
Nutritional indices of juvenile Caribbean spiny lobsters in a Mexican reef lagoon: Are changes over...
7
Nutritional indices of juvenile Caribbean spiny lobsters in a Mexican reef lagoon: Are changes over a 10-year span related to the emergence of Panulirus argus Virus 1 (PaV1)? Patricia Briones-Fourzán ⁎, Karla Baeza-Martínez, Enrique Lozano-Álvarez Universidad Nacional Autónoma de México, Instituto de Ciencias del Mar y Limnología, Unidad Académica Puerto Morelos. Prol. Av. Niños Héroes s/n, Domicilio Conocido, Puerto Morelos, Quintana Roo, 77580 México abstract article info Article history: Received 30 July 2008 Received in revised form 27 November 2008 Accepted 3 December 2008 Keywords: Artificial shelters Casitas Disease Nutritional condition PaV1 Starvation We compared two indices of nutritional condition, the relative weight of the digestive gland (RWDG) and the ratio of weight to carapace length (weight/CL ratio), of the first three benthic phases, algal juveniles (7– 25 mm CL), postalgal juveniles (25–45 mm CL), and subadults (45–79 mm CL) of Caribbean spiny lobster (Panulirus argus) dwelling in a shelter-poor reef lagoon before (1995) and after (2005) deployment of large artificial shelters (“casitas”) in 1998. To provide an experimental baseline, in 2005 we also conducted a laboratory experiment to assess the effect of starvation on both nutritional indices. Through 25-day trials, starvation significantly decreased the mean RWDG of all three juvenile phases, but did not affect their weight/CL ratios. In the field, casitas significantly enhanced juvenile lobsters relative to control sites and to prior values, but between 1995 and 2005 there was a significant decrease in condition of algal juveniles (both indices) and postalgal juveniles (only the RWDG), but not of subadults. These results would appear to counter previous findings that, for P. argus, food is not limiting and nutritional condition is not density-dependent. However, in 2000, a new pathogenic virus that has a distinct predilection for small juveniles, Panulirus argus Virus 1 (PaV1), emerged in the reef lagoon. Among other effects, this lethal virus causes a marked atrophy of the digestive gland. Through the laboratory trials and in the field, prevalence of visibly diseased individuals was highest in algal juveniles and lowest in subadults. These individuals were excluded from statistical analyses, but the disease progression may affect the digestive gland progressively given that newly infected lobsters take several weeks to become visibly diseased. Up to 2006, PaV1 had not significantly impacted lobster density on casita sites, but because P. argus is highly gregarious and transmission of PaV1 appears to be mainly by contact, futher investigation is required. These issues are relevant to enhancement of spiny lobsters and assessment of ecological impacts of marine diseases. © 2009 Elsevier B.V. All rights reserved. 1. Introduction Understanding factors that regulate the abundance of decapod crustaceans of economic or ecological consequence is of interest to fisheries biologists and ecologists. For example, nutritional condition — the extent to which an animal has accumulated reserves to allow normal physiological function and growth (Moore et al., 2000)— can affect survival, behavior, growth rates, and fecundity of crustaceans (Thomas et al., 2003; Wahle and Fogarty, 2006). Nutritional condition is most obviously affected by starvation, but also by availability of other vital resources (e.g., shelter) and diseases (Pollock et al., 1997; McGarvey et al., 1999; Behringer and Butler, 2006). In crustaceans, changes in nutritional condition are not reflected in visible external changes because their length remains fixed between successive molts (Dall, 1974), but their condition can be assessed through several indices. Some nutritional indices focus on changes in concentrations of metabolic compounds in different body tissues (e.g., Wang and Stickle, 1986; Musgrove, 2001; Parslow-Williams et al., 2001) or on changes in the hemolymph refractive index, which reflects protein concentration in the hemolymph (Moore et al., 2000; Oliver and MacDiarmid, 2001; Behringer and Butler, 2006), whereas others are based on morphometric measures. For example, the hepa- tosomatic index or relative weight of the digestive gland (RWDG) (weight of the digestive gland (= hepatopancreas)/body weight×100) has been used to assess nutritional condition in freshwater crayfish (McClain, 1995; Mannonen and Henttonnen, 1995; Lucic and Erben, 2006), snow crabs (Godbout et al., 2002), American lobsters (Stewart et al., 1967), and spiny lobsters (Dall, 1974; Lozano-Álvarez and Aramoni-Serrano, 1996; Tsvetnenko et al., 1999; Briones-Fourzán et al., 2003; Castañeda-Fernández de Lara et al., 2005). More recently, the ratio of body weight to carapace length (weight/CL ratio) has also been used to assess nutritional condition in spiny lobsters (Robertson Journal of Experimental Marine Biology and Ecology 370 (2009) 82–88 ⁎ Corresponding author. Unidad Académica Puerto Morelos, ICML-UNAM, P.O. Box 1152, Cancún, Q.R. 77500 México. Tel.: +52 998 8710367x151; fax: +52 998 8710138. E-mail address: [email protected] (P. Briones-Fourzán). 0022-0981/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.jembe.2008.12.004 Contents lists available at ScienceDirect Journal of Experimental Marine Biology and Ecology journal homepage: www.elsevier.com/locate/jembe
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Nutritional indices of juvenile Caribbean spiny lobsters in a Mexican reef lagoon: Are changes over...

Journal of Experimental Marine Biology and Ecology 370 (2009) 82–88
Contents lists available at ScienceDirect
Journal of Experimental Marine Biology and Ecology
j ourna l homepage: www.e lsev ie r.com/ locate / jembe
Nutritional indices of juvenile Caribbean spiny lobsters in a Mexican reeflagoon: Are changes over a 10-year span related to the emergenceof Panulirus argus Virus 1 (PaV1)?
Patricia Briones-Fourzán ⁎, Karla Baeza-Martínez, Enrique Lozano-ÁlvarezUniversidad Nacional Autónoma de México, Instituto de Ciencias del Mar y Limnología, Unidad Académica Puerto Morelos. Prol. Av. Niños Héroes s/n, Domicilio Conocido,Puerto Morelos, Quintana Roo, 77580 México
⁎ Corresponding author. Unidad Académica Puerto M1152, Cancún, Q.R. 77500 México. Tel.: +52 998 871 036
E-mail address: [email protected] (P. Brio
0022-0981/$ – see front matter © 2009 Elsevier B.V. Adoi:10.1016/j.jembe.2008.12.004
a b s t r a c t
a r t i c l e i n f oArticle history:
We compared two indices o Received 30 July 2008Received in revised form 27 November 2008Accepted 3 December 2008Keywords:Artificial sheltersCasitasDiseaseNutritional conditionPaV1Starvation
f nutritional condition, the relative weight of the digestive gland (RWDG) and theratio of weight to carapace length (weight/CL ratio), of the first three benthic phases, algal juveniles (7–25 mm CL), postalgal juveniles (25–45 mm CL), and subadults (45–79 mm CL) of Caribbean spiny lobster(Panulirus argus) dwelling in a shelter-poor reef lagoon before (1995) and after (2005) deployment of largeartificial shelters (“casitas”) in 1998. To provide an experimental baseline, in 2005 we also conducted alaboratory experiment to assess the effect of starvation on both nutritional indices. Through 25-day trials,starvation significantly decreased the mean RWDG of all three juvenile phases, but did not affect theirweight/CL ratios. In the field, casitas significantly enhanced juvenile lobsters relative to control sites and toprior values, but between 1995 and 2005 there was a significant decrease in condition of algal juveniles (bothindices) and postalgal juveniles (only the RWDG), but not of subadults. These results would appear to counterprevious findings that, for P. argus, food is not limiting and nutritional condition is not density-dependent.However, in 2000, a new pathogenic virus that has a distinct predilection for small juveniles, Panulirus argusVirus 1 (PaV1), emerged in the reef lagoon. Among other effects, this lethal virus causes a marked atrophy ofthe digestive gland. Through the laboratory trials and in the field, prevalence of visibly diseased individualswas highest in algal juveniles and lowest in subadults. These individuals were excluded from statisticalanalyses, but the disease progression may affect the digestive gland progressively given that newly infectedlobsters take several weeks to become visibly diseased. Up to 2006, PaV1 had not significantly impactedlobster density on casita sites, but because P. argus is highly gregarious and transmission of PaV1 appears tobe mainly by contact, futher investigation is required. These issues are relevant to enhancement of spinylobsters and assessment of ecological impacts of marine diseases.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Understanding factors that regulate the abundance of decapodcrustaceans of economic or ecological consequence is of interest tofisheries biologists and ecologists. For example, nutritional condition—
the extent to which an animal has accumulated reserves to allownormal physiological function and growth (Moore et al., 2000)— canaffect survival, behavior, growth rates, and fecundity of crustaceans(Thomas et al., 2003; Wahle and Fogarty, 2006). Nutritional conditionis most obviously affected by starvation, but also by availability ofother vital resources (e.g., shelter) and diseases (Pollock et al., 1997;McGarvey et al., 1999; Behringer and Butler, 2006).
In crustaceans, changes in nutritional condition are not reflected invisible external changes because their length remains fixed between
orelos, ICML-UNAM, P.O. Box7x151; fax: +52 998 871 0138.nes-Fourzán).
ll rights reserved.
successive molts (Dall, 1974), but their condition can be assessedthrough several indices. Some nutritional indices focus on changes inconcentrations of metabolic compounds in different body tissues (e.g.,Wang and Stickle, 1986; Musgrove, 2001; Parslow-Williams et al.,2001) or on changes in the hemolymph refractive index, whichreflects protein concentration in the hemolymph (Moore et al., 2000;Oliver and MacDiarmid, 2001; Behringer and Butler, 2006), whereasothers are based on morphometric measures. For example, the hepa-tosomatic index or relative weight of the digestive gland (RWDG)(weight of the digestive gland (= hepatopancreas)/body weight×100)has been used to assess nutritional condition in freshwater crayfish(McClain, 1995; Mannonen and Henttonnen, 1995; Lucic and Erben,2006), snow crabs (Godbout et al., 2002), American lobsters (Stewartet al., 1967), and spiny lobsters (Dall, 1974; Lozano-Álvarez andAramoni-Serrano, 1996; Tsvetnenko et al., 1999; Briones-Fourzánet al., 2003; Castañeda-Fernández de Lara et al., 2005). More recently,the ratio of body weight to carapace length (weight/CL ratio) has alsobeen used to assess nutritional condition in spiny lobsters (Robertson

83P. Briones-Fourzán et al. / Journal of Experimental Marine Biology and Ecology 370 (2009) 82–88
et al., 2000; Oliver and MacDiarmid, 2001; Behringer and Butler,2006).
In species that have wide foraging ranges and/or undergoontogenetic habitat shifts, such as the Caribbean spiny lobster, Panu-lirus argus (Latreille, 1804), differing foraging returns and energyexpenditure in different habitats may affect nutritional condition.Panulirus argus is a major fisheries resource across the greater Carib-bean region (Lipcius and Eggleston, 2000) that has a complex lifehistory (reviewed by Butler and Herrnkind, 2000). Briefly, following a5 to 9 month-long pelagic larval phase, postlarvae of P. argus settlein shallow, vegetated habitats, where the solitary, widely dispersedearly juveniles (“algal juveniles”, 5 to 15–25 mm carapace length, CL)dwell. A few months later, the “postalgal juveniles” (15–25 mm CLto 35–45 mm CL) emerge from the vegetation to seek daytime refugein crevices within or adjacent to the vegetated habitats (e.g., solu-tion holes and crevices, rocky outcrops, small coral heads, and thebases of large sponges and soft corals) and become socially gregarious.Subsequent habitat shifts include a subadult (35–45 mm to 75–80 mm CL) shift to patch reefs and shallow coral reef habitats, fol-lowed by an eventual shift to deeper reefs by the adults (75–80 toN200 mm CL).
All benthic phases of P. argus feed upon a wide array of smallinvertebrates (Herrnkind et al., 1975; Colinas-Sánchez and Briones-Fourzán, 1990; Herrera et al., 1991; Briones-Fourzán et al., 2003).Lobsters emerge from their shelters at dusk and disperse to foragesolitarily across seagrass and rubble beds (Cox et al., 1997). Foragingdistances increase from b1m (algal juveniles) to a fewmeters (postalgaljuveniles) to several tens or hundreds of meters (subadults-adults)(Herrnkind et al., 1975; Butler and Herrnkind, 2000; Lozano-Álvarezet al., 2003). Because this foraging behavior, in conjunction with thescattered prey assemblage in tropical habitats, prevents the depressionof prey abundance (Behringer and Butler, 2006; Nizinski, 2007), food isusually not limiting for P. argus (Andrée, 1981; Marx and Herrnkind,1985; Butler et al.,1997; Briones-Fourzán et al., 2003). Crevice shelter, bycontrast, is an important limiting resource, especially for juveniles,which are more vulnerable to predation (Butler and Herrnkind, 2000).
For example, the reef lagoon at Puerto Morelos, Mexico (centeredat 20° 51′ N and 86° 51.5′ W), is covered by a well-developed seagrasscommunity that harbors abundant algal juveniles of Panulirus argus(Briones-Fourzán and Lozano-Álvarez, 2001a), but is remarkably poorin crevice shelter, thus causing a demographic bottleneck for furthercrevice-dwelling phases (Briones-Fourzán and Lozano-Álvarez, 2001b).These features allowed for a controlled field experiment to test wheth-er the deployment of large artificial crevice shelters (“casitas”, whichprovidea shelterareaof ~ 1m2) could help relieve the local demographicbottleneck. Casitas were chosen because P. argus lobsters fare betterwhere available crevices allow for gregariousness, which increases per-capita survival rates (Eggleston and Lipcius,1992; Briones-Fourzán et al.,2007). In 1995, prior to the casita experiment, Briones-Fourzán et al.(2003) studied the feeding ecology of local juveniles of P. argus. Aftercontrolling for molt stage (postmolt individuals had lower RWDGs),they found that sex, sampling site, and season did not affect the RWDG,but that subadults had a significantly lower mean RWDG than algaland postalgal juveniles. These results were attributed to the paucity ofsuitable crevice shelter for subadults, which could prevent these largerindividuals from efficiently foraging across the reef lagoon. Analternative hypothesis, however, is that the RWDG may change withontogeny.
Following deployment of casitas (July 1998) and up to November2002, the density, biomass, survival, and persistence of lobstersincreased significantly in sites with casitas (“casita sites”) relative tocontrol sites and to previous values (Briones-Fourzán et al., 2007).However, during 2000, a new pathogenic viral disease emerged inthe lobster population (Huchin-Mian et al., 2008). This disease, causedby an unclassified virus named Panulirus argus Virus 1, or PaV1, wasfirst discovered in 1999 in juveniles of P. argus from Florida Bay, USA
(Shields and Behringer, 2004). PaV1 shows a distinct predilection forsmall juveniles (Behringer et al., 2006; Shields et al., 2006; Lozano-Álvarez et al., 2008) and appears to be transmitted mainly by contact(Butler et al., 2008).
After November 2002, the casita sites, but not the control sites,were maintained for other research purposes. Lozano-Álvarez et al.(2008) found that, despite a significant increase in the averageprevalence of PaV1 on casita sites between 2001 and 2006, lobsterdensity had not been significantly impacted, but remarked that thedisease might be affecting lobsters in other measures, including theirnutritional condition. Indeed, Behringer et al. (2008) reported that, inFlorida Bay, visibly diseased Panulirus argus lobsters had a significantlylower hemoplymph refractive index than visibly healthy lobsters.Because lobsters with overt PaV1 infections undergo a markedatrophy of the the digestive gland (Shields and Behringer, 2004), thedisease would also be expected to affect the RWDG of lobsters.
In 2005, we collected algal juveniles, postalgal juveniles, andsubadults of Panulirus argus from casita sites to compare their meanRWDGs with those estimated during 1995. If paucity of shelter forsubadults was the main factor underlying the pattern observed in1995 (lower RWDG in subadults, higher in algal and postalgal juve-niles), we would expect an increase in the RWDG of subadult lobstersrelative to algal and postalgal juveniles by 2005 due to the mitigatingeffect of casitas. If the pattern reflects an ontogenetic trait, we wouldexpect to find a similar pattern in both years. But if PaV1 affects theRWDG of lobsters, we would expect the more vulnerable algal andpostalgal juveniles, but not the less vulnerable subadults, to showlower RWDGs in 2005. Our data bases also allowed us to compare theweight/CL ratios of lobsters between 1995 and 2005. These ratios,however, have been used in the laboratory to test the effects of dif-ferent feeding regimes on condition of lobsters over relatively narrowsize ranges (e.g., Robertson et al., 2000; Oliver and MacDiarmid, 2001;Behringer and Butler, 2006), but not to assess and compare conditionof wild lobsters across ontogenetic phases. Therefore, to provide anexperimental baseline, we tested the influence of starvation (themost obvious factor potentially affecting condition) on both the RWDGand the weight/CL ratios of lobsters in the three juvenile phases inlaboratory trials.
2. Methods
2.1. Experiment to assess the influence of starvation onnutritional indices
The experiment to assess the influence of starvation on the RWDGand the weight/CL ratio of juvenile P. argus was conducted in theUnidad Académica Puerto Morelos (Universidad Nacional Autónomade México). Using Scuba diving, we collected lobsters by hand acrossthe Puerto Morelos reef lagoon and transferred them to the laboratorywithin one hour of collection. Lobsters were acclimatized for twoweeks in shaded tanks with multiple shelters. During this period,lobsters were fed to satiation every other day with a frozen-freshmixture of fish, shrimp, squid, mussels, and crabs. An open seawatersystem maintained ambient temperature in all acclimatization tanksand experimental units (EUs).
Each juvenile phase was tested in a separate trial. In each trial, sixEUs, eachwith two communal shelters appropriate to the size range ofthe experimental lobsters, were randomized between two treat-ments: “fed lobsters” (lobsters fed to excess), and “starved lobsters”(lobsters deprived of food). The EUs for algal juveniles were glassaquaria, 1.0 m in length×0.4 m in width×0.4 m in depth, with a waterlevel of 0.15 m. The EUs for postalgal juveniles and subadults werefiberglass tanks (1.0 m in diameter×0.6 m in height, and 2.0 m indiameter×1.0 m in height, respectively), with a water level of 0.3 m.
Ten visibly healthy lobsters (nine in the postalgal juvenile trial)were introduced into each EU 24 h before the trial began. The mean

Table 1Panulirus argus
Fed lobsters Starved lobsters
Juvenile phase EU Initial Final Healthy EU Initial Final Healthy
Algal juveniles A 10 7 6 A 10 7 7B 10 8 7 B 10 8 5C 10 9 9 C 10 10 6
Postalgal juveniles A 9 9 9 A 9 9 7B 9 9 8 B 9 9 9C 9 8 7 C 9 9 5
Subadults A 10 10 9 A 10 10 10B 10 10 10 B 10 10 10C 10 10 10 C 10 10 10
Summary of data on numbers of lobsters that survived through the experimental trials(final) and that were visibly healthy (healthy) at the end of the trials. Trials consisted ontwo treatments, lobsters fed to excess (fed lobsters) and lobsters deprived of food(starved lobsters), with three experimental units (EU) per juvenile phase and treatment.Initial size range of lobsters were: algal juveniles (7.0–25.0 mm carapace length, CL),postalgal juveniles (25.4–44.0 mm CL), subadults (45.1–78.7 mm CL).
Table 2Panulirus argus
Effect df MS F p
Intercept 1 548.492 1345.453 b0.001Treatment [T] 1 104.714 266.933 b0.001Juvenile phase [JP] 2 3.804 9.697 0.003T × JP 2 2.370 6.042 0.015Experimental unit(T × JP) 12 0.392 1.979 0.032Error 122 0.198
Analysis of variance to test for differences in relative weight of the digestive gland(RWDG) between fed and starved lobsters in each of three juvenile phases: algaljuveniles (7.0–25.0 mm carapace length, CL), postalgal juveniles (25.4–44.0 mm CL) andsubadults (45.1–78.7 mm CL). For each juvenile phase and treatment there were threereplicate experimental units.
Fig. 1. Panulirus argus. (a) Mean (+ SD) relative weight of the digestive gland (RWDG)and (b) mean (+ SD) ratio of weight to carapace length (weight/CL ratio, g/mm) ofexperimental lobsters [algal juveniles (7.0–25.0 mm CL), postalgal juveniles (25.4–44.0 mm CL) and subadults (45.1–78.7 mm CL)] subjected for 25 days to two feedingtreatments: fed lobsters (lobsters fed to excess) and starved lobsters (lobsters deprivedof food). In panel (a), letters above bars denote statistically similar groups (mixed-modelANOVA, followed by Tuckey test on the Treatment× Juvenile Phase interaction). In panel(b), comparisons were made for each juvenile phase with an individual nested ANOVA(ns: not significant).
84 P. Briones-Fourzán et al. / Journal of Experimental Marine Biology and Ecology 370 (2009) 82–88
size of lobsters did not differ significantly among the six EUs (one-wayanalyses of variance, all p-values N0.950). The trials for algal andpostalgal juveniles began in late September 2005. These trials werescheduled to run for six weeks, followed immediately by the subadulttrial, but by mid-October hurricane “Wilma” was fast approachingPuerto Morelos, forcing the termination of the trials after 25 days.“Wilma” struck Puerto Morelos as a category 4 hurricane on 21October 2005, causing extensive damage to our research facilities.Therefore, the subadult trial, which also ran for 25 days, began in lateFebruary 2006. Water temperature ranged from 28.8 °C to 29.8 °Cduring the algal-postalgal juvenile trials and from 25.0 °C to 25.7 °Cduring the subadult trial, but season has no significant effect on themean RWDGs of local juvenile lobsters (Briones-Fourzán et al., 2003).
The optimum amount of food for juveniles of P. argus held at watertemperatures of 24–30 °C is up to 15% of their body weight (Lellis andRussel, 1991), but we doubled that amount for the lobsters fed toexcess. To minimize handling stress, we estimated the individualweight of lobsters with the equation Log weight (g)=2.4306 Log CL(mm) – 2.0048 (Briones-Fourzán et al., 2003) and totaled the weightsof all individuals in each EU to obtain the lobster biomass for that EU.Every day at dusk, we distributed across each EU an amount of food(the same mixture used during acclimatization) equivalent to 30% ofits lobster biomass. All food remains were siphoned out the followingmorning. Starved lobsters were subjected to similar levels ofdisturbance by mimicking food distribution and by also siphoningtheir EUs. Except for the time it took to conduct these activities, all EUsremained covered with a double black mesh.
Lobsters that died during the trials were quantified and examinedfor gross signs of the PaV1 disease (lethargy and morbidity, a pinkdiscoloration of the clear marks over the carapace, and a milkyhemolymph that does not clot) (Shields and Behringer, 2004). At theend of the trials, surviving lobsters were sorted into visibly healthyand visibly diseased. For each trial and EU, the proportion of lobstersthat survived and the proportion of those that were visibly healthywere arcsine transformed. The transformed data were subjected toseparate factorial ANOVAs to test for effects of treatment and juvenilephase (Hurlbert and White, 1993).
Lobsters that survived were stored at –4 °C overnight (Briones-Fourzán et al., 2003). The following morning, each lobster wasmeasured, molt-staged by microscopic observation of the tip of onepleopod (Lyle and MacDonald, 1983), blotted and weighed on a digitalscale (wet weight,±0.01 g), and dissected to extract the digestivegland, which was also blotted and weighed (wet weight of digestivegland, ±0.01 g). For each individual, we estimated the RWDG (=weightof the digestive gland/weight×100) and the weight/CL ratio. Lobstersin postmolt and those that were visibly diseased were not included in
further analyses. The RWDG data were subjected to a mixed-modelANOVA, with Treatment (T, with two levels, fed and starved) andJuvenile Phase (JP, with three levels, algal juveniles, postalgaljuveniles, and subadults) as main (fixed) factors. EU was considereda random factor, nested within the T×JP interaction (Underwood,1981). The final model included the terms T, JP, T× JP, and EU(T×JP).

Table 3Panulirus argus
Effect Algal juveniles Postalgal juveniles Subadults
df MS F p df MS F p df MS F p
Intercept 1 8.979 485.98 b0.001 1 66.804 647.71 b0.001 1 554.83 999.94 b0.001Treatment [T] 1 0.005 0.290 0.617 1 0.181 1.755 0.255 1 0.224 0.403 0.559EU(T) 4 0.018 0.896 0.477 4 0.103 0.606 0.661 4 0.553 0.588 0.673Error 33 0.021 38 0.170 51 0.940
Separate analyses of variance to test for differences in weight/carapace length ratios between fed and starved lobsters in each of three juvenile phases: algal juveniles (7.0–25.0 mmcarapace length, CL), postalgal juveniles (25.4–44.0 mm CL), and subadults (45.1–78.7 mm CL). For each juvenile phase and treatment there were three replicate experimental units(EU).
85P. Briones-Fourzán et al. / Journal of Experimental Marine Biology and Ecology 370 (2009) 82–88
The weight/CL ratios were not subjected to a similar analysis because,given that weight increases as a power funtion of CL, the varianceswere highly heterogeneous and could not be homogenized despiteseveral transformations. Therefore, the weight/CL ratios of eachjuvenile phase were subjected to a nested ANOVA, with T as themain (fixed) factor and EU as a random factor nested within T.
2.2. Comparison of nutritional indices in wild lobsters between 1995and 2005
Wild lobsters were collected by hand from casita sites betweenJune and September 2005 and in late January 2006 to compare theirnutritional indices with those from lobsters collected during 1995(Briones-Fourzán et al., 2003). Immediately after collection, lobsterswere placed in a container with iced seawater to reduce theirmetabolism and transferred to the laboratory. Lobsters were storedovernight at –4 °C and processed the following morning in thesame way as lobsters used in the laboratory trials. Visibly diseasedindividuals were quantified and processed, but data from these lob-sters and from lobsters in postmolt or with obvious errors in anymeasurement (gross outliers) were excluded from further analyses.The RWDG values were subjected to a factorial ANOVA (Underwood,1981). The main (fixed) factors were Year (Y, with two levels, 1995 and2005) and Juvenile Phase (JP, with three levels, algal juveniles,postalgal juveniles, and subadults). The final models included theterms Y, JP, and Y×JP. Theweight/CL ratios of each juvenile phasewerecompared between years with a Student's t-test.
3. Results
Throughout the text, results are expressed as mean±standarddeviation unless stated otherwise. Statistical results were consideredas significant if pb0.05.
Table 4Panulirus argus
Juvenile phase ncollected
n excluded from analyses n includedin analysesdiseased postmolt outliers
(a) 1995Algal juveniles 48 0 9 6 33Postalgal juveniles 91 0 7 9 75Subadults 41 0 3 5 33Total 182 0 19 20 141
(b) 2005Algal juveniles 65 11 (16.9%) 3 10 41Postalgal juveniles 82 3 (3.7%) 4 7 68Subadults 66 1 (1.5%) 6 2 57Total 213 15 (7.0%) 13 19 166
Sample sizes (n) used to compare nutritional indices of the three juvenile phases [algaljuveniles (12.0–25.0 mm carapace length, CL), postalgal juveniles (25.2–45.0 mm CL),subadults (45.7–69.0 mm CL)] of wild lobsters collected from the Puerto Morelos reeflagoon (a) during 1995 and (b) during 2005. Data from lobsters showing gross signs ofinfection by PaV1 (diseased), in postmolt, or that showed obvious errors in somemeasurement (outliers) were excluded from statistical analyses. In parentheses,percentages (prevalence) of visibly diseased lobsters.
3.1. Influence of starvation on nutritional indices
Through the trials, survival differed significantly among juvenilephases (F2, 12=11.388, p=0.002), but was not significantly affected bytreatment (F1,12=0.001, p=0.979) or by the T×JP interaction (F2, 12=0.733,p=0.501). After pooling the data from both treatments, survival did notdiffer between subadults (100%) and postalgal juveniles (98.1%±4.5%),but was significantly lower for algal juveniles (81.7%±11.7%) (Table 1).All lobsters that died showed gross signs of the PaV1 disease. Thesesigns were also observed in a few lobsters that survived (Table 1), butthe proportion of surviving lobsters that were visibly healthy was notsignificantlyaffectedby treatment (F1,12=0.509, p=0.489), juvenile phase(F2, 12=2.319, p=0.141), or the T×JP interaction (F2, 12=0.629, p=0.550).
All factors significantly affected the mean RWDG of experimentallobsters (Table 2). Despite differences among EUs, starved lobsters inevery juvenile phase had a significantly lower mean RWDG than fedlobsters, but the T×JP interactionwas also significant (Table 2). A post-hoc Tukey test for unequal sample sizes revealed a more pronouncedreduction in mean RWDG with starvation for algal juveniles (0.85±0.22 versus 2.59±0.45 in fed algal juveniles) and postalgal juveniles(0.91±0.20 versus 3.12±0.59) than for subadults (1.62±0.53 versus2.98±0.53) (Fig. 1a).
The mean weight/CL ratios increased considerably between sub-sequent juvenile phases (Fig. 1b), but starvation did not significantlyaffect the weight/CL ratios in any juvenile phase (Table 3). There-fore, we pooled the data from both treatments to obtain an over-all mean weight/CL ratio for each juvenile phase. These ratios were0.50±0.14 g/mm for algal juveniles, 1.23±0.41 g/mm for postalgaljuveniles, and 3.14±0.95 g/mm for subadults. The mean weight/CLratios increased by a factor of ~2.5 between subsequent juvenilephases.
3.2. Comparison of nutritional indices in wild lobsters between 1995and 2005
Table 4 summarizes the effective sample sizes of wild lobsters ineach juvenile phase after discarding data from lobsters thatwere visiblydiseased, in postmolt, or with measurement errors. In 1995 all lobsterswere visibly healthy, but there were 7.0% visibly diseased lobsters in2005. However, the prevalence of visibly diseased individuals was
Table 5Panulirus argus
Effect df MS F p
Intercept 1 131.275 25570.578 b0.001Year [Y] 1 0.194 37.759 b0.001Juvenile phase [JP] 2 0.008 1.490 0.227Y × JP 2 0.031 6.018 0.003Error 301 0.005
Analysis of variance to test for changes in the relative weight of the digestive gland(RWDG) of wild lobsters in each of three juvenile phases, algal juveniles (12.0–25.0 mmcarapace length, CL), postalgal juveniles (25.2–45.0 mm CL) and subadults (45.7–69.0 mm CL), collected from the Puerto Morelos reef lagoon during the years 1995 and2005.
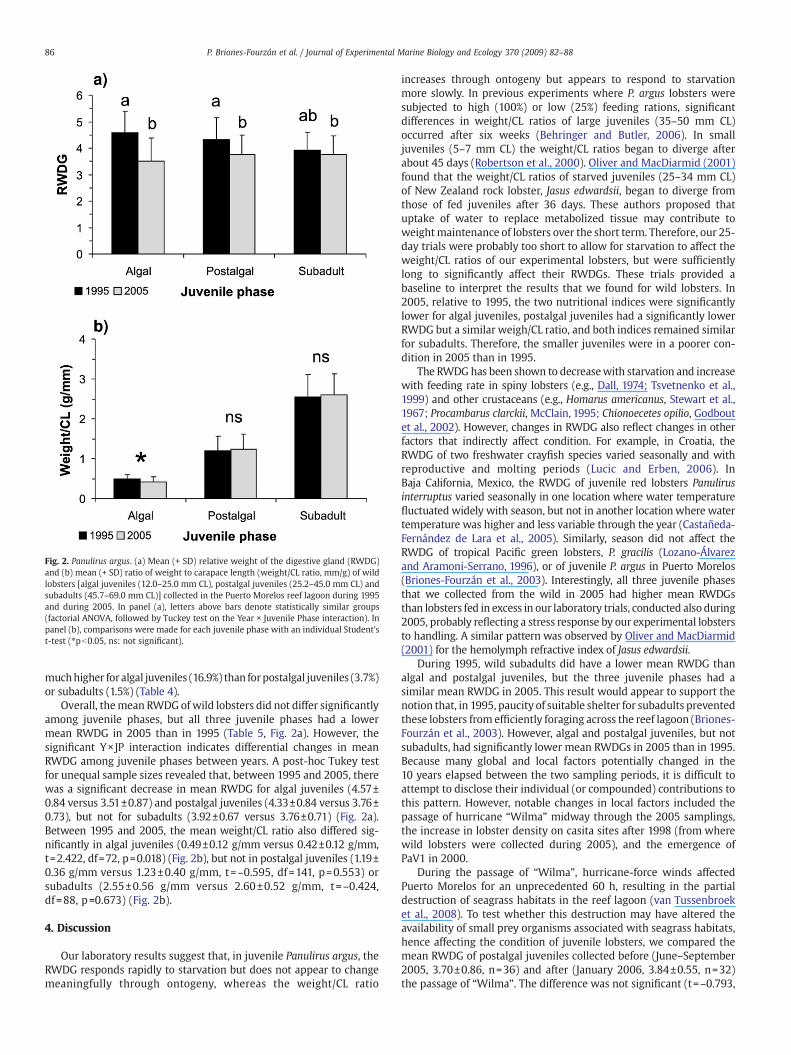
Fig. 2. Panulirus argus. (a) Mean (+ SD) relative weight of the digestive gland (RWDG)and (b) mean (+ SD) ratio of weight to carapace length (weight/CL ratio, mm/g) of wildlobsters [algal juveniles (12.0–25.0 mm CL), postalgal juveniles (25.2–45.0 mm CL) andsubadults (45.7–69.0 mm CL)] collected in the Puerto Morelos reef lagoon during 1995and during 2005. In panel (a), letters above bars denote statistically similar groups(factorial ANOVA, followed by Tuckey test on the Year × Juvenile Phase interaction). Inpanel (b), comparisons were made for each juvenile phase with an individual Student'st-test (⁎pb0.05, ns: not significant).
86 P. Briones-Fourzán et al. / Journal of Experimental Marine Biology and Ecology 370 (2009) 82–88
muchhigher foralgal juveniles (16.9%) than forpostalgal juveniles (3.7%)or subadults (1.5%) (Table 4).
Overall, themean RWDGof wild lobsters did not differ significantlyamong juvenile phases, but all three juvenile phases had a lowermean RWDG in 2005 than in 1995 (Table 5, Fig. 2a). However, thesignificant Y×JP interaction indicates differential changes in meanRWDG among juvenile phases between years. A post-hoc Tukey testfor unequal sample sizes revealed that, between 1995 and 2005, therewas a significant decrease in mean RWDG for algal juveniles (4.57±0.84 versus 3.51±0.87) and postalgal juveniles (4.33±0.84 versus 3.76±0.73), but not for subadults (3.92±0.67 versus 3.76±0.71) (Fig. 2a).Between 1995 and 2005, the mean weight/CL ratio also differed sig-nificantly in algal juveniles (0.49±0.12 g/mm versus 0.42±0.12 g/mm,t=2.422, df=72, p=0.018) (Fig. 2b), but not in postalgal juveniles (1.19±0.36 g/mm versus 1.23±0.40 g/mm, t=–0.595, df=141, p=0.553) orsubadults (2.55±0.56 g/mm versus 2.60±0.52 g/mm, t=–0.424,df=88, p=0.673) (Fig. 2b).
4. Discussion
Our laboratory results suggest that, in juvenile Panulirus argus, theRWDG responds rapidly to starvation but does not appear to changemeaningfully through ontogeny, whereas the weight/CL ratio
increases through ontogeny but appears to respond to starvationmore slowly. In previous experiments where P. argus lobsters weresubjected to high (100%) or low (25%) feeding rations, significantdifferences in weight/CL ratios of large juveniles (35–50 mm CL)occurred after six weeks (Behringer and Butler, 2006). In smalljuveniles (5–7 mm CL) the weight/CL ratios began to diverge afterabout 45 days (Robertson et al., 2000). Oliver and MacDiarmid (2001)found that the weight/CL ratios of starved juveniles (25–34 mm CL)of New Zealand rock lobster, Jasus edwardsii, began to diverge fromthose of fed juveniles after 36 days. These authors proposed thatuptake of water to replace metabolized tissue may contribute toweightmaintenance of lobsters over the short term. Therefore, our 25-day trials were probably too short to allow for starvation to affect theweight/CL ratios of our experimental lobsters, but were sufficientlylong to significantly affect their RWDGs. These trials provided abaseline to interpret the results that we found for wild lobsters. In2005, relative to 1995, the two nutritional indices were significantlylower for algal juveniles, postalgal juveniles had a significantly lowerRWDG but a similar weigh/CL ratio, and both indices remained similarfor subadults. Therefore, the smaller juveniles were in a poorer con-dition in 2005 than in 1995.
The RWDGhas been shown to decreasewith starvation and increasewith feeding rate in spiny lobsters (e.g., Dall, 1974; Tsvetnenko et al.,1999) and other crustaceans (e.g., Homarus americanus, Stewart et al.,1967; Procambarus clarckii, McClain, 1995; Chionoecetes opilio, Godboutet al., 2002). However, changes in RWDG also reflect changes in otherfactors that indirectly affect condition. For example, in Croatia, theRWDG of two freshwater crayfish species varied seasonally and withreproductive and molting periods (Lucic and Erben, 2006). InBaja California, Mexico, the RWDG of juvenile red lobsters Panulirusinterruptus varied seasonally in one location where water temperaturefluctuated widely with season, but not in another locationwhere watertemperature was higher and less variable through the year (Castañeda-Fernández de Lara et al., 2005). Similarly, season did not affect theRWDG of tropical Pacific green lobsters, P. gracilis (Lozano-Álvarezand Aramoni-Serrano, 1996), or of juvenile P. argus in Puerto Morelos(Briones-Fourzán et al., 2003). Interestingly, all three juvenile phasesthat we collected from the wild in 2005 had higher mean RWDGsthan lobsters fed in excess in our laboratory trials, conducted also during2005, probably reflecting a stress response by our experimental lobstersto handling. A similar pattern was observed by Oliver and MacDiarmid(2001) for the hemolymph refractive index of Jasus edwardsii.
During 1995, wild subadults did have a lower mean RWDG thanalgal and postalgal juveniles, but the three juvenile phases had asimilar mean RWDG in 2005. This result would appear to support thenotion that, in 1995, paucity of suitable shelter for subadults preventedthese lobsters from efficiently foraging across the reef lagoon (Briones-Fourzán et al., 2003). However, algal and postalgal juveniles, but notsubadults, had significantly lower mean RWDGs in 2005 than in 1995.Because many global and local factors potentially changed in the10 years elapsed between the two sampling periods, it is difficult toattempt to disclose their individual (or compounded) contributions tothis pattern. However, notable changes in local factors included thepassage of hurricane “Wilma” midway through the 2005 samplings,the increase in lobster density on casita sites after 1998 (from wherewild lobsters were collected during 2005), and the emergence ofPaV1 in 2000.
During the passage of “Wilma”, hurricane-force winds affectedPuerto Morelos for an unprecedented 60 h, resulting in the partialdestruction of seagrass habitats in the reef lagoon (van Tussenbroeket al., 2008). To test whether this destruction may have altered theavailability of small prey organisms associated with seagrass habitats,hence affecting the condition of juvenile lobsters, we compared themean RWDG of postalgal juveniles collected before (June–September2005, 3.70±0.86, n=36) and after (January 2006, 3.84±0.55, n=32)the passage of “Wilma”. The difference was not significant (t=–0.793,

87P. Briones-Fourzán et al. / Journal of Experimental Marine Biology and Ecology 370 (2009) 82–88
df=66, p=0.430), supporting previous findings that wild juvenile Pa-nulirus argus are not food-limited (see Introduction).
In sedentary lobsters, which tend to forage close to their dens(e.g., Wynne and Côté, 2007), an increase in population density mayincrease competition for food and affect nutritional condition. Forexample, at a Mexican Pacific location, the condition of blue lob-sters Panulirus inflatus (a sedentary species, Lozano et al., 1982) wassignificantly reduced following a considerable increase in local pop-ulation density (Lozano-Álvarez and Aramoni-Serrano, 1996). How-ever, in highly mobile lobsters such as P. argus, growth rates andnutritional condition appear to be density-independent. For example,in Florida Bay, deployment of artificial crevice shelters increased thelocal density of juvenile P. argus but did not affect their hemolymphrefractive index (Behringer and Butler, 2006) and, in Puerto Morelos, a6-fold increase in mean lobster density on casita sites relative tocontrol sites did not affect their growth rates (Briones-Fourzán et al.,2007).
We suspect that the lower RWDGs of wild algal and postalgaljuveniles during 2005 were related to the PaV1 disease. This diseasedid not exist in 1995 and, prior to its emergence, wild populations ofspiny lobsters were remarkably free of pathogenic diseases (Evanset al., 2000). In 2005, PaV1 prevalence was highest in algal juveniles,followed by postalgal juveniles. Consistent with the findings byBehringer et al. (2008) for the hemolymph refractive index, we foundthat, in algal juveniles, the mean RWDG was significantly lower invisibly diseased individuals (2.86±1.73, n=11) than in visibly healthyindividuals (3.54±0.86, n=47) (Mann-Whitney U-test, U=132, p=0.012), but this cannot explain our field results because we excludeddata from all visibly diseased lobsters from our analyses. Behringeret al. (2006), however, found that it took six weeks on average forjuvenile P. argus inoculated with infected hemolymph to developgross signs of the disease and an additional two weeks to becomeinfectious. It is possible then that some of the wild lobsters that wesampled in 2005 were already infected but had not yet developedgross signs of the disease. If so, the lower mean RWDGs of algal andpostalgal juveniles in 2005 relative to 1995 would suggest that thedigestive gland is affected progressively as the disease progresses.
Similarly, we only used visibly healthy lobsters in our 25-day trialsyet a few died from PaV1 and others developed gross signs of thedisease through the trials. Bothmortality and the proportion of visiblydiseased lobsters were independent of feed treatment, but the highestmortality occurred in algal juveniles, which are more susceptible toPaV1. In a 75-day trial, poor nutritional condition did not increasesusceptibility to PaV1 in subadult Panulirus argus (50–65 mm CL)challenged with the virus (Behringer et al., 2008). However, subadultsare overall less vulnerable to the virus. Subadults also probably have alower metabolic expenditure than algal and postalgal juveniles (e.g.,they molt less often). In conjunction, these features of subadults couldexplain why, in our trials, starved subadults had a significantly highermean RWDG than starved algal and postalgal juveniles.
5. Conclusion
Between 1995 and 2005, the nutritional condition of the threejuvenile phases of Panulirus argus in the Puerto Morelos reef lagoonunderwent differential changes. Algal juveniles showed the greatestdecrease in condition, followed by postalgal juveniles, while thecondition of subadults remained similar. These differences appearrelated to the PaV1 disease, which emerged locally in 2000 (Huchin-Mian et al., 2008) and whose prevalence has since increased.Interestingly, though, up to 2006 lobster density on casita sites hadnot been impacted (Lozano-Álvarez et al., 2008). However, given thatP. argus is highly gregarious and transmission of PaV1 appears to bemainly by contact, whether the enhancement effect of casitas onjuvenile spiny lobsters could be potentially jeopardized over a longerterm by the PaV1 disease requires further investigation.
Acknowledgements
We thank C. Barradas-Ortiz and F. Negrete-Soto for their invaluableassistance in field and laboratory activities. Additional help wasprovided by E. Ramírez-Zaldívar, M. Pérez-Ortiz, J. Valladárez-Cob, andE. Magallón-Gayón. This study was funded by Consejo Nacional deCiencia y Tecnología (Grant 40159-Q, which included a Bachellor'sthesis scholarship for KBM), and Universidad Nacional Autónoma deMéxico. Annual permits to collect lobsters from the Puerto Morelosreef lagoon were issued by Comisión Nacional de Acuacultura y Pesca,México. [ST]
References
Andrée, S.W., 1981. Locomotory activity patterns and food items of benthic postlarvalspiny lobsters, Panulirus argus. M. Sc. Thesis, Florida State University, Tallahasse.
Behringer, D.C., Butler, M.J., 2006. Density-dependent population dynamics in juvenilePanulirus argus (Latreille): the impact of artificial density enhancement. J. Exp. Mar.Biol. Ecol. 334, 84–95.
Behringer, D.C., Butler, M.J., Shields, J.D., 2006. Avoidance of disease by social lobsters.Nature 441, 421.
Behringer, D.C., Butler, M.J., Shields, J.D., 2008. Ecological and physiological effects ofPaV1 infection on the Caribbean spiny lobster (Panulirus argus Latreille). J. Exp. Mar.Biol. Ecol. 359, 26–33.
Briones-Fourzán, P., Lozano-Álvarez, E., 2001a. The importance of Lobophora variegata(Phaeophyta: Dictyotales) as a habitat for small juveniles of the spiny lobster, Pa-nulirus argus (Decapoda: Palinuridae) in a tropical reef lagoon. Bull. Mar. Sci. 68,207–219.
Briones-Fourzán, P., Lozano-Álvarez, E., 2001b. Effects of artificial shelters (Casitas) onthe abundance and biomass of juvenile spiny lobsters Panulirus argus in a habitat-limited tropical reef lagoon. Mar. Ecol. Prog. Ser. 221, 221–232.
Briones-Fourzán, P., Lozano-Álvarez, E., Negrete-Soto, F., Barradas-Ortiz, C., 2007. En-hancement of juvenile Caribbean spiny lobsters: an evaluation of multiple responsevariables with the addition of large artificial shelters. Oecologia 151, 401–416.
Briones-Fourzán, P., Castañeda-Fernández de Lara, V., Lozano-Álvarez, E., Olivo-Estrada,J., 2003. Feeding ecology of the three juvenile phases of the spiny lobster Panulirusargus in a tropical reef lagoon. Mar. Biol. 142, 855–865.
Butler,M.J., Herrnkind,W.F., 2000. Puerulus and juvenile ecology, In: Phillips, B.F., Kittaka, J.(Eds.), Spiny lobsters: fisheries and culture, 2nd ed. Blackwell, Oxford, pp. 276–301.
Butler, M.J., Herrnkind, W.F., Hunt, J.H., 1997. Factors affecting the recruitment ofjuvenile Caribbean spiny lobsters dwelling in macroalgae. Bull. Mar. Sci. 61, 3–19.
Butler, M.J., Behringer, D.C., Shields, J.D., 2008. Transmission of Panulirus argus Virus 1(PaV1) and its effect on the survival of juvenile Caribbean spiny lobster. Dis. Aquat.Org. 79, 173–182.
Castañeda-Fernández de Lara, V., Servière-Zaragoza, E., Hernández-Vázquez, S., Butler,M.J., 2005. Feeding ecology of juvenile spiny lobster, Panulirus interruptus, on thePacific coast of Mexico. N.Z. J. Mar. Freshw. Res. 39, 425–435.
Colinas-Sánchez, F., Briones-Fourzán, P., 1990. Alimentación de las langostas Panulirusargus y P. guttatus (Latreille, 1804) en el Caribe mexicano. An. Inst. Cienc. Mar yLimnol. Univ. Nal. Autón. México 17 (1), 89–109.
Cox, C., Hunt, J.H., Lyons, W.G., Davis, G.E., 1997. Nocturnal foraging of the Caribbean spinylobster (Panulirus argus) onoffshore reefs of Florida, USA.Mar. Freshw. Res. 48, 671–679.
Dall, W., 1974. Indices of nutritional state in the western rock lobster, Panulirus longipes(Milne-Edwards). I: Blood and tissue constituents and water content. J. Exp. Mar.Biol. Ecol. 16, 167–180.
Eggleston, D.B., Lipcius, R.N., 1992. Shelter selection by spiny lobster under variablepredation risk, social conditions, and shelter size. Ecology 73, 992–1101.
Evans, L.H., Jones, J.B., Brock, J.A., 2000. Diseases of spiny lobsters, In: Phillips, B.F.,Kittaka, J. (Eds.), Spiny lobsters: fisheries and culture, 2nd ed. Blackwell, Oxford,pp. 586–600.
Godbout, G., Dutil, J.D., Hardy, D., Munro, J., 2002. Growth and condition of post-moultmale snow crab (Chionoecetes opilio) in the laboratoy. Aquaculture 206, 323–340.
Herrera, A., Ibarzábal, D., Foyo, J., Espinosa, J., 1991. Alimentación natural de la langostaPanulirus argus en la región de Los Indios (plataforma SWde Cuba) y su relación conel bentos. Rev. Investig. Mar. 12, 172–182.
Herrnkind, W.F., van der Walker, J.A., Barr, L., 1975. Population dynamics, ecology andbehavior of spiny lobsters, Panulirus argus, of St John, U.S.V.I. IV: Habitation, patterns ofmovement and general behavior. Nat. Hist. Mus. Los Angeles Cty. Sci. Bull. 20, 31–45.
Huchin-Mian, J., Rodríguez-Canul, R., Arias-Bañuelos, E., Simá-Álvarez, R., Pérez-Vega, J.A.,Briones-Fourzán, P., Lozano-Álvarez, E., 2008. Presence of Panulirus argus Virus 1(PaV1) in juvenile spiny lobsters, Panulirus argus, from the Caribbean coast of Mexico.Dis. Aquat. Org. 79, 153–156.
Hurlbert, S.H., White, M.D., 1993. Experiments with freshwater invertebrate zooplank-tivores: quality of statistical analyses. Bull. Mar. Sci. 53, 128–154.
Lellis, A.W., Russell, J.A., 1991. Effect of temperature on survival, growth and feed intakeof postlarval spiny lobsters, Panulirus argus. Aquaculture 90, 1–9.
Lipcius, R.N., Eggleston, D.B., 2000. Ecology and fishery biology of spiny lobsters, In:Phillips, B.F., Kittaka, J. (Eds.), Spiny lobsters: fisheries and culture, 2nd ed. Blackwell,Oxford, pp. 1–41.
Lozano, E., Briones, P., Santarelli, L., Gracia, A., 1982. Densidad poblacional de Panulirusgracilis Streets y Panulirus inflatus (Bouvier) (Crustacea: Palinuridae) en dos áreascercanas a Zihuatanejo, Gro., México. Cienc. Pesq. Inst. Nal. Pesca México 3, 61–73.

88 P. Briones-Fourzán et al. / Journal of Experimental Marine Biology and Ecology 370 (2009) 82–88
Lozano-Álvarez, E., Aramoni-Serrano, G., 1996. Alimentación y estado nutricional de laslangostas Panulirus inflatus y Panulirus gracilis (Decapoda: Palinuridae) en Guerrero,México. Rev. Biol. Trop. 44/45, 453–461.
Lozano-Álvarez, E., Briones-Fourzán, P., Ramos-Aguilar, M.E., 2003. Distribution, shelterfidelity, and movements of subadult spiny lobsters (Panulirus argus) in areas withartificial shelters. J. Shellfish Res. 22, 533–540.
Lozano-Álvarez, E., Briones-Fourzán, P., Ramírez-Estévez, A., Placencia-Sánchez, D.,Huchin-Mian, J.P., Rodríguez-Canul, R., 2008. Prevalence of Panulirus argus Virus 1(PaV1) and habitation patterns of healthy and diseased Caribbean spiny lobsters inshelter-limited habitats. Dis. Aquat. Org. 80, 95–104.
Lucic, A., Erben, R., 2006. Seasonal changes of hepatosomatic indices andmoisture contentin the noble crayfish (Astacus astacus) and the stone crayfish (Austropotamobiustorrentium). Freshw. Crayfish 15, 117–125.
Lyle, W.G., MacDonald, C.D., 1983. Molt stage determination in the Hawaiian spinylobster, Panulirus marginatus. J. Crustac. Biol. 3, 208–216.
Mannonen, A., Henttonen, P., 1995. Some observations on the condition of crayfish(Astacus astacus (L.)) in a river affected by peat mining in central Finland. Freshw.Crayfish 10, 274–281.
McClain, W.R., 1995. Effects of population density and feeding rate on growth andfeeding consumption of red swamp crawfish Procambarus clarkii. J. World Aquac.Soc. 26, 14–22.
McGarvey, R., Ferguson, G.J., Prescott, J.H., 1999. Spatial variation in mean growth ratesat size of southern rock lobster, Jasus edwardsii, in South Australian waters. Mar.Freshw. Res. 50, 333–342.
Marx, J., Herrnkind, W.F., 1985. Factors regulating microhabitat use by young juvenilespiny lobsters, Panulirus argus: food and shelter. J. Crustac. Biol. 5, 650–657.
Moore, L.E., Smith, D.M., Loneragan, N.R., 2000. Blood refractive index and whole bodylipid content as indicators of nutritional condition for penaeid prawns (Decapoda:Penaeidae). J. Exp. Mar. Biol. Ecol. 244, 131–143.
Musgrove, R.J.B., 2001. Interactions between haemolymph chemistry and condition inthe southern rock lobster, Jasus edwardsii. Mar. Biol. 139, 891–899.
Nizinski, M.S., 2007. Predation in subtropical soft-bottom systems: spiny lobsters andmolluscs in Florida Bay. Mar. Ecol. Prog. Ser. 345, 185–197.
Oliver, M.D., MacDiarmid, A.B., 2001. Blood refractive index and ratio of weight tocarapace length as indices of nutritional condition in juvenile rock lobsters (Jasusedwardsii). Mar. Freshw. Res. 52, 1395–1400.
Parslow-Williams, P.J., Atkinson, R.J.A., Taylor, A.C., 2001. Nucleic acids as indicators ofnutritional condition in the Norway lobster Neprhops norvegicus. Mar. Ecol. Prog.Ser. 211, 235–243.
Pollock, D.E., Cockcroft, A.C., Goosen, P.C., 1997. A note on reduced rock lobster growthrates and related environmental anomalies in the southern Benguela, 1988–1995.S. Afr. J. Mar. Sci. 18, 287–293.
Robertson, D.N., Butler, M.J., Dobbs, F.C., 2000. An evaluation of lipid- and morpho-metric-based indices of nutritional condition for early benthic stage spiny lobsters,Panulirus argus. Mar. Freshw. Behav. Physiol. 33, 161–171.
Shields, J.D., Behringer, D.C., 2004. A new pathogenic virus in the Caribbean spinylobster Panulirus argus from the Florida Keys. Dis. Aquat. Org. 59, 109–118.
Shields, J.D., Stephens, F.J., Jones, B., 2006. Pathogens, parasites and other symbionts. In:Phillips, B.F. (Ed.), Lobsters: biology, management, aquaculture and fisheries. Black-well, Oxford, pp. 146–204.
Stewart, J.E., Cornick, J.W., Foley, D.M., Li, M.F., Bishop, C.M., 1967. Muscle weightrelationship to serum proteins, hemocytes, and hepatopancreas in the lobster,Homarus americanus. J. Fish. Res. Board Can. 23, 2339–2354.
Thomas, C.W., Carter, C.G., Crear, B.J., 2003. Feed availability and its relationship tosurvival, growth, dominance and the agonistic behaviour of the southern rocklobster, Jasus edwardsii in captivity. Aquaculture 215, 45–65.
Tsvetnenko, E., Brown, J., Glencross, B.D., Evans, L.H., 1999. Measures of condition indietary studies on western rock lobster post-pueruli. Proc. Internat. Symp. LobsterHealth Management, Adelaide, Australia, September 1999, pp. 100–109.
Underwood, A.J., 1981. Techniques of analysis of variance in experimental marinebiology and ecology. Oceanogr. Mar. Biol. Annu. Rev. 19, 513–605.
van Tussenbroek, B.I., Barba-Santos, M.G., van Dijk, J.K., Sanabria-Alcaraz, S.N.M., Téllez-Calderón, M.L., 2008. Selective elimination of rooted plants from a tropical seagrassbed in a back-reef lagoon: a hypothesis tested by HurricaneWilma (2005). J. CoastalRes. 24, 278–281.
Wahle, R.A., Fogarty, M.J., 2006. Growth and development: understanding and modellinggrowth variability in lobsters. In: Phillips, B.F. (Ed.), Lobsters: biology, management,aquaculture and fisheries. Blackwell, Oxford, pp. 1–44.
Wang, S.Y., Stickle, W.B., 1986. Changes in nucleic acid concentration with starvation inthe blue crab Callinectes sapidus Rathbun. J. Crustac. Biol. 6, 49–56.
Wynne, S.P., Côté, I.M., 2007. Effects of habitat quality and fishing on Caribbean spottedspiny lobster populations. J. Appl. Ecol. 44, 488–494.




















