
Ovine bone marrow mesenchymal stem cells: isolation and characterization of the cells and their...
11
Ovine bone marrow mesenchymal stem cells: isolation and characterization of the cells and their osteogenic differentiation potential on embroidered and surface-modified polycaprolactone-co-lactide scaffolds C. Rentsch & R. Hess & B. Rentsch & A. Hofmann & S. Manthey & D. Scharnweber & A. Biewener & H. Zwipp Received: 30 October 2009 / Accepted: 31 March 2010 / Published online: 20 May 2010 / Editor: J. Denry Sato # The Society for In Vitro Biology 2010 Abstract The current study was undertaken with the goal being isolation, cultivation, and characterization of ovine mesenchymal stem cells (oMSC). Furthermore, the objective was to determine whether biological active polycaprolactone-co-lactide (trade name PCL) scaffolds support the growth and differentiation of oMSC in vitro. The oMSC were isolated from the iliac crest of six merino sheep. Three factors were used to demonstrate the MSC properties of the isolated cells in detail. (1) Their ability to proliferate in culture with a spindle-shaped morpho- logy, (2) presence of specific surface marker proteins, and (3) their capacity to differentiate into the three classical mesenchymal pathways, osteoblastic, adipogenic, and chondrogenic lineages. Furthermore, embroidered PCL scaffolds were coated with collagen I (coll I) and chondroitin sulfate (CS). The porous structure of the scaffolds and the coating with coll I/CS allowed the oMSC to adhere, proliferate, and to migrate into the scaffolds. The coll I/CS coating on the PCL scaffolds induced osteogenic differentiation of hMSC, without differ- entiation supplements, indicating that the scaffold also has an osteoinductive character. In conclusion, the isolated cells from the ovine bone marrow have similar morphologic, immunophenotypic, and functional characteristics as their human counterparts. These cells were also found to differentiate into multiple mesenchymal cell types. This study demonstrates that embroidered PCL scaffolds can act as a temporary matrix for cell migration, proliferation, and differentiation of oMSC. The data presented will provide a reliable model system to assess the translation of MSC-based therapy into a variety of valuable ovine experimental models under autologous settings. Keywords Ovine mesenchymal stem cells . Bone marrow . Multilineage differentiation . Polycaprolactone-co-lactide scaffold . Tissue engineering Introduction The reconstruction of post-traumatic or tumor-related large bone defects is still a significant clinical problem. To overcome the disadvantages linked with autografts or allografts, numerous bone tissue engineering strategies have been developed to create functional and structural tissue alternatives (Hollinger et al. 1996; Li et al. 2005; Rentsch et al. 2008). The concept of tissue engineering incorporates the combination of living cells, biologically active molecules, and a structural scaffold to promote the repair and regeneration of tissue (Hutmacher et al. 2007). C. Rentsch & R. Hess contributed equally to this work. C. Rentsch (*) : S. Manthey : A. Biewener : H. Zwipp Department of Trauma and Reconstructive Surgery, University Hospital Carl Gustav Carus Dresden, Fetscher Straße 74, 01307 Dresden, Germany e-mail: [email protected] R. Hess (*) : A. Hofmann : D. Scharnweber Max Bergmann Center of Biomaterials, Budapester Straße 27, 01069 Dresden, Germany e-mail: [email protected] B. Rentsch Catgut GmbH, Gewerbepark 18, 08258 Markneukirchen, Germany In Vitro Cell.Dev.Biol.—Animal (2010) 46:624–634 DOI 10.1007/s11626-010-9316-0
-
Upload
uniklinikum-dresden -
Category
Documents
-
view
1 -
download
0
Transcript of Ovine bone marrow mesenchymal stem cells: isolation and characterization of the cells and their...

Ovine bone marrow mesenchymal stem cells: isolationand characterization of the cells and their osteogenicdifferentiation potential on embroidered and surface-modifiedpolycaprolactone-co-lactide scaffolds
C. Rentsch & R. Hess & B. Rentsch & A. Hofmann &
S. Manthey & D. Scharnweber & A. Biewener & H. Zwipp
Received: 30 October 2009 /Accepted: 31 March 2010 /Published online: 20 May 2010 / Editor: J. Denry Sato# The Society for In Vitro Biology 2010
Abstract The current study was undertaken with the goalbeing isolation, cultivation, and characterization of ovinemesenchymal stem cells (oMSC). Furthermore, theobjective was to determine whether biological activepolycaprolactone-co-lactide (trade name PCL) scaffoldssupport the growth and differentiation of oMSC in vitro.The oMSC were isolated from the iliac crest of six merinosheep. Three factors were used to demonstrate the MSCproperties of the isolated cells in detail. (1) Their abilityto proliferate in culture with a spindle-shaped morpho-logy, (2) presence of specific surface marker proteins, and(3) their capacity to differentiate into the three classicalmesenchymal pathways, osteoblastic, adipogenic, andchondrogenic lineages. Furthermore, embroidered PCLscaffolds were coated with collagen I (coll I) andchondroitin sulfate (CS). The porous structure of thescaffolds and the coating with coll I/CS allowed
the oMSC to adhere, proliferate, and to migrate into thescaffolds. The coll I/CS coating on the PCL scaffoldsinduced osteogenic differentiation of hMSC, without differ-entiation supplements, indicating that the scaffold also has anosteoinductive character. In conclusion, the isolated cellsfrom the ovine bone marrow have similar morphologic,immunophenotypic, and functional characteristics as theirhuman counterparts. These cells were also found todifferentiate into multiple mesenchymal cell types. Thisstudy demonstrates that embroidered PCL scaffolds can actas a temporary matrix for cell migration, proliferation, anddifferentiation of oMSC. The data presented will provide areliable model system to assess the translation of MSC-basedtherapy into a variety of valuable ovine experimental modelsunder autologous settings.
Keywords Ovine mesenchymal stem cells . Bone marrow .
Multilineage differentiation . Polycaprolactone-co-lactidescaffold . Tissue engineering
Introduction
The reconstruction of post-traumatic or tumor-related largebone defects is still a significant clinical problem. Toovercome the disadvantages linked with autografts orallografts, numerous bone tissue engineering strategieshave been developed to create functional and structuraltissue alternatives (Hollinger et al. 1996; Li et al. 2005;Rentsch et al. 2008). The concept of tissue engineeringincorporates the combination of living cells, biologicallyactive molecules, and a structural scaffold to promote therepair and regeneration of tissue (Hutmacher et al. 2007).
C. Rentsch & R. Hess contributed equally to this work.
C. Rentsch (*) : S. Manthey :A. Biewener :H. ZwippDepartment of Trauma and Reconstructive Surgery, UniversityHospital Carl Gustav Carus Dresden,Fetscher Straße 74,01307 Dresden, Germanye-mail: [email protected]
R. Hess (*) :A. Hofmann :D. ScharnweberMax Bergmann Center of Biomaterials,Budapester Straße 27,01069 Dresden, Germanye-mail: [email protected]
B. RentschCatgut GmbH,Gewerbepark 18,08258 Markneukirchen, Germany
In Vitro Cell.Dev.Biol.—Animal (2010) 46:624–634DOI 10.1007/s11626-010-9316-0

The scaffold is expected to support cell adhesion,proliferation, and differentiation. The scaffold incorpo-rates essential characteristics such as biocompatibility,osteoinductivity, osteoconductivity, interconnecting pores,appropriate mechanical properties, and degradation (Gugalaand Gogolewski 2004; Salgado et al. 2004; Hutmacher etal. 2007).
Aliphatic polyesters such as poly(lactide) and poly(ε-caprolactone) have attracted considerable attention inbiomedicine because of their biodegradability and biocom-patibility (Salgado et al. 2004). Former studies have beenshown that an embroidered polycaprolactone-co-lactide(trade name PCL, Catgut GmbH) scaffold can act astemporary matrix for cell migration, proliferation, anddifferentiation of human mesenchymal stem cells (Rentschet al. 2009).
Considerable effort has been expended in mimickingthe extracellular matrix (ECM) to guide morphogenesisand tissue repair (Hubbell 2003). Type I collagen (coll I),the main organic composition of the ECM, can bereconstituted on the surface of certain polymers. Coll Iis augmented in its biomechanical and biochemicalfunction by glycosaminoglycans, such as chondroitinsulfate (CS; Bierbaum et al. 2006; Wollenweber et al.2006; Douglas et al. 2007). Previous studies have shownthat the use of ECM components enhances cell attach-ment and proliferation. It was shown that CS, as part ofthe artificial matrix, could induce the osteogenic differen-tiation of human mesenchymal stem cells (Rentsch et al.2008; 2009).
The next step, after the development of an adequatebiological active scaffold, is to choose a reliable source ofcells. Tissue engineering of bone can be performed withdifferent cell types. At present, concentrated bone marrow,mesenchymal stem cells (MSC), progenitor cells, osteo-blasts, or genetically manipulated cells are applied(Castano-Izquierdo et al. 2007). Due to their multipotency,MSC have a great potential for cell-based applications(Chamberlain et al. 2007).
Before clinical trials are accepted, the safety profile andsuccess of new drugs or implants are usually tested in largeanimal models (Perel et al. 2007). Sheep are an ideal modelfor bone tissue engineering because their anatomy andweight bearing are close to human conditions (Guo et al.2004). Despite the advantages of sheep as pre-clinicalmodel for bone tissue engineering, limited informationregarding ovine MSC (oMSC) are available. Numerousstudies have improved the understanding of the phenotype,physiology, differentiation potential, and possible clinicalapplication of MSC from different species. The majority ofresearch on MSC has been done on cells derived fromrodent and human (Phinney et al. 1999; Pittenger et al.1999; Woodbury et al. 2000).
The current study was undertaken with the goal beingisolation, cultivation, and characterization of oMSC for abetter comprehension of their biology in vitro. Furthermore,the objective was to determine whether biological activePCL scaffolds, designed as an implant for the treatment oflarge bone defects, support the growth and differentiation ofoMSC in vitro.
Materials and Methods
Isolation and cultivation of oMSC. This study has beenlicensed by the ethic commission of the University ofDresden (24D-9168.11-1-2006-17). All experimentswere carried out under written consent according tothe declaration of Helsinki for good scientific practice inits current form. During reconstructive surgery undertotal anesthesia, 10 ml of ovine bone marrow aspirateswere obtained from iliac crest of six merino sheep. TheoMSC were isolated using a density gradient (Percoll,Biochrom AG, Berlin, Germany) and cultured in Dulbecco'sModified Essential Medium [DMEM, Biochrom (F0415)],which contains 10% fetal calf serum (FCS) and 1%penicillin/streptomycin (all Biochrom; expansion medium)in a humidified atmosphere with 5% CO2 at 37°C. Thefirst culture medium change took place after 5 d and thenevery 3 d after that. Unless not otherwise mentioned, cellsof six merino sheep (oMSC CL-1 to 6) and hMSC controlcells were used between passage 2 and 4 for the followingexperiments.
Cell proliferation assay of oMSC. To determine thedoubling time and the growth characteristics of oMSC,the cells were seeded with a density of 2,000 cells/cm2
and cultured in the expansion medium for up to 21 d.After washing the cells twice in phosphate buffered saline(PBS), the cells were frozen, and cell number was deter-mined via measurement of the lactate dehydrogenase.
Lactate dehydrogenase assay. After cell lysis (1% triton X-100 in PBS), the lactate dehydrogenase (LDH) activity wasmeasured using the Cytotoxicity Kit (Takara; Otsu, Japan)according to the manufacturer's instructions. The findingswere correlated with the number of viable cells using acalibration line.
Immunofluorescence staining of surface markers. For sur-face marker staining, 4,000 cells/cm2 were seeded onglass slides and cultivated for 3 d in expansion medium.Fixation was carried out by using cold acetone/methanol(1:2). After fixation, cells were washed in PBS, blockedwith blocking solution (1% BSA in PBS, 1 h, RT) and
OMSC CHARACTERIZATION AND OSTEOGENESIS 625

incubated with the antibody solution (1:10 in blockingsolution, 1 h RT). The antibody solution contained a directfluorescent dye label or biotin [CD9-FITC (Immunotech,Marseille, France), CD44-biotin, CD45-APC, CD54-APC,CD73-PE, CD90-Biotin (BD Bioscience, San Jose, CA),CD105-FITC (Ancell, Bayport, MN), CD166-PE (RD-systems, Minneapolis, MN)]. In case of biotin labeling, asecond incubation with streptavidin–PerCP (BD Bioscience)took place (1:100 in blocking solution, 1 h RT). The slideswere counterstained with DAPI (Merck, Darmstadt, Ger-many) for 10 min. Lastly, the samples were covered withmounting medium and visualized under the fluorescencemicroscope (Leica Camera AG, Solms, Germany).
Chondrogenic differentiation, alcian blue and hematoxylinand eosin staining. Chondrogenic differentiation was in-duced by using the pellet cultivation system (Jackson et al.2007). For 21 d, 1,000,000 cells were cultivated inchondrogenic differentiation medium [DMEM containing1% penicillin/streptomycin, 1% insulin–transferin–selenit,10 ng/ml TGF-ß1, 210 µM L-ascorbic-acid-2-phosphateand 10 nM dexamethasone (all Sigma Aldrich, St. Louis,MO)]. Cells cultured in the expansion medium were used as acontrol. The medium was completely exchanged every 3–4 d.After cultivation, pellets were fixed in 4% formalin, imbeddedin paraffin blocks, and sectioned into 5-µm sections.
For Alcian blue staining, the sections were washed with3% acetic acid, stained with 1% Alcian blue (Sigma) for30 min, and finally washed with water. For hematoxylinand eosin (HE) staining, the sections were first incubatedfor 10 min in hematoxylin (Merck), followed by adifferentiation with 0.3% acid alcohol, and finally stainedwith eosin (Merck) for 5 min.
Adipogenic differentiation and oil red O staining. Foradipogenic differentiation, the cells were seeded with a densityof 30,000 cells/cm2. After 24 h, the expansion medium wasreplaced by adipogenic differentiation medium [DMEM,containing 10% FCS, 1% penicillin/streptomycin, 1 µMdexamethasone, 1.7 µM insulin, 0.2 mM indomethacin and0.5 mM 3-isobutylxanthin (Sigma)]. The cells were culturedfor 14 d. Cells cultured in expansion medium were used as acontrol. Adipogenic differentiated and control cells werefixed with 4% paraformaldehyde, washed with PBS, andincubated with 100% propylene glycol (Sigma) for 5 min.The lipids were stained with 0.1% Oil red O solution (Sigma)in isopropanol (10 min, 60°C), incubated in 85% propyleneglycol (10 min, RT), and finally rinsed in water.
Osteogenic differentiation. For osteogenic differentiation,the cells were seeded with a density of 2,000 cells/cm2.After 24 h, the expansion medium was replaced by theosteogenic differentiation medium [DMEM containing 10%
FCS, 1% penicillin/streptomycin, 100 nM dexamethasone,10 mM β-glycerophosphate, and 0.05 mM L-ascorbic acid-2-phosphate (Sigma)]. As a negative control, an equalnumber of cells were maintained in the expansion medium.All cells were cultured for up to 28 d with mediumchanges every 3–4 d. Afterward, the cell number and thecalcium content were measured, and the expression ofosteocalcin was visualized immunohistologically.
Calcium deposition. The calcium content was determinedcolorimetrically (absorbance 570 nm) by using theo-Cresolphthalein Complexone Kit (Rolf Greiner, Biochemica,Flacht, Germany). After measurement of the LDH, thelysate was freeze-dried, followed by a decalcification with0.2 M HCl (12 h, 4°C). The absorption was measured byusing a UV VIS spectrometer (Tecan Spectrafluor Plus) at570 nm. A standard curve was generated each time usingthe standards provided in the kit. Negative controlsincluded equivalent samples without cells. The calciumcontent of the cells was normalized by the LDH activity.Von Kossa staining (15 min in 5% silver nitrate and 2 minin 5% sodium thiosulfate) was used to characterize thebiological mineralization of the cells qualitatively.
Immunohistological analyses of osteocalcin. For osteocal-cin staining, the cells were fixed with 4% paraformaldehyde,washed with PBS, and incubated in blocking solution (1:2,serum and 2% dry milk in PBS). The primary osteocalcinantibody (Zymed, San Francisco, CA) was incubated overnight at 4°C (1:75), followed by an incubation with biotin-conjugated antibody (Vectastain Elite Kit, Vector Laborato-ries, Burlingame, Canada) for 30 min and finally withavidin–biotin conjugated peroxidase (Vectastain Elite Kit,Vector Laboratories). The immunoreaction was developedby Romulin (Biocare Medical, Concord, CA). The slideswere counterstained with HE.
Production and coating of the of PCL scaffolds. Thecopolymer was synthesized, by ring-opening copolymerisa-tion of L-lactide and ε-caprolactone and spun melted aspreviously described (Wollenweber et al. 2006; Rentsch etal. 2009). The resulted synthetic, resorbable, monofilamentpolycaprolactone-co-lactide fiber is commercially availableas PCL suture (Catgut GmbH, Markneukirchen, Germany).Textile scaffolds were made on a computer-aided embroi-dery machine and were coated with 914 µg (±157 µg) coll I(Porcine skin, MBP Neustadt-Glewe, Germany) or coll Iand 33,5 µg (±7) CS A and CS B (Sigma Aldrich) perscaffold as previously described (Wollenweber et al. 2006;Rentsch et al. 2009).
Scanning electron microscopy. After sputtering the non-coated or coll I/CS-coated PCL scaffolds with 50 nm gold
626 RENTSCH ET AL.
layer (Baltec sputter coater), the samples were observed in aPhilips FEG ESEM in a HiVak mode with accelerationvoltages of 2–10 kV.
Seeding and culture of oMSC on PCL scaffolds. Beforeseeding the scaffolds were beaded on gauge needles asdescribed before, pre-incubated in expansion medium for2–4 h at 37°C, in silan-coated spinner flasks (Wollenweberet al. 2006; Rentsch et al. 2009). Four hundred seventy-fivethousand cells per scaffold were added to the preparedspinner flask and incubated in a humidified atmospherewith 5% CO2 at 37°C with permanent stirring for 48 h.After seeding, the scaffolds were placed in 12-wellpolystyrene plates and incubated in expansion medium upto 28 d. The medium was changed every 3 d. At differentpoints in time, samples were analyzed for cell adherence,proliferation, and osteogenic differentiation.
Data analysis. Unless otherwise noted, all experimentswere run in triplicate and repeated at least threedifferent times. The results were analyzed using thestandard Student's t test. Cells from six merino sheep(oMSC CL-1 to 6) were characterized and oMSC CL-1were used to show the results representative for all ovinecell lines.
Results
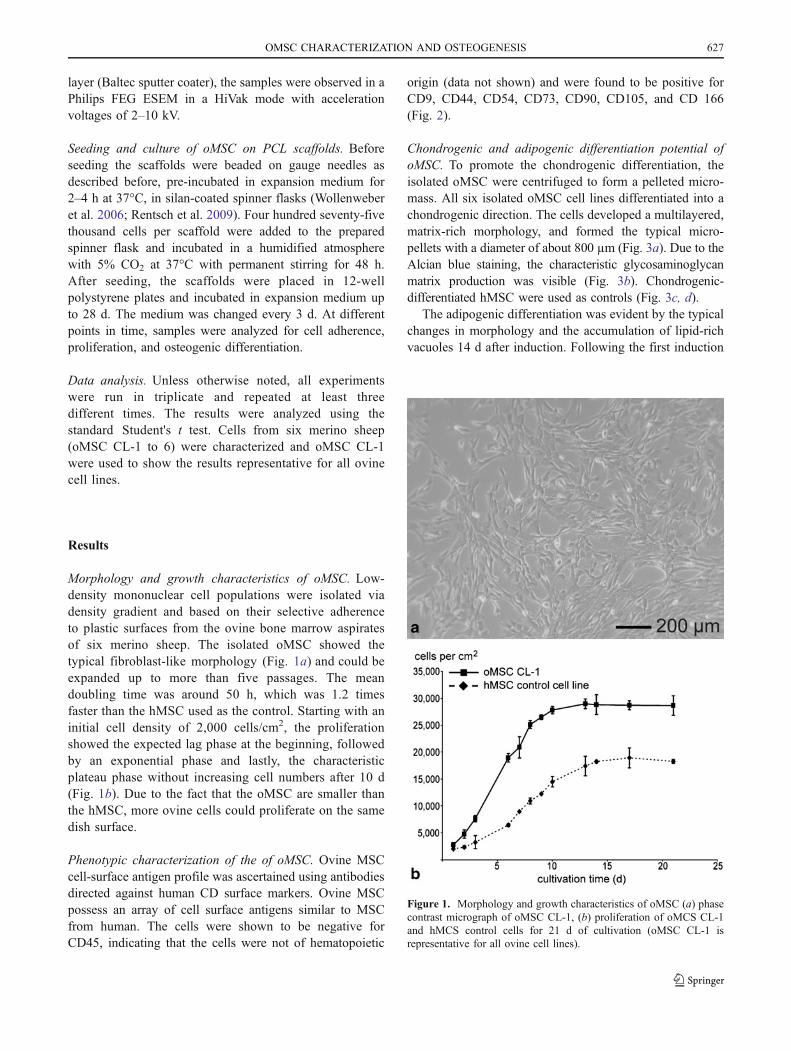
Morphology and growth characteristics of oMSC. Low-density mononuclear cell populations were isolated viadensity gradient and based on their selective adherenceto plastic surfaces from the ovine bone marrow aspiratesof six merino sheep. The isolated oMSC showed thetypical fibroblast-like morphology (Fig. 1a) and could beexpanded up to more than five passages. The meandoubling time was around 50 h, which was 1.2 timesfaster than the hMSC used as the control. Starting with aninitial cell density of 2,000 cells/cm2, the proliferationshowed the expected lag phase at the beginning, followedby an exponential phase and lastly, the characteristicplateau phase without increasing cell numbers after 10 d(Fig. 1b). Due to the fact that the oMSC are smaller thanthe hMSC, more ovine cells could proliferate on the samedish surface.
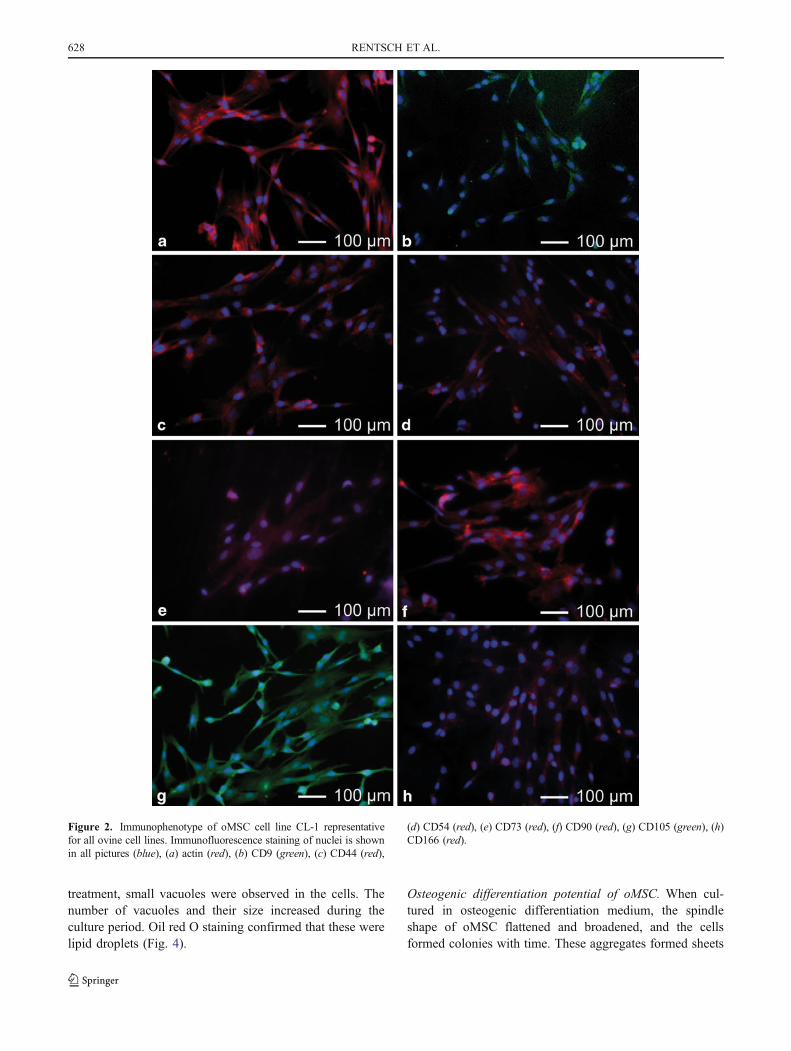
Phenotypic characterization of the of oMSC. Ovine MSCcell-surface antigen profile was ascertained using antibodiesdirected against human CD surface markers. Ovine MSCpossess an array of cell surface antigens similar to MSCfrom human. The cells were shown to be negative forCD45, indicating that the cells were not of hematopoietic
origin (data not shown) and were found to be positive forCD9, CD44, CD54, CD73, CD90, CD105, and CD 166(Fig. 2).
Chondrogenic and adipogenic differentiation potential ofoMSC. To promote the chondrogenic differentiation, theisolated oMSC were centrifuged to form a pelleted micro-mass. All six isolated oMSC cell lines differentiated into achondrogenic direction. The cells developed a multilayered,matrix-rich morphology, and formed the typical micro-pellets with a diameter of about 800 µm (Fig. 3a). Due to theAlcian blue staining, the characteristic glycosaminoglycanmatrix production was visible (Fig. 3b). Chondrogenic-differentiated hMSC were used as controls (Fig. 3c, d).
The adipogenic differentiation was evident by the typicalchanges in morphology and the accumulation of lipid-richvacuoles 14 d after induction. Following the first induction
Figure 1. Morphology and growth characteristics of oMSC (a) phasecontrast micrograph of oMSC CL-1, (b) proliferation of oMCS CL-1and hMCS control cells for 21 d of cultivation (oMSC CL-1 isrepresentative for all ovine cell lines).
OMSC CHARACTERIZATION AND OSTEOGENESIS 627
treatment, small vacuoles were observed in the cells. Thenumber of vacuoles and their size increased during theculture period. Oil red O staining confirmed that these werelipid droplets (Fig. 4).
Osteogenic differentiation potential of oMSC. When cul-tured in osteogenic differentiation medium, the spindleshape of oMSC flattened and broadened, and the cellsformed colonies with time. These aggregates formed sheets
Figure 2. Immunophenotype of oMSC cell line CL-1 representativefor all ovine cell lines. Immunofluorescence staining of nuclei is shownin all pictures (blue), (a) actin (red), (b) CD9 (green), (c) CD44 (red),
(d) CD54 (red), (e) CD73 (red), (f) CD90 (red), (g) CD105 (green), (h)CD166 (red).
628 RENTSCH ET AL.

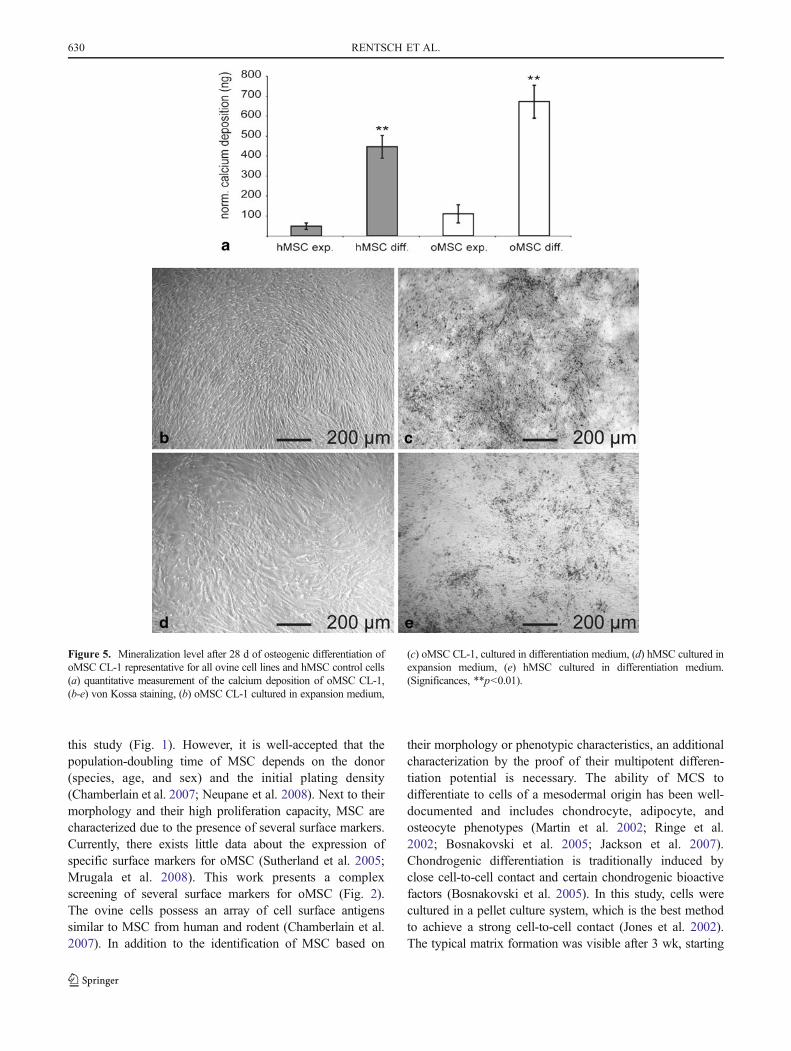
and detached easily from the culture disk. The cellphenotype did not change in the untreated cells. In thepresence of expansion medium, only a small amount ofcalcium deposition (oMSC CL-1=109 ng (±45)) could beobserved at day 28. By contrast, oMSC cultured inosteogenic differentiation medium showed a significantincrease in calcium deposition as quantitatively determined(oMSC CL-1=670 ng (±82)) and observed by von Kossastaining (Fig. 5).
Immunohistological staining of the osteogenic markerosteocalcin was performed on day 14. The expression ofosteocalcin was only observed in the cells cultured inosteogenic differentiation medium (Fig. 6).
Scaffold morphology. The round and approximately 1-mmhigh PCL scaffolds with a 19 mm outer diameter and10 mm inner diameter (Fig. 7a), showed a triaxial structure
(deposition of the thread in a triangular assembly; 0°/60°/120°-netting) with a stitch length of 1.4 mm and a meshspacing of 1.2 mm. The pore size in the scaffold was withinthe range of 0.2 to 1 mm, as assessed from SEM images(Fig. 7b). Reticular PCL scaffolds were coated with collI/CS. During the fibrillogenesis, the coll I/CS mixtureadsorbed on the scaffold surface fibers and coveredpartially the pores (Fig. 7c).
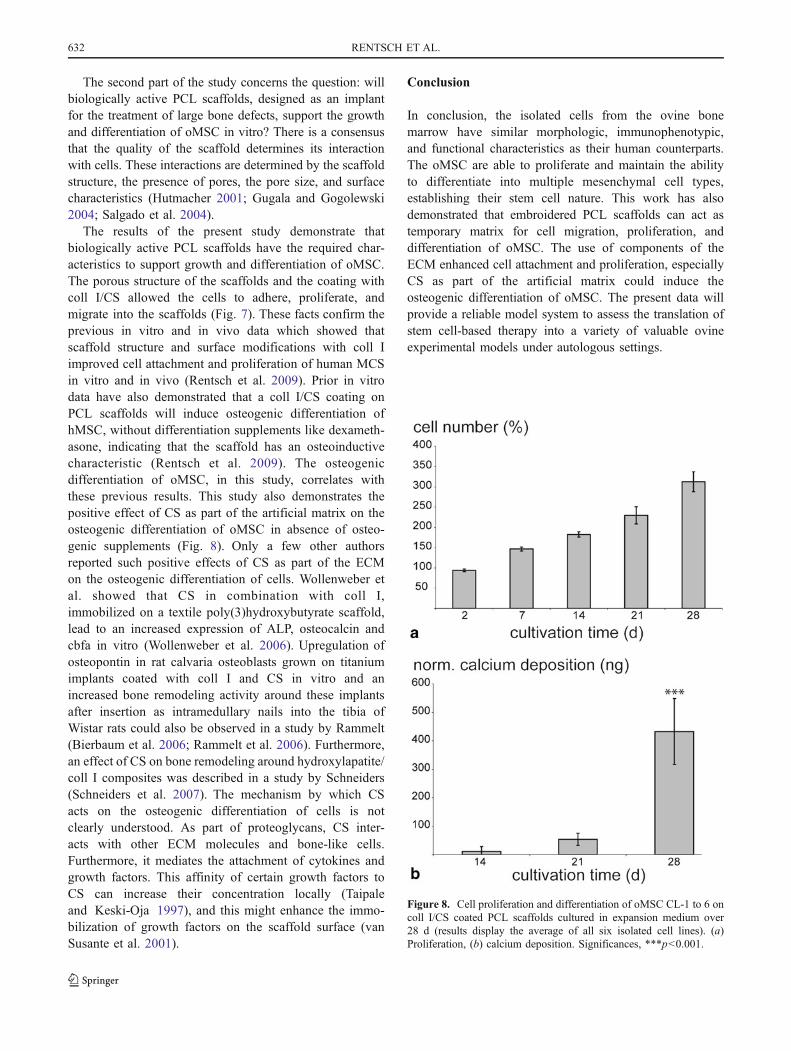
Cell adherence, growth, and osteogenic differentiation ofoMSC on PCL scaffolds. The scaffolds coated with collI/CS were seeded, as described above, and incubated inexpansion medium without any differentiation additivesover 28 d. At different points in time, samples wereanalyzed for cell adherence, proliferation, and osteogenicdifferentiation. The coll I/CS-coated scaffolds allowed theoMSC to adhere. The cells attached to the polymer fiberand to the coll I/CS coating with a flattened morphology.They developed podia, which linked the cells with eachother. A penetration of the cells into the plane 3D scaffoldcould be observed (Fig. 7d). After 48 h, a maximum cellnumber of around 447,187 cells (displays a seedingefficiency of 94%) could adhere on one scaffold. Afteradherence, a proliferation to higher amounts of cells waspossible (Fig. 8a). The calcium deposition was evaluatedas a marker for the osteogenic differentiation of oMSC onthe coll I/CS scaffolds. After 28 d of cultivation in theexpansion medium, the cells showed a highly significantincrease up to 432.7 ng (+/− 116.1 ng) of calciumdeposition (Fig. 8b). This indicates that the osteogenicdifferentiation of the oMSC on coll I/CS-coated scaffoldswas induced by the surface modification.
Discussion
Tissue-engineered transplants are mainly composed oftissue-specific cells, biologically active molecules, and astructural scaffold on which cells can develop (Ringe et al.2002; Hutmacher et al. 2007). Despite the popularity ofsheep as a model for in vitro and in vivo experiments intissue engineering applications, there is little data withrespect to oMSC. First of all, this report presents dataconcerning the isolation, cultivation, and characterization ofa fibroblast-like population from adult sheep bone marrow.Furthermore, the object of the study was to determinewhether a biological active PCL scaffold, designed as animplant for the treatment of large bone defects, supports thegrowth and differentiation of oMSC in vitro.
Based on their typical, spindle-shaped fibroblast-likemorphology, the isolated oMSC are very similar to MCSfrom rodent or human sources. The proliferation of oMSCwas faster in comparison to hMSC used as the control in
Figure 3. Chondrogenic differentiation for 21 d of oMSC CL-1representative for all ovine cell lines and hMSC control cells (a) HEstaining of oMSC CL-1, (b) Alcian blue staining of oMSC CL-1, (c) HEstaining of hMSC (d) Alcian blue staining of hMSC (magnification ×10).
Figure 4. Oil red O staining of lipid vacuoles after adipogenicdifferentiation for 14 d, (a) oMSC CL-1 representative for all ovinecell lines, (b) hMSC control cells.
OMSC CHARACTERIZATION AND OSTEOGENESIS 629
this study (Fig. 1). However, it is well-accepted that thepopulation-doubling time of MSC depends on the donor(species, age, and sex) and the initial plating density(Chamberlain et al. 2007; Neupane et al. 2008). Next to theirmorphology and their high proliferation capacity, MSC arecharacterized due to the presence of several surface markers.Currently, there exists little data about the expression ofspecific surface markers for oMSC (Sutherland et al. 2005;Mrugala et al. 2008). This work presents a complexscreening of several surface markers for oMSC (Fig. 2).The ovine cells possess an array of cell surface antigenssimilar to MSC from human and rodent (Chamberlain et al.2007). In addition to the identification of MSC based on
their morphology or phenotypic characteristics, an additionalcharacterization by the proof of their multipotent differen-tiation potential is necessary. The ability of MCS todifferentiate to cells of a mesodermal origin has been well-documented and includes chondrocyte, adipocyte, andosteocyte phenotypes (Martin et al. 2002; Ringe et al.2002; Bosnakovski et al. 2005; Jackson et al. 2007).Chondrogenic differentiation is traditionally induced byclose cell-to-cell contact and certain chondrogenic bioactivefactors (Bosnakovski et al. 2005). In this study, cells werecultured in a pellet culture system, which is the best methodto achieve a strong cell-to-cell contact (Jones et al. 2002).The typical matrix formation was visible after 3 wk, starting
Figure 5. Mineralization level after 28 d of osteogenic differentiation ofoMSC CL-1 representative for all ovine cell lines and hMSC control cells(a) quantitative measurement of the calcium deposition of oMSC CL-1,(b-e) von Kossa staining, (b) oMSC CL-1 cultured in expansion medium,
(c) oMSC CL-1, cultured in differentiation medium, (d) hMSC cultured inexpansion medium, (e) hMSC cultured in differentiation medium.(Significances, **p<0.01).
630 RENTSCH ET AL.

from the pellet borders (Fig. 3). This development is alsotypical for chondrogenic differentiated MSC derived fromhuman, canine, equine, bovine, and rodents models(Pittenger et al. 1999; Woodbury et al. 2000; Worster et al.2000; Ringe et al. 2002; Bosnakovski et al. 2005; Neupaneet al. 2008). Adipogenic differentiation could be induced(Fig. 4), in this study, by a routinely used method to
stimulate the adipogenic differentiation of human stem cells(Pittenger et al. 1999; Chamberlain et al. 2007). Osteo-genesis was analyzed by observing changes in cell morpho-logy, the mineralization of the matrix, and the expression ofosteocalcin. During the cultivating of oMSC in the osteoin-ductive medium, the morphology of the cells changed from afibroblastic to a cuboidal shape, which is also characteristicfor osteogenesis of other species (Bosnakovski et al. 2005;Neupane et al. 2008). These changes in morphology werefollowed by the formation of highly mineralized cellaggregates. Interestingly, ALP activity could not be detectedin oMSC using different methods (data not shown). Thiseffect goes in line with the group of Kon (Kon et al. 2000).Currently, there is no literature reported regarding theexpression of ALP by oMSC under osteogenic conditions.This effect could also be found in feline MSC, by the groupof Martin (Martin et al. 2002). They explained thephenomenon by the unusually short half-life of the felineenzyme and expected that the feline alkaline phosphatasewas simply undetectable with the assay system used. Thiscould be also the case for oMSC. However, the expression ofthe osteogenic marker osteocalcin, as well as the highcapacity of oMSC to mineralize the matrix (Figs. 5 and 6),demonstrates the osteogenic differentiation potential of theisolated cells.
Figure 6. Osteocalcin staining after osteogenic differentiation for 14 d ofoMSC CL-1 representative for all ovine cell lines and hMSC controlcells, positive osteocalcin staining is shown in red-brown and markedwith arrows, (a) oMSC CL-1 cultured in expansion medium, (b) oMSCCL-1 cultured in differentiation medium, (c) hMSC cultured in expansionmedium, (d) hMSC cultured in differentiation medium.
Figure 7. Embroidered PCL scaffold. a photograph of PCL scaffold, b SEM micrograph of non-coated PCL scaffold, c SEM micrograph of collI/CS-coated PCL scaffold, d oMSC adherence on coll I/CS-coated PCL scaffolds after 24 h of cultivation in expansion medium, DAPI staining.
OMSC CHARACTERIZATION AND OSTEOGENESIS 631
The second part of the study concerns the question: willbiologically active PCL scaffolds, designed as an implantfor the treatment of large bone defects, support the growthand differentiation of oMSC in vitro? There is a consensusthat the quality of the scaffold determines its interactionwith cells. These interactions are determined by the scaffoldstructure, the presence of pores, the pore size, and surfacecharacteristics (Hutmacher 2001; Gugala and Gogolewski2004; Salgado et al. 2004).
The results of the present study demonstrate thatbiologically active PCL scaffolds have the required char-acteristics to support growth and differentiation of oMSC.The porous structure of the scaffolds and the coating withcoll I/CS allowed the cells to adhere, proliferate, andmigrate into the scaffolds (Fig. 7). These facts confirm theprevious in vitro and in vivo data which showed thatscaffold structure and surface modifications with coll Iimproved cell attachment and proliferation of human MCSin vitro and in vivo (Rentsch et al. 2009). Prior in vitrodata have also demonstrated that a coll I/CS coating onPCL scaffolds will induce osteogenic differentiation ofhMSC, without differentiation supplements like dexameth-asone, indicating that the scaffold has an osteoinductivecharacteristic (Rentsch et al. 2009). The osteogenicdifferentiation of oMSC, in this study, correlates withthese previous results. This study also demonstrates thepositive effect of CS as part of the artificial matrix on theosteogenic differentiation of oMSC in absence of osteo-genic supplements (Fig. 8). Only a few other authorsreported such positive effects of CS as part of the ECMon the osteogenic differentiation of cells. Wollenweber etal. showed that CS in combination with coll I,immobilized on a textile poly(3)hydroxybutyrate scaffold,lead to an increased expression of ALP, osteocalcin andcbfa in vitro (Wollenweber et al. 2006). Upregulation ofosteopontin in rat calvaria osteoblasts grown on titaniumimplants coated with coll I and CS in vitro and anincreased bone remodeling activity around these implantsafter insertion as intramedullary nails into the tibia ofWistar rats could also be observed in a study by Rammelt(Bierbaum et al. 2006; Rammelt et al. 2006). Furthermore,an effect of CS on bone remodeling around hydroxylapatite/coll I composites was described in a study by Schneiders(Schneiders et al. 2007). The mechanism by which CSacts on the osteogenic differentiation of cells is notclearly understood. As part of proteoglycans, CS inter-acts with other ECM molecules and bone-like cells.Furthermore, it mediates the attachment of cytokines andgrowth factors. This affinity of certain growth factors toCS can increase their concentration locally (Taipaleand Keski-Oja 1997), and this might enhance the immo-bilization of growth factors on the scaffold surface (vanSusante et al. 2001).
Conclusion
In conclusion, the isolated cells from the ovine bonemarrow have similar morphologic, immunophenotypic,and functional characteristics as their human counterparts.The oMSC are able to proliferate and maintain the abilityto differentiate into multiple mesenchymal cell types,establishing their stem cell nature. This work has alsodemonstrated that embroidered PCL scaffolds can act astemporary matrix for cell migration, proliferation, anddifferentiation of oMSC. The use of components of theECM enhanced cell attachment and proliferation, especiallyCS as part of the artificial matrix could induce theosteogenic differentiation of oMSC. The present data willprovide a reliable model system to assess the translation ofstem cell-based therapy into a variety of valuable ovineexperimental models under autologous settings.
Figure 8. Cell proliferation and differentiation of oMSC CL-1 to 6 oncoll I/CS coated PCL scaffolds cultured in expansion medium over28 d (results display the average of all six isolated cell lines). (a)Proliferation, (b) calcium deposition. Significances, ***p<0.001.
632 RENTSCH ET AL.

Acknowledgments The authors would like to acknowledge theSächsische Aufbaubank (11633/1848) and the team of Dr. Jung andDr. Speckl from the Animal Care Unit of the Dresden UniversityHospital “Carl Gustav Carus” for their cooperation in conduction ofthe animal experiments.
References
Bierbaum S.; Douglas T.; Hanke T.; Scharnweber D.; Tippelt S.;Monsees T. K.; Funk R. H. W.; Worch H. Collageneous matrixcoating on titanium implants modified with decorin and chondroi-tin sulfate: Characterization and influence on osteoblastic cells.J Biomed Mat Res A 1(77): 551–562; 2006.
Bosnakovski D.; Mizuno M.; Kim G.; Takagi S.; Okumura M.;Fujinaga T. Isolation and multilineage differentiation of bovinebone marrow mesenchymal stem cells. Cell Tissue Res 319(2):243–53; 2005.
Castano-Izquierdo H.; Alvarez-Barreto J.; van den Dolder J.; Jansen J.A.; Mikos A. G.; Sikavitsas V. I. Pre-culture period ofmesenchymal stem cells in osteogenic media influences their invivo bone forming potential. J Biomed Mater Res A 82(1): 129–38; 2007.
Chamberlain G.; Fox J.; Ashton B.; Middleton J. Concise review:mesenchymal stem cells: their phenotype, differentiation capac-ity, immunological features, and potential for homing. Stem Cells25(11): 2739–49; 2007.
Douglas T.; Heinemann S.; Mietrach C.; Hempel U.; Bierbaum S.;Scharnweber D.; Worch H. Interactions of collagen types I and IIwith chondroitin sulfates A-C and their effect on osteoblastadhesion. Biomacromolecules 8(4): 1085–92; 2007.
Gugala Z.; Gogolewski S. Differentiation, growth and activity of ratbone marrow stromal cells on resorbable poly(L/DL-lactide)membranes. Biomaterials 25: 2299–2307; 2004.
Guo X.; Wang C.; Duan C.; Descamps M.; Zhao Q.; Dong L.; LüS.; Anselme K.; Lu J.; Song Y. Q. Repair of osteochondraldefects with autologous chondrocytes seeded onto bioceramicscaffold in sheep. Tissue Eng 10(11–12): 1830–40; 2004.
Hollinger J. O.; Brekke J.; Gruskin E.; Lee D. Role of bonesubstitutes. Clin Orthop Relat Res 324: 55–65; 1996.
Hubbell J. A. Materials as morphogenetic guides in tissue engineering.Curr Opin Biotechnol 14(5): 551–8; 2003.
Hutmacher D. W. Scaffold design and fabrication technologies forengineering tissue – state of the art and future perspectives.J Biomater Sci 12(1): 107–124; 2001.
Hutmacher D. W.; Schantz J. T.; Lam C. X.; Tan K. C.; Lim T. C.State of the art and future directions of scaffold-based boneengineering from a biomaterials perspective. J Tissue Eng RegenMed 1(4): 245–60; 2007.
Jackson L.; Jones D. R.; Scotting P.; Sottile V. Adult mesenchymalstem cells: differentiation potential and therapeutic applications.J Postgrad Med 53(2): 121–7; 2007.
Jones E. A.; Kinsey S. E.; English A.; Jones R. A.; Straszynski L.;Meredith D. M.; Markham A. F.; Jack A.; Emery P.; McGonagleD. Isolation and characterization of bone marrow multipotentialmesenchymal progenitor cells. Arthritis Rheum 46(12): 3349–60;2002.
Kon E.; Muraglia A.; Corsi A.; Bianco P.; Marcacci M.; Martin I.;Boyde A.; Ruspantini I.; Chistolini P.; Rocca M.; Giardino R.;Cancedda R.; Quarto R. Autologous bone marrow stromal cellsloaded onto porous hydroxyapatite ceramic accelerate bone repair
in critical-size defects of sheep long bones. J Biomed Mater Res49(3): 328–37; 2000.
Li W. J.; Tuli R.; Huang X.; Laquerriere P.; Tuan R. S. Multilineagedifferentiation of human mesenchymal stem cells in a three-dimensional nanofibrous scaffold. Biomaterials 26(25): 5158–66;2005.
Martin D. R.; Cox N. R.; Hathcock T. L.; Niemeyer G. P.; Baker H. J.Isolation and characterization of multipotential mesenchymalstem cells from feline bone marrow. Exp Hematol 30(8): 879–86;2002.
Mrugala D.; Bony C.; Neves N.; Caillot L.; Fabre S.; Moukoko D.;Jorgensen C.; Noël D. Phenotypic and functional characterisationof ovine mesenchymal stem cells: application to a cartilage defectmodel. Ann Rheum Dis 67(3): 288–95; 2008.
Neupane M.; Chang C. C.; Kiupel M.; Yuzbasiyan-Gurkan V.Isolation and Characterization of Canine Adipose-Derived Mes-enchymal Stem Cells. Tissue Eng Part A Apr 17; 2008.
Perel P.; Roberts I.; Sena E.; Wheble P.; Briscoe C.; SandercockP.; Macleod M.; Mignini L. E.; Jayaram P.; Khan K. S.Comparison of treatment effects between animal experimentsand clinical trials: systematic review. BMJ 334(7586): 197;2007.
Phinney D. G.; Kopen G.; Isaacson R. L.; Prockop D. J. Plasticadherent stromal cells from the bone marrow of commonly usedstrains of inbred mice: variations in yield, growth, anddifferentiation. J Cell Biochem 72(4): 570–85; 1999.
Pittenger M. F.; Mackay A. M.; Beck S. C.; Jaiswal R. K.; DouglasR.; Mosca J. D.; Moorman M. A.; Simonetti D. W.; Craig S.;Marshak D. R. Multilineage potential of adult human mesenchy-mal stem cells. Science 284(5411): 143–7; 1999.
Rammelt S.; Illert T.; Bierbaum S.; Scharnweber D.; Zwipp H.;Schneiders W. Coating of titanium implants with collagen, RGDpeptide and chondroitin sulfate. Biomaterials 27(32): 5561–71;2006.
Rentsch B.; Hofmann A.; Breier A.; Rentsch C.; Scharnweber S.Embroidered and surface modified polycaprolactone-co-lactide scaffolds as bioartificial bone substitute—in vitrocharacterization. Annals of Biomedical Engineering, In press;2009.
Rentsch C.; Rentsch B.; Breier A.; Hofmann A.; Manthey S.;Scharnweber D.; Biewener A.; Zwipp H. Evaluation of theosteogenic potential and vascularization of 3D poly(3)hydrox-ybutyrate Scaffolds Subcutaneously Implanted in Nude Rats.Biomed Mater Res A, In press; 2008.
Ringe J.; Kaps C.; Schmitt B.; Büscher K.; Bartel J.; Smolian H.;Schultz O.; Burmester G. R.; Häupl T.; Sittinger M. Porcinemesenchymal stem cells. Induction of distinct mesenchymal celllineages. Cell Tissue Res 307(3): 321–7; 2002.
Salgado A. J.; Coutinho O. P.; Reis R. L. Bone tissue engineering:state of the art and future trends. Maccromol Biosci 4: 743–765; 2004.
Schneiders W.; Reinstorf A.; Ruhnow M.; Rehberg S.; Heineck J.;Hinterseher I.; Biewener A.; Zwipp H.; Rammelt S. Effect ofchondroitin sulphate on material properties and bone remodellingaround hydroxyapatite/collagen composites. J Biomed Mater ResA; Sep 5; 2007.
Sutherland F.W.; Perry T. E.; Yu Y.; SherwoodM. C.; Rabkin E.; MasudaY.; Garcia G. A.; McLellan D. L.; Engelmayr Jr. G. C.; Sacks M. S.;Schoen F. J.; Mayer Jr. J. E. From stem cells to viable autologoussemilunar heart valve. Circulation 111(21): 2783–91; 2005.
Taipale J.; Keski-Oja J. Growth factors in the extracellular matrix.FASEB J 11(1): 51–9; 1997.
van Susante J. L. C.; Pieper J.; Buma P.; van Kuppevelt T. H.; vanBeuningen H.; van Der Kraan P. M.; Veerkamp J. H.; van denBerg W. B.; Veth R. P. H. Linkage of chondroitin-sulfate to type
OMSC CHARACTERIZATION AND OSTEOGENESIS 633

I collagen scaffolds stimulates the bioactivity of seeded chon-drocytes in vitro. Biomaterials 22(17): 2359–69; 2001.
Wollenweber M.; Domaschke H.; Hanke T.; Boxberger S.; SchmackG.; Gliesche K.; Scharnweber D.; Worch H. Mimicked bioartifi-cial matrix containing chondroitin sulfate on a textile scaffold ofpoly(3-hydroxybutyrate) alters the differentiation of adult humanmesenchymal stem cells. Tissue Eng 12(2): 345–59; 2006.
Woodbury D.; Schwarz E. J.; Prockop D. J.; Black I. B. Adult rat andhuman bone marrow stromal cells differentiate into neurons.J Neurosci Res 61(4): 364–70; 2000.
Worster A. A.; Nixon A. J.; Brower-Toland B. D.; Williams J. Effectof transforming growth factor beta1 on chondrogenic differenti-ation of cultured equine mesenchymal stem cells. Am J Vet Res61(9): 1003–10; 2000.
634 RENTSCH ET AL.




















