
Optimization of the HepaRG cell model for drug metabolism and toxicity studies
8
Optimization of the HepaRG cell model for drug metabolism and toxicity studies Sébastien Anthérieu a,b , Christophe Chesné c , Ruoya Li c , Christiane Guguen-Guillouzo a,b , André Guillouzo a,b,⇑ a Inserm UMR 991, Rennes, France b Université de Rennes 1, Rennes, France c Biopredic International, Saint-Grégoire, France article info Article history: Available online 27 May 2012 Keywords: HepaRG cells Hepatocytes CYP P450 Inducers Hepatotoxicity Drug metabolism abstract The HepaRG cell line is the first human cell line able to differentiate in vitro into mature hepatocyte-like cells. Our main objective within the framework of the EEC-LIINTOP project was to optimize the use of this cell line for drug metabolism and toxicity studies, especially after repeat treatments. The main results showed that differentiated HepaRG cells: (i) retained their drug metabolism capacity (major CYPs, phase 2 enzymes, transporters and nuclear receptors) and responsiveness to prototypical inducers at relatively stable levels for several weeks at confluence. The levels of several functions, including some CYPs such as CYP3A4, were dependent on the addition of dimethyl sulfoxide in the culture medium; (ii) sustained the different types of chemical-induced hepatotoxicity, including steatosis, phospholipidosis and cholestasis, after acute and/or repeat treatment with reference drugs. In particular, drug-induced vesicular steatosis was demonstrated in vitro for the first time. In conclusion, our results from the LIINTOP project, together with other studies reported concomitantly or more recently in the literature, support the conclusion that the metabolically competent human HepaRG cells represent a surrogate to primary human hepatocytes for investigating drug metabolism parameters and both acute and chronic effects of xenobiotics in human liver. Ó 2012 Elsevier Ltd. All rights reserved. 1. Introduction Most xenobiotics are lipophilic and cannot be directly excreted by the organism. The liver is the principal organ involved in the biotransformation of exogenous substances; it possesses rich enzy- matic equipment that can effect conversion of hydrophobic sub- stances into water-soluble products that can then be readily excreted from the body. However, a number of xenobiotics are potentially hepatotoxic either directly or more frequently after bioactivation, leading to the formation of chemically reactive metabolites and/or generation of reactive oxygen species. The capacity of a chemical to be excreted or to produce liver damage often results from the intersection of a series of complex cellular processes that are involved in its absorption, distribution, metabo- lism, excretion and/or toxicity (ADMET). Drug-induced liver injury represents a major clinical problem and is a major cause for drugs being withdrawn from the market and of attrition in drug develop- ment. It is an infrequent event that mostly occurs after several weeks or months of treatment and is, with few exceptions, unpre- dictable (idiosyncratic) (Lee, 2003; Uetrecht, 2008). Incidence of drug-induced hepatotoxicity caused by some drugs can be high as 1 in 100 patients (e.g., chlorpromazine), but in most cases is esti- mated at 1 in 10,000 to 1 in 100,000 treated patients. More than 1000 drugs and herbal products have been associated with idio- syncratic hepatotoxicity (Biour et al., 2000). Such unpredictable hepatotoxicity is challenging to investigate because of its rarity and the lack of experimental models. The low prediction of drug- induced liver toxicities in human patients from studies in animals is explained in particular by frequent and large variations in the rates and routes of drug metabolism between humans and labora- tory animals (Olson et al., 2000). Estimation of ADMET properties is an important consideration for the development of novel therapeu- tic agents. Thus, predicting the ability of a drug to modulate cytochrome P450 (CYP) expression at an early stage of its discovery and development should reduce the risk of failure in the clinic and, more importantly, permit the identification of alternative non- inducing/non-inhibiting chemical structures. 0887-2333/$ - see front matter Ó 2012 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.tiv.2012.05.008 Abbreviations: ADMET, absorption, distribution, metabolism, excretion and/or toxicity; AFB1, aflatoxin B1; CL int , intrinsic clearance; CYP, cytochrome P450; DMSO, dimethyl sulfoxide; LD, low density; HD, high density; MeiQx, 2-amino-3,8- dimethylimidazo[4,5-f]quinoxaline; PhiP, 2-amino-1-methyl-6-phenylimidazo[4,5- b]pyridine; PPAR, peroxisome proliferator-activated receptor; PB, phenobarbital; OME, omeprazole; RIF, rifampicin; RT-qPCR, real time-quantitative polymerase chain reaction; TG, triglycerides. ⇑ Corresponding author. Address: Inserm UMR 991, Faculté des Sciences Pharmaceutiques et Biologiques, 35043 Rennes Cedex, France. Tel.: +33 223234791; fax: +33 223235385. E-mail addresses: [email protected] (S. Anthérieu), Andre.Guillouzo@ univ-rennes1.fr (A. Guillouzo). Toxicology in Vitro 26 (2012) 1278–1285 Contents lists available at SciVerse ScienceDirect Toxicology in Vitro journal homepage: www.elsevier.com/locate/toxinvit
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Optimization of the HepaRG cell model for drug metabolism and toxicity studies

Toxicology in Vitro 26 (2012) 1278–1285
Contents lists available at SciVerse ScienceDirect
Toxicology in Vitro
journal homepage: www.elsevier .com/locate / toxinvi t
Optimization of the HepaRG cell model for drug metabolism and toxicity studies
Sébastien Anthérieu a,b, Christophe Chesné c, Ruoya Li c, Christiane Guguen-Guillouzo a,b,André Guillouzo a,b,⇑a Inserm UMR 991, Rennes, Franceb Université de Rennes 1, Rennes, Francec Biopredic International, Saint-Grégoire, France
a r t i c l e i n f o a b s t r a c t
Article history:Available online 27 May 2012
Keywords:HepaRG cellsHepatocytesCYP P450InducersHepatotoxicityDrug metabolism
0887-2333/$ - see front matter � 2012 Elsevier Ltd. Ahttp://dx.doi.org/10.1016/j.tiv.2012.05.008
Abbreviations: ADMET, absorption, distribution, mtoxicity; AFB1, aflatoxin B1; CLint, intrinsic clearanDMSO, dimethyl sulfoxide; LD, low density; HD, highdimethylimidazo[4,5-f]quinoxaline; PhiP, 2-amino-1-mb]pyridine; PPAR, peroxisome proliferator-activatedOME, omeprazole; RIF, rifampicin; RT-qPCR, real tchain reaction; TG, triglycerides.⇑ Corresponding author. Address: Inserm UMR
Pharmaceutiques et Biologiques, 35043 Rennes223234791; fax: +33 223235385.
E-mail addresses: [email protected] (S. Auniv-rennes1.fr (A. Guillouzo).
The HepaRG cell line is the first human cell line able to differentiate in vitro into mature hepatocyte-likecells. Our main objective within the framework of the EEC-LIINTOP project was to optimize the use of thiscell line for drug metabolism and toxicity studies, especially after repeat treatments. The main resultsshowed that differentiated HepaRG cells: (i) retained their drug metabolism capacity (major CYPs, phase2 enzymes, transporters and nuclear receptors) and responsiveness to prototypical inducers at relativelystable levels for several weeks at confluence. The levels of several functions, including some CYPs such asCYP3A4, were dependent on the addition of dimethyl sulfoxide in the culture medium; (ii) sustained thedifferent types of chemical-induced hepatotoxicity, including steatosis, phospholipidosis and cholestasis,after acute and/or repeat treatment with reference drugs. In particular, drug-induced vesicular steatosiswas demonstrated in vitro for the first time. In conclusion, our results from the LIINTOP project, togetherwith other studies reported concomitantly or more recently in the literature, support the conclusion thatthe metabolically competent human HepaRG cells represent a surrogate to primary human hepatocytesfor investigating drug metabolism parameters and both acute and chronic effects of xenobiotics in humanliver.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction processes that are involved in its absorption, distribution, metabo-
Most xenobiotics are lipophilic and cannot be directly excretedby the organism. The liver is the principal organ involved in thebiotransformation of exogenous substances; it possesses rich enzy-matic equipment that can effect conversion of hydrophobic sub-stances into water-soluble products that can then be readilyexcreted from the body. However, a number of xenobiotics arepotentially hepatotoxic either directly or more frequently afterbioactivation, leading to the formation of chemically reactivemetabolites and/or generation of reactive oxygen species. Thecapacity of a chemical to be excreted or to produce liver damageoften results from the intersection of a series of complex cellular
ll rights reserved.
etabolism, excretion and/orce; CYP, cytochrome P450;
density; MeiQx, 2-amino-3,8-ethyl-6-phenylimidazo[4,5-
receptor; PB, phenobarbital;ime-quantitative polymerase
991, Faculté des SciencesCedex, France. Tel.: +33
nthérieu), Andre.Guillouzo@
lism, excretion and/or toxicity (ADMET). Drug-induced liver injuryrepresents a major clinical problem and is a major cause for drugsbeing withdrawn from the market and of attrition in drug develop-ment. It is an infrequent event that mostly occurs after severalweeks or months of treatment and is, with few exceptions, unpre-dictable (idiosyncratic) (Lee, 2003; Uetrecht, 2008). Incidence ofdrug-induced hepatotoxicity caused by some drugs can be highas 1 in 100 patients (e.g., chlorpromazine), but in most cases is esti-mated at 1 in 10,000 to 1 in 100,000 treated patients. More than1000 drugs and herbal products have been associated with idio-syncratic hepatotoxicity (Biour et al., 2000). Such unpredictablehepatotoxicity is challenging to investigate because of its rarityand the lack of experimental models. The low prediction of drug-induced liver toxicities in human patients from studies in animalsis explained in particular by frequent and large variations in therates and routes of drug metabolism between humans and labora-tory animals (Olson et al., 2000). Estimation of ADMET properties isan important consideration for the development of novel therapeu-tic agents. Thus, predicting the ability of a drug to modulatecytochrome P450 (CYP) expression at an early stage of its discoveryand development should reduce the risk of failure in the clinic and,more importantly, permit the identification of alternative non-inducing/non-inhibiting chemical structures.

S. Anthérieu et al. / Toxicology in Vitro 26 (2012) 1278–1285 1279
To overcome poor correlation between studies of human andanimal toxicity of chemicals and reduce the costs of preclinicaldrug development, the use of in vitro human liver model systemsis highly desirable for investigating ADMET properties. Primary hu-man hepatocyte cultures are usually considered the most pertinentmodel to mimic the human liver in vivo. They are routinely used toanalyze drug uptake and kinetics parameters, estimate hepaticclearance, generate and identify metabolites, demonstrate inter-species differences and predict potential drug-drug interactions.In most cases these cells generate metabolites identical to thoseformed in vivo but in vitro-in vivo quantitative variations arefrequently observed (Le Bigot et al., 1987). Moreover, scarce andunpredictable availability, large inter-donor functional variabilityand occurrence of early phenotypic alterations when placed inculture, greatly limit their use (Guillouzo, 1998). Hepatic cell linescould represent an alternative to primary hepatocytes but mosthave limited bioactivation capacity and consequently, are oflimited interest. There is, however, at least one exception that isrepresented by the HepaRG cell line derived from a hepato-cholangiocarcinoma of a female patient and established by one ofthe authors’ group (Gripon et al., 2002). This cell line is composedof a homogeneous cell population with a karyotype exhibiting fewalterations (Gripon et al., 2002). When seeded at low density, He-paRG cells express markers of hepatic bipotent progenitors andafter proliferation the cultures contain differentiated hepatocytesand primitive biliary cells. This human liver cell line possessesthe unique property to transdifferentiate (Cerec et al., 2007). Hepa-tocyte-like cells express various phase I and phase II xenobioticmetabolizing enzyme activities as well as transporters and nuclearreceptors at levels close to those measured in primary humanhepatocytes (Aninat et al., 2006; Le Vee et al., 2006), thereby sup-porting the view that HepaRG cells possess both the metabolic per-formances of primary hepatocytes and growth capacity of hepaticcell lines (Guillouzo et al., 2007).
We examined whether these differentiated cells remained rela-tively stable functionally at confluence and could consequently beused for toxicity studies following either acute or repeat treat-ments. Within the 3-year EU framework LIINTOP project (2007–2009) our aim was to optimize the HepaRG cell model for thispurpose. More precisely, we (i) compared transcript and activitylevels of the major CYPs and their responsiveness to prototypicalinducers over a 4-week period in differentiated HepaRG cells,and (ii) evaluated the suitability of these cells for studying the ef-fects of several types of hepatotoxic drugs, with special referenceto liver-specific lesions. In the present review the main data ob-tained during this FP6 European project are summarized and dis-cussed in the light of other studies performed concomitantly ormore recently on HepaRG cells.
2. Long-term maintenance of drug metabolism capacity inHepaRG cells
2.1. Early studies
Previous studies from our laboratory had shown that whenseeded onto plastic at low density (LD) HepaRG cells took the mor-phology of elongated undifferentiated cells that exhibit markers ofliver bipotent progenitor cells and actively divided until they reachconfluence at around day 10. Then, they progressively formed typ-ical colonies of hepatocyte-like cells surrounded by clearer andmore flattened cells. Addition of 2% dimethyl sulfoxide (DMSO)from days 15 to 30 resulted in the appearance of more granularcells closely resembling mature hepatocytes delineating typicalbile canaliculus-like structures and expressing liver-specific mark-ers. The other flattened cells exhibited markers of primitive biliary
epithelial cells, such as cytokeratin 19 (Cerec et al., 2007) (Fig. 1A).Aninat and co-workers analyzed expression of various genesencoding phase 1 (CYPs) and phase 2 enzymes, nuclear receptorsand other liver-specific functions by real time-quantitative poly-merase chain reaction (RT-qPCR) and found that transcript levelsof many of these genes were close to those measured in primaryhuman hepatocyte cultures (Aninat et al., 2006). A similar conclu-sion was made by measuring corresponding activities. CYP1A1/2and CYP2E1 transcripts and activities were however lower thanin primary hepatocytes. Noticeably, expression and activity ofother CYPs, especially CYP2B6 and CYP3A4, rapidly dropped afterDMSO deprivation (Aninat et al., 2006). As observed in vivo (Gedet al., 1989), fold-inductions of several CYPs by prototypical induc-ers were directly dependent on basal activity levels; the lower thebasal activity, the higher was the induction level. The mechanismsby which DMSO increases some liver-specific functions are notfully understood (Guguen-Guillouzo and Guillouzo, 2010). DMSOacts as an inducer of several genes involved in xenobiotic detoxica-tion; it could also act as a reactive oxygen species scavenger, ananti-apoptotic agent (Gilot et al., 2002) and also likely a chromatinremodeling agent. Reversion of HepaRG cells to progenitors follow-ing enzymatic detachment can be avoided by high density (HD)seeding. In such conditions after a transient and limited decreasetranscripts and/or activity levels of the main CYPs and other func-tions fully recover (Aninat et al., 2008).
The first demonstration of the ability of HepaRG cells to retaintheir drug metabolism capacity for several weeks at confluencewas provided by Josse and co-workers (Josse et al., 2008). No sig-nificant changes were observed in the transcript levels of a subsetof genes related to drug metabolism in differentiated HepaRG cellsover a 4-week period. Moreover, activity and responsiveness toprototypical inducers of two major CYPs, CYP3A4 and CYP1A2,were also found to remain relatively stable during the same period(Josse et al., 2008).
2.2. LIINTOP studies
In order to extend our first observations on the maintenance ofdrug metabolism activities and responsiveness to inducers in con-fluent differentiated HepaRG cells, basal mRNA levels of twentygenes were determined by RT-qPCR in both LD- and HD-seededHepaRG cells over a 4-week period and the values were comparedto those measured in 1-day cultured primary human hepatocytesobtained from five donors (Antherieu et al., 2010). HepaRG cellswere used between passages 10 and 20. Indeed, only limitedchanges in functional activities are observed during the first 20passages. Tested genes included ten CYPs (CYP1A1, CYP1A2,CYP2A6, CYP2B6, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1,and CYP3A4), two phase II enzymes [glutathione transferase A1/A2 (GSTA1/A2) and UDP-glucuronosyl transferase 1A1 (UGT1A1)],five membrane transporters [breast cancer resistance protein(BCRP), bile salt export pump (BSEP), multidrug resistance pro-tein-1 (MDR1), multidrug resistance-associated protein-2 (MRP2),and Na+-dependent taurocholic cotransporting polypeptide(NTCP)] and three nuclear receptors (aryl hydrocarbon receptor(AhR), constitutive androstane receptor (CAR) and pregnane Xreceptor (PXR)). Two different media were tested: a medium con-taining 10% fetal calf serum (FCS) and 2% DMSO, and a DMSO-freemedium supplemented with 2% FCS. Briefly, all the genes were ex-pressed in DMSO-exposed HepaRG cultures. Compared to 1-dayprimary hepatocyte cultures the levels of transcripts were eitherstrongly lower (CYP1A2, CYP2C8, CYP2D6, CYP2E1, BSEP, NTCP),representing 30–50% (CYP1A1, CYP2A6, CYP2B6, PXR) or compara-ble or even higher (CYP2C9, CYP2C19, CYP3A4, GSTA1/2, UGT1A1,MDR1, MRP2, AhR, CAR), in agreement with our previous data(Aninat et al., 2006). When HepaRG cells were shifted to a

Fig. 1. Light microscopic examination of differentiated HepaRG cell cultures. HepaRG cells were incubated with: (A) the standard medium and observed under phase-contrastmicroscopy: H, hepatocyte-like cells; EC, epithelial-like cells; (B) carboxydichlorofluorescein di-acetate (fluorescent substrate of MRP2) for 30 min and then observed underfluorescence microscopy. The fluorescent substrate has accumulated in bile canaliculi. (C) 50 lM tetracycline for 14 days followed by Oil Red O staining. Note lipid dropletsaccumulation in hepatocyte-like cells; (D) 20 lM amiodarone for 24 h followed by Oil Red O staining. Unstained vesicles corresponding to lamellar bodies are visible in bothhepatocyte-like and biliary epithelial-like cells.
1280 S. Anthérieu et al. / Toxicology in Vitro 26 (2012) 1278–1285
DMSO-free medium containing 2% FCS for 72 h before mRNA quan-tification transcripts of CYP1A1, CYP2A6, CYP2B6, CYP3A4, andUGT1A1 were strongly decreased whereas those of CYP1A2,CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1, GSTA1-A2, AhR, CARand transporters were only slightly altered, indicating that the for-mer group of genes was the most sensitive to DMSO. Only CYP1A1was highly expressed in poorly differentiated HepaRG cells cul-tured for 5–15 days in the absence of DMSO after LD seeding. Geneexpression levels of all tested drug metabolizing enzymes andtransporters did not exhibit significant changes in differentiatedHepaRG cells from both LD- and HD-seeded cultures maintainedat confluence for up to 4 weeks (Antherieu et al., 2010).
CYP activities were simultaneously estimated (Antherieu et al.,2010) using two different cocktail substrate settings developed byLahoz and co-workers and Turpeinen and co-workers, and namedUAM and NM cocktails, respectively (Turpeinen et al., 2005;Lahoz et al., 2008). These UAM and NM cocktails contained eightand 10 substrates, respectively. The major differences betweenthe two assay procedures were the use of lower substrateconcentrations and longer incubation times in the NM assay. Foursubstrates were in common in the two cocktails and their concen-trations were 5-fold (bupropion, chlorzoxazone, midazolam) or2.5-fold (coumarin) higher in the UAM than that in the NM assay.As a rule, lower values were frequently obtained with the NMcocktail that was previously designed for drug-drug interactionstudies. While CYP activities in HepaRG cells were found to beeither lower (CYP1A2, CYP2A6, CYP2C9, CYP2E1) or relativelyclose (CYP2B6, CYP2C19, CYP2D6, CYP3A4) in DMSO-supple-mented medium, they were lower, especially CYP2B6 andCYP3A4, in DMSO-free medium, compared to levels in 1-day pri-mary hepatocyte cultures. In agreement with an undifferentiatedmorphology, CYP activities were all quite low, if detectable, in 5-and 15-day LD-seeded cultures. All activities were maintainedover a 28-day period at confluence in DMSO-supplementedmedium. Using the NM cocktail CYP activity values were either
comparable (CYP1A2, CYP2B6, CYP2C19, CYP2D6) to or lower(CYP2C9, CYP3A4) than those estimated with the UAM one. Fur-thermore, low CYP2A6 and CYP2E1 activities were detected withthe UAM cocktail only (Antherieu et al., 2010).
In addition, distribution of carboxydichlorofluorescein di-ace-tate and BODIPY FL vinblastine, fluorescent substrates of MRP2and BSEP/MDR1, respectively, were analyzed (Fig. 1B). Thirty min-utes after addition of either carboxydichlorofluorescein di-acetateor BODIPY FL vinblastine, bile canaliculi of HepaRG hepatocyteswere strongly labeled demonstrating the functional activity ofthese canalicular transporters (Antherieu et al., 2010).
To further estimate the functional capacity of HepaRG cells atconfluence their responsiveness to the three prototypical inducers,phenobarbital (PB), rifampicin (RIF), and omeprazole (OME), wasalso assessed over a 4-week period (Antherieu et al., 2010). At dif-ferent time-points HepaRG cell cultures were shifted to a DMSO-free medium supplemented with 2% FCS for 72 h and then treatedwith a single exposure of the prototypical inducers for 48 and 72 hfor mRNA and activity measurements, respectively. Comparable re-sults were obtained in LD- and HD-seeded HepaRG cell culturesand the levels of CYP inductions were relatively well maintainedfor 4 weeks in differentiated HepaRG cells in both culture condi-tions (Figs. 2 and 3). As expected, CYP1A2, CYP2B6, and CYP3A4transcripts and activities were strongly increased by their proto-typical inducers in both LD- and HD-seeded HepaRG cells. In addi-tion, various other CYPs were modulated by PB, RIF and/or OME.Thus, expression of CYP1A1 and 1A2 was augmented by OMEand CYP2A6 was induced by PB while CYP2E1 transcripts were de-creased by the three inducers. Noticeably, CYP2A6 and CYP2E1activities were not detected in DMSO-free HepaRG cell cultureswhatever the inducer used. Several CYP2C genes were overexpres-sed by PB and more weakly by RIF in HepaRG cells, whereas onlyRIF induced CYP2C8 and CYP2C19 expression in human hepato-cytes. Noticeably, when observed, induction rates were much low-er in human hepatocytes than in HepaRG cells. Thus, CYP1A1,
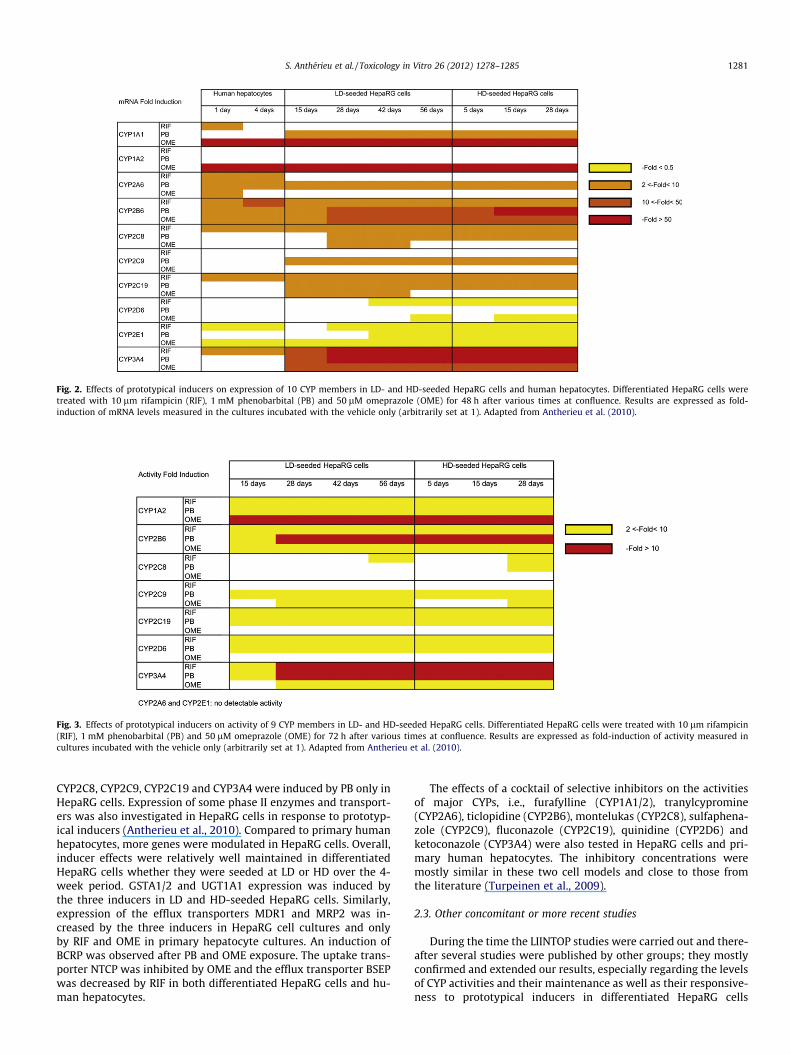
Fig. 2. Effects of prototypical inducers on expression of 10 CYP members in LD- and HD-seeded HepaRG cells and human hepatocytes. Differentiated HepaRG cells weretreated with 10 lm rifampicin (RIF), 1 mM phenobarbital (PB) and 50 lM omeprazole (OME) for 48 h after various times at confluence. Results are expressed as fold-induction of mRNA levels measured in the cultures incubated with the vehicle only (arbitrarily set at 1). Adapted from Antherieu et al. (2010).
Fig. 3. Effects of prototypical inducers on activity of 9 CYP members in LD- and HD-seeded HepaRG cells. Differentiated HepaRG cells were treated with 10 lm rifampicin(RIF), 1 mM phenobarbital (PB) and 50 lM omeprazole (OME) for 72 h after various times at confluence. Results are expressed as fold-induction of activity measured incultures incubated with the vehicle only (arbitrarily set at 1). Adapted from Antherieu et al. (2010).
S. Anthérieu et al. / Toxicology in Vitro 26 (2012) 1278–1285 1281
CYP2C8, CYP2C9, CYP2C19 and CYP3A4 were induced by PB only inHepaRG cells. Expression of some phase II enzymes and transport-ers was also investigated in HepaRG cells in response to prototyp-ical inducers (Antherieu et al., 2010). Compared to primary humanhepatocytes, more genes were modulated in HepaRG cells. Overall,inducer effects were relatively well maintained in differentiatedHepaRG cells whether they were seeded at LD or HD over the 4-week period. GSTA1/2 and UGT1A1 expression was induced bythe three inducers in LD and HD-seeded HepaRG cells. Similarly,expression of the efflux transporters MDR1 and MRP2 was in-creased by the three inducers in HepaRG cell cultures and onlyby RIF and OME in primary hepatocyte cultures. An induction ofBCRP was observed after PB and OME exposure. The uptake trans-porter NTCP was inhibited by OME and the efflux transporter BSEPwas decreased by RIF in both differentiated HepaRG cells and hu-man hepatocytes.
The effects of a cocktail of selective inhibitors on the activitiesof major CYPs, i.e., furafylline (CYP1A1/2), tranylcypromine(CYP2A6), ticlopidine (CYP2B6), montelukas (CYP2C8), sulfaphena-zole (CYP2C9), fluconazole (CYP2C19), quinidine (CYP2D6) andketoconazole (CYP3A4) were also tested in HepaRG cells and pri-mary human hepatocytes. The inhibitory concentrations weremostly similar in these two cell models and close to those fromthe literature (Turpeinen et al., 2009).
2.3. Other concomitant or more recent studies
During the time the LIINTOP studies were carried out and there-after several studies were published by other groups; they mostlyconfirmed and extended our results, especially regarding the levelsof CYP activities and their maintenance as well as their responsive-ness to prototypical inducers in differentiated HepaRG cells

1282 S. Anthérieu et al. / Toxicology in Vitro 26 (2012) 1278–1285
(Kanebratt and Andersson, 2008a,b; Kaneko et al., 2009; McGinnityet al., 2009; Hoekstra et al., 2011; Lubberstedt et al., 2011). Inagreement with our results (Aninat et al., 2006; Josse et al.,2008; Antherieu et al., 2010), they showed that basal CYP levelsand fold inductions were dependent on several parameters, suchas culture conditions, especially DMSO addition, baseline levels,choice of the substrate, concentration of the inducer and durationof exposure. Moreover, intrinsic clearances were compared in He-paRG cells and primary hepatocytes and good correlations wereobtained (Kanebratt and Andersson, 2008a; Lubberstedt et al.,2011).
Noteworthy, cultures from seeded differentiated cryopreservedHepaRG cells can also be used. The freeze/thaw process does notalter their functional activities and cryopreserved HepaRG cellsare commercially available (Biopredic International). Intrinsicclearance CLint of 26 CYP substrates with diverse properties (clear-ance, charge and lipophilicity) was compared in cryopreserved He-paRG cells, cryopreserved pooled human hepatocytes and in vivoby other LIINTOP partners and the results were recently published(Zanelli et al., 2012). CLint in HepaRG cells was on average equal toCLint in the hepatocytes and directly proportional throughout therange of in vivo CLint (1–2000 ml/min/ kg). However, an underpre-diction of up to 5-fold was observed for the most highly cleareddrugs with both in vitro cell models.
HepaRG cells have also been cultured in three dimensional bio-reactors which enabled HD cell culture under dynamic conditions.As found in 2-D cultures, most CYP activities remained stable overseveral weeks in differentiated HepaRG cells cultured in 3-Ddimension (Darnell et al., 2011).
3. Assessment of drug hepatotoxicity in HepaRG cells
Since HepaRG cells can retain their hepatocyte-specific func-tions over a 4-week duration, it makes them suitable candidatesfor repeated dose studies with hepatotoxins, the toxicity of whichis seen after chronic exposure in humans.
One of the most common causes of hepatotoxicity is the CYP-dependent formation of reactive metabolites that are directly hep-atotoxic or form adducts with hepatic proteins potentially trigger-ing an immune response. Other mechanisms include disruption ofmitochondrial functions, inhibition of xenobiotic metabolism path-ways and inhibition of bile acid transport. Consequently, drug-in-duced liver injury encompasses a large spectrum of lesions thatinclude necrosis/apoptosis, steatosis, phospholipidosis and chole-stasis (Lee, 2003).
3.1. Early studies
Only few studies had been dedicated to chemical-induced hep-atotoxicity in HepaRG cells before the LIINTOP project. It was firstdemonstrated that the two compounds acetaminophen and afla-toxin B1 (AFB1), the toxicity of which is caused by toxic metabo-lites formed by CYP-catalyzed reactions, were more cytotoxic indifferentiated HepaRG than in HepG2 cells after either 24 or 72 hof treatment (Aninat et al., 2006). Moreover, AFB1 was found tocause toxicity at decreasing concentrations by repeated exposuresand as expected, AFB1-induced damage was first restricted tohepatocyte-like cells (Josse et al., 2008).
3.2. LIINTOP studies
Within the 3-year EU framework LIINTOP project our mainobjective was to determine whether HepaRG cells represent a suit-able in vitro cell model for the study of the different types of chem-
ical-induced hepatic lesions, and consequently to focus mainly onsteatosis, phospholipidosis and cholestasis.
3.2.1. SteatosisMany drugs have been classified as steatogenic. Hepatic steato-
sis refers as to an accumulation of triglycerides (TG) into intracyto-plasmic vesicles in hepatocytes. It represents a reversible state ofmetabolic dysfunction that can possibly progress to inflammatorysteatohepatitis, fibrosis, cirrhosis, and even hepatocellular carci-noma (Farrell and Larter, 2006; Reddy and Rao, 2006). Accumula-tion of such typical lipid vesicles following treatment withsteatogenic drugs had never been demonstrated in human liver cellcultures. Only increased TG content had been evidenced (Donatoet al., 2009; Moya et al., 2010). By using the Oil Red O assay, wefound that the steatogenic drug tetracycline at 50 lM inducedaccumulation of neutral lipids in hepatocyte-like HepaRG cells(but not in primitive biliary-like cells) after a 24-h treatment(Antherieu et al., 2011). This accumulation was amplified after re-peated exposure every 2–3 days for 14 days (Fig. 1C) and was alsoobserved after repeated treatment with 20 lM amiodarone for14 days. These results were confirmed by quantification of TG bygas/liquid chromatography. Transcriptomic analysis showed thatgenes involved in de novo lipogenesis (e.g., sterol regulatory ele-ment-binding protein 1 (SREBP1), peroxisome proliferator-acti-vated receptors gamma (PPARG), thyroid hormone-induciblehepatic protein (THRSP), fatty acid synthase (FASN) and ATP-citratesynthase (ACLY)) and lipid droplet formation (adipose differentia-tion-related protein (ADFP) and perilipin-4 (PLIN4)) were up-regu-lated by both amiodarone and tetracycline. Moreover a reductionof fatty acid oxidation was evidenced by measuring the [14C]-la-beled acid-soluble b-oxidation products in HepaRG cells treatedwith the two drugs. These results represent the first demonstrationthat drugs can induce vesicular steatosis in vitro and show a directrelationship between TG accumulation and enhanced expression oflipogenic genes (Antherieu et al., 2011). Noticeably, severe micro-vesicular steatosis has been reported in patients with high serumand liver (1–2 mM) concentrations of amiodarone (Rotmenschet al., 1984; Brien et al., 1987) and tetracycline (Whalley et al.,1964; Freneaux et al., 1988) after chronic use in humans. Com-pared to these in vivo data, it appears that steatosis can be inducedin HepaRG cells at relatively low drug concentrations.
Microvesicular steatosis was also induced by a 24-h exposure ofHepaRG cells to various polyunsaturated fatty acids and deriva-tives (Antherieu et al., 2011; Madec et al., 2011). Noteworthy, theliver-specific CYP4F3B, that plays a major role in energy homeosta-sis and lipid metabolism, was highly expressed in differentiatedHepaRG cells (Madec et al., 2011).
3.2.2. PhospholipidosisPhospholipidosis is characterized by an excessive intracellular
accumulation of phospholipids typified as lamellar bodies thatare easily detected by electron microscopy examination. More than50 cationic amphiphilic drugs, including amiodarone and chlor-promazine, have been reported to induce phospholipidosis in fewpatients (Shaikh et al., 1987; Halliwell, 1997). This lesion can bevisualized in various hepatic (Poucell et al., 1984) and nonhepaticcell types (Lullmann et al., 1975). Typical intracytoplasmic concen-tric lamellar structures of both hepatocyte-like and biliary-likecells were observed in HepaRG cell cultures after a 24-h treatmentwith 20 lM amiodarone (Fig. 1D). They were enhanced over a 2-week period by exposing the cells every 2–3 days to this non cyto-toxic amiodarone concentration. Accumulation of phospholipidswas evidenced by determination of various phospholipid entitiesby high-performance liquid chromatography. Several genes wereidentified as deregulated by amiodarone treatment. They includedup-regulated ones such as acyl-CoA desaturase (SCD), a fatty acid

S. Anthérieu et al. / Toxicology in Vitro 26 (2012) 1278–1285 1283
biosynthesis-related gene; glycerophosphodiester phosphodiester-ase domain containing 3 (GDPD3) and acid sphingomyelinase-likephosphodiesterase 3a (ASML3A), two genes involved in phospho-lipid degradation, and lipoprotein lipase (LPL), that were specifi-cally modulated after repeat exposure to amiodarone andprobably corresponded to a defense mechanism to reduce phos-pholipids accumulation (Antherieu et al., 2011). Another cationicamphiphilic drug, chlorpromazine, which has been reported to in-duce phospholipidosis in a few patients (Shaikh et al., 1987; Halli-well, 1997), was also recently shown to induce typical lamellarbodies in both hepatocyte-like and biliary-like HepaRG cells. Theselamellar bodies were associated with an overexpression of ADFPand PLIN4, two genes involved in the formation of such vesicles.In addition, SCD1 and lipin-1 (LPIN1) that both are involved in syn-thesis of phospholipids were up-regulated. All these genes whichwere similarly deregulated by amiodarone represent potential bio-markers of drug-induced phospholipidosis in humans (Antherieuet al., submitted).
3.2.3. CholestasisCholestasis is a clinical syndrome that results from the hepatic
retention of products normally excreted into bile, in particular bilesalts. This pathology is induced by various factors, including anumber of drugs. The mechanisms of drug-induced cholestasisare varied and poorly understood in humans. Current models usedfor safety assessment in drug development do not accurately pre-dict cholestasis in humans. Inhibition of the activity of transportersresponsible for efflux of bile acids and other biliary constituentsinto canalicular spaces of hepatocytes is thought to represent animportant mechanism of cholestatic drug hepatotoxicity (Paddaet al., 2011). A well-known cholestatic drug, the anticonvulsantchlorpromazine, was investigated in differentiated HepaRG cells.As expected, an inhibition of bile acids efflux was observed earlyafter chlorpromazine treatment as shown by [H3]-taurocholic acidaccumulation. It was associated with induction of an oxidativestress. In addition, increased efflux basolateral transport and de-creased bile acid synthesis were observed after 24 h; these lattereffects could represent compensatory hepatoprotective mecha-nisms aiming at reducing intrahepatic accumulation of toxic bileacids (Antherieu et al., manuscript submitted).
3.3. Other concomitant or more recent studies
The conclusion that the HepaRG cell line is the closest in vitrocell model to primary hepatocytes and can be used as a surrogatehas been largely based on drug metabolism studies. Our work de-scribed above also supports the view that HepaRG cells are prom-ising for hepatotoxicity studies. Interestingly, other investigationshave come to the same conclusion. Indeed, several studies havecompared HepaRG gene expression profiles with other liver mod-els, and changes in gene expression profiles induced by hepato-toxic compounds. Other works have been dedicated tomechanisms of chemical cytotoxicity and evaluation of HepaRGcells for genotoxicity testing as well as effects of acute and repeatexposures to low concentrations of environmental contaminantseither individually or in mixtures.
The close equivalence of the HepaRG cell line with primaryhepatocytes has also been demonstrated by several transcriptomicanalyses (Lambert et al., 2009; Hart et al., 2010; Jennen et al., 2010;Rogue et al., 2011, 2012). HepaRG cells were shown to expressmost of the genes normally active in human hepatocytes. Thus,we have reported that HepaRG cells expressed from 81% to 92%of the genes active in 2-day cultures of primary human hepato-cytes from different donors and could be classified as an ‘‘averagehuman hepatocyte population’’ (Rogue et al., 2012). However, itmust be borne in mind that in addition these cells express a spe-
cific gene subset mainly related to their transformed status, somechromosomal abnormalities, and the presence of progenitors/primitive biliary epithelial cells (Rogue et al., 2012).
Differentiated HepaRG cells have served to analyze geneexpression profiles induced by several potential hepatotoxic com-pounds, including phenobarbital, PPARc and PPARa/c agonists andAFB1, in comparison with primary human hepatocytes. Most of themain deregulated genes after 20-h treatment with 1 mM pheno-barbital were the same in both in vitro cell models (Lambertet al., 2009). Phenobarbital-responsive genes included the well-recognized CAR- and PXR-dependent responsive cytochromesP450 (CYP2B6, CYP3A4), sulfotransferase 2A1 and plasma mem-brane transporters (ABCB1, ABCC2), as well as a number of genescritically involved in various metabolic pathways, including lipid(CYP4A11, CYP4F3), vitamin D (CYP24A1) and bile (CYP7A1 andCYP8B1) metabolism (Lambert et al., 2009). Gene expression pro-files induced by different concentrations of two PPARc (troglitaz-one and rosiglitazone) and two PPARa/c (muraglitazar andtesaglitazar) agonists were also compared in HepaRG cells and pri-mary human hepatocytes from several donors after a 24-h treat-ment (Rogue et al., 2011). Many genes involved in lipid,carbohydrate, xenobiotic and cholesterol metabolism, as well asinflammation and immunity, were regulated by both PPARc andPPARa/c agonists in at least a number of human hepatocyte popu-lations and/or HepaRG cells. Very few genes were selectivelyderegulated by glitazars when compared to glitazones, indicatingthat PPARc and PPARa/c agonists share most of their target genes.Hierarchical clustering analysis showed that some human hepato-cyte populations were closer to HepaRG cells than to others (Rogueet al., 2011). Recently, the transcriptomic approach was used toanalyze early gene expression profiles induced by AFB1 and toidentify target genes. Several genes related to the p53 pathwayand many other genes, including (proto)-oncogenes and tumorsuppressor genes, were found to be altered in both cell models.Moreover, several target genes, especially fragile histidine triadgene (FHIT), breast carcinoma amplified sequence 3 (BCAS3) andSET and MYND domain containing 3 (SMYD3), were found to beresponsive to direct genotoxins, as well as genotoxic compoundsrequiring bioactivation and could represent potent biomarkers todiscriminate genotoxic from non-genotoxic and other compounds(Josse et al., 2012a).
Mechanisms of acetaminophen-induced cell death have beenanalyzed in differentiated HepaRG cells and were found to be thesame as those characterized in rodent in vivo models and primarymouse hepatocyte cultures, i.e., glutathione depletion, acetamino-phen protein adducts, mitochondrial dysfunction and lactate dehy-drogenase release (McGill et al., 2011). However, no caspaseactivity increase was observed; one explanation could be the highconcentrations used; however, the authors arrived them basedupon concentrations found in clinical cases, adjusted for empiricaldata on acetaminophen’s in vivo half-life. HepaRG cells have alsobeen used to compare the toxicity of three thiopurines (azathiopu-rine, 6-mercaptopurine and 6-thiopurine) (Petit et al., 2008) and todemonstrate that hypoxia down-regulates drug-metabolizing en-zymes (Legendre et al., 2009).
Metabolically competent HepaRG cells have served to assess thegenotoxic potential of indirect chemical mutagens using the cometand micronucleus assays and the results obtained with a limitedset of compounds were promising (Josse et al., 2008; Le Hegaratet al., 2010). Recently, we proposed an adaptation of the in vitromicronucleus assay for the HepaRG cells that consisted in the useof cultures containing more than 80% mature hepatocytes andtreatment in situ with various chemicals for 24 h followed by a3-day mitogenic stimulation with EGF without cytokinesis block.In such conditions the test was more sensitive with certain indirectmutagens, such as AFB1 (Josse et al., 2012b).

1284 S. Anthérieu et al. / Toxicology in Vitro 26 (2012) 1278–1285
HepaRG cells have also been used to investigate the effects ofacute and repeated exposures to low concentrations of environ-mental contaminants either individually or in mixtures, usingvarious experimental approaches, including transcriptomic tech-nologies. In a first study, two common heterocyclic aromaticamines, 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP)and 2-amino-3,8-dimethylimidazo[4,5-f]quinoxaline (MeIQx)were compared (Dumont et al., 2010b). After a 24-h treatment,PhIP and MeIQx, individually and in combination, exerted differen-tial effects on apoptosis, oxidative stress, DNA damage and CYPactivities. Only PhIP induced DNA damage. It was also a strongerinducer of CYP1A1 and CYP1B1 expression and activity than MeI-Qx. In contrast, only MeIQx exposure resulted in a significantinduction of CYP1A2 activity. The equimolar combination of PhIPwith MeIQx induced an oxidative stress and showed synergistic ef-fects on apoptosis but inhibitory effects on DNA damage. In addi-tion, gene expression profiles were analyzed after either a single(24-h) or a repeated (28-day) exposure to PhIP or MeIQx (Dumontet al., 2010a). The most responsive genes to both HAA were down-stream targets of the AhR, i.e., CYP1A1 and CYP1A2 after both timepoints and CYP1B1 and aldehyde dehydrogenase 3 family, memberA1 (ALDH3A1) after 28 days. Several other genes exhibited bothtime-dependent and compound-specific gene expression changeswith, however, a smaller magnitude than that obtained for AhRtarget genes. These findings highlight the need for investigationson both acute and chronic effects of contaminants, singly and inmixtures, as well as on their interactions when assessing risks forhuman health and demonstrate the unique properties of HepaRGcells as a metabolically competent cell line for acute and chronicchemical toxicity testing.
4. Conclusions
In the course of the LIINTOP project we performed an extensivecharacterization of long-term maintenance of drug metabolismcapacity and responsiveness to prototypical inducers in HepaRGcells and showed that these cells remained functionally relativelystable for several weeks at confluence, allowing evaluation of bothacute and chronic drug-induced liver injury. Reproducible datawere obtained and the levels of functions could be modulated byusing different culture conditions. If in standard culture conditionsdifferentiated HepaRG cells contain mature hepatocytes and prim-itive biliary epithelial cells in around equal percentages (Cerecet al., 2007), nearly pure HepaRG hepatocyte cultures (containingup to 80–90% hepatocytes) can be obtained in appropriate experi-mental conditions based on the use of DMSO during cell detach-ment and high-density seeding (Pernelle et al., 2011). Althoughnon-significant changes were observed in expression and activityof drug metabolizing enzymes during the long-term maintenanceat confluence, recent observations have evidenced that the cellsaged after having reached confluence, as shown by a time-depen-dent increase of H6-dependent b-galactosidase activity in absenceof DMSO (Pernelle et al., 2011). Depending on the scientific objec-tive, culture conditions have to be optimized. Obviously, DMSO fa-vored formation of typical bile canaliculi and induced somefunctions such as CYP3A4, CYP2B6 and CYP1A2. However, DMSO-free cultures can be more suitable for the expression of some func-tions. Thus, a 2-fold increase in ammonia elimination was obtainedby adding carbamoyl-glutamate in the absence of DMSO (Hoekstraet al., 2011).
Since drug-induced liver injury usually requires metabolism ofthe drug to form reactive metabolites and free radicals (Parket al., 2005), the use of metabolically competent human hepaticcells for hepatotoxicity studies is critical. Moreover, it mostly oc-curs after several weeks or months of treatment. HepaRG cells pos-
sess the unique properties to remain metabolically active forseveral weeks and to permit repeat drug treatments. Accordingly,the different types of chemical-induced hepatotoxicities, includingsteatosis, phospholipidosis and cholestasis, can be induced bychemicals in HepaRG cells. In particular, for the first time vesicularsteatosis was obtained in vitro after repeat treatment with steato-genic drugs using these cells. HepaRG cells also respond to cyto-kines (Aninat et al., 2008). Therefore, inflamed and diseasedHepaRG cells represent attractive cell models to mimic the effectsof drugs in human diseases.
A new potential source of unlimited numbers of differentiatedhepatocytes is hoped to arise from stem cells either isolated fromembryos or adult tissues or obtained by reprogramming somaticcells but presently, only hepatocyte-like cells expressing low levelsof drug metabolizing and detoxifying enzymes, are usually ob-tained, making them still unsuitable as metabolically competentcells for toxicity studies (Guguen-Guillouzo et al., 2010). Therefore,HepaRG cells remain the sole liver cell model pertinent for repro-ducible studies on acute and chronic effects of chemicals in meta-bolically competent human liver cells.
Acknowledgements
The studies related to the LIINTOP project that are described inthis review, were supported by the EEC contract LIINTOP-STREP-037499. We thank Dr. Flavia Zucco for her excellent co-ordinationof the LIINTOP project, Dr. Agustin Lahoz (Avancell, Valencia) andDr. Jouko Uusitalo (Novamass Ltd, Oulu) and their colleagues fortheir involvement in the original work published in Drug Metabo-lism and Disposition (2010) and Dr. David Steen for critical readingof the manuscript.
References
Aninat, C., Piton, A., Glaise, D., Le Charpentier, T., Langouet, S., Morel, F., Guguen-Guillouzo, C., Guillouzo, A., 2006. Expression of cytochromes P450, conjugatingenzymes and nuclear receptors in human hepatoma HepaRG cells. Drug Metab.Dispos. 34, 75–83.
Aninat, C., Seguin, P., Descheemaeker, P.N., Morel, F., Malledant, Y., Guillouzo, A.,2008. Catecholamines induce an inflammatory response in human hepatocytes.Crit. Care Med. 36, 848–854.
Antherieu, S., Chesne, C., Li, R., Camus, S., Lahoz, A., Picazo, L., Turpeinen, M.,Tolonen, A., Uusitalo, J., Guguen-Guillouzo, C., Guillouzo, A., 2010. Stableexpression, activity, and inducibility of cytochromes P450 in differentiatedHepaRG cells. Drug Metab. Dispos. 38, 516–525.
Antherieu, S., Rogue, A., Fromenty, B., Guillouzo, A., Robin, M.A., 2011. Induction ofvesicular steatosis by amiodarone and tetracycline is associated with up-regulation of lipogenic genes in heparg cells. Hepatology 53, 1895–1905.
Biour, M., Poupon, R., Grange, J.D., Chazouilleres, O., 2000. Drug-inducedhepatotoxicity. The 13th updated edition of the bibliographic database ofdrug-related liver injuries and responsible drugs. Gastroenterol. Clin. Biol. 24,1052–1091.
Brien, J.F., Jimmo, S., Brennan, F.J., Ford, S.E., Armstrong, P.W., 1987. Distribution ofamiodarone and its metabolite, desethylamiodarone, in human tissues. Can. J.Physiol. Pharmacol. 65, 360–364.
Cerec, V., Glaise, D., Garnier, D., Morosan, S., Turlin, B., Drenou, B., Gripon, P.,Kremsdorf, D., Guguen-Guillouzo, C., Corlu, A., 2007. Transdifferentiation ofhepatocyte-like cells from the human hepatoma HepaRG cell line throughbipotent progenitor. Hepatology 45, 957–967.
Darnell, M., Schreiter, T., Zeilinger, K., Urbaniak, T., Soderdahl, T., Rossberg, I.,Dillner, B., Berg, A.L., Gerlach, J.C., Andersson, T.B., 2011. Cytochrome P450-dependent metabolism in HepaRG cells cultured in a dynamic three-dimensional bioreactor. Drug Metab. Dispos. 39, 1131–1138.
Donato, M.T., Martinez-Romero, A., Jimenez, N., Negro, A., Herrera, G., Castell, J.V.,O’Connor, J.E., Gomez-Lechon, M.J., 2009. Cytometric analysis for drug-inducedsteatosis in HepG2 cells. Chem. Biol. Interact. 181, 417–423.
Dumont, J., Josse, R., Lambert, C., Antherieu, S., Laurent, V., Loyer, P., Robin, M.A.,Guillouzo, A., 2010a. Preferential induction of the AhR gene battery in HepaRGcells after a single or repeated exposure to heterocyclic aromatic amines.Toxicol. Appl. Pharmacol. 249, 91–100.
Dumont, J., Josse, R., Lambert, C., Antherieu, S., Le Hegarat, L., Aninat, C., Robin, M.A.,Guguen-Guillouzo, C., Guillouzo, A., 2010b. Differential toxicity of heterocyclicaromatic amines and their mixture in metabolically competent HepaRG cells.Toxicol. Appl. Pharmacol. 245, 256–263.
Farrell, G.C., Larter, C.Z., 2006. Nonalcoholic fatty liver disease: from steatosis tocirrhosis. Hepatology 43, S99–S112.

S. Anthérieu et al. / Toxicology in Vitro 26 (2012) 1278–1285 1285
Freneaux, E., Labbe, G., Letteron, P., The Le, D., Degott, C., Geneve, J., Larrey, D.,Pessayre, D., 1988. Inhibition of the mitochondrial oxidation of fatty acids bytetracycline in mice and in man: possible role in microvesicular steatosisinduced by this antibiotic. Hepatology 8, 1056–1062.
Ged, C., Rouillon, J.M., Pichard, L., Combalbert, J., Bressot, N., Bories, P., Michel, H.,Beaune, P., Maurel, P., 1989. The increase in urinary excretion of 6 beta-hydroxycortisol as a marker of human hepatic cytochrome P450IIIA induction.Br. J. Clin. Pharmacol. 28, 373–387.
Gilot, D., Loyer, P., Corlu, A., Glaise, D., Lagadic-Gossmann, D., Atfi, A., Morel, F.,Ichijo, H., Guguen-Guillouzo, C., 2002. Liver protection from apoptosis requiresboth blockage of initiator caspase activities and inhibition of ASK1/JNK pathwayvia glutathione S-transferase regulation. J. Biol. Chem. 277, 49220–49229.
Gripon, P., Rumin, S., Urban, S., Le Seyec, J., Glaise, D., Cannie, I., Guyomard, C., Lucas,J., Trepo, C., Guguen-Guillouzo, C., 2002. Infection of a human hepatoma cell lineby hepatitis B virus. Proc. Natl. Acad. Sci. USA 99, 15655–15660.
Guguen-Guillouzo, C., Guillouzo, A., 2010. General review on in vitro hepatocytemodels and their applications. Methods Mol. Biol. 640, 1–40.
Guguen-Guillouzo, C., Corlu, A., Guillouzo, A., 2010. Stem cell-derived hepatocytesand their use in toxicology. Toxicology 270, 3–9.
Guillouzo, A., 1998. Liver cell models in in vitro toxicology. Environ. HealthPerspect. 106 (Suppl 2), 511–532.
Guillouzo, A., Corlu, A., Aninat, C., Glaise, D., Morel, F., Guguen-Guillouzo, C., 2007.The human hepatoma HepaRG cells: a highly differentiated model for studies ofliver metabolism and toxicity of xenobiotics. Chem. Biol. Interact. 168, 66–73.
Halliwell, W.H., 1997. Cationic amphiphilic drug-induced phospholipidosis. Toxicol.Pathol. 25, 53–60.
Hart, S.N., Li, Y., Nakamoto, K., Subileau, E.A., Steen, D., Zhong, X.B., 2010. Acomparison of whole genome gene expression profiles of HepaRG cells andHepG2 cells to primary human hepatocytes and human liver tissues. DrugMetab. Dispos. 38, 988–994.
Hoekstra, R., Nibourg, G.A., van der Hoeven, T.V., Ackermans, M.T., Hakvoort, T.B.,van Gulik, T.M., Lamers, W.H., Elferink, R.P., Chamuleau, R.A., 2011. The HepaRGcell line is suitable for bioartificial liver application. Int. J. Biochem. Cell Biol. 43,1483–1489.
Jennen, D.G., Magkoufopoulou, C., Ketelslegers, H.B., van Herwijnen, M.H., Kleinjans,J.C., van Delft, J.H., 2010. Comparison of HepG2 and HepaRG by whole-genomegene expression analysis for the purpose of chemical hazard identification.Toxicol. Sci. 115, 66–79.
Josse, R., Aninat, C., Glaise, D., Dumont, J., Fessard, V., Morel, F., Poul, J.M., Guguen-Guillouzo, C., Guillouzo, A., 2008. Long-term functional stability of humanHepaRG hepatocytes and use for chronic toxicity and genotoxicity studies. DrugMetab. Dispos. 36, 1111–1118.
Josse, R., Dumont, J., Fautrel, A., Robin, M.A., Guillouzo, A., 2012a. Identification ofearly target genes of aflatoxin B1 in human hepatocytes, inter-individualvariability and comparison with other genotoxic compounds. Toxicol. Appl.Pharmacol. 258, 176–187.
Josse, R., Rogue, A., Lorge, E., Guillouzo, A., 2012b. An adaptation of the humanHepaRG cells to the in vitro micronucleus assay. Mutagenesis 27, 295–304.
Kanebratt, K.P., Andersson, T.B., 2008a. Evaluation of HepaRG cells as an in vitromodel for human drug metabolism studies. Drug Metab. Dispos. 36, 1444–1452.
Kanebratt, K.P., Andersson, T.B., 2008b. HepaRG cells as an in vitro model forevaluation of cytochrome P450 induction in humans. Drug Metab. Dispos. 36,137–145.
Kaneko, A., Kato, M., Sekiguchi, N., Mitsui, T., Takeda, K., Aso, Y., 2009. In vitro modelfor the prediction of clinical CYP3A4 induction using HepaRG cells. Xenobiotica39, 803–810.
Lahoz, A., Donato, M.T., Picazo, L., Castell, J.V., Gomez-Lechon, M.J., 2008.Assessment of cytochrome P450 induction in human hepatocytes using thecocktail strategy plus liquid chromatography tandem mass spectrometry. DrugMetab. Lett. 2, 205–209.
Lambert, C.B., Spire, C., Claude, N., Guillouzo, A., 2009. Dose- and time-dependenteffects of phenobarbital on gene expression profiling in human hepatomaHepaRG cells. Toxicol. Appl. Pharmacol. 234, 345–360.
Le Bigot, J.F., Begue, J.M., Kiechel, J.R., Guillouzo, A., 1987. Species differences inmetabolism of ketotifen in rat, rabbit and man: demonstration of similarpathways in vivo and in cultured hepatocytes. Life Sci. 40, 883–890.
Lee, W.M., 2003. Drug-induced hepatotoxicity. N. Engl. J. Med. 349, 474–485.Legendre, C., Hori, T., Loyer, P., Aninat, C., Ishida, S., Glaise, D., Lucas-Clerc, C.,
Boudjema, K., Guguen-Guillouzo, C., Corlu, A., Morel, F., 2009. Drug-metabolising enzymes are down-regulated by hypoxia in differentiatedhuman hepatoma HepaRG cells: HIF-1alpha involvement in CYP3A4repression. Eur. J. Cancer 45, 2882–2892.
Le Hegarat, L., Dumont, J., Josse, R., Huet, S., Lanceleur, R., Mourot, A., Poul, J.M.,Guguen-Guillouzo, C., Guillouzo, A., Fessard, V., 2010. Assessment of thegenotoxic potential of indirect chemical mutagens in HepaRG cells by the cometand the cytokinesis-block micronucleus assays. Mutagenesis 25, 555–560.
Le Vee, M., Jigorel, E., Glaise, D., Gripon, P., Guguen-Guillouzo, C., Fardel, O., 2006.Functional expression of sinusoidal and canalicular hepatic drug transporters inthe differentiated human hepatoma HepaRG cell line. Eur. J. Pharm. Sci. 28,109–117.
Lubberstedt, M., Muller-Vieira, U., Mayer, M., Biemel, K.M., Knospel, F., Knobeloch,D., Nussler, A.K., Gerlach, J.C., Zeilinger, K., 2011. HepaRG human hepatic cellline utility as a surrogate for primary human hepatocytes in drug metabolismassessment in vitro. J. Pharmacol. Toxicol. Methods 63, 59–68.
Lullmann, H., Lullmann-Rauch, R., Wassermann, O., 1975. Drug-inducedphospholipidosis II. Tissue distribution of the amphiphilic drugchlorphentermine. CRC Crit. Rev. Toxicol. 4, 185–218.
Madec, S., Cerec, V., Plee-Gautier, E., Antoun, J., Glaise, D., Salaun, J.P., Guguen-Guillouzo, C., Corlu, A., 2011. CYP4F3B expression is associated withdifferentiation of HepaRG human hepatocytes and unaffected by fatty acidoverload. Drug Metab. Dispos. 39, 1987–1996.
McGill, M.R., Yan, H.M., Ramachandran, A., Murray, G.J., Rollins, D.E., Jaeschke, H.,2011. HepaRG cells: a human model to study mechanisms of acetaminophenhepatotoxicity. Hepatology 53, 974–982.
McGinnity, D.F., Zhang, G., Kenny, J.R., Hamilton, G.A., Otmani, S., Stams, K.R., Haney,S., Brassil, P., Stresser, D.M., Riley, R.J., 2009. Evaluation of multiple in vitrosystems for assessment of CYP3A4 induction in drug discovery: humanhepatocytes, pregnane X receptor reporter gene, and Fa2N-4 and HepaRGcells. Drug Metab. Dispos. 37, 1259–1268.
Moya, M., Gomez-Lechon, M.J., Castell, J.V., Jover, R., 2010. Enhanced steatosis bynuclear receptor ligands: a study in cultured human hepatocytes and hepatomacells with a characterized nuclear receptor expression profile. Chem. Biol.Interact. 184, 376–387.
Olson, H., Betton, G., Robinson, D., et al., 2000. Concordance of the toxicity ofpharmaceuticals in humans and in animals. Regul. Toxicol. Pharmacol. 32, 56–67.
Padda, M.S., Sanchez, M., Akhtar, A.J., Boyer, J.L., 2011. Drug-induced cholestasis.Hepatology 53, 1377–1387.
Park, B.K., Kitteringham, N.R., Maggs, J.L., Pirmohamed, M., Williams, D.P., 2005. Therole of metabolic activation in drug-induced hepatotoxicity. Annu. Rev.Pharmacol. Toxicol. 45, 177–202.
Pernelle, K., Le Guevel, R., Glaise, D., Stasio, C.G., Le Charpentier, T., Bouaita, B., Corlu,A., Guguen-Guillouzo, C., 2011. Automated detection of hepatotoxic compoundsin human hepatocytes using HepaRG cells and image-based analysis ofmitochondrial dysfunction with JC-1 dye. Toxicol. Appl. Pharmacol. 254, 256–266.
Petit, E., Langouet, S., Akhdar, H., Nicolas-Nicolaz, C., Guillouzo, A., Morel, F., 2008.Differential toxic effects of azathioprine, 6-mercaptopurine and 6-thioguanineon human hepatocytes. Toxicol. In Vitro 22, 632–642.
Poucell, S., Ireton, J., Valencia-Mayoral, P., Downar, E., Larratt, L., Patterson, J.,Blendis, L., Phillips, M.J., 1984. Amiodarone-associated phospholipidosis andfibrosis of the liver. Light, immunohistochemical, and electron microscopicstudies. Gastroenterology 86, 926–936.
Reddy, J.K., Rao, M.S., 2006. Lipid metabolism and liver inflammation. II. Fatty liverdisease and fatty acid oxidation. Am. J. Physiol. Gastrointest. Liver Physiol. 290,G852–G858.
Rogue, A., Lambert, C., Josse, R., Antherieu, S., Spire, C., Claude, N., Guillouzo, A.,2011. Comparative gene expression profiles induced by PPARgamma andPPARalpha/gamma agonists in human hepatocytes. PLoS ONE 6, e18816.
Rogue, A., Lambert, C., Spire, C., Claude, N., Guillouzo, A., 2012. Interindividualvariability in gene expression profiles in human hepatocytes and comparisonwith HepaRG cells. Drug Metab. Dispos. 40, 151–158.
Rotmensch, H.H., Belhassen, B., Swanson, B.N., Shoshani, D., Spielman, S.R.,Greenspon, A.J., Greenspan, A.M., Vlasses, P.H., Horowitz, L.N., 1984. Steady-state serum amiodarone concentrations: relationships with antiarrhythmicefficacy and toxicity. Ann. Intern. Med. 101, 462–469.
Shaikh, N.A., Downar, E., Butany, J., 1987. Amiodarone–an inhibitor ofphospholipase activity: a comparative study of the inhibitory effects ofamiodarone, chloroquine and chlorpromazine. Mol. Cell. Biochem. 76, 163–172.
Turpeinen, M., Tolonen, A., Chesne, C., Guillouzo, A., Uusitalo, J., Pelkonen, O., 2009.Functional expression, inhibition and induction of CYP enzymes in HepaRGcells. Toxicol. In Vitro 23, 748–753.
Turpeinen, M., Uusitalo, J., Jalonen, J., Pelkonen, O., 2005. Multiple P450 substratesin a single run: rapid and comprehensive in vitro interaction assay. Eur. J.Pharm. Sci. 24, 123–132.
Uetrecht, J., 2008. Idiosyncratic drug reactions: past, present, and future. Chem. Res.Toxicol. 21, 84–92.
Whalley, P.J., Adams, R.H., Combes, B., 1964. Tetracycline toxicity in pregnancy.Liver and pancreatic dysfunction. JAMA 189, 357–362.
Zanelli, U., Caradonna, N.P., Hallifax, D., Turlizzi, E., Houston, J.B., 2012. Comparisonof cryopreserved HepaRG cells with cryopreserved human hepatocytes forprediction of clearance for 26 drugs. Drug Metab. Dispos. 40, 104–110.




















