
New aspects concerning to the characterization and the relationship with the immune response in vivo...
11
New aspects concerning to the characterization and the relationship with the immune response in vivo of the spiny lobster Panulirus argus nitric oxide synthase Tania Rodríguez-Ramos a , Yamila Carpio b , Laida Ramos a , Tirso Pons c , Omar Farnós b , Concepción Iglesias d , Iván Sánchez de Melo d , Yassel Ramos b , Carlos Pendón d,⇑ , Mario Pablo Estrada b,⇑ , Jorge Bolívar d,⇑ a Centro de Investigaciones Marinas, Universidad de La Habana (CIM-UH), Cuba b Centro de Ingeniería Genética y Biotecnología (CIGB), Cuba c Facultad de Biología, Universidad de La Habana (Fbio), Cuba d Facultad de Ciencias, Universidad de Cádiz (UCA), Spain article info Article history: Received 7 July 2011 Revised 16 September 2011 Available online 28 September 2011 Keywords: Panulirus argus Spiny lobster Invertebrate immunity Innate immunity Nitric oxide synthase Hemocyte abstract Nitric oxide (NO) is a short-lived radical generated by nitric oxide synthases (NOS). NO is involved in a variety of functions in invertebrates, including host defense. In a previous study, we isolated and sequenced for the first time the NOS gene from hemocytes of Panulirus argus, demonstrating the induc- ibility of this enzyme by lipopolysaccharide (LPS) in vitro. In the present work, lobster hemocytes and gills exposed to Escherichia coli O55:B5 LPS showed an increase in both NOS activity and NOS gene expression in vivo. This response was dose and time dependent. The 3D NOS structure was predicted by comparative modeling showing the oxygenase and reductase domains. These domains contain the conserved binding motifs of NOS already found in a variety of organisms. The 3D structure prediction analysis allowed the selection of a fragment of 666 bp that was cloned and subsequently expressed in E. coli BL21, in which a recombinant product of around 31 KDa was obtained. Hyperimmune serum obtained from immunized rabbits was tested and employed to specifically detect the recombinant polypeptide or the endogenous NOS from lobster hemocytes by western blot and immunofluorescence. This study contributes to enlarge the existing knowledge related to NOS structure and NOS participation in the immune response in lob- sters. The evaluation of an antibody capable to recognize NOS from lobsters constitutes a novel and inter- esting tool for the implementation of further studies on NOS functions in crustaceans. Ó 2011 Elsevier Inc. All rights reserved. Introduction Spiny lobsters (Palinuridae) are interesting models for the study of the crustacean immune system. This is due their huge economic importance as they are among the most valuable seafood in the world, and also because these organisms have large amounts of hemolymph that can be collected, in which important humoral and cellular effectors of the crustacean immunity can be measured [1–4]. However, most of the lobster fisheries are being managed for their maximum sustainable yield. Therefore, the capture of juve- niles from the wild and on-growing them to market size, have offered a means by which their supply can realistically be increased until hatchery production becomes commercially viable [5,6]. Surprisingly, immunity in spiny lobsters remains poorly studied as compared to other crustaceans and few immunity effectors and mechanisms have been examined [7–12]. Thus, the lobster industry must give priority to finding ways to combat pathogens, in order to avoid economic losses in the future. Nitric oxide (NO) is generated by nitric oxide synthases (NOS), a group of evolutionarily conserved cytosolic or membrane bound isoenzymes that convert the amino acid L-arginine to citrulline and NO. In mammals, three isoforms of NOS have been identified. Two of these are constitutively expressed, mainly in the brain (nNOS) and endothelial cells (eNOS) and they are Ca 2+ /calmodulin dependent. A third enzyme, the inducible Ca 2+ -independent iso- form (iNOS), is expressed in some cell types after stimulation with Escherichia coli lipopolysaccharide (LPS) and/or different cytokines such as interferon-c (IFNc), interleukin-1b (IL-1b), or tumor necro- sis factor-a (TNFa) [13,14]. In invertebrates, NO has been involved as a modulator of nerve and muscle activity [15–21]. However, there is now an increasing number of references on NO that show, also, multiple biological 1089-8603/$ - see front matter Ó 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.niox.2011.09.002 ⇑ Corresponding authors. Addresses: Departamento de Biomedicina, Biotecnología y Salud Pública, Universidad de Cádiz (UCA), Facultad de Ciencias, Av. República Saharaui s/n, E11519, Puerto Real, Cádiz, Spain (C. Pendón, J. Bolívar), Center for Genetic Engineering and Biotechnology, Ave. 31 entre 158 y 190, Cubanacán, Playa, P.O. Box 6162, Habana 10600, Cuba (M.P. Estrada). E-mail addresses: [email protected] (C. Pendón), [email protected] (M.P. Estrada), [email protected] (J. Bolívar). Nitric Oxide 25 (2011) 396–406 Contents lists available at SciVerse ScienceDirect Nitric Oxide journal homepage: www.elsevier.com/locate/yniox
Transcript of New aspects concerning to the characterization and the relationship with the immune response in vivo...

Nitric Oxide 25 (2011) 396–406
Contents lists available at SciVerse ScienceDirect
Nitric Oxide
journal homepage: www.elsevier .com/locate /yniox
New aspects concerning to the characterization and the relationship with theimmune response in vivo of the spiny lobster Panulirus argus nitric oxide synthase
Tania Rodríguez-Ramos a, Yamila Carpio b, Laida Ramos a, Tirso Pons c, Omar Farnós b,Concepción Iglesias d, Iván Sánchez de Melo d, Yassel Ramos b, Carlos Pendón d,⇑,Mario Pablo Estrada b,⇑, Jorge Bolívar d,⇑a Centro de Investigaciones Marinas, Universidad de La Habana (CIM-UH), Cubab Centro de Ingeniería Genética y Biotecnología (CIGB), Cubac Facultad de Biología, Universidad de La Habana (Fbio), Cubad Facultad de Ciencias, Universidad de Cádiz (UCA), Spain
a r t i c l e i n f o a b s t r a c t
Article history:Received 7 July 2011Revised 16 September 2011Available online 28 September 2011
Keywords:Panulirus argusSpiny lobsterInvertebrate immunityInnate immunityNitric oxide synthaseHemocyte
1089-8603/$ - see front matter � 2011 Elsevier Inc. Adoi:10.1016/j.niox.2011.09.002
⇑ Corresponding authors. Addresses: DepartBiotecnología y Salud Pública, Universidad de CádizAv. República Saharaui s/n, E11519, Puerto Real, CádizCenter for Genetic Engineering and BiotechnologyCubanacán, Playa, P.O. Box 6162, Habana 10600, Cub
E-mail addresses: [email protected] (C. Pendó(M.P. Estrada), [email protected] (J. Bolívar).
Nitric oxide (NO) is a short-lived radical generated by nitric oxide synthases (NOS). NO is involved in avariety of functions in invertebrates, including host defense. In a previous study, we isolated andsequenced for the first time the NOS gene from hemocytes of Panulirus argus, demonstrating the induc-ibility of this enzyme by lipopolysaccharide (LPS) in vitro. In the present work, lobster hemocytes and gillsexposed to Escherichia coli O55:B5 LPS showed an increase in both NOS activity and NOS gene expressionin vivo. This response was dose and time dependent. The 3D NOS structure was predicted by comparativemodeling showing the oxygenase and reductase domains. These domains contain the conserved bindingmotifs of NOS already found in a variety of organisms. The 3D structure prediction analysis allowed theselection of a fragment of 666 bp that was cloned and subsequently expressed in E. coli BL21, in which arecombinant product of around 31 KDa was obtained. Hyperimmune serum obtained from immunizedrabbits was tested and employed to specifically detect the recombinant polypeptide or the endogenousNOS from lobster hemocytes by western blot and immunofluorescence. This study contributes to enlargethe existing knowledge related to NOS structure and NOS participation in the immune response in lob-sters. The evaluation of an antibody capable to recognize NOS from lobsters constitutes a novel and inter-esting tool for the implementation of further studies on NOS functions in crustaceans.
� 2011 Elsevier Inc. All rights reserved.
Introduction
Spiny lobsters (Palinuridae) are interesting models for the studyof the crustacean immune system. This is due their huge economicimportance as they are among the most valuable seafood in theworld, and also because these organisms have large amounts ofhemolymph that can be collected, in which important humoraland cellular effectors of the crustacean immunity can be measured[1–4]. However, most of the lobster fisheries are being managed fortheir maximum sustainable yield. Therefore, the capture of juve-niles from the wild and on-growing them to market size, haveoffered a means by which their supply can realistically be
ll rights reserved.
amento de Biomedicina,(UCA), Facultad de Ciencias,, Spain (C. Pendón, J. Bolívar),
, Ave. 31 entre 158 y 190,a (M.P. Estrada).
increased until hatchery production becomes commercially viable[5,6]. Surprisingly, immunity in spiny lobsters remains poorlystudied as compared to other crustaceans and few immunityeffectors and mechanisms have been examined [7–12]. Thus, thelobster industry must give priority to finding ways to combatpathogens, in order to avoid economic losses in the future.
Nitric oxide (NO) is generated by nitric oxide synthases (NOS), agroup of evolutionarily conserved cytosolic or membrane boundisoenzymes that convert the amino acid L-arginine to citrullineand NO. In mammals, three isoforms of NOS have been identified.Two of these are constitutively expressed, mainly in the brain(nNOS) and endothelial cells (eNOS) and they are Ca2+/calmodulindependent. A third enzyme, the inducible Ca2+-independent iso-form (iNOS), is expressed in some cell types after stimulation withEscherichia coli lipopolysaccharide (LPS) and/or different cytokinessuch as interferon-c (IFNc), interleukin-1b (IL-1b), or tumor necro-sis factor-a (TNFa) [13,14].
In invertebrates, NO has been involved as a modulator of nerveand muscle activity [15–21]. However, there is now an increasingnumber of references on NO that show, also, multiple biological

T. Rodríguez-Ramos et al. / Nitric Oxide 25 (2011) 396–406 397
roles of this molecule related to feeding, defense, environmentalstress, learning, metamorphosis, swimming, symbiosis, hemocyteaggregation and regulation of blood pressure [22,23].
Recently, there is an increasing interest in the study of crusta-cean NOS genes. According to our knowledge, NOS full length cDNAhas been cloned and sequenced from seven crustaceans. Except theNOS cDNA from the decapod Gecarcinus lateralis [24], the remain-ing sequences have been reported during the last two years. Thisis the case of the NOS sequences obtained from the decapodsPanulirus argus [12], Litopenaeus vannamei [25], Marsupenausjaponicus [26], Carcinus maenas [27], and the cladocerans Daphniapulex [28] and Daphnia magna [29]. However the studies on NOSexpression and its relation to the immune system in crustaceanare still seldom reported.
We have previously demonstrated the presence of NOS inhemocytes of P. argus (Pa-NOS) as an inducible enzyme by LPSin vitro. Additionally, the expression of NOS gene was detected ina variety of organs, including gills, without previous stimulation[12]. In the present study, we have evaluated the effect of theLPS in vivo on NOS activity and NOS gene expression in hemocytesand gills of the spiny lobster P. argus. The 3D structure of Pa-NOSfrom hemocytes was also predicted showing the conserved do-mains and binding motifs of NOS. The 3D structure predictionallowed the selection of a hydrophilic and predominantly exposedNOS fragment that was expressed as a recombinant polypeptide ofapproximately 31 KDa in E. coli BL21. Finally the presence of theenzyme was verified in hemocytes using a new polyclonal antiPa-NOS antibody produced in rabbits immunized with the NOS re-combinant fragment. This study contributes to enlarge theknowledge about the presence, the control of expression and spe-cific role of NOS in the immune system in crustaceans, taking as amodel species with a high economic value.
Table 1Primers used to: (a) Real time quantitative PCR, (b) Expression of NOS in E. coli.
Primers Sequence 50–30 bp Direction
(a) Real time qPCRqFNOS1 CACCGGCAGAAGAGAGAAAGTG 22 ForwardqRNOS2 GCTGCCTGAGAAAGTGTAGGTG 22 ReverseqFbact1 CAGGAATTGCCGATAGGATGC 21 ForwardqRbAct2 TACTTGCGTTCAGGGGGAGC 20 ReverseqFEFa1 CCAGTAGACAAACCACTTCG 20 ForwardqREFa2 CATACCTGGCTTCAAGATGC 20 Reverse
(b) Expression in E. colieFNOS1 CACCGAGCTCGTCGGTTGTGAGAC 24 ForwardeRNOS2 CCGTCGACCTAGTCTAGTGACACAC 25 Reverse
Material and methods
Biological material and sample collection
P. argus lobsters weighting from 0.4 to 0.6 kg were collected bydiving at Golfo de Batabanó, located in the southwestern part ofthe Cuban archipelago. They were kept for one week in the exper-imental area of the Center for Marine Research the University ofHavana. In this area the tanks were connected to a flow throughseawater supply system, using biological–mechanical filtration,seawater UV-sterilization, central aeration system, and controlledphotoperiod of 12:12 h light:darkness. All the experiments weremade with P. argus lobsters, except the detection of NOS by immu-nofluorescence that was made using hemocytes of Panulirus regius.The P. regius lobster were collected by diving at Morocco coasts andkept in the Laboratory of Marine Cultures of the University of Cadizunder similar conditions. Experimental animals were treatedaccording with the European Union Directive (EEC, 1986) for theprotection of animals used for experimental and other scientificpurposes.
To obtain hemocytes, 5 ml of hemolymph from five lobsters wasextracted using a modified Citrate-EDTA buffer (400 mM NaCl,0.1 M glucose, 30 mM trisodium citrate, 26 mM citric acid and20 mM EDTA, pH 4.6) as anticoagulant [30]. The hemolymph wasextracted from the fourth walking leg coxa using a pyrogen-freedisposable syringe containing the same volume of pre-cooled anti-coagulant solution. The mixture was centrifuged at 700g for 10 minat 4 �C. Samples of gills and plasma were stored at �20 �C prior toNOS activity determination. The cell pellet was washed twice withanticoagulant and frozen immediately in liquid nitrogen. Samplesof gills and the hemocytes were stored at �80 �C prior to RNAextraction and gene expression analysis.
Effect of E .coli LPS on in vivo activity and expression of Pa-NOSin hemocytes and gills
To evaluate the effect of LPS on NOS expression and activity atotal of 15 lobsters (five lobsters per group) were labeled and ran-domly allocated to the three flow-through tanks. The LPS concen-trations used were: 2 and 4 mg/kg wet weight (mg/kgWW)dissolved in 400 mM NaCl. The animals were injected in the coxesof the fourth pair of pereiopods. Hemocytes form each lobster wassampled at 0, 3 and 9 h after the injection, while gills werecollected after 9 h of the injection. In the negative control group,the LPS was substituted by 400 mM NaCl.
Gills were homogenized in 10 ml of PBS (137 mM NaCl, 2.7 mMKCl, 4.3 mMNa2HPO47H2O, pH 7.3) per gram of tissue using the Ul-tra-Turrax T25 homogenizer (IKA-Labortechnik, Germany) andcentrifuged at 5000g for 15 min at 4 �C. The supernatants werecollected and stored at �70 �C until use.
Plasma and gills extract supernatant were used to measure ni-trites as an indicator of NOS activity in vivo. To achieve this objec-tive nitrates were first converted to nitrites using the nitratereductase enzyme (Boheringer, Germany). Briefly, 50 ll of sampleswere added to each well and mixed with 50 ll of a freshly pre-pared reaction mixture containing nitrate reductase (2 U/mL)(Boehringer, UK), reduced nicotinamide adenine dinucleotidephosphate (NADPH: 0.344 mM) and flavine adenine dinucleotide(FAD: 0.044 mM) in phosphate buffer. Two fold serial dilutions ofsodium nitrate (250 lM) were used as standard curve. The plateswere incubated at 37 �C for 1 h. Total nitrite was determinedadding 200 ll of Griess reagents (1% sulphanilamide, 0.1%naphthylethylene diamine dihydrochloride, 2.5% H3PO4). Thismethod is based on the reaction of nitrites with Griess reagents,which generates a purple-azo dye product that can be monitoredby a spectrophotometer at 540 nm [31].
To quantify the expression of NOS, total RNA was isolated by theacid guanidinium thiocyanate–phenol–chloroform extractionmethod [32]. Its concentration and quality were determined byUV absorbance at 260 nm, the A260/A280 ratio and by denaturinggel electrophoresis. Contaminating genomic DNA was eliminatedby DNase I digestion (USB, USA).
One and a half micrograms of total RNA from gills and hemo-cytes were reverse-transcribed into cDNA using the iScript cDNAsynthesis kit (Bio-Rad, USA) to a final volume of 30 ll, accordingto the manufacturer’s instructions.
Expression analysis of NOS gene in hemocytes and gills was per-formed by Real-time qPCR as previously reported [12]. Briefly: spe-cific-gene primers (qFNOS1–qRNOS2) (Table 1) were used toamplify an 85 bp fragment from lobster gills and hemocytes. Allreactions were performed in triplicate. Amplification of NOS cDNAsamples was carried out in a MiniOpticom Real Time PCR System(Bio-Rad, USA), using the 2� Sensimix dU SYBR Green Kit(Quantace, UK) in 20 lL final volume and the following conditions:10 min at 94 �C for enzyme activation and initial denaturing, 35

398 T. Rodríguez-Ramos et al. / Nitric Oxide 25 (2011) 396–406
cycles of 15 s at 95 �C, 30 s at 64 �C and 10 s at 72 �C. Standardcurves were generated using five 10-fold serial dilutions of the lin-ear pCR 4-TOPO vector plasmid containing the target/reference se-quence. After each reaction, a melting curve was performed toconfirm that a single PCR product was amplified. NOS expressionwas quantified using relative quantification using the comparativemethod (DDCt) [33]. The values of Ct were first normalized usingthe Ct value of the reference genes, and then they were normalizedusing the expression at time 0. Elongation factor alpha (EFa) and b-actin (b-act) genes were used as references. Quantitative analyseswere performed using MJ OpticonMonitorTM Analysis Softwarev.3.1 (Bio-Rad, USA). For NOS amplification cDNA from 200 ng oftotal RNA per replicate were assayed, whereas cDNA from 50 ngof RNA were enough for EFa and b-act amplifications. The qPCRconditions for the reference genes were the same as abovedescribed for NOS, except that an annealing step at 60 �C was usedfor EFa. Amplified products were sequenced to verify that theycorresponded to the specific Pa-NOS sequence.
Three-dimensional structure by comparative modeling
All databases and software used in the present study arepublicly available on the world-wide web. Sequences and three-dimensional (3D) structures of NOS were retrieved from theUniProt/Swiss-Prot and the Protein Data Bank (PDB) databases,respectively. Position-Specific Iterated BLAST (PSI-BLAST) againstthe National Center for Biotechnology Information (NCBI) non-redundant (nr) database (http://www.ncbi.nlm.nih.gov) was usedto identify P. argus NOS (Pa-NOS) related sequences. Sequencealignments with E-value less than 10�3 and with bit score greaterthan 100 were considered significant.
The profile-alignment option of the CLUSTALX program [34]was used to compare lobster, human inducible (PDB: 4nos) andrat neuronal (PDB: 2tll) NOS sequences. The aligned sequenceswere inspected and adjusted manually to minimize the numberof gaps and insertions. These manual adjustments were based onthe results from sequence similarities, and on the secondary struc-ture elements observed in human inducible and rat neuronal NOS.In search for alternative structural templates from known struc-tures in the PDB, and a sequence-to-structure alignment ofPa-NOS reductase and oxygenase domain sequences we used theMETASERVER available at http://meta.bioinfo.pl/ [35]. Also, wecomplemented the METASERVER predictions with those obtainedby PHYRE [36] in order to gain success in the fold recognitionapproach.
The predicted 3D-models of Pa-NOS reductase and oxygenasedomains were subjected to a series of tests for evaluating theirinternal consistency and reliability. Backbone conformation wasevaluated by the inspection of the Psi/Phi Ramachandran plot ob-tained from PROCHECK analysis [37]. Packing quality of the 3Dmodel was investigated by the calculation of WHATCHECK Z-scorevalue [38]. Finally, sequence–structure compatibility wasevaluated by VERIFY-3D [39]. All these programs were executedfrom the structure analysis and verification servers’ web-site atUCLA (http://www.doe-mbi.ucla.edu/Services/SV/).
In addition, based on the 3D model of the reductase domain, the‘‘protrusion index’’ was calculated according to Thornton et al. [40],to select the fragment of Pa-NOS to be expressed in E. coli.
Bacterial expression and purification of a fragment of Pa-NOS
For expression in E. coli, a NOS fragment of 666 bp was ampli-fied from plasmid DNA by PCR using specific primers (eFNOS1and eRNOS2) (Table 1). These primers contained Sac I and Sal Irestriction sites to insert the amplified fragment into thecorresponding cloning sites in of the PBiEx1 expression vector
(Novagen, USA). The final vector (PBiEx1-NOS) was assayed byrestriction endonucleases site analysis and sequenced to confirmthat variations had not taken place in the nucleotide sequence. Inthis construct, the inserted gene was under the control of theinducible T7 promoter and yielded the polypeptide with a C-termi-nal fusion His (6) tail. Single clones of BL21 transformed withPBiEx1-NOS were grown overnight at 37 �C in Luria Bertani (LB)medium containing 50 lg/mL of ampicillin. Cultures were then di-luted (1:20) in fresh LB medium and grown at 37 �C until the OD600 nm reached approximately 0.5. The expression of recombinantpolypeptides was initiated by the Isopropyl b-D-1-thiogalactopyra-noside (IPTG) (Sigma, USA) to a final concentration of 0.5 mM andincubation continued during 1, 3, 5 h and O/N for induction of therecombinant polypeptide expression.
E. coli cells were lysed in buffer column (100 mM NaH2PO4,10 mM Tris–HCl, pH 8) by pressure using a french hydraulic press(SLM INstrument, Inc.). The whole-cell lysate was fractioned intosoluble and insoluble fractions by centrifugation. Then, the insolu-ble pellet was homogenized in buffer column to test both fractionsfor polypeptide expression in SDS–PAGE and western blotting.
The affinity chromatography was performed under native con-ditions employing 5 ml of Ni–NTA resin (Qiagen, USA) accordingthe manufacturer’s instructions. Briefly, 20 ml of supernatant wereloaded into the column using 100 mM NaH2PO4, 10 mM Tris–HCland 5 mM Imidazole, pH 8. Then, the column was washed with50 ml of 100 mM NaH2PO4, 10 mM Tris–HCl and 40 mM ImidazolepH 8. Finally the polypeptide elution was accomplished with 20 mlof 100 mM NaH2PO4, 10 mM Tris–HCl and 250 mM Imidazole, pH8, by collecting four fractions of 5 ml.
Samples from the induction and purification processes were re-solved in 15% sodium dodecyl sulfate–polyacrylamide gel electro-phoresis (SDS–PAGE) [41] under reducing (1% glycerol, 0.4% SDS,0.1% b-mercaptoethanol, 12.5 mM Tris–HCl, pH 6.6) conditionsand were electrotransferred onto nitrocellulose membranes(Amersham, UK) using a TransBlotTM transfer semi-dry (BioRad,USA).
The nitrocellulose membrane was blocked for 1 h with 5%skimmed milk (Oxoid, UK), and washed with PBST (PBS plus 0.1%Tween-20). The membrane was incubated with an anti-His horse-radish peroxidase conjugate (Sigma, USA) for 1 h, subsequentlywashed, and diaminobenzidine was used for visualization of pro-tein bands. The Broad Range Standard (Bio-Rad, USA) was usedfor visualization of markers on western blot membranes.
Mass spectrometry analysis of the recombinant polypeptide
The Coomassie blue-stained protein band corresponding to re-combinant NOS was treated with 250 mM ammonium bicarbonatein 30% acetonitrile (v/v). Gel bands were cut in 1 mm3 cubes anddehydrated with acetonitrile 90%. Gels were rehydrated with50 mM ammonium bicarbonate, containing 12.5 ng/lL of trypsin(Promega, USA) in a thermomixer, and incubated overnight at37 �C.
Tryptic peptides from selected spots were obtained and pro-cessed as described by González et al. [42]. ESI–MS spectra wereacquired using a QTof-2TM (Micromass, UK) fitted with a Z-spraynanoflow electrospray ion source operated at 80 �C with a dryinggas flow at 50 L/h. Peptides eluted with 60% acetonitrile (v/v) in1% formic acid (v/v) from ZipTips (Millipore, USA) were loaded intothe borosilicate nanoflow tips and submitted to 900 and 35 V ofcapillary and cone voltage, respectively.
Data acquisition and processing were performed using Mass-Lynx v3.5 (Micromass, UK). The ESI-MS/MS spectra were manuallyinterpreted and the sequences of the peptides were successfullyaligned with the deduced amino acid sequence obtained from the

T. Rodríguez-Ramos et al. / Nitric Oxide 25 (2011) 396–406 399
cDNA, using the MASCOT software (http://www.matrix science.com).
Polyclonal antibody production
The purified polypeptide was dialyzed against PBS and quanti-fied using the BCA Protein Assay Kit (Pierce) according to themanufacturer’s instructions. New Zealand white rabbits wereimmunized subcutaneously with three doses (weeks 0, 3 and 7)containing 100 lg of purified recombinant polypeptide (Pa31 KDa-NOS) per dose in Freund’s complete adjuvant (Sigma,USA) at week 0 and 200 lg of purified recombinant polypeptidein Freund’s incomplete adjuvant (Sigma, USA) at weeks three andseven. Rabbit were bled at week 7th.
Immunodetection of recombinant polypeptide (Pa 31 KDa-NOS)and native NOS from hemocytes by western blotting
The detection of the Pa 31 KDa-NOS polypeptide was achievedafter 15% SDS–PAGE in western blotting. The membrane was incu-bated for 1 h with the rabbit serum diluted 1:10,000 in PBS, and agoat anti-rabbit IgG-horseradish peroxidase conjugate (Amersham,UK) as secondary antibody.
Polypeptides from hemocytes were extracted using the followingsolution: 400 mM NaCl, 10 mM KCl and Triton 1%, pH 7. 0. Sampleswere resolved in 10% SDS–PAGE [41] under reducing and non-reducing (1% glycerol, 0.4% SDS, 12.5 mM Tris–HCl, pH 6.6) condi-tions and were electrotransferred overnight onto nitrocellulosemembranes (Amersham, UK) using a TransBlotTM transfer (BioRad,USA). The membrane was blocked for 2 h with 5% skimmed milk(Oxoid, UK), incubated overnight with the anti-NOS immune serumdiluted 1:10000 in PBS and washed with PBST three times for30 min. Afterward, the membrane was incubated with a goat anti-rabbitIgG-horseradish peroxidase conjugate (Amersham, UK) for 1 h, sub-sequently washed, and the enhanced chemiluminescence detectionsystem (Amersham Biosciences, UK) was used for visualization ofprotein bands.
Anti b-actin and anti a-tubulin monoclonal mouse antibodiesfrom Sigma, USA and Abcam, UK respectively, were used at a finaldilution of 1:10000 to detect the reference proteins in the extractof hemocytes. In both case an anti-mouse IgG diluted 1:2000 wasused as secondary antibody (Sigma, USA). The membranes wereautoradiographed with Curix RP2 film (Agfa, Spain).
Immunodetection of native NOS from hemocytesby immunofluorescence
For indirect immunofluorescence staining, hemocytes from sixlobsters (P. regius) were washed twice with anticoagulant and fi-nally exposed in vitro to E. coli LPS as previously described [12].The LPS concentrations used were 0.25 and 0.5 mg/ml. After onehour the hemocytes were extended on poly-lysine treated cover-slips and then fixed in cold acetone for 10 min. Cells were thenwashed with PBS, blocked with BSA 3% in PBS for 30 min and incu-bated with the primary antibody diluted in PBS (1:400 of anti-Pa31 KDa-NOS antibody) at 37 �C for 75 min. Cells were then washedwith PBS for 30 min at room temperature and incubated withAlexa fluor 488 labeled secondary antibodies (Molecular Probes)at 37 �C for 45 min. Finally, cells were washed twice in PBS andmounted in PBS–glycerol containing DAPI at 0.1 lg/ml. A ZeissAxiophot microscope equipped with a 63 � NA 1.3 oil-immersionobjective was routinely used. Images were taken using a SPOTCamera (Diagnostic Instruments Inc.) using the same settings,and processed using Adobe Photoshop software.
Statistical analysis
STATISTICA 6.0 software (StatSoft, Inc. 2001, Tulsa, OK, USA)was used for statistical analyses. ANOVA, followed by Newman–Keuls test was used to test for significant differences. Data of enzy-matic activity and expression are reported as the mean ± standarddeviation. The level of significance was set at 60.05 for all analyses.
Results
Effect of E .coli LPS on in vivo activity of Pa-NOS in hemocytes and gills
The effect of E. coli LPS on NOS metabolites production (nitra-tes + nitrites) in vivo in lobster hemocytes (3 and 9 h after expo-sure) and gills (9 h after exposure) is shown in Figs. 1 and 2A,respectively. The combination of a 4 mg/kgWW dose of LPS and9 h of exposure to the inductor produced the highest concentrationof nitrates and nitrites in both hemocytes and gills. This increasewas higher than the one of the control group at the same time ofexposure (p 6 0.05).
Effect of E .coli LPS on in vivo gene expression of P. argus NOSin hemocytes and gills
There were not significant changes in b-act or EF-a geneexpression in response to LPS. Thus, these genes were used asreference genes.
NOS gene expression showed a significant increase (two to four-fold) (p 6 0.05) in lobster hemocytes at 3 and 9 h after exposure to4 mg/kgWW of LPS with respect to the hemocytes from unexposedlobsters (Fig. 1B, C). NOS expression also showed a significant in-crease (three to four-fold, p 6 0.05) in gills of lobsters exposed tothe higher dose of LPS (Fig. 2B,C). Very similar results were ob-tained when b-act or EF-a were used for normalizing the qPCR re-sults, however, when EF-a was used, the dose of 2 mg/kgWW ofLPS also produced an increase of NOS expression in gills with re-spect to the control group (Fig. 2B).
In silico analysis of Pa-NOS 3D protein structure
Since the 3D-structure of P. argus NOS sequence is not available,and despite the significant sequence similarity between Pa-NOS,and rat neuronal reductase (PDB ID: 2tll; % identity = 46%)/humaninducible oxygenase (PDB ID: 4nos; % identity = 61%) domains, weused fold-recognition methods to search for alternative structuraltemplates in the Protein Data Bank (PDB), and a sequence-to-struc-ture alignment. The alignment provided by PSI-BLAST is usuallynot optimal and often includes only regions of high similaritybetween the query sequence and the database hits, so it wasnecessary to realign the selected template [43].
According to the structure prediction methods METASERVERand PHYRE, the 2tll and 4nos structures match ranked highest withscores greater than the threshold. Therefore, we calculatedindividual 3D-models of Pa-NOS reductase/oxidase domains basedon a consensus sequence-to-structure alignment derived by META-SERVER and PHYRE (Fig. 3).
The predicted 3D NOS structure showed the oxygenase andreductase domains which contain the conserved binding motifs ofNOS found in a variety of organisms. The 3D structure predictionallowed the selection of a hydrophilic and predominantly exposedNOS fragment of 222 aa. (Fig. 3C). This fragment (Val680–Asp901) alsoshowed the higher values in the ‘‘protrusion index’’ plot, which isused to predict continuous antigenic determinants (Fig. 3D) [40].
The 3D models were analyzed by different structure validationprograms including PROCHECK for the evaluation of the
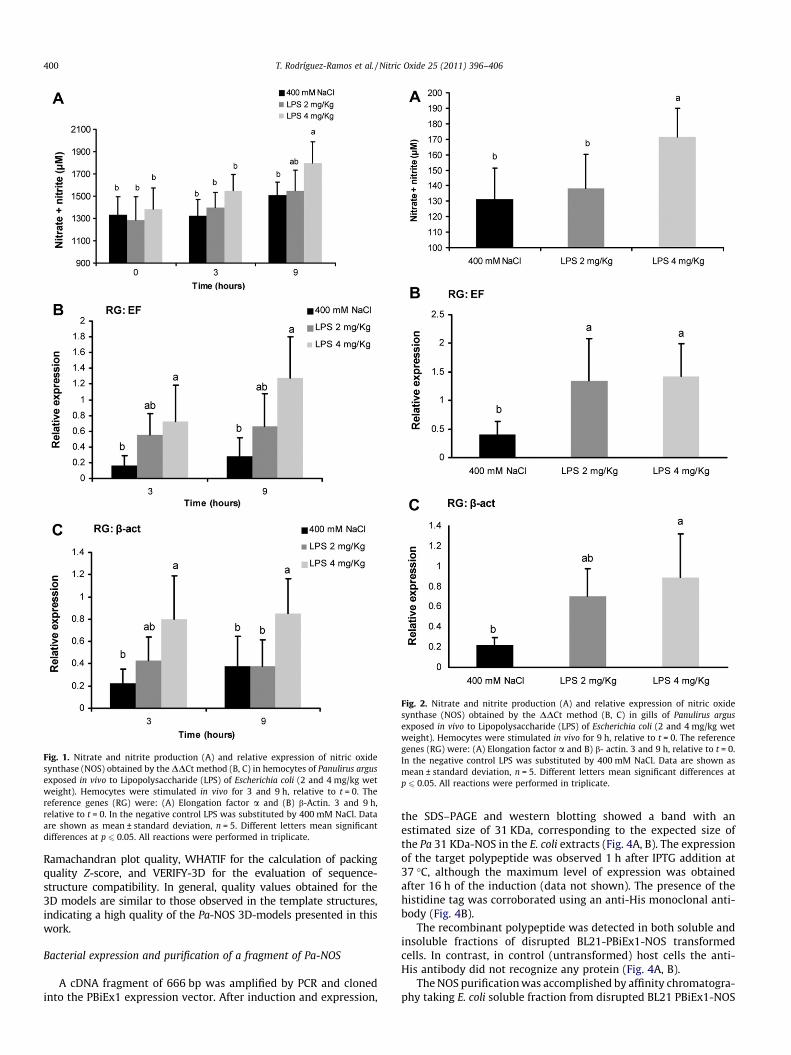
Fig. 1. Nitrate and nitrite production (A) and relative expression of nitric oxidesynthase (NOS) obtained by the DDCt method (B, C) in hemocytes of Panulirus argusexposed in vivo to Lipopolysaccharide (LPS) of Escherichia coli (2 and 4 mg/kg wetweight). Hemocytes were stimulated in vivo for 3 and 9 h, relative to t = 0. Thereference genes (RG) were: (A) Elongation factor a and (B) b-Actin. 3 and 9 h,relative to t = 0. In the negative control LPS was substituted by 400 mM NaCl. Dataare shown as mean ± standard deviation, n = 5. Different letters mean significantdifferences at p 6 0.05. All reactions were performed in triplicate.
Fig. 2. Nitrate and nitrite production (A) and relative expression of nitric oxidesynthase (NOS) obtained by the DDCt method (B, C) in gills of Panulirus argusexposed in vivo to Lipopolysaccharide (LPS) of Escherichia coli (2 and 4 mg/kg wetweight). Hemocytes were stimulated in vivo for 9 h, relative to t = 0. The referencegenes (RG) were: (A) Elongation factor a and B) b- actin. 3 and 9 h, relative to t = 0.In the negative control LPS was substituted by 400 mM NaCl. Data are shown asmean ± standard deviation, n = 5. Different letters mean significant differences atp 6 0.05. All reactions were performed in triplicate.
400 T. Rodríguez-Ramos et al. / Nitric Oxide 25 (2011) 396–406
Ramachandran plot quality, WHATIF for the calculation of packingquality Z-score, and VERIFY-3D for the evaluation of sequence-structure compatibility. In general, quality values obtained for the3D models are similar to those observed in the template structures,indicating a high quality of the Pa-NOS 3D-models presented in thiswork.
Bacterial expression and purification of a fragment of Pa-NOS
A cDNA fragment of 666 bp was amplified by PCR and clonedinto the PBiEx1 expression vector. After induction and expression,
the SDS–PAGE and western blotting showed a band with anestimated size of 31 KDa, corresponding to the expected size ofthe Pa 31 KDa-NOS in the E. coli extracts (Fig. 4A, B). The expressionof the target polypeptide was observed 1 h after IPTG addition at37 �C, although the maximum level of expression was obtainedafter 16 h of the induction (data not shown). The presence of thehistidine tag was corroborated using an anti-His monoclonal anti-body (Fig. 4B).
The recombinant polypeptide was detected in both soluble andinsoluble fractions of disrupted BL21-PBiEx1-NOS transformedcells. In contrast, in control (untransformed) host cells the anti-His antibody did not recognize any protein (Fig. 4A, B).
The NOS purification was accomplished by affinity chromatogra-phy taking E. coli soluble fraction from disrupted BL21 PBiEx1-NOS

Fig. 3. Three-dimensional model of the oxygenase (A) and reductase (B) domains of Pa-NOS, showing the binding sites to heme (green), tetrahydrobiopterin (BH4) (blue), FMN(yellow), flavin (dark blue), FAD (purple) and NADPH (red). Different residues are distinguished because of their relevance in other NOS previously studied: Cysteines in thepositions 34 and 39 of the oxygenase domain form part of a zinc-tetrathiolate (dimerization domain) that plays a key role in stabilizing the NOS dimer structure. Trp383 is believedto be the key for stabilization of BH4 binding. Glu297 has been found to be critical for substrate binding. The presence of Asn290 and Asp302 is characteristic of the active site withinthe inducible NOS. (C) Reductase domain showing the fragment Val680–Asp901 that was expressed in E. coli. (D) Calculation of the protrusion index based on a 3D model of thepredicted reductase domain of P. argus NOS. The fragment Val680–Asp901 have the higher values in the ‘‘protrusion index’’ plot. The discontinuous line indicates the average valuefor the reductase domain.
T. Rodríguez-Ramos et al. / Nitric Oxide 25 (2011) 396–406 401
as starting material. The purified polypeptide was analyzed by SDS–PAGE under reducing conditions and was specifically recognizedwith an anti-His monoclonal antibody (Fig. 4C, D).
The band of 31 KDa was then sequenced by MS. Table 2 showsthe results obtained after the manual identification and interpreta-tion of the spectrums. The analysis shows that the extracted
peptide sequences corresponded to a fragment of NOS fromP. argus. The signals at m/z 1362.67 were assigned to the C-termi-nal peptide of the designed polypeptides which includes the histi-dine tag useful for protein purification and immunodetection.Differences between m/z observed and calculated were associatedwith the internal error of the mass spectrometer (below 50 ppm).

Fig. 4. Expression and purification of Pa 31 KDa-NOS. (A) SDS–PAGE 10% showing the proteins of disruption supernatant and pellet from BL21 cells. Lane 1 representsproteins from the non-transformed BL21 disruption pellet; Lane 2 represents proteins from the non-transformed BL21 disruption supernatant; Lane 3 represents proteinsfrom the BL21-PBiEx1-NOS disruption pellet; and Lane 4 represents proteins from the BL21-PBiEx1-NOS disruption supernatant. Arrows indicate the expression of therecombinant polypeptide. (B) Lanes from 1 to 4 show the proteins specifically recognized by the anti-His monoclonal antibody in the same samples showed in lanes 1–4 fromA. (C) SDS–PAGE and (D) western blotting showing the recombinant NOS after purification employing a Ni–NTA (Quiagen, USA) affinity gel column. In all cases (lines A5 andC3), the molecular weight marker Broad Range Standard (Bio-Rad, USA) was used.
Table 2Sequences obtained from mass spectrometry (MS). Bold letters indicate the sequence of the expression vector included in the recombinant polypeptide. Cursive and underlinedletters indicate the sequence of the recombinant NOS experimentally demonstrated. Asterisk indicates the sequence corresponding to the C-terminal fragment of the recombinantpolypeptide.
Recombinant NOS Sequences obtained by MS m/z E m/z T Z
MAHHHHHHSSGKETAAAKFERQHMDSPPPSGLVPRGSAGSGTIDDDDKISGPGSDPNWELVGCETFCLDDDVAMKEATAALKTEASASADKIRLSPCTK
TDDIALGLSRAHGKRVRSCQVLVSRNLHGEKASR
WTQQVILSTDSINDLNYVPGDHVAILPANRQDLVDAVLARLD
NCPDPDQPIQVLLLKEIHSLNGVSQTWEPHER
LPSASVRELLTRYLDITTPPTSNFLHLLAEYAYDNDQRTRLDQL
AMDPHEYEEWKHMRYPHLKEVLEEFPSVSLD
TDDIALGLSR 1059.57 1059.56 2QDLVDAVLAR 1098.63 1098.60 2EIHSLNGVSQTWEPHER 2017.97 2017.96 3EVLEEFPSVSLD⁄ 1362.67 1362.66 2LDNCPDPDQPIQVLLLK 1991.05 1991.03 2YLDITTPPTSNFLHLLAEYAYDNDQR 369.50 369.47 3WTQQVILSTDSINDLNYVPGDHVAILPANR 3348.71 3348.71 3
402 T. Rodríguez-Ramos et al. / Nitric Oxide 25 (2011) 396–406
Immunodetection of NOS in hemocytes by western blotting andimmunofluorescence
Immune sera from three rabbits were collected and thespecificity was tested by western blotting. The sera specificallyrecognized the recombinant Pa 31 KDa (Fig 5A).
In order to detect the endogenous NOS from hemocytes of thelobster P. argus the cell homogenates were analyzed by SDS–PAGEunder reducing and non reducing conditions followed by westernblotting. Under reducing conditions, a single band correspondingto the monomer was observed (Fig. 5C lanes 1). In contrast, undernon-reducing conditions a predominant band was obtained withthe expected size of the lobster NOS homodimer, while anotherband of lower intensity was observed with the expected size ofthe monomer (Fig. 5C lane 3).
On the other hand, when the anti-NOS serum was pre-incubated for 2 h with the recombinant polypeptide, the intensityof the bands decreased, under both reducing and non reducingconditions, indicating the specificity of the serum (Fig. 5C, lanes2 and 4). NOS from shrimp L. vannamei hemocytes were also spe-cifically recognized by anti-NOS serum (data not shown).
Native NOS was specifically recognized by immunofluorescencemicroscopy in the hemocytes of P. regius using the polyclonalantibody anti-Pa-NOS (Fig. 6). The intensity of the fluorescencewas higher in the hemocytes exposed to LPS of E. coli with respectto the unexposed hemocytes. In all cases NOS was localized in thecytoplasm of these cells with higher fluorescence intensities whenthe LPS concentration was increased. The hemocytes exposedin vitro to the higher dose of LPS (0.5 mg/ml) also showed aggrega-tion and changes in their morphology. Control hemocytes incu-bated with pre-immune serum showed no background fluorescence (data not shown).
Discussion
In this study we have shown that the hemocytes and gills of thecrustacean P. argus were able to express NOS and produce NO,following an injection of E. coli LPS in vivo. The increase obtainedin nitrites (an indicator of NO production) in plasma was similarto the increase obtained in a variety of invertebrates challengedwith pathogen or microbial components. For instance, inDrosophila larvae after the infection with Leptopilina boulardi[44]; in the shrimp Fenneropenaeus chinensis and M. japonicus afterchallenge with WSSV [45] and in the crayfish P. clarkii [46], thesnail V. ater [47] and the shrimp Litopenaeus schmitti [48] afterexposure to LPS.
The hemocytes are the main cellular effectors of immunity incrustaceans and they have several different functions such as adhe-sion, phagocytosis, encapsulation and melanization [1–4,49,50].The relationship between NOS and these functions are still un-der-studied in invertebrates, although there are already someavailable data. In Procambarus clarkii, the hemocyte-derived Ca-independent NOS were involved in promoting bacterial adhesionto hemocytes and enhancing bactericidal activity of hemocytes[46]. In vitro bacterial clumping activity in the hemolymph ofMytilus edulis, in the presence or absence of inhibitory drugs orstimulation by LPS, revealed the release of NO by the hemocytes[51]. Moreover, in Macrobrachium rosenbergii the hemocytes hadthe ability to generate NO during phagocytosis of erythrocytes,and there was an enhancement in the generation (75% increase)of NO during agglutinin-mediated phagocytosis [52].
In the present study, NOS gene expression was higher in hemo-cytes and gills of the lobster after injection with LPS. This variation,detected by real-time RT-PCR, was observed at 3 and 9 h after theinjection with the inductor. Recently Yao et al. [25] also
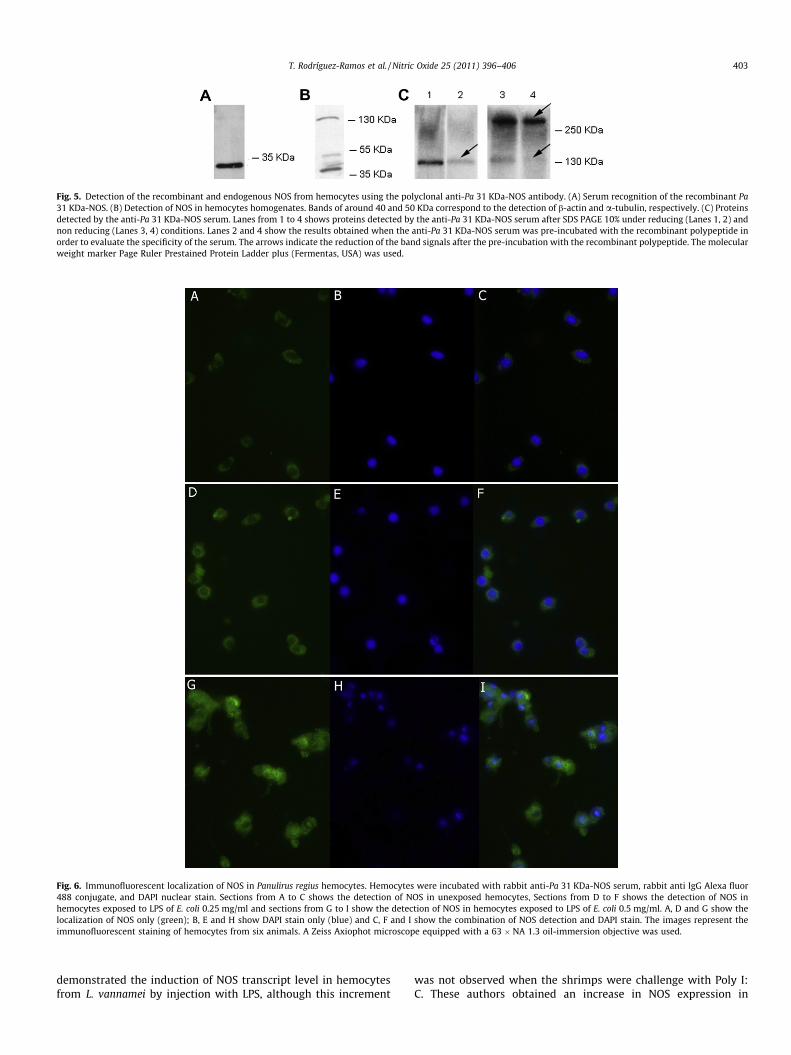
Fig. 6. Immunofluorescent localization of NOS in Panulirus regius hemocytes. Hemocytes were incubated with rabbit anti-Pa 31 KDa-NOS serum, rabbit anti IgG Alexa fluor488 conjugate, and DAPI nuclear stain. Sections from A to C shows the detection of NOS in unexposed hemocytes, Sections from D to F shows the detection of NOS inhemocytes exposed to LPS of E. coli 0.25 mg/ml and sections from G to I show the detection of NOS in hemocytes exposed to LPS of E. coli 0.5 mg/ml. A, D and G show thelocalization of NOS only (green); B, E and H show DAPI stain only (blue) and C, F and I show the combination of NOS detection and DAPI stain. The images represent theimmunofluorescent staining of hemocytes from six animals. A Zeiss Axiophot microscope equipped with a 63 � NA 1.3 oil-immersion objective was used.
Fig. 5. Detection of the recombinant and endogenous NOS from hemocytes using the polyclonal anti-Pa 31 KDa-NOS antibody. (A) Serum recognition of the recombinant Pa31 KDa-NOS. (B) Detection of NOS in hemocytes homogenates. Bands of around 40 and 50 KDa correspond to the detection of b-actin and a-tubulin, respectively. (C) Proteinsdetected by the anti-Pa 31 KDa-NOS serum. Lanes from 1 to 4 shows proteins detected by the anti-Pa 31 KDa-NOS serum after SDS PAGE 10% under reducing (Lanes 1, 2) andnon reducing (Lanes 3, 4) conditions. Lanes 2 and 4 show the results obtained when the anti-Pa 31 KDa-NOS serum was pre-incubated with the recombinant polypeptide inorder to evaluate the specificity of the serum. The arrows indicate the reduction of the band signals after the pre-incubation with the recombinant polypeptide. The molecularweight marker Page Ruler Prestained Protein Ladder plus (Fermentas, USA) was used.
T. Rodríguez-Ramos et al. / Nitric Oxide 25 (2011) 396–406 403
demonstrated the induction of NOS transcript level in hemocytesfrom L. vannamei by injection with LPS, although this increment
was not observed when the shrimps were challenge with Poly I:C. These authors obtained an increase in NOS expression in

404 T. Rodríguez-Ramos et al. / Nitric Oxide 25 (2011) 396–406
hemocytes 3 h after LPS injection that later decreased in time untilno differences at 12 h. Then, it peaked again at 24 h post-injection.The nature of the apparent differences in the ability of crustaceanhemocytes to respond to LPS requires further study. Discrepanciesin the results might be also attributable to the different experimen-tal conditions and procedures used, as well as to differences due tospecies specificity.
The results presented herein demonstrated NOS transcriptionincrement after LPS injection also in gills. These findings are con-sistent with the high NOS expression found previously [12,24]and the fact this organ is an important point for entry of pathogens.It is therefore reasonable to expect that gills have a high level ofcellular and humoral effectors of the immune response in crusta-cean species. For instance, an invasion of hemocytes in the gillswas observed in Penaeus monodon upon White Spot SyndromeVirus (WSSV) infection, possibly caused by the adherence of hemo-cytes to the hemolymph vessels [53]. In the Pacific white shrimp L.vannamei, nine genes relevant to immune defense were expressedin gills, especially the genes encoding prophenoloxidase (proPO),lysozyme, crustin, transglutaminase (TGase), penaeidin-3 andcytosolic manganese superoxide dismutase (cMnSOD) [54]. In theland crab G. lateralis gills, NOS gene expression was also detectedand the NOS protein was localized by immunohistochemistry tothe epithelium lining the central axis between the gill lamellaeand in the pillar cells [24]. Also, in a time course experiment onMarsupenaeus japonicus NOS expression, it was demonstrated anincrease in the relative expression level of NOS in the gill afterinjection of Vibrio penaeicida [26].
Purified LPS has been used by many researchers as an effectivestimulator of NOS production [45–48,55]. In this study the effectof LPS on NOS activity and expression in vivo was found to be doseand time dependent. Accordingly, the dose of 4 mg/kgWW of LPSproduced the highest NOS gene expression and also the higherconcentration of nitrates and nitrites in hemocytes and gills. Thelower dose (2 mg/kgWW) was insufficient to induce the variationof NOS expression or NOS activity when compared to the controlgroup, although the analysis in gills using EF a as reference gene alsoshowed a significant increment of NOS gene expression in this organat the lower dose. In addition, 9 h of exposure to the inductorproduced the highest concentration of nitrates and nitrites in hemo-cytes while 3 h after challenge were enough to show the induction ofNOS gene expression in LPS treated lobsters. The higher sensitivity ofReal time PCR technique and the fact that the inducible NOS arehighly regulated by post-transcriptional events could be the causesof the differences observed in the results [13,14].
We have performed previous studies in which the effect of theinhibitor L-NMMA on LPS-treated hemocytes was tested in vitro[12]. The use of L-NMMA caused a decrease in nitrite concentra-tion: 1.7-fold-decrease with the lowest dose of inhibitor assayed(1.5 mM) and 2.4-fold-decrease with the highest dose (3.0 mM).These results demonstrated a direct relationship between an in-crease in the concentration of nitrites and NOS activity after treat-ment with E. coli LPS, given that nitrites are one of the stable endproducts of NO metabolism.
In all cases the increase in NOS gene expression or NOS activitywere low increased. It is known that for iNOS expression andactivity in vertebrates the response to LPS could be up-regulatedby pro-inflammatory cytokines such as interferon-c, TNF andIL-1. Therefore, it would be interesting to test the effect of acombined treatment with LPS and cytokines on NOS expressionin crustacean, despite the fact that studies about the presence ofcytokines homologs in this group still need more attention [56].
In general, the inducible NOS have been demonstrated inarthropods but still it remains to clarify the function and structureof this enzyme [46,57–59]. In this study the 3D structure of thehuman inducible NOS was used as the template for the oxygenase
domain. The predicted 3D-models of Pa-NOS reductase and oxy-genase domains showed the structure of the conserved bindingmotifs of NOS [12]. Previous studies regarding NOS structure havedemonstrated that the substrate (L-Arg or N-hydroxyarginine)binds above the heme iron atom in the distal pocket, whereasthe cofactor (H4B) binds along the side of the heme [60].
The crystal structures of NOS previously obtained have shownthat the heme is coordinated by a cysteine residue on the proximalside [60–62], which seems to be the Cys120 in P. argus. The structureof the heme domain in the human inducible nitric-oxide synthase(iNOS) also showed two symmetry-related cysteine residues thatform a disulfide bond in the zinc-free structure state, and the sametwo cysteine residues forming part of a zinc-tetrathiolate (ZnS(4))center in the Zn-bound state. This Cys-(X)4-Cys motif is conservedin most of the NOS sequences known to date, showing that the theZnS(4) plays a key role in stabilizing the NOS dimer structure andin maintaining the integrity of the cofactor tetrahydrobiopterin(BH4) binding site of NOS [61,63]. Pa-NOS shows these cysteines inthe positions 34 and 39 of the protein. Besides, among residues inthe BH4 binding site that have been studied by mutagenesis,Trp457 in murine iNOS that correspond to the Trp383 in P. argus seemsto be the most critical, and is believed to be the key for stabilizationof the BH4 binding in all three isoforms [64].
Additionally, in the structure of the murine iNOS oxygenase do-main, the Glu371 has been found to be critical for substrate bindingto murine iNOS. The L-arginine guanidinium group makes twohydrogen bonds to the carboxy oxygens of this residue, localizingthe substrate over the heme [61]. The same residue is found inPa-NOS corresponding to the position 297. Also, there are two dif-ferences found in the structure of the active site of the inducibleNOS with respect to the constitutive NOS. One of these is an Asnresidue on the b-strand in murine iNOS that replaces a Ser in bothnNOS and eNOS [64]. The other difference in the arginine bindingsite is at iNOS Asp, which is an Asn in eNOS and lies some 9 Å fromthe substrate guanidinium [61]. These residues correspond to theAsn290 and Asp302 in P. argus NOS, respectively.
This finding is very interesting and probably ratifies the exis-tence of an inducible NOS in P. argus. However, it is known thatinvertebrate NOS can show characteristics of both inducible andCa2+/CaM-dependent enzymes. In vertebrates, the presence of aninhibitory loop in the middle of the FMN-binding subdomain con-trols the Ca2+/CaM-dependent electron transfer in the constitutiveisoforms. The neuronal NOS also has a PDZ domain in the N-termi-nal region [65]. Sequence alignments of lobster and human iso-forms indicated that the deduced NOS protein of P. argus lackedthe PDZ domain although it has a sequence in the position of theinhibitory loop of the constitutive isoforms. Further functionalstudies will ratify or not the inhibitory role of this loop in lobsters.
The predicted 3D-models of the reductase domain helped us toselect a fragment of 222 aa that was expressed in E. coli using theexpression vector PBiEx1. The MS analysis showed that the ex-pressed polypeptide sequences corresponded unequivocally to afragment of NOS from P. argus. The anti-NOS serum obtained fromrabbits specifically recognized the recombinant Pa 31 KDa-NOSand also the endogenous NOS from hemocytes and gills of P. argus.
In vertebrates, iNOS is catalytically active only in its dimericform. The heme group, the BH4 cofactor, as well as the substrateL-Arg contribute to dimer stability. However the monomeric formcan be found in cells as well. For example, in human hepatocytesiNOS exists in a least two pools: a soluble pool composed of boththe active dimers and monomers and also a peroxisomal pool ofmonomeric iNOS that can be found sequestered in peroxisomes[66]. Probably something similar occurs in hemocytes and there-fore under non reducing conditions it was possible to detect thepresence of the dimeric and monomeric forms of the enzyme inthe total cell lysates, although most of the protein seems to be in

T. Rodríguez-Ramos et al. / Nitric Oxide 25 (2011) 396–406 405
its dimeric form. As expected, when the samples were run underreducing conditions Pa-NOS dimer was nearly absent and the bandcorresponding to the monomeric enzyme was stronger.
The results obtained by immunofluorescence microscopyshowed the localization of NOS in the cytoplasm of the lobsterhemocytes, even when the cells were unexposed to LPS. To ourknowledge this is the first report of the subcellular localization ofNOS in hemocytes of a crustacean. NOS has been previouslydetected by immunostaining in Y-organ of the crab G. lateralis[24] and C. maenas [27] using an universal NOS antibody.
The intensity of the fluorescence was higher in the hemocytesthat were exposed to LPS of E. coli, and this increment was dosedependent. The hemocytes exposed in vitro to the higher dose ofLPS (0.5 mg/ml) also showed an aggregation process and changesin their morphology, probably caused by the degranulation of thecells. It is known that in invertebrates the recognition of the LPSby pattern-recognition proteins (PRPs), induces the degranulationof haemocytes and several proteins of the immune system are re-leased, leading to melanin production, cell adhesion, encapsula-tion, and phagocytosis [2]. Further studies have to demonstrate ifNOS is released during the degranulation of the hemocytes andhow this protein is related to other effectors of the immune systemin crustacean.
In this work, we have produced and evaluated an anti-lobsterNOS serum that constitute a valuable tool for the design of futureexperiments concerning to the localization and functions of nitricoxide synthase in lobsters and probably in close taxonomic groups.This antiserum allowed the characterization of the recombinantNOS fragment expressed in E. coli and the identification of theendogenous protein in hemocytes and gills. To date, localizationstudies of NOS in various tissues of non-mammalian vertebrateshave been performed with a universal anti-NOS antibody[24,27,67]. Until this study, no previous published data havedescribed the production and usefulness of an antiserum against acrustacean nitric oxide synthase. To our knowledge, this is also thefirst report on the effect of LPS in vivo on nitric oxide production ina lobster. The results demonstrated that NOS expression is inducedin hemocytes and gills, causing an increment in the NOS activity inresponse to a microbial elicitor of the immune system. Therefore,our previous in vitro [12] and present in vivo results strongly suggestthat NO has a role in the immune responses of lobsters.
Acknowledgments
This study was supported by a grant from the InternationalFoundation of Science, (IFS grantee A/4609-1) to TRR and by theAECID Grant to CP and LR (PCI, A/024958/10). The author thankRolando Perdomo from the Center for Research and Drug Develop-ment of Cuba for the donation of E. coli LPS and the Aquacultureteam from the Center for Marine Research of Cuba, especially ErickPerera, for the essential support in animal care and collecting sam-ples. Sequencing analyses were performed in ‘‘Servicio Central deCiencia y Tecnología’’, University of Cadiz, Spain.
References
[1] P. Jiravanichhpaisal, B.L. Lee, K. Söderhäll, Cell-mediated immunity inarthropods: hematopoiesis, coagulation, melanization and opsonization,Immunobiology 211 (4) (2006) 213–236.
[2] S.Y. Lee, K. Söderhäll, Early events in crustacean innate immunity, Fish &Shellfish Immunology 12 (2002) 421–437.
[3] J. Rodríguez, G. Le Moullac, State of the art of immunological tools and healthcontrol of penaeid shrimp, Aquaculture 2000 (191) (2000) 109e19.
[4] K. Söderhäll, P.O. Thörnqvist, Crustacean immunity - a short review, FishVaccinology 90 (1997) 45–51.
[5] B.F. Phillips, Perspectives, in: B.F. Phillips, J. Kittaka (Eds.), Spiny Lobsters:Fisheries and Culture, 2nd ed., Blackwell Science Ltd., London, UK, 2000, pp.667–672.
[6] B.F. Phillips, R. Melville-Smith, Y.W. Cheng, Estimating the effects of removingPanulirus cygnus pueruli on the fishery stock, Fisheries Research 65 (2003) 89–101.
[7] H. Aono, K. Mori, Interaction between hemocytes and plasma is necessary forhemolymph coagulation in the spiny lobster, Panulirus japonicus, ComparativeBiochemistry and Physiology. Part A 113 (3) (1996) 301–305.
[8] J. Jussila, S. Mc Bride, J. Lago, L.H. Evans, Hemolymph clotting time as anindicator of stress in western rock lobster (Panulirus cignus Geoge),Aquaculture 199 (2001) 185–193.
[9] R. Perdomo-Morales, V. Montero-Alejo, E. Perera, Z. Pardo-Ruiz, E. Alonso-Jiménez, Phenoloxidase activity in the hemolymph of the spiny lobsterPanulirus argus, Fish & Shellfish Immunology 23 (2007) 1187–1195.
[10] R. Perdomo-Morales, V. Montero-Alejo, E. Perera, Z. Pardo-Ruiz, E. Alonso-Jiménez, Hemocyanin-derived phenoloxidase activity in the spiny lobsterPanulirus argus (Latreille, 1804), Biochimica et Biophysica Acta 1780 (4) (2008)652–658.
[11] B. Verghese, E.V. Radhakrishnan, A. Padhi, Effect of environmental parameterson immune response of the Indian spiny lobster, Panulirus homarus (Linnaeus,1758), Fish & Shellfish Immunology 23 (2007) 928–938.
[12] T. Rodríguez-Ramos, Y. Carpio, J. Bolívar, G. Espinosa, J. Hernández-López, T.Gollas Galván, L. Ramos, C. Pendón, M.P. Estrada, An inducible nitric oxidesynthase (NOS) is expressed in hemocytes of the spiny lobster Panulirus argus:cloning, characterization and expression analysis, Fish & Shellfish Immunology29 (2010) 469–479.
[13] C. Bogdan, Nitric oxide and the regulation of gene expression, Trends in CellBiology 11 (2) (2001) 66–75.
[14] A. Pautz, J. Art, S. Hahn, S. Nowag, C. Voss, H. Kleinert, Regulation of theexpression of inducible nitric oxide synthase, Nitric Oxide 23 (2010) 75–93.
[15] M. Araki, H. Schuppe, S. Fujimoto, T. Nagayama, P.L. Newland, Nitric oxidemodulates local reflexes of the tailfan of the crayfish, Journal of Neurobiology60 (2) (2004) 176–186.
[16] J.L. Benton, D.C. Sandeman, B.S. Beltz, Nitric oxide in the crustacean brain:regulation of neurogenesis and morphogenesis in the developing olfactorypathway, Developmental Dynamics 236 (11) (2007) 3047–3060.
[17] F.M. Casares, A. McElroy, K. Mantione, G. Baggermann, W. Zhu, G.B. Stefano,The American lobster, Homarus americanus, contains morphine that is coupledto nitric oxide release in its nervous and immune tissues: evidence forneurotransmitter and hormonal signaling, Neuroendocrinology Letter 26 (2)(2005) 89–97.
[18] M.F. Goy, Nitric oxide: an inhibitory retrograde modulator in the crustaceanheart, Comparative Biochemistry and Physiology A 142 (2005) 151–163.
[19] A. Hermann, C. Erxleben, Nitric oxide activates voltage dependent potassiumcurrents of crustacean skeletal muscle, Nitric Oxide (5) (2001) 361–369.
[20] K.U. Johansson, M. Carlberg, NADPH-diaphorase histochemistry and nitricoxide synthase activity in deutocerebrum of the crayfish, Pacifastacusleniusculus (Crustacea, Decapoda), Brain Research 649 (1994) 36–42.
[21] W. Stein, C.C. Eberle, U.B. Hedrich, Motor pattern selection by nitric oxide inthe stomatogastric nervous system of the crab, European Journal ofNeuroscience 21 (10) (2005) 2767–2781.
[22] A. Palumbo, Nitric oxide in marine invertebrates: a comparative perspective,Comparative Biochemistry and Physiology. Part A 142 (2005) 241–248.
[23] E.S. Chang, D.L. Mykles, Regulation of crustacean molting: a review and ourperspectives, General and Comparative Endocrinology 172 (2011) 323–330.
[24] H.W. Kim, L.A. Batista, J.L. Hoppes, K.J. Lee, D.L. Mykles, A crustacean nitricoxide synthase expressed in nerve ganglia, Y-organ, gill and gonad of thetropical land crab, Gecarcinus lateralis, Journal of Experimental Biology 207(16) (2004) 2845–2857.
[25] C.l. Yao, P.F. Ji, Z.Y. Wang, F.H. Li, J.H. Xiang, Molecular cloning and expressionof NOS in shrimp, Litopenaeus vannamei, Fish & Shellfish Immunology 28 (3)(2010) 553–560.
[26] M. Inada, T. Mekata, R. Sudhakaran, S. Okugawa, T. Kono, A.M. El Asely, et al.,Molecular cloning and characterization of the nitric oxide synthase gene fromkuruma shrimp Marsupenaeus japonicus, Fish & Shellfish Immunology 28 (4)(2010) 701–711.
[27] A.A. McDonald, E.S. Chang, D.L. Mykles, Cloning of a nitric oxide synthase fromgreen shore crab, Carcinus maenas: a comparative study of the effects ofeyestalk ablation on expression in the molting glands (Y-organs) of C. maenas,and blackback land crab, Gecarcinus lateralis, Comparative Biochemistry andPhysiology Part A 158 (2011) 150–162.
[28] S.J. McTaggart, C. Conlon, J.K. Colbourne, M.L. Blaxter, T.J. Little, Thecomponents of the Daphnia pulex immune system as revealed by completegenome sequencing, BMC Genomics 10 (2009) 175.
[29] P. Labbé, S.J. McTaggart, T.J. Little, An ancient immunity gene duplication inDaphnia magna: RNA expression and sequence analysis of two nitric oxidesynthase genes, Developmental and Comparative Immunology 33 (2009)1000–1010.
[30] K. Söderhäll, V.J. Smith, Separation of the hemocyte populations of Carcinusmaenus and other marine decapods, and prophenoloxidase distribution,Developmental and Comparative Immunology 7 (1983) 229–239.
[31] A.M. Leone, P. Rhodes, V. Furst, S. Moncada, Techniques for the measurementof nitric oxide, in: D.S. Kendall, S.J. Hill (Eds.), Signal Transduction Protocols,Human Press, Totowa, New Jersey, 1995, pp. 285–299.
[32] P. Chomczynski, N. Sacchi, Single-step method of RNA isolation by acidguanidinium thiocyanate-phenol-chloroform extraction, AnalyticalBiochemistry 162 (1) (1987) 156–159.

406 T. Rodríguez-Ramos et al. / Nitric Oxide 25 (2011) 396–406
[33] K.J. Livak, T.D. Schmittgenb, Analysis of relative gene expression data usingrealtime quantitative PCR and the 2-deltadeltaCT method, Methods 4 (25)(2001) 402–408.
[34] J.D. Thompson, T.J. Gibson, F. Plewniak, F. Jeanmougin, D.G. Higgins, TheCLUSTAL_X windows interface. flexible strategies for multiple sequencealignment aided by quality analysis tools, Nucleic Acids Research 25 (1997)4876–4882.
[35] K. Ginalski, A. Elofsson, D. Fischer, L. Rychlewski, 3D-Jury: a simple approach toimprove protein structure predictions, Bioinformatics 19 (2003) 1015–1018.
[36] L.A. Kelley, M.J. Sternberg, Protein structure prediction on the Web: a casestudy using the Phyre server, Nature Protocols 4 (2009) 363–371.
[37] R.A. Laskowski, M.W. MacArthur, D. Moss, J.M. Thornton, Procheck – a programto check the stereochemical quality of protein structures, Journal of AppliedCrystallography 26 (1993) 283–291.
[38] R.W. Hooft, G. Vriend, C. Sander, E.E. Abola, Errors in protein structures, Nature381 (1996) 272.
[39] J.U. Bowie, R. Lüthy, D.A. Eisenberg, A method to identify protein sequencesthat fold into a known three-dimensional structure, Science 253 (1991) 164–170.
[40] J.M. Thornton, M.S. Edwards, W.R. Taylor, D.J. Barlow, Location of ‘continuous’antigenic determinants in the protruding regions of proteins, EMBO Journal 5(2) (1986) 409–413.
[41] U.K. Laemmli, Cleavage of structural proteins during the assembly of the headof bacteriophage T4, Nature 227 (1970) 680–685.
[42] L.J. González, L. Castellanos-Serra, V. Badock, M. Díaz, A. Moro, S. Perea, et al.,Identification of nuclear proteins of small cell lung cancer cell line H82: animproved procedure for the analysis of silver-stained proteins, Electrophoresis24 (2003) 237–252.
[43] A. Tramontano, Homology modeling with low sequence identity, Methods 14(1998) 293–300.
[44] A.J. Nappi, E. Vass, F. Frey, Y. Carton, Nitric oxide involvement in Drosophilaimmunity, Nitric Oxide 4 (4) (2000) 423e30.
[45] G. Jiang, R. Yu, M. Zhou, Studies on nitric oxide synthase activity in hemocytesof shrimps Fenneropenaeus chinensis and Marsupenaeus japonicus after whitespot syndrome virus infection, Nitric Oxide 14 (3) (2006) 219–227.
[46] F.C. Yeh, S.H. Wu, C.Y. Lai, C.Y. Lee, Demonstration of nitric oxide synthaseactivity in crustacean hemocytes and anti-microbial activity of hemocyte-derived nitric oxide, Comparative Biochemistry and Physiology. Part B 144 (1)(2006) 11–17.
[47] A. Franchini, A. Conte, E. Ottaviani, Nitric oxide: an ancestral immunocyteeffector molecule, Advances in Neuroimmunology 5 (1995) 463–478.
[48] T. Rodríguez-Ramos, E. Espinosa, J. Hernández-López, T. Gollas-Galván, J.Marrero, Y. Borrell, et al., Effects of Echerichia coli lipopolysaccharides anddissolved ammonia on immune response in southern white shrimpLitopenaeus schmitti, Aquaculture 274 (2008) 118–125.
[49] N.A. Ratcliffe, A.F. Rowley, S.W. Fitzgerald, C.P. Rhodes, Invertebrate immunity:basic concepts and recent advances, International Review of Cytology 97(1985) 183–350.
[50] K. Söderhäll, L. Cerenius, Crustacean immunity, Annual Review of Fish Diseases(1992) 3–23.
[51] E. Ottaviani, L.R. Paemen, P. Cadet, G.B. Stefano, Evidence for nitric oxideproduction and utilization as a bacteriocidal agent by invertebrate
immunocytes, European Journal of Pharmacology: Environment Toxicologyand Pharmacology 248 (1993) 319–324.
[52] T. Raman, M. Arumugam, P. Mullainadhan, Agglutinin-mediated phagocytosisassociated generation of superoxide anion and nitric oxide by the hemocytesof the giant freshwater prawn Macrobrachium rosenbergii, Fish & ShellfishImmunology 24 (3) (2008) 337–345.
[53] J.A.J. Arts, A.J. Taverne-Thiele, H.F.J. Savelkoul, J.H.W.M. Rombout, Haemocytereactions in WSSV immersion infected Penaeus monodon, Fish & ShellfishImmunology 23 (1) (2007) 164–170.
[54] Yu.-Chi.Wang. YCh, Chang. PSh, H.Y. Chen, Tissue expressions of nine genesimportant to immune defence of the Pacific white shrimp Litopenaeusvannamei, Fish & Shellfish Immunology 23 (2007) 1161–1177.
[55] J. Weiske, A. Wiesner, Stimulation of NO synthase activity in theimmunecompetent Lepidopteran Estigmene acraea hemocyte line, NitricOxide: Biology and Chemistry 3 (2) (1999) 123–131.
[56] D.R. Rosa, M. Barracco, Shrimp interferon is rather a portion of themitochondrial F0-ATP synthase than a true alpha-interferon, MolecularImmunology 45 (12) (2008) 3490–3493.
[57] M. Imamura, J. Yang, M. Yamakawa, CDNA cloning, characterization and geneexpression of nitric oxide synthase from the silkworm, Bombyx mori, InsectMolecular Biology 11 (2002) 257–265.
[58] S. Luckhart, Y. Vodovotz, L.W. Cui, R. Rosenberg, The mosquito Anophelesstephensi limits malaria parasite development with inducible synthesis ofnitric oxide, Proceedings of the National Academy of Sciences of the UnitedStates of America 95 (10) (1998) 5700–5705.
[59] E. Foley, P.H. O’Farrell, Nitric oxide contributes to induction of innate immuneresponses to gram negative bacteria in Drosophila, Genes Development 17 (1)(2003) 115–125.
[60] D. Li, E.Y. Hayden, K. Panda, D.J. Stuehr, H. Deng, D.L. Rousseau, S.-R. Yeh,Regulation of the Monomer-Dimer Equilibrium in Inducible Nitric-oxideSynthase by Nitric Oxide, The Journal of Biological Chemistry 281 (12)(2006) 8197–8204.
[61] W.K. Alderton, C.E. Cooper, R.G. Knowles, Nitric oxide synthases: structure,function and inhibition, Biochemistry Journal 357 (2001) 593–615.
[62] B.R. Crane, A.S. Arvai, D.K. Ghosh, C. Wu, E.D. Getzoff, D.J. Stuehr, J.A. Tainer,Structure of nitric oxide synthase oxygenase dimer with pterin and substrate,Science 279 (1998) 2121–2126.
[63] C.S. Raman, H. Li, P. Martasek, V. Kral, S.S.B. Masters, T.L. Poulos, CrystalStructure of constitutive endothelial nitric oxide synthase: A paradigm forpterin function involving a novel metal center, Cell 95 (1998) 939–950.
[64] H. Li, T.L. Poulos, Structure–function studies on nitric oxide synthases, Journalof Inorganic Biochemistry 99 (2005) 293–305.
[65] N. Andreakis, S. D́Aniello, R. Albalat, F.P.. Patti, J. García-Fernández, G.Procaccini, P. Sordino, A. Palumbo, Evolution of the Nitric Oxide SynthaseFamily in Metazoans, Molecular Biology and Evolution 28 (1) (2011) 163–179.
[66] P.A. Loughran, D.B. Stolz, Y. Vodovotz, S.C. Watkins, R.L. Simmons, T.R. Billiar,Monomeric inducible nitric oxide synthase localizes to peroxisomes inhepatocytes, PNAS 102 (39) (2005) 13837–13842.
[67] N.L. Scholz, J.S. Labenia, J. De Vente, K. Graubard, M. Goy, Expression of nitricoxide synthase and nitric oxide-sensitive guanylate cyclase in the crustaceancardiac ganglion, The Journal of Comparative Neurology 454 (2) (2002) 158–167.




















