
Laser microdissection on Norway spruce bark tissue: A suitable protocol for subsequent real-time...
7
Laser microdissection on Norway spruce bark tissue: A suitable protocol for subsequent real-time reverse transcription–polymerase chain reaction (RT-PCR) analysis N. LUCHI 1 , P. CAPRETTI 1 , C. G. FOSSDAL 2 , M. PAZZAGLI 3 , & P. PINZANI 3 1 Dipartimento Biotecnologie Agrarie, Sez. Protezione delle Piante, Universita ` degli Studi di Firenze, Firenze, Italy, 2 The Norwegian Forest and Landscape Institute, A ˚ s, Norway, and 3 Dipartimento Fisiopatologia Clinica, Sez. Biochimica Clinica, Universita ` degli Studi di Firenze, Firenze, Italy Abstract The development of new tools able to select specific plant tissue is crucial for gene expression studies. During the last years, the use of laser microdissection, mainly tested on herbaceous plant tissue, has been found to be a useful technique for these purposes. This method is poorly tested on woody species, and so far no studies of gene expression have been applied on forest trees. For this reason the present work proposes the optimization of a functional protocol using laser microdissection pressure catapulting (LMPC) and real-time reverse transcription–polymerase chain reaction (RT-PCR) in bark stem tissue of Norway spruce (Picea abies). Bark tissue fragments were collected from Norway spruce trees and sliced with a cryostat. RNA was extracted from both whole cross-sections and microdissected bark cells. The feasibility of the method was confirmed by the amplification of the a-tubulin, an endogenous gene of P. abies, with efficiency comparable to that obtained from non-microdissected tissue. The proposed protocol, here adapted for bark tissue of woody species, represents a useful tool in a wide range of hosts that, unlike herbaceous plants, have scarcely been considered up to now. Keywords: a-Tubulin gene, bark, laser microdissection pressure catapulting, Picea abies, RT-PCR, woody plants Abbreviations: LAM, laser-assisted microdissection; LMPC, laser microdissection pressure catapulting; RT-PCR, reverse transcription–polymerase chain reaction. Introduction During the last years laser-assisted microdissection (LAM) has been devised to dissect morphologically homogeneous cells from microscopic tissue sections (Day et al. 2005; Pinzani et al. 2006), facilitating the quest for cell-specific information (Kehr 2003; Brandt 2005). LAM includes ‘‘laser microdissection pressure catapulting’’ (LMPC) – (P.A.L.M. Micro- laser Technologies AG, Bernried, Germany). This system uses an inverted microscope and a nitrogen laser for the selection, microdissection, and subse- quent recovery of a single cell or a small group of morphologically defined cells by catapulting them into a collection cap using the same laser source. The collected cells can then be used for RNA and DNA extraction and for downstream applications such as expression studies or gene copy number evaluation using real-time quantitative methods (Pinzani et al. 2006, 2008). LAM has been widely used, principally in medi- cine, where, combined with real-time RT-PCR quantification, it makes possible expression studies of tumor cells buried among other cells in biopsy tissues (Gelmini et al. 2006; Morrogh et al. 2007). However, devising useful protocols for the micro- dissection of tissue for clinical studies has turned out to be a laborious undertaking, the major difficulties being in the preparation of the specimens for LAM, and even more in retrieving and amplifying the nucleic acids isolated from the microportions of fixed tissue (Pinzani et al. 2006). LAM has been used much less frequently in plants, where it is mostly applied to herbaceous Correspondence: N. Luchi, Dipartimento Biotecnologie Agrarie, Sezione Protezione delle Piante, Universita ` degli Studi di Firenze, Piazzale delle Cascine 28, 50144 Firenze, Italia. Tel: þ39 0553288275. Fax: þ39 0553288273. Email: nicola.luchi@unifi.it or [email protected] Plant Biosystems, 2011; 1–7, iFirst article ISSN 1126-3504 print/ISSN 1724-5575 online ª 2011 Societa ` Botanica Italiana DOI: 10.1080/11263504.2010.505017
Transcript of Laser microdissection on Norway spruce bark tissue: A suitable protocol for subsequent real-time...

Laser microdissection on Norway spruce bark tissue: A suitable protocolfor subsequent real-time reverse transcription–polymerase chainreaction (RT-PCR) analysis
N. LUCHI1, P. CAPRETTI1, C. G. FOSSDAL2, M. PAZZAGLI3, & P. PINZANI3
1Dipartimento Biotecnologie Agrarie, Sez. Protezione delle Piante, Universita degli Studi di Firenze, Firenze, Italy, 2The
Norwegian Forest and Landscape Institute, As, Norway, and 3Dipartimento Fisiopatologia Clinica, Sez. Biochimica Clinica,
Universita degli Studi di Firenze, Firenze, Italy
AbstractThe development of new tools able to select specific plant tissue is crucial for gene expression studies. During the last years,the use of laser microdissection, mainly tested on herbaceous plant tissue, has been found to be a useful technique for thesepurposes. This method is poorly tested on woody species, and so far no studies of gene expression have been applied onforest trees. For this reason the present work proposes the optimization of a functional protocol using laser microdissectionpressure catapulting (LMPC) and real-time reverse transcription–polymerase chain reaction (RT-PCR) in bark stem tissueof Norway spruce (Picea abies). Bark tissue fragments were collected from Norway spruce trees and sliced with a cryostat.RNA was extracted from both whole cross-sections and microdissected bark cells. The feasibility of the method wasconfirmed by the amplification of the a-tubulin, an endogenous gene of P. abies, with efficiency comparable to that obtainedfrom non-microdissected tissue. The proposed protocol, here adapted for bark tissue of woody species, represents a usefultool in a wide range of hosts that, unlike herbaceous plants, have scarcely been considered up to now.
Keywords: a-Tubulin gene, bark, laser microdissection pressure catapulting, Picea abies, RT-PCR, woody plants
Abbreviations: LAM, laser-assisted microdissection; LMPC, laser microdissection pressure catapulting; RT-PCR,reverse transcription–polymerase chain reaction.
Introduction
During the last years laser-assisted microdissection
(LAM) has been devised to dissect morphologically
homogeneous cells from microscopic tissue sections
(Day et al. 2005; Pinzani et al. 2006), facilitating the
quest for cell-specific information (Kehr 2003;
Brandt 2005). LAM includes ‘‘laser microdissection
pressure catapulting’’ (LMPC) – (P.A.L.M. Micro-
laser Technologies AG, Bernried, Germany). This
system uses an inverted microscope and a nitrogen
laser for the selection, microdissection, and subse-
quent recovery of a single cell or a small group of
morphologically defined cells by catapulting them
into a collection cap using the same laser source. The
collected cells can then be used for RNA and DNA
extraction and for downstream applications such as
expression studies or gene copy number evaluation
using real-time quantitative methods (Pinzani et al.
2006, 2008).
LAM has been widely used, principally in medi-
cine, where, combined with real-time RT-PCR
quantification, it makes possible expression studies
of tumor cells buried among other cells in biopsy
tissues (Gelmini et al. 2006; Morrogh et al. 2007).
However, devising useful protocols for the micro-
dissection of tissue for clinical studies has turned out
to be a laborious undertaking, the major difficulties
being in the preparation of the specimens for LAM,
and even more in retrieving and amplifying the
nucleic acids isolated from the microportions of fixed
tissue (Pinzani et al. 2006).
LAM has been used much less frequently in
plants, where it is mostly applied to herbaceous
Correspondence: N. Luchi, Dipartimento Biotecnologie Agrarie, Sezione Protezione delle Piante, Universita degli Studi di Firenze, Piazzale delle Cascine 28,
50144 Firenze, Italia. Tel: þ39 0553288275. Fax: þ39 0553288273. Email: [email protected] or [email protected]
Plant Biosystems, 2011; 1–7, iFirst article
ISSN 1126-3504 print/ISSN 1724-5575 online ª 2011 Societa Botanica Italiana
DOI: 10.1080/11263504.2010.505017

individuals, since woody tissues such as those of the
stems of trees, are more difficult to prepare and cut
with a laser. Among studies on herbaceous plants,
some have dealt with gene expression in rice and
maize (Zhang et al. 2007; Hobo et al. 2008) some
with transcriptomic and proteomic analysis in maize
and Arabidopsis thaliana (L.) Heynh. (Dembinsky
et al. 2007; Day et al. 2008), and others with the
metabolites and chemical compounds in A. thaliana,
nettle, and alfalfa (Schad et al. 2005; Angeles et al.
2006; Nakashima et al. 2008). There are also some
studies on gene expression involved in plant–patho-
gen interactions, such as the interaction of nema-
todes on soybean (Klink et al. 2010), the powdery
mildew on Arabidopsis (Chandran et al. 2010), and
that of the anthracnose stalk rot fungus on maize
(Tang et al. 2006).
Up to now few studies, carried out on olive and
citrus, have examined laser microdissection to study
gene expression of woody plants (Corpas et al. 2006;
Agustı et al. 2009). However these studies examined
leaves and not woody tissue. A protocol for the LAM
of a conifer tree such as Norway spruce, which has
thicker lignified tissues than the herbaceous plants
mentioned above, represents a new challenge in
RNA studies. The aim of this study was to optimize
LMPC combined with real-time RT-PCR in order to
detect a-tubulin gene expression in Norway spruce
bark tissue. Effectiveness of the method on micro-
dissected tissue has been tested with particular
attention to the efficiency of amplification in whole
cross-sections on a-tubulin transcript amount. The
a-tubulin gene has previously been used as an
endogenous reference transcript in gene expression
studies in Norway spruce because its expression
levels are relatively stable (Johnsen et al. 2005;
Yakovlev et al. 2006) and little affected by treatment
such as stem wounding and inoculation with fungal
pathogens (Hietala et al. 2004; Johnk et al. 2005).
Here it was used as an example of a constitutively
expressed gene.
Materials and methods
Plant material and preparation of sections
Three healthy 30-year-old Norway spruce (Picea
abies (L.) Karst.) trees were selected in the Mon-
tesenario forest (Florence, Italy). Rectangular strips
of living bark (ca. 26 5 cm, 1-cm thick) were
excised with a scalpel from the base of the trees
down to the sapwood surface. Samples were im-
mediately frozen in liquid nitrogen and stored at
7808C until use. The samples were divided into
smaller plugs (ca. 56 56 5 mm) and then sliced to
a thickness of 10, 15, and 30 mm with a cryostat
(Frigomat, Jung, Germany) at 7208C to obtain an
optimal thickness. Three cross-sections were cut for
each tree, floated on diethylpyrocarbonate (DEPC)-
treated H2O, and then stretched on PALM1
membrane slides (1-mm glass microscope slides
coated with a 1.35-mm poly-ethylene-naphthalene
membrane). The slides were washed two times in
DEPC water and once in RNaseZap (Ambion,
Cambridgeshire, UK), and then placed for 30 min
in a UV light chamber, in order to remove the
nucleases and increase their adherence to the
membrane. Sections were then stained in 0.1%
toluidine-O-blue and dehydrated in an ethanol series
(containing DEPC water) at 70%, 95%, and 100%
for 2 min for each step.
Laser microdissection pressure catapulting
Bark cells were microdissected from sliced sections
using the PALM1 Microbeam System (P.A.L.M.
Microlaser Technologies) which employs laser pres-
sure catapulting and cold ablation by a UV laser
source (337 nm nitrogen laser), with an inverted
microscope (Carl Zeiss, Milan, Italy). The laser was
used with pulse energy higher than 270 mJ, 3 ns pulse
duration, and a cutting speed of 30 pulses s71. The
area selected for cutting was visualized on a
computer screen, and a mass of cells was dissected
with the laser beam at 406 magnification and
directly catapulted into a cap tube (Eppendorf,
Hamburg, Germany) containing 20 ml of buffer
RLT (Qiagen, Milan, Italy). The tubes were then
centrifuged at full speed for 5 s before RNA
extraction.
In order to estimate the number of cells present in
the microdissected samples, the area of the indivi-
dual cells was measured with the microscope system,
and expressed as mm2. This information was then
used to calculate the number of cells present in the
microdissected samples.
RNA extraction and reverse transcription (RT) reaction
Total RNA was extracted both from the whole cross-
sections and from microdissected Norway spruce
tissue, following the protocol of the RNeasy Micro
kit Total RNA isolation from microdissected cryo-
sections (Qiagen). To ascertain whether the tolui-
dine-O-Blue stain interfered with the RNA isolation
yield, RNA was extracted from both stained and
unstained whole cross-sections.
The RNA was extracted with an RNeasy MinElute
spin column, DNase treated, and eluted in 14 ml
of RNAse-free water. To each sample, 0.4 U ml71 of
RNAse inhibitor was added. The concentration of
the extracted RNA was measured using a Nano-
drop1 ND-1000 spectrophotometer (NanoDrop
Technologies, Wilmington, DE, USA).
2 N. Luchi et al.

Eight microliters of the total extracted RNA was
reverse-transcribed in a final volume of 20 ml
containing: 106 TaqMan RT Buffer, 5.5 mM
MgCl2, 2.5 mM of each dNTP, 1.25 U ml71
Multiscribe RT, and 1.25 mM random hexamers.
All RT reagents were purchased from Applied
Biosystems (Foster City, CA, USA). The RT
reaction was carried out on a Bio-Rad Thermal
Cycler System (Bio-Rad, Milan, Italy) kept at 258Cfor 10 min, at 488C for 30 min for reverse transcrip-
tion (RT), and at 958C for 5 min to denature the
enzyme.
Real-time PCR and the a-tubulin assay
The level of the Norway spruce a-tubulin gene
transcript used as an endogenous reference for real-
time studies (Hietala et al. 2004; Johnk et al. 2005)
was assayed using SYBR green chemistry. Each
reaction was performed in a total volume of 25 ml
containing 12.5 ml SYBR1 Green PCR Master Mix
(Applied Biosystems), 1.25 mM of each of the reverse
(50-AAGTTGTTGGCGGCGTCTT-30) and for-
ward (50-GGCATACCGGCAGCTCTTC-30) a-
tubulin primers (described in Hietala et al. 2004),
and 5 ml of cDNA. Real-time PCR was performed on
an ABI PRISM 7700 (Applied Biosystems) as
follows: a first cycle at 958C for 10 min; and 50
cycles at 958C for 15 s and at 608C for 1 min. Each
sample was measured in triplicate. Three wells
without cDNA were used as the no-template
controls, representing a negative reference.
Four 10-fold serial dilutions of RNA extracted
from Norway spruce bark tissue were used as a
standard to obtain a calibration curve to calculate the
amount of a-tubulin transcript in the unknown
microdissected tissues. The standard curve ranged
from 25 ng to 25 pg of total RNA extracted from
Norway spruce. The choice of a bulk tissue RNA
extract as a standard has the advantage to exhibit
features very close to those of the unknown samples
and thus to amplify with comparable efficiency.
PCR efficiency of sample amplification (whole and
microdissected cross-sections) was calculated by
using the program LinRegPCR 11.0, for the analysis
of quantitative RT-PCR data (Ruijter et al. 2009).
For each sample the program determines a window-
of-linearity and then uses linear regression analysis to
fit a straight line through the PCR data set. From the
slope of this line the PCR efficiency of each
individual sample is calculated (Ruijter et al. 2009).
In order to evaluate the effectiveness of LMPC
method, the estimated total RNA amount, based on
the transcript level of a-tubulin, was normalized per
area of extracted tissue (pg mm72). Quantification
was carried out for whole sections (stained and
unstained) and for the LMPC samples.
Results
Cross-sections in Norway spruce bark
The size of the cryosections of Norway spruce bark
tissue was adjusted to a thickness of 15 mm. This was
the optimal thickness that allowed the target cells to
be preserved without any solvent or cryoprotection,
leaving the single cortical cells structurally intact
before LMPC. In some cases cross-sections thicker
than 15 mm (i.e. at 30 mm) failed to stretch out on
the PEN membrane of the slide used in LMPC. In
other cases, sections thinner than 15 mm (at 10 mm)
were extremely fragile. Even at a thickness of 15 mm
there were some cell damage and cracking in the
sections. This was particularly evident for the resin
duct cells.
Laser pressure catapulting microdissection
Analyzed single bark cell had an average area of
2148.65 mm2, (n¼ 20; SD¼ 629.87), ranging from
1147 to 3294 mm2 (Figure 1a) as evaluated using the
PALM1 Microbeam System. Three distinct micro-
areas, measuring about 100,000 mm2, were micro-
dissected from each tissue slice. In some cases the
whole microdissected area (corresponding to ap-
proximately 50 bark cells) could not be catapulted in
one go, so it was decided to divide the total area to be
dissected into microareas of about 40,000 mm2 each.
For each of these microareas the tissue was selected
(Figure 1b), cut twice by a laser microbeam (Figure
1c), and catapulted into the lid of a collection tube
(Figure 1d).
RNA isolation and a-tubulin detection by real-time PCR
Total RNA was isolated with values ranging between
51.5 ng ml71 (mean value in whole cross-sections)
and 6.5 ng ml71 (mean value in microdissected tissue
that correspond *0.13 ng total RNA per cell). We
also performed preliminary extraction tests on bark
cells using Trizol, following the protocol described
by Kerk et al. (2003), which gave comparable results
(data not shown), but we decided to use the Qiagen
kit because it was less time-consuming.
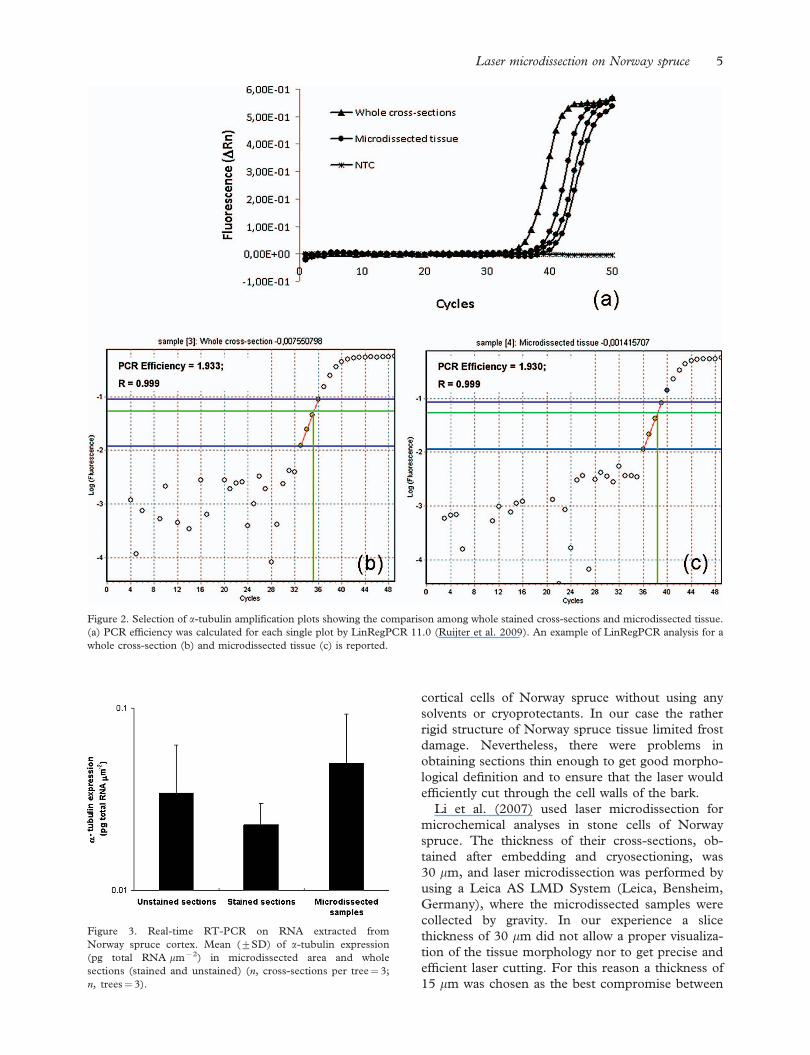
LMPC did not affect the amount of the detected a-
tubulin transcript with respect to the whole cross-
sections. All the samples gave positive results with
the real-time RT-PCR assay for a-tubulin (Figure
2a). In view of the fact that in RT and subsequent
amplification the volumes of RNA and cDNA
employed are portions of the total volume, the
average Ct value obtained for the microdissected
sample corresponded to the amount of transcript
extracted from five to six bark cells. The standard
curve used to quantify the a-tubulin transcript
exhibited a slope¼7 3.321; a Y-intercept¼ 40.44,
Laser microdissection on Norway spruce 3

and R2¼ 0.998. PCR efficiency calculated on the
kinetic obtained from microdissected samples
(1.930; R¼ 0.999) did not differ significantly
(p4 0.05) from those obtained in whole cross-
sections, both stained (1.933; R¼ 0.999) and un-
stained (1.974; R¼ 0.999) (Figure 2b and c).
Similarly, the amount of a-tubulin mRNA per area
did not differ significantly (p4 0.05) between
LMPC and the whole cross-sections, whether treated
with toluidine or not (Figure 3).
Discussion
In the present work it was demonstrated that LMPC
could be successfully used to obtain RNA for real-
time RT-PCR quantification in a woody plant such as
Norway spruce. So far, two methods have mainly
been employed to prepare sections for laser micro-
dissection in plants (mostly in herbaceous plants):
paraffin-embedded sectioning and cryosectioning
(Inada & Wildermuth 2005; Scanlon et al. 2009).
The first gives excellent histological resolution, but
sample preparation (fixation, dehydratation, and
embedding) requires a large number of steps and
this may lead to a partial loss of macromolecules such
as RNA, due to fixation, diffusion and oxidative
processes. The second method often gives a lower
resolution than the previous one from a histological
point of view but is more effective when the main
objective is RNA or DNA isolation (Kerk et al. 2003).
In herbaceous plants, cryosectioning typically
causes ice crystals to form inside the vacuoles,
damaging part of the sample (Huang et al. 2002);
for this reason, to obtain optimal histological
sections, immersion in sucrose has been suggested
(Casson et al. 2005). However, Schad et al. (2005),
in a study of the vascular bundles of Arabidopsis
found that good cryosections could also be obtained
without any immersion in sucrose. This simpler
procedure improved preservation of the cellular
structure and minimized loss of metabolites and
macromolecules such as RNA and proteins.
Taking this last study as our starting point, we
developed a protocol for the microdissection of
Figure 1. Laser microdissection pressure catapulting (LMPC) in Norway spruce cortical cells. (a) Selection and measure of the area
(expressed in mm2) of each single cell. (b) Cells, encircled on the computer screen, can be cut by LMPC. (c) After cutting, an empty space
was observed on the slide and microdissected tissue was catapulted (d) into the cap of an Eppendorf tube. Magnitude (a) and (d):640; (b)
and (c):610.
4 N. Luchi et al.
cortical cells of Norway spruce without using any
solvents or cryoprotectants. In our case the rather
rigid structure of Norway spruce tissue limited frost
damage. Nevertheless, there were problems in
obtaining sections thin enough to get good morpho-
logical definition and to ensure that the laser would
efficiently cut through the cell walls of the bark.
Li et al. (2007) used laser microdissection for
microchemical analyses in stone cells of Norway
spruce. The thickness of their cross-sections, ob-
tained after embedding and cryosectioning, was
30 mm, and laser microdissection was performed by
using a Leica AS LMD System (Leica, Bensheim,
Germany), where the microdissected samples were
collected by gravity. In our experience a slice
thickness of 30 mm did not allow a proper visualiza-
tion of the tissue morphology nor to get precise and
efficient laser cutting. For this reason a thickness of
15 mm was chosen as the best compromise between
Figure 2. Selection of a-tubulin amplification plots showing the comparison among whole stained cross-sections and microdissected tissue.
(a) PCR efficiency was calculated for each single plot by LinRegPCR 11.0 (Ruijter et al. 2009). An example of LinRegPCR analysis for a
whole cross-section (b) and microdissected tissue (c) is reported.
Figure 3. Real-time RT-PCR on RNA extracted from
Norway spruce cortex. Mean (+SD) of a-tubulin expression
(pg total RNA mm72) in microdissected area and whole
sections (stained and unstained) (n, cross-sections per tree¼ 3;
n, trees¼ 3).
Laser microdissection on Norway spruce 5

histological integrity and laser cutting suitability,
even if the cell walls on account of their lignification
sometimes still resisted cutting.
In addition, problems during laser cutting may be
due to an inadequately performed tissue preparation,
such as the dehydration. This step is very important
because it avoids the formation of small water bub-
bles, which may prevent tissue adhesion to the slide,
making laser microdissection impossible (Pinzani
et al. 2006). In our case the ethanol series used was
suitable for Norway spruce bark tissue.
The use of stain is particularly important in LAM
because it facilitates identification of the various cell
types and brings the retrieved sample into better
view. Our results showed that toluidine, already used
in conifer tissue of Pinus nigra Arn. (Luchi et al.
2005), did not significantly affect the yield of total
RNA, as demonstrated by the results obtained with
real-time RT-PCR from whole tissue sections
(Figure 3).
In the present study, in which LMPC was
combined with RNA extraction, it was also necessary
to find a balance between reasonable histological
resolution and macromolecule extraction. The use of
cryosections was helpful to detect specific cells for
harvesting and allowed the macromolecules to be
isolated. RNA was successfully extracted from both
whole sections and the microdissected areas, by a
spin-column based extraction method (RNeasy
Micro kit) commonly used for animal tissues
(Pinzani et al. 2006). The effectiveness of the
extraction method has been demonstrated, as re-
ported in Figure 3, where the amount of a-tubulin
expression did not differ between microdissected and
whole samples. This result has also been confirmed
after PCR efficiency analysis that showed no
differences not only between these two kinds of
samples (Figure 2b and c), but also between whole
stained and unstained cross-section, showing that
toluidine did not affect the PCR efficiency.
The microdissection protocol combined with real
time RT-PCR in Norway spruce bark samples
demonstrated the suitability of the method in heavily
lignified plant tissue. Consequently, the objective of
the present study was fully reached in devising a
protocol for laser microdissection, followed by RNA
quantitative determination for forest trees. In the last
few years the use of laser microdissection has
revealed itself to be a useful tool to study gene
expression in plant–microbe interactions (Balestrini
& Bonfante 2008; Gomez & Harrison 2009;
Chandran et al. 2010). The development of this
protocol for the bark stem tissue of forest trees
represents an important new way to study gene
modulation at the level of specific cells during the
interaction of cells of Norway spruce with patho-
genic microorganisms.
Acknowledgements
This study was carried out by grants from the
University of Florence ‘Fondi di Ateneo’. We grate-
fully acknowledge the Section of Plant Biology (DiBA,
University of Florence) for cryostat sectioning.
References
Agustı J, Merelo P, Cercos M, Tadeo FR, Talon M. 2009.
Comparative transcriptional survey between laser-microdis-
sected cells from laminar abscission zone and petiolar cortical
tissue during ethylene-promoted abscission in citrus leaves.
BMC Plant Biol 9: 127.
Angeles G, Berrio-Sierra J, Joseleau JP, Lorimier P, Lefebvre A,
Ruel K. 2006. Preparative laser capture microdissection and
single-pot cell wall material preparation: A novel method for
tissue-specific analysis. Planta 224: 228–232.
Balestrini R, Bonfante P. 2008. Laser microdissection (LM):
Application to plant materials. Plant Biosyst 142: 331–336.
Brandt SP. 2005. Microgenomics: Gene expression analysis at the
tissue-specific and single-cell levels. J Exp Bot 56: 495–505.
Casson S, Spencer M, Walker K, Lindsey K. 2005. Laser capture
microdissection for the analysis of gene expression during
embryogenesis of Arabidopsis. Plant J 42: 111–123.
Chandran D, Inada N, Hather G, Kleindt CK, Wildermuth MC.
2010. Laser microdissection of Arabidopsis cells at the powdery
mildew infection site reveals site-specific processes and
regulators. Proc Natl Acad Sci USA 107: 460–465.
Corpas FJ, Fernandez-Ocana A, Carreras A, Valderrama R, Luque
F, Esteban FJ, et al. 2006. The expression of different
superoxide dismutase forms is cell-type dependent in olive
(Olea europaea L.) leaves. Plant Cell Physiol 47: 984–994.
Day RC, Grossniklaus U, Macknight RC. 2005. Be more specific!
Laser microdissection of plant cells. Trends Plant Sci 10: 397–
406.
Day RC, Herridge RP, Ambrose BA, Macknight RC. 2008.
Transcriptome analysis of proliferating Arabidopsis endosperm
reveals biological implications for the control of syncytial
division, cytokinin signalling, and gene expression regulation.
Plant Physiol 148: 1964–1984.
Dembinsky D, Woll K, Saleem M, Liu Y, Fu Y, Borsuk LA, et al.
2007. Transcriptomic and proteomic analyses of pericycle cells
of the maize primary root. Plant Physiol 145: 575–588.
Gelmini S, Poggesi M, Pinzani P, Mannurita SC, Cianchi F,
Valanzano R, et al. (2006). Distribution of Tankyrase-1
mRNA expression in colon cancer and its prospective
correlation with progression stage. Oncol Rep 16: 1261–1266.
Gomez SK, Harrison MJ. 2009. Laser microdissection and its
application to analyze gene expression in arbuscular mycor-
rhizal symbiosis. Pest Manag Sci 65: 504–511.
Hietala AM, Kvaalen H, Schmidt A, Johnk N, Solheim H, Fossdal
CG. 2004. Temporal and spatial profiles of chitinase expression
by Norway spruce in response to bark colonization by Hete-
robasidion annosum. Appl Environ Microbiol 70: 3948–3953.
Hobo T, Suwabe K, Aya K, Suzuki G, Yano K, Ishimizu T, et al.
2008. Various spatiotemporal expression profiles of anther-
expressed genes in rice. Plant Cell Physiol 49: 1417–1428.
Huang LE, Luzzi V, Ehrig T, Holtschlag V, Watson MA. 2002.
Optimized tissue processing and staining for laser capture
microdissection and nucleic acid retrieval. Meth Enzymol 356:
49–62.
Inada N, Wildermuth MC. 2005. Novel tissue preparation method
and cell-specific marker for laser microdissection of Arabidopsis
mature leaf. Planta 221: 9–16.
6 N. Luchi et al.

Johnk N, Hietala AM, Fossdal CG, Collinge DB, Newman M.
2005. Defense-related genes expressed in Norway spruce roots
after infection with the root rot pathogen Ceratobasidium bicorne
(anamorph: Rhizoctonia sp.). Tree Physiol 25: 1533–1543.
Johnsen Ø, Fossdal CG, Nagy N, Mølmann J, Dæhlen O, Skrøppa
T. 2005. Climatic adaptation in Picea abies progenies is affected
by the temperature during zygotic embryogenesis and seed
maturation. Plant Cell Environ 28: 1090–1102.
Kehr J. 2003. Single cell technology. Curr Op Plant Biol 6: 617–
621.
Kerk NM, Ceserani T, Tausta SL, Sussex IM, Nelson TM. 2003.
Laser capture microdissection of cells from plant tissues. Plant
Physiol 132: 27–35.
Klink VP, Hosseini P, Matsye PD, Alkharouf NW, Matthews BF.
2010. Syncytium gene expression in Glycine max ([PI 88788])
roots undergoing a resistant reaction to the parasitic nematode
Heterodera glycines. Plant Physiol Biochem 48: 176–193.
Li SH, Schneider B, Gershenzon J. 2007. Microchemical analysis
of laser-microdissected stone cells of Norway spruce by
cryogenic nuclear magnetic resonance spectroscopy. Planta
225: 771–779.
Luchi N, Ma R, Capretti P, Bonello P. 2005. Systemic induction
of traumatic resin ducts and resin flow in Austrian pine by
wounding and inoculation with Sphaeropsis sapinea and
Diplodia scrobiculata. Planta 221: 75–84.
Morrogh M, Olvera N, Bogomolniy F, Borgen PI, King TA. 2007.
Tissue preparation for laser capture microdissection and RNA
extraction from fresh frozen breast tissue. BioTechniques 43:
41–48.
Nakashima J, Chen F, Jackson L, Shadle G, Dixon RA. 2008.
Multi-site genetic modification of monolignol biosynthesis in
alfalfa (Medicago sativa): Effects on lignin composition in
specific cell types. New Phytol 179: 738–750.
Pinzani P, Lind K, Malentacchi F, Nesi G, Salvianti F, Villari D,
et al. 2008. Prostate-specific antigen mRNA and protein levels
in laser microdissected cells of human prostate measured by
real-time reverse transcriptase-quantitative polymerase chain
reaction and immuno-quantitative polymerase chain reaction.
Hum Pathol 39: 1474–1482.
Pinzani P, Orlando C, Pazzagli M. 2006. Laser-assisted micro-
dissection for real-time PCR sample preparation. Mol Asp
Med 27: 140–159.
Ruijter JM, Ramakers C, Hoogaars W, Bakker O, van den Hoff
MJB, Karlen Y, et al. 2009. Amplification efficiency: linking
baseline and bias in the analysis of quantitative PCR data.
Nucleic Acids Res. 37: e45.
Scanlon MJ, Ohtsu K, Timmermans MC, Schnable PS. 2009.
Laser microdissection-mediated isolation and in vitro tran-
scriptional amplification of plant RNA. Curr Protoc Mol Biol
(Unit 25A.3): 1–15. DOI: 10.1002/0471142727.mb25a03s87.
Schad M, Mungur R, Fiehn O, Kehr J. 2005. Metabolic profiling
of laser microdissected vascular bundles of Arabidopsis thaliana.
Plant Meth 1: 1–10.
Tang W, Coughlan S, Crane E, Beatty M, Duvick J. 2006. The
application of laser microdissection to in planta gene expres-
sion profiling of the maize anthracnose stalk rot fungus
Colletotrichum graminicola. Mol Plant Micr Inter 19: 1240–
1250.
Yakovlev I, Fossdal CG, Johnsen Ø, Junttila O, Skrøppa T. 2006.
Analysis of gene expression during bud burst initiation in
Norway spruce via ESTs from subtracted cDNA libraries. Tree
Gen Genomes 2: 39–52.
Zhang X, Madi S, Borsuk L, Nettleton D, Elshire RJ, Buckner B,
et al. 2007. Laser microdissection of narrow sheath mutant
maize uncovers novel gene expression in the shoot apical
meristem. PLoS Genet 3: e101.
Laser microdissection on Norway spruce 7




















