
Ovipositional Behavior in the Context of Mass Rearing of Anopheles arabiensis
8
Journal of the American Mosquito Control Association, 26(4):365-372, 2010 Copyright (C) 2010 by The American Mosquito Control Association, Inc. 190365 OVIPOSITIONAL BEHAVIOR IN THE CONTEXT OF MASS REARING OF ANOPHELES ARABIENSIS FABRIZIO BALESTRINO, SHARON M. SOLIBAN, JIRIMIE GILLES, CLELIA OLIVA AND MARK Q. BENEDICT Entomology Unit, FA O/IAEA Agriculture & Biotechnology Laboratory, Joint FAO/IAEA Programme, International Atomic Energy Agency, PO Box 100, Wagramerstrasse 5, A-1400 Vienna, Austria ABSTRACT. Large-scale production of mosquitoes is a key factor for a successful sterile insect technique program. A manageable mass-production cage must contain appropriate features for adult resting, mating, feeding, and ovipositional activities. In order to maximize egg collections, tests were conducted to determine the physical characteristics of ovipositional sites for caged Anopheles arabiensis. Effects of texture, shade, height, and shape of the ovipositional container on female behavior were investigated. Results indicate a strong preference for oviposition on humid substrates over free-standing water. The shade and texture of the cup's walls also influenced site choice, with black rough inner vertical walls of the cup resulting in the largest number of eggs. Ovipositional sites with square shape were preferred rather than circular cups, and in the square cups, >60% of the eggs were laid in the corners. Height also affected oviposition, as An. arabiensis significantly favored the lowest sites even though some oviposition occurred at higher sites. Based on this study and from available literature, we determined the characteristics of an artificial ovipositional site that includes all these characteristics, which will yield large numbers of eggs required for mass production of this species. KEY WORDS Oviposition site characteristics, oviposition preference, mass rearing, sterile insect technique INTRODUCTION The sterile insect technique (SIT) is an insect pest control method based on the releases of massive numbers of sterile conspecific males, typically in an isolated target area. The SIT is usually part of a pest management strategy in an integrated approach over wide areas. The insects released should perform well in the field environ- ment, maintaining as much natural behavior as possible with high competitiveness for mating activities. The International Atomic Energy Agen- cy (IAEA) has long worldwide experience in supporting and developing SIT programs target- ing major insect pests of agricultural and veteri- nary importance. In 2004, the IAEA started a collaboration with the Tropical Medicine Re- search Institute in Khartoum, Sudan, in order to support their feasibility studies for the application of SIT against Anopheles arabiensis Patton along the Nile River in the Northern State. As part of this project, efforts are underway to develop mass rearing of mosquitoes for field release (Dyck et al. 2005, Benedict et al. 2009a), and the optimization of each step will bring the SIT program on An. arabiensis closer to operational status. The pro- duction of a large number of sterile males is one of the main technical obstacles in a mosquito SIT program and consumes the majority of time and cost. For mass rearing, a large production cage is needed, and one component is an oviposition system that is attractive to female mosquitoes for oviposition so that the efforts of production efficiently recover a maximum number of eggs. Such a system must also be amenable to simple construction, cleaning, and operation. Oviposition site choice in anopheline mosqui- toes is influenced by several factors: olfactory (Munga et al. 2006), tactile, temperature, chem- ical, and visual cues (Bentley and Day 1989, Otienoburu et al. 2007) are known to play a role in oviposition site selection. The visual cues of a potential breeding site are assessed by gravid females for reflectance, color, and contrast between the ovipositional sites and its immediate surroundings (Clements 1999). Studies conducted on Anopheles gambiae Giles s.1. showed that visual contrast strongly influenced the attractive- ness of sites and dark places stimulated oviposi- tion (McCrae 1984, Huang et al. 2005). Besides, anopheline eggs remain viable only on wet substrates (Clements 1999) and gravid females often choose a moist surface on which to oviposit their eggs in the laboratory and in the field (Olson and Meek 1977). In these experiments, the characteristics of oviposition sites were varied, with a view toward an egg collection device that not only elicited a strong ovipositional response that increased the net productivity of the mass-rearing cage but also met the requirements of predictability, manage- ability, and economy. We evaluated factors that influenced oviposition, including substrate mois- ture, shape, color, and height. MATERIALS AND METHODS Mosquito stocks and rearing methods Anopheles arabiensis Dongola strain (available from the Malaria Research and Reference Re- agent Resource Center, MR4, as MRA-856) was 365
Transcript of Ovipositional Behavior in the Context of Mass Rearing of Anopheles arabiensis

Journal of the American Mosquito Control Association, 26(4):365-372, 2010Copyright (C) 2010 by The American Mosquito Control Association, Inc.
190365
OVIPOSITIONAL BEHAVIOR IN THE CONTEXT OF MASS REARING OFANOPHELES ARABIENSIS
FABRIZIO BALESTRINO, SHARON M. SOLIBAN, JIRIMIE GILLES, CLELIA OLIVA ANDMARK Q. BENEDICT
Entomology Unit, FAO/IAEA Agriculture & Biotechnology Laboratory, Joint FAO/IAEA Programme, InternationalAtomic Energy Agency, PO Box 100, Wagramerstrasse 5, A-1400 Vienna, Austria
ABSTRACT. Large-scale production of mosquitoes is a key factor for a successful sterile insect techniqueprogram. A manageable mass-production cage must contain appropriate features for adult resting, mating,feeding, and ovipositional activities. In order to maximize egg collections, tests were conducted to determinethe physical characteristics of ovipositional sites for caged Anopheles arabiensis. Effects of texture, shade,height, and shape ofthe ovipositional container on female behavior were investigated. Results indicate a strongpreference for oviposition on humid substrates over free-standing water. The shade and texture of the cup'swalls also influenced site choice, with black rough inner vertical walls of the cup resulting in the largest numberof eggs. Ovipositional sites with square shape were preferred rather than circular cups, and in the square cups,>60% of the eggs were laid in the corners. Height also affected oviposition, as An. arabiensis significantlyfavored the lowest sites even though some oviposition occurred at higher sites. Based on this study and fromavailable literature, we determined the characteristics of an artificial ovipositional site that includes all thesecharacteristics, which will yield large numbers of eggs required for mass production of this species.
KEY WORDS Oviposition site characteristics, oviposition preference, mass rearing, sterile insect technique
INTRODUCTION
The sterile insect technique (SIT) is an insectpest control method based on the releases ofmassive numbers of sterile conspecific males,typically in an isolated target area. The SIT isusually part of a pest management strategy in anintegrated approach over wide areas. The insectsreleased should perform well in the field environ-ment, maintaining as much natural behavior aspossible with high competitiveness for matingactivities. The International Atomic Energy Agen-cy (IAEA) has long worldwide experience insupporting and developing SIT programs target-ing major insect pests of agricultural and veteri-nary importance. In 2004, the IAEA started acollaboration with the Tropical Medicine Re-search Institute in Khartoum, Sudan, in order tosupport their feasibility studies for the applicationof SIT against Anopheles arabiensis Patton alongthe Nile River in the Northern State. As part ofthis project, efforts are underway to develop massrearing of mosquitoes for field release (Dyck et al.2005, Benedict et al. 2009a), and the optimizationof each step will bring the SIT program on An.arabiensis closer to operational status. The pro-duction of a large number of sterile males is one ofthe main technical obstacles in a mosquito SITprogram and consumes the majority of time andcost. For mass rearing, a large production cage isneeded, and one component is an ovipositionsystem that is attractive to female mosquitoes foroviposition so that the efforts of productionefficiently recover a maximum number of eggs.Such a system must also be amenable to simpleconstruction, cleaning, and operation.
Oviposition site choice in anopheline mosqui-toes is influenced by several factors: olfactory(Munga et al. 2006), tactile, temperature, chem-ical, and visual cues (Bentley and Day 1989,Otienoburu et al. 2007) are known to play a rolein oviposition site selection. The visual cues of apotential breeding site are assessed by gravidfemales for reflectance, color, and contrastbetween the ovipositional sites and its immediatesurroundings (Clements 1999). Studies conductedon Anopheles gambiae Giles s.1. showed thatvisual contrast strongly influenced the attractive-ness of sites and dark places stimulated oviposi-tion (McCrae 1984, Huang et al. 2005). Besides,anopheline eggs remain viable only on wetsubstrates (Clements 1999) and gravid femalesoften choose a moist surface on which to oviposittheir eggs in the laboratory and in the field (Olsonand Meek 1977).
In these experiments, the characteristics ofoviposition sites were varied, with a view towardan egg collection device that not only elicited a
strong ovipositional response that increased thenet productivity of the mass-rearing cage but alsomet the requirements of predictability, manage-ability, and economy. We evaluated factors thatinfluenced oviposition, including substrate mois-ture, shape, color, and height.
MATERIALS AND METHODS
Mosquito stocks and rearing methods
Anopheles arabiensis Dongola strain (availablefrom the Malaria Research and Reference Re-agent Resource Center, MR4, as MRA-856) was
365

366 JOURNAL OF THE AMERICAN MOSQUITO CONTROL ASSOCIATION VOL. 26, No. 4
used in all experiments. The strain originatedA
from the Northern State of Sudan and has beenin the colony of the IAEA Laboratories since2005. Mosquitoes were reared in a climate-controlled room operating at 27 __ IC, 60 ___10% RH with a 12:12 h (light:dark) photocyclethat included periods of dusk (1 h)and dawn _
,.(1 h). Larvae were reared in white Acrylonitrile- |Butadiene-Styrene plastic trays (41 30 8 cm)containing liter of deionized water and fed withpowdered koi food (Floating Blend(R); Aquari-Care, Victor, NY), powdered and sieved througha 224-micron mesh. Pupae were collected andplaced in white plastic cups and placed inside thecages for emergence. Adults were held in 30 30
30-cm white polypropylene cages (BugDorm E .G H
1(R); MegaView, Taichung, Taiwan) with constant
access to 10% (w/v) sucrose solution containing0.2% (w/v)methylparaben (CAS 99-76-3; Sigma,St. Louis, MO; Benedict et al. 2009b). Defibrin-ated bovine blood was provided twice weekly forapproximately 15 min, each feeding using a water-heated aluminum plate covered with parafilm"M" (Pechney Plastic Packaging, Menasha, WI).
B, C D
General experimental conditions
Fig. 1. Materials and cups used in experiments: (A)circular translucent plastic cup without lining; (B)circular cup lined with black and white fabric; (C)circular cup w,ith black fabric lining; (D) square cupwith black fabric lining; (E) grid to quantify the eggs onsquare and circular cups; (F) flat rectangular plate; (G)rectangular plate with ridge in the middle covered inblack fabric lining; (H) rectangular plate with closedlong sides covered in black fabric lining; and (I)rectangular plate with closed short and long sidescovered in fabric lining.
On the 4th day following emergence, femalemosquitoes that had been caged with males at lowdensity (---500 adults per cage, 1:1 ratio) wereoffered a blood meal. Approximately 24 h afterfeeding, 50 bloodfed females were transferred to
new cages with an equal number of males fromthe same cohort using a mouth aspirator.
All circular oviposition cups were made oftranslucent polypropylene, 10.5 cm diam and4.5 cm deep (Fig. 1A; manufacturer not identifi-able) and are a common type used for foodstorage. A moist oviposition substrate inside cupswas made from a sponge cloth (Vileda(R), Wein-heim, Ge..--rnany) overlaid with white cellulosefilter paper (Cat No. 1001 090, 597; Whatman(R),Maidstone, United Kingdom). Prior to use, thesponge cloths were soaked in water until theywere fully saturated. In experiments where thewalls of the oviposition sites were covered, awaterproof roofing fabric (Novoflor Top 1, Linz,Austria), hereafter referred to as fabric, was used.In all experiments, the first egg collection wasdone 2 days after bloodfeeding beginning in mid-to late afternoon; egging dishes were removedaround 9:00 a.m. on day 3. The eggs werecounted under a dissection microscope.
Standing water versus moist substrate
Two circular cups (Fig. 1A), one with standingwater and the other with a moist ovipositionsubstrate, were introduced simultaneously intoeach cage. The 2 cups were placed in the center ofthe cage 5 cm apart. The cup with standing water
was created by adding 40 ml of deionized water tothe cup. The amount of water in this cup wasdetermined in relation to the thickness of the wetsubstrate presented in the other cup: 40 ml wasneeded to achieve the same distance between therims and the ovipositional surfaces in both cupsand was found to be adequate to maintain a layerof water throughout the entire duration of thetest. To make the appearance of the 2 oviposi-tional sites similar, a disk of white cellulose filterpaper was attached to the bottom of the cupcontaining standing water. In this experiment, 3replicates were performed.
Effects of shade and texture
Circular cups (Fig. 1A) were used in thisexperiment and modified in such a way that eachcup placed in a cage was divided into 2 equalsections (Fig. 1B). Each section of the cup wasdefined by the characteristics of the inner verticalwalls: black and white fabric was glued inside tocreate 3 cups with "black-rough" and "white-rough" sections. Black and white fabric was gluedoutside to create 3 cups with "black-smooth" and"white-smooth" sections. One cup was introducedinto each of 6 cages. All cups used were providedwith a moist substrate as described above.
Influence of oviposition cup heightIn these trials, circular oviposition cups
(Fig. C) whose inner walls were lined with black

DECEMBER 2010 OvIr,OSIa'IOy IN MASS REARING OF ANOPHELES ARABIENSIS 367
fabric and had moist ovipositional substrates wereused. Two types of experiments were performed.First, in a choice assay, females were given 4ovipositional containers in a cage: 1) beneath, sothat only the mouth of the cup was exposed to theinside of the cage; 2) on the floor; 3) elevated to10 cm; and 4) at 20 cm--the cups at 10 and 20 cmwere supported using transparent Plexiglas (acryl-ic) cylinders. Four cages were tested simulta-neously so that each height was replicated 4 times.Then, in a no-choice assay, a single cup was
provided in each cage at of the 4 heights testedpreviously. Each height was replicated 4 times.After the 1st egg collection (2 days after feeding),a new cup was offered to each cage (3 days afterfeeding). This time, all cups were placed on thefloor in the center of the cages. The cups wereremoved the following morning (4 days afterfeeding) and the size of the second egg batcheswas determined.
Square versus circular cup
A circular oviposition cup (Fig. 1C) wascompared with a square cup (10.5 cm wide,4.5 cm deep; Fig. 1D) in a choice assay. Both cuptypes had inner walls lined with black fabric andmoist substrates as described above. The cupswere equally wide (10.5 cm) but differed inperimeter wall length and in substrate area. Eachof the 4 replicates contained 250 bloodfed femalesand 250 males. The square and the circular cupswere introduced into the centers of the cages 5 cmapart 2 days after feeding and were recovered thefollowing day. To count the eggs, a griddedtransparent plastic sheet (2.5 2.5-cm squares;Fig. 1E) was laid over the filter paper and thenumber of eggs present in each cell of the grid wasrecorded. The cells were categorized as follows:C--corner cells, B--border cells, and M--middlecells. For the purpose of standardization in thechoice assay, the outcome was expressed in termsof the number of eggs per square centimeter ofthe substrate.
Effect of various container shapesThree variants of an initial oviposition tray
design (Fig. IF) were created and described asfollows: 1) open tray with a wall in the middlecovered in black fabric (Fig. 1G); 2) semi-opentray (U-shaped) with the inner sides of the longwalls lined with black fabric while the short endswere left open (Fig. 1H); and 3) closed tray withthe inner side of all 4 walls covered with blackfabric (Fig. I).
All tray models created were 20 cm long and4 cm wide. All the vertical walls were 4.5 cm high.Each tray was compared with a circular ovipo-sition cup with black fabric (Fig. 1C) in choice-test experiments. Cups and trays were set with
1000
900
800
%" 700
, 6oo
- 500
400
300
200
100
Fig. 2.
Rough cup Smooth cup
Mean number (_ SEM) of eggs laid asfunction of the texture (rough or smooth) and color(black or white)of the cup (* P < 0.05; P < 0.01).
moist substrates and placed in the center of thecage 5 cm apart. Six choice-test replications werecarried out for each tray model.
Statistical analysisAll analyses were conducted using Minitab
(Minitab(R) Inc., State College, PA). The effect ofphysical characteristics of the ovipositional siteon the number of laid eggs was determined using1-way analysis of variance (ANOVA) except inExperiment 4 where a 2-way ANOVA wasperformed. Pair-wise mean comparisons weredone with the Tukey method. Data were trans-formed (log x + 1).
RESULTS
Standing water versus moist substrate
A very strong effect of the oviposition con-tainer's horizontal surface was observed. Gravidfemales laid significantly more eggs (F 35.76;df 1; P < 0.05) on the moist substrate (spongecloth overlaid with white filter paper, mean +/-SEM, 3,544 +/- 680) than on standing water (mean+/- SEM, 471 + 107).
Effects of shade and texture
The number of eggs laid by the females differedsignificantly according to the shade (F 22.25;df 1; P < 0.001) and the texture (F 13.92;df 1; P < 0.005) of the inner wall of the cup(Fig. 2). Rough texture and black color were
more attractive to ovipositing females (t -3.73;P < 0.005 and -4.72; P < 0.001,
respectively). No interaction between the effectsof shade (black or t'hite) and texture (rough or
smooth) of the cup was observed.
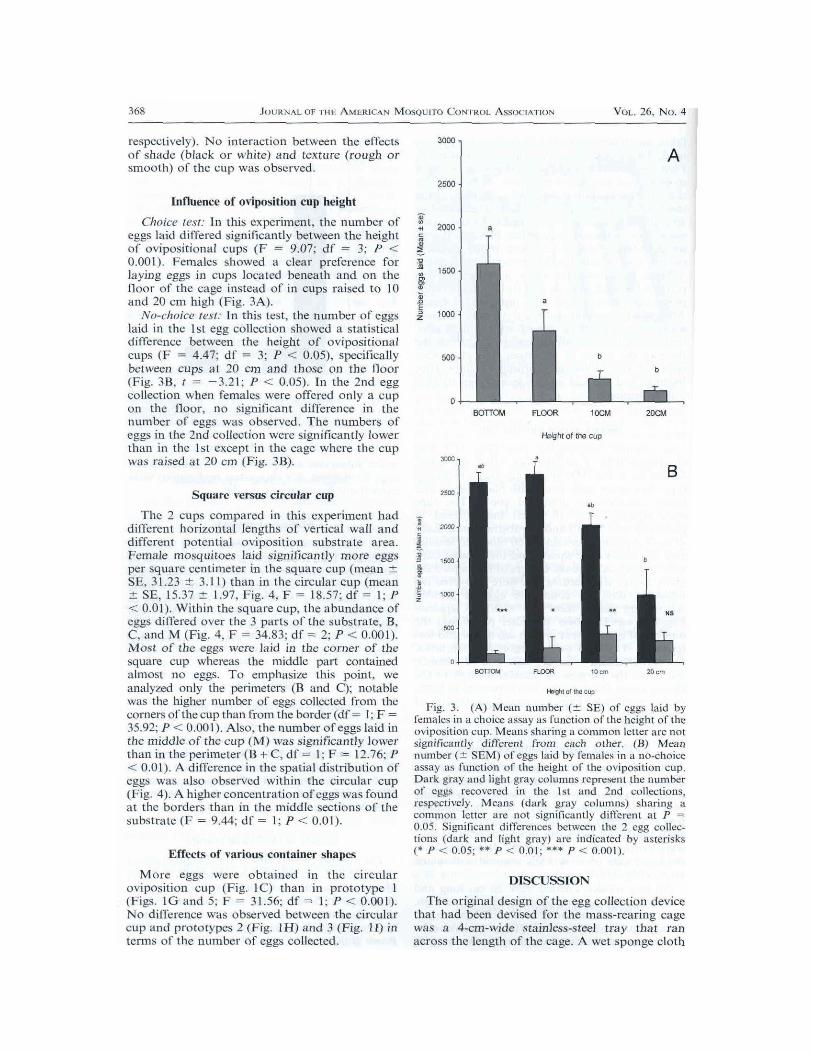
Influence of oviposition cup heightChoice test: In this experiment, the number of
eggs laid differed significantly between the heightof ovipositional caps (F 9.07: df 3; P <
0.001). Females showed a clear prclbrence forlaying eggs in cups located beneath and on thefloor of the cage instead of in cups raised to 10and 20 cm high (Fig. 3A).
No-choice test: In this test, the number of eggslaid in the 1st egg collection showed a statisticaldifference between the height of ovipositionalcups (F 4.47: df 3: P < 0.05), specificallybetween cups al 20 cm and those on the floor(Fig. 3B, -3.21; P < 0.05). In the 2nd eggcollection when females were offered only a ctlpon the floor, no significant difference in thenumber of eggs was observed. The numbers of
eggs in the 2nd collection were significantly lowerthan in the 1st except in the cage where the cupwas raised at 20 cm (Fig. 3B).
The 2 cups compared in this experiment haddiflbrent horizontal lengths of vertical wall and
Female mosquitoes laid significantly more eggs
SE, 31.23 +/- 3.11) than in the circular cup (mean_+ SE, 15.37 +_ 1.97, Fig. 4, F 18.57; df 1; P< 0.01). Within the square cup, the abundance of
C, and M (Fig. 4, F 34.83: df 2: P < 0.001).Most of the eggs were laid in the corner of thesquare cup whereas the middle part containedalmost no eggs. To emphasize this point, we
analyzed only the perimeters (B and C); notablewas the higher number of eggs collected from the
corners ofthe cup than from the border (dr- l: F35.92; P < 0.001 ). Also, tile number of eggs laid inthe middle of the cup (M) vas significantly lowerthan in the perimeter (B + C, df F 12.76; P< 0,01). A difference in the spatial distribution ofeggs was also observed within the circular cup(Fig. 4). A higher concentration ofeggs was foundat the borders than in the middle sections of thesubstratc (F 9.44; df I; P < 0.01).
Effects of various container shapes
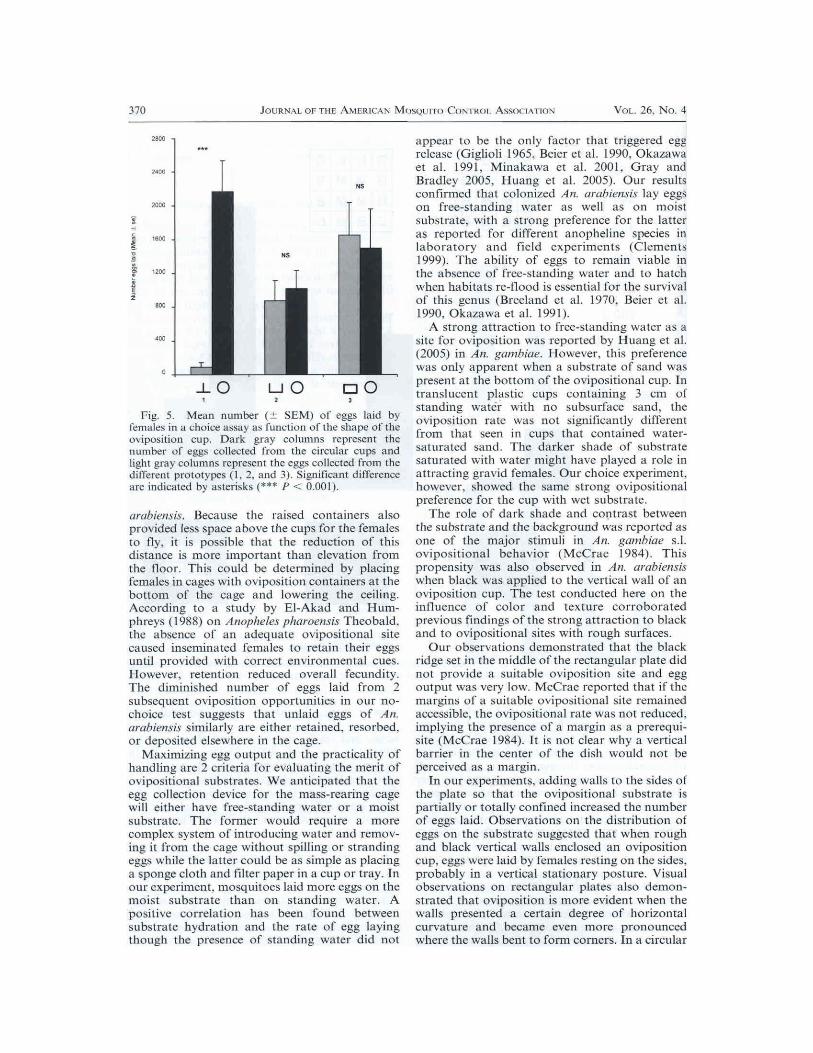
More eggs were obtained in the circularoviposition cup (Fig. IC) than in prototype(Figs. IG and 5; F 31.56; df 1; P < 0.001).No difference was observed between the circalarcup and prototypes 2 (/-'ig. IH) and 3 (Fig, 11) interms of the number of eggs collected.
A
Fig. 3. (A) Mean number (+/- SE) of eggs laid byt?malcs in choice assay function of the height of theoviposition cup. Meatls sharing colllllaon letter are notsignificantly different From each other. B) Meannmnber (-- SEM) of eggs laid by l?rnalcs in no-choiceassay function of the height of the oviposition cup.Dat gray and light gray columns represent the numberof eggs recovered in the Ist and 2nd collections.respeclively. Meas (dark gray columns) sharingcommon letter not significantly different at P0.05. Signil]callt differences between the 2 egg collec-tions (dark and light gray) indicated by asterisks
DISCUSSION
The original design of the egg collection devicethat had been devised for the mass-rearing cagewas a 4-cm-wide stainless-steel tray that ran
across the length of the cage. A wet sponge cloth

600
550
500
450
400
350
300
250
200
150
100
50
0
BMM
Location within the cup
Fig. 4. Mean number (+/- SEM} of eggs laid by females according to different locations inside the }vipositioncups. Significant difference between black and gray indicated by asterisks (** P < 0.05: P < 0.001}.
covered with a strip of white cellulose filter paperwas laid over the tray for egg collection. The traywas pushed in and drawn out through a narrow
aperture in the short side of the cage. Preliminarytrials coaducted ia the mass-rearing cage as well
as in the small cage consistently resulted in verypoor oviposition. Because our immediate purposefor these trials was to create a design for a mass-
rearing cage containing ovipositional sites thatwould collect the largest numbers of eggs, variousparameters wcrc tested that nught have affectedthe original design's poor performance. Results ofexperiments where the heights of the cups were
tested showed that the artilicial environment andcolonization did not alter the natural preferenceof An. arahiensis for ovipositional sites near theground. In nature, the types of habitats thatharbor high densities of larwe are slow-moving
streams (especially stream edges and beds) andtransitory habitats such as puddles, wateringholes, and tracks on the ground (Robert et al,1998, Gimnig et al. 2001. Shililu ct al. 2003).Exceptions have been observed in An. ,gambiae s,1.that was found occupying habitats in holes ofllamboyant trees (Delot*iv regiu (Bojer ex Hook.)RaE) up to 212 cm high where Aedes larvae are
typically foand (OmIin et al. 2007}. The authorsattributed the unusual presence of Anopheles inthe tree holes to the absentee of appropriatelocations in which to lay eggs that forced themosquitoes to resort to otherwise unsuitableplaces.
In our experiments, the mean numbers ol'eggscollected at different heights in the choice testsuggested a negative correlation between sub-
strate elevation and ovipositional output in
..LO uO
Fig. 5. Mean number (+ SEM) of eggs laid byfemales in choice assay function of the shape of theoviposition cup, Dark gray colunms represent thenumber of eggs collected from the circular cups andlight gray columns represent the eggs collected from thedifferent prototypes I. 2, and 3). Significant difference
indicated by asterisks (*** P < 0.0011.
provided less space above the cups for the females
to fly, it is possible that the reduction of thisdistance is more important than elevation fromthe floor. This could be determined by placingfemales in cages with oviposition containers at thebottom of the cage and lowering the ceiling.According to a study by EI-Akad and Hum-phreys (1988) on Anopheles pharoensis Theobald.the absence of an adeqaate ovipositional sitecaused inseminated lbmales to retain lheir eggsuntil provided with correct environmental cues,
However, retention reduced overall fecundity.The diminished number of eggs laid from 2
choice lest suggests thai unlaid eggs of An.arabiensis similarly are either retained, resorbed,
or deposited elsewhere in the cage.Maximizing egg output and the practicality of
handling are 2 criteria for evaluating the merit ofovipositionul substrates. We anticipated that the
egg collection device for the mass-rearing cagewill either have free-standing water or a moisl
ing it from the cage without spilling or strandingeggs while the latter could be as simple as placinga sponge cinth and filter paper in a cup or tray. In
our experiment, mosquitoes laid more eggs on the
positive correlalion has been found betweensubslrate hydration and the rate of egg layingthough the presence of standing water did not
appear to be the only factor that triggered eggrelease (Giglioli 1965, Beier et al. 1990, Okazawaet al. 1991, Minakawa ctal. 2001, Gray andBradley 2005, Huang el al. 2005), Our resultsconfirmed that colonized An. arabiensi lay eggson free-standing water as well as on moistsubstrate, with a strong preference for the latter
as reported for different anopheline species inlaboratory and field experiments (Clcmcnts1999). The ability of eggs to reluain viable inthe absence of frcc-standing water and to hatchwhen habitats re-flood is essential for the survivalof this genus (Breeland et al. 1970, Beier et al.i990. Okazawa et al. 19911.A strong attraction to free-standing water as a
site for oviposition was reported by tfuang et al.(2005) in An. gambiae. However, this preferencewas only apparent when a substrate of sand was
present at the bottom of the ovipositional cup. Intranslucent plastic cups containing 3 cm ofsttmding watdr with no subsurface sand. lheoviposition rate was not significantly differentfrom that seen in cups that contained water-
saturated sand. The darker shade of substratesaturated with water might have played a role inattracting gravid females. Our choice expefnnent.however, showed the same strong ovipositionalpreference for the cup with wet substrate.The role of dark shade and co0trast between
tile substtate and the background was reported as
one of the major stimuli in An. gambiae s.l.ovipositional behavior (McCrac 1984). Thispropensity was also observed in An. arabiensiswhen black was applied to the vertical wall of anoviposition cup. The test condncted here on theinflaence of color and texture corroboratedprevious findings of the stroag attraction to blackand to ovipositionaI sites with rough surfaces.Onr observations demonstrated that the black
ridge set in the middle of the rectaagular plate didnot provide a suitable oviposition site and eggoutput was very low. McCrae reported that if themargins of a suitable ovipositional site remainedaccessible, the ovipositional rate was not reduced,implying the presence of a margin as a prerequi-site (McCrae 1984). It is not clear why a verticalbarrier in the center of the dish would not beperceived as a margin.
In our experiments, adding walls to the sides ofthe plate so that the ovipositional substrate ispartially or totally confined increased the numberof eggs laid. Observations on the distribution of
eggs on the substrate suggested that when roughand black vertical wails enclosed an ovipositioncup. eggs were laid by females resting on the sides,probably in a vertical stationary poslnre. Visualobservations on rectangular plates also demon-strated that oviposition is more evident when thewalls presented a certain degree of horizontalcurvature and became even more pronouncedwhere the walls bent to form corners. In a circular

DECEMBER 2010 OVIPOSITION IN MASS REARING OF ANOPHELES ARABIENSIS 371
cup, with its constant curvature, the eggs formed acontinuous mound along the entire perimeterwithout any further discreet aggregation. In arectangular tray, a straight black wall did notappear conducive to resting, and oviposition fromthe wall was not evident. The small number ofeggs released in the open prototypes (Figs. 1G,1H) were not distributed in any apparent patternon the substrate. When the rectangular plate wassurrounded by black walls on all 4 sides, it wasapparent that the corner was the most attractivelocation for oviposition (data not shown).The propensity to lay eggs in corners of square
dishes was detectable when mosquitoes were keptin high density (500 mosquitoes per cage, sexratio 1:1, cage dimension 30 30 30 cm)butwas not as pronounced when there were only 50mosquitoes in the cage (data not shown). Furtherstudies are needed to confirm this preliminaryobservation, but one might speculate that thereare oviposition attractants when a number abovesome threshold is reached. Alternatively, ovipo-sition dish crowding and adult irritation at highdensity might affect behavior differently thanwhen adults are kept at a low density.
In the pre-oviposition behavior model de-scribed for different anopheline species, femaleslocate an ovipositional site by flying at differentheights above the ground, changing their flightpattern as they approach a target (Clements 1999).After testing the ovipositional site, they begin tohover over its immediate vicinity, intermittentlydipping to touch the substrate. Even whilehovering some distance above the water surface,the females were observed to strictly keep withinthe margins of the ovipositional site (Clements1999), suggesting once again the important roleplayed by the presence of a margin. The absenceof a vertical wall (Fig. 1F) or its position(Fig. 1G) could be the reason for the poor eggcollections in open trays. In addition, the phase ofinspection performed by gravid females inside andaround the oviposition area is most likelycompleted earlier in the closed square cup due tothe presence of differential geometry that inducesthe gravid females to rest and lay eggs. This mightalso explain the aggregates of eggs in the cornersof the cups. In small trials that we conducted, veryfew eggs were recovered when a white screen wasplaced a few millimeters above the moist whitesubstrate in circular cups lined with black fabric(data not shown). It would appear that when thecontact with the substrate is obstructed, or whenthe appearance of the substrate is altered, theoviposition process is strongly affected. Theabsence of eggs in this cup is consistent withobservations that oviposition in a stationaryposition happens after the female appraises thesite by direct contact with the substrate. Ovipo-sition from a stationary posture was identified as a
response to an optimal oviposition stimulus;
indeed, females that lay their eggs while sittingtend to release all their eggs simultaneously(McCrae 1984).
This behavior could be exploited in massrearing where eggs need to be collected at apredetermined time. If, with the right set of cues,the females can be made to lay their eggssynchronously, the eggs can be collected andprocessed further with minimal time and effort.Based on our observations, for mass productionit is anticipated that the egg cups will have a moistsubstrate enclosed by black and rough-texturedwalls that are approximately 4.5 cm high and willbe located on the cage floor. In the earlierprototypes of the mass-rearing cage, the eggcollection device was a flat open rectangular trayat 10 cm high to allow free space at the bottom ofthe cage where maintenance work would be done.It was not anticipated that this height wouldnegatively affect oviposition, which emphasizesthat experimental observations on the basicbehavior of mosquitoes are necessary to success-fully develop mass rearing.
REFERENCES CITED
Beier JC, Copeland R, Oyaro C, Masinya A, OdagoWO, Odour S, Koech DK, Roberts CR. 1990.Anopheles gambiae complex egg-stage survival indry soil from larval development sites in westernKenya. J Am Mosq Control Assoc 6:105-109.
Benedict MQ, Knols BGJ, Bossin HC, Howell PI,Mialhe E, Caceres C, Robinson AS. 2009a. Coloni-zation and mass rearing: learning from others.Malar J 8:$4.
Benedict MQ, Hood-Nowotny RC, Howell PI, WilkinsEE. 2009b. Methylparaben in Anopheles gambiae s.l.sugar meals increases longevity and malaria oocystabundance but is not a preferred diet. J Insect Physiol55:197-204.
Bentley MD, Day JF. 1989. Chemical ecology andbehavioral aspects of mosquito oviposition. Annu RevEntomol 34:401-421.
Breeland SG, Pletsch DJ, Quarterman KD. 1970.Laboratory studies on the suitability of mud as anoviposition substrate for Anopheles albimanus Wie-demann. Mosq News 30:81-88.
Clements AN. 1999. The biology of mosquitoes.Volume 2. London, United Kingdom: CABI.
Dyck VA, Hendrichs J, Robinson AS. 2005. The sterileinsect technique." principles and practice in area-wideintegratedpest management. Dordrecht, Netherlands:Springer.
E1-Akad AS, Humphreys JG. 1988. Factors affectingoviposition and egg production in laboratory-rearedAnopheles pharoensis Theobald. Bull Soc Vector Ecol13:243-247.
Giglioli MEC. 1965. Oviposition by Anopheles melasand its effect on egg survival during the dry season inthe Gambia, West Africa. Ann Entomol Soc Am58:885-891.
Gimnig JE, Ombok M, Kamau L, Hawley WA. 2001.Characteristics of larval anopheline (Diptera: Culic-idae) habitats in western Kenya. J Med Entomol38:282-288.

372 JOURNAL OF THE AMERICAN MOSQUITO CONTROL ASSOCIATION VOL. 26, No. 4
Gray EM, Bradley TJ. 2005. Physiology of desiccationresistance in Anopheles gambiae and Anophelesarabiensis. Am J Trop Med Hyg 73:553-559.
Huang J, Walker ED, Giroux PY, Vulule J, Miller JR.2005. Ovipositional site selection by Anophelesgambiae: influences of substrate moisture and texture.Med Vet Entomol 19:442-450.
McCrae AWR. 1984. Oviposition by African malariavector mosquitoes. II. Effects of site tone, water typeand conspecific immatures on target selection byfreshwater Anopheles gambiae Giles, sensu lato. AnnTrop Med Parasitol 78:307-318.
Minakawa N, Githure JI, Beier JC, Yan G. 2001.Anopheline mosquito survival strategies during thedry period in western Kenya. J Meal Entomol38:388-392.
Munga S, Minakawa N, Zhou G, Barrack O-OJ,Githeko AK, Yan G. 2006. Effects of larvalcompetitors and predators on oviposition site selec-tion of Anopheles gambiae sensu stricto. J MedEntomol 43:221-224.
Okazawa T, Vaisui L, Suzuki H, Kere N. 1991.Resistance of Anopheles farauti eggs to desiccation.Jpn J Sanit Zool 42:65-67.
Olson JK, Meek CL. 1977. Soil moisture conditionsthat are most attractive to ovipositing females ofPsorophora columbiae in Texas rice lands. Mosq News37:19-26.
Omlin FX, Carlson JC, Ogbunugafor CB, Hassanali A.2007. Anopheles gambiae exploits the treehole eco-
system in western Kenya: a new urban malaria risk?Am J Trop Med Hyg 77:264-269.
Otienoburu PE, Bayoh N, Gimnig J, Huang J, WalkerED, Otieno MF, Vulule J, Miller JR. 2007. Anophelesgambiae (Diptera: Culicidae) oviposition as influ-enced by type of water infused into black and redsoils of western Kenya. Int J Trop Insect Sci 27:2-5.
Robert V, Awono-Ambene HP, Thioulouse J. 1998.Ecology of larval mosquitoes, with special referenceto Anopheles arabiensis (Diptera: Culicidae) inmarket-garden wells in urban Dakar, Senegal. J MedEntomol 35:948-955.
Shililu J, Ghebremeskel T, Seulu F, Mengistu S,Fakadu H, Zerom M, Ghebregziabiher A, SintasathD, Bretas G, Mbogo C, Githure J, Brantly E, NovakR, Beier JC. 2003. Larval habitat diversity andecology oftnopheline larvae in Eritrea. J MedEntomol 40:921-929.
















![Rearing Healthy Calves Manual 2nd ed (1)[2] copy](https://static.fdokumen.com/doc/165x107/6326a762051fac18490ddddd/rearing-healthy-calves-manual-2nd-ed-12-copy.jpg)



