
On the early emergence of reverse transcription: Theoretical basis and experimental evidence
13
J Mol Evol (1992) 35:524-536 Journal of Molecular Evolution © Springer-Verlag NewYorkInc. 1992 On the Early Emergence of Reverse Transcription: Theoretical Basis and Experimental Evidence A. Lazcano, x V. Valverde, 1'3 G. Hern~mdez, 1 P. Gariglio, 2'3 G.E. Fox, 4 and J. Or6 4 1 Departamento de Biologfa, Facultad de Ciencias-UNAM, Apartado Postal 70-407, Cd. Universitaria, M6xico 04511, D.F., M6xico 2 Departamento de Gen6tica y Biologia Molecular, CINVESTAV-IPN, Apartado Postal 14-740, M6xico 07000, D.F., M6xico 3 Departamento de Gen6tica y Patologfa Experimental, Instituto Nacional de Salud POblica, CISEI, Cuernavaca, Morelos, M6xico 4 Department of Biochemical & Biophysical Sciences, University of Houston, Houston, TX 77204-5500, USA Summary. Reverse transcriptase (RT) was first discovered as an essential catalyst in the biological cycle of retroviruses. However, in the past years evidence has accumulated showing that RTs are in- volved in a surprisingly large number of RNA- mediated transpositional events that include both viral and nonviral genetic entities. Although it is probable that some RT-bearing genetic elements like the different types of AIDS viruses and the mammalian LINE family have arisen in recent geo- logical times, the possibility that reverse transcrip- tion first took place in the early Archean is sup- ported by (1) the hypothesis that RNA preceded DNA as cellular genetic material; (2) the existence of homologous regions of the subunit tau of the E. coli DNA polymerase III with the simian immuno- deficiency virus RT, the hepatitis B virus RT, and the 13' subunit of the E. coli RNA polymerase (McHenry et al. 1988); (3) the presence of several conserved motifs, including a 14-amino-acid seg- ment that consists of an Asp-Asp pair flanked by hydrophobic amino acids, which are found in all RTs and in most cellular and viral RNA poly- merases. However, whether extant RTs descend from the primitive polymerase involved in the Abbreviations: RT, reverse transcriptase; MMLV, Moloney rou- tine leukemia virus; MMTV, mouse mammary tumor virus; HIV, human immunodeficiency virus; SNV, spleen necrosis vi- rus; ASLV, avian sarcoma leukosis virus; RSV, Rous sarcoma virus; NTPs, ribonucleotides; dNTPs, deoxyribonucleotides Offprint requests to: A. Lazcano RNA-to-DNA transition remains unproven. Sub- strate specificity of the AMV and HIV-1 RTs can be modified in the presence of Mn 2÷ , a cation which allows them to add ribonucleotides to an oligo (dG) primer in a template-dependent reaction. This change in specificity is comparable to that observed under similar conditions in other nucleic acid poly- merases. This experimentally induced change in RT substrate specificity may explain previous observa- tions on the misincorporation of ribonucleotides by the Maloney murine sarcoma virus RT in the minus and plus DNA of this retrovirus (Chen and Temin 1980). Our results also suggest that HIV-infected macrophages and T-cell cells may contain mixed polynucleotides containing both ribo- and deoxyri- bonucleotides. The evolutionary significance of these changes in substrate specificities of nucleic acid polymerases is also discussed. Key words: Reverse transcriptase -- RNA ge- nomes -- Reverse transcriptase substrate ambigu- ities -- Early biological evolution Introduction Reverse transcriptase (RT) is an RNA-dependent DNA polymerase (Fig. 1) first discovered in exog- enous retroviruses (Baltimore 1970; Temin and Mi- zutani 1970), and later found to be encoded by a wide variety of genetic elements that are known as retroids (Fuetterer and Hohn 1987). It is known that
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of On the early emergence of reverse transcription: Theoretical basis and experimental evidence

J Mol Evol (1992) 35:524-536
Journal of Molecular Evolution © Springer-Verlag New York Inc. 1992
On the Early Emergence of Reverse Transcription: Theoretical Basis and Experimental Evidence
A. Lazcano, x V. Valverde, 1'3 G. Hern~mdez, 1 P. Gariglio, 2'3 G.E. F o x , 4 and J. Or6 4
1 Departamento de Biologfa, Facultad de Ciencias-UNAM, Apartado Postal 70-407, Cd. Universitaria, M6xico 04511, D.F., M6xico 2 Departamento de Gen6tica y Biologia Molecular, CINVESTAV-IPN, Apartado Postal 14-740, M6xico 07000, D.F., M6xico 3 Departamento de Gen6tica y Patologfa Experimental, Instituto Nacional de Salud POblica, CISEI, Cuernavaca, Morelos, M6xico 4 Department of Biochemical & Biophysical Sciences, University of Houston, Houston, TX 77204-5500, USA
Summary. Reverse transcriptase (RT) was first discovered as an essential catalyst in the biological cycle of retroviruses. However, in the past years evidence has accumulated showing that RTs are in- volved in a surprisingly large number of RNA- mediated transpositional events that include both viral and nonviral genetic entities. Although it is probable that some RT-bearing genetic elements like the different types of AIDS viruses and the mammalian LINE family have arisen in recent geo- logical times, the possibility that reverse transcrip- tion first took place in the early Archean is sup- ported by (1) the hypothesis that RNA preceded DNA as cellular genetic material; (2) the existence of homologous regions of the subunit tau of the E. coli DNA polymerase III with the simian immuno- deficiency virus RT, the hepatitis B virus RT, and the 13' subunit of the E. coli RNA polymerase (McHenry et al. 1988); (3) the presence of several conserved motifs, including a 14-amino-acid seg- ment that consists of an Asp-Asp pair flanked by hydrophobic amino acids, which are found in all RTs and in most cellular and viral RNA poly- merases. However, whether extant RTs descend from the primitive polymerase involved in the
Abbreviations: RT, reverse transcriptase; MMLV, Moloney rou- tine leukemia virus; MMTV, mouse mammary tumor virus; HIV, human immunodeficiency virus; SNV, spleen necrosis vi- rus; ASLV, avian sarcoma leukosis virus; RSV, Rous sarcoma virus; NTPs, ribonucleotides; dNTPs, deoxyribonucleotides Offprint requests to: A. Lazcano
RNA-to-DNA transition remains unproven. Sub- strate specificity of the AMV and HIV-1 RTs can be modified in the presence of Mn 2÷ , a cation which allows them to add ribonucleotides to an oligo (dG) primer in a template-dependent reaction. This change in specificity is comparable to that observed under similar conditions in other nucleic acid poly- merases. This experimentally induced change in RT substrate specificity may explain previous observa- tions on the misincorporation of ribonucleotides by the Maloney murine sarcoma virus RT in the minus and plus DNA of this retrovirus (Chen and Temin 1980). Our results also suggest that HIV-infected macrophages and T-cell cells may contain mixed polynucleotides containing both ribo- and deoxyri- bonucleotides. The evolutionary significance of these changes in substrate specificities of nucleic acid polymerases is also discussed.
Key words: Reverse transcriptase - - RNA ge- nomes - - Reverse transcriptase substrate ambigu- ities - - Early biological evolution
Introduction
Reverse transcriptase (RT) is an RNA-dependent DNA polymerase (Fig. 1) first discovered in exog- enous retroviruses (Baltimore 1970; Temin and Mi- zutani 1970), and later found to be encoded by a wide variety of genetic elements that are known as retroids (Fuetterer and Hohn 1987). It is known that

A Mg
tRNA primer ~ M g
H .Zn"] / \ , ~ ~.m,,,=lP '~ - DNA strond \ j " - ~ ~ PPi
#T~-ff [ I I [ 0 dNTP_.Mg
[IL LELIIJIIII, , retrovirol RNA
B M Z n ~ M n n Nn Oligo(dG) H primer poly(G) I" \ . PPi
5' l l I I I I I I 1 O" "~2Mn G_p- t /
3,,L ! L L LtttttL tl , poly (C)
Fig. 1. A Reverse transcriptase activity in vivo, B RT using a ribonucleotide substrate under in vitro conditions as described in the text (RT drawing modified after Mildvan and Loeb 1979).
RTs are directly involved in the replication of a wide spectrum of genetic elements that includes both viral and nonviral retrotransposons (Weiner et al. 1986).
It is likely that some retroids, such as the mam- malian LINE family (Weiner et al. 1986) and the AIDS viruses (Li et al. 1988; Smith et al. 1988), have arisen in recent geological times. Neverthe- less, the hypothesis that RNA was used as an in- format iona l mac romolecu le prior to double- stranded DNA in early Archean cells (Lazcano et al. 1988a) implies an early emergence of reverse transcription (Lazcano et al. 1988b) and led to the prediction of prokaryotic RTs (Lazcano et al. 1989). This suggestion was shown to be correct with the discovery of eubacterial RTs (Inouye et al. 1989; Lampson et al. 1989a,b). Analysis of codon usage in the gene coding for the RT found in Escherichia coli B suggests that this protein may be a recent evolu- tionary novelty in this particular bacterial strain (Lampson et al. 1989a; Inouye et al. 1989; Temin 1989), probably acquired by conjugation or trans- duction (Varmus 1989). However, this is not the case for the RT found in Myxococcus xanthus (Lampson et al. 1989a), where codon usage analysis suggests that the gene has been part of the myxo- bacterial genophore for a considerable period of time (Inouye et al., 1989). The existence of prokary-
525
otic RTs is also consistent with the protovirus hy- pothesis, i.e., the idea that reverse transcriptase is more ancient than retroviruses (Temin 1970, 1971), and is in excellent agreement with the phylogenetic trees derived by Doolittle et al. (1989) from RT amino acid sequences that clearly indicate that ret- roviruses are restricted to the Animal Kingdom.
The evidence suggesting that early cellular ge- nomes were RNA molecules (Lazcano et al. 1988b) implies that some type of reverse transcription ac- tivity must be almost as ancient as life itself, i.e., at least 3.5 x 10 9 years old (Oro et al. 1990). The ex- istence of a primordial proteinic RT has been sug- gested independently by several authors (Darnell 1985; Darnell and Doolittle 1986; Varmus 1987; Weiner 1987; Wintersberger and Wintersberger 1987; Lazcano et al. 1988a). Whether or not con- temporary RTs were descended from that hypothet- ical early enzyme is still an open question. The pur- pose of this paper is to discuss in detail the possibility that among extant RTs vestiges of an early Archean polymerase involved in nucleic acid replication can still be found. Therefore, this paper has been divided into two parts. First, the theoret- ical arguments that support the existence of early Archean reverse transcription are reviewed. Sec- ond, experimental evidence is presented on the sub- strate ambiguities of RTs, and the evolutionary sig- nificance of these results is discussed.
RNA Prior to DNA
The hypothesis that RNA preceded both DNA and proteins in primordial biological systems (Woese 1967; Crick 1968; Orgel 1968) has received consid- erable support from the discovery of RNA mole- cules with catalytic properties, which has implica- tions for early biological evolution (Pace and Marsh 1985; Cech 1989; Joyce 1989). Experimental and theoretical evidence suggests that RNA cellular ge- nomes were preceded during early biological evo- lution by polymers made of flexible, acyclic, prob- ably prochiral nucleotide analogues of prebiotic origin (Joyce et al. 1987). Detailed models describ- ing the evolutionary transition from primordial bio- logical systems based on catalytic and replicative RNA molecules into cells with DNA, RNA, and proteins have been discussed elsewhere (Alberts 1986; Gilbert 1986; Lazcano 1986; Lazcano et al. 1992).
Evidence indicating that RNA preceded DNA as an informational macromolecule in early Archean ceils has been reviewed recently (Lazcano et al. 1988a, 1992) and includes the following facts: (1) protein biosynthesis can take place in the absence of DNA but not of RNA; (2) the existence of viroids

526
and many different types of RNA viruses clearly indicates that replication systems can use either sin- gle- or double-stranded RNA molecules to store ge- netic information; (3) RNA molecules have been shown to exhibit catalytic properties; (4) ribonucle- otide cofactors containing purines, pyrimidines, and pyridines are ubiquitous and essential to pri- mary metabolism in all extant life forms; (5) the 2-OH' group of ribose, which is absent in deoxyri- bose, is involved in a number of RNA-catalyzed reactions, including phosphorylation and amino acid condensation reactions of prebiotic signifi- cance; (6) the biosynthesis of deoxyribonucleotides always proceeds via the enzymatic reduction of ri- bonucleotides; (7) RNA genomes lack repair mech- anisms, such as those involved in UV-light damage repair; (8) the 2'-deoxy-containing phosphodiester bonds of the DNA backbone are more stable to ba- sic hydrolysis; (9) DNA is more resistant than RNA to chemical degradation, including photochemical changes induced by UV irradiation, a form of en- ergy that was more abundant in early Archean en- vironments.
According to the "RNA prior to DNA hypothe- sis" (Lazcano et al. 1988a, 1992), the monophyletic origin and evolution of the biosynthetic pathways leading to the reduction of ribose to deoxyribose, the appearance of thymine, and the presence of DNA polymerases with proofreading activity evolved in the ancestors of contemporary cell lines in which the use of double-stranded DNA as an informational macromolecule enhanced the stability of genetic information and allowed the further de- velopment of repair mechanisms (Lazcano 1986; Lazcano et al. 1988a).
However, the DNA genome of pararetroviruses suggests that evolutionary transitions from RNA to DNA genomes have happened more than once in the history of the biosphere. Pararetroviruses lack an integrated replicative form and include both the caulimoviruses, which infect plants (Matthews 1981), and the hepadnaviruses (Temin 1985). Hep- adnavirus are infectious retroids that have been identified as the etiological agents of hepatitis B in man, rodents, and birds (Mandart et al. 1984), and replicate via RNA intermediates that are tran- scribed into DNA (Kaplan et al. 1973; Summers et al. 1975; Summers and Mason 1982). A similar rep- licative pattern is found in caulimoviruses (Hull and Covey 1983; Marco and Howell 1984).
Comparisons of available DNA-encoded RT pri- mary sequences of several hepadnaviruses (Ono et al. 1983; Mandart et al. 1984; Kadama et al. 1985; Miller and Robinson 1986) and caulimoviruses (Hull et al. 1986) have shown that they are homologous with retroviral RTs (Doolittle et al. 1989). This lends strong support to the hypothesis that animal
pararetroviruses descend from typical retroviruses (Temin 1985; Miller and Robinson 1986), i.e., that in the ancestors of hepadnaviruses a change from the use of RNA as packaged genome to DNA took place. As noted by Doolittle et al. (1989), genuine retroviruses must have emerged long after the ori- gin of vertebrates, and caulimoviral RTs are closely related to those coded by Drosophila melanogaster mobile genetic elements gypsy, 412, 297, and 17.6. Therefore, it may be concluded that at least two more independent evolutionary transitions from RNA to DNA genomes took place several billions of years after the appearance of double-stranded DNA in Archean cells:
Molecular Evolution of Reverse Transcriptases
The suggestion that prior to the evolutionary devel- opment of DNA cellular genomes were made of RNA implies that reverse transcription events took place in the early Archean history of the biosphere (Darnell 1985; Darnell and Doolittle 1986; Varmus 1987; Weiner 1987; Wintersberger and Winters- berger 1987; Lazcano et al. 1988a). The functional similarities between template-directed enzymatic synthesis of both DNA and RNA catalyzed by nu- cleic acid polymerases led to the hypothesis that their related functions evolved from a single gene originally coding for an ancestral small monomeric RNA-dependent RNA polymerase gene, which may be represented today by the [~' subunit of the eu- bacterial DNA-dependent RNA polymerase and its homologues (Lazcano et al. 1988b).
This hypothesis implies the existence of amino acid sequence similarities and/or tertiary structure resemblances (Zuckerkandl and Pauling 1965) among the catalytic domains of RTs and the eubac- terial RNA polymerase [3' subunit and its homo- logues. This possibility is in fact supported by the results of McHenry et al. (1988), who have shown the existence of conserved domains in the primary structure of RTs, a E. coli DNA polymerase Ill subunit, and the DNA-dependent RNA polymerase [3' subunit from the same bacterium. McHenry et al. (1988) have looked for homologues of the E. coli dnazx protein products (which correspond to the ~/ and the "r subunits of the DNA polymerase III), us- ing the Intelligenetics IFind and Align programs, with a 20-amino-acid window and a gap penalty of 2. The Align program allows the alignment of two proteins in regions of maximum similarity, and by using it McHenry and his co-workers were able to show the existence of homologous regions shared by the "r subunit of the E. coli DNA polymerase III with (1) the [3' subunit of the E. coli RNA polymer- ase; (2) the hepatitis B virus DNA polymerase; and

(3) the s imian i m m u n o d e f i c i e n c y viral RT (McHenry et al. 1988).
The monophyletic origin of extant RTs is indi- cated by the statistically significant amino acid se- quence similarities of several domains that are present in all of them (Johnson et al. 1986; Xiong and Eickbush 1988; Doolittle et al. 1989). The pol region exhibits some of the lowest mutational rates in the retroviral genomes (Coffin 1986; Li et al. 1988; McClure et al. 1988; Smith et al. 1988). Com- parison of available RT sequences with RNA- dependent RNA polymerases encoded by plus-, mi- nus-, and double-stranded RNA viruses has shown the existence of four consensus sequences, which may be part of an ancestral polymerase module (Poch et al. 1989). Further comparison of these polymerases with viral and cellular DNA poly- merases has allowed the identification of three con- served domains that apparently interact with nucle- otides, supporting the hypothesis that the different families of polymerases diverged from a common ancestor (Delarue et al. 1990; Blanco et al. 1991) that may have emerged in cells with RNA genomes (Lazcano et al. 1992). This possibility is supported by the DNA-dependent DNA polymerase encoded by the LaBelle mitochondrial plasmid of Neurospo- ra intermedia (Schulte and Lambowitz 1991), which exhibits some sequence similarity to RTs (Pande et al. 1989).
The most highly conserved region of all extant RTs is found toward the amino-terminal one-third part of the proteins (Varmus and Brown 1989), where a 14-amino-acid-residue segment consisting of an Asp-Asp pair flanked by hydrophobic amino acids is located (Fig. 2). This same type of domain centered around the Asp-Asp pair (also referred to as the DD domain) is found in almost all viral and cellular RNA polymerases (Kamer and Argos 1984; Argos 1988; Lazcano et al. 1988a; Velasco et al. 1992). A comparable segment is located in many viral and cellular DNA-dependent DNA poly- merases, where the hydrophobic region is centered around an Asp-Thr-Asp-Ser sequence (Argos 1988; Wong et al. 1988; Wang et al. 1989; Bernard et al. 1990). We interpret the similarity between these two highly conserved domains as the result of ad- aptation of the ancestral polymerase to different templates and substrates during the early Archean transition from RNA to DNA cellular genomes (Lazcano et al. 1998b).
Mutational studies on the domain structure of the Moloney murine leukemia virus (MMLV) RT have shown that the active site involved in reverse tran- scription is located toward the amino-terminal part of the enzyme (Tanese and Goff 1988), where the Asp-Asp domain is found. This conclusion is sup- ported by (1) the comparison of amino acid se-
527
Asp Asp
! I Met Leu !
Fig. 2. The HIV-1 reverse transcriptase DD domain (modified after Argos 1988).
quences of several retroviral and nonretroviral RTs, which suggest that the DNA polymerase catalytic domain is located within the first 300 amino acid residues of the RT-amino terminus (Gerard et al. 1986; Johnson et al. 1986); (2) experimental deter- mination of the MMLV RT substrate-binding site, which has shown that one lysine directly involved in dNTP binding is located in this same region (Basu et al. 1988); and (3) evidence that a region consisting of 37 amino acids located in the carboxy-end of the avian sarcoma-leukosis virus (ASLV) RT is not re- quired for viral replication (Katz and Skalka 1988); i.e., the catalytic site is not located in this part of the protein.
Experimental determinations and theoretical analyses of the mutational rates of genetic informa- tion encoded by RNA viruses have shown that RNA genomes change 10 6 times more rapidly than DNA genomes (Holland et al. 1982; Reanney 1982; Domingo et al. 1985; Steinhauer and Holland 1987). Although the mutational rate of different RTs is known to be half the value of that of other parts of the retroviral genome (Coffin 1986; Li et al. 1988; McClure et al. 1988; Smith et al. 1988), it is still one million times higher than the equivalent value for cellular genomes. Given this high number of muta- tions, the evolutionary conservation of the DD do- main implies that mutants are strongly selected against. However, the Asp-Asp domain is so short that the possibility that its almost universal distri-

528
bution among polymerases is due to convergent evolution cannot be dismissed altogether.
As shown in Fig. 2, theoretical calculations sug- gest that both the DD and the DTDS domains adopt a [3-hairpin configuration (Argos 1988). Since the carboxylic side chain of aspartic acid (1) can inter- act with Mg 2+ (Argos 1988), Zn z +, and other cat- ions and (2) can form hydrogen bonds with nucleic acid bases (Helene and Maurizot 1981), the highly conserved Asp-Asp domain may be a binding site, a catalytic region, or a regulatory structure. Although a detailed genetic analysis of the conserved regions of RTs is still lacking (Varmus and Brown 1989), there are several additional indications of the func- tional role of the Asp-Asp domain in RT, RNA rep- licases, and prokaryotic DNA-dependent RNA polymerases (Velasco et al. 1992). These include the following:
1. Enzymat ic activit ies of the E. coli DNA- dependent RNA polymerase and the avian my- eloblastosis RT were strongly inhibited by anti- bodies raised against a small protein (PGll) of the phage T7 tail appendage, containing the se- quence Leu-Ile-Val-Tyr-Ser-Asp-Asp-Tyr-Leu- Ser-Leu-Met (Zavriev and Borisova 1987).
2. Polymerizing activity of the Q[3 replicase was lost in vivo by mutant coliphages in which the Gly adjacent to the D-D pair wag changed to Ala, Ser, Pro, Met, or Val (Inokuchi and Hirashima 1987).
3. Site-especific mutagenesis studies of the HIV-1 RT expressed in E. coli have shown that the sub- stitution by histidine of one of the aspartic acids in the D-D pair led to a complete inhibition of the enzymatic activity of the polymerase (Larden et al. 1987).
4. Removal of the conserved Tyr of the HIV-1 RT D-D domain and its substitution by a small se- quence consisting of five amino acid residues led to a complete inhibition of its polymerizing ac- tivity (Hizi et al. 1988).
The Substrate Ambiguities of Reverse Transcriptases
RTs are RNA-dependent DNA polymerases that form a minus single-stranded DNA molecule ( - D N A ) complementary to the positive RNA ret- roviral genome and then read the - D N A molecule and behave as DNA-dependent DNA polymerases (cf. Varmus and Brown 1989). Retroviral RTs are zinc metalloenzymes (Poiesz et al. 1974; Auld et al. 1975) that also require other divalent metallic cat- ions. The Rous sarcoma virus (RSV), the mouse
mammary tumor virus (MMTV), and the human im- munodeficiency virus (HIV) RTs u s e M g 2+ , al- though the spleen necrosis virus (SNV) requires Mn z+ (Varmus and Brown 1989). DNA synthesis catalyzed by MMLV RT in the template-primer transcription of poly(A)-oligo(dT) is enhanced by a factor of four in the presence of Mn 2+ as compared with the effect of Mg z + (Verma 1977). The catalytic role of Mn z+ in oligomerization reactions of nucle- otide analogues of possible prebiotic significance has also been demonstrated (Visscher and Schwartz 1989). Although several retroviral RTs are Mn 2+ dependent, the cationic requirements of other RTs, like those encoded by the RSV, MMTV, and HIV, are comparable to those exhibited by most RNA and DNA polymerases, since without metallic cat- ions the syntheses of RNA and DNA cannot take place (Mildvan and Loeb, 1979, 1981).
Substitution of M g 2+ for Mn 2 + is known to in- crease the misincorporation of dNTPs by DNA- dependent DNA polymerases (Loeb and Mildvan 1981) and to alter their substrate specificities. The same effect has been reported for DNA-dependent RNA polymerases (Table 1). Since these changes in substrate specificities have been interpreted as a vestigial property of the times when cellular ge- nomes were made of RNA (Llaca et al. 1987), we have studied the role of Mn 2 + in the polymerizing properties of the avian myeloma virus (AMV) RT. The results we report here suggest that the speci- ficity of the AMV RT can be modified in vitro by Mn 2+ , allowing it to polymerase ribonucleotides.
Materials and Methods
Materials. AMV-RT, Tris-HCl, KCI, MnC12, disodium hydrogen phosphate 12-hydrate, tetrasodium pyrophosphate 10-hydrate, sodium dodecyl sulphate, ethylendiamintetracetate (Na2EDTA), toluene, ethanol, and unlabeled dGTP and GTP were acquired from Sigma Chem. Co.; dithiothreitol, from Calbiochem; (c~-32p)dGTP and (c~-32P)GTP were purchased from Amersham; poly(rC) and oligo(dG), from Pharmacia. Whatman DE-81 filters were used; 2,5-diphenyloxazol (PPO) and 1,4-bis(5-phenyl-2,- oxazolyl)bencene (POPOP) were acquired from New England Nuclear Co.
Solutions. The DE-81 filters were washed with a solution consisting of 0.8% sodium pyrophosphate, 10% disodium hydro- gen phosphate, and 0.1% sodium dodecyl sulphate prepared in bidistiUed water washing solution. Scintillating solution: PPO 0.5% POPOP 0.3% dissolved in toluene.
Activity Assays. Reverse transcription activity assays were made using a final volume of 50 I~1 of a reaction mixture consist- ing of 50 mM Tris-HC1 (pH 8.3), 1 mM DTT, 40 mM KCI, 10 ixg/ml poly(rC), 5 ixg/ml oligo(dG), 5 IxM dGTP, and 1 ixCi of (ct-32P)dGTP. The same mixture was used to study the AMV-RT

Table 1. Alternate functions of nucleic acid polymerases
529
Enzyme Normal activity Alternate role References
E. coli DNA pol 1
E. coli RNA pol
Phage T7 RNA pol
E. coli DNA primase
"R-DNA-polymerase" from healthy Xenopus, chicken embryos, rat liver and spleen cells, murine cells, and human lymphocytes
"R-DNA polymerase" from healthy HeLa and human embryonic lung cells
E. coli polynucleotide phosphorylase
DNA-dependent DNA polymerase
DNA-dependent RNA polymerase
DNA-dependent RNA polymerase
DNA-dependent RNA polymerase
?
Nontemplate-directed polymerization of NDPs
Incorporates NTPs in nascent DNA chains (with Mn 2+)
Reverse transcriptase
RNA-dependent RNA polymerase (with Mn z+)
RNA-dependent RNA polymerase
Incorporates NTPs producing mixed primers
DNA-dependent DNA polymerase (with Mg z+)
Reverse transcriptase (with Mn 2+)
Reverse transcriptase (with Mn z+)
Nontemplate-directed polymerization of dNDPs (with Mn 2 ÷)
Berg et al. 1963
Karkas et al. 1972; Karkas 1973; Loeb et al. 1973
Llaca et al. 1987
Konarska & Sharp 1989
Kornberg 1977; Rowen and Kornberg 1978
Bolden et al. 1972; Spadari and Weissbach 1974
Fridlender et al. 1972
Hsleh 1971; Gillam et al. 1978
changes in specificity, but dGTP was substituted by 5 ~xM GTP, and 1 vCi of (a-3Zp)GTP was used instead of (a-3zp)dGTP. MnC12 and MgCI2 concentrations were varied as shown in Fig. 3. AMV-RT was added to these mixtures and incubated at 37°C for 10 min. Reactions were stopped with 4 izl of EDTA 0.5 M, and the final mixture was then added to DE-81 filters. These filters were washed at 5-min intervals, eight times with washing solu- tion, then once with bidistillated water and twice with ethanol. Dry filters were treated with scintillation liquid and counted in a Miniaxi Beta 260 Tricarb 4000 (United Technological Packard) liquid scintillation counter. Incorporation of dGTP or GTP was estimated from the counts per minute (cpm). According to our estimates, 8,880 cpm are equivalent to 1 pmol of incorporated nucleotides.
ResuRs
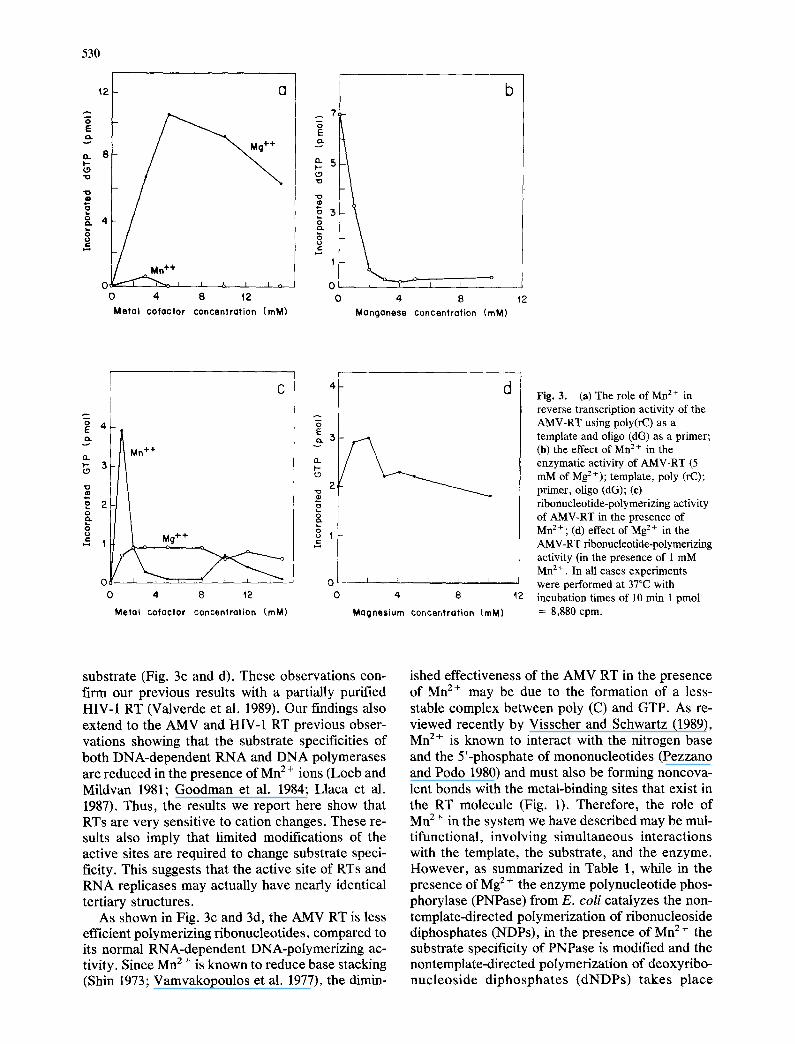
Results are shown graphically in Fig. 3. Figure 3a shows that AMV-RT dNTP-polymerizing activity is strongly Mg 2 +-dependent, reaching a peak of ac- tivity at 5 raM. Minor dGTP-polymerizing activity was observed at 3 mM Mn 2 +. In Fig. 3b the inhibi- tion of reverse transcriptase activity by Mn z+ (at 5 mM M g 2+) is shown. The polymerization of the ri- bonucleotide substrate induced by Mn z+ under varying cation concentrations is shown in Fig. 3c and d.
Discussion
It is well known that RTs are enzymatically active on different RNA and DNA templates provided a complementary oligo(dNTP) is present to act as a primer (Varmus and Swanstrom 1982). An assay for
measuring in vitro reverse transcription activity has been deve loped using the complex poly(C)- oligo(dG) as a template-primer (Gerard et al. 1976; Kiessling and Goulian 1976) that is specific for sev- eral RTs but does not work for other polymerases. Since prokaryotic and eukaryotic DNA-dependent DNA polymerases are unable to transcribe the poly(C)-oligo(dG) complex (Gerard et al. 1976), we have used this system to study the effect of Mn z+ on the AMV RT substrate specificity.
Some retroviral RTs are Mn2+-dependent, and there are also some reports that mammalian type C retroviral RT activity is enhanced in the presence of Mn 2+ (Sarngadharan et al. 1976). However, our re- sults clearly show that this is not the case either for HIV-1 (Valverde et al. 1989) or for the AMV RTs. As shown in Fig. 3b, under in vitro conditions Mn 2+ is a strong inhibitor of reverse transcription. This conclusion is in agreement with previous ob- servations in which reverse transcription was stud- ied in the presence of Mg 2+ and Mn 2+ (Marcus and Modak 1976; Vamvakopoulos et al. 1977). Our re- sults also help us to unders t and the Mn z+- dependent enhancement of the inhibitory effects of the antiviral drugs phosphonoformate (Sundquist and Oberg 1979) and pyr idoxal 5 ' -phospha te (Modak and Dumaswala 1981) over the catalytic ac- tivity of the AMV and the Rauscher leukemia virus RTs, respectively.
The results presented here suggest that the sub- stitution of Mg z+ by Mn z+ modifies the specifity of the AMV RT, allowing it to use a ribonucleotide
12 b
o E o .
a. 8 k -
"1o
"1o o
2 o 4
M n + +
0_..- • . ~ o i I ~) I I
0 4 8 t2
Metal cofactor concentration (raM) 0 4 8 12
Manganese concentration (mM)
530
C
4
3
2
I
O, 0
Mn ++
0 4 B 12 0
Metal cofactor concentration (raM)
d
I I r I I
4 8
Magnesium Concentration (raM)
~Z
Fig. 3. (at The role of Mn z + in reverse transcription activity of the AMV-RT using poly(rC) as a template and oligo (dG) as a primer; (b) the effect of Mn 2 ÷ in the enzymatic activity of AMV-RT (5 mM of MgZ÷); template, poly (rC); primer, oligo (dG); (c) ribonucleotide-polymerizing activity of AMV-RT in the presence of Mn z + ; (d) effect of Mg 2 + in the AMV-RT ribonucleotide-polymerizing activity (in the presence of 1 mM Mn 2+ . In all cases experiments were performed at 37°C with incubation times of 10 rain 1 pmol = 8,880 cpm.
substrate (Fig. 3c and d). These observations con- firm our previous results with a partially purified HIV-I RT (Valverde et al. 1989). Our findings also extend to the AMV and HIV-1 RT previous obser- vations showing that the substrate specificities of both DNA-dependent RNA and DNA polymerases are reduced in the presence of Mn 2 ÷ ions (Loeb and Mildvan 1981; Goodman et al. 1984; Llaca et al. 1987). Thus, the results we report here show that RTs are very sensitive to cation changes. These re- suits also imply that limited modifications of the active sites are required to change substrate speci- ficity. This suggests that the active site of RTs and RNA replicases may actually have nearly identical tertiary structures.
As shown in Fig. 3c and 3d, the AMV RT is less efficient polymerizing ribonucleotides, compared to its normal RNA-dependent DNA-polymerizing ac- tivity. Since Mn 2 + is known to reduce base stacking (Shin 1973; Vamvakopoulos et al. 1977), the dimin-
ished effectiveness of the AMV RT in the presence of Mn 2+ may be due to the formation of a less- stable complex between poly (C) and GTP. As re- viewed recently by Visscher and Schwartz (1989), Mn z+ is known to interact with the nitrogen base and the 5'-phosphate of mononucleotides (Pezzano and Podo 1980) and must also be forming noncova- lent bonds with the metal-binding sites that exist in the RT molecule (Fig. 1). Therefore, the role of Mn z+ in the system we have described may be mul- tifunctional, involving simultaneous interactions with the template, the substrate, and the enzyme. However, as summarized in Table 1, while in the presence of Mg z+ the enzyme polynucleotide phos- phorylase (PNPase) from E. coli catalyzes the non- template-directed polymerization of ribonucleoside diphosphates (NDPs), in the presence of Mn 2 + the substrate specificity of PNPase is modified and the nontemplate-directed polymerization of deoxyribo- nucleoside diphosphates (dNDPs) takes place

Table 2. Substrate ambiguities of reverse transcriptases
531
Enzyme Abnormal product References
MMLV-RT ASV-RT
MMTV-RT
HIVI-RT
AMV-RT
Minus-strand RNA molecules complementary to the MMLV genome (in vivo) Small amounts of minus-strand RNA molecules complementary to the RSV
genome (in vivo) Small amounts of minus-strand RNA molecules complementary to the MMTV
genome (in vivo) Poly(rG) complementary to a poly(rC) template (in vitro, in the presence of
Mn 2+) Poly(rG) complementary to a poly(rC) template (in vitro, in the presence of
Mn 2+)
Biswal and Benyesh-Melnick 1969 Stavnezer et al. 1976
Stavnezer et al. 1976
Valverde et al. 1989
This paper
(Hsieh 1971; Gillam et al. 1978). Thus, the effect of Mn z+ in the reaction we have studied could be re- stricted solely to the substrates and the poly- merases.
As noted by Jensen (1976), erroneous reactions of highly specific enzymes are maximized by con- ditions involving either (1) high levels of enzyme or (2) abnormal substrates or (3) low concentrations of normal substrate. To these factors we can clearly add the presence of cations like Mn 2+ that are ca- pable of altering the substrate specificity of all ma- jor types of nucleic acid polymerases (Tables 1 and 2). Thus, it is likely that previous reports summa- rized in Table 1 of the so-called eukaryotic "R- d e p e n d e n t D N A p o l y m e r a s e s " , i .e . , RNA- dependent DNA polymerases (Bolden et al. 1972; Fridlender et al. 1972; Spadari and Weissbach 1974) are in effect artifacts, since in all these cases re- verse transcription was observed in the presence of Mn 2+, and at least in two cases (Bolden et al. 1972; Spadari and Weissbach 1974) the enzymes behaved as DNA-dependent DNA polymerases in the pres- ence of Mg 2+.
Data summarized in Tables 1 and 2 also suggest that the incorporation of ribonucleotides in a DNA chain by polymerases may depend on the amount of available nucleotides. A local high concentration of ribonucleotides in the replicative cellular microen- vironment will promote their incorporation in the nascent DNA strand by otherwise deoxyribonucle- otide-specific DNA polymerases. There have been previous reports of r ibonucleotides covalently linked to DNA from mitochondria (Grossman et al. 1973), coliphages T4 (Speyer et al. 1972) and T5 (Rosenkranz 1973), and bovine rhinotracheitis virus (Babiuk and Rouse 1976). It is possible that their presence is due to defective DNA polymerases or localized high concentrations of ribonucleotides.
The relatively low substrate specificity of the AMV and the HIV-1 RTs reported here suggests an additional source of retroviral biological diversity and may also help explain the presence of misincor- porated ribonucleotides in the minus and the plus strands of SNV DNA reported by Chen and Temin
(1980). Since the change in substrate specificity of RTs appears to be at least in part due to minor conformational changes induced by Mn 2+, the ob- servations of Chen and Temin (1980) may be ex- plained in terms of RNA-directed RNA synthesis catalyzed by a defective, i.e., mutant, SNV-RT. This conclusion and the results summarized in Ta- ble 2 also suggest that similar mixed ribo- and deoxyribonucleic single-stranded polymers may be found in HIV-infected human T-cells and macro- phages. The hydrolytic lability of 3'---~5' bonds in which ribose is involved leads us to predict the presence not only of mixed polynucleotides con- taining both ribo- and deoxyribonucleotides in HIV- infected cells but also of HIV-1 + D N A fragments of random length with terminal ribonucleosides lo- cated preferentially at their 5' ends.
The above explanation may also account for the existence of small amounts of large intracellular - R N A molecules complementary to the genomes of the RSV, the MMTV (Stavnezer et al. 1976), and the Moloney murine sarcoma virus (Biswal and Benyesh-Melnick 1969; Knesek et al. 1980) found in infected cells (Table 2). It is also possible that in these cases the cellular dNTP biosynthetic enzyme complex was crippled by the retroviral infection, leading to localized high concentrations of ribonu- cleotides which are polymerized into a - R N A strand by the RSV, MMTV, and MMLV RTs.
The results presented here and those summa- rized in Tables 1 and 2 on the substrate ambiguities of RTs and other nucleic acid polymerases may also have evolut ionary implications. As discussed above, the existence of different DNA and RNA polymerases has been explained as the result of gene duplication and further diversification events of the gene coding for an ancestral replicase (Laz- cano et al. 1988b). As reviewed by Li (1983), sev- eral hypotheses have been proposed to account for the emergence of new genes from redundant dupli- cates. The step-wise model of the evolutionary de- velopment of metabolic pathways suggested by Wa- ley (1969) proposes that the evolution of a duplicate copy of a gene of an enzyme resulting in minimal

532
changes in conformation could lead to a new en- zyme capable of catalyzing a similar type of reac- tion. Moreover, as an alternative to Horowitz 's (1945, 1965) hypothesis of retrograde evolution of metabolic pathways, Jensen (1976) and Jensen and Byng (1982) suggested a model quite comparable to that of Waley (1969), arguing that primitive en- zymes were unregulated, substrate-ambiguous cat- alysts. Gene duplication and further divergence opened the possibility for specialization. Similar ideas on the broad substrate specificities of early enzymes and the appearance of new functions in proteins have been proposed independently by Zuckerkandl (1970, 1975), Cove (1974), Koshland (1976), and Fink (UC-Santa Cruz, personal commu- nication). These hypotheses imply that in the course of evolution primitive biological catalysts have become highly specific, regulated, and in many cases, oligomeric enzymes.
The results reported here and in a previous arti- cle (Llaca et al. 1987) support Waley's (1969) and Jensen's (1976) models of enzyme evolution. Al- though it is not known/how many mutational steps would be required to achieve the adaptation of an- cestral polymerases to different templates and sub- strates, the sensitivity of RTs and other poly- merases to cation changes (Tables 1 and 2) implies that very simple modifications of their substrate- binding and active sites are required for a change in catalytic properties. Thus, we can conclude that early polymerases had rather broad substrate and template specificities, and that the evolutionary transition from a primitive RNA-dependent RNA polymerase into the extant DNA-dependent en- zymes via ancestral reverse transcription activity was mediated by low-specificity nucleic acid poly- merases in which novel catalytic properties were acquired by few conformational modifications.
Finally, it should be pointed out that the findings summarized in Table 2 are consistent with the no- tion that the transition from cellular RNA genome to DNA genomes (Lazcano et al. 1988b) need not have been a sharp one. It is conceivable that a pe- riod of cellular evolution existed in which genetic information may have been carried by mixed poly- mers. If so, the final transition to double-stranded DNA was probably rapid, due to the enhanced chemical stability of DNA, which makes it more suitable for information storage.
Conclusions
Although there are several indications that some retroids, like the different AIDS viruses, are recent biological novelties (Li et al. 1988; Smith et al. 1988; Doolittle 1989), it is proposed here that reverse
transcription itself is a very ancient process that was mediated in early Archean times by poly- merases with broad template and substrate specific- ities. The evidence that is discussed in this paper can be summarized as follows:
1. The hypothesis that during the early Archean the cellular genomes were made of RNA molecules prior to DNA implies an early existence of re- verse transcription activity (Lazcano et al. 1988b).
2. All retroviral RTs share significant amino acid sequence similarities with each other and with the RTs encoded by other retroids and by prokaryotes, suggesting a monophyletic origin for all RTs. Furthermore, the comparison of pri- mary structures of RTs, RNA-dependent RNA polymerases, and cellular and viral DNA poly- merases has led to the identification of several conserved domains (Poch et al. 1989; Delarue et al. 1990), supporting the hypothesis of a common origin of all extent template-dependent poly- merases (Lazcano et al. 1988a, 1991). Whether or not the common ancestor of contemporary RTs was an early Archean polymerase involved in the RNA-to-DNA transition is still an open question. One common feature shared by all RTs and most cellular and viral RNA polymerases is the DD domain, which may be involved in cata- lytic and/or regulatory functions, or may repre- sent an essential structural feature common to all these enzymes.
3. The results presented here suggest that previous observations that had shown that the presence of Mn z + can alter the substrate specificities of both DNA-dependent DNA and RNA polymerases can be extended to the RTs of AMV and HIV-1 (Table 2). These results indirectly confirm the existence of structural similarities among RTs and RNA and DNA polymerases and suggest that few mutations were required to adapt the polymerases during the evolutionary transition from RNA to DNA cellular genomes in early Archean times. In addition, these results support the models of enzyme evolution developed by Waley (1969), Jensen (1976), and Jensen and Byng (1982) that have proposed that ancestral enzymes had broad substrate specificities.
4. Most RTs are small monomeric polymerases of approximately 70 kd (Varmus and Swanstrom 1982) that are relatively inefficient catalysts with a rate of elongation 10- to 1,000-fold slower than most DNA polymerases (Edenberg and Huber- man 1975). Since RTs can read both DNA and RNA templates, lack proofreading activity (Preston et al. 1988; Roberts et al. 1988), and can polymerize both dNTPs and NTPs in vivo and

533
u n d e r s l ightly modi f ied l a b o r a t o r y cond i t i ons ,
they cons t i tu t e an exce l len t expe r imen ta l mode l
o f the gene t ica l ly e n c o d e d pr imi t ive po lyme r a se s wh ich may have b e e n e v o l u t i o n a r y in te rmedi -
ates b e t w e e n the small ca ta ly t ic pept ides (Oro and L a z c a n o - A r a u j o 1984; Shen et al. 1989) and
r i b o z y m e s (Sharp 1985; Pace and Marsh 1985;
Cech 1989) that m a y have p receded the complex mul t iun i t D N A p o l y m e r a s e s i nvo lved in c on t e m-
pora ry ce l lu lar r ep roduc t ion .
Acknowledgments. We are especially grateful to Drs. Jaime Septilveda Amor and Mauricio Hern~indez of the Mexican Min- istry of Health for their constant support and encouragement. We are indebted to Drs. Howard M. Temin and Carlos Larralde for their critical comments, and to Dr. S.F.J. Le Grice (Hoffman- La Roche and Co.) for his kind gift of the plasmid pRT/CG with gene coding for the HIV-I reverse transcriptase. Miles. Elena Hilario and Aria Velasco and Monsieur Ervin Silva kindly pro- vided us with several useful references. We are indebted to Thomas Mills for his help in preparing the figures and Tables 1 and 2 and for his careful reading of the manuscript. This work has been supported in part by CONASIDA-M6xico and Proyecto UNAM.IN 105289 (A.L.), COSNET, Aaron Saenz Assoc., and Givaudan de Mrxico (P.G.), NASA grant NS6-7440 (G.E.F.), and NASA grant 44-005-002 (J.O.).
References
Alberts BM (1986) The function of the hereditary materials: bi- ological catalyses reflect the cell's evolutionary history. Am Zool 26:781-796
Argos P (1988) A sequence motif in many polymerases. Nucleic Acid Res 16:9909-9916
Auld DS, Kawagachi H, Livingston DM, Vallee BL (1975) Re- verse transcriptase from avian myeloblastosis virus: a zinc metalloenzyme. Biochem Biophys Res Comm 62:296--302
Babiuk LA, Rouse BT (1976) Ribonucleotides in infectious bo- vine rhinotracheitis virus DNA. J Gen Virol 31:221-230
Baltimore D (1970) RNA-dependent DNA polymerase in virions of RNA tumour viruses. Nature 226:1209-1211
Basu A, Nanduri VB, Gerard GF, Modak MJ (1988) Substrate binding domain of Murine leukemia virus reverse tran- scriptase. J Biol Chem 263:1648--1653
Berg P, Fancher H, Chamberlin M (1963) The synthesis of mixed polynucleotides containing ribo- and deoxyribonucleotides by purified preparations of DNA polymerase from Escheri- chia coli. In: Vogel H J, Bryson V, Lampen JO (eds) infor- mational macromolecules. Academic Press, New York, pp 467-483
Bernard A, Lazaro JM, Salas M, Blanco L (1990) The highly conserved amino acid sequence motif Tyr-Gly-Asp-Thr-Asp- Ser in alpha-like DNA polymerases is required by phage o 29 DNA polymerase for protein-primed initiation and polymer- ization. Proc Natl Acad Sci USA 87:4610--4614
Biswal N, Benyesh-Melnick M (1969) Complementary nuclear RNAs of murine sarcoma-leukemia virus complex in trans- formed cells. Proc Natl Acad Sci USA 64:1372-1379
Blanco L, Bernard A, Blasco MA, Salas M (1991) A general structure for DNA-dependent DNA polymerases. Gene 100: 27-38
Bolden A, Fry M, Muller R, Citarella R, Weissbach A (1972) The presence of a polyriboadenylic acid-dependent DNA poly- merase in eukaryotic cells. Arch Biochem Biophys 153:26-33
Cech T (1989) Ribozyme self-replication? Nature 339:507-508 Chen IS, Temin HE (1980) Ribonucleotides in unintegrated lin-
ear spleen necrosis virus DNA. J Virol 33:1058--1073 Coffin JM (1986) Genetic variation in AIDS viruses. Cell 46:1-4 Cove DJ (1974) Evolutionary significance of autogenous regula-
tion. Nature 251:256 Crick FHC (1968) The origin of the genetic code. J Mol Biol
38:367-379 Darnell JE (1985) RNA. Sci Am 253:54-64 Daruell JE, Doolittle WF (1986) Speculations on the early course
of evolution. Proc Natl Sci USA 83:1271-1275 Delarue M, Poch M, Tordo N, Moras D, Argos P (1990) An
attempt to unify the structure of polymerases. Protein Engi- neering 3:461-467
Domingo E, Martinez-Salas E, Sobrino F, de la Torre JC, Portela A, Ortin J, Lopez-Galindez C, Perez-Brena P, Vil- lanueva N, Najera R, Van de Pol S, Steinhauer D, DePolo N, Holland JJ (1985) The quasispecies (extremely heteroge- neous) nature of viral RNA genome populations: biological relevance--a review. Gene 40:1-8
Doolittle RF (1989) Immunodeficiency viruses: the simian- human connection. Nature 339:338-339
Doolittle RF, Feng DF, Johnson MS, McClure MA (1989) Ori- gins and evolutionary relationships of retroviruses. Quart Rev Biol 64:1-30
Edenberg H J, Huberman JA (1975) Eukaryotic chromosome rep- lication. Ann Rev Genet 9:245-284
Fridlender B, Fry M, Bolden M, Weissbach A (1972) A new synthetic RNA-dependent DNA polymerase from human tis- sue culture cells. Proc Natl Acad Sci USA 69:452-455
Fuetterer J, Hohn T (1987) Involvement of nucleocapsids in re- verse transcription: a general phenomenon? Trends Biochem Sci 12:92-95
Gerard GF, D'Alessio JM, Kotewicz ML, Noon MC (1986) In- fluence on stability in Escherichia coli of the carboxy- terminal structure of cloned Moloney murine leukemia virus reverse transcriptase. DNA 5:271-279
Gerard GF, Rottman F, Green M (1976) Poly (2'-0-methyl- cytidylate) oligodeoxyguanylate as a template for the ribonu- cleic acid directed deoxyribonucleic acid polymerase in ribo- nucleic acid tumor virus particles and a specific probe for the rihonucleic acid directed enzyme in transformed murine cell. Biochemistry 13:1632-1641
Gilbert W (1986) The RNA world. Nature 319:618 Gillam S, Jahnke P, Smith M (1978) Enzymatic synthesis of oli-
godeoxyribonucleotides of defined sequence. J Biol Chem 253:2532-2539
Goodman TC, Nagel L, Rappold W, Klotz G, Riesner D (1984) Viroid replication: equilibrium association constant and com- parative activity measurements for the viroid-polymerase in- teraction. Nucleic Acid Res 12:6231--6246
Grossman L1, Watson R, Vinograd J (1973) The presence of ribonucleotides in mature closed circular mitochondrial DNA. Proc Natl Acad Sci USA 70:3339-3343
Helene C, Maurizot JC (1981) Interactions of oligopeptides with nucleic acids. CRC Critical Rev Biochem 10:213-258
Hizi A, McGill C, Hughes SH (1988) Expression of soluble, en- zymatically active, human immunodeficiency virus reverse transcriptase in Escherichia coli and analysis of mutants. Proc Natl Acad Sci USA 85:1218-1222
Holland J, Spindler K, Horodyski F, Graban E, Nichols S, VandePol S (1982) Rapid evolution of RNA genomes. Sci- ence 215:1577-1585
Horowitz NH (1945) On the evolution of biochemical syntheses. Proc Natl Acad Sci USA 31:153-157
Horowitz NH (1965) The evolution of biochemical syntheses-- retrospect and prospect. In: Bryson V, Vogel HJ (eds) Evolv- ing genes and proteins. Academic Press, New York, pp 15-23

534
Hsieh WT (1971) Polymerization of deoxyribonucleoside diphos- phates from and Escherichia coli mutant lacking deoxyribo- nucleic acid polyrnerase activity. J Biol Chem 246:1780~1784
Hull R, Covey SN (1983) Does cauliflower mosaic virus replicate by reverse transcription? Trends Biochem Sci 8:119-121
Hull R, Sadler J, Longstaff M (1986) The sequence of carnation etched ring virus DNA: comparison with cauliflower mosaic virus and retroviruses. EMBO J 5:3083-3090
Inokuchi Y, Hirashima A (1987) Interference with viral infection by defective RNA replicase. J Virol 61:3946-3949
Inouye S, Hsu M-Y, Eagle S, Inouye M (1989) Reverse tran- scriptase associated with the biosynthesis of the branched RNA-linked ms DNA in Myxococcus xanthus. Cell 56:706- 717
Jensen RA (1976) Enzyme recruitment in evolution of new func- tion. Annu Rev Microbiol 30:409-425
Jensen RA, Byng GS (1982) The partioning of biochemical path- ways with isosymes systems. In: Rattazi MC, Scandalios JG, Whitt GS (eds) Isozymes: current topics in biological and medical research, vol 5. Alan R Liss, New York, pp 143-175
Johnson MS, McClure MA, Feng DF, Gray J, Doolittle RF (1986) Computer analysis of retroviral pol genes: assignment of enzymatic functions to specific sequences and homologies with nonviral enzymes. Proc Natl Acad Sci USA 83:7648- 7652
Joyce GF (1989) RNA evolution and the origins of life. Nature 338:217-224
Joyce GF, Schwartz AW, Miller SL, Orgel LE (1987) The case for an ancestral genetic system involving simple analogues of the nucleotides. Proc Natl Acad Sci USA 84:4398-4402
Kadama K, Ogasawara N, Yoshikawa H, Murakami S (1985) Nucleotide sequence of a cloned woodchuck hepatitis virus genome: evolutionary relationships between hepadnaviruses. J Virol 56:978-986
Kamer G, Argos P (1984) Primary structural comparisons of RNA-dependent polymerases from plant, animal and bacte- rial viruses. Nucleic Acid Res 12:7269-7282
Kaplan PN, Greeman RL, Geirn JL, Purcell RH, Robinson WS (1973) DNA polymerase associated with human hepatitis B antigen. J Virol 12:995-1005
Karkas JD (1973) Reverse transcription by Escherichia coli poly- merase I. Proc Natl Acad Sci USA 70:3834-3838
Karkas JD, Jannis GS, Chargaff E (1972) Action of DNA poly- merase I of Escherichi coli with DNA-RNA hybrids as tem- plates. Proc Natl Acad Sci USA 69:398-402
Katz RA, Skalka AM (1988) A C-terminal domain in the Avian Sarcoma-Leukosis virus pol gene product is not essential for viral replication. J Virol 62:528-533
Kiessling AA, Goulian M (1976) A comparison of the enzymatic responses of the DNA polymerases from four RNA tumor viruses. Biochem Biophys Res Comm 71:1069-1077
Knesek JE, Nash MA, Chan JC, Bartlett RJ, Bowen JM, East JL (1980) Intracellular RNA complementary to the RNA genome of the Moloney routine sarcoma virus complex. Virology 100: 288-299
Konarska MM, Sharp PA (1989) Replication of RNA by the DNA-dependent RNA polymerase of Phage T7. Cell 57:423- 431
Kornberg A (1977) Multiple stages in the enzymic replication of DNA. Biochem Soc Trans 5:359-374
Koshland DE (1976) The evolution of function in enzymes. Fed Proc 35:2104-2111
Lampson BC, Inouye M, Inouye S (1989a) Reverse transcriptase with concomitant ribonuclease H activity in the cell-free syn- thesis of branched RNA-linked msDNA of Myxococcus xan- thus. Cell 56:701-707
Lampson BC, Sun J, Hsu MY, Vallejo-Ramirez J, Inouye S, Inouye M (1989b) Reverse transcriptase in a clinical strain of
Escherichia coli: production of branched RNA-linked msDNA. Science 243:1033-1037
Larden BA, Purifoy DJM, Powell KL, Darby G (1987) Site spe- cific mutagenesis of AIDS virus reverse transcriptase. Nature 327:716--717
Lazcano A (1986) Prebiotic evolution and the organ of cells. Treballs Soc Cat Biol 39:73-103
Lazcano A, Fastag J, Gariglio P, Ramirez C, Oro J (1988a) On the early evolution of RNA polymerase. J Mol Evol 27:365- 376
Lazcano A, Guerrero R, Margulis L, Oro J (1988b) The evolu- tionary transition from RNA to DNA in early cells. J Mol Evol 27:283-290
Lazcano A, Gariglio P, Orozco E, Oro J (1989) On the early evolution of reverse transcriptase. Abstracts of the 6th Meet- ing of the International Society for the Study of the Origins of Life, Prague, Checoslovakia, July 3--8, 1989, 166-167
Lazcano A, Fox GE, Oro J (1992) Life before DNA: the origin and evolution of early Archean cells. In: Mortlock RP (ed) The evolution of metabolic function. CRC Press, Boca Ra- ton, pp. 237-295
Li WH (1983) Evolution of duplicate genes and pseudogenes. In: Nei M, Koehn RK (eds) Evolution of genes and proteins. Sinauer Press, Sunderland, MA, pp 14-37
Li WH, Tanimura M, Sharp PM (1988) Rates and dates of diver- gence between AIDS virus nucleotide sequences. Mol Biol Evol 5:313-330
Llaca V, Silva E, Lazcano A, Rangel LM, Gariglio P, Oro J (1987) In search of the ancestral RNA polymerase: an exper- imental approach. In: F Eirich, C Ponnamperuma (eds) Pre- biological organization: proceedings of the VIII College Park Colloquium on Chemical Evolution. A Deeker Publ, Hamp- ton, VA, pp 247-260
Loeb LA, Mildvan AS (1981) The role of metal ions in the fidelity of DNA and RNA synthesis. In: Eichhorn GL, Manzilli LG (eds) Metal ions in genetic information transfer. Elsevier/ North Holland, New York, pp 125-142
Loeb LA, Tartof KD, Travaglini EC (1973) Copying natural RNAs with E. coli DNA polymerase I. Nature 242:66-69
Mandart E, Kay A, Galibert F (1984) Nucleotide sequence of a cloned duck hepatitis B virus genome: comparison with woodchuck and human hepatitis B virus sequences. J Virol 49:782-792
Marco Y, Howell SH (1984) Intracellular forms of viral DNA consistent with a model of reverse transcriptional replication of the cauliflower mosaic virus genome. Nucleic Acid Res 12:1517-1528
Marcus SL, Modak MJ (1976) Observations on template specific conditions for DNA synthesis by avian myeloblastosis virus DNA polymerase. Nucleic Acid Res 3:1473-1486
Matthews REF (1981) Plant virology. Academic Press, New York
McHenry CS, Flower AM, Hawker JR (1988) Coordination of leading with lagging strand synthesis by the asymmetric dimeric DNA polymerase III holoenzyme of Escherichia coli. Cancer Cells 6:35-41
McClure MA, Johnson MS, Feng DF, Doolittle RF (1988) Se- quence comparisons of retroviral proteins: relative rates of change and general phylogeny. Proc Natl Acad Sci USA 85: 2469-2473
McMacken R, Koruberg A (1978) A multienzyme system for priming the replication of OX174 viral DNA. J Biol Chem 253:3313-3319
Mildvan AS, Loeb LA (1979) The role of metal ions in the mech- anisms of DNA and RNA polymerases. CRC Crit Rev Bio- chem 6:219-244
Mildvan AS, Loeb LA (1981) The role of metal ions in the mech- anism of DNA and RNA polymerases. In: Eichhorn GL,

Marzilli LG (eds) Metal ions in genetic information transfer. Elsevier/North Holland, New York, pp 103-123
Miller RH, Robinson WS (1986) Common evolutionary origin of hepatitis B virus and retroviruses. Proc Natl Acad Sci USA 83:2531-2535
Modak MJ, Dumaswala UJ (1981) Divalent cation-dependent py- ridoxal 5'-phosphate inhibition of Rauscher leukemia virus DNA polymerase. Biochem Biophys Acta 654:227-235
Ono Y, Onda H, Sasada R, Igarashi K, Sugino Y, Nishioka K (1983) The complete nucleotide sequences of the cloned hep- atitis B virus DNA: subtypes adr and adw. Nucleic Acid Res 11:1747-1757
Orgel LE (1968) Evolution of the genetic apparatus. J Mol Biol 38:380-393
Oro J, Lazcano-Araujo A (1984) A minimal living system and the origin of a protocell. Adv Space Res 4:167-176
Oro J, Miller SL, Lazcano A (1990) The origin and early evolu- tion of life on Earth. Annu Rev Earth Planet Sci 18:317-356
Pace NR, Marsh TL (1985) RNA catalysis and the origin of life. Origins Life 16:97-116
Pande S, Lemire EG, Nargang FE (1989) The mitochondrial plasmid from Neurospora intermedia strain LaBelle-lb con- tains a long open reading frame with blocks of amino acids characteristic of reverse transcriptases and related proteins. Nuclei Acid Res 17:2023-2042
Pezzano H, Podo F (1980) Structure of binary complexes of mono- and polynucleotides with metal ions of the first tran- sition group. Chem Rev 80:365-401
Poch O, Sauvaget I, Delarue M, Tordo N (1989) Identification of four conserved motifs among the RNA-dependent polymer- ase encoding elements. EMBO J 8:3867-3874
Poiesz BJ, Battula N, Loeb LA (1974) Zinc in reverse tran- scriptase. Biochem Biophys Res Comm 56:959-964
Preston BD, Poiesz BJ, Loeb LA (1988) Fidelity of HIV-1 re- verse transcriptase. Science 242:1168-1171
Reanney DC (1982) The evolution of RNA viruses. Annu Rev Microbiol 36:47-73
Roberts JD, Bebenek K, Kunkel TA (1988) The accuracy of reverse transcriptase from HIV-1. Science 242:1171-1173
Rosenkranz HS (1973) RNA in coliphage T5. Nature 242:327-329 Rowen L, Koruberg A (1978) A ribo-deoxyribonucleotide primer
synthesized by primase. J Biol Chem 253:770-774 Sarngadharan MG, Allaudeen HS, Gallo RC (1976) Reverse tran-
scriptase activity of RNA tumor viruses and animal cells. Methods Cancer Res 12:3-47
Schulte U, Lambowitz AM (1991) The LaBelle mitochondrial plasmid of Neurospora intermedia encodes a novel DNA polymerase that may be derived from a reverse transcriptase. Mol Cell Biol 11:1696-1706
Sharp PA (1985) On the origin of RNA splicing and introns. Cell 42:397-400
Shen C, Lazcano A, Oro J (1989) Enhancing effects of histidyl- histidine in some prebiotic reactions. J Mol Evol (submitted)
Shin Y (1973) Interaction of metal ions with polynucleotides and related compounds. XXII. Effect of divalent metal ions on the conformational changes of polyribonucleotides. Biopoly- mers 12:245%2475
Smith TF, Srinivasan A, Schochetman G, Marcus M, Myers G (1988) The phylogenetic history of immunodeficiency vi- ruses. Nature 333:573-575
Spadari S, Weissbach A (1974) HeLa cell R-deoxyribonucleic acid polymerase. J Biol Chem 249:5809-5815
Speyer JF, Chao J, Chao L (1972) Ribonucleotides covalently linked to deoxyribonucleic acid in T4 bacteriophage. J Virol 10:902-908
Stavnezer E, Ringold G, Varmus HE, Bishop M (1976) RNA complementary to the genome of RNA tumor viruses in vi- rions and virus-producing cells. J Virol 20:342-347
535
Steinhauer DA, Holland JJ (1987) Rapid evolution of RNA vi- ruses. Annu Rev Microbiol 41:409--433
Summers JA, Mason WS (1982) Replication of the genome of a hepatitis B-like virus by reverse transcription of an RNA in- termediate. Cell 29:403-415
Summers JA, O'Connell A, Millman I (1975) Genome of hepatitis B virus: restriction enzyme cleavage and structure of DNA extracted from Dane particles. Proc Natl Acad Sci USA 72: 4597-4601
Sundquist B, Oberg B (1979) Phosphonoformate inhibits reverse transcription. J Gen Virol 45:273-281
Tanese N, Goff SP (1988) Domain structure of the Maloney mu- rine leukemia virus reverse transcriptase: mutational analysis and separate expression of the DNA polymerase and RNAse H activities. Proc Natl Acad Sci USA 85:1777-1781
Temin HM (1970) Malignant transformation of cells by viruses. Perspect Biol Med 14:11-26
Temin HM (1971) The protovirus hypothesis. J Natl Cancer Inst 46:III-VII
Ternin HM (1985) Reverse transcription in the eukaryotic ge- nome: retroviruses, pararetroviruses, retrotransposons, and retrotranscripts. Mol Biol Evol 2:455-468
Temin HM (1989) Retrons in bacteria. Nature 339:254-255 Temin HM, Mizutani M (1970) RNA-directed DNA polymerase
in virions of Rous sarcoma virus. Nature 226:1211-1213 Toh H, Hayashida H, Miyata T (1983) Homology of reverse
transcriptase of retrovirus with putative polymerase gene products of hepatitis B virus and cauliflower mosaic virus. Nature 305:827-829
Valverde V, Greco-Hernandez, O1"o J, Gariglio P, Lazcano A (1989) Caracterizacion de la actividad de RNA replicasa que la reverso transcriptasa del VIH-1 presenta in vitro. Re- sumenes del II Congreso Nacional sobre SIDA. Mexico, DF, Noviembre 13-16, 1989
Vamvakopoulos NC, Vournakis JN, Marcus SL (1977) The ef- fect of magnesium and manganese ions on the structure and template activity for reverse transcriptase of polyribocyti- dylate and its 2'-o-methyl derivative. Nucleic Acid Res 4: 358%3597
Varmus H (1987) Reverse transcription. Sci AM 257:56-64 Varmus H (1989) Reverse transcription in bacteria. Cell 56:721-724 Varmus HE, Swanstrom R (1982) Replication of retroviruses. In:
Weiss R, Teich N, Varmus H, Coffin J (eds) RNA tumor viruses, vol 1. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, pp 369-512
Varmus H, Brown P (1989) Retroviruses. In: Howe M, Berg, D (eds) Mobile DNA elements. American Society of Microbi- ology, Washington, DC, pp 53-108
Velasco AM, Medrano L, Lazcano A, Or6 J (1992) A redefini- tion of the Asp-Asp domain of reverse transcriptases. J Mol Evol 35:551-556
Verma IM (1977) The reverse transcriptase. Biochem Biophys Acta 473:1-38
Visscher J, Schwartz AW (1989) Manganese-catalyzed oligomer- izations of nucleotide analogs. J Mol Evol 29:284-287
Waley SG (1969) Some aspects of the evolution of metabolic pathways. Comp Biochem Physiol 30:1-11
Wang TSF, Wong SW, Korn D (1989) Human DNA polymerase alpha: predicted functional domains and relationships with viral DNA polymerases. FASEB J 3:14-21
Weiner AM (1987) The origins of life. In Watson JD, Hopkins NH, Roberts JW, Steitz JA, Weiner AM (eds) Molecular bi- ology of the gene, vol II. Benjamin Cummings, Menlo Park CA, pp 1098-1160
Weiner AM, Deininger PL, Efstratiadis A (1986) Nonviral retro- posons: genes, pseudogenes and transposable elements gen- erated by the reverse flow of genetic information. Ann Rev Biochem 55:631-661

536
Wintersberger U, Wintersberger E (1987) RNA makes DNA: a speculative view of the evolution of DNA replication mech- anisms. Trends Genet 3:198--202
Woese CR (1967) The origins of the genetic code. Harper and Row, New York
Wong SW, Wahl AF, Yuan MP, Arai N, Pearson BE, Arai K, Korn D, Hunkapiller MW, Wang TSF (1988) Human DNA polymerase alpha gene expression is cell proliferation depen- dent and its primary structure is similar to both prokaryotic and eukaryotic replicative DNA polymerases. EMBO J 7:37- 47
Xiong Y, Eickbush TH (1988) Similarity of reverse transcriptase- like sequences of viruses, transposable elements, and mito- chondrial introns. Mol Biol Evol 5:675-690
Zavriev SK, Borisova OV (1987) Possible functional role of the "DD-domain" of RNA-dependent polymerases. Moleku- yarnaya Biologiya 21:229-241 (English translation)
Zuckerkandl E (1970) The cell machinery in its relation to bio- genesis. Rev Eur Etud Clin Biol XV:369-374
Zuckerkandl E (1975) The appearance of new structures and functions in proteins during evolution. J Mol Evol 7:1-57
Zuckerkandl E, Pauling L (1965) Molecules as documents of evolutionary history. J Theor Biol 8:357-366
Received March, 1990/Revised December 5, 1991/Accepted May 10, 1992




















