
Evolving dispersal and age at death
9
Evolving dispersal and age at death Calvin Dytham and Justin M. J. Travis Dytham, C. and Travis, J. M. J. 2006. Evolving dispersal and age at death. Oikos 113: 530 538. Traditional, and often competing, theories on ageing agree that a programmed age at death must have arisen as a side effect of natural selection, and that it can have no adaptive value of its own. However, theoretical models suggest that ageing and programmed death can be adaptive. Travis J. M. J. suggested that iffecunditydeclines with age, aprogrammed age of death evolves through kin selection and that the nature of dispersal is crucial as it determines the degree of spatial structure and hence the strength of kin selection. Here, using a similar model, we consider the interplay between dispersal and age of death. We incorporate more realistic dispersal kernels and allow both dispersal and age of death to evolve. Our results showeach trait can evolve in response to the other: earlier age of death evolves when individuals disperse less and greater dispersal distances evolvewhen individuals are programmed to die later. When we allow dispersal and age of death to evolve at the same time we typically find that dispersal evolves more rapidly, and that ageing then evolves in response to the new dispersal regime. The cost of dispersal is crucial in determining the evolution of both traits. We argueboth that ageing is an overlooked ecological process, and that the field of gerontology could learn a lot from evolutionary ecology. We suggest that it is time to develop the field of ecological gerontology and we highlight a few areas where future work might be particularly rewarding. C. Dytham, Dept of Biology, Univ. of York, York, UK, YO10 5YW ([email protected]). J. M. J. Travis, Centre for Ecology and Hydrology, Hill of Brathens, Banchory, Aberdeenshire, UK, AB31 4BW. There are three generally accepted theories of ageing: mutation accumulation (Medawar 1952), antagonistic pleiotropy (Williams 1957) and disposable soma (Kirkwood 1977). All three of these theories in some way imply that ageing is the consequence of evolution acting to promote fitness early in life at the expense of fitness later in life. Mitteldorf (2004) summarised these traditional views on ageing writing: ‘‘Standard theo- ries ... regard senescence as an epiphenomenon of selection, with no adaptive value of its own.’’ Recent discoveries in nematodes (Kenyon et al. 1993, Kimura et al. 1997, Friedman and Johnson 1998), insects (Bartke 2001, Tatar et al. 2001) and mammals (Migliaccio et al. 1999, Holzenberger et al. 2003, Lithgow and Gill 2003) of genes that, when mutated, increase life span challenge these conventional theories, and there is a need to reassess the merit of Alfred Russel Wallace’s early ideas regarding adaptive death (Wallace 1889). Travis (2004) demonstrated that in a spatial model a gene that results in deterministic death at a specified age can evolve, and does so through kin selection. The spatial scale of dispersal is a key determinant of the age of death that evolves (Travis 2004), and in this paper we consider in more detail how dispersal influences the age of death to evolve. Specifically we are interested in the evolutionary interplay between dispersal and age of death. We are aware that, despite some recent interest in the ecological consequences of ageing (Bonsall and Mangel 2004, Hendry et al. 2004, Broussard et al. 2005), for many ecologists theories on ageing may be a new topic. First Accepted 28 October 2005 Subject Editor: Tim Benton Copyright # OIKOS 2006 ISSN 0030-1299 OIKOS 113: 530 538, 2006 530 OIKOS 113:3 (2006)
Transcript of Evolving dispersal and age at death

Evolving dispersal and age at death
Calvin Dytham and Justin M. J. Travis
Dytham, C. and Travis, J. M. J. 2006. Evolving dispersal and age at death. � Oikos 113:530�538.
Traditional, and often competing, theories on ageing agree that a programmed age atdeath must have arisen as a side effect of natural selection, and that it can have noadaptive value of its own. However, theoretical models suggest that ageing andprogrammed death can be adaptive. Travis J. M. J. suggested that if fecundity declineswith age, a programmed age of death evolves through kin selection and that the natureof dispersal is crucial as it determines the degree of spatial structure and hence thestrength of kin selection. Here, using a similar model, we consider the interplaybetween dispersal and age of death. We incorporate more realistic dispersal kernels andallow both dispersal and age of death to evolve. Our results show each trait can evolvein response to the other: earlier age of death evolves when individuals disperse less andgreater dispersal distances evolve when individuals are programmed to die later. Whenwe allow dispersal and age of death to evolve at the same time we typically find thatdispersal evolves more rapidly, and that ageing then evolves in response to the newdispersal regime. The cost of dispersal is crucial in determining the evolution of bothtraits. We argue both that ageing is an overlooked ecological process, and that the fieldof gerontology could learn a lot from evolutionary ecology. We suggest that it is time todevelop the field of ecological gerontology and we highlight a few areas where futurework might be particularly rewarding.
C. Dytham, Dept of Biology, Univ. of York, York, UK, YO10 5YW ([email protected]).� J. M. J. Travis, Centre for Ecology and Hydrology, Hill of Brathens, Banchory,Aberdeenshire, UK, AB31 4BW.
There are three generally accepted theories of ageing:
mutation accumulation (Medawar 1952), antagonistic
pleiotropy (Williams 1957) and disposable soma
(Kirkwood 1977). All three of these theories in some
way imply that ageing is the consequence of evolution
acting to promote fitness early in life at the expense of
fitness later in life. Mitteldorf (2004) summarised these
traditional views on ageing writing: ‘‘Standard theo-
ries . . . regard senescence as an epiphenomenon of
selection, with no adaptive value of its own.’’ Recent
discoveries in nematodes (Kenyon et al. 1993, Kimura
et al. 1997, Friedman and Johnson 1998), insects (Bartke
2001, Tatar et al. 2001) and mammals (Migliaccio et al.
1999, Holzenberger et al. 2003, Lithgow and Gill 2003)
of genes that, when mutated, increase life span challenge
these conventional theories, and there is a need to
reassess the merit of Alfred Russel Wallace’s early ideas
regarding adaptive death (Wallace 1889). Travis (2004)
demonstrated that in a spatial model a gene that results
in deterministic death at a specified age can evolve, and
does so through kin selection. The spatial scale of
dispersal is a key determinant of the age of death that
evolves (Travis 2004), and in this paper we consider in
more detail how dispersal influences the age of death to
evolve. Specifically we are interested in the evolutionary
interplay between dispersal and age of death. We are
aware that, despite some recent interest in the ecological
consequences of ageing (Bonsall and Mangel 2004,
Hendry et al. 2004, Broussard et al. 2005), for many
ecologists theories on ageing may be a new topic. First
Accepted 28 October 2005Subject Editor: Tim Benton
Copyright # OIKOS 2006ISSN 0030-1299
OIKOS 113: 530�538, 2006
530 OIKOS 113:3 (2006)

we outline the traditional theories, and then give more
detail on novel perspectives on adaptive death.
Why do organisms age and die?
This question has long vexed biologists. Alfred Russel
Wallace first suggested that ageing and death might be
adaptive (Weismann 1882, Wallace 1889). In the 1860s
Wallace wrote ‘‘Natural selection . . . in many cases
favours such races as die almost immediately after they
have left successors’’. Despite some early support, this
adaptive view of ageing and death was soon dismissed,
to such an extent that in the 1920s it was labelled a
‘‘perverse extension of the theory of natural selection’’
(Pearl 1922). This has remained the case since with
almost all biological gerontologists believing that ‘‘long-
evity determination is under genetic control only indir-
ectly’’, and that ‘‘ . . . ageing is a product of evolutionary
neglect, not evolutionary intent’’ (Olshansky et al. 2002).
Today, there are three largely competing theories used to
explain ageing; mutation accumulation, antagonistic
pleiotropy and disposable soma. The evolution of ageing
has been reviewed by Gavrilov and Gavrilova (2002),
Goldsmith (2003) and Kirkwood (2005). Below, we
summarise the key theories on ageing, and for compar-
ison also describe the main drivers of dispersal.
Traditional theories
The mutation accumulation theory for ageing (Medawar
1952) suggests that ageing occurs due to the accumula-
tion of deleterious mutation that result in negative effects
late in life. Later age classes contribute less to lifetime
fitness and therefore mutations acting late in life
are under weaker selection. Antagonistic pleiotropy
(Williams 1957) is an alternative theory suggesting that
ageing is the result of genes that have beneficial effects
when an organism is young, but also have adverse effects
when the organism is older. This theory relies on the idea
that the beneficial fitness effects early in life will be under
stronger selection than the adverse effects later in life.
For some species, it is likely that genes with compara-
tively small early-life fitness benefits can be selected even
if they cause a major reduction in late-life survival.
A third theory of ageing is disposable soma theory
(Kirkwood 1977) which posits that organisms have to
budget their energy among metabolism, growth, activity
and reproduction. Repairing damaged proteins and
error-checking in DNA replication both make demands
on energy resources, and allocation to these resources
has to be balanced with that to other functions. Failure
to do a perfect job in these and other repair and
maintenance functions leads to an accumulation of
damage, which results in ageing.
Adaptive death
The widely accepted view that ageing has no adaptive
value of its own has been challenged (Guarente and
Kenyon 2000, Skulachev 2001, 2002, Goldsmith 2003,
Mitteldorf 2004, Travis 2004). Two individual-based
models provide theoretical support for the concept of
adaptive ageing and programmed death (Travis 2004,
Mitteldorf 2006), the latter invoking a form of group
selection and the former kin selection.
Mitteldorf (2006) developed a lattice model in which
an array of sites is connected by occasional dispersal.
Each site can support a population whose dynamics are
simulated using the discrete logistic equation. Simulation
experiments were conducted in which individals carry
genes that determine their rate of ageing. Mitteldorf’s
results indicate that parameter space exists where ageing
is selected. He suggests that ageing is selected at the
population level as an adaptation that stabilises popula-
tion dynamics. Within a population, selection favours
individuals leaving the most offspring (i.e. have a lower
rate of ageing). However, this individual-level selection
will lead to an increase in the population’s growth rate
and potentially drive the population into chaotic fluc-
tuations which increases the risk of local extinction. On
average, sites where individuals age more slowly will go
extinct more often, and those vacant sites will, on
average, be colonised by individuals with more rapid
ageing. There is a tension between selection at the level
of the individual and at the level of the population. In
many respects, Mitteldorf’s argument represents classic
group selection but with the twist that selection acts at
the higher level to promote demographic homeostasis.
Travis (2004) developed a rather different spatial
population model to explore the evolutionary ecology
of programmed organism death. As in Mitteldorf (2006)
a lattice is used to represent space, but in Travis (2004)
each site on the lattice supports a single individual
(rather than a population). The model simulates a
viscous population rather than a collection of subpopu-
lations each of which is freely mixing. Results from this
model demonstrate that if an individual’s reproductive
fitness declines with age, then an age of death gene is
selected through kin selection. By dying, an individual
frees resources (in this case, space) that can be used by a
younger, fitter individual. If dispersal is local, such that
local spatial structure develops, the individual that
benefits from another’s death is likely to be a relative.
Therefore if dispersal is local, a critical age exists at
which an individual maximises its inclusive fitness by
dieing. The age of death that evolves is sensitive to
dispersal distance, although even for relatively long
distance dispersal, individuals are selected to have a
finite lifespan (Travis 2004). The interplay between
dispersal and age of death is likely to be relatively
complex, and will be mediated through the spatial
population dynamics. Travis (2004) only touched on
OIKOS 113:3 (2006) 531

this issue and in this paper we aim to extend this first age
of death model to incorporate the evolution of dispersal.
Drivers of dispersal
A number of mechanisms have been suggested to play an
important role in driving the evolution of dispersal
strategies. Here, we outline these mechanisms, and
consider how they may together shape the dispersal
behaviour of individuals within a population.
Increased dispersal is likely to be selected for if local
population extinctions are likely. A population will only
survive in the long-term if it is able to recolonise patches
from where it has become locally extinct. The probability
of local extinction will depend upon a combination of
temporal variability in habitat quality, demographic
stochasticity and the nature of the population dynamics.
Extreme temporal variability in habitat quality may
deterministically drive local populations extinct, and in
this case a population will only survive if it is able to
track the dynamic habitat matrix. There is good
empirical evidence that the temporal persistence of
habitat patches is a determinant of dispersal behaviour
for wing-dimorphic insects (Roff 1986, 1994). Local
populations may go extinct through demographic
stochasticity even when habitat quality remains constant
through time. For populations prone to frequent local
extinctions through demographic stochasticity we would
expect higher rates of dispersal to evolve. Demographic
stochasticity will be most important when effective local
population sizes are small. Population dynamics can
play an important role in determining a population’s
persistence within a patch. For example, populations
that exhibit intrinsically unstable dynamics may become
locally extinct very frequently, driving selection for high
rates of dispersal.
At least two mechanisms can promote dispersal even
when local populations are not at risk of extinction.
Hamilton and May (1977) demonstrated that even in
spatially and temporally stable habitats dispersal is
selected. They demonstrated that dispersal is favoured
because it reduces competition between kin. Dispersal
may also be selected for as a mechanism reducing the
effects of inbreeding depression (Bengtsson 1978, Perrin
and Mazalov 1999, Cockburn et al. 2003). Whatever the
mechanism, it is expected that higher dispersal rates
should evolve when local population equilibrium den-
sities are low, for example when habitat patch sizes are
small, because kin competition is more intense and
inbreeding more severe in small populations.
Local population extinction, kin competition and
inbreeding all act to select for increased dispersal. The
evolution of increased dispersal is always constrained by
one or more costs, for example mortality risks during
travelling, investments in dispersal morphology, or a
tradeoff between dispersal and competitive abilities.
Linking age of death and dispersal in a simple patch
occupancy model
Here, we construct a simple patch occupancy model, in
which fertility declines with age and an individual’s
genotype determines both its dispersal propensity and its
programmed age of death. We look first at how different
dispersal strategies influence the age of death that
evolves, second at how different age of deaths influence
the evolution of dispersal, and third at how the two
evolve.
Method
We develop an individual-based, simulation model to
investigate the evolution of programmed age at death
and dispersal distance. Simulations take place in an
arena of cells with individuals located in continuous
space. For simplicity, each cell can only support one
individual. The model is event-based with events taking
place in series. An individual and a type of event are
selected at random and time is incremented by an
infinitesimal amount in a manner similar to that
described in Renshaw (1991). Events are either deaths
or birth and dispersal. This approach overcomes poten-
tial problems with the ordering of events (discussed in
Palmer 1992). We consider a species that has only one
dispersal event in its life and this occurs immediately
after birth. The model is agent-based with each
individual carrying an exact location in space (x, y co-
ordinates), its age, a dispersal distance assigned to any
offspring and an age at death. Individuals can suffer
mortality form a) a background mortality rate indepen-
dent of age or density which is usually set at 0.01 (i.e. 1%
probability of death in a ‘year’); or b) through senescence
when an individual exceeds its assigned age at death.
Following Travis (2004), births occur with a probability
that declines with age and dispersal immediately follows
birth. Organisms are iteroparous and in each birth event
there is only one offspring ‘seed’. The seed moves in a
random direction from the adult selected from a uniform
random variate between 0 and 2p radians. The distance
is drawn from a negative exponential distribution with a
mean taken from the genotype of the individual. So, an
individual that has a birth event may have a dispersal
distance of 4.2 which means that any offspring will move
in a random direction a random distance drawn from a
negative exponential distribution with a mean of 4.2.
Any individual that fails to leave the parental cell is
moved exactly one unit in a cardinal direction from its
532 OIKOS 113:3 (2006)

parent’s location (Fig. 1). A disperser arriving at an
occupied cell is not able to establish.
As this is an event-based model there will be some
individuals who have several events occur to them in a
‘year’ while others have no events. Events occur at
random, so they will be Poisson distributed amongst
individuals.
The arena is a torus so any individual leaving the
arena will reappear on the opposite side. We also use a
mortality associated with dispersal. Following Murrell
et al. (2002) this is applied as a linear, increasing
mortality with distance during dispersal. A value of 20
equates to a mortality of 100% when moving a distance
of 20 units, or 50% for 10 units, and so on. In the
simulations presented here we have dispersal mortality
values of 5, 10 or � (no mortality during dispersal).
There may be mutation associated with a birth event.
Mutations are rare (usually 0.3%). Mutations can affect
either age at death or dispersal distance. Mutations to
age at death are drawn from a uniform distribution from
�/3 to �/3. Mutation to dispersal distance is also
uniform from �/0.5 to �/0.5. Both characters have a
limit at zero and any mutation that gives a value less than
zero is set to 0.01. An individual may only have a
mutation to one character. While there is no explicit
calculation of the relatedness of individuals limited
dispersal leads to population viscosity and the clustering
of individuals sharing a genotype (i.e. kin).
Simulations were carried out on square grids of 200
cell side lengths (40 000 cells) with approximately 1/3 of
cells occupied at the start. Individuals are assigned either
a) random age at death (between 1 and 100), dispersal
distance (between 0.01 and 10) and current age (between
0 and 100) at initialisation or b) set values for one or
both characters.
Simulations are run for up to 150 000 years and where
confidence intervals are shown there were ten realisa-
tions of the model for each parameter value.
Results
When dispersal distance is fixed and age at death
allowed to evolve there is a clear positive relationship
between evolved age at death and dispersal distance
(Fig. 2a). Although the mean evolved values for age at
death for the population are very consistent between
realisations there is a range of evolved values remaining
within a single realisation even after a long simulation
(Fig. 2b). Clusters of relatives (i.e. individuals sharing a
common strategy) are clearly seen as patches sharing a
common age at death.
When age at death is fixed and dispersal distance
allowed to evolve there is an increase in dispersal
distance with age at death (Fig. 3) although the relation-
ship is not linear.
Fixing both characters for all individuals in the
population allows the effect of age at death and dispersal
distance on total population size to be seen. Within a
single dispersal distance, as shown in Fig. 4, there is a
much lower population size with early age at death, this
rises rapidly to a peak and then declines slowly as age at
death increases further.
When both age at death and dispersal distance are
allowed to evolve with no cost to dispersal the dispersal
distance rises quickly and the age at death slowly with
no apparent limit. Simulations run for a very long time
(�/250 000 years) still show age at death drifting slowly
upwards. Mortality during dispersal keeps dispersal
distances down and higher costs to dispersal reduce
both age at death and dispersal distance.
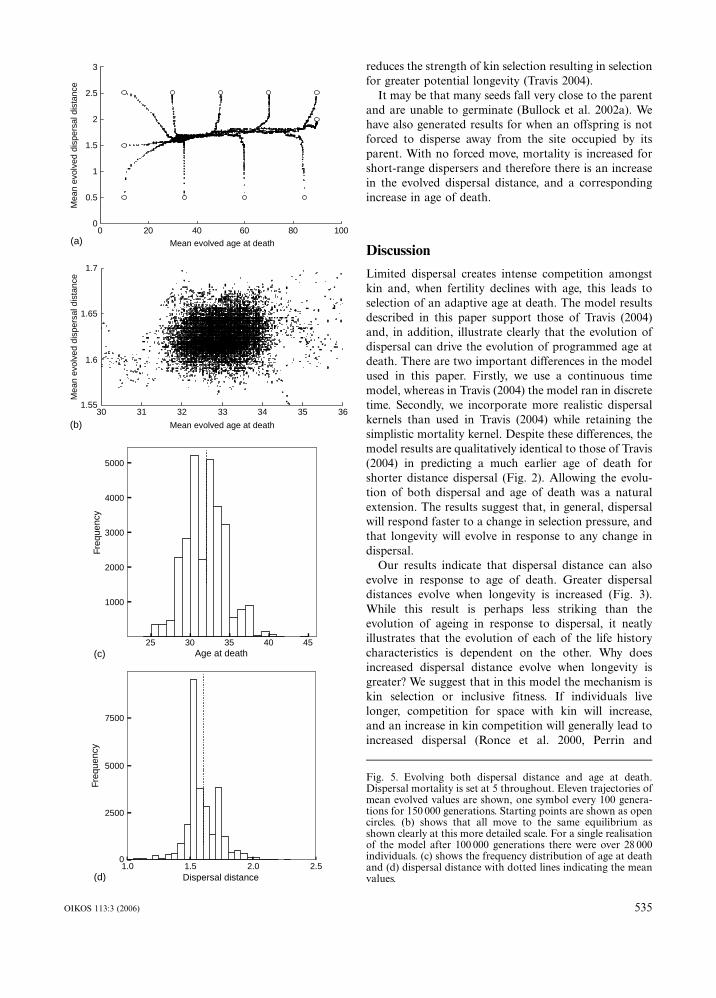
If all individuals start with identical strategies and
both characters are allowed to evolve the population
means reach the same values regardless of starting
values. This is shown for 11 trajectories from different
starting values in Fig. 5a and, in more detail around the
equilibrium value in 5b. Using this range of starting
conditions we only found a single attractor. It is clear
from the trajectories that dispersal distance is under
stronger selection pressure than age at death except when
age at death is very low. We determined this as a
proportion of the final change in a character that
was achieved in the first 1000 years. This is because
old individuals are under very limited selection pressure.
x
(a) (b)
(c)
Fig. 1. This shows a stylised version of the event-based model.In (a) a dispersal event (arrow) fails because the destination cellis occupied. In (b) an individual dies. In (c) a short distancedispersal event (dotted arrow) fails to leave the cell, so adispersal distance of 1 (arrow) is selected for a successfuldispersal event.
OIKOS 113:3 (2006) 533

Figure 5c and 5d show that at equilibrium the
mutation�selection balance is allowing quite a large
variation between individuals.
Sensitivity analysis
Results are not affected by mutation rate or mutation
ranges except in the speed at which an equilibrium level
is reached. Changing boundary conditions to absorbing
or reflecting boundaries also has little effect, except in
arenas much smaller than those reported here and when
mortality during dispersal is very low and more indivi-
duals are lost from the edge of the arena.
Adjusting the background rate of mortality and
changing the shape of the decline in reproductive fitness
with age both have the same qualitative impacts as
reported by Travis (2004). When reproductive senescence
is more rapid, selection favours an earlier age of death.
As background mortality is increased, selection acts to
increase the programmed age of death. This can be
explained by considering the population dynamic con-
sequences of higher background mortality. As extrinsic
mortality is increased population abundance declines
and this reduces the competition of space. This in turn
60
50
Mea
n ev
olve
d ag
e at
dea
th
40
30
20.5 1.0 1.5 2.0 2.5 3.0 5.0 6.0 8.0
Distance7.04.0
(b)
(a)
Fig. 2. The evolved age at death after 10 000 generations. (a)shows mean and 95% confidence interval of 10 realisations ofthe model for a range of fixed values for dispersal distance.Dispersal mortality is 10 throughout. The dotted vertical lineindicates a change in the x-scale. (b) a snapshot from the end ofa single realisation where dispersal distance is 1 and the meanevolved age at death is 31, showing the level of variation andclustering of evolved strategies.
Fig. 3. The evolved dispersal distance after 10 000 generationsfor a range of fixed values age at death. Dispersal mortality is 10throughout. Mean and 95% confidence interval of 10 realisationsof the model are shown.
3.0
Mea
n ev
olve
d di
stan
ce
2.8
2.6
2.4
2.2
2.010 20 30 40 50 60
Age at death70 80 90 100
10 20 30 40 50 60 70 80 90 100
Age at death
18000
19000
20000
21000
22000
23000
Pop
ulat
ion
size
Fig. 4. The mean and 95% confidence interval of populationsize is shown for a range of fixed age at death. Dispersalmortality is 5, dispersal fixed at 1.5, age at death fixed for eachrealisation but varied from 10 to 100 in steps of two. The dottedvertical line indicates the age at death that evolves whendispersal is fixed at 1.5 and cost is 5.
534 OIKOS 113:3 (2006)
reduces the strength of kin selection resulting in selection
for greater potential longevity (Travis 2004).
It may be that many seeds fall very close to the parent
and are unable to germinate (Bullock et al. 2002a). We
have also generated results for when an offspring is not
forced to disperse away from the site occupied by its
parent. With no forced move, mortality is increased for
short-range dispersers and therefore there is an increase
in the evolved dispersal distance, and a corresponding
increase in age of death.
Discussion
Limited dispersal creates intense competition amongst
kin and, when fertility declines with age, this leads to
selection of an adaptive age at death. The model results
described in this paper support those of Travis (2004)
and, in addition, illustrate clearly that the evolution of
dispersal can drive the evolution of programmed age at
death. There are two important differences in the model
used in this paper. Firstly, we use a continuous time
model, whereas in Travis (2004) the model ran in discrete
time. Secondly, we incorporate more realistic dispersal
kernels than used in Travis (2004) while retaining the
simplistic mortality kernel. Despite these differences, the
model results are qualitatively identical to those of Travis
(2004) in predicting a much earlier age of death for
shorter distance dispersal (Fig. 2). Allowing the evolu-
tion of both dispersal and age of death was a natural
extension. The results suggest that, in general, dispersal
will respond faster to a change in selection pressure, and
that longevity will evolve in response to any change in
dispersal.
Our results indicate that dispersal distance can also
evolve in response to age of death. Greater dispersal
distances evolve when longevity is increased (Fig. 3).
While this result is perhaps less striking than the
evolution of ageing in response to dispersal, it neatly
illustrates that the evolution of each of the life history
characteristics is dependent on the other. Why does
increased dispersal distance evolve when longevity is
greater? We suggest that in this model the mechanism is
kin selection or inclusive fitness. If individuals live
longer, competition for space with kin will increase,
and an increase in kin competition will generally lead to
increased dispersal (Ronce et al. 2000, Perrin and
0
0.5
1
1.5
2
2.5
3
0 20 40 60 80 100
Mean evolved age at death
Mea
n ev
olve
d di
sper
sal d
ista
nce
1.55
1.6
1.65
1.7
30 31 32 33 34 35 36
Mean evolved age at death
Mea
n ev
olve
d di
sper
sal d
ista
nce
(a)
(b)
25 30(c)
(d)
35 40 45Age at death
1000
2000
3000
4000
5000
Freq
uenc
y
1.0 1.5 2.0 2.5Dispersal distance
0
2500
5000
7500
Freq
uenc
y
Fig. 5. Evolving both dispersal distance and age at death.Dispersal mortality is set at 5 throughout. Eleven trajectories ofmean evolved values are shown, one symbol every 100 genera-tions for 150 000 generations. Starting points are shown as opencircles. (b) shows that all move to the same equilibrium asshown clearly at this more detailed scale. For a single realisationof the model after 100 000 generations there were over 28 000individuals. (c) shows the frequency distribution of age at deathand (d) dispersal distance with dotted lines indicating the meanvalues.
OIKOS 113:3 (2006) 535

Goudet 2001). Our model has a dispersal rate indepen-
dent of age, although Ronce et al. (1998) showed that
fewer offspring should disperse from an older individual
than a younger one.
Any change in the cost of dispersal, is likely to
ultimately impact both on the dispersal characteristics
and the programmed age of death found in a population.
From Fig. 5 it is clear that the dispersal distance evolves
more rapidly than the age at death. Dispersal evolves to
an equilibrium level and then age at death responds. We
predict that, in nature, any change in cost to dispersal,
for example through landscape alteration, will first affect
dispersal rates and distances first and impact on age at
death much more slowly. As we have previously demon-
strated (Travis and Dytham 1998, 1999) unintelligent
dispersers, such as a plants with wind-dispersed seed, are
likely to evolve shorter dispersal distances in response to
habitat fragmentation, and results from this paper lead
us to predict that habitat loss will eventually lead to
younger age at death for individuals in these popula-
tions. We could also speculate on the effect of habitat
persistence and habitat quality on age at death from
what we know about its effect on dispersal distance
(Travis et al. 1999, Travis 2001, Murrell et al. 2002,
Poethke and Hovestadt 2002, Parvinen 2004). However,
changes in habitat persistence can substantially alter the
nature of the spatial population dynamics and this in
turn can modify the strength of kin selection. We suspect
that the interplay between dispersal, age at death and the
spatial population dynamics may in this instance be
particularly complex, and highlight it as an area where
further theory would be beneficial.
In this paper we have considered the evolution of
dispersal and programmed age of death. Theoretical and
empirical work has together contributed to a major
advance in our understanding of both the causes and
consequences of different dispersal behaviours (Clobert
et al. 2001, Bullock et al. 2002b, Bowler and Benton
2005). Most often, work investigating the evolution of
dispersal, considers the process in the absence of
evolution acting on other life history characteristics.
An important feature of this paper is that it investigates
the evolution of dispersal with another character. In
reality, dispersal is likely to evolve with a wide range of
traits such as competitive ability, mating strategy, age of
maturity and resistance to or virulence of pathogens. For
example dispersal may evolve with kin-recognition
mechanisms (Perrin and Lehmann 2001, Lehmann and
Perrin 2003). Theoretical and empirical work dedicated
to investigating the evolution of different life history
characteristics would be valuable and Mittledorf (2004)
reviews some of the existing studies.
Whereas, in the dispersal literature there is a general
recognition that the evolved strategy depends upon a
suite of drivers acting together, in ageing research the
various theories are considered alternates, and in general
it is not considered that the different processes may act
together to shape the life history of an organism. Our
view is that ageing is likely to be determined by a
combination of these processes, and that the relative
importance of each driver will vary depending upon the
organism concerned and the environment it inhabits. For
example, although adaptive age at death may be
important when only a few individuals reach old age, it
is likely to become more important when survival is
higher. It is also worth considering that if selection
favours a finite lifespan, pleiotropic genes may confer a
selective advantage both because they confer increased
fitness early in life and because they promote ageing
later. Rather than the late-life ‘detrimental effect’ being
viewed as an unfortunate indirect effect it may actually
confer an evolutionary benefit of its own. It is possible
that a gene that has the ‘negative’ late life effect may
be selected over a gene with the same early-life
consequences without the pleiotropic effect. Future
work should seek to integrate traditional and adaptive
theories on ageing.
Another outcome of our model that appears robust is
that the evolved age at death is not necessarily the one
that generates the highest population density (Fig. 4).
Evolution operates at a level below that of the popula-
tion (in this model it acts at both individual and kin
group levels) so there is no reason to expect that the
evolved and optimal (in terms of maximising population
size) strategies should coincide. It is worth comparing
the result shown here with those presented by Parvinen
(2004; see also Olivieri and Gouyon 1997). Parvinen
(2004) clearly illustrates that an evolved dispersal rate
can be either higher or lower than the one that optimises
population size, and that the direction of deviation
depends upon the spatial scale of dispersal.
This paper adds to the recent ideas relating to
adaptive ageing and programmed death (Skulachev
2001, 2002, Mitteldorf 2004, Travis 2004). To date, this
work has consisted of verbal arguments illustrated by
judicious use of empirical evidence (Mitteldorf 2004)
and individual-based simulation models (Travis 2004,
Mitteldorf 2005). Theoretically, programmed death can
be adaptive. The key question now, is to what extent is
the ageing of organisms determined by selection acting
directly on it, and to what extent is it a side effect of
selection acting to maximise fitness early in life. Teasing
apart these two is likely to be challenging, and will
demand a mix of theoretical and experimental work.
Studies on mammals (Broussard et al. 2003, 2005) have
made some progress in determining the relative invest-
ment in reproduction or soma (i.e. longevity) through
life, but sample sizes are inevitably small. We believe that
carefully planned microcosm experiments offer consider-
able potential: organisms such as Caenorhabditis
elegans (Gardner et al. 2004), Drosophila melanogaster
(Kennington et al. 2003) and mites (Benton et al. 2004)
536 OIKOS 113:3 (2006)

are all amenable to microcosm work. They are readily
cultured, and have relatively short generation times
allowing evolutionary processes to be studied in the
confines of the laboratory. In addition it is easy to
acquire genotyes of C. elegans that have different
dispersal and ageing behaviours. Experiments could be
designed to test specific predictions of the models. For
example, to test the idea that population viscosity
determines the evolution of ageing, a simple experiment
could be established involving competition between
different ageing genotypes. The spatial structure of a
set of populations could be manipulated by manually
moving individuals and thus artificially increasing dis-
persal, while in the control the same proportion of
individuals could be picked up and replaced in the same
position. An added complication to the interpretation of
results from such a study is that fertility profile will also
evolve under such selection (Rose 1984).
Ageing research, or gerontology, is booming. While
this has no doubt been fuelled by a desire to better
understand and deal with the consequences of an ageing
human population, considerable progress has been made
that can be applied to a broad range of questions,
including ecological ones. Additionally, ecological in-
sights may help elucidate the importance of various
potential drivers of ageing under different environmental
conditions. We can predict, for example, that habitat
fragmentation will have an impact on evolved age at
death through its impact on dispersal because highly
fragmented populations are likely to be inbred.
Although traditionally quite separate fields there
may be much that gerontologists can learn from
ecologists and vice versa. It is our hope that this paper
will help to stimulate interest in the field of ecological
gerontology.
References
Bartke, A. 2001. Mutations prolong life in flies; implications foraging in mammals. � Trends Endocrinol. Metab. 12: 233�234.
Bengtsson, B. O. 1978. Avoiding inbreeding: at what cost? � J.Theor. Biol. 73: 439�444.
Benton, T. G., Cameron, T. C. and Grant, A. 2004. Populationresponses to perturmbations: predictions and responsesfrom laboratory mite populations. � J. Anim. Ecol. 73:983�995.
Bonsall, M. B. and Mangel, M. 2004. Life-history tradeoffs andecological dynamics in the evolution of longevity. � Proc. R.Soc. Lond. 271: 1143�1150.
Bowler, D. E. and Benton, T. G. 2005. Causes and consequencesof animal dispersal strategies: relating individual behaviourto spatial dynamics. � Biol. Rev. 80: 205�225.
Broussard, D. R., Risch, T. S., Dobson, F. S. et al. 2003.Senescence and age-related reproduction of female Colum-bian ground squirrels. � J. Anim. Ecol. 72: 212�219.
Broussard, D. R., Michener, G. S., Risch, T. S. et al. 2005.Somatic senescence: evidence from female Richardson’sground squirrels. � Oikos 108: 591�601.
Bullock, J., Kenward, R. and Hails, R. 2002a. Dispersalecology. � Blackwell Science.
Bullock, J. M., Moy, I. L., Pywell, R. F. et al. 2002b. Plantdispersal and colonisation processes at local and landscapescales. � In: Bullock, J. M., Kenward, R. E. and Hails, R. S.(eds), Dispersal. Blackwell Science.
Clobert, J., Danchin, E., Dhont, A. A. et al. 2001. Dispersal.� Oxford Univ. Press.
Cockburn, A., Osmond, H. L., Mulder, R. A. et al. 2003.Divorce, dispersal and incest avoidance in the cooperativelybreeding superb fairy-wren Malurus cyaneus. � J. Anim.Ecol. 72: 189�202.
Friedman, D. B. and Johnson, T. E. 1988. Three mutants thatextend both the mean and maximum life span of thenematode, Caenorhabditis elegans, define the age-1 gene.� J. Gerentol. 43: 102�109.
Gardner, M., Gems, D. and Viney, M. 2004. Aging in a veryshort-lived nematode. � Exp. Gerontol. 39: 1267�1276.
Gavrilov, L. A. and Gavrilova, N. S. 2002. Evolutionarytheories of aging and longevity. � Sci. World J. 2: 339�356.
Goldsmith, T. C. 2003. The evolution of aging: how Darwin’sdilemma is affecting your chance for a longer and healthierlife. � Universe Inc, Lincoln, USA.
Guarente, L. and Kenyon, C. 2000. Genetic pathways thatregulate ageing in model organisms. � Nature 408: 255�262.
Hamilton, W. D. and May, R. M. 1977. Dispersal in stablehabitats. � Nature 269: 578�581.
Hendry, A. P., Morbey, Y. E., Berg, O. K. et al. 2004. Adaptivevariation in senescence: reproductive lifespan in a wildsalmon population. � Proc. R. Soc. 271: 259�266.
Holzenberger, M., Dupont, J., Ducos, B. et al. 2003. IGF-1receptor regulates lifespan and resistance to oxidative stressin mice. � Nature 421: 182�187.
Kennington, W. J., Killeen, J. R., Goldstein, D. B. et al. 2003.Rapid laboratory evolution of adult body size in Drosophilamelanogaster in response to humidity and temperature.� Evolution 57: 932�936.
Kenyon, C., Chang, J., Gensch, E. et al. 1993. A C. elegansmutant that lives twice as long as wild type. � Nature 366:461�464.
Kimura, K. D., Tissenbaum, H. A., Liu, Y. et al. 1997. Daf-2,an insulin receptor-like gene that regulates longevity anddiapause in Caenorhabditis elegans. � Science 277: 942�946.
Kirkwood, T. B. L. 1977. Evolution of ageing. � Nature 270:301�304.
Kirkwood, T. B. L. 2005. Understanding the odd science ofaging. � Cell 120: 437�447.
Lehmann, L. and Perrin, N. 2003. Inbreeding avoidancethrough kin recognition. Choosy females boost maledispersal. � Am. Nat. 162: 638�652.
Lithgow, G. J. and Gill, M. S. 2003. Cost-free longevity in mice?� Nature 421: 125�126.
Medawar, P. B. 1952. An unsolved problem of biology. � H. K.Lewis.
Migliaccio, E., Giorgio, M., Mele, S. et al. 1999. The p66shc
adaptor protein controls oxidative stress response andlifespan in mammals. � Nature 402: 309�313.
Mitteldorf, J. 2004. Ageing selected for its own sake. � Evol.Ecol. Res. 6: 937�953.
Mitteldorf, J. 2006. Demographic homeostasis and the evolu-tion of senescence. � Evol. Ecol. Res. (in press).
Murrell, D. J., Travis, J. M. J. and Dytham, C. 2002. Theevolution of dispersal distance in spatially-structured popu-lations. � Oikos 97: 229�236.
Olivieri, I. and Gouyon, P. H. 1997. Evolution of migration rateand other traits: the metapopulation effect. � In: Hanski, I.and Gilpin, M.E. (eds), Metapopulation biology: ecology,genetics and evolution. Academic Press, pp. 293�323.
Olshansky, S. J., Halflick, L. and Carnes, B. A. 2002. The truthabout human aging [position statement on human aging].� J. Gerontol. 57A: B292�B297.
OIKOS 113:3 (2006) 537

Pearl, R. 1922. The biology of death. � J. B. Lippincott,Philadelphia.
Palmer, J. B. 1992. Hierarchical and concurrent individual basedmodelling. � In: DeAngelis, D. L. and Gross, L. J. (eds),Individual-based models and approaches in ecology:populations, communities and ecosystems. Chapman andHall, pp. 188�207.
Parvinen, K. 2004. Adaptive responses to landscape distur-bances: theory. � In: Ferriere, R., Dieckmann, U. andCouvet, D. (eds), Evolutionary conservation biology.� Cambridge Univ. Press, pp. 265�286.
Perrin, N. and Mazalov, V. 1999. Dispersal and inbreedingavoidance. � Am. Nat. 154: 282�292.
Perrin, N. and Goudet, J. 2001. Inbreeding, kinship and theevolution of natal dispersal. � In: Clobert, J. et al. (eds),Dispersal. Oxford Univ. Press, pp. 155�167.
Perrin, N. and Lehmann, L. 2001. Is sociality driven by thecosts of dispersal or the benefits of philopatry? A role forkin-discrimination mechanisms. � Am. Nat. 158: 471�583.
Poethke, H. J. and Hovestadt, T. 2002. Evolution of density andpatch-size-dependent dispersal rates. � Proc. R. Soc. Lond.269: 637�645.
Renshaw, E. 1991. Modelling biological populations in spaceand time. � Cambridge Univ. Press.
Roff, D. A. 1986. Evolution of wing polymorphism and itsimpact on life cycle adaptation in insects. � In: Taylor, F.and Karban, K. (eds), The evolution of insect life cycles.Springer-Verlag, pp. 204�221.
Roff, D. A. 1994. The evolution of flightlessness: is historyimportant? � Evol. Ecol. 8: 629�657.
Ronce, O., Clobert, J. and Massot, M. 1998. Natal dispersal andsenescence. � Proc. Nat. Acad. Sci. 95: 600�605.
Ronce, O., Gandon, S. and Rousset, F. 2000. Kin selection andnatal dispersal in an age-structured population. � Theor.Popul. Biol. 58: 143�159.
Rose, M. R. 1984. Laboratory evolution of postponed senes-cence in Drosophila melanogaster. � Evolution 38: 1004�1010.
Skulachev, V. P. 2001. The programmed death phenomena,aging, and the Samurai law of biology. � Exp. Gerontol. 36:995�1024.
Skulachev, V. P. 2002. Programmed death phenomena: fromorganelle to organism. � Ann. N. Y. Acad. Sci. 959: 214�237.
Tatar, M., Kopelman, A., Epstein, D. et al. 2001. A mutantDrosophila insulin receptor homolog that extends life-spanand impairs neuroendocrine function. � Science 292: 107�110.
Travis, J. M. J. 2001. The color of noise and the evolution ofdispersal. � Ecol. Res. 16: 157�163.
Travis, J. M. J. 2004. The evolution of programmed death in aspatially structured population. � J. Gerontol. 59A: 301�305.
Travis, J. M. J. and Dytham, C. 1998. The evolution of dispersalin a metapopulation: a spatially explicit, individual-basedmodel. � Proc. R. Soc. Lond. 265: 17�23.
Travis, J. M. J. and Dytham, C. 1999. Habitat persistence,habitat availability and the evolution of dispersal. � Proc. R.Soc. Lond. 266: 723�728.
Travis, J. M. J., Murrell, D. J. and Dytham, C. 1999. Theevolution of density-dependent dispersal. � Proc. R. Soc.Lond. 266: 1837�1842.
Wallace, A. R. 1889. The action of natural selection inproducing old age, decay and death. A note by Wallacewritten ‘some time between 1865 and 1870’ and included inWeismann, A., Essays upon heredity and kindred biologicalproblems. � Clarendon Press.
Weismann, A. 1882. Uber die Dauer des Lebens. � Verlag vonGustav Fisher, Jena.
Williams, G. C. 1957. Pleiotropy, natural selection and theevolution of senescence. � Evolution 11: 398�411.
538 OIKOS 113:3 (2006)




















