
Ostrich expansion into India during the Late Pleistocene: Implications for continental dispersal...
36
Ostrich expansion into India during the Late Pleistocene: Implications for continental dispersal corridors James Blinkhorn, Hema Achyuthan, Michael D. Petraglia PII: S0031-0182(14)00537-9 DOI: doi: 10.1016/j.palaeo.2014.10.026 Reference: PALAEO 7063 To appear in: Palaeogeography, Palaeoclimatology, Palaeoecology Received date: 2 July 2014 Revised date: 14 October 2014 Accepted date: 21 October 2014 Please cite this article as: Blinkhorn, James, Achyuthan, Hema, Petraglia, Michael D., Ostrich expansion into India during the Late Pleistocene: Implications for continen- tal dispersal corridors, Palaeogeography, Palaeoclimatology, Palaeoecology (2014), doi: 10.1016/j.palaeo.2014.10.026 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Transcript of Ostrich expansion into India during the Late Pleistocene: Implications for continental dispersal...

�������� ����� ��
Ostrich expansion into India during the Late Pleistocene: Implications forcontinental dispersal corridors
James Blinkhorn, Hema Achyuthan, Michael D. Petraglia
PII: S0031-0182(14)00537-9DOI: doi: 10.1016/j.palaeo.2014.10.026Reference: PALAEO 7063
To appear in: Palaeogeography, Palaeoclimatology, Palaeoecology
Received date: 2 July 2014Revised date: 14 October 2014Accepted date: 21 October 2014
Please cite this article as: Blinkhorn, James, Achyuthan, Hema, Petraglia, Michael D.,Ostrich expansion into India during the Late Pleistocene: Implications for continen-tal dispersal corridors, Palaeogeography, Palaeoclimatology, Palaeoecology (2014), doi:10.1016/j.palaeo.2014.10.026
This is a PDF file of an unedited manuscript that has been accepted for publication.As a service to our customers we are providing this early version of the manuscript.The manuscript will undergo copyediting, typesetting, and review of the resulting proofbefore it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers thatapply to the journal pertain.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
1
Ostrich expansion into India during the Late Pleistocene: Implications for continental dispersal
corridors
Blinkhorn, James1, Achyuthan, Hema
2, Petraglia, Michael D.
3
1UMR5199 PACEA, Université de Bordeaux, Avenues de Facultes, Cedex Pessac, France
2Department of Geology, Anna University, Chennai 600 025, India
3School of Archaeology, Research Laboratory for Archaeology and the History of Art, University of
Oxford, Oxford OX1 3QY, United Kingdom
Correspondance: J. Blinkhorn, UMR5199 PACEA, Université de Bordeaux, Avenues de Facultes,
Cedex Pessac, France; Email: [email protected] Phone : 00 33 5 40 00 25 45
ABSTRACT
New evidence is presented for the earliest occurrence of ostrich (Struthio sp.) in India during the Late
Pleistocene along with a synthesis on the evidence for ostrich populations in the subcontinent. Direct
dating of ostrich eggshell using Accelerator Mass Spectrometry (AMS) radiocarbon methods on
excavated samples from Katoati, Rajasthan, India, are supported by Optically Stimulated
Luminesence (OSL) dating of associated sediments to demonstrate the arrival of ostrich in India
before 60 thousand years ago (ka). In addition, the first stable isotope studies on ostrich eggshell from
India have been conducted, yielding a new form of palaeoenvironmental proxy data for the Late
Pleistocene. The geographic expansion of ostrich into India corresponds with the distribution of
Sahel-like environments, bordering but not substantially colonising endemic Indian vegetation zones.
The dispersal of ostrich into India marks a rare introduction of megafauna into the subcontinent
during the Late Pleistocene, the longevity of which spans a period more than 40 ka. The timespan and
range of this colonisation indicates the availability and exploitation of suitable habitats in India. The
continental dispersal of ostrich into India during the Late Pleistocene offers useful insights into the

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
2
debate surrounding the dispersal of modern humans, and contrasts with the hypothesized coastal
movement of Homo sapiens into India.
Keywords: Dispersal; India; Katoati; Late Pleistocene; stable isotope; Struthio
Highlights
Oldest directly dated evidence for Late Pleistocene expansion of Struthio into India
First stable isotope study of Late Pleistocene ostrich eggshell in India
Synthesis of existing evidence for Struthio populations in South Asia
Struthio expansion associated with extension of Sahel-like habitats

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
3
1. Introduction
Broad continuity in the presence of megafauna can be observed in the Indian subcontinent
throughout the Middle Pleistocene and extending into the Late Pleistocene (Roberts et al., 2014). The
Pakistani Siwalik formations yield repeated evidence for the presence of Struthiodiae (family of
flightless ratite birds), spanning the Middle Miocene to Middle Pleistocene (Dennell, 2004; Patnaik et
al., 2009b; Stern et al., 1994). However, a significant change in the distribution of Struthiodiae is
evident in the Late Pleistocene, becoming absent in the Himalayan forelands and instead appearing in
western and central India for the first time (Andrews, 1911; Badam, 2005; Bidwell, 1910). The timing
of this significant range expansion is poorly constrained and has yet to be set within the broader
context of Late Pleistocene faunal dynamics and their relationship with palaeoenvironmental changes.
The only osteological remains of Struthiodiae from South Asia were collected from unspecified
Siwalik strata and described as a new species, Struthio asiaticus, by Milne-Edwards (1871) on the
basis of size differences with the extant African ostrich, S. camelus. A metric study of these
specimens noted that despite presenting a more robust neck, the Siwalik specimens were comparable
in form and size with a large male specimen of S. camelus (Davies, 1880). Further description by
Lydekker (1884) suggested that the stouter neck of the Siwalik specimen should be considered as
within the realm of individual variation, rather than at a species level, thus suggesting S. asiaticus be
treated as a ‘preliminary’ taxon. Since this time, a number of osteological remains and, more
problematically, eggshell specimens, have been attributed to S. asiaticus spanning from the Pliocene
to the Late Pleistocene and ranging from South Africa to East Asia (Kurochkin et al., 2010; Manegold
et al., 2013; Mikhailov, 1991; Mourer-Chauviré and Geraads, 2008), making it the most widespread
species in the genus Struthio. Osteological studies of specimens attributed to S. asiaticus are noted to
be between 20-50% more robust than contemporary African ostrich, although not necessarily taller
(Mourer-Chauvire and Geraads 2008).
In contrast to the paucity of osteological remains, ostrich eggshells (OES) have been recovered
from a number of contexts in the Siwalik Range (Dennell, 2005; Stern et al., 1994) and in peninsula

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
4
India (Badam 2005). Ratite eggshells with aepyornithoid-type pore patterns are reported from the
mid-Miocene Dhok Pathan formations near Hasnot (Patnaik et al., 2009b; Sauer, 1972), and a range
of sites in the Siwalik formations dating between 11.35-1.25 millon years ago (Ma.) (Stern et al.,
1994). Eggshells with struthionid-type pore patterns are found in younger Siwalik deposits ranging in
age from 2.24–0.5 Ma. (Stern et al., 1994) and at a number of sites in the Indian states of Rajasthan,
Uttar Pradesh, Madhya Pradesh and Maharashtra dating between >43–18 thousand years ago (ka)
(Badam 2005). More detailed description of eggshell morphology and putative taxonomic affiliations
of these OES samples have been restricted to the Late Pleistocene samples (Sahni et al., 1990). The
earliest samples examined, from Ken River (Uttar Pradesh), were considered as “practically
indistinguishable” from egg-shell of modern Somali ostrich (S. camelus molydophanes)(Andrews
1911; Bidwell 1910), though noted as thicker than contemporary examples, with more recent studies
reaching the same conclusions based upon wider sampling from peninsula India (Sahni et al., 1990).
The extinction of Late Pleistocene Struthio sp. from India has been briefly discussed (e.g. Badam
2005) yet the timing and potential causes for their expansion into western and central India has not be
addressed. Pertinent to this are patterns of climatic amelioration during the Late Pleistocene that may
have permitted the extension of favourable habitats for ostrich from their endemic regions into India.
Here, evidence is presented for the earliest dated Late Pleistocene Struthio samples from western
India, which is placed in its palaeoenvironmental context along with all known samples of
Struthiodiae from South Asia. Factors that may have affected this Late Pleistocene expansion are
discussed
2. Materials and Methods
Ostrich eggshell has been recovered from both surface and excavated contexts at Katoati,
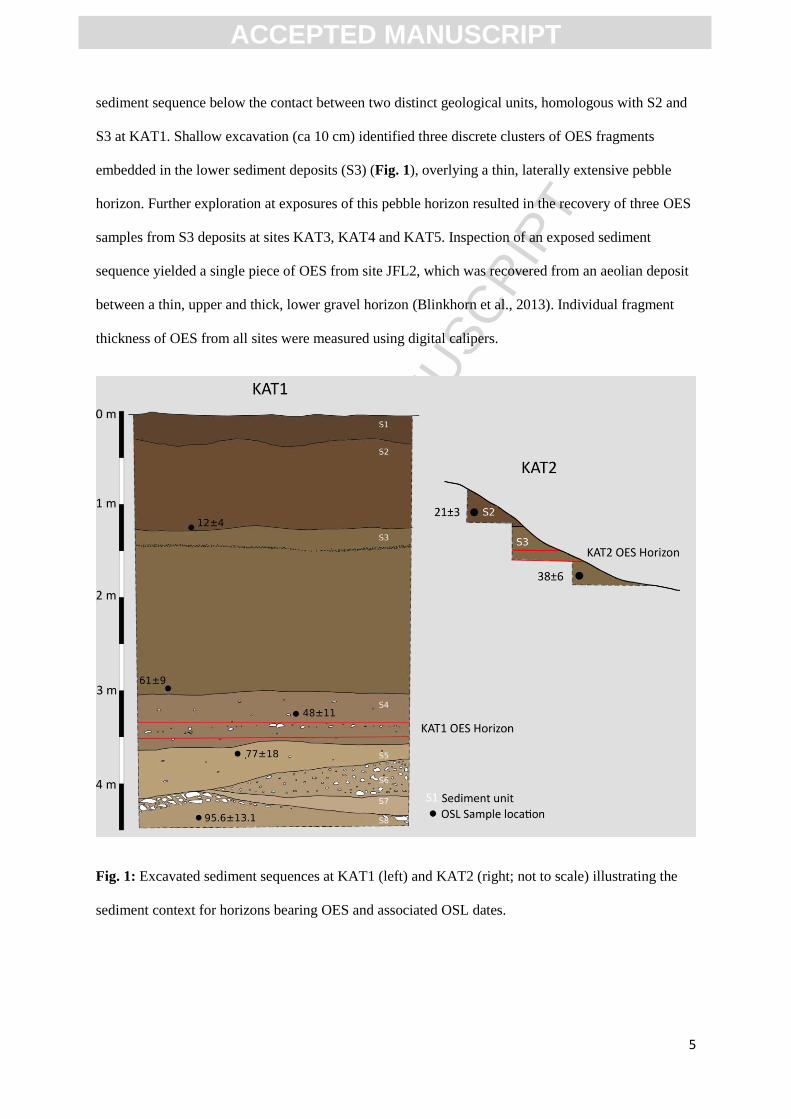
Rajasthan, India. OES fragments have been recovered from six sites, labelled KAT1–5 and JFL2. The
excavation of a 4.5 m trench at KAT1 yielded two pieces of OES. The two pieces were recovered in
10mm screens from sediment excavated at a depth of 3.4 m from surface in horizon S4 (Fig.
1)(Blinkhorn et al., 2013). At KAT2 a cluster of OES was observed eroding from an exposed
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
5
sediment sequence below the contact between two distinct geological units, homologous with S2 and
S3 at KAT1. Shallow excavation (ca 10 cm) identified three discrete clusters of OES fragments
embedded in the lower sediment deposits (S3) (Fig. 1), overlying a thin, laterally extensive pebble
horizon. Further exploration at exposures of this pebble horizon resulted in the recovery of three OES
samples from S3 deposits at sites KAT3, KAT4 and KAT5. Inspection of an exposed sediment
sequence yielded a single piece of OES from site JFL2, which was recovered from an aeolian deposit
between a thin, upper and thick, lower gravel horizon (Blinkhorn et al., 2013). Individual fragment
thickness of OES from all sites were measured using digital calipers.
Fig. 1: Excavated sediment sequences at KAT1 (left) and KAT2 (right; not to scale) illustrating the
sediment context for horizons bearing OES and associated OSL dates.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
6
A total of nine OES samples were submitted and processed for AMS radiocarbon dating.
Eight samples were processed in the Oxford Radiocarbon Accelerator Unit (ORAU). These samples
were sandblasted in aluminium oxide powder, etched in ~0.2 M HCl for 2 minutes, rinsed in distilled
water and dried prior to acid hydrolysis for producing CO2 for graphitisation for AMS dating and
mass spectrometer analysis (see Bronk-Ramsey et al., 2002, 2004a, 2004b for further details). One of
four samples recovered from KAT2 was analysed by Prof. Jay Quade, University of Arizona, Tucson.
Additional dating has been undertaken at KAT2 using Optically Stimulated Luminescence (OSL).
OSL samples were recovered from sediment deposits ca 5 cm above and below the excavated OES
deposits at KAT2, relating to units S2 (above) and S3 (below) from KAT1, and submitted to the
Wadia Institute of Himalayan Geology for analysis (see Supplementary Information for
methodology).
Mass spectrometer analysis of 12 OES samples (including seven samples analysed by ORAU)
was undertaken without physical or chemical abrasion of the surface. Subsamples of OES pieces were
selected and rinsed in ethanol to remove any adhering sediments prior to crushing in an agate pestle
and mortar and drying at 40°C. Oxygen and carbon stable isotopic results were obtained using a VG
Isogas Prism II mass spectrometer with an on-line VG Isocarb common acid bath preparation system.
Each sample was reacted with purified phosphoric acid (H3PO4) at 90°C with the liberated carbon
dioxide cryogenically distilled prior to admission to the mass spectrometer. Both oxygen and carbon
isotopic ratios are reported relative to the VPDB international standard. Calibration was against the in-
house NOCZ Carrara Marble standard with a reproducibility of better than 0.2%. Stable isotope ratios
are expressed using the notation as difference in parts per thousand (permil, ‰) relative to the
standard, calculated as δ‰([Rsample/Rstandard]-1)x1000, where R = 13C/12C or 18O/16O.
3. Results
3.1 Ostrich Eggshell Thickness and Form
Thirty-six measurements of OES thickness indicate a mean thickness of 2.27 mm, with a
range between 2.13 and 2.38 mm. This range matches closely with the reported thicknesses of

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
7
samples from the Indian peninsula, including from Ken River (2.4 mm) (Andrews 1910) and from the
sites of Ledkheri, Chandresal and Anjar (2.1 to 2.2 mm) (Sahni et al., 1990). Collectively, these
samples are all notably larger than a single measurement of 1.7 mm from a sample from Pakistan
(Grellet-Tinner, 2006), but substantially smaller than aepyornithoid-type OES from the Dhok Pathan
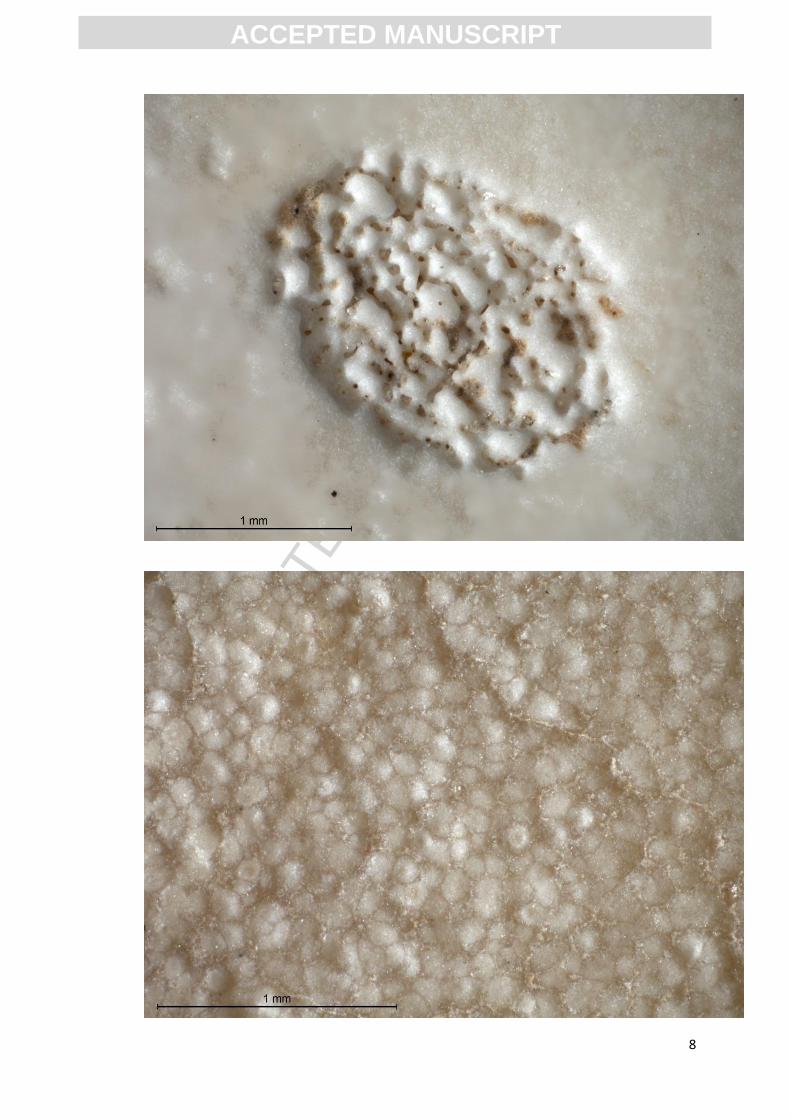
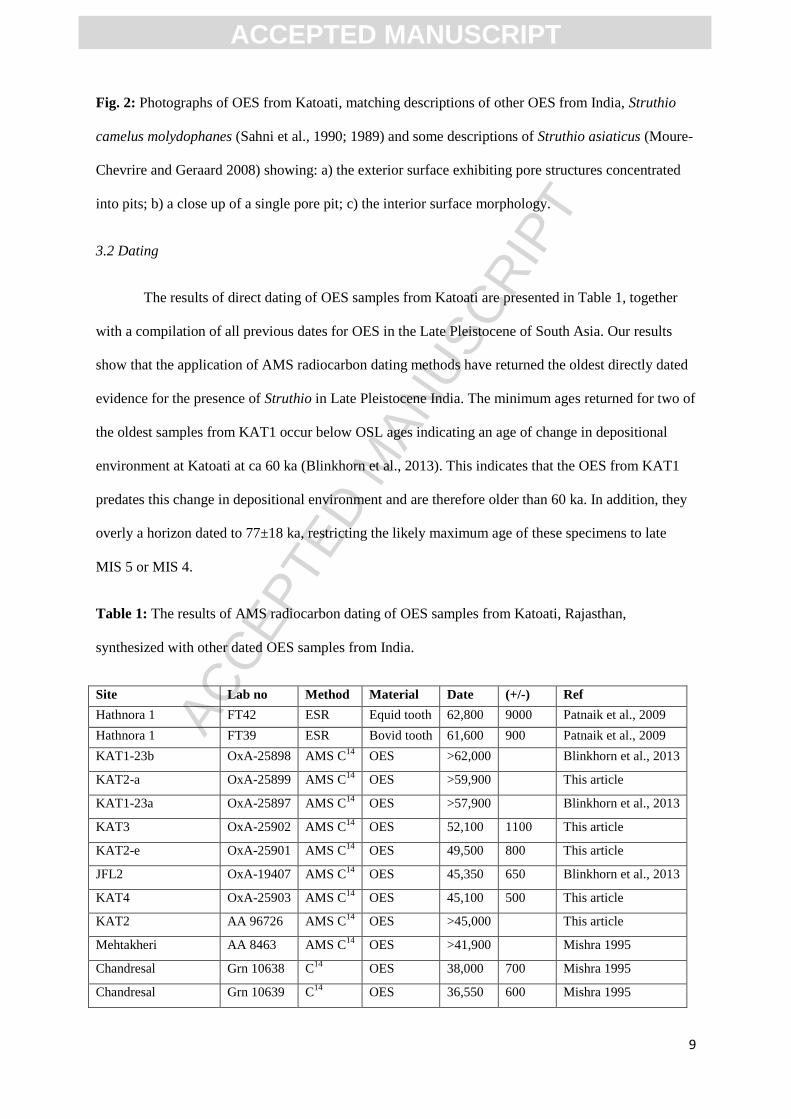
formation at Hasnot with a mean of 2.76 mm (range=2.5 to 2.9 mm)(Patnaik et al., 2009). Pores are
concentrated in spherical to sub-spherical pits (Fig. 2) matching descriptions of previously studied
Late Pleistocene OES from peninsular India (Sahni et al., 1990, 1989), and appear analogous in form
to the pore structures of S. camelus molydophanes (Sahni et al., 1990, 1989) and descriptions of S.
asiaticus (Moure-Chevrire and Geraard 2008).
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
8
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
9
Fig. 2: Photographs of OES from Katoati, matching descriptions of other OES from India, Struthio
camelus molydophanes (Sahni et al., 1990; 1989) and some descriptions of Struthio asiaticus (Moure-
Chevrire and Geraard 2008) showing: a) the exterior surface exhibiting pore structures concentrated
into pits; b) a close up of a single pore pit; c) the interior surface morphology.
3.2 Dating
The results of direct dating of OES samples from Katoati are presented in Table 1, together
with a compilation of all previous dates for OES in the Late Pleistocene of South Asia. Our results
show that the application of AMS radiocarbon dating methods have returned the oldest directly dated
evidence for the presence of Struthio in Late Pleistocene India. The minimum ages returned for two of
the oldest samples from KAT1 occur below OSL ages indicating an age of change in depositional
environment at Katoati at ca 60 ka (Blinkhorn et al., 2013). This indicates that the OES from KAT1
predates this change in depositional environment and are therefore older than 60 ka. In addition, they
overly a horizon dated to 77±18 ka, restricting the likely maximum age of these specimens to late
MIS 5 or MIS 4.
Table 1: The results of AMS radiocarbon dating of OES samples from Katoati, Rajasthan,
synthesized with other dated OES samples from India.
Site Lab no Method Material Date (+/-) Ref
Hathnora 1 FT42 ESR Equid tooth 62,800 9000 Patnaik et al., 2009
Hathnora 1 FT39 ESR Bovid tooth 61,600 900 Patnaik et al., 2009
KAT1-23b OxA-25898 AMS C14
OES >62,000 Blinkhorn et al., 2013
KAT2-a OxA-25899 AMS C14
OES >59,900 This article
KAT1-23a OxA-25897 AMS C14
OES >57,900 Blinkhorn et al., 2013
KAT3 OxA-25902 AMS C14
OES 52,100 1100 This article
KAT2-e OxA-25901 AMS C14
OES 49,500 800 This article
JFL2 OxA-19407 AMS C14
OES 45,350 650 Blinkhorn et al., 2013
KAT4 OxA-25903 AMS C14
OES 45,100 500 This article
KAT2 AA 96726 AMS C14
OES >45,000 This article
Mehtakheri AA 8463 AMS C14
OES >41,900 Mishra 1995
Chandresal Grn 10638 C14
OES 38,000 700 Mishra 1995
Chandresal Grn 10639 C14
OES 36,550 600 Mishra 1995
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
10
KAT2-c OxA-25900 AMS C14
OES 35,210 220 This article
Nagda PRL-854 C14
OES >31,000 Agrawal et al., 1991
Ramgada (Ramnagar) PRL-1196 C14
OES >31,000 Kumar et al., 1990
Bori AA 8461 AMS C14
OES 30,000 420 Mishra et al., 2003
Patne Grn 7200 C14
OES 25,000 200 Sali, 1985
Morgaon AA 8846 AMS C14
OES 22,485 320/310 Mishra et al., 2003
Khapardkhera A 9446 C14
Charcoal 15,680 440/415 Mishra et al., 2003
The chronometric evidence provides robust support for the earlier age estimate from KAT2
(>59.9 ka), and for the overall antiquity of all samples analysed, which occur at the limit of AMS
radiocarbon range. The ages of the four samples from KAT2 either predate or fall within the error
range of the OSL date associated with S3 sediments at the site (38±6 ka), which corroborates the
antiquity of the samples, clearly indicating that they are not related with the younger (21±3 ka) S2
sediments (see Table 2). The dating results from KAT 3 (52.1±1.1 ka) and KAT4 (45.1±0.5 ka), also
derived from the upper part of S3 sediment deposits, yielding similar age estimates. The single sample
collected from JFL2, occurring in a slightly different sediment sequence, returned an age of 45.3±0.6
ka.
Table 2: OSL ages and supporting De and dose rate data for the sediment samples from KAT2.
Lab
Sample
Field
Sample
U
(ppm)
Th
(ppm)
K
%
Mean
De
(Gy)
Least
De
(Gy)
Wt.
mean
De
(Gy)
Dose
Rate
(Gy/ka)
Mean
Age
(ka)
Least
Age
(ka)
Wt.
Mean
Age
(ka)
LD-
1099
KAT2 –
Above
1.5 11.6 1.3 48±5 42±1 46±5 2.2±0.1 21±3 19±1 21±3
LD-
1100
KAT2 –
Below
1.9 13.5 1.3 98±19 79±7 93±13 2.4±0.2 40±8 32±4 38±6
We present the eight oldest directly dated samples of OES from India. Our findings support
similar evidence for the antiquity of OES from gravel deposits of the Surajkund Formation at
Hathnora 1 (Patnaik et al., 2009a). At Hathnora 1 samples of OES pieces were found in association
with an equid and a bovid tooth, which were ESR dated to 61.6 ka and 62.8 ka respectively; a third,
reworked tooth from the same sediment context was dated to 131 ka. As a result, the maximum age of
the sediment context, and therefore the OES, is ca 62 ka. This corroborates the antiquity of previously

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
11
dated samples for which only minimum age estimates could be returned, including at Mehtakheri
(Mishra, 1995), Nagda (Agrawal et al., 1991) and Ramgada (Ramnagar)(Kumar et al., 1990).
Elsewhere, OES samples have been dated between >42 to 22 ka, with the exception of Khapardkhera,
where charcoal associated with an archaeological horizon containing OES beads post-dates the Last
Glacial Maximum (LGM). However, it may be anticipated that numerous other previously dated
samples would return older dates if subject to the same pre-treatment protocols and AMS dating
methods that have been employed at Katoati.
3.3 Stable Isotopes
The results of the first stable isotope analyses of Late Pleistocene OES in India are presented
in Table 3 and Fig. 3. The isotopic composition of OES is characterized as a “snap-shot” of diet,
averaged over 3–5 days, with breeding occurring after the rainy season when sufficient nutrient stores
have been acquired (Johnson et al., 1998). As non-obligate drinkers, a range of fractionation processes
within either surface or plant water sources as well as physiological responses of ostrich to humidity
may affect oxygen isotope data preserved in OES, suggesting these results may be best used only as a
qualitative measure of palaeoclimate (Johnson et al., 1998). δ18
O results from the Katoati sites can be
readily separated into three groups. The majority of samples range between -0.5‰ to 0.5‰, whereas
the two samples from the KAT1 excavated sequence show a large step in enrichment of δ18
O,
returning results of ~6‰, and a similarly large step is observed with KAT3a, returning results of
~12‰. As a result, it can be suggested that the majority of samples relate to eggs produced during
more humid conditions, with those from KAT1 and KAT3a indicating increasing aridity.
Table 3: Results of stable isotope studies of OES samples from Rajasthan; a carbon isotope results
reported from physically and chemically abraded samples produced during AMS radiocarbon dating
at University of Oxford; bstable isotopes results reported from sample dated at University of Arizona.
Sample δ13C‰ δ
13C‰
a δ
18O‰ Age (BP)
KAT1-23a -3.277 -5.1 6.057 >57,900
KAT1-23b -3.413 -4.1 6.391 >62,000
KAT2-a -2.902 -3.9 0.173 >59,900
KAT2-b -2.948 -0.007
KAT2-c -2.969 -2.8 0.139 35,210±220
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
12
KAT2-d -3.095 -0.062
KAT2-e -2.655 -2.8 -0.538 49,500±800
KAT2-f -2.904 0.338
KAT2b -2.57 -0.086 >45,000
KAT3-a -9.917 -9.9 11.937 52,100±1,100
KAT3-b -3.311 0.218
KAT4 -3.015 -2.6 0.038 45,100±500
KAT5 -2.921 0.058
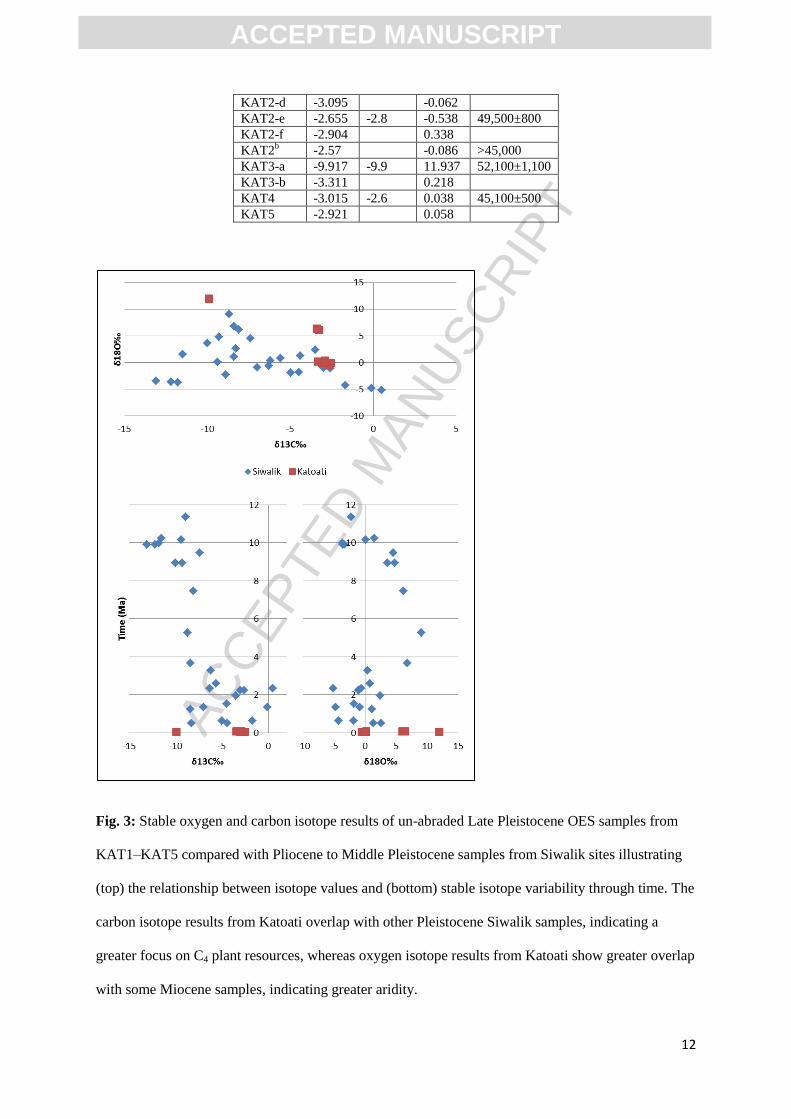
Fig. 3: Stable oxygen and carbon isotope results of un-abraded Late Pleistocene OES samples from
KAT1–KAT5 compared with Pliocene to Middle Pleistocene samples from Siwalik sites illustrating
(top) the relationship between isotope values and (bottom) stable isotope variability through time. The
carbon isotope results from Katoati overlap with other Pleistocene Siwalik samples, indicating a
greater focus on C4 plant resources, whereas oxygen isotope results from Katoati show greater overlap
with some Miocene samples, indicating greater aridity.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
13
Carbon isotope results from OES appear to directly reflect the isotopic composition of the
diet, with an enrichment of ~16‰ (Johnson et al., 1998; Ségalen et al., 2006) such that a pure C3 diet
would produce an average δ13C of -9.5‰ and a pure C4 diet would return results of 4.2‰. Ostriches
show a slight preference for C3 shrubs and select against some vegetation (Johnson et al., 1998),
which could relate to requirement for plant water intake where surface water supplies are not
available. Overall, the results from most samples indicate that the estimated ostrich diets at Katoati
show relatively equal proportions of C3 and C4 input with δ13C of -2 to -3.5‰ relating to a slight bias
toward C3 consumption forming 51.1 to 55.6% of consumed vegetation. The results from KAT3a are
substantially different, indicating a pure C3 diet. The carbon isotope results of samples that were
physically and chemically abraded match well, although not exactly, with un-abraded samples
suggesting potential contamination from secondary carbonate sources have been effectively limited.
Overall, no clear diachronic trends in isotopic composition are evident in this Late Pleistocene
sample.
When compared to isotope results from earlier Siwalik Struthiodiae (Stern et al., 1994), it is
evident that the majority of results from Katoati occupy a tight range within that observed in the
Quaternary period (Fig. 2). Oxygen isotope results from Katoati, which indicated increased aridity
(KAT 23a; KAT 23b; KAT 3a), extend beyond the typical range observed in Indian ratite populations.
Instead, these match results from a limited number of aepyornithoid-type Pliocene specimens. This
may represent aridity experienced at an individual, rather than population-wide, scale. Similarly, only
the carbon isotope results from KAT3a extend beyond the variability observed in ratite δ13
C‰ from
the Siwalik Quaternary sequence, but within the range of Miocene samples, again potentially the
result of individual scale variation.
3.4 Geographic Distribution
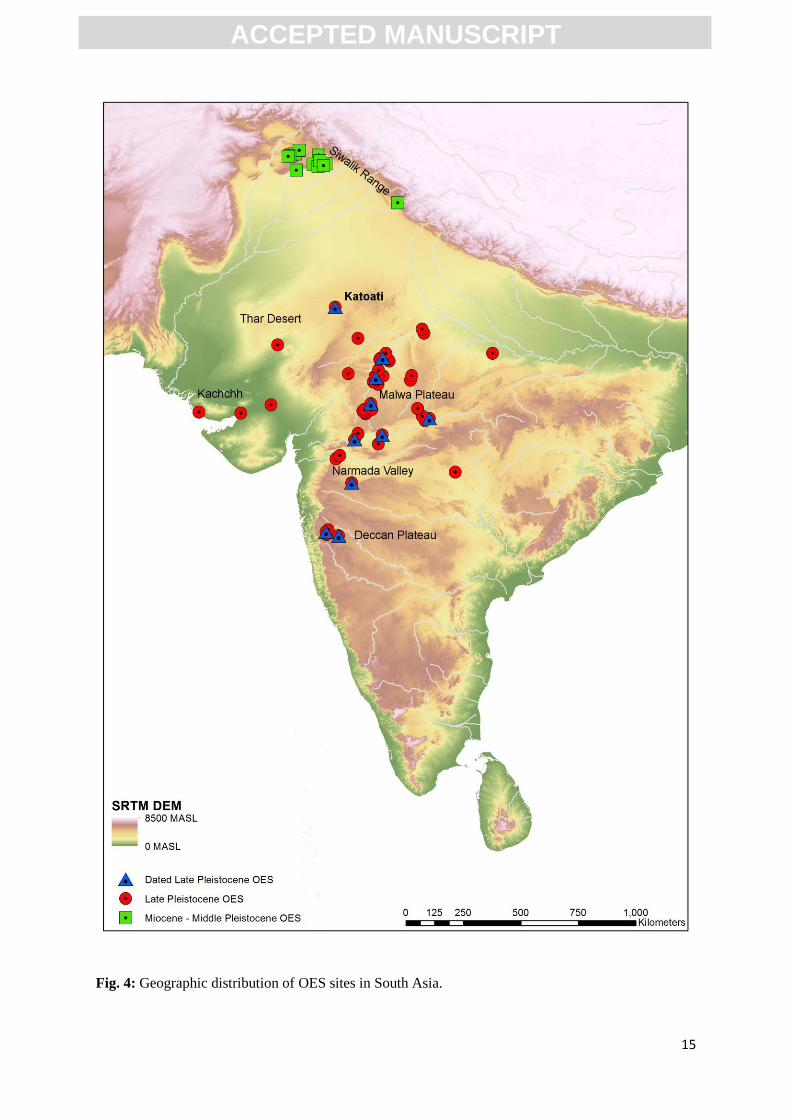
The distribution of sites with OES in South Asia is presented in Fig. 4. This illustrates a clear
spatial distinction between the range of Late Pleistocene sites in peninsular India and the earlier

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
14
Siwalik sites. The majority of Late Pleistocene occurrences of OES in India occur in the Chambal and
Narmada valleys, associated with the Malwa Plateau of west-central India. Fewer sites are reported
from the Tapi valley and between the headwaters of the Krishna and Bhima Rivers on the Deccan
Plateau in south-central India. A small number of sites, including Katoati, are notable in that they not
located in the immediate vicinity of one of the subcontinents large drainage basins. Nevertheless, the
landscape at Katoati indicates that the excavated sites occur at the edge of a braided stream network
that may have ultimately fed into the Luni River. Three sites located in Kachchh, Gujarat, are located
close to smaller fluvial regimes in near-coastal locations. Sites bearing OES are reported in both in
lowland or wide valley floor contexts, as well as within regions displaying more relief, occurring at
the edge of different watersheds. In contrast, Pliocene to Middle Pleistocene OES sites are restricted
to the foothills of the Himalaya and the Siwalik ranges and occur in the upper reaches of the Indus
drainage system.
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
15
Fig. 4: Geographic distribution of OES sites in South Asia.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
16
Despite the occurrence of Late Pleistocene OES in a wide variety of landscape contexts, their
distribution appears geographically limited, predominately associated with the Malwa Plateau and
associated drainage systems. Occupation of the Deccan Plateau is significantly more limited to a
cluster of sites close to Pune, Mahrashtra, and a single site at the watershed of the Tapi River.
Combined, these sites present a SW–NE trend, although Katoati, the Kachchh sites, Ken River and
Shindi are clear exceptions to this. The earliest dated examples of OES in India, dating between >60–
35 ka at Katoati and Hathnora, appear in river systems that flow to the west/south-west. The
concentration of sites on the Malwa Plateau that are associated with eastward flowing drainage
networks mostly date between >40 to >30ka, with the exception of Khapardkhera, which dates to
16ka. The dated sites from the Deccan Plateau, also relating to eastward flowing river systems, are
younger than both of these groups, dating to 30–22 ka.
4. Discussion
The presence of ostrich in Late Pleistocene India appears to be restricted both temporally and
spatially, and marks a significant change in geographic range compared to the Miocene to Middle
Pleistocene. The earliest evidence for ostrich in Late Pleistocene India occurs in contexts dating to
>60 ka, in MIS 4, at the margins of the Thar Desert and the westward draining Narmada basin. While
ostrich are present at Katoati and the Narmada basin during MIS 3, the range of ostrich expands in
this period to include the Malwa Plateau. With the onset of MIS 2, evidence for ostrich is only found
on the north western Deccan Plateau. Only a single example of ostrich, at Khapardkhera, on the
Malwa Plateau, is known from India post-dating the LGM. In order to further assess the timing and
nature of ostrich expansion into India, it is necessary to situate these findings in broader context.
4.1 Chronology
The presence of ostrich in India currently appears restricted from MIS 4 to MIS 2 (>60–16
ka). The earliest dated evidence for ostrich, occurring before 60 ka (MIS 4), stretches beyond the
range of traditional radiocarbon methods, although earlier dates from Katoati are suggested by OSL
dating. Therefore the use of dating methods, such as OSL, is necessary to corroborate the antiquity of

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
17
the samples. As this has typically not been the case in South Asia, expansion of ostrich into western
and central India prior to MIS 4 cannot be precluded. Nevertheless, both environmental and
ecological evidence reviewed below may further support the appearance of ostrich in western and
central India during MIS 4.
Khaparkeda presents the only evidence for the survival of ostrich in India <20 ka. The
regional extinction of ostrich from India is suggested to have occurred across the LGM, perhaps
resulting from extreme climatic pressure and increased competition and human predation (see below).
The presence of numerous OES samples dating to before the LGM and the scarcity of similar samples
dating after the LGM supports the suggestion of significant collapse of ostrich populations at this
time, which ultimately leads to their regional extinction. Based on examples of Struthio extirpation in
North Asia (Janz et al., 2009) and numerous megafaunal taxa at a global scale (Koch and Barnosky,
2006), an alternative hypothesis may suggest the regional extinction of ostrich around the
Pleistocene–Holocene boundary with the return to interglacial climates. However, as yet there is no
evidence to support continuity of ostrich populations in India into the Holocene, and the unique,
mosaic nature of Indian habitats may indicate that the global collapse of megafaunal diversity ca 10
ka is not a suitable analogy (Roberts et al., 2014).
4.2 Taxonomy
The Late Pleistocene ostrich population represented by OES in peninsula India can be most
securely referred to as Struthio sp. In their comparison of OES samples from late Miocene and
Pliocene of East Africa and Namibia, Harrison and Msuya (2005) identify a pattern of decreasing
shell thickness and pore diameter and increasing pore density through time between S. karingarbensis
(shell thickness 2.9–3.2; pore density 2.2/cm2; pore diameter 2.7 mm; age range - 6.5–4.2 Ma.), S.
kakesiensis (Shell thickness 2.5–4.4 mm, mean=3.2; pore density=4.6/cm2; pore diameter =2.5 mm;
age range 4.5–3.6 Ma.), S. daberasensis (Shell thickness 1.7–2.6 mm; mean=2.3; pore density=4–
5/cm2; pore diameter=2.2 mm; age range 6.5–3 Ma.) and S. camelus (Shell thickness 1.5–2.8 mm;
mean = 2.1; pore density 10.8/cm2; pore diameter=1 mm; age range 3.8 Ma. to present). It is worth

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
18
noting that dated OES from Siwalik deposits bearing struthionid-type morphotypes only overlap
chronologically with S. camelus. OES comparable to S. camelus molydophanes associated with
osteological remains identified as S. asiaticus in Late Pliocene deposits at Ahl al Oughlam, Morocco,
exhibit a shell thickness (2.3 to 2.7 mm; mean=2.54 mm) that overlaps the range of S. daberasensis
but a greater pore density (10–20/cm2) comparable to S. camelus, thus fitting the broad trend (Moure-
Chevrire and Geraard 2008). The decreased shell thickness in the Late Pleistocene ostrich population
in India with high pore density continues this African trend. However, while the thickness range
overlaps with S. asiaticus samples from Ahl al Oughlam, the Indian samples are notably thinner.
The occurrence of OES in Central and East Asia share some similarities with South Asia.
The recovery of aepyornithoid-type eggshells from Miocene deposits followed by struthionid forms
appearing in the Pliocene deposits in Mongolia and reported as Struthiolithus (Sauer 1972) parallel
the records from the Siwalik Range. Notably, the samples, originally reported as Struthiolithus,
exhibit thinner shells than their African counterparts, ranging between 1.8–2.4 mm. In addition, the
struthionid-type eggshells exhibit needle point pores rather than pore-pits observed in the
contemporary African sample. A rich collection of Upper Pleistocene specimens dated using C14
methods are reported from China, Mongolia and Transbaikal Russia (Janz et al., 2009; Kurochkin et
al., 2010). Further descriptions of their morphology (i.e. shell thickness, pore structure and density)
are required before meaningful comparison can be made with the South Asian sample.
The direct ancestry and taxonomic affiliation of Late Pleistocene Struthio in western and
central India is difficult to assess based on ootaxonomy alone, particularly as a number of potential
source populations exist. They may represent descendants of the Siwalik populations, a more
widespread population of S. asiaticus that extends from the Siwalik Range to Morocco and potentially
including South Africa, more recent Struthio populations found within this range, such as those
known from Arabia (Potts, 1999) including Late Pleistocene samples from the Mundafan depression
(Crassard et al., 2013), or of shared source population with Central and East Asian Struthio
populations. While it may not be possible to identify which source population, and therefore which

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
19
taxon, expanded into western and central India in the Late Pleistocene, it is possible to explore why
this expansion occurred.
4.3 Neogene and Quaternary habitats of Struthiodiae in South Asia
Significant changes to the geographic and ecological make up of South Asia have occurred
since the earliest known appearance of ostrich, and particularly in the mountainous northern region
which has yielded all evidence for ostrich prior to the Late Pleistocene. Himalayan orogeny and
associated changes to Eurasian climate systems, particularly the evolution of the Asian monsoons, are
likely to have had significant impacts upon ostrich populations. Studies of Himalayan exhumation
rates indicate a decrease in monsoonal intensity after 10 Ma., following the mid Miocene climatic
optimum, followed by an enhanced monsoon system after 4 Ma. (Clift et al., 2008). Isotopic studies of
pedogenic carbonates have indicated a sharp change to C4 dominated ecologies in the Late Miocene
(Cerling et al., 1997; Quade and Cerling 1995), but more recent studies have suggested lateral
variability in floral gradients prior to C4 expansions and greater heterogeneity in floral change
(Behrensmeyer et al., 2007; Morgan et al., 2009). Similarly the expansion of C4 flora has a notable but
complex effect on faunal isotopic signatures (Barry et al., 2002; Morgan et al., 1994).
Two distinct phases in Siwalik OES isotope chemistry can clearly be identified. Mid-Miocene
samples older than ca 9 Ma., exclusively comprising specimens with aepyornithoid-type pore patterns,
present a strong C3 signature associated with relatively humid conditions. Pleistocene samples,
predominately comprising specimens with struthionid-type pore patterns, present a strong C4
signature associated with relatively humid conditions. Limited sample size prevents a clear
characterisation of Late Miocene and Pliocene OES isotope chemistry, although the carbon isotope
signature is intermediate between the range of the mid-Miocene and Pleistocene groups whereas the
oxygen isotope signature indicates increased aridity in contrast to both.
These results broadly corroborate existing trends identified in other isotope archives,
particularly highlighting the appearance of C4 plants in the Siwaliks and their exploitation by
Pleistocene ostrich populations. The apparent synchronicity of the appearance of a C4 signature in

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
20
OES isotope chemistry and the appearance of struthionid-type pore patterns may indicate that the
appearance of new ostrich populations in the Siwalik sequence was related to wide ranging ecological
reconfiguration. However, the continuity of specimens with aepyornithoid-type pore patterns that
substantially overlaps the appearance of C4 flora and OES with struthionid-type pore patterns suggests
that existing ostrich populations were not rapidly replaced. In this context, the Late Pleistocene
expansion of ostrich into western and central India exhibits the exploitation of a similar resource base
with earlier Pleistocene ostrich populations, focused upon C4 habitats.
Extirpation of Siwalik ostrich populations may have been the result of changing relief,
environments, impacts of climate change on seasonality or a combination of these factors, and
identifying which factor may have been dominant in this process is beyond resolution of data
available. However, these Siwalik habitats differed significantly in their geographic structure and
climatic seasonality from the regions of western and central India into which ostrich populations
expanded during the Late Pleistocene.
4.4 Geographic and Palaeoenvironmental context of Late Pleistocene ostrich expansion in western
and central India
Given the known distribution of Struthiodiae spanning the Miocene–Pleistocene, and the
geographical and ecological makeup of north India and Pakistan, the Late Pleistocene expansion of
ostrich into western and central India is most likely to have been the result of dispersals from the
north-west of the subcontinent. Indeed, the NW–SE cline noted in the dated occurrences of OES in
western and central India appears to support this. Geographic and environmental conditions in the
Thar Desert, marking the most north-westerly extremity of the Late Pleistocene ostrich range in India,
are therefore critical to understanding the timing and nature of ostrich expansions.
An expansion into the Thar Desert, whether eastwards from Africa and Arabia or southeast-
wards from the Siwalik Range, requires a crossing of the Indus River. A GIS model of eastward
dispersal routes across southern Asia, based on least cost surface models, indicates the Indus as a
major barrier, indicating that any expansions would likely be directed in to the interior of the Thar

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
21
Desert to be capable of traversing these river systems (Field et al., 2007). Increased humidity
experienced across southern Asia during MIS 5 would have both provided an important context both
for population expansion and an increased potential for river systems to prevent major changes to
geographic ranges. While the Indus may have been a major physical barrier to dispersals, it is likely to
have also acted as a refugium for flora and fauna alike during periods of increased aridity, such as
MIS 4, delivering a constant supply of water from the Himalaya through landscapes lacking direct
precipitation. In this context, the Indus corridor provides the most likely direct source for ostrich
expansions into western and central India. Indeed, during glacial conditions such as MIS 4, changes to
river channel morphology in the mid and lower Indus valley modulated by varying flow and sediment
supply may have provided the most suitable conditions for the crossing of this physical barrier. This
provides the immediate context for the appearance of Late Pleistocene ostrich in India.
The Thar Desert provides the best recorded evidence for Late Pleistocene environmental
change in South Asia and suggests humid conditions were present for much of MIS 3, with evidence
for faltering fluvial systems and the onset of dune mobility occurring by 35 ka (see Blinkhorn 2014).
During this period of faltering humidity in MIS 3, ostrich appear to successfully expand to the
majority of its known range in India, encompassing all dated sites within and to the north of the
Narmada Valley. The occurrence of ostrich in Arabia in this time period has also been recently
reported, dating to 49.8 ka and >50 ka (Crassard et al., 2013). Increasing aridity occurs during leading
up to the LGM, evident in the onset of sand dune mobility commencing in the central Thar Region
and extending to the mouth of the Narmada. It is during the transition between MIS 3–2 and prior to
the LGM that the geographic range of ostrich in India reaches its southernmost extent on the north-
west Deccan Plateau. Only the site of Kharpakeda indicates the limited continuity of ostrich
populations on the Marwa plateau across the LGM.
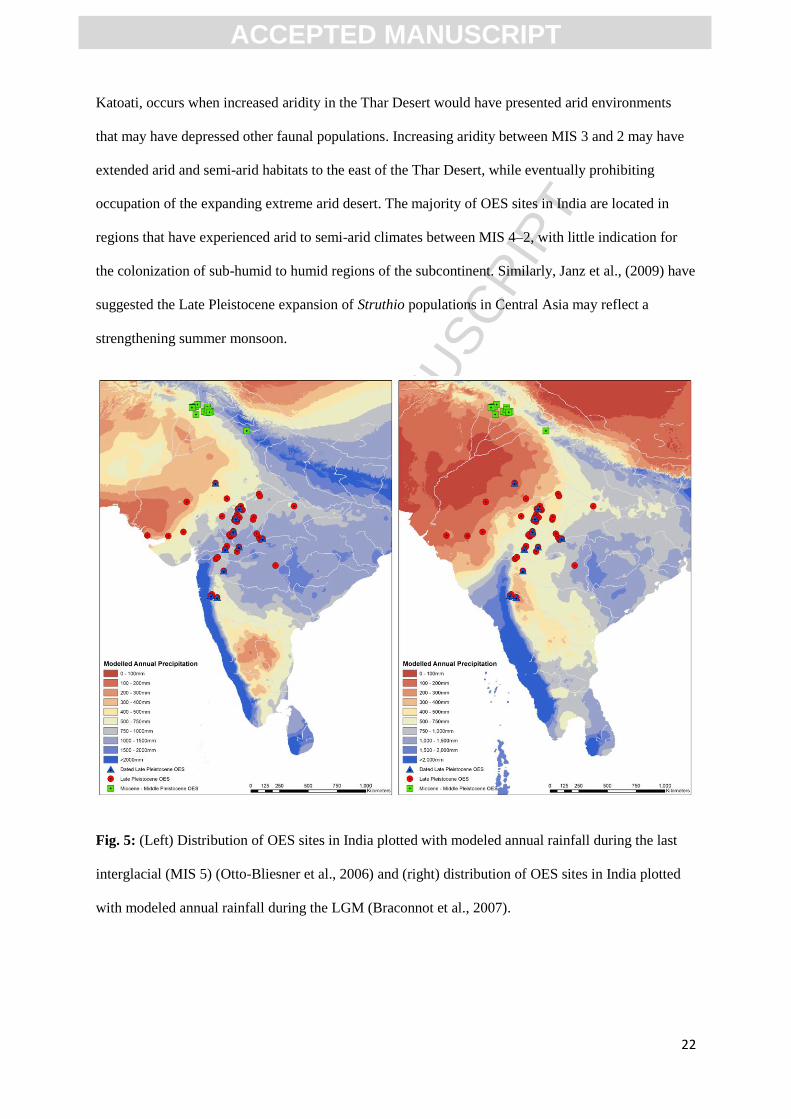
Fig. 5 presents the distribution of OES sites in India plotted with modelled rainfall during
peak humidity in MIS 5 (which may also serve as an appropriate analogy for early MIS 3 given the
presence of terrestrial proxies directly indicating humid conditions) and during the LGM (a suitable,
but more arid analogue for MIS 4). The earliest evidence for Late Pleistocene ostrich in India, at
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
22
Katoati, occurs when increased aridity in the Thar Desert would have presented arid environments
that may have depressed other faunal populations. Increasing aridity between MIS 3 and 2 may have
extended arid and semi-arid habitats to the east of the Thar Desert, while eventually prohibiting
occupation of the expanding extreme arid desert. The majority of OES sites in India are located in
regions that have experienced arid to semi-arid climates between MIS 4–2, with little indication for
the colonization of sub-humid to humid regions of the subcontinent. Similarly, Janz et al., (2009) have
suggested the Late Pleistocene expansion of Struthio populations in Central Asia may reflect a
strengthening summer monsoon.
Fig. 5: (Left) Distribution of OES sites in India plotted with modeled annual rainfall during the last
interglacial (MIS 5) (Otto-Bliesner et al., 2006) and (right) distribution of OES sites in India plotted
with modeled annual rainfall during the LGM (Braconnot et al., 2007).
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
23
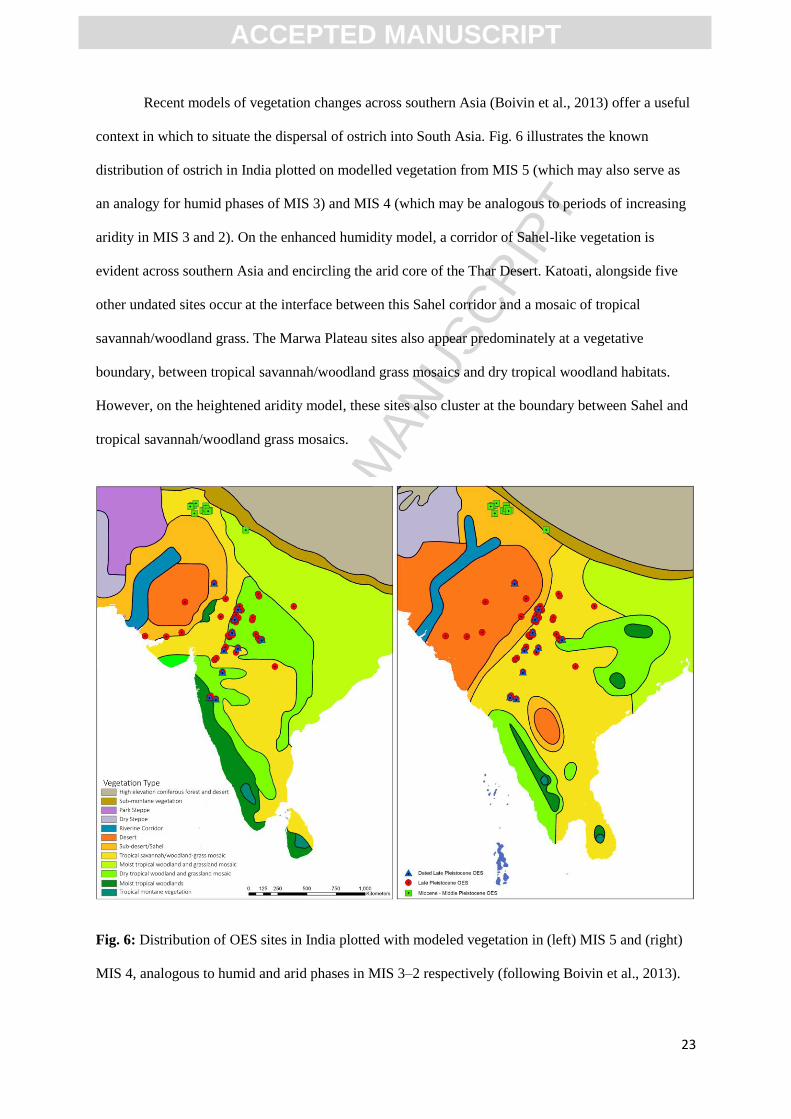
Recent models of vegetation changes across southern Asia (Boivin et al., 2013) offer a useful
context in which to situate the dispersal of ostrich into South Asia. Fig. 6 illustrates the known
distribution of ostrich in India plotted on modelled vegetation from MIS 5 (which may also serve as
an analogy for humid phases of MIS 3) and MIS 4 (which may be analogous to periods of increasing
aridity in MIS 3 and 2). On the enhanced humidity model, a corridor of Sahel-like vegetation is
evident across southern Asia and encircling the arid core of the Thar Desert. Katoati, alongside five
other undated sites occur at the interface between this Sahel corridor and a mosaic of tropical
savannah/woodland grass. The Marwa Plateau sites also appear predominately at a vegetative
boundary, between tropical savannah/woodland grass mosaics and dry tropical woodland habitats.
However, on the heightened aridity model, these sites also cluster at the boundary between Sahel and
tropical savannah/woodland grass mosaics.
Fig. 6: Distribution of OES sites in India plotted with modeled vegetation in (left) MIS 5 and (right)
MIS 4, analogous to humid and arid phases in MIS 3–2 respectively (following Boivin et al., 2013).

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
24
The heterogeneous habitats of the Indian subcontinent appear to have been critical to the
continuity of large faunal taxa in India. Faunal records from the Billasurgum caves, south India,
suggest 20 of 21 large animal species have remained present in the Indian subcontinent over the Late
Pleistocene, with the disappearance of Theropithecus marking the only example of extinction (Roberts
et al., 2014). Nevertheless, a number of species, such as wild ass and Indian rhinoceros, now only
appear in restricted distributions within the Indian subcontinent (Roberts et al., 2014). Fragmentation
and spatial redistribution of India’s habitat mosaic appears to have played a critical role in faunal
mobility and long term continuity in the region (Roberts et al., 2014). The arrival of ostrich in western
and central India before 60 ka appears to have occurred during such a period of habitat redistribution.
The high levels of mobility of the ostrich populations may have been critical to effectively exploit
changing habitat boundaries and compete with Indian fauna, particularly with the extension of Sahel-
like vegetation in the region. Nevertheless, there is little evidence to suggest the successful ostrich
colonization of endemic Indian habitats. It remains unclear what caused the regional extinction of
ostrich from India, although the lack of an endemic source population, competition for marginal
resources and increased predation from Indian fauna over the LGM, as well as exploitation by rapidly
growing modern human populations may all have played some role.
4.5 The importance of ostrich expansion into India for the human dispersal debate
The ostrich is one of two megafaunal species known to have expanded into western and
central India during the Late Pleistocene, the other being Homo sapiens. The dispersal of ostrich into
western and central India before 60 ka offers an important opportunity to assess a number of aspects
of the modern human dispersal debate. Meanwhile, the demographic expansion of human populations
within India may be significant in explaining the regional extinction of ostrich, either through
predation or indirect impacts upon regional ecology.
The appearance of ostrich in Late Pleistocene India prior to 60 ka occurs before the time
frame suggested by some for the appearance of modern humans in India, between 60 to 50 ka (e.g.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
25
Mellars et al., 2013). The argument for a dispersal of modern humans after 60 ka favours a rapid
coastal dispersal in to India on account of advanced technological and social adaptations, with a
timeframe based on mtDNA evidence (Mellars et al., 2013). Ostrich expansions into India during the
Late Pleistocene indicate continental routes of dispersal into India were also possible. Predation of
ostrich and use of OES by contemporary and prehistoric human populations indicate significant
geographic overlap of humans and ostriches in Sahel-like and savannah habitats (Cooper et al., 2009).
This suggests that the continental routes of expansion exploited by ostrich populations prior to 60ka,
focused upon Sahel-like and savannah habitats, may also have been habitable to human populations
and offer an alternative dispersal route to those proposed by coastal models.
Ostrich beads and symbolic pieces play a role in the identification of behavioural modernity
in the archaeological record, both in Africa (e.g. d’Errico and Stringer, 2011) and in South Asia
(James and Petraglia, 2005; Mellars et al., 2013). The presence of OES beads, and fragments incised
with hatched patterns, such as at Patne (Sali, 1985), is suggested to support a dispersal of human
populations associated with these material behaviours at the onset of MIS 3 (Mellars et al., 2013).
Yet, alternate models argue for earlier expansions of modern humans into South Asia during MIS 5
(see Blinkhorn and Petraglia 2014), possibly preceding the arrival of ostrich. Resolving the timeframe
of the earliest expansion of ostrich into western and central India is therefore critical to assess whether
OES may have been available to human populations throughout MIS 5. An earlier arrival of human
populations would preclude the use of OES, and the appearance of symbolic material culture
produced using OES may indicate regional innovations in response to local demographic and
environmental pressures (Petraglia et al., 2009) rather than an innate feature of modern human
behaviour. Nevertheless, ostrich appear to have been recorded, albeit rarely, in the rich corpus of rock
art known from South Asia, such as at the site of Firengi, south of Bhopal (Badam, 2005). Evidence
for ostrich regional extinction prior to the Holocene supports the suggestion that rock art was being
created in India during the late phases of the Late Pleistocene (Taçon et al., 2010).
If the extension of Sahel-like vegetation across South Asia was a key factor in the dispersal of
both ostrich and humans, potential dispersals of both taxa during MIS 5 should be considered based

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
26
on available palaeoenvironmental reconstructions. However, modern humans appear to have been
uniquely successful in expanding beyond these Sahel-like habitats, to which ostrich appear restricted.
The successful adaptation of modern humans to new habitats and changing environments may have
also impacted upon the regional extinction of ostrich. Overlapping factors of environmental
deterioration and demographic expansion appear to have provided a context for technological
innovation around the MIS 3–2 boundary, associated with the emergence of more efficient hunting
technologies and the earliest evidence for human-modified OES (Petraglia et al., 2009). Whether
through direct predation of ostrich and their eggs, or more indirect ecological impacts on increasingly
fragmented habitats, the growing population of modern humans in South Asia is likely to have been a
factor contributing to the regional extinction of ostrich from India.
5. Summary:
Our results of direct dating of OES samples from Katoati, along with dating of their sedimentary
context, present the first robust evidence to indicate a significant Late Pleistocene range expansion of
ostrich in India before 60 ka. By synthesising these results with existing reports and placing them
within their palaeoenvironmental and ecological context, a restricted timespan for the occurrence of
ostrich expansion into peninsula India, limited to MIS 3–2, has been demonstrated. The range of Late
Pleistocene ostrich in India now appears to track the margin of Sahel-like habitats, with limited
evidence to suggest any extensive colonisation of regions with endemic Indian vegetation. The
dispersal of ostrich into India during the Late Pleistocene is notable in the context of long-term
stability in the diversity of mega-fauna in the region, as well as potential analogies for the more
successful dispersals of modern humans.
Acknowledgements
The fieldwork was supported by the Emslie Horniman Scholarship (Royal Anthropological Institute,
London) awarded to JB. AMS radiocarbon dating at ORAU was funded by the NERC Radiocarbon
Facility (2011/2/16) awarded to MP and JB. JB is supported by a Fondation Fyssen Post-Doctoral
Fellowship. MP is supported by a grant from the European Research Council (no. 295719). We thank

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
27
Jay Quade (University of Arizona) for AMS dating an OES sample from KAT2, Peter Ditchfield
(University of Oxford) for facilitating the stable isotope analysis, and N. Suresh (Wadia Institute of
Himalayan Studies, Dheradun) for dating the OSL samples from KAT2 and providing the
methodology and figures reported in SI. JB and HA thank the Anna University for their support and
facilities. We thank Francesco d’Errico for Fig. 2.
References
Agrawal D.P., Kusumgar S., Yadava M., 1991. Physical Research Laboratory Radiocarbon Date List
VI. Radiocarbon 33, 329 –344.
Andrews C.W., 1911. Note on some Fragments of the Fossil Egg-shell of a large Struthious Bird from
Southern Algeria, with some Remarks on some Pieces of the Egg-shell of an Ostrich from
Northern India. Verhandlungen V Internationale Ornithologie Kongress, Berlin, 1910, pp 169 –
174.
Badam G.L., 2005. A note on the Ostrich in India since the Miocene. Man and Environment 30, 97 –
104.
Barry, J. C., Morgan, M. E., Flynn, L. J., Pilbeam, D., Behrensmeyer, A. K., Raza, S. M., Khan, I. A.,
Badgley, C., Hicks, J., Kelley, J., 2002. Faunal and environmental change in the late Miocene
Siwaliks of northern Pakistan. Paleobiology 28, 1-71.
Behrensmeyer, A. K., Quade, J., Cerling, T. E., Kappelman, J., Khan, I. A., Copeland, P., Roe, L.,
Hicks, J., Stubblefield, P., Willis, B. J., Latorre, C., 2007. The structure and rate of late Miocene
expansion of C4 plants: Evidence from lateral variation in stable isotopes in paleosols of the
Siwalik Group, northern Pakistan. Geological Society of America Bulletin 119, 1486-1505.
Bidwell E., 1910. Remarks on some fragments of egg shell from a fossil ostrich in India. Ibis IV, 759–
761.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
28
Blinkhorn, J., 2014. Late Middle Palaeolithic surface sites occurring on dated sediment formations in
the Thar Desert. Quaternary International in press.
Blinkhorn J., Achyuthan H., Petraglia M., Ditchfield P., 2013. Middle Palaeolithic occupation in the
Thar Desert during the Upper Pleistocene: the signature of a modern human exit out of Africa?
Quaternary Science Reviews 77, 233 – 238.
Blinkhorn, J., Petraglia, M. D., 2014. Assessing Models for the Dispersal of Modern Humans to South
Asia. In: Dennell, R., Porr, M. (Eds.), Southern Asia, Australia and the Search for Human
Origins. Cambridge University Press, Cambridge, pp. 64 – 75.
Boivin N., Fuller D.Q., Dennell R., Allaby R., Petraglia M.D., 2013. Human dispersal across diverse
environments of Asia during the Upper Pleistocene. Quaternary International 300, 32–47.
Braconnot P., Otto-Bliesner B., Harrison S., Joussaume S., Peterchmitt J.-Y., Abe-Ouchi A., Crucifix
M., Driesschaert E., Fichefet T., Hewitt C.D., Kageyama M., Kitoh A., Laîné A., Loutre M.-F.,
Marti O., Merkel U., Ramstein G., Valdes P., Weber S.L., Yu Y., Zhao Y., 2007. Results of
PMIP2 coupled simulations of the Mid-Holocene and Last Glacial Maximum – Part 1:
experiments and large-scale features. Climate of the Past 3, 261–277.
Bronk-Ramsey C., Higham T.F.G., Bowles A., Hedges R.E.M., 2004a. Improvements to the
pretreatment of bone at Oxford. Radiocarbon 46, 155–163.
Bronk-Ramsey C., Higham T.F.G., Leach P., 2004b. Towards high-precision AMS: progress and
limitations. Radiocarbon 46, 17–24.
Bronk-Ramsey C., Higham T.F.G., Owen D.C., Pike A.W.G., Hedges R.E.M., 2002. Radiocarbon
Dates from the Oxford Ams System: Archaeometry Datelist 31. Archaeometry 44, 1–150.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
29
Cerling, T. E., Harris, J. M., MacFadden, B. J., Leakey, M. G., Quade, J., Eisenmann, V., Ehleringer,
J. R., 1997. Global vegetation change through the Miocene/Pliocene boundary. Nature 389, 153-
158.
Clift, P. D., Hodges, K. V., Heslop, D., Hannigan, R., Van Long, H., Calves, G., 2008. Correlation of
Himalayan exhumation rates and Asian monsoon intensity. Nature Geoscience 1, 875-880.
Cooper R.G., Mahrose K.M.A., Horbanczuk J.O., Villegas-Vizcaíno R., Kennou Sebei S., Faki
Mohammed A.E., 2009. The wild ostrich Struthio camelus : a review. Tropical Animal Health
and Productivity 41, 1669–1678.
Crassard R., Petraglia M.D., Drake N. A, Breeze P., Gratuze B., Alsharekh A., Arbach M., Groucutt
H.S., Khalidi L., Michelsen N., Robin C.J., Schiettecatte J., 2013. Middle palaeolithic and
neolithic occupations around Mundafan Palaeolake, Saudi Arabia: implications for climate
change and human dispersals. PloS one 8, e69665.
d’Errico F., Stringer C.B., 2011. Evolution, revolution or saltation scenario for the emergence of
modern cultures? Philosophical transactions of the Royal Society of London. Series B Biological
Sciences 366, 1060–9.
Davies W., 1880. On some fossil bird-remains from the Siwalik Hills in the British Museum. The
Geological Magazine 7, 18–27.
Dennell R., 2004. Hominid dispersals and asian biogeography during the lower and early Middle
Pleistocene, c. 2.0-0.5 mya. Asian Perspectives 43, 205 – 226.
Dennell R.W., 2005. The Solo Ngandong Homo erectus assemblage : a taphonomic assessment.
Archaeology in Oceania 40, 81–90.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
30
Field J.S., Petraglia M.D., Lahr M.M., 2007. The southern dispersal hypothesis and the South Asian
archaeological record: Examination of dispersal routes through GIS analysis. Journal of
Anthropological Archaeology 26, 88–108.
Grellet-Tinner G., 2006. Phylogenetic interpretation of eggs and eggshells: implications for
phylogeny of Palaeognathae. Alcheringa: An Australasian Journal of Palaeontology 30, 141–
182.
Harrison T., Msuya C.P., 2005. Fossil struthionid eggshells from Laetoli , Tanzania : Taxonomic and
biostratigraphic significance. Journal of African Earth Sciences 41, 303–315.
Janz L., Elston R.G., Burr G.S., 2009. Dating North Asian surface assemblages with ostrich eggshell:
implications for palaeoecology and extirpation. Journal of Archaeological Science 36, 1982–
1989.
Johnson B.J., Fogel M.L., Miller G.H., 1998. Stable isotopes in modern ostrich eggshell : A
calibration for paleoenvironmental applications in semi-arid regions of southern Africa.
Geochimica et Cosmochimica Acta 62, 2451–2461.
Koch P.L., Barnosky A.D., 2006. Late Quaternary extinctions: state of the debate. Annual Review of
Ecology, Evolution, and Systematics, 37, 215–250.
Kumar G., Sahni A., Pancholi R.K., Narvare, G., 1990. Archaeological discoveries and a study of
Late Pleistocene Ostrich egg shells and egg shell objects in India. Man and Environment, 15, 29
– 40.
Kurochkin E.N., Kuzmin Y. V., Antoshchenko-Olenev I. V., Zabelin V.I., Krivonogov S.K., Nohrina
T.I., Lbova L. V., Burr G.S., Cruz R.J., 2010. The timing of ostrich existence in Central Asia:
AMS 14C age of eggshells from Mongolia and southern Siberia a pilot study. Nuclear
Instruments and Methods in Physics Research Section B: Beam Interactions with Materials and
Atoms, 268, 1091–1093.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
31
Lydekker R., 1884. Vol. III Siwalik Birds. Palaeontologia Indica: Indian Tertiary and Post-Tertiary
Vertebrata. Geological Survey of India, Calcutta.
Manegold A., Louchart A., Carrier J., Elzanowski A., 2013. The Early Pliocene avifauna of
Langebaanweg South Africa : a review and update. Proceedings 8th International Meeting
Society of Avian Paleontology and Evolution 135–152.
Mellars P., Gori K.C., Carr M., Soares P. A., Richards M.B., 2013. Genetic and archaeological
perspectives on the initial modern human colonization of southern Asia. Proceedings of the
National Academy of Sciences of the United States of America 110, 10699 – 10704.
Mikhailov K.E., 1991. Classification of fossil eggshells of amniotic vertebrates. Acta Palaeotologica
Polonica 36, 1991.
Milne-Edwards A., 1871. Recherches anatomiques et paléontologiques pour servir à l ’ histoire des
oiseaux fossiles de la France. Librairie de G. Masson, Paris.
Mishra S., 1995. Chronology of the Indian Stone Age: The impact of recent absolute and relative
dating attempts. Man and Environment 20, 11–16.
Mishra S., Naik S., Rajaguru S., 2003. Fluvial response to late Quaternary climatic change: case
studies from upland Western India. Proceedings of the Indian National Science Academy 69,
185 – 200.
Morgan, M. E., Kingston, J. D., Marino, B. D., 1994. Carbon isotopic evidence for the emergence of
C4 plants in the Neogene from Pakistan and Kenya. Nature 367, 162 – 165.
Morgan, M. E., Behrensmeyer, A. K., Badgley, C., Barry, J. C., Nelson, S., Pilbeam, D., 2009. Lateral
trends in carbon isotope ratios reveal a Miocene vegetation gradient in the Siwaliks of Pakistan.
Geology 37, 103-106.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
32
Mourer-Chauviré C., Geraads D., 2008. The Struthionidae and Pelagornithidae Aves :
Struthioniformes, Odontopterygiformes from the late Pliocene of Ahl Al Oughlam , Morocco. 7,
169–194.
Otto-Bliesner B.L., Marshall S.J., Overpeck J.T., Miller G.H., Hu A., 2006. Simulating Arctic
climate warmth and icefield retreat in the last interglaciation. Science 311, 1751–3.
Patnaik R., Chauhan P.R., Rao M.R., Blackwell B. A. B., Skinner A. R., Sahni A., Chauhan M.S.,
Khan H.S., 2009a. New geochronological, paleoclimatological, and archaeological data from the
Narmada Valley hominin locality, central India. Journal of Human Evolution 56, 114–33.
Patnaik R., Sahni A., Cameron D., Pillans B., Chatrath P., Simons E., Williams M., Bibi F., 2009b.
Ostrich-like eggshells from a 10 . 1 million-yr-old Miocene ape locality , Haritalyangar ,
Himachal Pradesh. Current Science 96, 1485–1495.
Petraglia M., Clarkson C., Boivin N., Haslam M., Korisettar R., Chaubey G., Ditchfield P., Fuller D.,
James H., Jones S., Kivisild T., Koshy J., Lahr M.M., Metspalu M., Roberts R., Arnold L., 2009.
Population increase and environmental deterioration correspond with microlithic innovations in
South Asia ca. 35,000 years ago. Proceedings of the National Academy of Sciences of the
United States of America 106, 12261–6.
Potts, D. T., 2001. Ostrich distribution and exploitation in the Arabian peninsula. Antiquity 75: 182-
19
Roberts P., Delson E., Miracle P., Ditchfield P., Roberts R.G., Jacobs Z., Blinkhorn J., Ciochon R.L.,
Fleagle J.G., Frost S.R., Gilbert C.C., Gunnell G.F., Harrison T., Korisettar R., Petraglia M.D.,
2014. Continuity of mammalian fauna over the last 200,000 y in the Indian subcontinent.
Proceedings of the National Academy of Sciences of the United States of America 111, 5848 –
5853.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
33
Quade, J., Cerling, T. E., 1995. Expansion of C4 grasses in the late Miocene of northern Pakistan:
evidence from stable isotopes in Paleosols. Palaeogeography, Palaeoclimatology, Palaeoecology
115, 91-116.
Sahni, A., Kumar G., Bajpai S., Srinivasan S., 1989. Ultrastructure and texonomy of ostrich eggshells
from Upper Palaeolithic sites of India. Jounral of the Palaeontological Society of India 34, 91 –
98.
Sahni A., Kumar G., Bajpai S., Srinivasan S., 1990. A review of Late Pleistocene Ostriches Struthio
sp. in India. Man and Environment 15, 41 – 47.
Sali S., 1985. The Upper Palaeolithic culture at Patne, District Jalgaon, Maharashtra. In: Misra, V,
Bellwood, P. (Eds.), Recent Advances in Indo-Pacific Prehistory. Indo-Pacific Prehistory
Association, New Delhi, pp. 137–145.
Sauer E.G.F., 1972. Ratitie Eggshells and Phylogenetic Questions. Bonner Zoologische Beitrage 23, 3
– 48.
Ségalen L., Renard M., Lee-Thorp J.A., Emmanuel L., Le Callonnec L., de Rafélis M., Senut B.,
Pickford M., Melice J.-L., 2006. Neogene climate change and emergence of C4 grasses in the
Namib, southwestern Africa, as reflected in ratite 13C and 18O. Earth and Planetary Science
Letters 244, 725–734.
Stern L.A., Johnson G.D., Chamberlain C.P., 1994. Carbon isotope signature of environmental change
found in fossil ratite eggshells from a South Asian Neogene sequence Carbon isotope signature
of environmental change found in fossil ratite eggshells from a South Asian Neogene sequence.
Geology 22, 419–422.
Taçon P.S.C., Boivin N., Hampson J., Blinkhorn J., Korisettar R., Petraglia M., 2010. New rock art
discoveries in the Kurnool District, Andhra Pradesh, India. Antiquity 84, 335–350.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
34
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online version of this article:
Appendix S1: Quartz Optically Stimulated Luminescence (OSL dating)

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
35
Ostrich expansion into India during the Late Pleistocene: Implications for continental dispersal
corridors
Highlights
Oldest directly dated evidence for Late Pleistocene expansion of Struthio into India
First stable isotope study of Late Pleistocene ostrich eggshell in India
Synthesis of existing evidence for Struthio populations in South Asia
Struthio expansion associated with extension of Sahel-like habitats




















