
O R I G I N A L A R T I C L E Technological properties and probiotic potential of Enterococcus...
13
ORIGINAL ARTICLE Technological properties and probiotic potential of Enterococcus faecium strains isolated from cow milk K. Banwo 1,2 , A. Sanni 1 and H. Tan 2 1 Department of Microbiology, University of Ibadan, P.M.B 1 Ibadan, Oyo State, Nigeria 2 State Key Laboratory of Microbial Resources, Institute of Microbiology, Chinese Academy of Sciences, Beijing, China Keywords enterocin A, Enterococcus faecium, milk, Nono, safety. Correspondence Kolawole Banwo, Department of Microbiology, University of Ibadan, P.M.B 1 Ibadan, Oyo State, Nigeria. E-mail: [email protected] 2012/1083: received 14 June 2012, revised 29 August 2012 and accepted 29 September 2012 doi:10.1111/jam.12031 Abstract Aim: To identify enterococci from the fermentation of milk for the production of nono, an African fermented dairy product, to determine the technological properties for suitability as starter cultures and safety as probiotics. Methods and Results: Enterococcus faecium CM4 and Enterococcus faecium 2CM1 were isolated from raw cow’s milk. The strains were phenotypically and genotypically identified. Technological properties, safety investigations, in vitro adherence properties and antimicrobial characteristics were carried out. Strong acidification and tolerance to bile salts were recorded. The strains were bile salts hydrolytic positive and no haemolysis. There was no resistance to clinically relevant antibiotics. The strains exhibited adherence to human collagen type IV, human fibrinogen and fibronectin. The bacteriocins were active against Bacillus cereus DSM 2301, Bacillus subtilis ATCC 6633, Micrococcus luteus and Listeria monocytogenes. Bacteriocins were stable at pH 4 –9 and on treatment with lipase, catalase, a-amylase and pepsin, while their activity was lost on treatment with other proteases. The bacteriocins produced were heat stable at 100°C for 10 min. The bacteriocin produced by the strains was identified as enterocin A. Conclusions: The E. faecium strains in this study exhibited probiotic activity, and the safety investigations indicate their suitability as good candidates for a starter culture fermentation process. Significance and Impact of the Study: The use of bacteriocin-producing E. faecium strains as starter cultures in fermented foods is beneficial but, however, their safety investigations as probiotics must be greatly emphasized. Introduction Enterococcus species are Gram-positive bacteria, faculta- tive anaerobic cocci that occur singly, in pairs or in chains (Giraffa 2003). They are commensal micro-organ- isms that colonize in the gastrointestinal tract of humans and animals and are also found in several different food sources, such as meats, milk and cheese. These bacteria are able to survive extreme environments, such as 6 5% NaCl, pH of 9 6, high heat as well as being able to grow and survive under other harsh environmental conditions, like those found in various soils, surface water, raw plants and animal products (Giraffa 2003; Johnston and Jaykus 2004). Enterococci are utilized in many diverse roles in a multi- tude of different processes. For instance, they serve as an important contributor in the ripening and flavour enhancement of several types of food such as cheeses and sausages (Giraffa 2003). In dairy products, their major role is in the development of organoleptic characteristics during the development and maturation process (Giraffa 2003; Cocolin et al. 2007). These bacteria have also been utilized as probiotics to improve the microbial balance of the intes- tine and to treat gastroenteritis in humans and animals Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology 229 Journal of Applied Microbiology ISSN 1364-5072
Transcript of O R I G I N A L A R T I C L E Technological properties and probiotic potential of Enterococcus...

ORIGINAL ARTICLE
Technological properties and probiotic potential ofEnterococcus faecium strains isolated from cow milkK. Banwo1,2, A. Sanni1 and H. Tan2
1 Department of Microbiology, University of Ibadan, P.M.B 1 Ibadan, Oyo State, Nigeria
2 State Key Laboratory of Microbial Resources, Institute of Microbiology, Chinese Academy of Sciences, Beijing, China
Keywords
enterocin A, Enterococcus faecium, milk,
Nono, safety.
Correspondence
Kolawole Banwo, Department of
Microbiology, University of Ibadan, P.M.B 1
Ibadan, Oyo State, Nigeria.
E-mail: [email protected]
2012/1083: received 14 June 2012, revised
29 August 2012 and accepted 29 September
2012
doi:10.1111/jam.12031
Abstract
Aim: To identify enterococci from the fermentation of milk for the
production of nono, an African fermented dairy product, to determine the
technological properties for suitability as starter cultures and safety as
probiotics.
Methods and Results: Enterococcus faecium CM4 and Enterococcus faecium
2CM1 were isolated from raw cow’s milk. The strains were phenotypically and
genotypically identified. Technological properties, safety investigations, in vitro
adherence properties and antimicrobial characteristics were carried out. Strong
acidification and tolerance to bile salts were recorded. The strains were bile
salts hydrolytic positive and no haemolysis. There was no resistance to
clinically relevant antibiotics. The strains exhibited adherence to human
collagen type IV, human fibrinogen and fibronectin. The bacteriocins were
active against Bacillus cereus DSM 2301, Bacillus subtilis ATCC 6633,
Micrococcus luteus and Listeria monocytogenes. Bacteriocins were stable at pH 4
–9 and on treatment with lipase, catalase, a-amylase and pepsin, while their
activity was lost on treatment with other proteases. The bacteriocins produced
were heat stable at 100°C for 10 min. The bacteriocin produced by the strains
was identified as enterocin A.
Conclusions: The E. faecium strains in this study exhibited probiotic activity,
and the safety investigations indicate their suitability as good candidates for a
starter culture fermentation process.
Significance and Impact of the Study: The use of bacteriocin-producing
E. faecium strains as starter cultures in fermented foods is beneficial but,
however, their safety investigations as probiotics must be greatly emphasized.
Introduction
Enterococcus species are Gram-positive bacteria, faculta-
tive anaerobic cocci that occur singly, in pairs or in
chains (Giraffa 2003). They are commensal micro-organ-
isms that colonize in the gastrointestinal tract of humans
and animals and are also found in several different food
sources, such as meats, milk and cheese. These bacteria
are able to survive extreme environments, such as 6�5%NaCl, pH of 9�6, high heat as well as being able to grow
and survive under other harsh environmental conditions,
like those found in various soils, surface water, raw plants
and animal products (Giraffa 2003; Johnston and Jaykus
2004).
Enterococci are utilized in many diverse roles in a multi-
tude of different processes. For instance, they serve as an
important contributor in the ripening and flavour
enhancement of several types of food such as cheeses and
sausages (Giraffa 2003). In dairy products, their major role
is in the development of organoleptic characteristics during
the development and maturation process (Giraffa 2003;
Cocolin et al. 2007). These bacteria have also been utilized
as probiotics to improve the microbial balance of the intes-
tine and to treat gastroenteritis in humans and animals
Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology 229
Journal of Applied Microbiology ISSN 1364-5072

(Kayser 2003). Furthermore, enterococci harbour some
useful biotechnological and functional properties, such as
production of antimicrobials with antilisterial activity
(Foulquie-Moreno et al. 2003; Cocolin et al. 2007).
The antimicrobial properties of the strains have been
ascribed to the production of bacteriocins (Todorov and
Dicks 2006), which are defined as small proteins or pep-
tides with bactericidal or bacteriostatic activity against
genetically closely related species (Klaenhammer 1993).
There have been published reports on bacteriocin-pro-
ducing enterococci, mainly among the strains of Entero-
coccus faecium associated with food ecosystems and dairy
products (Cocolin et al. 2007).
There have been numerous reports on bacteriocin-pro-
ducing bacteria, primarily among strains of lactic acid bac-
teria (LAB) associated with food systems (Park et al.
2003), but there is little information on Enterococcus spe-
cies from African fermented food sources (Yousif et al.
2005) especially traditional fermented dairy products
(Olasupo et al. 1999; Yousif et al. 2005). Nono is fermented
cow milk taken with fura which is made from ground
millet. Hence, the name Fura de nono, African fermented
milk curd. This is consumed mainly by the northern region
of Nigeria and sub-Saharan Africa. The milk is either
consumed immediately after milking from the cow or left
to the next day. The method employed by the indigenous
people in the preservation of the milk is usually back slop-
ping (Eka and Ohaba 1977). Hence, the importance in the
development of starter culture fermentation process to
ensure consistency in the product and safety.
Bacteriocin production by LAB is believed to confer an
ecological advantage over competitors present in the
same ecosystem, as Enterococcus species form part of the
autochthonous flora of the intestinal tract of humans and
animals (Fuller 1989; Javed et al. 2011) and have also
been isolated from fermenting plant and dairy products
(Villani and Coppola 1994), they show potential for
application as probiotics. Their antibiotic susceptibility
profile also ensures their safety as probiotics especially
the absence of antibiotics and multidrug resistance genes
(Yousif et al. 2005). Enterococci from food sources have
relatively low virulence, and there are some beneficial
activities of some strains; they are also considered as nos-
ocomial pathogens, which cause bacteraemia, endocarditis
and other infections (Ben-Omar et al. 2004).
Production of bacteriocins by enterococci maybe an
advantage in the use as a probiotic because the bacterioc-
ins add up to antimicrobial effect of other substances
such as hydrogen peroxide and organic acids and con-
tribute to the competition in the gastrointestinal tract
(Strompfova and Laukova 2007). LAB strains that possess
certain functional properties such as tolerance to bile
salts, acidification, absence of antibiotic resistance genes,
in vitro adhesion capabilities and also production of bac-
teriocins render them good candidates for starter culture
fermentation process and safety as probiotics (Giraffa
2003; Yousif et al. 2005; Strompfova and Laukova 2007;
Javed et al. 2011).
In our study, we evaluated the probiotic potentials and
the bacteriocin production of two newly isolated Ent. fae-
cium strains from fermenting raw cow’s milk in Nigeria.
The aim of the study was to investigate the probiotic
properties, functionality and safety of the Ent. faecium
strains, to employ them as starter cultures for the
production of nono African fermented dairy product.
Materials and methods
Identification of bacteriocin-producing strains
Seven isolates from raw milk were characterized pheno-
typically as Enterococcus species. The strains were grown
in De Man, Rogosa, Sharpe agar (Oxoid, Basingstoke,
UK) at 37°C for 24–48 h. They were presumptively iden-
tified by the following tests: observation of colonial char-
acteristics and cell morphology, Gram staining, catalase,
growth at 15 and 45°C, growth in the presence of 6�5%NaCl and at pH 9�6 and fermentation of wide range of
sugars. Genetic identification to species level was per-
formed by 16S rRNA sequencing, as described by Brosius
et al. (1978) and Kostinek et al. (2005).
Acidification
The Enterococcus faecium strains were inoculated (1% of
an overnight culture) into MRS broth adjusted to pH
6�75 before autoclaving (pH 6�54 after autoclaving) and
grown aerobically at 30°C. Acid production was deter-
mined by measuring the pH of the culture after 6, 24
and 48 h. The MRS broth medium for all acid produc-
tion tests was prepared from a single batch that was pH
adjusted and then dispensed into tubes of 10 ml each
before autoclaving (Kostinek et al. 2005).
Resistance to bile salts
The ability of the strains to grow in the presence of bile
was determined according to the method of Walker and
Gilliland (1993) as modified by Vinderola and Reinhei-
mer (2003). Each strain was inoculated (2% v/v) into
MRS broth with 0�3, 0�5 and 1% (w/v) of bile (Sigma-
Aldrich, Saint Louis, MO, USA). Cultures were incubated
at 37°C for 24 h; optical density (OD) of the inoculated
tube was measured at OD560 nm wavelength and
compared with a control culture (without bile salts). This
was expressed as the percentage of growth at OD560 nm in
230 Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology
Enterococcus faecium strains from milk K. Banwo et al.

the presence of bile salts compared with the control. Each
determination was carried out in triplicates.
Bile salts hydrolytic activity
Bile salt hydrolytic (BSH) plates were prepared by adding
0�5% (w/v) sodium glycodeoxycholate (GCDA) and 0�5%(w/v) taurodeoxycholate (TDCA) to MRS agar. The
organisms were streaked on the plates and incubated at
37°C for 24–48 h, and the activities were determined by
the precipitation of deconjugated bile salts around the col-
onies, which was recorded as positive (Minelli et al. 2004).
Haemolysis and production of gelatinase
The strains were cultured in MRS broth at 37°C for 12
–18 h and then transferred onto blood agar (Difco, Michi-
gan, USA) plates supplemented with 5% defibrinated
whole sheep blood as described by Yoon et al. (2008).
After 24–48 h, the haemolytic reaction was recorded by the
observation of a partial hydrolysis of red blood cells and
greening zone (a-haemolysis), clear zone around bacterial
growth (b-haemolysis) and no reaction (c-haemolysis).
To determine the presence of gelatinase activity, the
plate assay method on Todd–Hewitt agar containing 30 g
of gelatin l�1 was carried out as described by Ben-Omar
et al. (2004).
Antibiotic susceptibility testing of the Enterococcus
faecium strains
The antibiotic strips for the determination of susceptibil-
ity profile were used according to the manufacturer’s
instructions. The susceptibility was determined towards
ten antibiotics, namely amoxycillin (25 lg), ofloxacin
(5 lg), streptomycin (10 lg), chloramphenicol (30 lg),cefriazone (30 lg), gentamicin (10 lg), pefloxacin (5 lg),cotrimaxazole (25 lg), ciprofloxacin (10 lg) and erythro-
mycin (5 lg). Tetracycline and vancomycin were tested
separately in a dilution of 30 lg ml�1 and 30 lg ml�1 in
sterile distilled water, respectively.
This was determined semiquantitatively using the agar
overlay diffusion methods of National Committee for Clin-
ical Laboratory Standards (NCCLS 1990). A bacterial sus-
pension was made by picking colonies from MRS agar
plates using a sterile loop and making a suspension in ster-
ile MRS broth and left for 18–24 h. With the use of a sterile
swab, the suspension was applied on the surface of Mueller
–Hinton agar in sterile Petri dishes, and the strips were
placed on the surface using a sterile scalpel. For tetracycline
and vancomycin, a 3-mm sterile cork borer was used after
which 100 ll of the antibiotic solution was dispensed into
the holes. The plates were then incubated at 30°C for 48 h
after which the readings were taken (Mueller and Hinton
1941; Hummel et al. 2007; Mathara et al. 2008).
In vitro adherence assay
This assay was carried out according to the study by Schil-
linger et al. (2005) with slight modification. Commercial
precoated 96-well human fibronectin (Fn) and human col-
lagen type IV cell culture plates were obtained from
Becton-Dickinson (San Jose, CA, USA) and human plasma
fibrinogen (Fb) (Sigma-Aldrich). Human plasma fibrino-
gen was dissolved in 0�1 mol l�1 phosphate-buffered saline
(PBS, pH 7�4) at a concentration of 50 lg ml�1 and used
to coat 96-well cell culture microtiter plates from Corning
(Lowell, MA, USA). The strains were grown in Dulbecco’s
modified Eagle’s medium (DMEM; Invitrogen, Beijing
China) supplemented with 2% (v/v) foetal bovine serum
(FBS; MDgenics, USA) for 24 h at 37°C. Cells were har-
vested by centrifugation at 10 000 g for 5 min and the pel-
lets resuspended in DMEM and diluted in 10�4 dilution
factor in Tris/borate/EDTA (TBE) buffer pH 8�0, and Syto
62 red (Molecular Probes, Eugene, OR, USA) was added as
the fluorescent dye for detection in the flow cytometer.
The cell count was determined using a BD FACS cell sorter
(BD Biosciences, San Jose, CA, USA). A 1-ml aliquot of the
bacterial suspension was adjusted to 1 9 108 cfu ml�1.
The wells were filled with 200 ll of extracellular matrices
(ECM) protein (Fb, Fn and collagen type IV) and incu-
bated overnight at 4°C. The protein solution was removed
and wells washed twice with PBS. Then, FBS (200 ll of a2% solution in DMEM) was added to the wells to block
unoccupied sites and to prevent nonspecific binding of the
bacteria. The FBS was removed using a pipette after incu-
bation at 37°C for 1 h. Two hundred microlitres of the ini-
tial bacterial suspension in DMEM was added to the ECM-
coated cell culture plates. The plates were centrifuged at
2000 g for 120 s and incubated at 37°C for 1 h. The
unbound bacteria were removed with a pipette into an Ep-
pendorf tube diluted in TBE buffer and the cell count per-
formed with a BD FACS cell sorter (USA). The number of
nonadherent and adherent bacteria was established and
expressed as a percentage of the initial bacterial suspension.
Each determination was performed in triplicates. Probiotic
strain Lactobacillus acidophilus CNRZ 1923 and Lactobacil-
lus rhamnosus GG were used as controls.
Determination of bacteriocin production and
antimicrobial spectrum
The preparation of the cell-free culture supernatant was
carried out, as described by Cocolin et al. (2007). The
strains Ent. faecium CM4 and 2CM1 were grown in MRS
broth (Oxoid) at 37°C for 18–24 h. Cultures were centri-
Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology 231
K. Banwo et al. Enterococcus faecium strains from milk

fuged at 13 000 g for 15 min at 4°C (Eppendorf 5810 R,
California, USA), and supernatants were collected and
adjusted to pH 7 with NaOH and filtered through a 0�2-lm membrane filter (Millipore, Billerica, MA, USA). The
antimicrobial spectrum of the cell-free culture supernatant
was carried out using agar well diffusion assay, as
described by Schillinger and Lucke (1989). The indicator
strains were grown in appropriate media such as MRS
(Oxoid) for LAB and brain heart infusion agar (Difco) for
non-LAB. Wells were punctured in the soft agar with a
sterile cork borer (5 mm) after solidification. These wells
were inoculated with 100 ll of the cell-free supernatants ofthe Ent. faecium strains, and the assay plates were incu-
bated at 30–37°C, aerobically for non-LAB and anaerobi-
cally for LAB for 18–24 h. At the end of the incubation
period, the inhibition zones were detected by a clear region
around the well. MRS broth without micro-organisms was
used as a control. Each determination was carried out in
duplicates.
Effect of enzymes, heat and pH on bacteriocin activity
The stability of the bacteriocins was tested after treatment
with catalase, proteinase K, trypsin, pepsin, lysozyme,
pronase E, lipase and a-amylase (Sigma-Aldrich). The
enzymes were added to 1 ml of neutralized cell-free cul-
ture supernatant to a final concentration of 2 mg ml�1 in
all cases, and 100 ll was inoculated into assayed plates
(Cocolin et al. 2007). The plates were incubated at 37°Cfor 18–24 h, and the antimicrobial activity spectrum was
carried out by measuring the zones of inhibition against
indicator strains Bacillus cereus DSM 2301 and Bacillus
subtilis ATCC 6633. The untreated cell-free supernatants
were used as controls. Each determination was carried
out in duplicates.
To determine the effect of pH and heat treatment on
the bacteriocin activity, final pH values of the cell-free cul-
ture supernatants, ranging from 4�0 to 9�0, were prepared,using 1 mol l�1 NaOH and HCl, and the bacteriocins
were treated at different temperatures and time: 121°C for
15 min, 100°C for 10 min and 30 min, 60°C for 10 min
and 30 min and 45°C for 10 and 30 min (Cocolin et al.
2007). The plates were incubated at 37°C for 18–24 h,
and the antimicrobial spectrum of activity was carried out
by measuring the zones of inhibition against indicator
strains B. cereus DSM 2301 and B. subtilis ATCC 6633.
The untreated cell-free supernatants were used as controls.
Each determination was carried out in duplicates.
Partial purification of the bacteriocin
For partial purification of the bacteriocins, the modified
method of Callewaert et al. (1999) and Strompfova and
Laukova (2007) was employed. The cultures were grown
for 18–24 h at 37°C, and the supernatant was obtained
by centrifugation at 10 000 g at 4°C for 15 min. The pH
was adjusted to pH 7�0 with 1 mol l�1 NaOH and the
supernatant cooled in an ice-water bath. Ammonium sul-
fate was added to the supernatant to a final saturation of
60% and held overnight at 4°C with slow and continuous
stirring on a magnetic stirrer. The suspension was centri-
fuged at 5500 g for 30 min at 4°C (Eppendorf centrifuge
5415 R), and the precipitate (surface pellicle) was resus-
pended in 50 mmol l�1 sodium phosphate buffer (pH
6�5). The dissolved precipitate was desalted by dialysis for
24 h at 4°C against 50 mmol l�1 sodium phosphate buf-
fer (pH 6�5) with two changes of buffer over a magnetic
stirrer. The molecular weight cut-off of the membrane
used was 1000 Da. The dialysate was resuspended in ster-
ile Milli Q water and frozen at �20°C until use. The
inhibitory activity of the partially purified bacteriocin was
quantified using a modified method of Yamamoto et al.
(2003) and Prasad et al. (2005). This was performed by
the preparation of 10 ll of twofold serial dilution of the
partially purified bacteriocin in sterile Milli Q water that
was spotted onto the inoculated lawn of the indicator
strain (B. cereus DSM 2301) and incubated at 30°C for 18
–24 h. The bacteriocin titre was expressed as reciprocal of
the highest dilution that exhibited inhibition. The activity
was calculated in arbitrary units per ml (AU ml�1) using
the formula: 1 AU ml�1 = 2n 9 (1000 ll per 10 ll�1),
where AU ml�1 is the arbitrary unit per ml and n is
reciprocal of the highest dilution that gave inhibition.
PCR amplification of the enterocin genes
Genes encoding for the bacteriocins were targeted by
PCR using the primers and conditions described by De
Vuyst et al. (2003) and Cocolin et al. (2007). The specific
primers for enterocins A and B were used in the amplifi-
cation. For enterocin A, the forward primer was made up
of EntFa (5′ GGT ACC ACT CAT AGT GGA AA 3′) andreverse primer EntFb (5′ CCC TGG AAT TGC TCC ACC
TAA 3′). PCR amplification conditions were as follows:
initial denaturation for 5 min at 95°C, 30 cycles of dena-
turation for 30 s at 95°C, annealing at 58°C for 30 s and
extension at 72°C for 30 s and a final elongation step of
5 min at 72°C.For enterocin B, the forward primer was made up of
Ent1 (5′ CAA AAT GTA AAA GAA TTA AGT ACG 3′) andreverse primer Ent2 (5′ AGA GTA TAC ATT TGC TAA
CCC 3′). PCR amplification conditions were as follows:
initial denaturation of 5 min at 95°C, 30 cycles of denatur-ation at 95°C for 30 s, annealing at 56°C for 30 s,
extension at 72°C for 30 s and final extension at 72°C for
5 min.
232 Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology
Enterococcus faecium strains from milk K. Banwo et al.
The PCR products were subjected to electrophoresis
using a 1�8% agarose gel and 19 TBE buffer containing
1 lg ml�1 ethidium bromide (Sambrook et al. 1989).
Results
Identification of the bacteriocin-producing strains
The bacteriocin-producing strains Enterococcus faecium
CM4 and Ent. faecium 2CM1 were characterized to be
Gram-positive, cocci-shaped, catalase-negative bacteria
that did not produce gas from glucose and grew at pH
9�6. Further confirmation was carried out by 16S rRNA
sequencing as stated in the Materials and methods. The
sequences are deposited in the GenBank database under
the following accession numbers JN104687 and
JN104688, respectively.
Acidification
The strain Ent. faecium CM4 reduced the pH to 4�05,while Ent. faecium 2CM1 reduced to pH 3�85 after 48-h
incubation. The initial pH of the medium was 6�54. Thestrains showed strong acidification properties by lowering
the pH of the MRS medium to <5.
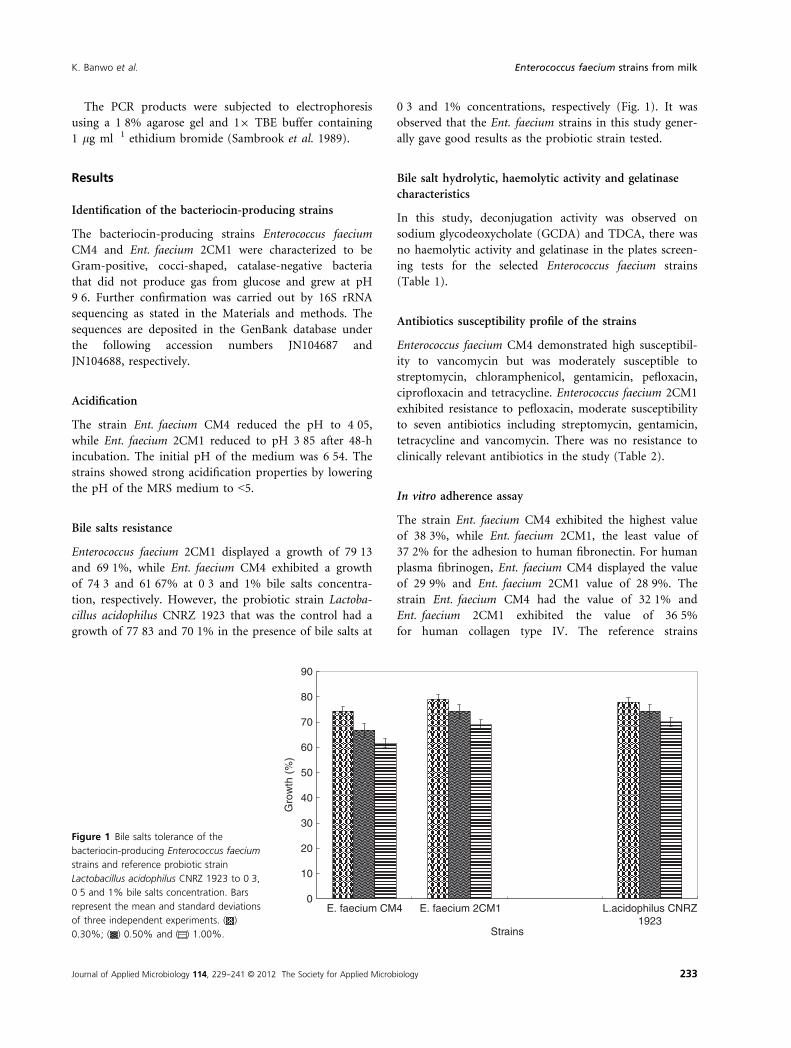
Bile salts resistance
Enterococcus faecium 2CM1 displayed a growth of 79�13and 69�1%, while Ent. faecium CM4 exhibited a growth
of 74�3 and 61�67% at 0�3 and 1% bile salts concentra-
tion, respectively. However, the probiotic strain Lactoba-
cillus acidophilus CNRZ 1923 that was the control had a
growth of 77�83 and 70�1% in the presence of bile salts at
0�3 and 1% concentrations, respectively (Fig. 1). It was
observed that the Ent. faecium strains in this study gener-
ally gave good results as the probiotic strain tested.
Bile salt hydrolytic, haemolytic activity and gelatinase
characteristics
In this study, deconjugation activity was observed on
sodium glycodeoxycholate (GCDA) and TDCA, there was
no haemolytic activity and gelatinase in the plates screen-
ing tests for the selected Enterococcus faecium strains
(Table 1).
Antibiotics susceptibility profile of the strains
Enterococcus faecium CM4 demonstrated high susceptibil-
ity to vancomycin but was moderately susceptible to
streptomycin, chloramphenicol, gentamicin, pefloxacin,
ciprofloxacin and tetracycline. Enterococcus faecium 2CM1
exhibited resistance to pefloxacin, moderate susceptibility
to seven antibiotics including streptomycin, gentamicin,
tetracycline and vancomycin. There was no resistance to
clinically relevant antibiotics in the study (Table 2).
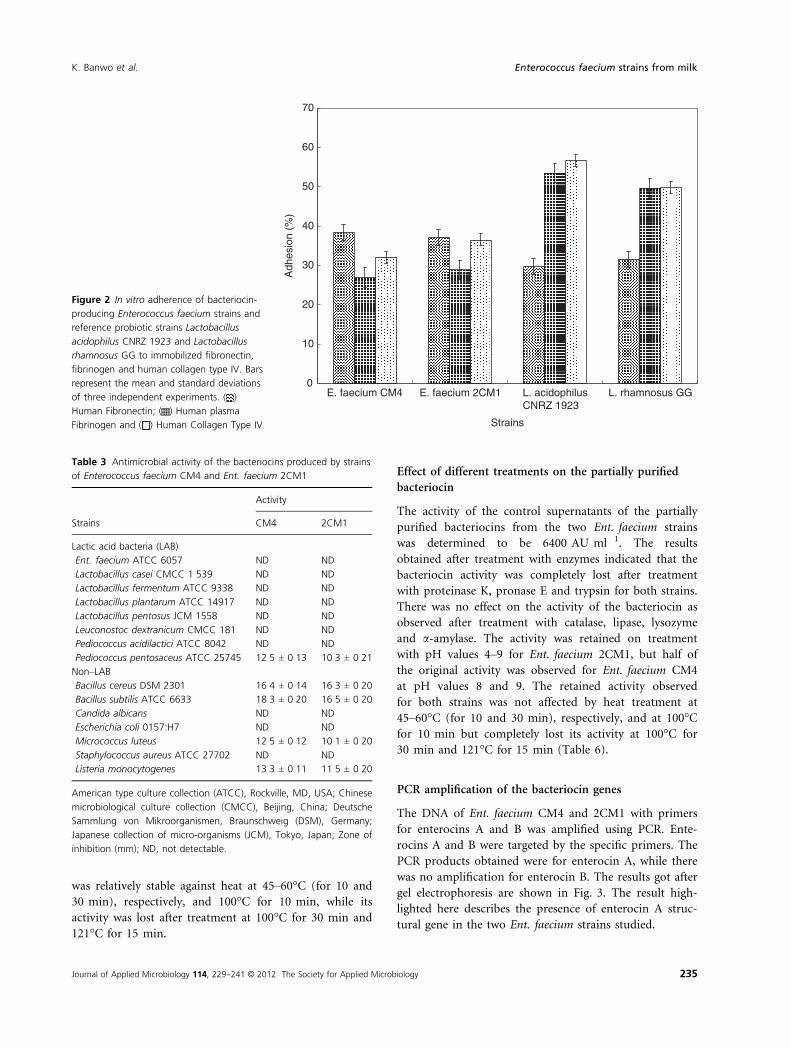
In vitro adherence assay
The strain Ent. faecium CM4 exhibited the highest value
of 38�3%, while Ent. faecium 2CM1, the least value of
37�2% for the adhesion to human fibronectin. For human
plasma fibrinogen, Ent. faecium CM4 displayed the value
of 29�9% and Ent. faecium 2CM1 value of 28�9%. The
strain Ent. faecium CM4 had the value of 32�1% and
Ent. faecium 2CM1 exhibited the value of 36�5%for human collagen type IV. The reference strains
0
10
20
30
40
50
60
70
80
90
L.acidophilus CNRZ1923
E. faecium 2CM1E. faecium CM4
Strains
Gro
wth
(%
)
Figure 1 Bile salts tolerance of the
bacteriocin-producing Enterococcus faecium
strains and reference probiotic strain
Lactobacillus acidophilus CNRZ 1923 to 0�3,0�5 and 1% bile salts concentration. Bars
represent the mean and standard deviations
of three independent experiments. ( )
0.30%; ( ) 0.50% and ( ) 1.00%.
Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology 233
K. Banwo et al. Enterococcus faecium strains from milk

demonstrated higher values than the selected strains for
human plasma fibrinogen and human collagen type IV,
while the Ent. faecium strains exhibited higher value for
human fibronectin (Fig. 2).
Detection of bacteriocin produced by Enterococcus
faecium CM4 and 2CM1 and the antimicrobial
spectrum
The determination of the spectrum of activity of the
bacteriocin produced by the Ent. faecium strains consid-
ered in this study, using LAB and non-LAB as indicator
organisms, is shown in Table 3. Both strains did not
show any antibacterial activity towards the representa-
tive LAB strains Enterococcus faecium ATCC 6057, Ped-
iococcus acidilactici ATCC 8042, Lactobacillus fermentum
ATCC 9338, Lactobacillus plantarum ATCC 14917, Lac-
tobacillus pentosus JCM 1558, Lactobacillus casei CMCC
1.539 and Leuconostoc dextranicum CMCC 181, while
activity was observed against Pediococcus pentosaceus
ATCC 25745. In the non-LAB strains, activity was
observed in Bacillus cereus DSM 2301, Bacillus subtilis
ATCC 6633, Micrococcus luteus and Listeria monocytoge-
nes for both strains.
Effect of enzymes, pH and temperature on bacteriocin
activity
On treatment with enzymes, the cell-free culture super-
natant demonstrated that the inhibitory activity of bac-
teriocin produced by Ent. faecium CM4 and
Ent. faecium 2CM1 was completely eliminated by pro-
teinase K and pronase E, while some activity was
observed to be retained for Ent. faecium CM4 on treat-
ment with trypsin, as shown in Tables 4 and 5. How-
ever, the activity was not affected by treatment with
catalase, pepsin, a-amylase, lysozyme and lipase for both
strains. The inhibitory activity of both Ent. faecium
strains was retained after exposure to a pH range of
4–9. The inhibitory activity of both Ent. faecium strains
Table 1 Bile salt hydrolase (BSH), Haemolytic and gelatinase
activities
Strains
BSH
Haemolysis GelatinaseGCDA TDCA
Enterococcus faecium
CM4
+ + c �
Ent. faecium 2CM1 + + c �
GCDA, glycodeoxycholic acid; TDCA, taurodeoxycholic acid; +: posi-
tive/precipitation of bile salts deconjugation; �/c: negative/no hae-
molysi.
Table
2Antibiotics
suscep
tibility
profile
oftheEn
terococcusfaecium
strains
Antibiotics
Strains
Amoxycillin
Ofloxacin
Streptomycin
Chloramphen
icol
Cefriazone
Gen
tamicin
Pefloxacin
Cotrim
axazole
Ciprofloxacin
Erythromycin
Tetracycline
Van
comycin
Ent.faecium
CM4
+(9)
�++(13)
++(12)
+(5)
++(14)
++(13)
+(5)
++(14)
�++(14)
+++(18)
Ent.faecium
2CM1
+(5)
+(7)
++(11)
++(14)
+(9)
++(14)
�++(13)
++(13)
++(12)
++(13)
++(14)
+++(15–2
0mm):highly
suscep
tible;++(10–1
4mm):moderatelysuscep
tible;+(1–9
mm):suscep
tible;�:
resistan
ce;Su
scep
tibility:presence
ofazoneofinhibition(m
m);Resistance:ab
sence
of
azoneofinhibition(m
m).
234 Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology
Enterococcus faecium strains from milk K. Banwo et al.
was relatively stable against heat at 45–60°C (for 10 and
30 min), respectively, and 100°C for 10 min, while its
activity was lost after treatment at 100°C for 30 min and
121°C for 15 min.
Effect of different treatments on the partially purified
bacteriocin
The activity of the control supernatants of the partially
purified bacteriocins from the two Ent. faecium strains
was determined to be 6400 AU ml�1. The results
obtained after treatment with enzymes indicated that the
bacteriocin activity was completely lost after treatment
with proteinase K, pronase E and trypsin for both strains.
There was no effect on the activity of the bacteriocin as
observed after treatment with catalase, lipase, lysozyme
and a-amylase. The activity was retained on treatment
with pH values 4–9 for Ent. faecium 2CM1, but half of
the original activity was observed for Ent. faecium CM4
at pH values 8 and 9. The retained activity observed
for both strains was not affected by heat treatment at
45–60°C (for 10 and 30 min), respectively, and at 100°Cfor 10 min but completely lost its activity at 100°C for
30 min and 121°C for 15 min (Table 6).
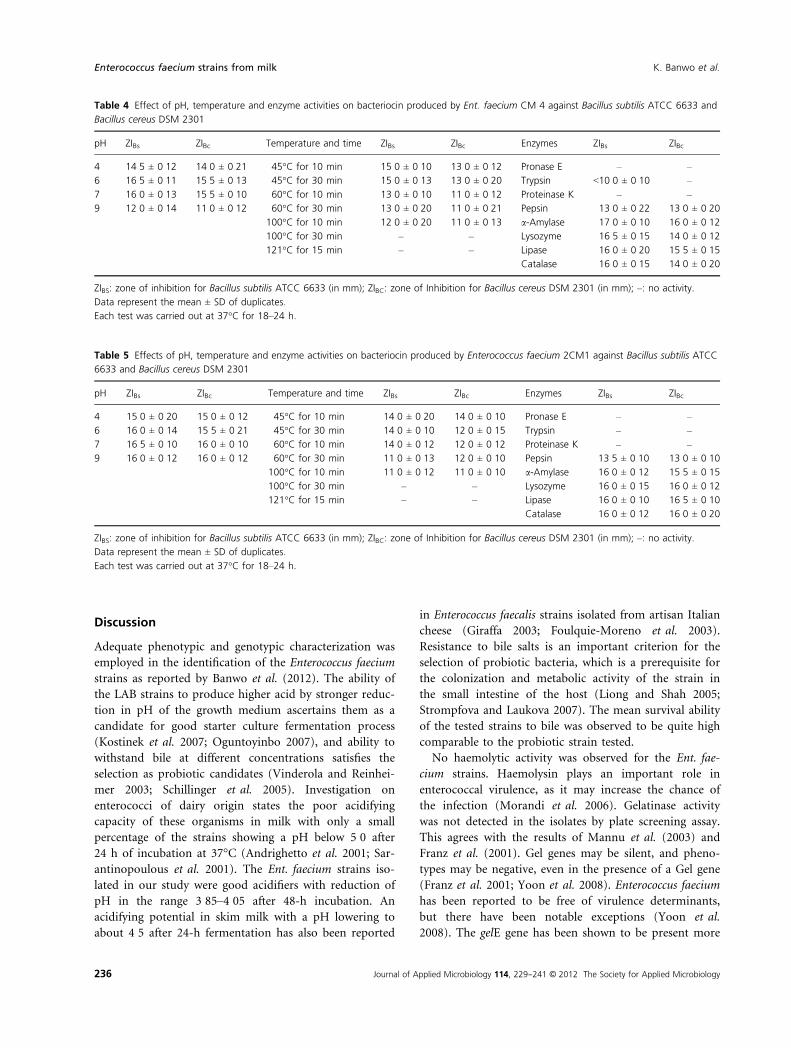
PCR amplification of the bacteriocin genes
The DNA of Ent. faecium CM4 and 2CM1 with primers
for enterocins A and B was amplified using PCR. Ente-
rocins A and B were targeted by the specific primers. The
PCR products obtained were for enterocin A, while there
was no amplification for enterocin B. The results got after
gel electrophoresis are shown in Fig. 3. The result high-
lighted here describes the presence of enterocin A struc-
tural gene in the two Ent. faecium strains studied.
0
10
20
30
40
50
60
70
E. faecium CM4 E. faecium 2CM1 L. acidophilusCNRZ 1923
L. rhamnosus GG
Strains
Adh
esio
n (%
)Figure 2 In vitro adherence of bacteriocin-
producing Enterococcus faecium strains and
reference probiotic strains Lactobacillus
acidophilus CNRZ 1923 and Lactobacillus
rhamnosus GG to immobilized fibronectin,
fibrinogen and human collagen type IV. Bars
represent the mean and standard deviations
of three independent experiments. ( )
Human Fibronectin; ( ) Human plasma
Fibrinogen and ( ) Human Collagen Type IV
Table 3 Antimicrobial activity of the bacteriocins produced by strains
of Enterococcus faecium CM4 and Ent. faecium 2CM1
Strains
Activity
CM4 2CM1
Lactic acid bacteria (LAB)
Ent. faecium ATCC 6057 ND ND
Lactobacillus casei CMCC 1�539 ND ND
Lactobacillus fermentum ATCC 9338 ND ND
Lactobacillus plantarum ATCC 14917 ND ND
Lactobacillus pentosus JCM 1558 ND ND
Leuconostoc dextranicum CMCC 181 ND ND
Pediococcus acidilactici ATCC 8042 ND ND
Pediococcus pentosaceus ATCC 25745 12�5 ± 0�13 10�3 ± 0�21Non–LAB
Bacillus cereus DSM 2301 16�4 ± 0�14 16�3 ± 0�20Bacillus subtilis ATCC 6633 18�3 ± 0�20 16�5 ± 0�20Candida albicans ND ND
Escherichia coli 0157:H7 ND ND
Micrococcus luteus 12�5 ± 0�12 10�1 ± 0�20Staphylococcus aureus ATCC 27702 ND ND
Listeria monocytogenes 13�3 ± 0�11 11�5 ± 0�20
American type culture collection (ATCC), Rockville, MD, USA; Chinese
microbiological culture collection (CMCC), Beijing, China; Deutsche
Sammlung von Mikroorganismen, Braunschweig (DSM), Germany;
Japanese collection of micro-organisms (JCM), Tokyo, Japan; Zone of
inhibition (mm); ND, not detectable.
Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology 235
K. Banwo et al. Enterococcus faecium strains from milk
Discussion
Adequate phenotypic and genotypic characterization was
employed in the identification of the Enterococcus faecium
strains as reported by Banwo et al. (2012). The ability of
the LAB strains to produce higher acid by stronger reduc-
tion in pH of the growth medium ascertains them as a
candidate for good starter culture fermentation process
(Kostinek et al. 2007; Oguntoyinbo 2007), and ability to
withstand bile at different concentrations satisfies the
selection as probiotic candidates (Vinderola and Reinhei-
mer 2003; Schillinger et al. 2005). Investigation on
enterococci of dairy origin states the poor acidifying
capacity of these organisms in milk with only a small
percentage of the strains showing a pH below 5�0 after
24 h of incubation at 37°C (Andrighetto et al. 2001; Sar-
antinopoulous et al. 2001). The Ent. faecium strains iso-
lated in our study were good acidifiers with reduction of
pH in the range 3�85–4�05 after 48-h incubation. An
acidifying potential in skim milk with a pH lowering to
about 4�5 after 24-h fermentation has also been reported
in Enterococcus faecalis strains isolated from artisan Italian
cheese (Giraffa 2003; Foulquie-Moreno et al. 2003).
Resistance to bile salts is an important criterion for the
selection of probiotic bacteria, which is a prerequisite for
the colonization and metabolic activity of the strain in
the small intestine of the host (Liong and Shah 2005;
Strompfova and Laukova 2007). The mean survival ability
of the tested strains to bile was observed to be quite high
comparable to the probiotic strain tested.
No haemolytic activity was observed for the Ent. fae-
cium strains. Haemolysin plays an important role in
enterococcal virulence, as it may increase the chance of
the infection (Morandi et al. 2006). Gelatinase activity
was not detected in the isolates by plate screening assay.
This agrees with the results of Mannu et al. (2003) and
Franz et al. (2001). Gel genes may be silent, and pheno-
types may be negative, even in the presence of a Gel gene
(Franz et al. 2001; Yoon et al. 2008). Enterococcus faecium
has been reported to be free of virulence determinants,
but there have been notable exceptions (Yoon et al.
2008). The gelE gene has been shown to be present more
Table 4 Effect of pH, temperature and enzyme activities on bacteriocin produced by Ent. faecium CM 4 against Bacillus subtilis ATCC 6633 and
Bacillus cereus DSM 2301
pH ZIBs ZIBc Temperature and time ZIBs ZIBc Enzymes ZIBs ZIBc
4 14�5 ± 0�12 14�0 ± 0�21 45°C for 10 min 15�0 ± 0�10 13�0 ± 0�12 Pronase E – –
6 16�5 ± 0�11 15�5 ± 0�13 45°C for 30 min 15�0 ± 0�13 13�0 ± 0�20 Trypsin <10�0 ± 0�10 –
7 16�0 ± 0�13 15�5 ± 0�10 60°C for 10 min 13�0 ± 0�10 11�0 ± 0�12 Proteinase K – –
9 12�0 ± 0�14 11�0 ± 0�12 60°C for 30 min 13�0 ± 0�20 11�0 ± 0�21 Pepsin 13�0 ± 0�22 13�0 ± 0�20100°C for 10 min 12�0 ± 0�20 11�0 ± 0�13 a-Amylase 17�0 ± 0�10 16�0 ± 0�12100°C for 30 min – – Lysozyme 16�5 ± 0�15 14�0 ± 0�12121°C for 15 min – – Lipase 16�0 ± 0�20 15�5 ± 0�15
Catalase 16�0 ± 0�15 14�0 ± 0�20
ZIBS: zone of inhibition for Bacillus subtilis ATCC 6633 (in mm); ZIBC: zone of Inhibition for Bacillus cereus DSM 2301 (in mm); –: no activity.
Data represent the mean ± SD of duplicates.
Each test was carried out at 37°C for 18–24 h.
Table 5 Effects of pH, temperature and enzyme activities on bacteriocin produced by Enterococcus faecium 2CM1 against Bacillus subtilis ATCC
6633 and Bacillus cereus DSM 2301
pH ZIBs ZIBc Temperature and time ZIBs ZIBc Enzymes ZIBs ZIBc
4 15�0 ± 0�20 15�0 ± 0�12 45°C for 10 min 14�0 ± 0�20 14�0 ± 0�10 Pronase E – –
6 16�0 ± 0�14 15�5 ± 0�21 45°C for 30 min 14�0 ± 0�10 12�0 ± 0�15 Trypsin – –
7 16�5 ± 0�10 16�0 ± 0�10 60°C for 10 min 14�0 ± 0�12 12�0 ± 0�12 Proteinase K – –
9 16�0 ± 0�12 16�0 ± 0�12 60°C for 30 min 11�0 ± 0�13 12�0 ± 0�10 Pepsin 13�5 ± 0�10 13�0 ± 0�10100°C for 10 min 11�0 ± 0�12 11�0 ± 0�10 a-Amylase 16�0 ± 0�12 15�5 ± 0�15100°C for 30 min – – Lysozyme 16�0 ± 0�15 16�0 ± 0�12121°C for 15 min – – Lipase 16�0 ± 0�10 16�5 ± 0�10
Catalase 16�0 ± 0�12 16�0 ± 0�20
ZIBS: zone of inhibition for Bacillus subtilis ATCC 6633 (in mm); ZIBC: zone of Inhibition for Bacillus cereus DSM 2301 (in mm); –: no activity.
Data represent the mean ± SD of duplicates.
Each test was carried out at 37°C for 18–24 h.
236 Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology
Enterococcus faecium strains from milk K. Banwo et al.
frequently in clinical isolates than in noninfectious strains
(Yoon et al. 2008). Eaton and Gasson (2001) reported
that Ent. faecium strains isolated from dairy products and
ham could test positive for one or more virulence deter-
minants. This determinant is found at similar frequencies
in the Ent. faecalis and Ent. faecium strains and appears
in all culture types, including isolated food, starter and
medical cultures (Yoon et al. 2008).
Enterococci are well known to be intrinsically resistant
to cephalosporins, b-lactams, sulfonamides and low levels
of clindamycin and aminoglycosides (Yousif et al. 2005).
Multiple resistant strains of enterococci had emerged in
the last decade, which showed resistance to tetracycline,
chloramphenicol and vancomycin (Yousif et al. 2005).
High levels of antibiotics resistance are related to a
combination of nonprescription antibiotic usage and the
circulation of resistant isolates in the environment with
limited sanitation facilities (Al-Jabouri and Al-Meshhada-
ni 1985). According to the findings of Mathur and Singh
(2005), there was a possible baseline for the reference
probiotic strain Ent. faecium 68, used as a probiotic for
man as well as for animal and as silage inoculant. This
strain isolated in the pre-antibiotic era is susceptible to
erythromycin (15 lg), streptomycin/penicillin (streptopen
35 lg), gentamicin (10 lg), chloramphenicol (30 lg) andtetracycline (30 lg). It also possesses intrinsic resistance
to kanamycin (30 lg), streptomycin (10 lg) and oxacillin
(5 lg) (Mathur and Singh 2005).
The Enterococcus strains were moderately sensitive to
streptomycin, gentamicin and ciprofloxacin. Enterococcus
faecium CM4 showed resistance to erythromycin (5 lg),which agreed with the intrinsic resistance characteristics
of reference probiotic strain of Ent. faecium 68. However,
it was observed that Ent. faecium CM4 showed high sus-
ceptibility to vancomycin, while Ent. faecium 2CM1 mod-
erately susceptible to this antibiotic. In another study,
enterococci isolated from Portuguese dairy products such
as milk and cheese were screened for gentamicin resis-
tance (Lopes et al. 2003). Although enterococci are gener-
ally regarded as being intrinsically resistant to low levels
of gentamicin, a high-level gentamicin resistance was
detected in many dairy isolates (Giraffa 2003; Hummel
et al. 2007).
Lactic acid bacteria are reported to bind to ECM such
as human collagen, fibronectin and human fibrinogen,
which may be exposed if the epithelial layer is injured
(Kapczynski et al. 2000; Lorca et al. 2002; Schillinger
et al. 2005). The ECM proteins binding to enterococci
Table 6 Effect of different treatments on the partially purified bacte-
riocin produced by Enterococcus faecium CM4 and Ent. faecium
2CM1
Treatment
Residual activity (AU ml�1)
Ent. faecium CM4 Ent. faecium 2CM1E
Control 6400 6400
Enzymes
Catalase 6400 6400
Trypsin – –
Pepsin 3200 3200
Pronase E – –
Proteinase K – –
Lipase 6400 6400
Lysozyme 6400 6400
a-Amylase 6400 6400
pH
4 6400 6400
5 6400 6400
6 6400 6400
7 6400 6400
8 3200 6400
9 3200 6400
Temperature (min)
45°C (10) 6400 6400
45°C (30) 6400 6400
60°C (10) 6400 6400
60°C (30) 6400 6400
100°C (10) 6400 6400
100°C (30) – –
121°C (15) – –
–: no activity.
Bacillus cereus DSM 2301 was used as the indicator strain.
AU ml�1: Arbitrary Unit per ml defined as reciprocal of highest
dilution showing inhibition of indicator strain 9 100.
Each test was carried out at 37°C for 18–24 h and the residual
bacteriocin activity determined.
Data represent test in duplicates (n = 2).
Figure 3 Polymerase chain reaction (PCR) products obtained from
the amplification using the specific primer for enterocins A and B.
Lane 1: molecular ladder (50 bp); lanes 2 and 3: enterocin A amplifi-
cation for Enterococcus faecium CM4 and Ent. faecium 2CM1; lanes
4 and 5: no amplification for enterocin B for Ent. faecium CM4 and
Ent. faecium, respectively.
Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology 237
K. Banwo et al. Enterococcus faecium strains from milk

could be involved in tissue colonization by enterococci as
shown for other Gram-positive pathogens (Wadstrom
et al. 1987; Kostrzynska et al. 1992). Our results showed
the ability of enterococci to bind to fibronectin at 30% in
contradiction with the findings of Zareba et al. (1997)
who reported that enterococci are nonbinders of rela-
tively low binding capacities of 5% to 7%. The strongest
binding capacities were observed in the human fibronec-
tin, but binding to human collagen type IV and fibrino-
gen was lower for the strains tested. As reported by
Morelli (2000), adherent strains of probiotic bacteria are
more likely to stay longer in the intestinal tract and
therefore may have a better opportunity to exhibit meta-
bolic and immunomodulatory effects than the nonadher-
ing counterparts. The enterococci exhibited sufficient
binding rate for probiotics. Moreover, as reported by
Strompfova and Laukova (2007), probiotic strains that
may not be able to show in vitro and/or in vivo adhesion
characteristics can still show positive effects in the hosts.
In addition, the rate of equality of in vitro and in vivo
capabilities is also questionable.
Numerous strains of enterococci associated with food
systems, mainly Ent. faecium and Ent. faecalis, are capable
of producing a variety of bacteriocin called enterocin
with broad spectrum activity (Franz et al. 1996; Ennahar
et al. 2001; Giraffa 2003). Enterocins usually belong to
class II bacteriocins, which are small, heat stable and
nonlantibiotics (Giraffa 2003). In this study, two Entero-
coccus strains isolated from cow’s milk show antimicro-
bial activity against Gram-positive bacteria, including
Bacillus cereus DSM 2301, Bacillus subtilis ATCC 6633,
Micrococcus luteus, Listeria monocytogenes and Pediococcus
pentosaceus ATCC 25745. The inhibitory activity of the
bacteriocin produced by the Ent. faecium strains com-
prises Gram-positive bacteria including List. monocytoge-
nes but showed limited or no activity against Gram-
negative indicators. This observation is in agreement with
previous workers on enterocins (Cocolin et al. 2007;
Javed et al. 2011). Enterococcal bacteriocins that show
antilisterial activity are of great importance in food and
dairy industries (Rodriguez et al. 2000; De Vuyst et al.
2003). Olasupo et al. (1999) isolated a bacteriocin-pro-
ducing Ent. faecium NA01 strain from wara, a fermented
skimmed cow milk product from West Africa. Enterocins
characterization and screening from Ent. faecium of dif-
ferent origins has been an important area of research, to
enhance the safety of this strain as starter culture for food
products (Foulquie-Moreno et al. 2003; Cocolin et al.
2007).
The effect of pH on enterocin production and appar-
ent degradation was as intense as the effect on bacterial
growth (Foulquie-Moreno et al. 2003). This effect was
similar in our study for Ent. faecium CM4, which
expressed a reduction in activity at pH 8 and 9 and still
had a very good activity at pH 4–7; however, Ent. fae-cium 2CM1 exhibited good activity. This agrees with the
work of Parente and Ricciardi (1999), who reported the
bacteriocin activity for Enterocin 1146, which displayed a
decrease in the early stationary phase at pH values higher
than 4�5. The decrease of activity was ascribed to the
adsorption of bacteriocin molecules on the cell surface of
producer cells; and this depends on the pH of cell envi-
ronment being more pronounced at higher pH (Parente
and Ricciardi 1999; Leroy and De Vuyst 2003; Vizoso-
Pinto et al. 2006). The bacteriocins produced lost its
activity after exposure to heat at 100°C for 30 min but
still had full activity at 100°C for 10 min. This indicates
sensitivity to heat, which was also reported by Du Toit
et al. (2000) and Cocolin et al. (2007) who observed that
Enterocin 1170 was sensitive to heat as its activity
decreased at 80 and 100°C after 30 min, and the activity
was retained after 100 and 60°C for 10 min for Ent. fae-
cium M241 and M249, respectively. pH values affecting
the activity of enterocins produced are in accordance
with those reported by Du Toit et al. (2000). The activity
of Enterococcus over a wide range of pH may be advanta-
geous when produced in the gastrointestinal tract, where
pH levels are known to vary from pH 3 in the stomach
to >pH 7�0 in the large intestine as reported by Du Toit
et al. (2000) and Strompfova and Laukova (2007).
The effect of enzyme was investigated on the bacterio-
cin produced; it was observed that the activity of the
enterocin was completely lost after the action of protein-
ase K and pronase E and trypsin for Ent. faecium 2CM1.
This indicates that the bacteriocin produced is proteina-
ceous in nature. The ability to retain some activity after
treatment with trypsin in Ent. faecium CM4 and pepsin
for both strains agrees with the work of Cocolin et al.
(2007), in which some activity was retained for strains
Ent. faecium M241 and M249 after treatment with pep-
sin. This is in agreement with report of other works on
enterocin production by Ent. faecium strains (Cocolin
et al. 2007; Strompfova and Laukova 2007). However, the
enterocins produced in our study retained its full activity
on treatment with a-amylase, lysozyme, lipase and cata-
lase and reduced activity for pepsin. The neutralized cul-
ture supernatants were not inactivated by lysozyme and
lipase, which suggest that the Enterococcus in this study
do not require a lipid or carbohydrate moiety for activity
as also reported by Du Toit et al. (2000).
The genetic determinants of the bacteriocin genes for
the bacteriocin-producing organisms were targeted for
enterocins. The primer specific for enterocins A and B
were used as described by De Vuyst et al. (2003) and
Cocolin et al. (2007). The PCR product of the expected
size for the enterocin produced by the bacteriocin genes
238 Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology
Enterococcus faecium strains from milk K. Banwo et al.

was observed to be about 140 bp for enterocin A. This is
in agreement with the size observed also by Cocolin et al.
(2007). After amplification, it was confirmed that the
bacteriocin-producing Enterococcus species (Ent. faecium
CM 4 and Ent. faecium 2 CM1) had the presence of
enterocin A but not enterocin B. In the works of Cocolin
et al. (2007), there was the presence of both enterocins A
and B in Ent. faecium M 241 and Ent. faecium M 249.
The structural gene of enterocin A is widely distributed
among Ent. faecium strains, whereas that of enterocin B
always occurs in the presence of enterocin A as reported
by De Vuyst et al. (2003). This implies that there are no
transport genes for enterocin B production as reported
by Franz et al. (1999) in De Vuyst et al. (2003). The
authors suggested that these genes may be lost due to
mutations, which could mean that the structural gene of
enterocin B was lost by several strains, resulting in the
observation that the structural gene of enterocin A occurs
solely in some strains (De Vuyst et al. 2003). The bacte-
riocin genes were observed to be chromosomal because
there was no plasmid isolated from the Ent. faecium
strains (results not shown). This indicates that the genes
for bacteriocin activities are located on the chromosomal
DNA. This is in agreement with the work of De Vuyst
et al. (2003); they further stated that chromosomal locali-
zation of enterocin genes can be an intrinsic characteris-
tic, while plasmid localization is a result of acquired
antibacterial potential through conjugal gene transfer.
The application and importance of bacteriocin-produc-
ing Ent. faecium strains is of immense usefulness in food
and dairy industries (Giraffa 2003; Cocolin et al. 2007).
The use of these strains as starter cultures, cocultures or
protective cultures is becoming an important aspect that
can lead to the production of safer food (Knorr 1998;
Giraffa 2003; Cocolin et al. 2007). The use as a coculture
was addressed by Cocolin et al. (2007) who cocultured
the bacteriocin-producing Ent. faecium M 241 and
Ent. faecium M 249 in skimmed milk with Lact. monocyt-
ogenes. Both enterococci strains studied by the authors
showed a notable inhibition towards List. monocytogenes
with delay in growth of the pathogen (Cocolin et al.
2007). In this study, the enterocins produced by the
strains of Enterococcus faecium showed considerable inhi-
bition against List. monocytogenes. It has also been
reported that enterocin-producing strains of Ent. faecium
is of great potential in dairy technology (Giraffa 2003).
Therefore, enterocins or the producers show a potential
for a diary application as biopreservatives or protective
cultures as reported by Cocolin et al. (2007).
In conclusion, the Ent. faecium strains exhibited a good
probiotic activity because of their tolerance to low pH,
BSH activity, safety and in vitro adherence properties and
the enterocins produced displayed inhibitory activities
against food pathogens and spoilage micro-organisms.
The use of bacteriocin-producing Enterococcus strains as
starter cultures, protective agents in fermented foods and
as probiotics is strain specific as reported by De Vuyst
et al. (2003). The authors also reiterated that it is safer to
use Ent. faecium strains than Ent. faecalis strains. The
bacteriocin produced by the strains demonstrates their
ability to be included in a starter culture for food and
dairy fermentation.
Acknowledgements
The award of a postgraduate fellowship by CAS-TWAS to
Kolawole Banwo for the research period at Institute of
Microbiology, Chinese Academy of Sciences, Beijing,
China, is gratefully acknowledged. The authors appreciate
Dr. Jin Zhong for kindly providing some of the reference
strains used in this study.
References
Al-Jabouri, M.M. and Al-Meshhadani, N.S. (1985) A note on
antibiotic-resistant Escherichia coli in adult man, raw
sewage and sewage polluted River Tigris in Mosul (Iraq).
J Appl Bacteriol 59, 513–518.
Andrighetto, C., Knijff, E., Lombardi, A., Torriani, S.,
Vancanneyt, M., Kersters, K., Swings, J. and Dellaglio, F.
(2001) Phenotypic and genetic diversity of enterococci
isolated from Italian Cheeses. J Dairy Res 68, 303–
316.
Banwo, K., Sanni, A.I., Tan, H.R. and Tian, Y.Q. (2012)
Phenotypic and genotypic characterization of lactic acid
bacteria isolated from some Nigerian traditional fermented
foods. Food Biotechnol, 26(2), 124–142.
Ben-Omar, N., Castro, A., Lucas, R., Abriouel, H., Yousif, N.
M.K., Franz, C.M.A.P., Holzapfel, W.H., Perez-Pulido, R.
et al. (2004) Functional and safety aspects of enterococci
isolated from different Spanish foods. Syst Appl Microbiol
27, 118–130.
Brosius, J., Palmer, M.L., Kennedy, P.J. and Noller, H.F.
(1978) Complete nucleotide sequence of a 16S ribosomal
RNA gene from Escherichia coli. Proc Natl Acad Sc USA
75, 4801–4805.
Callewaert, R., Holo, H., Devreese, B., Van Beeumen, J., Nes,
I. and De Vuyst, L. (1999) Characterization and
production of amylovorin L471, a bacteriocin purified
from Lactobacillus amylovorus DCE 471 by a novel three-
step method. Microbiol 145, 2559–2568.
Cocolin, L., Foschino, R., Comi, G.M. and Fortina, G. (2007)
Description of the bacteriocins produced by two strains of
Enterococcus faecium isolated from Italian goat milk. Food
Microbiol 24, 752–758.
De Vuyst, L., Foulquie Moreno, M.R. and Revets, H. (2003)
Screening for enterocins and detection of hemolysin and
Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology 239
K. Banwo et al. Enterococcus faecium strains from milk

vancomycin resistance in enterococci of different origins.
Int J Food Microbiol 84, 299–318.
Du Toit, M., Franz, C.M.A.P., Dicks, L.M.T. and Holzapfel,
W.H. (2000) Preliminary characterization of bacteriocins
produced by Enterococcus faecium and Enterococcus faecalis
isolated from pig faeces. J Appl Microbiol, 88, 482–494.
Eaton, T.J. and Gasson, M.J. (2001) Molecular screening of
enterococcus virulence determinants and potential for
genetic exchange between food and medical isolates. Appl
Environ Microbiol, 67, 1628–1635.
Eka, O.U. and Ohaba, J.A. (1977) Microbiological examination
of Fulani milk (nono) and butter (man shanu). Nig J Sci,
11, 113–122.
Ennahar, S., Sashihara, T., Sonomoto, K. and Isahizaki, A.
(2001) Class IIa bacteriocins; biosynthesis, structure and
activity. FEMS Microbiol Rev 24, 85–106.
Foulquie-Moreno, M.R., Rea, M.C., Cogan, T.M. and De
Vuyst, L. (2003) Applicability of a bacteriocin-producing
Enterococcus faecium as co-culture in Cheddar cheese
manufacture. Int J Food Microbiol 81, 73–84.
Franz, C.M.A.P., Schillinger, U. and Holzapfel, W.H. (1996)
Production and characterization of enterocin 900, a
bacteriocin produced by Enterococcus faecium BFE 900
from black olives. Int J Food Microbiol 29, 255–270.
Franz, C.M.A.P., Worobo, R.W., Quadri, L.E.N., Schillinger,
U., Holzapfel, W.H., Vedras, J.C. and Stiles, M.E. (1999)
Atypical genetic locus associated with constitutive
production of enterotoxin B by Enterococcus faecium BFE
900. Appl Environ Microbiol 65, 2170–2178.
Franz, C.M.A.P., Muscholl-Silberhorn, A.B., Yousif, N.M.K.,
Vancanneyt, M., Swings, J. and Holzapfel, W.H. (2001)
Incidence of virulence factors and antibiotic resistance
among enterococci isolated from food. Appl Environ
Microbiol, 6, 4385–4389.
Fuller, R. (1989) A review: probiotics in man and animals. J
Appl Bacteriol 66, 365–378.
Giraffa, G. (2003) Functionality of enterococci in dairy
products. Int J Food Microbiol 88, 215–222.
Hummel, A.S., Holzapfel, W.H. and Franz, C.M.A.P. (2007)
Characterisation and transfer of antibiotic resistance genes
from enterococci isolated from food. Syst Appl Microbiol
30, 1–7.
Javed, I., Ali, M.I., Ahmed, B., Ghumro, P.B., Abdul, H. and
Safia, A. (2011) Optimization and partial purification of
bacteriocins from Enterococcus spp indigenous to Pakistan.
Food Biotechnol 25, 130–139.
Johnston, L.M. and Jaykus, L. (2004) Antimicrobial resistance
to Enterococcus species isolated from produce. Appl
Environ Microbiol, 70, 3133–3137.
Kapczynski, D.R., Meinersmann, R.J. and Lee, M.D. (2000)
Adherence of Lactobacillus to intestinal 407 cells in culture
correlates with fibronectin binding. Curr Microbiol 41,
136–141.
Kayser, F.H. (2003) Safety aspects of enterococci from the
medical point of view. Int J Food Microbiol, 88, 255–262.
Klaenhammer, T.R. (1993) Genetics of bacteriocins produced
by lactic acid bacteria. FEMS Microbiol Rev 12, 39–86.
Knorr, D. (1998) Technological aspects related to
microorganisms in functional foods. Trends Food Sc
Technol 9, 295–306.
Kostinek, M., Specht, I., Edward, V.A., Schillinger, U., Hertel,
C., Holzapfel, W.H. and Franz, C.M.A.P. (2005) Diversity
and technological properties of predominant lactic acid
bacteria from fermented cassava used for the preparation
of Gari, a traditional African food. Syst Appl Microbiol 28,
527–540.
Kostinek, M., Specht, I., Edward, V.A., Pinto, C., Egounlety,
M., Sossa, C., Mbugua, S., Dorti, C. et al. (2007)
Characterization and biochemical properties of
predominant lactic acid bacteria from fermenting cassava
for selection as starter cultures. Int J Food Microbiol, 114,
342–351.
Kostrzynska, M., Paulsson, M., Schmidt, K.H. and Wadstrom,
T. (1992) Comparative studies on binding of vitronectin
and fibronectin to groups A and C streptococci. Microbios
71, 179–192.
Leroy, F. and De Vuyst, L. (2003) A combined model to
predict the functionality of the bacteriocin-producing
Lactobacillus sakei strain CTC494. Appl Environ Microbiol
69, 1093–1099.
Liong, M.T. and Shah, N.P. (2005) Acid, bile tolerance and
cholesterol removal ability of lactobacilli strains. J Dairy Sc
88, 55–66.
Lopes, M.F.S., Ribeiro, T., Martins, M.P., Tenreiro, R. and
Crespo, M.T.B. (2003) Gentamicin resistance in dairy and
clinical Enterococcal isolates and in reference strains. J Ant
Chem 52, 214–219.
Lorca, G., Torino, M.I., Font de Valdez, G. and Ljungh, A.
(2002) Lactobacilli express cell surface proteins which
mediate binding of immobilized collagen and Fibronectin.
FEMS Microbiol Lett 206, 31–37.
Mannu, L., Paba, A., Daga, E., Comunian, R., Zanetti, S.,
Dupre, I. and Sechi, L.A. (2003) Comparison of the
incidence of virulence determinants and antibiotic
resistance between Enterococcus faecium strains of dairy,
animal and clinical origin. Int J Food Microbiol, 88,
291–304.
Mathara, J.M., Schillinger, U., Kutima, P.M., Mbugua, S.K.,
Guigas, C., Franz, C. and Holzapfel, W.H. (2008)
Functional properties of Lactobacillus plantarum strains
isolated from Maasai Traditional Fermented Milk
Products in Kenya. Curr Microbiol 56, 315–321.
Mathur, S. and Singh, R. (2005) Antibiotic resistance in food
lactic acid bacteria – a review. Int J Food Microbiol 105,
281–295.
Minelli, E.B., Benini, A., Marzotto, M., Sbarbati, A.,
Ruzzenente, O., Ferrario, R. and Hendriks, H. (2004)
Assessment of novel probiotic Lactobacillus casei strains
for the production of functional dairy foods. Int Dairy J,
14, 723–736.
240 Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology
Enterococcus faecium strains from milk K. Banwo et al.

Morandi, S., Brasca, M., Andrighetto, C., Lombardi, A. and
Lodi, R. (2006) Technological and molecular
characterization of enterococci isolated from north-west
Italian dairy products. Int Dairy J, 16, 867–875.
Morelli, L. (2000) In vitro selection of probiotic Lactobacilli: a
critical appraisal. Curr Issues Intestinal Microbiol 1(2),
59–67.
Mueller, J.H. and Hinton, J. (1941) A protein-free medium for
primary isolation of the Gonococcus and Meningococcus.
Proc Soc Exp Biol Med 48, 330–333.
National Committee for Clinical Laboratory Standards (1990)
Methods of Antimicrobial Susceptibility Testing of Anaerobic
Bacteria, 2nd edn. Approved standard MII-AZ.
Philadelphia, PA, Villanova, USA: National Committee for
Clinical Laboratory Standards.
Oguntoyinbo, F.A. (2007) Identification and functional
properties of dominant lactic acid bacteria isolated at
different stages of solid state fermentation of cassava
during Gari production. W J Microbiol Biotechnol, 23,
1425–1432.
Olasupo, N.A., Schillinger, U., Narbad, A., Dodd, H. and
Holzapfel, W.H. (1999) Occurrence of nisin Z production
in Lactococcus lactis BFE 1500 isolated from wara, a
traditional Nigerian cheese product. Int J Food Microbiol
53, 141–152.
Parente, E. and Ricciardi, A. (1999) Production, recovery and
purification of bacteriocins from lactic acid bacteria. Appl
Microbiol Biotechnol 52, 628–638.
Park, S.H., Itoh, K. and Fujisawa, T. (2003) Characteristics
and identification of enterocins produced by Enterococcus
faecium JCM 5804. J Appl Microbiol, 95, 294–300.
Prasad, S., Morris, P., Hansen, R., Meaden, P.G. and Austin,
B. (2005) A novel bacteriocin-like substance (BLIS) from a
pathogenic strain of Vibrio harveyi. Microbiol 151,
3051–3058.
Rodriguez, E., Gonzalez, B., Gaya, P., Nunez, M. and Medina,
M. (2000) Diversity of bacteriocins produced by lactic
acid bacteria isolated from raw milk. Int Dairy J 10, 7–15.
Sambrook, J.E., Eritsch, F. and Maniatis, J. (1989) Molecular
Cloning: A Laboratory Manual, 2nd edn. Cold spring
harbor, NY: Cold spring habour Laboratory Press.
Sarantinopoulous, P., Kalantzopoulous, G. and Tsakalodou, E.
(2001) Citrate metabolism by Enterococcus faecalis FAIR-
E229. Appl Environ Microbiol 67, 5482–5487.
Schillinger, U. and Lucke, F.K. (1989) Antibacterial activity of
Lactobacillus sake isolated from meat. Appl Environ
Microbiol 55, 1901–1906.
Schillinger, U., Guigas, C. and Holzapfel, W.H. (2005) In vitro
adherence and other properties of lactobacilli used in
probiotic yoghurt-like products. Int Dairy J 15,
1289–1297.
Strompfova, V. and Laukova, A. (2007) In vitro study on
bacteriocin production of enterococci associated with
chickens. Anaerobe, 13, 228–237.
Todorov, S.D. and Dicks, L.M.T. (2006) Screening for
bacteriocin producing lactic acid bacteria from boza, a
traditional cereal beverage from Bulgaria. Comparison of
the bacteriocins. Proc Biochem 41, 11–19.
Villani, F. and Coppola, S. (1994) Selection of enterococcal
strains for water-buffalo Mozzarella cheese manufacture.
Ann Rev Microbial Enzymol 44, 97–105.
Vinderola, C.G. and Reinheimer, J.A. (2003) Lactic acid starter
and probiotic bacteria, a comparative ‘in vitro’ study of
probiotic characteristics and biological barrier resistance.
Food Res Int 36, 895–904.
Vizoso-Pinto, M.G., Franz, C.M.A.P., Schillinger, U. and
Holzapfel, W.H. (2006) Lactobacillus spp. with in vitro
probiotic properties from human faeces and traditional
fermented products. Int J Food Microbiol 109, 205–214.
Wadstrom, T., Andersson, K., Sydow, M., Axelsson, L.,
Lindgren, S. and Gullmar, B. (1987) Surface properties of
lactobacilli isolated from the small intestine of pigs. J Appl
Bacteriol 62, 513–520.
Walker, D.K. and Gilliland, S.E. (1993) Relationships among
bile tolerance, bile salt deconjugation and assimilation of
cholesterol by Lactobacillus acidophilus. J Dairy Sc 76,
956–961.
Yamamoto, Y., Togawa, Y., Shimosaka, M. and Okazaki, M.
(2003) Purification and characterization of a novel
bacteriocin produced by Enterococcus faecalis strain RJ-11.
Appl Environ Microbiol 69, 5746–5753.
Yoon, M.Y., Kim, Y.J. and Hwang, H.J. (2008) Properties and
safety aspects of Enterococcus faecium strains isolated from
Chungkukjang, a fermented soy product. LWT, 41,
925–933.
Yousif, N.M.K., Dawyndt, P., Abriouel, H., Wijaya, A.,
Schillinger, U., Vancanneyt, M., Swings, J., Dirar, H.A.
et al. (2005) Molecular characterization, technological
properties and safety aspects of enterococci from
‘Hussuwa’, an African fermented sorghum product. J Appl
Microbiol 98, 216–228.
Zareba, T.W., Pascu, C., Hryniewicz, W. and Wadstrom, T.
(1997) Binding of extracellular matrix proteins by
enterococci. Curr Microbiol 34, 6–11.
Journal of Applied Microbiology 114, 229--241 © 2012 The Society for Applied Microbiology 241
K. Banwo et al. Enterococcus faecium strains from milk




















