
On the distribution and evaluation of Na, Mg and Cl in leaves of selected halophytes
6
On the distribution and evaluation of Na, Mg and Cl in leaves of selected halophytes Paula Pongrac a , Katarina Vogel-Mikuš a , Marjana Regvar a , Mitja Kaligaric ˇ b , Primoz ˇ Vavpetic ˇ c , Mitja Kelemen c , Nataša Grlj c , Oren Shelef d , Avi Golan-Goldhirsh d , Shimon Rachmilevitch d , Primoz ˇ Pelicon c,⇑ a Department of Biology, Biotechnical Faculty, University of Ljubljana, Vec ˇna pot 111, SI-1000 Ljubljana, Slovenia b Department of Biology, Faculty of Natural Sciences and Mathematics, Koroška c. 160, SI-2000 Maribor, Slovenia c Joz ˇef Stefan Institute, Jamova 39, SI-1000 Ljubljana, Slovenia d French Associates Institute for Agriculture & Biotechnology of Drylands, The Jacob Blaustein Institutes for Desert Research (BIDR), Ben-Gurion University of the Negev, 84990 Midreshet Ben-Gurion, Israel article info Article history: Received 23 July 2012 Received in revised form 29 November 2012 Accepted 5 December 2012 Available online xxxx Keywords: Light elements Micro-PIXE Salinity Salt tolerance abstract Diverse physiological, biochemical and morphological adaptations enable plants to survive in extreme sal- ine environments where osmotic and ionic stresses limit growth and development. Halophytes are salt-tol- erant plants that can withstand extraordinarily high levels of Na and Cl in their leaves. The tissue and cellular distribution patterns of salt ions can be linked to the underlying mechanisms of salt tolerance. Application of fast, reliable, multi-elemental and quantitative techniques such as micro-proton-induced X-ray emission (micro-PIXE) will significantly contribute to and accelerate studies of plant salt tolerance, especially as micro-PIXE also provides spatially resolved quantitative data for light elements, such as Na and Mg. The spatial concentration distributions of Na, Mg, Cl, K, P and S in leaves of four halophytes (Bassia indica, Atriplex prostrata, Spartina maritima and Limonium angustifolium) were determined using micro- PIXE, to study the salt-tolerance strategies of the selected halophytes. Different distribution patterns of the studied elements were seen in the leaves; however, in all four of these plant species, Na was excluded from photosynthetically active chlorophyl tissues. With the exception of L. angustifolium, Cl, P and S con- tents (representing chloride, phosphate and sulphate ionic forms, respectively) did not ensure charge bal- ance in the leaves, which suggests other anionic compounds, such as nitrate and organic anions, have crucial roles in maintaining electroneutrality in these halophytes. By increasing soil salinisation worldwide, the possibility to reliably complement spatial distributions of Na, Mg, Cl, K, P and S with plant structural mor- phology will contribute significantly to our understanding of plant tolerance mechanisms at the tissue and cell levels. In addition, these kinds of studies are of particular value for designing crop plants with high salt tolerance and for the development of phytoremediation technologies. Ó 2013 Elsevier B.V. All rights reserved. 1. Introduction High salinity is one of the major environmental factors that lim- it plant growth and productivity, through the consequent osmotic stress, ion toxicity and nutrient imbalance. Accumulation of Na + in the cytoplasm not only results in ion imbalance and affects critical biochemical processes, but also increases plasma-membrane in- jury, causes malondialdehyde accumulation, and impairs photo- synthesis. Nevertheless, some organisms have developed a combination of physiological, biochemical, and morphological adaptations that enable them to thrive under extreme salt condi- tions. Plants with these features are known as halophytes [1]. As a consequence of their multiple origins, halophytes differ widely in their salt-tolerance mechanisms. Overall tolerance to salinity relies on the uptake and compartmentalisation of Na + ,K + and Cl and the synthesis of organic anions (such as malate, citrate and glutamate), all of which serve to adjust to the external low water potential [2]. Previous studies into the spatial concentration distributions of these monovalent ions in halophytes have mainly focused on salt tolerance in crops, as generally glycophytes, which lack the genetic basis for salt tolerance [3]. In general, striking dif- ferences in concentrations between the Na + -rich vacuoles and the K + -rich cytoplasm have been reported for halophytes [4]. Extreme Na + gradients between salt exclusion cells, leaf epidermis and mesophyll cells have been observed [5,6]. It was suggested that vacuoles of non-photosynthetically active cells, such as epidermal, water storage and parenchyma cells, serve as the main storage pools of excess Na + and exchange of K + with the rest of the leaf 0168-583X/$ - see front matter Ó 2013 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.nimb.2012.12.057 ⇑ Corresponding author. Tel.: +386 1 5885294; fax: +386 1 4773151. E-mail address: [email protected] (P. Pelicon). Nuclear Instruments and Methods in Physics Research B xxx (2013) xxx–xxx Contents lists available at SciVerse ScienceDirect Nuclear Instruments and Methods in Physics Research B journal homepage: www.elsevier.com/locate/nimb Please cite this article in press as: P. Pongrac et al., On the distribution and evaluation of Na, Mg and Cl in leaves of selected halophytes, Nucl. Instr. Meth. B (2013), http://dx.doi.org/10.1016/j.nimb.2012.12.057
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of On the distribution and evaluation of Na, Mg and Cl in leaves of selected halophytes

Nuclear Instruments and Methods in Physics Research B xxx (2013) xxx–xxx
Contents lists available at SciVerse ScienceDirect
Nuclear Instruments and Methods in Physics Research B
journal homepage: www.elsevier .com/locate /n imb
On the distribution and evaluation of Na, Mg and Cl in leaves of selected halophytes
Paula Pongrac a, Katarina Vogel-Mikuš a, Marjana Regvar a, Mitja Kaligaric b, Primoz Vavpetic c,Mitja Kelemen c, Nataša Grlj c, Oren Shelef d, Avi Golan-Goldhirsh d, Shimon Rachmilevitch d,Primoz Pelicon c,⇑a Department of Biology, Biotechnical Faculty, University of Ljubljana, Vecna pot 111, SI-1000 Ljubljana, Sloveniab Department of Biology, Faculty of Natural Sciences and Mathematics, Koroška c. 160, SI-2000 Maribor, Sloveniac Jozef Stefan Institute, Jamova 39, SI-1000 Ljubljana, Sloveniad French Associates Institute for Agriculture & Biotechnology of Drylands, The Jacob Blaustein Institutes for Desert Research (BIDR), Ben-Gurion University of the Negev, 84990Midreshet Ben-Gurion, Israel
a r t i c l e i n f o a b s t r a c t
Article history:Received 23 July 2012Received in revised form 29 November 2012Accepted 5 December 2012Available online xxxx
Keywords:Light elementsMicro-PIXESalinitySalt tolerance
0168-583X/$ - see front matter � 2013 Elsevier B.V.http://dx.doi.org/10.1016/j.nimb.2012.12.057
⇑ Corresponding author. Tel.: +386 1 5885294; fax:E-mail address: [email protected] (P. Pelicon).
Please cite this article in press as: P. Pongrac et a(2013), http://dx.doi.org/10.1016/j.nimb.2012.1
Diverse physiological, biochemical and morphological adaptations enable plants to survive in extreme sal-ine environments where osmotic and ionic stresses limit growth and development. Halophytes are salt-tol-erant plants that can withstand extraordinarily high levels of Na and Cl in their leaves. The tissue andcellular distribution patterns of salt ions can be linked to the underlying mechanisms of salt tolerance.Application of fast, reliable, multi-elemental and quantitative techniques such as micro-proton-inducedX-ray emission (micro-PIXE) will significantly contribute to and accelerate studies of plant salt tolerance,especially as micro-PIXE also provides spatially resolved quantitative data for light elements, such as Naand Mg. The spatial concentration distributions of Na, Mg, Cl, K, P and S in leaves of four halophytes (Bassiaindica, Atriplex prostrata, Spartina maritima and Limonium angustifolium) were determined using micro-PIXE, to study the salt-tolerance strategies of the selected halophytes. Different distribution patterns ofthe studied elements were seen in the leaves; however, in all four of these plant species, Na was excludedfrom photosynthetically active chlorophyl tissues. With the exception of L. angustifolium, Cl, P and S con-tents (representing chloride, phosphate and sulphate ionic forms, respectively) did not ensure charge bal-ance in the leaves, which suggests other anionic compounds, such as nitrate and organic anions, have crucialroles in maintaining electroneutrality in these halophytes. By increasing soil salinisation worldwide, thepossibility to reliably complement spatial distributions of Na, Mg, Cl, K, P and S with plant structural mor-phology will contribute significantly to our understanding of plant tolerance mechanisms at the tissue andcell levels. In addition, these kinds of studies are of particular value for designing crop plants with high salttolerance and for the development of phytoremediation technologies.
� 2013 Elsevier B.V. All rights reserved.
1. Introduction
High salinity is one of the major environmental factors that lim-it plant growth and productivity, through the consequent osmoticstress, ion toxicity and nutrient imbalance. Accumulation of Na+ inthe cytoplasm not only results in ion imbalance and affects criticalbiochemical processes, but also increases plasma-membrane in-jury, causes malondialdehyde accumulation, and impairs photo-synthesis. Nevertheless, some organisms have developed acombination of physiological, biochemical, and morphologicaladaptations that enable them to thrive under extreme salt condi-tions. Plants with these features are known as halophytes [1].
All rights reserved.
+386 1 4773151.
l., On the distribution and evalu2.057
As a consequence of their multiple origins, halophytes differwidely in their salt-tolerance mechanisms. Overall tolerance tosalinity relies on the uptake and compartmentalisation of Na+, K+
and Cl� and the synthesis of organic anions (such as malate, citrateand glutamate), all of which serve to adjust to the external lowwater potential [2]. Previous studies into the spatial concentrationdistributions of these monovalent ions in halophytes have mainlyfocused on salt tolerance in crops, as generally glycophytes, whichlack the genetic basis for salt tolerance [3]. In general, striking dif-ferences in concentrations between the Na+-rich vacuoles and theK+-rich cytoplasm have been reported for halophytes [4]. ExtremeNa+ gradients between salt exclusion cells, leaf epidermis andmesophyll cells have been observed [5,6]. It was suggested thatvacuoles of non-photosynthetically active cells, such as epidermal,water storage and parenchyma cells, serve as the main storagepools of excess Na+ and exchange of K+ with the rest of the leaf
ation of Na, Mg and Cl in leaves of selected halophytes, Nucl. Instr. Meth. B

2 P. Pongrac et al. / Nuclear Instruments and Methods in Physics Research B xxx (2013) xxx–xxx
as demonstrated for epidermal cells of salt-tolerant Thellungiellahalophila [7].
Without doubt, more information on the spatial concentrationdistributions of salt ions/elements at the tissue, cell and subcellularlevels is needed in different halophytes to elucidate the general salttolerance mechanisms [2]. Application of micro-proton-induced X-ray emission (micro-PIXE) can accelerate such studies, especiallyas it provides spatially resolved quantitative data on biologicallyimportant elements. One of the first attempts to localise elementsin halophyte plant tissue with micro-PIXE was a study of Aster trip-olium roots [8]. In the non-mycorrhizal root cross-section the mainaccumulation of Na, P and S was seen in the rhizodermis area,whereas Cl was distributed reversly and was accompanied by a highK concentration, probably to maintain homeostasis under this saltstress [8]. There are no detailed studies of the spatial distributionsof salt ions/elements in leaves of halophytes with micro-PIXE. Thus,spatial distributions of Na, Mg, Cl, K, P and S in leaves of four selectedhalophytes were studied; namely, for two Chenopodiaceae speciesBassia indica and Atriplex prostrata, and for Spartina maritima (Poa-ceae) and Limonium angustifolium (Plumbaginaceae). B. indica is anannual shrub, and a tumbleweed, and it is frequently found on dis-turbed land in Israel [9]. B. indica accumulates salts mainly in itsleaves [10] and its roots grow towards higher salinity, thus exhibit-ing positive root halotropism [11]. The remaining three halophyteswere collected from or in the vicinity of seawater along the Adriaticcoast in Slovenia, and these are used as model halophyte species [2].A. prostrata is an annual plant, and a facultative halophyte with saltexcreting hairs on the leaf surface [12]. The other two halophytes, S.maritima and L. angustifolium, are herbaceous perennials. The mono-cotyledonous S. maritima is an important ecosystem model plant insalt marshes [13], and the dicotyledonous L. angustifolium is an orna-mental plant species that has salt glands [14,15].
The aims of the study were to: (i) illustrate the applicability ofmicro-PIXE in resolving spatial concentration distributions of Na,Mg, Cl, K, P and S in these four selected halophytes; (ii) relate thespatial distribution patterns of these elements to the morphologi-cal features that are characteristic for each of these halophytes;and (iii) assess the contributions of chloride, sulphate and phos-phate to the maintenance of the charge balance in the leaves ofthese tolerant plants using micro-PIXE.
2. Experimental
2.1. Plant material and sample preparation
B. indica (Wight) A.J. Scott (Kochia; Chenopodiaceae) plantswere grown in a salinity gradient [11]. Plant specimens of A. pro-strata Bouch. Ex DC. [A. hastate aucht.] subsp. prostrata (Chenopo-diaceae), S. maritima (Curt.) Fern. [S. stricta (Ait.) Roth] (Poaceae),and L. angustifolium (Tauch) Degen [Statice serotina Rchb., L. vulgareMill. subsp. serotinum (Rchb.) Gams] (Plumbaginaceae) were col-lected in the St. Jernej canal in the vicinity of the Secovlje salterns(Slovenia). The coordinates for the sampling points were as fol-lows: 45�490980 0 N, and 13�580800 0 E for A. prostrata, 45�490830 0 Nand 13�590350 0 E for S. maritima and 45�490990 0 N and 13�580950 0 Efor L. angustifolium.
The micrographic images of the leaf cross-sections were ob-tained with an Axiocam MRc colour digital camera attached toan Axioscop 2 MOT microscope (Carl Zeiss, Goettingen, Germany)using the AxioVision 4.1 software.
The plants were carefully dug from the substrate with the rhizo-sphere soil intact, and transferred to the laboratory, where fresh,fully developed leaves were detached, washed in deionised water,plunge-frozen in propane cooled with liquid nitrogen, cut using acryo-microtome, and freeze-dried [16–18].
Please cite this article in press as: P. Pongrac et al., On the distribution and evalu(2013), http://dx.doi.org/10.1016/j.nimb.2012.12.057
2.2. Micro-proton induced X-ray emission analysis
The H� beam used for these analyses was directly extractedfrom hydrogen plasma created in a duoplasmatron ion source,and injected into a tandem accelerator, where protons wereformed in a stripping process. The accelerated proton beam was fo-cused onto the object slit of the nuclear microprobe.
The X-rays emitted had energies from 1 keV up to 25 keV, andwere detected with a high-purity germanium detector with an ac-tive area of 95 mm2. This consisted of a 25-lm-thick berylliumwindow and a 100-lm-thick polyimide absorber, which were posi-tioned at 135� to the incoming beam. A Si(Li) detector with an areaof 10 mm2 and an 8-lm-thick Be window was positioned at 125�with respect to the incoming beam, and was simultaneously usedfor the detection of X-rays in the energy range from 0.8 keV to4 keV. The samples were irradiated with low-energy electronsfrom a hot tungsten filament to avoid sample charging during irra-diation, which eliminated need for the time-consuming specimencarbon coating.
The proton dose was determined by a rotating in-beam chopper[19]. Measurement and evaluation of the X-ray data was per-formed as described previously [16,19–22]. The calibration of thePIXE method was verified with standard reference materials NISTSRM 1573a (tomato leaves, homogenised powder, analysed in aform of pressed pellet), NIST SRM 1107 (Naval Brass B, alloy) andNIST SRM 620 (Soda-Lime Flat Glass). The inter-calibration of PIXEand STIM was verified using thin mono-elemental metallic foils.The description of the X-ray detection efficiency below the Si KX-ray absorption edge was improved [23].
The concentrations of elements in specific tissues of interestwere extracted from numeric concentration matrices generatedusing GeoPIXE II, by manually encircling the selected regions inthe elemental distribution map using the ImageJ programme(http://www.macbiophotonics.ca/imagej/). The co-localisation ele-mental maps were generated using GeoPIXE II.
3. Results and discussion
3.1. Morphological features of the selected halophytes
Leaf cross-sections of B. indica, but not A. prostrata (both Cheno-podiaceae), showed conspicuous water storage tissue (WST), to-gether with voluminous epidermal cells (Fig. 1A and B, top leftpanels). In B. indica, the WST was located below the vascular bundlein the centre of the leaf lamina, indicating the succulence nature ofthis species. On the other hand, in A. prostrata instead of a WST, therewas extensive midrib parenchyma tissue that enclosed the mainvascular bundle of the leaf. In comparison to A. prostrata, a C3 plantspecies, the leaf of B. indica has a Kranz anatomy; i.e., it has two typesof tissue where photosynthesis takes place, namely bundle sheathcells (BSCs), which are missing in A. prostrata and which containstarch-rich chloroplasts but lack grana, and mesophyll cells. In B. in-dica, the leaf BSCs create a semi-circle to surround the vascular bun-dles on one side and to border the mesophyll cells on the other.Kranz anatomy of the leaf was also seen in S. maritima (Poaceae)(Fig. 1C, top left panel). The leaf of S. maritima has a flat abaxialand a ridged adaxial side, which particularly resembles the leavesof S. alterniflora and S. anglica, where the BSCs surround vascularbundles and the cells extend, usually in two rows, up to the adaxialepidermis in each ridge [24]. In the monocotyledonous S. maritima,there is an additional collenchymatous cell layer just beneath theepidermis on the top of each ridge and parenchyma tissue underliesthe adaxial epidermis. Salt-excreting glands have been reported forseveral Spartina species [25]. However these glands could not be lo-cated in the analysed leaf cross-sections. The dicotyledonous L.
ation of Na, Mg and Cl in leaves of selected halophytes, Nucl. Instr. Meth. B
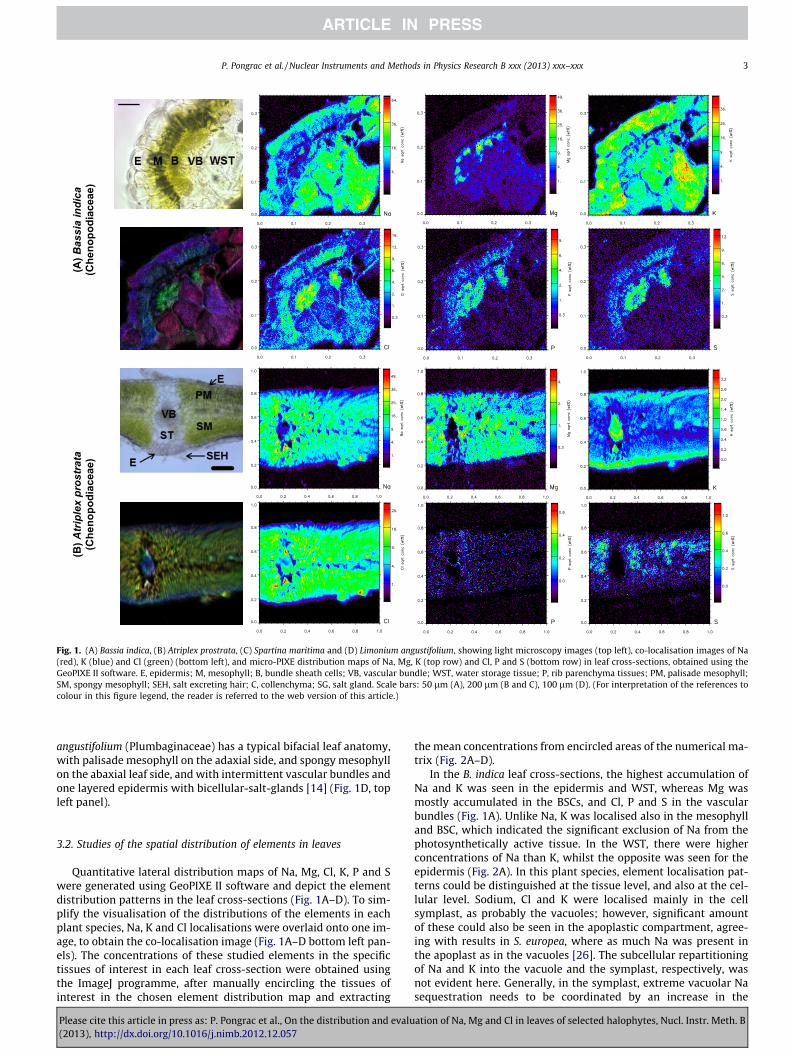
(A)B
assi
a in
dica
(Chenopodiaceae)
(B)A
trip
lex
pros
trat
a(Chenopodiaceae)
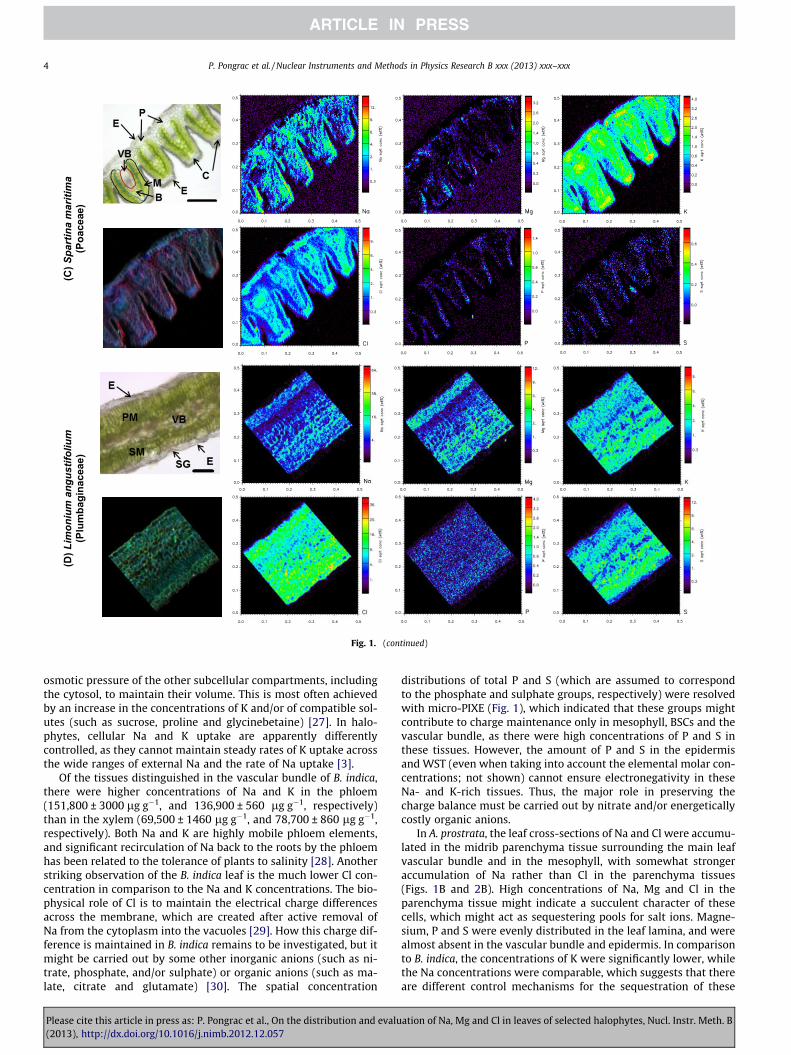
Fig. 1. (A) Bassia indica, (B) Atriplex prostrata, (C) Spartina maritima and (D) Limonium angustifolium, showing light microscopy images (top left), co-localisation images of Na(red), K (blue) and Cl (green) (bottom left), and micro-PIXE distribution maps of Na, Mg, K (top row) and Cl, P and S (bottom row) in leaf cross-sections, obtained using theGeoPIXE II software. E, epidermis; M, mesophyll; B, bundle sheath cells; VB, vascular bundle; WST, water storage tissue; P, rib parenchyma tissues; PM, palisade mesophyll;SM, spongy mesophyll; SEH, salt excreting hair; C, collenchyma; SG, salt gland. Scale bars: 50 lm (A), 200 lm (B and C), 100 lm (D). (For interpretation of the references tocolour in this figure legend, the reader is referred to the web version of this article.)
P. Pongrac et al. / Nuclear Instruments and Methods in Physics Research B xxx (2013) xxx–xxx 3
angustifolium (Plumbaginaceae) has a typical bifacial leaf anatomy,with palisade mesophyll on the adaxial side, and spongy mesophyllon the abaxial leaf side, and with intermittent vascular bundles andone layered epidermis with bicellular-salt-glands [14] (Fig. 1D, topleft panel).
3.2. Studies of the spatial distribution of elements in leaves
Quantitative lateral distribution maps of Na, Mg, Cl, K, P and Swere generated using GeoPIXE II software and depict the elementdistribution patterns in the leaf cross-sections (Fig. 1A–D). To sim-plify the visualisation of the distributions of the elements in eachplant species, Na, K and Cl localisations were overlaid onto one im-age, to obtain the co-localisation image (Fig. 1A–D bottom left pan-els). The concentrations of these studied elements in the specifictissues of interest in each leaf cross-section were obtained usingthe ImageJ programme, after manually encircling the tissues ofinterest in the chosen element distribution map and extracting
Please cite this article in press as: P. Pongrac et al., On the distribution and evalu(2013), http://dx.doi.org/10.1016/j.nimb.2012.12.057
the mean concentrations from encircled areas of the numerical ma-trix (Fig. 2A–D).
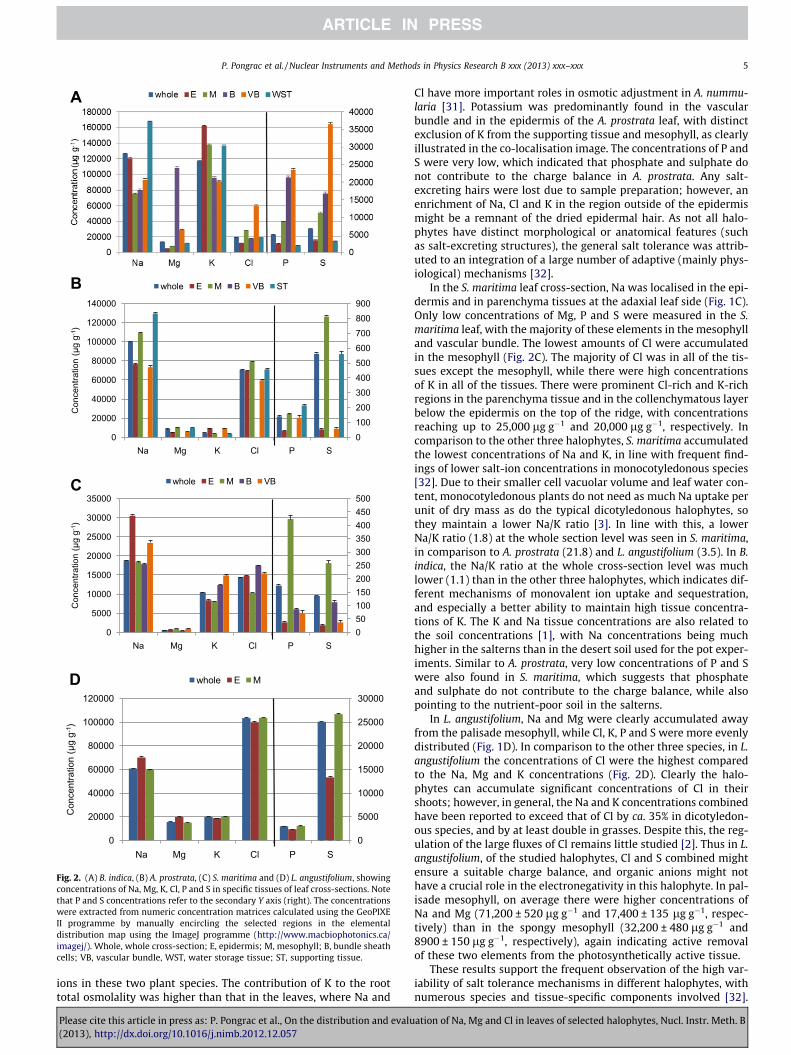
In the B. indica leaf cross-sections, the highest accumulation ofNa and K was seen in the epidermis and WST, whereas Mg wasmostly accumulated in the BSCs, and Cl, P and S in the vascularbundles (Fig. 1A). Unlike Na, K was localised also in the mesophylland BSC, which indicated the significant exclusion of Na from thephotosynthetically active tissue. In the WST, there were higherconcentrations of Na than K, whilst the opposite was seen for theepidermis (Fig. 2A). In this plant species, element localisation pat-terns could be distinguished at the tissue level, and also at the cel-lular level. Sodium, Cl and K were localised mainly in the cellsymplast, as probably the vacuoles; however, significant amountof these could also be seen in the apoplastic compartment, agree-ing with results in S. europea, where as much Na was present inthe apoplast as in the vacuoles [26]. The subcellular repartitioningof Na and K into the vacuole and the symplast, respectively, wasnot evident here. Generally, in the symplast, extreme vacuolar Nasequestration needs to be coordinated by an increase in the
ation of Na, Mg and Cl in leaves of selected halophytes, Nucl. Instr. Meth. B
(C)S
part
ina
mar
itim
a(Poaceae)
(D)L
imon
ium
ang
ustif
oliu
m(Plumbaginaceae)
Fig. 1. (continued)
4 P. Pongrac et al. / Nuclear Instruments and Methods in Physics Research B xxx (2013) xxx–xxx
osmotic pressure of the other subcellular compartments, includingthe cytosol, to maintain their volume. This is most often achievedby an increase in the concentrations of K and/or of compatible sol-utes (such as sucrose, proline and glycinebetaine) [27]. In halo-phytes, cellular Na and K uptake are apparently differentlycontrolled, as they cannot maintain steady rates of K uptake acrossthe wide ranges of external Na and the rate of Na uptake [3].
Of the tissues distinguished in the vascular bundle of B. indica,there were higher concentrations of Na and K in the phloem(151,800 ± 3000 lg g�1, and 136,900 ± 560 lg g�1, respectively)than in the xylem (69,500 ± 1460 lg g�1, and 78,700 ± 860 lg g�1,respectively). Both Na and K are highly mobile phloem elements,and significant recirculation of Na back to the roots by the phloemhas been related to the tolerance of plants to salinity [28]. Anotherstriking observation of the B. indica leaf is the much lower Cl con-centration in comparison to the Na and K concentrations. The bio-physical role of Cl is to maintain the electrical charge differencesacross the membrane, which are created after active removal ofNa from the cytoplasm into the vacuoles [29]. How this charge dif-ference is maintained in B. indica remains to be investigated, but itmight be carried out by some other inorganic anions (such as ni-trate, phosphate, and/or sulphate) or organic anions (such as ma-late, citrate and glutamate) [30]. The spatial concentration
Please cite this article in press as: P. Pongrac et al., On the distribution and evalu(2013), http://dx.doi.org/10.1016/j.nimb.2012.12.057
distributions of total P and S (which are assumed to correspondto the phosphate and sulphate groups, respectively) were resolvedwith micro-PIXE (Fig. 1), which indicated that these groups mightcontribute to charge maintenance only in mesophyll, BSCs and thevascular bundle, as there were high concentrations of P and S inthese tissues. However, the amount of P and S in the epidermisand WST (even when taking into account the elemental molar con-centrations; not shown) cannot ensure electronegativity in theseNa- and K-rich tissues. Thus, the major role in preserving thecharge balance must be carried out by nitrate and/or energeticallycostly organic anions.
In A. prostrata, the leaf cross-sections of Na and Cl were accumu-lated in the midrib parenchyma tissue surrounding the main leafvascular bundle and in the mesophyll, with somewhat strongeraccumulation of Na rather than Cl in the parenchyma tissues(Figs. 1B and 2B). High concentrations of Na, Mg and Cl in theparenchyma tissue might indicate a succulent character of thesecells, which might act as sequestering pools for salt ions. Magne-sium, P and S were evenly distributed in the leaf lamina, and werealmost absent in the vascular bundle and epidermis. In comparisonto B. indica, the concentrations of K were significantly lower, whilethe Na concentrations were comparable, which suggests that thereare different control mechanisms for the sequestration of these
ation of Na, Mg and Cl in leaves of selected halophytes, Nucl. Instr. Meth. B
0100200300400500600700800900
0
20000
40000
60000
80000
100000
120000
140000
Na Mg K Cl P S
Con
cent
ratio
n (µ
gg-
1 )
whole E M B VB STB
050100150200250300350400450500
0
5000
10000
15000
20000
25000
30000
35000
Na Mg K Cl P S
Con
cent
ratio
n (µ
gg-
1 )
whole E M B VBC
0
5000
10000
15000
20000
25000
30000
0
20000
40000
60000
80000
100000
120000
Na Mg K Cl P S
Con
cent
ratio
n (µ
gg-
1 )
whole E MD
A
Fig. 2. (A) B. indica, (B) A. prostrata, (C) S. maritima and (D) L. angustifolium, showingconcentrations of Na, Mg, K, Cl, P and S in specific tissues of leaf cross-sections. Notethat P and S concentrations refer to the secondary Y axis (right). The concentrationswere extracted from numeric concentration matrices calculated using the GeoPIXEII programme by manually encircling the selected regions in the elementaldistribution map using the ImageJ programme (http://www.macbiophotonics.ca/imagej/). Whole, whole cross-section; E, epidermis; M, mesophyll; B, bundle sheathcells; VB, vascular bundle, WST, water storage tissue; ST, supporting tissue.
P. Pongrac et al. / Nuclear Instruments and Methods in Physics Research B xxx (2013) xxx–xxx 5
ions in these two plant species. The contribution of K to the roottotal osmolality was higher than that in the leaves, where Na and
Please cite this article in press as: P. Pongrac et al., On the distribution and evalu(2013), http://dx.doi.org/10.1016/j.nimb.2012.12.057
Cl have more important roles in osmotic adjustment in A. nummu-laria [31]. Potassium was predominantly found in the vascularbundle and in the epidermis of the A. prostrata leaf, with distinctexclusion of K from the supporting tissue and mesophyll, as clearlyillustrated in the co-localisation image. The concentrations of P andS were very low, which indicated that phosphate and sulphate donot contribute to the charge balance in A. prostrata. Any salt-excreting hairs were lost due to sample preparation; however, anenrichment of Na, Cl and K in the region outside of the epidermismight be a remnant of the dried epidermal hair. As not all halo-phytes have distinct morphological or anatomical features (suchas salt-excreting structures), the general salt tolerance was attrib-uted to an integration of a large number of adaptive (mainly phys-iological) mechanisms [32].
In the S. maritima leaf cross-section, Na was localised in the epi-dermis and in parenchyma tissues at the adaxial leaf side (Fig. 1C).Only low concentrations of Mg, P and S were measured in the S.maritima leaf, with the majority of these elements in the mesophylland vascular bundle. The lowest amounts of Cl were accumulatedin the mesophyll (Fig. 2C). The majority of Cl was in all of the tis-sues except the mesophyll, while there were high concentrationsof K in all of the tissues. There were prominent Cl-rich and K-richregions in the parenchyma tissue and in the collenchymatous layerbelow the epidermis on the top of the ridge, with concentrationsreaching up to 25,000 lg g�1 and 20,000 lg g�1, respectively. Incomparison to the other three halophytes, S. maritima accumulatedthe lowest concentrations of Na and K, in line with frequent find-ings of lower salt-ion concentrations in monocotyledonous species[32]. Due to their smaller cell vacuolar volume and leaf water con-tent, monocotyledonous plants do not need as much Na uptake perunit of dry mass as do the typical dicotyledonous halophytes, sothey maintain a lower Na/K ratio [3]. In line with this, a lowerNa/K ratio (1.8) at the whole section level was seen in S. maritima,in comparison to A. prostrata (21.8) and L. angustifolium (3.5). In B.indica, the Na/K ratio at the whole cross-section level was muchlower (1.1) than in the other three halophytes, which indicates dif-ferent mechanisms of monovalent ion uptake and sequestration,and especially a better ability to maintain high tissue concentra-tions of K. The K and Na tissue concentrations are also related tothe soil concentrations [1], with Na concentrations being muchhigher in the salterns than in the desert soil used for the pot exper-iments. Similar to A. prostrata, very low concentrations of P and Swere also found in S. maritima, which suggests that phosphateand sulphate do not contribute to the charge balance, while alsopointing to the nutrient-poor soil in the salterns.
In L. angustifolium, Na and Mg were clearly accumulated awayfrom the palisade mesophyll, while Cl, K, P and S were more evenlydistributed (Fig. 1D). In comparison to the other three species, in L.angustifolium the concentrations of Cl were the highest comparedto the Na, Mg and K concentrations (Fig. 2D). Clearly the halo-phytes can accumulate significant concentrations of Cl in theirshoots; however, in general, the Na and K concentrations combinedhave been reported to exceed that of Cl by ca. 35% in dicotyledon-ous species, and by at least double in grasses. Despite this, the reg-ulation of the large fluxes of Cl remains little studied [2]. Thus in L.angustifolium, of the studied halophytes, Cl and S combined mightensure a suitable charge balance, and organic anions might nothave a crucial role in the electronegativity in this halophyte. In pal-isade mesophyll, on average there were higher concentrations ofNa and Mg (71,200 ± 520 lg g�1 and 17,400 ± 135 lg g�1, respec-tively) than in the spongy mesophyll (32,200 ± 480 lg g�1 and8900 ± 150 lg g�1, respectively), again indicating active removalof these two elements from the photosynthetically active tissue.
These results support the frequent observation of the high var-iability of salt tolerance mechanisms in different halophytes, withnumerous species and tissue-specific components involved [32].
ation of Na, Mg and Cl in leaves of selected halophytes, Nucl. Instr. Meth. B

6 P. Pongrac et al. / Nuclear Instruments and Methods in Physics Research B xxx (2013) xxx–xxx
The exclusion of Na from photosynthetically active tissues appearsto be a common mechanism of salt tolerance that is seen in all fourof these selected halophytes. With the exception of L. angustifolium,the concentrations of Cl, P and S (the latter representing phosphateand sulphate, respectively) cannot ensure the charge balance in theleaves, which suggests other anionic compounds have crucial rolesin the maintenance of electroneutrality.
These conclusions significantly contribute to the current knowl-edge of salt tolerance mechanisms. With the application of fast,reliable, multi-elemental and quantitative techniques such asthese, micro-PIXE studies of salt tolerance are expected to be accel-erated. Such studies respond to the need to understand the mech-anisms of salt tolerance, which will in future help to design cropplants with increased salt tolerance, and to discover other halo-phytes with this potential [3].
Acknowledgements
This study was supported by the European Community as anIntegrating Activity ‘Support of Public and Industrial ResearchUsing Ion Beam Technology (SPIRIT)’ under EC contract no.227012. The authors also acknowledge the support of the Slove-nian Research Agency for the P1-0112 and P1-0212 Research Pro-grammes, and for the research infrastructure of the MicroanalyticalCentre.
References
[1] I. Türkan, T. Demiral, Recent developments in understanding salinity tolerance,Environ. Exp. Bot. 67 (2009) 2–9.
[2] T.J. Flowers, T.D. Colmer, Salinity tolerance in halophytes, New Phytol. 179(2008) 945–963.
[3] E.P. Glenn, J.J. Brown, E. Blumwald, Salt tolerance and crop potential ofhalophytes, Crit. Rev. Plant Sci. 18 (1999) 227–255.
[4] C.J. Ye, K.F. Zhao, Osmotically active compounds and their localization in themarine halophyte eelgrass, Biol. Plant 46 (2003) 137–140.
[5] B.J. Barkla, R. Vera-Estrella, J. Camacho-Emiterio, O. Pantoja, Na+/H+ exchangein the halophyte Mesembryanthemum crystallinum is associated with cellularsites of Na+ storage, Funct. Plant Biol. 29 (2002) 1017–1024.
[6] R. Storey, M.G. Pitman, R. Stelzer, C. Carter, X-ray micro analysis of cells andcell components of Atriplex spongiosa 1. Leaves, J. Exp. Bot. 34 (1983) 778–794.
[7] V. Volkov, B. Wang, P.J. Dominy, W. Fricke, A. Amtmann, Thellungiella halophila,a salt tolerant relative of Arabidopsis thaliana, possesses effective mechanismsto discriminate between potassium and sodium, Plant Cell Environ. 27 (2003)1–14.
[8] S. Scheloske, M. Maetz, T. Schenider, U. Hildebrandt, H. Bothe, B. Povh, Elementdistribution in mycorrhizal and nonmycorrhizal roots of the halophyte Astertripolium determined by proton induced X-ray emission, Protoplasma 223(2004) 183–189.
[9] N. Feinbrun-Dothan, A. Danin, Analytical Flora of Eretz-Israel, third ed., CanaPublishing House, Jerusalem, 1998.
[10] O. Shelef, A. Gross, S. Rachmilevitch, The use of Bassia indica for saltphytoremediation in constructed wetlands, Water Res. 46 (2012) 3967–3976.
[11] O. Shelef, N. Lazarovitch, B. Rewald, A. Golan-Goldhirsh, S. Rachmilevitch, Roothalotropism: salinity effects on Bassia indica root, Plant Biosyst. 144 (2010)471–478.
Please cite this article in press as: P. Pongrac et al., On the distribution and evalu(2013), http://dx.doi.org/10.1016/j.nimb.2012.12.057
[12] L.-W. Wang, A.M. Showalter, I.A. Ungar, Effect of salinity on growth, ioncontent, and cell wall chemistry in Atriplex prostrata (Chenopodiaceae), Am. J.Bot. 84 (1997) 1247–1255.
[13] E.M. Castellanos, C. Heredia, M.E. Figueroa, A.J. Davy, Tiller dynamics ofSpartina maritima in successional and non-successional Mediterranean saltmarsh, Plant Ecol. 137 (1998) 213–225.
[14] W. Larcher, Physiological Plant Ecology: Ecophysiology and Stress Physiologyof Functional Groups, Springer-Verlag, Berlin, 2001.
[15] A. Poljakoff-Majber, J. Gale, Plants in Saline Environments, Springer-Verlag,Berlin, Heidelberg, New York, 1975.
[16] K. Vogel-Mikuš, J. Simcic, P. Pelicon, M. Budnar, P. Kump, M. Necemer, J.Mesjasz-Przybyłowicz, W.J. Przybyłowicz, M. Regvar, Comparison of essentialand non-essential element distribution in leaves of the Cd/Znhyperaccumulator Thlaspi praecox as revealed by micro-PIXE, Plant CellEnviron. 31 (2008) 1484–1496.
[17] K. Vogel-Mikuš, M. Regvar, J. Mesjasz-Przybyłowicz, W.J. Przybyłowicz, J.Simcic, P. Pelicon, M. Budnar, Spatial distribution of cadmium in leaves ofmetal hyperaccumulating Thlaspi praecox as revealed by micro-PIXE, NewPhytol. 179 (2008) 712–721.
[18] P. Pongrac, K. Vogel-Mikuš, P. Vavpetic, J. Tratnik, M. Regvar, J. Simcic, N. Grlj,P. Pelicon, Cd induced redistribution of elements within leaves of the Cd/Znhyperaccumulator Thlaspi praecox as revealed by micro-PIXE, Nucl. Instr. Meth.B 268 (2010) 2205–2210.
[19] K. Vogel-Mikuš, P. Pelicon, P. Vavpetic, I. Kreft, M. Regvar, Elemental analysis ofedible grains by micro-PIXE: common buckwheat case study, Nucl. Instr. Meth.B 267 (2009) 2884–2889.
[20] K. Vogel-Mikuš, P. Pongrac, P. Kump, M. Necemer, J. Simcic, P. Pelicon, M.Budnar, B. Povh, M. Regvar, Localisation of metals and minerals within Thlaspipraecox seeds by micro-PIXE reveals its reproductive success strategies atmetal polluted sites, Environ. Pollut. 147 (2007) 50–59.
[21] K. Vogel-Mikuš, P. Pongrac, P. Pelicon, P. Vavpetic, B. Povh, H. Bothe, M. Regvar,Micro-PIXE analysis for localisation and quantification of elements in roots ofmycorrhizal metal-tolerant plants, in: A. Varma, A.C. Kharkwal (Eds.),Symbiotic Fungi: Principles and Practice, Soil Biology, 18, Springer,Heidelberg, 2009, pp. 227–242.
[22] J.L. Campbell, T.L. Hopman, J.A. Maxwell, Z. Nejedly, The Guelph PIXE softwarepackage III: alternative proton database, Nucl. Instr. Meth. B 170 (2000) 193–204.
[23] P. Vavpetic, N. Grlj, L. Jeromel, N. Ogrinc, P. Pelicon, Quantification of micro-PIXE by Si(Li) X-ray detector for the elements with characteristic K X-rayenergies below the silicon absorption edge, Poster presentation (P-107) at theICNMTA 2012 conference.
[24] B.R. Maricle, N.K. Koteyeva, E.V. Voznesenskaya, J.R. Thomasson, G.E. Edwards,Diversity in leaf anatomy, and stomatal distribution and conductance,between salt marsh and freshwater species in the C4 genus Spartina(Poaceae), New Phytol. 184 (2010) 216–233.
[25] C.A. Levering, W.W. Thompson, The ultrastructure of the salt gland of Spartinafoliosa, Planta 97 (1971) 183–196.
[26] S. Lv, P. Jiang, X. Chen, P. Fan, X. Wang, Y. Li, Multiple compartmentalization ofsodium conferred salt tolerance in Salicornia europaea, Plant Physiol. Biochem.51 (2012) 47–52.
[27] R. Munns, M. Tester, Mechanisms of salinity tolerance, Annu. Rev. Plant Biol.59 (2008) 651–681.
[28] M. Tester, R. Davenport, Na+ tolerance and Na+ transport in higher plants, Ann.Bot. 91 (2003) 503–527.
[29] P.J. White, M.R. Broadley, Chloride in soils and its uptake and movementwithin the plant: a review, Ann. Bot. 88 (2001) 967–988.
[30] E. Martinoia, S. Meyer, A. De Angeli, R. Nagy, Vacuolar transporters in theirphysiological context, Annu. Rev. Plant Biol. 63 (2012) 183–213.
[31] J.A. Gomes Silveira, S.A. Marinho Araújo, J.P.M. Santos Lima, R. Almeida Viégas,Roots and leaves display contrasting osmotic adjustment mechanisms inresponse to NaCl-salinity in Atriplex nummularia, Environ. Exp. Bot. 66 (2009)1–8.
[32] S. Shabala, A. Mackay, Ion transport in halophytes, in: J.C. Kader, M. Delseny(Eds.), Advances in Botanical Research, vol. 57, Academic Press, Burlington,2011, pp. 151–199.
ation of Na, Mg and Cl in leaves of selected halophytes, Nucl. Instr. Meth. B




















