
1(n
=20
)�
0.31
7(n
=20
)0.
203
(n=
20)
MEI
OF
PHY
LS�
0.62
8**
(n=
20)
0.48
3*(n
=20
)0.
262
(n=
20)
�0.
701*
**(n
=20
)SI
MPS
ON
IND
EX�
.556
*(n
=20
)�
0.56
2**
(n=
20)
0.77
6***
(n=
20)
%LS
HEA
THS
.641
**(n
=11
4)�
0.47
6*(n
=20
)TH
IOP
B�
.596
**(n
=20
)
8 D. Gutiérrez et al. / Progress in Oceanography xxx (2008) xxx–xxx
ARTICLE IN PRESS
Please cite this article in press as: Gutiérrez, D. et al., Oxygenation episonogr. (2008), doi:10.1016/j.pocean.2008.10.025
accompanied by similar changes of the thermocline and pycno-cline, while some periods of strong oxygen deficiency (mostly after2002) occurred in spite of depressed thermocline and pycnocline.These differences might be explained by the complex and non-lin-ear interactions between coastal upwelling, wind mixing, and redtide formation (Codispoti et al., 1989; Smayda, 2000; Graco et al.,2001). For example, moderate warming of the water column mayresult in decreased surface primary production followed by lowersubsurface oxygen consumption and higher BWDO values.
On the other hand, from 2003 onwards Equatorial dynamicheight fluctuations have been weaker and less correlated withthe Callao oceanographic parameters, and coherence between theCallao parameters themselves have been non-significant (oxycline,thermocline and pycnocline depths). Since the monthly Callaocoastal wind speed exhibits a positive trend since 1992, especiallyfor March–April (F(1,26) = 12.68; p < 0.01, R2 = 0.33; slope t-test = 3.56; p < 0.01), it is likely that coastal upwelling and turbu-lent mixing have been intensified in late summer and early fall,interfering with any equatorial modulation of oxygen supply. Thesummer/fall approach of warm, stratified subtropical waters to-wards the coast (Zuta and Guillén, 1970; Morón, 2006) causes for-mation of a steep upwelling front during wind events. When windsrelax, the front and thermal stratification migrate towards thecoast, favoring the development of red tides (Sánchez and Delgado,1996; Smayda, 2000). In fact, a positive trend in summer/fallSeaWifs chlorophyll-a off Callao from 1997 to 2005, and a higherfrequency and longer permanency of red tides related with mas-sive algal blooms off the Central Peruvian coast since 2002, havebeen detected (F. Chávez and S. Sánchez, personal communication).During field surveys we frequently observed red tides during sum-mer and fall in 2004 and 2005. Red tide blooms end in massivephytodetrital sedimentation and decay (Smayda, 2000), whichcan produce local anoxia on the shelves. Such strong local eventsdecouple the remotely-driven oxygen temperature and densityrelations in the water column, and likely explain at least some ofthe variability between the oxycline/OMZ and the thermocline orpycnocline observed since 2002.
4.2. Benthic ecosystem succession and oxygenation
Changes in benthic ecosystems off Peru and Chile have previ-ously been related to ENSO (Arntz et al., 2006; Tarazona et al.,1988a,b, 2001; Gutiérrez et al., 2002, 2006a,b; Sellanes et al.,2007). In the Bay of Ancón, during non-EN years, shallow bottoms(34 m) are dominated by characteristic species that are tolerant tooxygen deficiency, such as the polychaetes Magelona phyllisae andChaetozone sp, Thioploca, or nematodes. During the EN, short pulsesof opportunistic species occur, some developing early (as the poly-chaete P. pinnata and the ophiurid Ophiactis kroyeri) and otherspersisting for at least a year after EN termination (Tarazona et al.,1988a,b, 2001). Off Concepcion (36�S, central Chile), the extreme1997–1998 EN altered the seasonal regime of oxygen deficiencyand carbon flux with a >1 year period of bottom oxygenation andreduced carbon flux. Consequently the macrofaunal assemblageswitched from tube-dwelling, interface feeders (P. pinnata and A.araucana) to burrowing, deposit feeders (the polychaetes Medioma-stus branchiferus or Cossura chilensis; Gutiérrez et al., 2002; Sellaneset al., 2007).
The biomass of the large sulphur-oxidizing bacteria Thioploca isstrongly reduced during EN. Off Peru, large decreases of biomasshave been associated with abundant empty sheaths in the surfacesediments (Zafra et al., 1988; Arntz et al., 1991, 2006; Gutiérrezet al., 2002). Off central Chile, Gallardo (1985) found that Thioplocamats dominated the macrobenthic biomass in 1975, a non-EN year,but almost disappeared during the 1982–1983 EN. Afterwards inthe same location, Schulz et al. (2000) found a reduction in
des on the continental shelf of central Peru: Remote ..., Prog. Ocea-

Table 4Properties of the oxygen regime and major community descriptors for the different benthic ecosystem states (Macrof = macrofaunal-dominated; Thiop = Thioploca-dominated;and Nemat = Nematoda-dominated) in the continental shelf sediments off Callao (see text). For BWDO, it is indicated the frequency distribution (number of cases and percent),according to range in concentrations, for each of the states. For the rest of cases, mean values ± 1 S.D., number of observations (in brackets), and range of values are given. Nodata = ‘n.d.’.
Properties and descriptors Benthic states
Macrof Thiop Nemat
Period February 1996–February 1997August 1997–June 1999 August 2001–January 2004 August 1999–September 2000
August 2004–July 2005BWDO 6 10 nmolL�1:n(%) 10(42%) 5(23%) 18(56%)BWDO 10–20 nmol L�1:n(%) 6(25%) 15(68%) 12(38%)BWDO > 20 nmol L�1:n(%) 8(33%) 2(9%) 2(6%)Z-OMZ 71 ± 47 (24) 58 ± 22 (22) 36 ± 27 (32)
10–159 24–108 9–110BWDO concentration (nmoL L�1) 34 ± 43 (24) 13 ± 5 (22) 10 ± 6 (32)
0–143 4–26 0–30Macrofaunal biomass (g m�2) 18.9 ± 17.4 (21) 0.4 ± 0.7 (19) 0.4 ± 0.8 (30)
1.0–74.5 0.0–2.4 0.0–3.4Macrofaunal biomass (g C m�2) 1.148 ± 1.163 (21) 0.029 ± 0.052(19) 0.021 ± 0.038 (30)
0.069–5.264 0.000–0.197 0.000–0.158Macrofaunal abundance (N m�2) 657 ± 506 (21) 266 ± 746 (19) 124 ± 178 (30)
160–2300 0–3313 0–833Macrofaunal species richness (No. spp. 0.04 m�2) 6 ± 2(21) 1 ± 1 (19) 1 ± 1 (30)
4–9 0–2 0–4Thioploca biomass (g m�2) 0.0 ± 0.0 (21) 65.6 ± 35.5 (19) 2.1 ± 2.8 (30)
0.0–0.1 22.0–131.9 0.0– 10.7Thioploca biomass (g C m�2) 0.000 ± 0.000 (21) 0.738 ± 0.400 (19) 0.023 ± 0.031 (30)
0.000–0.001 0.248– 1.484 0.000–0.120Thioploca sheats with trichomes (%) 0.9 ± 2.5 (21) 88.6 ± 4.8 (19) 23.2 ± 22.9 (30)
0–10 70–90 0–70Meiofaunal abundance, top 1-cm (N 10 cm�2) n.d. 91 ± 75 (8) 613 ± 732 (6)
35–265 111–2052Nematoda biomass, top 1-cm (g C m�2) n.d. 0.016 ± 0.011 (8) 0.152 ± 0.212 (6)
0.005–0.033 0.005–0.546Meiofaunal phyletic richness (No. taxa 10 cm�2) n.d. 5 ± 1 (8) 3 ± 2 (6)
2–7 1–5Meiofaunal phyletic groups Simpson Index n.d. 0.58 ± 0.21 (8) 0.92 ± 0.09 (6)
0.34–0.94 0.76–1.00
Table 5Differences between Thioploca (Thiop), macrofauna (Macrof) and Nematoda (Nemat)carbon biomasses at each of the benthic ecosystem states, according to the non-parametric Kruskal–Wallis test. Homogenous groups were estimated from the Mann–Whitney U test (underlined). Nematoda biomass data only correspond to the top onecentimeter (see text).
State Kruskal–Wallis ANOVA p-value Homogenous groups (oc = 0.05)
Macrof H(1,42) = 33.39 <0.001 Thiop MacrofThiop H(2,46) = 33.27 <0.001 Macrof Nemat ThiopNemat H(1,68) = 6.47 0.039 Thiop Macrof Nemat
0
10
20
30
40
50
60
70
80
10 10 -20 20 - 30 30 - 40 > 40
BWDO (µmol L-1) class intervals
Freq
uenc
y (%
)
Macrof Thiop Nemat
Fig. 5. Frequency distribution (%) of BWDO (lmol L�1) under the macrofauna state(Macrof), Thioploca–state (Thiop), and Nematoda state (Nemat).
D. Gutiérrez et al. / Progress in Oceanography xxx (2008) xxx–xxx 9
ARTICLE IN PRESS
Thioploca biomass during the 1997–1998 EN as normally occursduring winter when normally upwelling is less frequent and theOMZ is deeper. For meiofauna, off Central Chile, Sellanes and Neira(2006) have reported an overall increase in abundance during the1997–1998 EN, though this increase varied along a benthic tran-sect between the Bay of Concepcion and the outer shelf (28–120 m). Harpacticoid copepod abundance increased while nema-todes decreased (Neira et al., 2001b; Sellanes and Neira, 2006).
Off Callao the benthic ecosystem was also affected by ENSO–driven oxygen changes. The system varied among three differentstates, with biomass dominance by: (i) macrofauna; (ii) Thioploca;or (iii) Nematoda (Table 4). The total ecosystem biomass differedsignificantly between these states (Kruskal–Wallis ANOVA andMann–Whitney tests; see Table 5 and below). The temporal fre-quency distributions of BWDO also differed (Fig. 5). In the ‘macro-fauna state’, the frequency distribution was bimodal, with the firstmode at 610 lmol L�1, and the second mode at >40 lmol L�1
(occurring on 42% and 25% of station occupations, respectively).In none of the other states did the >40 lmol L�1 class occur. Inthe ‘Thioploca state’, the 10–20 lmol L�1 was most common(68%). Finally, in the ‘Nematoda state’, the 610 lmol L�1 occurredduring 56% of station occupations. After collapsing the frequencydistribution into three classes (610 lmol L�1, 10–20 lmol L�1,and >20 lmol L�1), there was a significant association betweenecosystem state and oxygen concentration (chi-square tests,p < 0.05).
The macrofauna state evolved after the first coastal trappedwave reached Callao early during the 1997–98 EN, when O2 valuesreached 143 lmol L�1 near the bottom). Surprisingly, the macrofa-una state persisted after EN terminated and into 1999 under the
Please cite this article in press as: Gutiérrez, D. et al., Oxygenation episonogr. (2008), doi:10.1016/j.pocean.2008.10.025
extreme oxygen deficiency or even anoxia that characterized the1998–2000 LN (Fig. 5). This prolonged survival of macrofaunamay be explained by the colonization of bioturbating species
des on the continental shelf of central Peru: Remote ..., Prog. Ocea-

OXIC
Thioploca mats+
Meiofauna:high phyletic
diversity
Frequency of oxygenation events(%)
SULPHIDIC OXICPREVAILING condition in the SW interface
STRONG EN
MODERATE EN
NEUTRAL
LN
(-)
EN
SO p
hase
(+)
Beggiatoa mats+
Meiofauna:Nematodadominance
‘Classic’ benthicassemblages
Polychaeteinfaunal
community
(+)
T
OC
flu
x (
-)
SUBOXIC
Thioploca mats+
Meiofauna:high phyletic
diversity
Fig. 6. Conceptual model that explains the succession of shelf benthic communitiesoff Central Peru forced by ENSO and the associated frequency and intensity ofoxygenation episodes.
10 D. Gutiérrez et al. / Progress in Oceanography xxx (2008) xxx–xxx
ARTICLE IN PRESS
(mostly enteropneust and nemertean worms) that continued toventilate surface sediment even with very low-oxygen concentra-tions in the overlying waters (Gutiérrez et al., 2002). The switchto extreme oxygen deficiency associated to LN also resulted inthe depletion of nitrate (that is used for oxidize hydrogen sulphideby Thioploca), likely due to enhanced denitrification in the watercolumn (Graco et al., 2006). It was just in late 2000, after a CTWreached Callao depressing moderately the OMZ and increasingsubsurface nitrate (Graco et al., 2006), whereby Thioploca couldcolonize the seafloor. Then a Thioploca state characterized 2002–2003, during a weak EN. These years were characterized by a mod-erately deep oxycline and high near-bottom nitrate, but the pre-vailing oxygen range near the bottom was not enough formacrofaunal colonization (10–20 lmol L�1; Table 4; Fig. 5). Byearly 2004 under sustained oxygen deficiency (Figs. 2 and 4), themats were mostly gone, despite a deepening of the thermocline.During this year the macrofauna was also depauperate, while mei-ofaunal nematodes increased. Our data only assess meiofaunal inthe top centimeter of sediment, but other observations indicatethat this centimeter contains about 60% of the total meiofauna(<10 cm; E. Enríquez, personal communication). Since nematodescomprise over 90% of the metazoan meiofauna, nematode biomassin the top centimeter represent about 50% of the total meiofaunalbiomass. These conditions define the Nematoda state, which wasestablished from 2004 to 2005 (Fig. 4). Similar absence of bothmacrobenthic animals and Thioploca mats occurred during the ex-treme oxygen-deficient 1999–2000 period (see above). Probablythe Nematode state characterized the benthic ecosystem here aswell, since Nematoda populations multiply in anoxic settings withhigh loads of organic matter (Neira et al., 2001a; Levin et al., 2002;Levin, 2003). The Nematoda state may also have developed during1996–1997, when similar, though less extreme, conditions andreductions of macrobiota were observed (Fig. 4, Table 4).
Other periods exhibited mixed characters. In 2001–2002 andearly 2004, the biota was apparently in transition between the Thi-oploca and Nematoda states. Before 1996, macrofauna and Thioplo-ca biomasses oscillated, and a general decreasing trend ofmacrofaunal diversity was observed. Since an extended but moder-ate EN persisted from 1991 to 1993, it is possible that the variabil-ity reflects a late evolution of the macrofauna state associated tothe EN (Arntz et al., 2006). Frequent alternation of near-bottomoxygenation and depletion probably precluded development ofThioploca mats during these years.
4.3. Response times of the macrobiota
The macrofauna responded to oxygenation episodes with rapidincreases in biomass or species richness. Such increases occurredfollowing passage of the mid-1993, early 1995, March 1997 andmid 2000 CTWs. However, the colonizations were short-lived anddid not result in a macrofaunal state, as the oxygenated conditionsrapidly dissipated, with the exception of the extreme 1997–1998EN. Direct observation of the macrobiota during these transientoxygenation episodes reveals that most are new juvenile recruitsinhabiting the top centimeter of sediments, suggesting that mostcolonization is via larval settlement rather than immigration ofjuveniles and adults (Tarazona, 1990).
Thioploca also showed rapid colonization. During the earlyphase of the Thioploca state the population doubling time averaged34 ± 19 days with a minimum doubling time of 8 days, faster thanlaboratory growth rates under simulated optimal conditions (26–52 days; Otte et al., 1999) or on Concepcion shelf sediments (22–35 days; Schulz et al., 2000). Given optimal environmental condi-tions and starting from biomass values �1 g m�2, it apparentlytakes <2 months for the establishment of 100 g m�2 mats. On aver-age however, mats appear to take about eight months to form. This
Please cite this article in press as: Gutiérrez, D. et al., Oxygenation episonogr. (2008), doi:10.1016/j.pocean.2008.10.025
calculation is in agreement with the nine-month observed dura-tion of the transition to the Thioploca state (Fig. 4, Table 4). The pro-nounced �monthly fluctuations of Thioploca abundance suggestthat favorable growth conditions are often not sustained. For in-stance, the rapid collapse of the mats in early 2004 was associatedwith the appearance of fluffy, sulphidic layers in the surface sedi-ments. These layers most likely result from massive sedimentationof organic matter, presumably due to senescence and sinking of thered tides mentioned above. Degradation and respiration of this or-ganic matter probably switched redox conditions to anoxic. Exper-imental and field observations have shown that when the bottomwater is oxygen and nitrate-depleted and hydrogen sulphide buildsto high concentrations in the sediment-water interface, the Thiop-loca filaments emerge from the sediment and may be entrained inbottom flows, dispersing from their previous habitat (Gallardo,1985; Huettel et al., 1996).
4.4. A conceptual model of the benthic succession triggered by ENSO-driven oceanographic regimes
Here we frame a conceptual model to explain the variability ofthe benthic assemblages under contrasting oceanographic regimes,as were recorded in the 1993–2005 period off Callao (Fig. 6). Theswitch from anoxic (e.g., sulphidic) to suboxic and oxic conditionsdepends on the frequency and intensity of oxygenation events. Theecological effects of changes in the frequency and in the duration ofoxygenation periods are related. For example, it has been describedthat characteristic infaunal species inhabiting the shelf sediments,such as P. pinnata, have respiratory anaerobic pathways (Gonzálezand Quiñones, 2000) that permit them to survive in the transitionbetween oxygen deficiency and anoxic conditions. Thioploca movesup in surface sediments to avoid sulphidic conditions, preferringlow-oxygen concentrations in the overlying water (see below;Huettel et al., 1996). In addition, the nitrate storage capacity inits vacuoles should enable Thioploca to survive short periods whenthe bottom waters are anoxic and nitrate-depleted (Huettel et al.,1996). Free sulphide also reduces the ability of benthic animalsto survive absence of oxygen (Diaz and Rosenberg, 1995; Gamenicket al., 1996). Therefore, the characteristic shelf macrobiotic speciesseem to exhibit metabolic and behavioral adaptations to cope withshort periods of anoxia and their ability to survive depends on oxy-genation episodes. The duration of these episodes is constrained bythe characteristic periods of the coastal trapped waves (Table 1)and their frequency is normally increased during the EN phase ofENSO (Strub et al., 1998; Pizarro et al., 2001). During periods ofnegative Equatorial dynamic height anomalies (e.g., LN conditions),
des on the continental shelf of central Peru: Remote ..., Prog. Ocea-

D. Gutiérrez et al. / Progress in Oceanography xxx (2008) xxx–xxx 11
ARTICLE IN PRESS
the OMZ shoals and continental shelf oxygen levels are reduced.Carbon fluxes are also likely higher due to a shallower nutriclineand enhanced near-surface productivity. Consequently the shelfsediments are loaded with decaying organic matter, become anoxicand become dominated by nematode populations. This Nematodastate is also characteristic of sediments permanently located inthe core of the OMZ, where some nematodes contain symbiontswhich permit sulphide oxidation (Levin, 2003; Neira et al., 2001b).
As long as the benthic environment is subjected to oxygenationevents, nitrate is replenished in the bottom water, free sulphideand other reduced compounds are chemically removed and a sub-oxic layer is established, favoring the colonization by Thioplocatrichomes, which are easily resuspended and redistributed (Gal-lardo, 1985). However, the formation of mats will occur only if sub-oxic conditions persist. A positive feedback may be establishedwhen Thioploca become abundant enough to significantly reducesulphide levels by J 30%, pushing anoxic conditions deeper intothe sediment (Ferdelman et al., 1997; Fossing et al., 1995; Otteet al., 1999).
More frequent as well as more intense oxygenation (>20lmol L�1) permits development of the macrofaunal state that canreach higher biomasses under oxic conditions, as it occurred dur-ing the 1997–1998 EN. The reduction and disappearance of Thiop-loca in oxygenated conditions is consistent with previous reportsfor the Peru–Chile coast (Zafra et al., 1988; Schulz et al., 2000),but causes are elusive. Though nitrate is the major electron accep-tor in Thioploca metabolism (Otte et al., 1999), recent experimentshave demonstrated that Thioploca spp. can also respire aerobically(N.P. Revsbech, personal communication). Others have shown thattrichomes retreat into the sediment when oxygen reached 15 lMand nitrate was low (<5 lM), but there was not an avoidance ofoxygen under nitrate-rich (>20 lM) conditions (Huettel et al.,1996). Hence, the collapse of Thioploca populations seems to beonly indirectly controlled by oxygenation. Oxygen can influence ni-trate cycling (nitrification or denitrification) and sulphide oxida-tion. It seems that bottom water was not nitrate-depleted duringthe oxygenated 1997–1998 EN, varying from 8.8 to 19.8 lM (M.Graco, personal communication). Inspection of the sediments re-vealed abundant burrows and galleries and no sulphide smell atall, in contrast to conditions in non-EN periods. Possibly a reduc-tion of carbon fluxes during EN and a subsequent decrease of thereactivity of the stored organic matter by aerobic microbial respi-ration resulted in decreased sulphate reduction rates. In addition,flushing of oxygen into the sediments by benthic fauna could havepushed sulphate reduction deep into the compact sediment layer,hampering Thioploca survival. Part of the negative correlation be-tween Thioploca and macrofauna biomasses (Table 3) can be inter-preted by a differential tolerance to oxygen deficiency. This issupported by the spatial trends of Thioploca and macrofauna alongthe Northern and Central Peruvian shelf, showing that Thioplocatends to replace macrofauna at increasing latitude with strongerbottom water oxygen deficiencies (Gutiérrez et al., 2006a).
The negative correlation or the lack of correlation between Thi-oploca, and macrofauna or meiofauna (Table 3) questions the effec-tiveness of the ‘sulphide detoxification’ performed by Thioploca,particularly for the metazoan communities (Jørgensen and Gal-lardo, 1999; Neira et al., 2001b; Arntz et al., 2006). The tendencyfor higher diversity of meiofaunal major taxa to occur under theThioploca-dominated state can also be attributed to similar re-duced tolerances to anoxia by Thioploca and minor meiofaunalgroups, as harpacticoid copepods, rotifers, and others (Neiraet al., 2001b; Levin, 2003).
At shallower sites off Callao, absence of macrofauna and Thioplo-ca spp., and presence of nematode populations co-occur withsheathless filamentous bacterial mats, probably Beggiatoa spp.(D.G. pers. obs.). Sheathless, nitrate-vacuolated, sulphur-oxidizing
Please cite this article in press as: Gutiérrez, D. et al., Oxygenation episonogr. (2008), doi:10.1016/j.pocean.2008.10.025
mats are common in the semi-enclosed and shallow ConcepcionBay (Gallardo, 1979; Schulz et al., 2000; Graco et al., 2001). Thesheathless condition may be an adaptation to increase exchange be-tween the bacteria and nitrate-rich and the sulphidic layers (Gracoet al., 2001). Under the fluctuating environment off central Peru, thiscondition is more likely to occur in shallow areas with very high or-ganic load, as coastal embayments, permitting the transient co-exis-tence of sulphide buildup in the sediment surface with oxygenationand nitrate availability in the bottom water. There, the ‘Nematodastate’ may coexist with mats of sheathless filamentous bacteria.
5. Conclusions
Remote forcing controls the oxygen regime on the continentalshelf off Callao at monthly to interannual time-scales, taking intoaccount the characteristic periods associated with the passage ofcoastal trapped waves. When such forcing is weak (lower fre-quency and/or energy of coastal trapped waves, e.g. the period2003–2005), local physical, biological and biogeochemical pro-cesses (as those related to the development and fate of red tides)appear to control the level of near-bottom oxygenation.
Both intensity and frequency of oxygenation episodes and an-oxia control development of the benthic ecosystems of the Peru-vian shelf sediments. Permanent anoxia (e.g., sulphidic)conditions are unfavorable for the macrobiota, including Thioplocapopulations, but nematode populations can attain their highestbiomasses under anoxia. Increasing frequency of oxygenation epi-sodes favors the development of Thioploca mats as the most char-acteristic benthic ecosystem ‘state’. More intense oxygenationpermits recruitment of macrofauna that finally replace the matsas the main component of the macrobiota. Ecological interactions,mediated by the biological activity, as enhanced bioturbation andbio-irrigation, can additionally affect the persistence or transitionbetween these three characteristic ecosystem states.
Acknowledgements
We deeply thank the Instituto del Mar del Perú (IMARPE) forfull support of this research. Also we acknowledge the Direcciónde Hidrografía y Navegación del Peru (DHN) for kindly providingsea level data and monthly wind records off Callao. We acknowl-edge Mrs. Víctor Aramayo and José Pasapera who supported ana-lytical and data processing tasks. Dr. Francisco Chávez and Dr.David Foley kindly provided Seawifs chlorophyll-a informationfrom the zone off Callao. The EU-project CENSOR (Climate variabil-ity and El Nino Southern Oscillation: Impacts for natural resourcesand management, contract 511071) financed the post-doc grant ofM. Graco. This study was conducted in the frame of the EU-projectCENSOR (Climate variability and El Nino Southern Oscillation: Im-pacts for natural resources and management, contract 511071) andis CENSOR publication 0236.
References
Andrassy, I., 1956. Die rauminhalts- und gewichtsbestimmung der fadenwürmer(Nematoden). Acta Zoologica Academia Scientiarum Hungaricae 2, 1–15.
Arntz, W., Tarazona, J., Gallardo, V.A., Flores, L.A., Salzwedel, H., 1991. Benthoscommunities in oxygen deficient shelf and upper slope areas of the Peruvianand Chilean Pacific coast and changes caused by El Niño. In: Tyson, R.V.,Pearson, T.H. (Eds.), Modern and Ancient Continental Shelf Anoxia, vol. 58.Geological Society Special Publication, London, pp. 131–154.
Arntz, W., Gallardo, V.A., Gutiérrez, D., Isla, E., Levin, L.A., Mendo, J., Neira, C., Rowe,G.T., Tarazona, J., Wolff, M., 2006. El Niño and similar perturbation effects on thebenthos of the Humboldt, California, and Benguela Current upwellingecosystems. Advances in Geosciences 6, 243–265.
Bohle-Carbonell, M., 1989. On the variability of the Peruvian upwelling system. InPauly, D., Muck, P., Mendo, J., Tsukayama, I. (Eds.). ICLARM ConferenceProceedings of The Peruvian Upwelling Ecosystem: 1953–1984, vol. 18, pp.14–32.
des on the continental shelf of central Peru: Remote ..., Prog. Ocea-

12 D. Gutiérrez et al. / Progress in Oceanography xxx (2008) xxx–xxx
ARTICLE IN PRESS
Brey, T., Rumohr, H., Ankar, S., 1988. Energy content of macrobenthic invertebrates:general conversion factors from weight to energy. Journal of ExperimentalMarine Biology and Ecology 117, 271–278.
Brink, K.H., 1982. A comparison of long coastal trapped wave theory withobservations off Peru. Journal of Physical Oceanography 12, 897–913.
Brink, K.H., Jones, B.H., Van Leer, J.C., Mores, D.W., Stuart, C.N.K., Stevenson, M.R.,Dugdale, R.C., Herbun, G.W., 1981. Physical and biological structure andvariability in an upwelling center off Peru near 15�S during march 1977. In:Richards, F.A. (Ed.), Coastal Upwelling. American Geophysical Union,Washington, DC, pp. 473–495.
Bruland, K., 2006. A review of the chemistries of redox sensitive elements withinsuboxic zones of oxygen minimum regions. Gayana (Concepc.) 70(Supplement), 6–13.
Camayo, R., Campos, E.J.D., 2006. Application of wavelet transform in the study ofcoastal trapped waves off the west coast of South America. GeophysicalResearch Letters 33, L22601.
Carrit, D., Carpenter, J., 1966. Comparison and evaluation of currently employedmodification of the Winkler method for determining dissolved oxygen in seawater. Journal of Marine Research 24, 286–318.
Chavez, F.P., Barber, R.T., Soldi, H., 1984. Propagated temperature changes duringonset and recovery of the 1982–83 El Niño. Nature 309, 47–49.
Codispoti, L.A., Barber, R.T., Friederich, G.E., 1989. Do nitrogen transformations inthe poleward undercurrent off Peru and Chile have a globally significantinfluence? In: Neshyba, S.J., Mooers, N.K., Smith, R.L., Barber, R. (Eds.), PolewardFlows Along Eastern Ocean Boundaries. Coastal and Estuarine Studies. Springer-Verlag, pp. 281–310.
Cravatte, S., Picaut, J., Eldin, G., 2003. Second and first baroclinic Kelvin modes in theequatorial Pacific at intraseasonal timescales. Journal of Geophysical Research108, 3266–3286.
Dewitte, B., Purca, S., Illig S., Reanult, L., Giese, B., in press. Low frequencymodulation of intraseasonal equatorial Kelvin wave activity in the Pacific fromSODA:1958-2001. Journal of Climate.
Diaz, R.J., Rosenberg, R., 1995. Marine benthic hypoxia: a review of its ecologicaleffects and the behavioral responses of benthic macrofauna. Oceanography andMarine Biology Annual Review 33, 245–303.
Emery, W.J., Thomson, R.E., 1998. Data Analysis Methods in Physical Oceanography.Pergamon, Oxford.
Ferdelman, T.G., Lee, C., Pantoja, S., Harder, J., Bebout, B.B., Fossing, H., 1997. Sulfatereduction and methanogenesis in a Thioploca-dominated sediment off the coastof Chile. Geochimica et Cosmochimica Acta 61, 3065–3079.
Fossing, H., 1990. Sulfate reduction in shelf sediments in the upwelling region offcentral Peru. Continental Shelf Research 10 (4), 355–367.
Fossing, H., Gallardo, V.A., Jørgensen, B.B., Huettel, M., Nielsen, L.P., Schulz, H.,Canfield, D.E., Forster, S., Glud, R.N., Gundersen, J.K., Kuver, J., Ramsing, N.B.,Teske, A., Thamdrup, B., Ulloa, O., 1995. Concentration and transport of nitrateby the mat-forming sulphur bacterium Thioploca. Nature 374, 713–715.
Gallardo, V.A., 1977. Large benthic microbial communities in sulfide biota underPeru–Chile subsurface countercurrent. Nature 268, 331–332.
Gallardo, V.A., 1979. Peculiaridades bentónicas sublitorales del Pacífico sur oriental.In: Proceedings of the Workshop ‘‘Perspectivas en la investigación ecológicamarina en Chile (Pacífico sur oriental y Antártica)”. Universidad Austral deChile, Valdivia, August 23–25, 93 p.
Gallardo, V.A., 1985. Efectos del fenómeno de El Niño sobre el bentos sublitoralfrente a Concepción, Chile. In: Arntz, W., Landa, A., Tarazona, J. (Eds.). El Niño ysu impacto en la fauna marina, vol. Extraordinario. Boletin Instituto del Mar delPerú, pp. 79–885.
Gamenick, I., Jahn, A., Vopel, K., Giere, O., 1996. Hypoxia and sulphide as structuringfactors in a macrozoobenthic community on the Baltic Sea shore: colonizationstudies and tolerance experiments. Marine Ecology Progress Series 144, 73–85.
González, R.R., Quiñones, R.A., 2000. Pyruvate oxidoreductases involved inglycolytic anaerobic metabolism of polychaetes from the continental shelf offcentral-south Chile Estuarine. Coastal and Shelf Science 51, 507–519.
Graco, M., Farías, L., Molina, V., Gutiérrez, D., Nielsen, L.P., 2001. Massivedevelopments of microbial mats following phytoplankton blooms in anaturally eutrophic bay: implications for nitrogen cycling. Limnology andOceanography 46, 821–832.
Graco, M., Flores, G., Ledesma, J., Purca, S., Gutiérrez, D., Girón, M., 2006.Biogeochemical variability in the oxygen minimum zone of the upwellingHumboldt system off central Peru. In: Extended abstracts book, InternationalConference on the Humboldt Current system, 27 November–2 December, Lima,Peru, IMARPE, IRD and FAO, unpublished.
Gutiérrez, D., Gallardo, V.A., Levin, L., Quipúzcoa, L., Solís, J., 2002. Biomass andbioturbation responses of macrobenthos during El Niño 1997–1998 incontinental margin sediments off the southeastern Pacific. In: Salinas, S.,Urban, H.J., Arntz, W.E. (Eds.), Impacts of El Niño and Basin-Scale ClimateChange on Ecosystems and Living Marine Resources, vol. 30(1). InvestigacionesMarinas, Viña del Mar Chile, p. 144.
Gutiérrez, D., Quipúzcoa, L., Enríquez, E., 2006a. Oxygen deficiency and benthiccommunities in the Peruvian upper continental margin. Gayana (Concepc.) 70(Supplement), 29–36.
Gutiérrez, D., Aronés, K., Chang, F., Quipúzcoa, L., Villanueva, P., 2006b. Impacto dela variación oceanográfica estacional e inter-anual sobre los ensambles demicrofitoplancton, mesozooplancton, ictioplancton y macrozoobentos de dosáreas costeras del norte del Perú entre 1994 y 2002. Boletin Instituto del Mardel Perú. 22 (1–2).
Please cite this article in press as: Gutiérrez, D. et al., Oxygenation episonogr. (2008), doi:10.1016/j.pocean.2008.10.025
Hormazabal, S., Shaffer, G., Letelier, J., Ulloa, O., 2001. Local and remote forcing ofsea surface temperature in the coastal upwelling system off Chile. Journal ofGeophysical Research 106, 657–671.
Huettel, M., Forster, S., Klösser, S., Fossing, H., 1996. Vertical migration in sediment-dwelling sulfur bacteria Thioploca spp. in overcoming diffusion limitations.Applied and Environmental Microbiology 62, 1863–1872.
Jørgensen, B.B., Gallardo, V.A., 1999. Thioploca spp.: filamentous sulfur bacteria withnitrate vacuoles. FEMS Microbiology Ecology 128, 301–313.
Kessler, W.S., McPhaden, M.J., 1995a. Oceanic equatorial waves and the 1991–1993El Niño. Journal of Climate 8, 1757–1774.
Kessler, W.S., McPhaden, M.J., 1995b. Forcing of intraseasonal Kelvin waves in theequatorial Pacific. Journal of Geophysical Research 100 (10), 613–631.
Kessler, W.S., Kleeman, R., 2000. Rectification of Madden–Julian oscillation into theENSO cycle. Journal of Climate 13, 3560–3575.
Levin, L.A., Gutiérrez, D., Rathburn, A., Neira, C., Sellanes, J., Muñoz, P., Gallardo, V.A.,Salamanca, M., 2002. Benthic processes on the Peru margin: a transect acrossthe oxygen minimum zone during the 1997–98 El Niño. Progress inOceanography 53, 1–27.
Levin, L.A., 2003. Oxygen minimum zone benthos: adaptation and communityresponse to hypoxia. Oceanography and Marine Biology: An Annual Review 41,1–45.
Neira, C., Gad, G., Arroyo, N.L., Decraemer, W., 2001a. Glochinema bathyperuviensissp. n. (Nematoda, Epsilonematidae): a new species from Peruvian bathyalsediments, SE Pacific Ocean. Contributions to Zoology 70, 147–159.
Neira, C., Sellanes, J., Levin, L.A., Arntz, W.E., 2001b. Meiofaunal distributions on thePeru margin: relationship to oxygen and organic matter availability. Deep-SeaResearch I 48, 2453–2472.
NOAA, 2003. NOAA gets US consensus for El Niño/La Niña index, definitions. NOAAmagazine. september 30. WWW page, <http://www.magazine.noaa.gov/>.
Morales, C.E., Hormazabal, S.E., Blanco, J.L., 1999. Interannual variability in themesoscale distribution of the depth of the upper boundary of the oxygenminimum layer off northern Chile (18–241S): implications for the pelagicsystem and biogeochemical cycling. Journal of Marine Research 57, 909–932.
Morón, O., 2006. Monthly sea surface salinity by marsden squares off the Peruviancoast (1960–2004). In: Extended abstracts book, International conference onthe Humboldt Current system, 27 November–2 December, Lima, Peru. IMARPE,IRD and FAO, unpublished.
Mosquera, K., 2006. Numerical study of the response of the ocean to a northerlywind-jet in the equatorial eastern Pacific. In: Extended abstracts book,International Conference on the Humboldt Current System, 27 November–2December, Lima, Peru. IMARPE, IRD and FAO, unpublished.
Otte, S., Kuenen, J.G., Nielsen, L.P., Paerl, H.W., Zopfi, J., Schulz, H.N., Teske, A.,Strotmann, B., Gallardo, V.A., Jørgensen, B.B., 1999. Nitrogen, carbon andsulphur metabolism in natural Thioploca samples. Applied and EnvironmentalMicrobiology 65, 3148–3157.
Pennington, J.T., Mahoney, K.L., Kuwahara, V.S., Kolber, D., Calienes, R., Chavez, F.P.,2006. Primary production in the eastern tropical Pacific: a review. Progress inOceanography 69, 285–317.
Pizarro, O., Clarke, A.J., Van Gorder, S., 2001. El Niño sea level and current along theSouth American coast: comparison of observations with theory. Journal ofPhysical Oceanography 31, 1891–1903.
Rowe, G.T., Howarth, R., 1985. Early diagenesis of organic matter in sediments offthe coast of Peru. Deep-Sea Research 32, 43–55.
Salonen, K., Sarlava, J., Hakala, I., Viljanen, M.L., 1976. The relation of energy andorganic carbon in aquatic invertebrates. Limnology and Oceanography 21, 724–730.
Salzwedel, H., Flores, L., De Flores, E., Zafra, A., Carbajal, G., 1988. Macrozoobentosdel sublitoral peruano, antes, durante y después de El Niño 1982–1983. In:Salzwedel, H., Landa, A. (Eds.), Recursos y dinámica del ecosistema deafloramiento peruano, Vol. Extraordinario. Boletin Instituto del Mar del Perú(pp 77–98).
Sánchez, S., Delgado, E., 1996. Mareas rojas en el área de Callao (12�S), 1980–1995.Informe Progresivo Instituto del Mar del Perú (44), 19–37.
Sellanes, J., Neira, C., 2006. ENSO as a natural experiment to understandenvironmental control of meiofaunal community structure. Marine Ecology27, 31–43.
Sellanes, J., Quiroga, E., Neira, C., Gutiérrez, D., 2007. Changes of macrobenthoscomposition under different ENSO cycle conditions on the continental shelf offcentral Chile. Continental Shelf Research. doi:10.1016/j/csr.2007.01.001.
Schulz, H., Jørgensen, B.B., Fossing, H., Ramsing, R., 1996. Community structure offilamentous, sheath–building sulfur bacteria, Thioploca spp., off the coast ofChile. Applied and Environmental Microbiology 62 (6), 1855–1862.
Schulz, H., Strotmann, B., Gallardo, V.A., Jørgensen, B.B., 2000. Population study ofthe filamentous sulfur bacteria Thioploca spp. off the Bay of Concepción, Chile.Marine Ecology Progress Series 200, 117–126.
Simpson, E.H., 1949. Measurement of diversity. Nature 163, 688.Smayda, T.J., 2000. Ecological features of harmful algal blooms in coastal upwelling
ecosystems. South African Journal of Marine Science 22, 219–253.Strub, P.T., Mesías, J.M., Montecino, V., Rutllant, J., 1998. Coastal ocean circulation
off western South America. In: Robinson, A.R., Brink, K.H. (Eds.), The GlobalCoastal Ocean: Regional Studies and Syntheses, The Sea, vol. 11. John Wiley &Sons, New York, pp. 237–313.
Tarazona, J., 1990. Disturbance and stress associated to El Niño and theirsignificance for the macrobenthos of shallow areas of the Peruvian upwellingsystem. Ph.D. Thesis, Bremen Univ., Germany, 181 pp.
des on the continental shelf of central Peru: Remote ..., Prog. Ocea-

D. Gutiérrez et al. / Progress in Oceanography xxx (2008) xxx–xxx 13
ARTICLE IN PRESS
Tarazona, J., Arntz, W.E., 1986. Bacterias filamentosas gigantes en fondossomeros de la Bahía de Ancón. Revista de Ciencias UNMSM 74, 178–180.
Tarazona, J., Salzwedel, H., Arntz, W.E., 1988a. Oscillations of macrobenthos inshallow waters of the Peruvian central coast induced by El Niño 1982–83.Journal of Marine Research 46, 593–611.
Tarazona, J., Salzwedel, H., Arntz, W.E., 1988b. Positive effects of ‘‘El Niño” onmacrozoobenthos inhabiting hypoxic areas of the Peruvian upwelling system.Oecologia 76, 184–190.
Tarazona, J., Arntz, W., Valle, S., Peña, T., 2001. Los índices de El Niño y del impactosobre las comunidades bentónicas. In: Tarazona, J., Arntz, W., Castillo, E. (Eds.),El Niño en América Latina. Impactos biológicos y sociales. Consejo Nacional deCiencia y Tecnología, Lima , Perú, pp. 113–126.
Thamdrup, B., Canfield, E., 1996. Pathways of carbon oxidation in continentalmargin sediments off central Chile. Limnology and Oceanography 41 (8), 1629–1650.
Torrence, C., Compo, G.P., 1998. A practical guide to wavelet analysis. Bulletin of theAmerican Meteorological Society 79, 61–78.
Please cite this article in press as: Gutiérrez, D. et al., Oxygenation episonogr. (2008), doi:10.1016/j.pocean.2008.10.025
Tyson, R.V., Pearson, T.H., 1991. Modern and ancient continental shelf anoxia: anoverview. In: Tyson, R.V., Pearson, T.H. (Eds.), Modern and Ancient ContinentalShelf Anoxia. Geol. Soc. Spec. Publ. Geological Society of London (Vol. 58, pp. 1–24).
Ulloa, O., Escribano, R., Hormazabal, S., Quiñones, R., Gonzalez, R., Ramos, M., 2001.Evolution and biological effects of the 1997–98 El Niño in the upwellingecosystem off northern Chile. Geophysical Research Letters 28 (8), 1591–1594.
Wieser, W., 1960. Benthic studies in Buzzards Bay. II. The meiofauna. Limnology andOceanography 5, 121–137.
Wooster, W., Gilmartin, M., 1961. The Peru–Chile undercurrent. Journal of MarineResearch 19, 97–122.
Zafra, A., Salzwedel, H., Flores, L.A., 1988. Distribución y biomasa de bacteriasfilamentosas (Thioploca spp.) en las costas norte del Perú durante 1976–1985.In: Salzweedel, H., Landa, A. (Eds.), Recursos y dinámica del ecosistema deafloramiento peruano, vol. Extraordinario. Boletin Instituto del Mar del Perú,Callao, pp. 99–105.
Zuta, S., Guillén, O., 1970. Oceanografía de las aguas costeras del Perú. BoletinInstituto del Mar del Perú 2, 157–324.
des on the continental shelf of central Peru: Remote ..., Prog. Ocea-
Copyright © 2022 FDOKUMEN




















