
The sensory basis of feeding behaviour in the Caribbean spiny lobster, Panulirus argus
12
Mar. Freshwater Res., 2001, 52, 1339–50 © CSIRO 2001 10.1071/MF01099 1323-1650/01/081339 The sensory basis of feeding behaviour in the Caribbean spiny lobster, Panulirus argus Charles D. Derby, Pascal Steullet, Amy J. Horner, and Holly S. Cate Department of Biology and Center for Behavioral Neuroscience, Georgia State University, Atlanta, Georgia 30302-4010, USA. email: [email protected] Abstract. A complex nervous system enables spiny lobsters to have a rich behavioural repertoire. The present paper discusses the ways in which the sensory systems of the Caribbean spiny lobster, Panulirus argus, particularly its chemosensory systems, are involved in feeding behaviour. It addresses the neural mechanisms of three aspects of their food-finding ability: detection, identification, and discrimination of natural food odours; the effect of learning on responses to food odours; the mechanisms by which spiny lobsters orient to odours from a distance under natural flow conditions. It demonstrates that the olfactory organ of spiny lobsters might use across- neuron response patterns in discriminating odour quality; that the hedonic value of food can be modified by experience, including associative and nonassociative conditioning; that spiny lobsters can readily orient to distant odour sources; and that both chemo- and mechanosensory antennular input are important in this behaviour. Either aesthetasc or nonaesthetasc chemosensory pathways can be used in identifying odour quality, mediating learned behaviours, and permitting orientation to the source of distant odours. Studying the neuroethology of feeding behaviour helps us understand how spiny lobsters are adapted to living in complex and variable environments. Extra keywords : Crustacea, olfaction, chemical sense, chemoreception, mechanoreception, orientation, discrimination, learning Introduction The nervous system of spiny lobsters contains an enormous number and diversity of neurons. For example, receptor neurons number well over 1 million (Grünert and Ache 1988; Laverack 1988a; Cate and Derby, 2000, 2001), and the brain has many sensory and integrative centres, each with a different, complex organization (Maynard 1966; Sandeman 1990; Sandeman et al. 1992; Mellon and Alones 1993; Schmidt and Ache 1996a, 1996b). These animals also perform a rich array of behaviours, show extensive behavioural plasticity, and have impressive learning abilities (Winn and Olla 1972; Krasne 1973; Abramson and Feinman 1990; Feinman et al. 1990; Pereyra et al. 1999; Derby 2000). Studying their nervous system and behavioural abilities, and the causal links between the two, gives us a better understanding of how spiny lobsters and other crustaceans have adapted to living in complex and variable environments. The chemosensory pathways are a prominent part of the nervous system of spiny lobsters and other crustaceans. The first antennae (antennules) alone have over 500,000 chemosensory neurons (Grünert and Ache 1988; Steullet et al. 2000a; Cate and Derby 2001), and six paired neuropils (olfactory lobes, lateral antennular neuropils, median antennular neuropils, accessory lobes, terminal medullae, and hemiellipsoid bodies) are directly or indirectly connected to this antennular input (Sandeman et al. 1992; Schmidt and Ache 1996a, 1996b). It is therefore not surprising that spiny lobsters use their chemical senses in many behaviours, including feeding (Reeder and Ache 1980; Zimmer-Faust et al. 1996) and shelter selection (Ratchford and Eggleston 1998, 2000; Zimmer-Faust 1987). Whether they also use their chemical senses in larval settlement, avoiding predators, mate selection, social interactions among conspecifics, or other intra- and interspecific interactions remains to be determined, although these functions have been demonstrated in other crustaceans (Caldwell and Dingle 1985; Carr 1988; Gleeson 1991; Hazlett 1994a; Hazlett and Schoolmaster 1998; Karavanich and Atema 1998a, 1998b; Zulandt Schneider and Moore 1999, 2000). Feeding behaviour is the best studied of the chemosensory behaviours in spiny lobsters, especially in terms of sensory mechanisms. The present paper summarizes our current understanding of this topic.
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of The sensory basis of feeding behaviour in the Caribbean spiny lobster, Panulirus argus

Mar. Freshwater Res.
,
2001,
52
,
1339–50
© CSIRO 2001 10.1071/MF01099 1323-1650/01/081339
The sensory basis of feeding behaviour in the Caribbean spiny lobster,
Panulirus argus
Charles D. Derby, Pascal Steullet, Amy J. Horner, and Holly S. Cate
Department of Biology and Center for Behavioral Neuroscience, Georgia State University, Atlanta, Georgia 30302-4010, USA. email: [email protected]
Abstract.
A complex nervous system enables spiny lobsters to have a rich behavioural repertoire. The presentpaper discusses the ways in which the sensory systems of the Caribbean spiny lobster,
Panulirus
argus
,particularly its chemosensory systems, are involved in feeding behaviour. It addresses the neural mechanisms ofthree aspects of their food-finding ability: detection, identification, and discrimination of natural food odours; theeffect of learning on responses to food odours; the mechanisms by which spiny lobsters orient to odours from adistance under natural flow conditions. It demonstrates that the olfactory organ of spiny lobsters might use across-neuron response patterns in discriminating odour quality; that the hedonic value of food can be modified byexperience, including associative and nonassociative conditioning; that spiny lobsters can readily orient to distantodour sources; and that both chemo- and mechanosensory antennular input are important in this behaviour. Eitheraesthetasc or nonaesthetasc chemosensory pathways can be used in identifying odour quality, mediating learnedbehaviours, and permitting orientation to the source of distant odours. Studying the neuroethology of feedingbehaviour helps us understand how spiny lobsters are adapted to living in complex and variable environments.
Extra
keywords
:
Crustacea, olfaction, chemical sense, chemoreception, mechanoreception, orientation,
discrimination, learning
Introduction
The nervous system of spiny lobsters contains an enormousnumber and diversity of neurons. For example, receptorneurons number well over 1 million (Grünert and Ache1988; Laverack 1988
a
; Cate and Derby, 2000, 2001), andthe brain has many sensory and integrative centres, eachwith a different, complex organization (Maynard 1966;Sandeman 1990; Sandeman
et
al
. 1992; Mellon and Alones1993; Schmidt and Ache 1996
a
, 1996
b
). These animals alsoperform a rich array of behaviours, show extensivebehavioural plasticity, and have impressive learningabilities (Winn and Olla 1972; Krasne 1973; Abramson andFeinman 1990; Feinman
et
al
. 1990; Pereyra
et
al
. 1999;Derby 2000). Studying their nervous system andbehavioural abilities, and the causal links between the two,gives us a better understanding of how spiny lobsters andother crustaceans have adapted to living in complex andvariable environments.
The chemosensory pathways are a prominent part of thenervous system of spiny lobsters and other crustaceans. Thefirst antennae (antennules) alone have over 500,000chemosensory neurons (Grünert and Ache 1988; Steullet
etal
. 2000
a
; Cate and Derby 2001), and six paired neuropils
(olfactory lobes, lateral antennular neuropils, medianantennular neuropils, accessory lobes, terminal medullae,and hemiellipsoid bodies) are directly or indirectlyconnected to this antennular input (Sandeman
et
al
. 1992;Schmidt and Ache 1996
a
, 1996
b
).It is therefore not surprising that spiny lobsters use their
chemical senses in many behaviours, including feeding(Reeder and Ache 1980; Zimmer-Faust
et
al
. 1996) andshelter selection (Ratchford and Eggleston 1998, 2000;Zimmer-Faust 1987). Whether they also use their chemicalsenses in larval settlement, avoiding predators, mateselection, social interactions among conspecifics, or otherintra- and interspecific interactions remains to bedetermined, a l though these funct ions have beendemonstrated in other crustaceans (Caldwell and Dingle1985; Carr 1988; Gleeson 1991; Hazlett 1994
a
; Hazlett andSchoolmaster 1998; Karavanich and Atema 1998
a
, 1998
b
;Zulandt Schneider and Moore 1999, 2000). Feedingbehaviour is the best studied of the chemosensorybehaviours in spiny lobsters, especially in terms of sensorymechanisms. The present paper summarizes our currentunderstanding of this topic.

1340 Charles D. Derby
et al.
To use its chemical senses to find food, an animal mustsuccessfully perform several tasks involving sensory,integrative, and motor functions. First, it must identify thenature of the chemical signals. It must detect and distinguishrelevant chemical signals against background chemicals,and it must determine the qualities (
i
.
e
., molecularstructures), quantities (
i
.
e
., concentrations), and locations(
i
.
e
., spatiotemporal dynamics) of these signals. Second, itmust decide whether or not to search for the sources of thosesignals. To do so, it must compare this chemosensory inputwith innate or learned neural templates and makebehavioural choices whether or not to respond, and if so,how to respond. Third, it must locate the source of thesignals by orienting to them from a distance using availablecues, such as the spatiotemporal distribution of chemicaland hydrodynamic cues.
Here, we review work that shows that spiny lobsters canaccomplish all of these tasks. We focus on the Caribbeanspiny lobster,
Panulirus
argus
, which has been an organismof choice in neurobiological studies of the chemical senses.First, we describe mechanisms of detection, identification,and discrimination of behaviourally relevant chemicalsignals, especially complex food-related mixtures. Second,we show that spiny lobsters can change their responses tochemical signals in adaptive ways, as a result of experiencessuch as associative learning. Third, we explain sensorymechanisms involved in orientation toward distant chemicalsignals under natural flow conditions.
Detection and discrimination of chemical signals
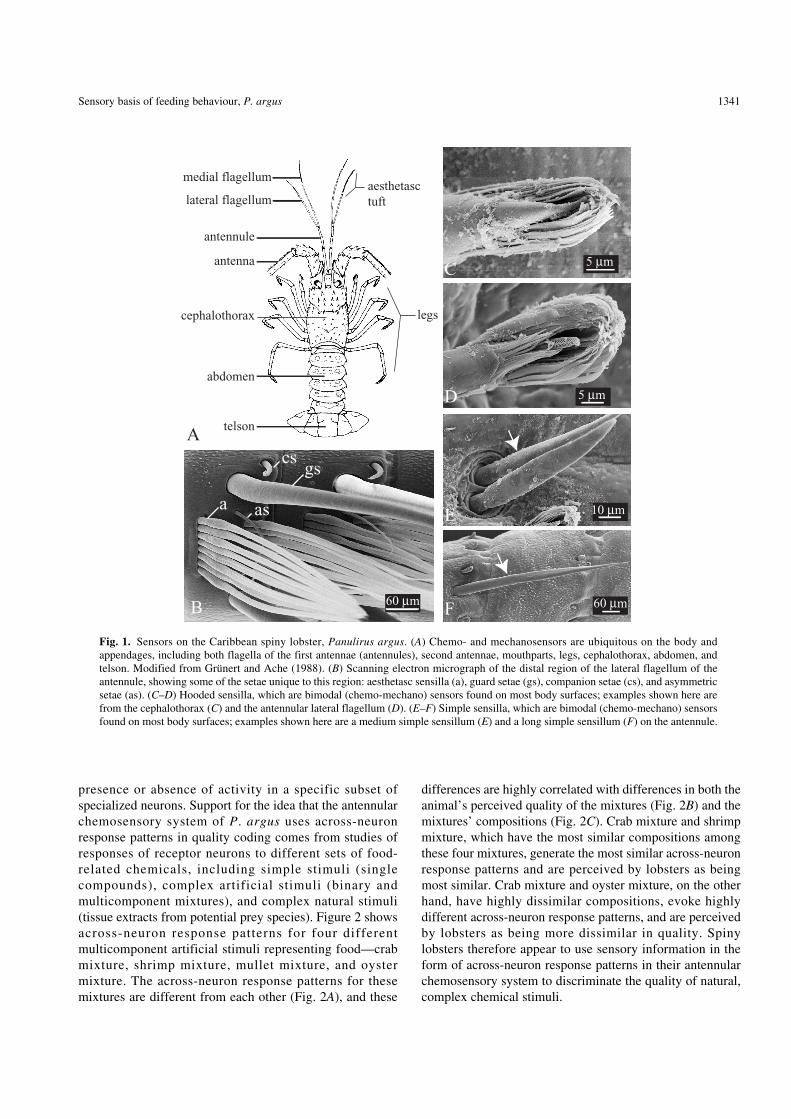
The first tasks that spiny lobsters must accomplish duringfeeding—detecting, identifying, and discriminating thenature of chemical cues—are accomplished through theirimpressive chemosensory systems. Chemical sensors arepresent on most body surfaces, especially the first antennae(= antennules), second antennae, legs, and mouthparts, butalso the cephalothorax, abdomen, and telson (Fig. 1
A
)(Laverack 1964, 1988
b
; Grünert and Ache 1988; Derby1989; Gleeson
et
al
. 1993; Cate and Derby 2000, 2001). Thechemosensors are organized as sensilla, which are cuticularextensions of the body surface that are innervated by thedendrites of chemosensory neurons. The aesthetascs are aprominent type of chemosensor that are located only on thedistal half of the antennular lateral flagella; they are the onlyknown unimodal chemosensors (Laverack 1964; Grünertand Ache 1988; Derby 1989; Cate and Derby 2001) (Figs1
A
and 1
B
). All of the other known chemosensilla on
P
.
argus
a re b imodal , be ing innervated by bothchemosensory neurons and mechanosensory neurons (Cateand Derby 2000, 2001). These include hooded sensilla (Figs1
C
and 1
D
) and simple sensilla (Figs 1
E
and 1
F
). Thehooded sensilla are a particularly interesting sensillar type,because they are abundantly located over most of theanimal’s body, including the lateral and medial antennular
flagella, antennae, legs, mouthparts, cephalothorax,abdomen, and telson (Cate and Derby 2000, 2001).
Different chemosensors act sequentially during thefeeding behaviour of lobsters: antennular chemoreceptorsinitiate searching and orientation toward the source of adistant chemical stimulus (Reeder and Ache 1980; Devineand Atema 1982; Steullet
et
al
. 2000
b
); leg chemoreceptorscon t ro l loca l g rasp ing re f l exes ; and mouthpar tchemoreceptors mediate the decision to ingest food (Derbyand Atema 1982). The roles of the chemoreceptors on thecephalothorax and abdomen have not been examined.
The neural basis for discrimination of food odours hasbeen extensively studied in the chemosensory neurons of theantennules. The first step in this process—the transductionof chemical information into electrical signals by antennularreceptor neurons—is known in cellular and molecular detailthat reveals the function of receptor proteins, G-proteins,second-messenger cascades, ion channels, transporters, andenzymes in detecting odorant molecules (Carr
et
al
. 1990;Ache and Zhainazarov 1995; Olson and Derby 1995;McClintock
et
al
. 1997; Xu
et
al
. 1998; Zhainazarov andAche 1999; Munger
et
al
. 2000). Beyond chemosensorytransduction, how chemical quality and quantity areencoded by the chemosensory neurons of the antennules hasalso been well studied in lobsters, both the spiny lobster
P
.
argus
and the American lobster
Homarus
americanus
(for reviews, see Atema
et
al
. 1989; Derby
et
al
. 1989;Derby and Atema 1988; Derby 2000). Each antennule hashundreds of thousands of chemoreceptor neurons (Grünertand Ache 1988; Cate and Derby 2000, 2001). The antennuleas a whole is sensitive to many different odorants,particularly to small, water-soluble molecules such as aminoacids, amines, nucleotides, and sometimes sugars andpeptides (Carr 1988). Yet each antennular chemoreceptorneuron responds to a limited subset of these chemicalstimulants. Individual neurons are not specifically tuned torespond to only one complex stimulus; most cells respond todifferent degrees to all tested complex food mixtures(Girardot and Derby 1990
b
). The responsiveness ofindividual neurons to the components of these mixtures isoften complex: typically, each neuron is excited by someodour compounds, is inhibited by others, and givescomplex, nonlinear, often unpredictable responses tocombinations of odour compounds (Ache 1989; Atema
et
al
.1989; Derby
et
al
. 1989; Cromarty and Derby 1998; Derbyand Atema 1988; Derby 2000). Thus, chemoreceptorneurons of spiny lobsters are complex peripheral processingunits.
How then is chemical quality determined? The likelyanswer is ‘across-neuron response patterns’, also called‘distributed’ or ‘ensemble’ neural response patterns. Anacross-neuron pattern distinguishes one odorant fromanother by differences in the relative activity among most orall members of a neuronal population, rather than by the
Sensory basis of feeding behaviour,
P. argus
1341
presence or absence of activity in a specific subset ofspecialized neurons. Support for the idea that the antennularchemosensory system of
P
.
argus
uses across-neuronresponse patterns in quality coding comes from studies ofresponses of receptor neurons to different sets of food-related chemicals, including simple stimuli (singlecompounds), complex artificial stimuli (binary andmulticomponent mixtures), and complex natural stimuli(tissue extracts from potential prey species). Figure 2 showsacross-neuron response patterns for four differentmulticomponent artificial stimuli representing food—crabmixture, shrimp mixture, mullet mixture, and oystermixture. The across-neuron response patterns for thesemixtures are different from each other (Fig. 2
A
), and these
differences are highly correlated with differences in both theanimal’s perceived quality of the mixtures (Fig. 2
B
) and themixtures’ compositions (Fig. 2
C
). Crab mixture and shrimpmixture, which have the most similar compositions amongthese four mixtures, generate the most similar across-neuronresponse patterns and are perceived by lobsters as beingmost similar. Crab mixture and oyster mixture, on the otherhand, have highly dissimilar compositions, evoke highlydifferent across-neuron response patterns, and are perceivedby lobsters as being more dissimilar in quality. Spinylobsters therefore appear to use sensory information in theform of across-neuron response patterns in their antennularchemosensory system to discriminate the quality of natural,complex chemical stimuli.
medial flagellum
lateral flagellum
antennule
antenna
cephalothorax legs
abdomen
aesthetasc tuft
telsonA
F
E
D
C
B
a as
gscs
10 µm
60 µm
5 µm
5 µm
60 µm
Fig. 1.
Sensors on the Caribbean spiny lobster,
Panulirus
argus
. (
A
) Chemo- and mechanosensors are ubiquitous on the body andappendages, including both flagella of the first antennae (antennules), second antennae, mouthparts, legs, cephalothorax, abdomen, andtelson. Modified from Grünert and Ache (1988). (
B
) Scanning electron micrograph of the distal region of the lateral flagellum of theantennule, showing some of the setae unique to this region: aesthetasc sensilla (a), guard setae (gs), companion setae (cs), and asymmetricsetae (as). (
C–D
) Hooded sensilla, which are bimodal (chemo-mechano) sensors found on most body surfaces; examples shown here arefrom the cephalothorax (
C
) and the antennular lateral flagellum (
D
). (
E–F
) Simple sensilla, which are bimodal (chemo-mechano) sensorsfound on most body surfaces; examples shown here are a medium simple sensillum (
E
) and a long simple sensillum (
F
) on the antennule.

1342 Charles D. Derby
et al.
(A) Neural Distributed Codes
(B) Behavioral Discrimination
(C) Composition
Fig. 2. Spiny lobsters might use across-neuron response patterns to assess differences in stimulus quality of four complex food-related chemicalmixtures. Stimuli are crab, shrimp, oyster, and mullet mixtures, which are 41-component artificial mixtures containing amino acids, amines,nucleotides, nucleosides, quaternary ammonium compounds, and organic acids at the concentrations that each occurs in the tissue fromrepresentative species (Carr 1988). (A) Relative similarities between mixtures according to across-neuron response patterns for a population of 30receptor neurons in the lateral antennular flagellum. Electrophysiological responses were recorded for each of the 30 neurons to the four mixtures,each at three concentrations (C1, S1, M1, and O1 are crab, shrimp, and oyster mixtures at 5 µM, respectively; C2, S2, M2, and O2 are these mixturesat 50 µM; and C3, S3, M3, and O3 are at 500 µM); responses were quantified as number of action potentials during 5 s of stimulation. The similaritybetween the across-neuron pattern responses of these 30 cells to each pair of mixtures was quantified by means of squared Euclidean distances; thispairwise analysis for all pairs of stimuli resulted in a 12 × 12 matrix of similarity measures. This matrix was then applied to multidimensionalscaling, which is a type of multivariate analysis, with the goal of reducing this matrix to as few ‘dimensions’ as possible while still explaining >90%of the variance in the matrix. In this case, the three-dimensional solution shown in this figure did so. Each of the three dimensions explains some,though differing amounts, of the variability in the across-neuron pattern responses due to stimulus type and/or concentration; thus, in this graph, asin all multidimensional scaling, the dimensions do not have ‘labels’. In this figure, each point represents the across-neuron response pattern for onestimulus (e.g., C3 is crab mixture at 500 µM), and each shaded triangle represents the stimulus space for a two-log-unit concentration range of eachmixture (for C, this is the stimulus space represented by 5–500 µM crab mixture). In this figure, the distance between stimuli or stimulus space iscorrelated with the similarity in across-neuron response patterns; thus, stimuli close to each other have relatively similar across-neuron responsepatterns, and stimuli distant from each other have relatively dissimilar across-neuron response patterns. Thus, this figure shows that the stimulusspace (and therefore across-neuron pattern responses) for crab mixture is relatively similar to that for shrimp mixture but relatively dissimilar to thatfor oyster mixture. From Girardot and Derby (1988). (B) Relative similarities between mixtures according to behavioural studies of discrimination.The data set used in this analysis was behavioural responses of lobsters to crab mixture (CM), shrimp mixture (SM), mullet mixture (MM), andoyster mixture (OM), each at two concentrations (50 and 500 µM). The data were collected from four groups of three animals; members of eachgroup were aversively conditioned to both concentrations of a single mixture before generalization testing for the other three mixtures. Behaviouralresponses were ‘aversion values’, based on changes in appetitive and aversion responses due to aversive conditioning. Aversive conditioningfollowed by generalization testing is a standard procedure for determining discrimination abilities of animals. In this analysis, similarities in theaversion values were determined for each pair of stimuli by means of Euclidean distances, and these similarities were used to construct an 8 × 8matrix. As in Fig. 2A, this matrix of similarities was used in multidimensional scaling to simplify the data set to the minimum number of‘dimensions’ sufficient to describe >90% of the data set’s variability. In this case, a two-dimensional solution sufficed. The dimensions of thisfigure do not have labels, as explained above under Fig. 2A. In this two-dimensional solution, distances between the stimuli are correlated withperceptual similarity; stimuli close to each other are perceived by lobsters as being more similar than are stimuli farther from each other. Thus,lobsters perceive the two concentrations of crab mixture as relatively similar to each other, and relatively similar to the two concentrations of shrimpmixture, but relatively different from oyster mixture or mullet mixture. From Fine-Levy et al. (1989). (C) Relative similarities in the chemicalcompositions of the mixtures. A cluster analysis, which is another type of multivariate analysis, was used to evaluate similarities in the chemicalcompositions of the four mixtures. The similarity in the chemical composition of each pair of mixtures was evaluated from Pearson product-moment correlation coefficients as distance measures based on the concentrations of the 41 chemical components of the mixtures. A 4 × 4 matrixof these similarity values was created and used in the cluster analysis, the results of which are depicted as a dendrogram. Mixtures joined at a shortcluster distance have more similar compositions than do mixtures joined at longer distances. Thus, crab and shrimp mixtures have compositionsrelatively similar to each other but different from oyster or mullet mixtures. Only four points appear in this figure, compared to the 12 points in Figs2A and 2B, because the multivariate analysis using Pearson correlation coefficients shows that three concentrations for each mixture type areidentical. From Fine-Levy et al. (1988).
Sensory basis of feeding behaviour,
P. argus 1343
Chemical concentration can be encoded by the overallintensity of responses of the antennular chemoreceptor cells(reviewed by Derby and Atema 1988; Ache 1989; Derby etal. 1989; Derby 2000). An independence of codes forchemical type and quantity would allow constancy in theperceived quality of an odour in spite of the largefluctuations in concentrations that occur in nature. Supportfor this independence in P. argus comes from threeobservations. First, the mean response from a population ofantennular chemoreceptor neurons is highly correlated withstimulus concentration (Girardot and Derby 1988, 1990a;Daniel et al . 1996). Second, changes in st imulusconcentration by as much as two orders of magnitude have arelatively small effect on the across-neuron responsepatterns for stimulus quality (Fig. 2A) (Girardot and Derby
1990a). Third, behavioural discriminations of chemicalquality are relatively insensitive to changes in chemicalintensity (Fig. 2B) (Fine-Levy et al. 1989; Fine-Levy andDerby 1991). An alternative theory of coding of stimulusquality has been proposed for H. americanus: that bothchemical concentration and quality are encoded by across-neuron response patterns (Johnson et al. 1992; Merrill et al.1994).
Whereas across-neuron response patterns are probablyused by spiny lobsters in encoding the quality of food-associated chemicals, other chemical signals, such aspheromones, might be encoded differently. In manyanimals, pheromones are encoded by specialized receptorneurons, which function as ‘labelled lines’ rather than inacross-neuron response patterns (Ache 1991, Hildebrand
% D
iffer
ence
in S
earc
h R
espo
nse
betw
een
Con
ditio
ned
and
Unc
ondi
tione
d Lo
bste
rs
-100
-80
-60
-40
-20
0
20
Intact LobstersAesthetasc-ablated LobstersNonaesthetasc-ablated Lobsters
Crab InverseCrab
Shrimp Mullet
% D
iffer
ence
in S
earc
h R
espo
nse
betw
een
Con
ditio
ned
and
Unc
ondi
tione
d Lo
bste
rs
-40
0
40
80
120
160
99.9:0.1 99:1 90:10 50:50
Blends of AMP:taurine (x 10-2 mM)
Intact LobstersAesthetasc-ablated Lobsters
A. Generalization Conditioning to Complex Mixture B. Discrimination Conditioning to Binary Mixture
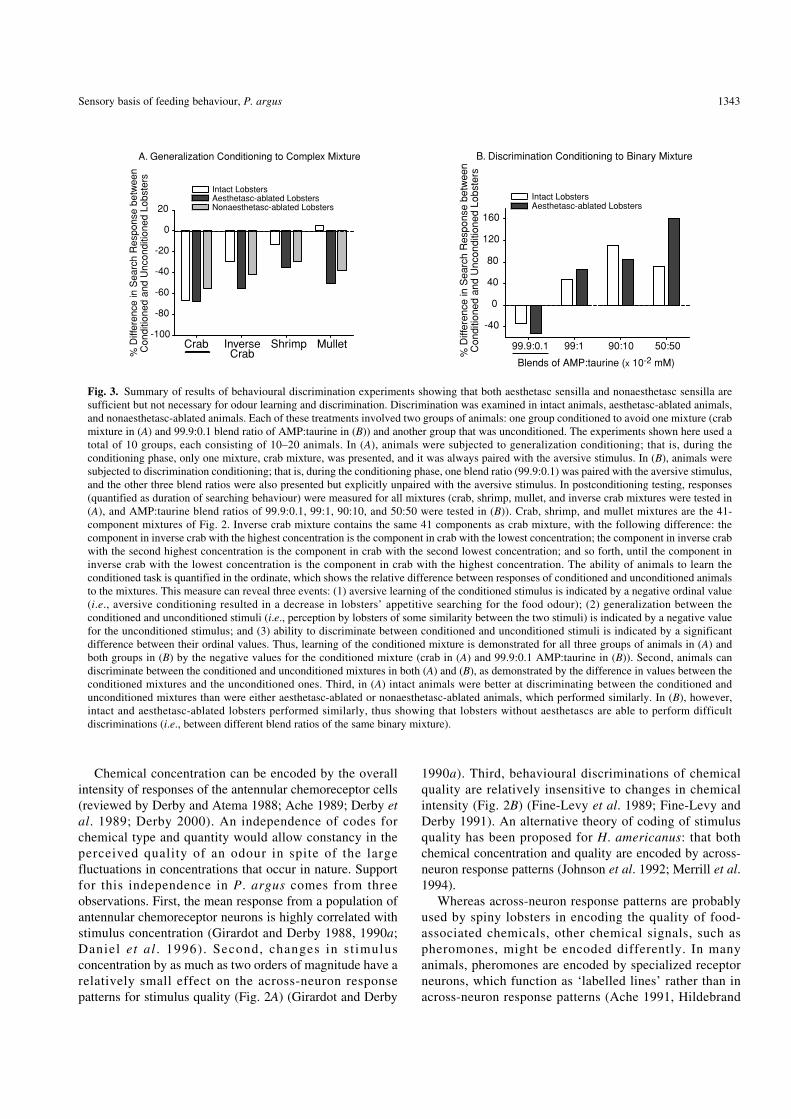
Fig. 3. Summary of results of behavioural discrimination experiments showing that both aesthetasc sensilla and nonaesthetasc sensilla aresufficient but not necessary for odour learning and discrimination. Discrimination was examined in intact animals, aesthetasc-ablated animals,and nonaesthetasc-ablated animals. Each of these treatments involved two groups of animals: one group conditioned to avoid one mixture (crabmixture in (A) and 99.9:0.1 blend ratio of AMP:taurine in (B)) and another group that was unconditioned. The experiments shown here used atotal of 10 groups, each consisting of 10–20 animals. In (A), animals were subjected to generalization conditioning; that is, during theconditioning phase, only one mixture, crab mixture, was presented, and it was always paired with the aversive stimulus. In (B), animals weresubjected to discrimination conditioning; that is, during the conditioning phase, one blend ratio (99.9:0.1) was paired with the aversive stimulus,and the other three blend ratios were also presented but explicitly unpaired with the aversive stimulus. In postconditioning testing, responses(quantified as duration of searching behaviour) were measured for all mixtures (crab, shrimp, mullet, and inverse crab mixtures were tested in(A), and AMP:taurine blend ratios of 99.9:0.1, 99:1, 90:10, and 50:50 were tested in (B)). Crab, shrimp, and mullet mixtures are the 41-component mixtures of Fig. 2. Inverse crab mixture contains the same 41 components as crab mixture, with the following difference: thecomponent in inverse crab with the highest concentration is the component in crab with the lowest concentration; the component in inverse crabwith the second highest concentration is the component in crab with the second lowest concentration; and so forth, until the component ininverse crab with the lowest concentration is the component in crab with the highest concentration. The ability of animals to learn theconditioned task is quantified in the ordinate, which shows the relative difference between responses of conditioned and unconditioned animalsto the mixtures. This measure can reveal three events: (1) aversive learning of the conditioned stimulus is indicated by a negative ordinal value(i.e., aversive conditioning resulted in a decrease in lobsters’ appetitive searching for the food odour); (2) generalization between theconditioned and unconditioned stimuli (i.e., perception by lobsters of some similarity between the two stimuli) is indicated by a negative valuefor the unconditioned stimulus; and (3) ability to discriminate between conditioned and unconditioned stimuli is indicated by a significantdifference between their ordinal values. Thus, learning of the conditioned mixture is demonstrated for all three groups of animals in (A) andboth groups in (B) by the negative values for the conditioned mixture (crab in (A) and 99.9:0.1 AMP:taurine in (B)). Second, animals candiscriminate between the conditioned and unconditioned mixtures in both (A) and (B), as demonstrated by the difference in values between theconditioned mixtures and the unconditioned ones. Third, in (A) intact animals were better at discriminating between the conditioned andunconditioned mixtures than were either aesthetasc-ablated or nonaesthetasc-ablated animals, which performed similarly. In (B), however,intact and aesthetasc-ablated lobsters performed similarly, thus showing that lobsters without aesthetascs are able to perform difficultdiscriminations (i.e., between different blend ratios of the same binary mixture).

1344 Charles D. Derby et al.
and Shepherd 1997). Neural processing of pheromones hasscarcely been studied in crustaceans, particularly thedecapods, because the molecular identity of theirpheromones is largely unknown (Gleeson 1991).
Antennular chemoreceptors provide the input that leadsto initial discrimination of stimuli and the decision whetheror not to initiate a search (Reeder and Ache 1980; Devineand Atema 1982), but the antennules bear many setal types(Fig. 1), including setae that are known to be innervated andare therefore called ‘sensilla’. Antennular chemosensillainclude aesthetascs, hooded sensilla, and several types ofsimple sensilla (Laverack 1964; Grünert and Ache 1988;Cate and Derby 2000, 2001). Other setae, whose innervationhas not been studied, include guard setae, companion setae,asymmetric setae, and plumose setae. Here we collectivelyca l l a l l an tennula r se tae o ther than aes the tascs‘nonaesthetasc setae’.
The identity of the setal types responsible for antennule-mediated odour discrimination is not obvious anddetermining it requires experimental manipulation. Weexamined this issue by studying the behaviour of spinylobsters in which specific antennular setal types (such asaesthetasc sensilla, nonaesthetasc setae, all antennular setae)had been deafferented and comparing the behaviours ofthese ablated animals with those of control animals that hadreceived no surgical treatment. We examined behaviouraldiscrimination by aversively conditioning lobsters to avoidpreviously stimulatory chemicals and subsequently testingtheir generalization to other chemical stimuli (Steullet et al.1999, 2000b). Intensity of response to odours was quantifiedas the duration of searching behaviour. Results showed thatantennular chemoreceptors are important for evoking thesesearching behaviours, because animals without themresponded significantly less than intact animals. Animalsfrom which either the aesthetasc or nonaesthetasc setae hadbeen removed still responded to the mixtures, learned theconditioning task, and discriminated between theconditioned and nonconditioned mixtures (Fig. 3), soaesthetasc chemoreceptors are sufficient but not necessaryfor lobsters to discriminate between highly related chemicalmix tu r e s . L ikewi se nonaes the t a sc an t ennu l a rchemoreceptors are sufficient but not necessary. We haveobserved the same results under a variety of experimentalconditions. These include using two stimulus sets—a set ofmulticomponent artificial food mixtures (Fig. 3A) and a setof binary mixtures containing the same components(adenosine-5’-monophosphate and taurine) but at differentblend ratios (from 99.9:0.1 to 50:50) (Fig. 3B)—and usingtwo training and discrimination tasks—generalizationconditioning (Fig. 3A) and discrimination conditioning (Fig.3B). Together, these experiments show that there is overlapin the function of the aesthetasc and nonaesthetasc receptorneurons in chemical discrimination, and this overlapfunctions in different biological contexts. There may be
differences in the discriminations mediated by theaesthetasc and nonaesthetasc systems of spiny lobsters, suchas in their responsiveness to social or sex pheromones, butthese differences have not yet been identified.
Learning about chemical signals
After identifying a chemical signal, a lobster must decidewhether to search for its source. In this respect, lobstersshow impressive behavioural plasticity; experience andlearning are important. This plasticity is expected from andbeneficial to animals such as lobsters that can live for manyyears, are omnivorous, live in many different environments,and consequently are exposed to different chemical stimulithat can assume of variety of meanings.
An example of plasticity in chemical responsiveness isthat the hedonic value of food can be modified byexperience. Spiny lobsters can learn through negativeconditioning to avoid naturally attractive chemicals, andthey can learn through positive conditioning to increasetheir attraction to other food-related chemicals. Forexample, as described in the previous section onchemosensory discrimination, spiny lobsters can beassociatively conditioned to stop responding to food odoursthat are normally excitatory and attractive if those stimuliare paired with aversive stimuli (Fine-Levy et al. 1988,1989; Lynn et al. 1994; Livermore et al. 1997). We used asthe aversive unconditioned stimulus a ‘pseudopredator’ (ablack object moved rapidly toward the animal) to simulatenatural conditions, such as a situation where lobsters areattracted to food odours but while searching are chased by apredator. The rapid learning of this conditioning task (5–10trials) and its several-day duration, together with our failuresto condition animals with unnatural aversive stimuli such aselectric shock (Fine-Levy et al. 1988), show that this is apowerful and biologically relevant form of learning. Spinylobsters can also learn by habituation to stop responding topreviously attractive food chemicals (Daniel and Derby1988). Such nonassociative learning requires many trials,however, presumably because food chemicals are normallyhighly attractive and have positive hedonic values, and it istherefore not adaptive for animals to habituate quickly toinnately positive stimuli that have not received concomitantnegative reinforcement.
Spiny lobsters can learn different features of food-relatedcomplex chemical cues, depending on the context oflearning and the available cues. A mixture might beperceived either as a combination of its individualcomponents (i.e., elemental cues) or as a mixture-uniquestimulus (i.e., configural cue). Which of these cuesdominates perception is influenced by the salience of each,including past experience with mixtures and theircomponents. Spiny lobsters are able to learn to respond toeither elemental or configural cues (Livermore et al. 1997),as can other animals from honey bees to rats (Rescorla et al.

Sensory basis of feeding behaviour, P. argus 1345
1985; Rudy and Sutherland 1992; Smith 1996). This abilityhighlights the sophistication of chemosensory processingand learning by lobsters.
Other interest ing forms of learning have beendemonstrated in some crustacean species but not in spinylobsters. These include one-trial food-aversion learning(Wight et al. 1990), learned preferences for the odour ofexperienced foods (Derby and Atema 1981; Hazlett 1994b),learning of the identity of individual conspecifics ordominance status during social encounters (Caldwell andDingle 1985; Hazlett 1994a; Hazlett and Schoolmaster1998; Karavanich and Atema 1998a, 1998b, ZulandtSchneider and Moore 1999, 2000), and other learnedbehaviours (Winn and Olla 1972; Krasne 1973; Abramson
and Feinman 1990; Feinman et al. 1990; Pereyra et al.1999).
Orientation to distant chemical signals
After sensing a food odour and deciding to search for it,spiny lobsters must locate that food using available cues.Their ability to do so has been studied in a laboratory flumethat produces controlled flow conditions mimicking thosefound in nature (Fig. 4A) (Horner et al. 2000). This studyshows that spiny lobsters can efficiently and reliably orientto odour sources 2 m away (Figs 4B and 5). Similar abilitieshave also been demonstrated in other crustaceans, includingAmerican lobsters (Moore et al. 1991; Grasso et al. 1998a),blue crabs (Weissburg and Zimmer-Faust 1993, 1994;
A. Orientation paths of intact lobsters (n = 14)
B. Orientation paths of lobsters with aesthetascsablated (n = 10)
D. Orientation paths of lobsters with non-aesthetasc chemoreceptors ablated (n = 4)
C. Orientation paths of lobsters with non-aesthetascchemo- and mechanoreceptors ablated (n = 5)
CageStimulus
5 cm/sec75 cm
200 cm
Sea Water5 cm/sec
Fig. 4. Paths of spiny lobsters in odour plumes produced in a laboratory flume. (A) A 2-m region of an 8000-L flume located at GeorgiaInstitute of Technology. The chemical stimulus was 3 gm/L shrimp extract released by a peristaltic pump at 5 cm/s, which is the same velocityat which sea water runs through the flume. The dimensions of the flume and lobster are shown to scale in all parts of this figure. A trialconsisted of placing an animal in the cage, releasing an odour from the source, and videotaping the behavioural response for 10 min with aCCD camera above the flume. (B–E) Orientation paths of individual animals from four treatment groups. The videotaped movements of eachlobster that successfully found the odour source were later digitized and plotted with Motion Analysis® software. The number of individualorientation tracks, each from a different lobster, is shown on each figure for each treatment group. (B) Intact animals: Animals with noablations. (C) Animals with aesthetascs ablated: only aesthetascs and asymmetric setae were surgically ablated; all other antennular setae(nonaesthetasc chemo/mechanoreceptors) were intact. (D) Animals with nonaesthetasc chemo- and mechanoreceptors ablated: all antennularnonaesthetasc setae were surgically ablated and/or covered with cyanoacrylate glue; aesthetasc chemoreceptors were intact. (E) Animals withnonaesthetasc chemoreceptors ablated: all antennular nonaesthetasc chemoreceptors were ablated through shaving and gluing ofnonaesthetasc sensilla in the distal region of the lateral flagella and through distilled-water ablation of those on the remainder of the flagella.The distilled-water ablation technique consists of exposing the area of interest to distilled water for 5 min, as described by Derby and Atema(1982). This technique inactivates all chemoreceptors while sparing at least some mechanoreceptor activity. These animals thus lack functionalnonaesthetasc chemoreceptors but have functional nonaesthetasc mechanoreceptors as well as aesthetasc chemoreceptors.
1346 Charles D. Derby et al.
Zimmer-Faust et al. 1995), and crayfish (Moore and Grills1999; T. Breithaupt, pers. comm.).
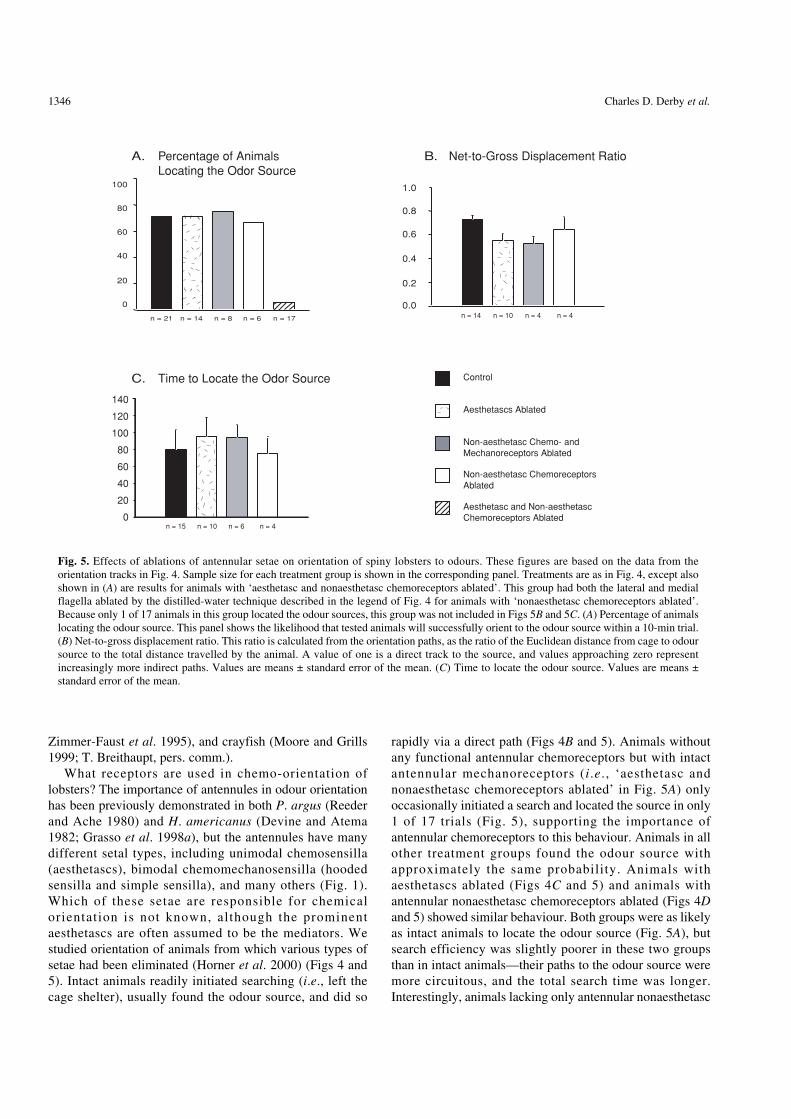
What receptors are used in chemo-orientation oflobsters? The importance of antennules in odour orientationhas been previously demonstrated in both P. argus (Reederand Ache 1980) and H. americanus (Devine and Atema1982; Grasso et al. 1998a), but the antennules have manydifferent setal types, including unimodal chemosensilla(aesthetascs), bimodal chemomechanosensilla (hoodedsensilla and simple sensilla), and many others (Fig. 1).Which of these setae are responsible for chemicalorientation is not known, although the prominentaesthetascs are often assumed to be the mediators. Westudied orientation of animals from which various types ofsetae had been eliminated (Horner et al. 2000) (Figs 4 and5). Intact animals readily initiated searching (i.e., left thecage shelter), usually found the odour source, and did so
rapidly via a direct path (Figs 4B and 5). Animals withoutany functional antennular chemoreceptors but with intactantennular mechanoreceptors (i.e., ‘aesthetasc andnonaesthetasc chemoreceptors ablated’ in Fig. 5A) onlyoccasionally initiated a search and located the source in only1 of 17 trials (Fig. 5), supporting the importance ofantennular chemoreceptors to this behaviour. Animals in allother treatment groups found the odour source withapproximately the same probability. Animals withaesthetascs ablated (Figs 4C and 5) and animals withantennular nonaesthetasc chemoreceptors ablated (Figs 4Dand 5) showed similar behaviour. Both groups were as likelyas intact animals to locate the odour source (Fig. 5A), butsearch efficiency was slightly poorer in these two groupsthan in intact animals—their paths to the odour source weremore circuitous, and the total search time was longer.Interestingly, animals lacking only antennular nonaesthetasc
1.0
0.8
0.6
0.4
0.2
0.0n = 10 n = 4n = 14 n = 4
100
80
60
40
20
0
n = 21 n = 8n = 14 n = 6 n = 17
Percentage of Animals Locating the Odor Source
A. Net-to-Gross Displacement RatioB.
Time to Locate the Odor Source
140
120
100
80
60
40
20
0n = 15 n = 6n = 10 n = 4
C. Control
Aesthetascs Ablated
Non-aesthetasc Chemo- and Mechanoreceptors Ablated
Non-aesthetasc Chemoreceptors Ablated
Aesthetasc and Non-aesthetasc Chemoreceptors Ablated
Fig. 5. Effects of ablations of antennular setae on orientation of spiny lobsters to odours. These figures are based on the data from theorientation tracks in Fig. 4. Sample size for each treatment group is shown in the corresponding panel. Treatments are as in Fig. 4, except alsoshown in (A) are results for animals with ‘aesthetasc and nonaesthetasc chemoreceptors ablated’. This group had both the lateral and medialflagella ablated by the distilled-water technique described in the legend of Fig. 4 for animals with ‘nonaesthetasc chemoreceptors ablated’.Because only 1 of 17 animals in this group located the odour sources, this group was not included in Figs 5B and 5C. (A) Percentage of animalslocating the odour source. This panel shows the likelihood that tested animals will successfully orient to the odour source within a 10-min trial.(B) Net-to-gross displacement ratio. This ratio is calculated from the orientation paths, as the ratio of the Euclidean distance from cage to odoursource to the total distance travelled by the animal. A value of one is a direct track to the source, and values approaching zero representincreasingly more indirect paths. Values are means ± standard error of the mean. (C) Time to locate the odour source. Values are means ±standard error of the mean.

Sensory basis of feeding behaviour, P. argus 1347
chemoreceptors (‘nonaesthetasc chemoreceptors ablated’:Figs 4E and 5) were better at orienting than were animalslacking both nonaesthetasc chemo- and mechanoreceptors(Figs 4D and 5). This result points to the importance ofantennular mechanoreceptors to orientation to odours inflow.
Taken together, our results on P. argus suggest thatantennules are necessary for initiating and probably formaintaining searching for distant odours and that bothantennular chemoreceptors and mechanoreceptorscontribute. The setae that mediate this behaviour aredistributed and diverse, because chemoreceptors in eitheraesthetasc or nonaesthetasc setae are sufficient but notnecessary for successful orientation behaviour. Antennularmechanoreceptors contribute to orientation to odours,presumably by providing information about flow cues. Thephysiology of the antennular neurons involved in orientationto distant chemical signals under natural flow conditions isbeginning to be understood, including their ability to resolvethe spatiotemporal distribution of chemical and mechanicalflow cues. Lobster antennular receptor neurons can followchemical stimulus pulses at frequencies as high as fourpulses per second (Marschall and Ache 1989; Gomez et al.1999). These cellular properties allow resolution of thespatiotemporal patterns of chemical signals produced intheir natural environment (Moore and Atema 1988; Atema1995; Weissburg 2000; Zimmer and Butman 2000). Evenbetter studied are assorted sensillar mechanoreceptorneurons, which have response properties that allow them tofollow dynamic changes in water flow under naturalcondit ions, including detect ing the presence andcharacteristics of eddies (Tautz and Sandeman 1980;Sandeman 1989; Breithaupt and Tautz 1990; Bleckmann etal. 1991; Weissburg 1997).
Exactly how chemo- and mechanoreceptors are used incombination in mediating orientation is not known,although several hypotheses based on work on othercrustaceans have been suggested. One proposed mechanism,based on experimental work on the clawed lobster Homarusamericanus, is that animals might orient in and find anodour source using only spatiotemporal features of chemicalcues (Moore and Atema 1988; Moore et al. 1991; Atema1996). This theory has been tested with modelling, robots,and animals (Grasso et al. 1998a, 1998b, 1999), and theresults have generally not supported it but have lead to asecond proposed mechanism—that the spatiotemporald i s t r ibu t ion of no t on ly chemica l cues bu t a l sohydrodynamic cues is used in orientation (Atema 1996;Grasso et al. 1998a, 1999; Moore and Grills 1999). Themechanism might be as relatively simple as an odour-activated rheotaxis, in which odour stimulation initiatessearch while the direction of current flow providesorientation cues (Weissburg and Zimmer-Faust 1993, 1994;Zimmer-Faust et al. 1995; T. Breithaupt, pers. comm.). The
mechanism could, however, be more complex. For example,substantial chemical and mechanical information might bepresent in the odour-scented eddies themselves that wouldindicate the location of the source (Atema 1996; Grasso eta l . 1998a , 1999; Moore and Gri l l s 1999) . Moreexperimentation is necessary to ascertain whether these orother mechanisms can explain orientation in crustaceans,including P. argus.
Acknowledgments
We thank V. Ngo, G. Hamidani, D. R. Krützfeldt, and T.Flavus for their technical assistance in the behaviouralexperiments; M. Weissburg for use of his flume at GeorgiaInstitute of Technology and T. Keller for advice andassistance in the flume studies; P. Harrison for helpfuldiscussions; and the staff of the Keys Marine Laboratory,Long Key, Florida, USA, for supplying lobsters. Thismaterial was based on work supported by the NationalScience Foundation under Grant IBN-0077474, the NationalInstitutes of Health under Grant DC00312, and the GeorgiaResearch Alliance.
References
Abramson, C. I., and Feinman, R. D. (1990). Operant conditioning incrustaceans. In ‘Frontiers in Crustacean Neurobiology’. (Eds K.Wiese, W. -D. Krenz, J. Tautz, H. Reichert, and B. Mulloney.) pp.207–14. (Birkhäuser: Basel.)
Ache, B. W. (1989). Central and peripheral bases for mixturesuppression in olfaction: a crustacean model. In ‘Perception ofComplex Smells and Tastes’. (Eds D. G. Laing, W. S. Cain, R. L.McBride, and B. W. Ache.) pp. 101–14. (Academic Press: Sydney.)
Ache, B. W. (1991). Phylogeny of smell and taste. In ‘Smell and Tastein Health and Disease’. (Eds T. V. Getchell, R. L. Doty, L. M.Bartoshuk, and J. B. Snow, Jr.) pp. 3–18. (Raven Press: NewYork.)
Ache, B. W., and Zhainazarov, A. B. (1995). Dual second-messengerpathways in olfactory transduction. Current Opinions inNeurobiology 5, 461–66.
Atema, J. (1995). Chemical signals in the marine environment:dispersal, detection, and temporal signal analysis. Proceedings ofthe National Academy of Sciences of the United States of America92, 62–66.
Atema, J. (1996). Eddy chemotaxis and odor landscapes: explorationof nature with animal sensors. Biological Bulletin (Woods Hole)191, 129–38.
Atema, J., Borroni, P. F., Johnson, B. R., Voigt, R., and Handrich, L.(1989). Adaptation and mixture interactions in chemoreceptorcells: mechanisms for diversity and contrast enhancement. In‘Perception of Complex Smells and Tastes’. (Eds D. G. Laing, W.S. Cain, R. L. McBride, and B. W. Ache.) pp. 83–100. (AcademicPress: Sydney.)
Bleckmann, H., Breithaupt, T., Blickhan, R., and Tautz, J. (1991). Thetime course and frequency content of hydrodynamic events causedby moving fish, frogs, and crustaceans. Journal of ComparativePhysiology A 168, 749–57.
Breithaupt, T., and Tautz, J. (1990). The sensitivity of crayfishmechanoreceptors to hydrodynamic and acoustic stimuli. In‘Frontiers in Crustacean Neurobiology’. (Eds K. Wiese, W. -D.Krenz, J. Tautz, H. Reichert, and B. Mulloney.) pp. 114–20.(Birkhäuser: Basel.)

1348 Charles D. Derby et al.
Caldwell, R. L., and Dingle, J. (1985). A test of individual recognitionin the stomatopod Gonodactylus festae. Animal Behaviour 33, 101–06.
Carr, W. E. S. (1988). The molecular nature of chemical stimuli in theaquatic environment. In ‘Sensory Biology of Aquatic Animals’.(Eds J. Atema, R. R. Fay, A. N. Popper, and W. N. Tavolga.) pp. 3–27. (Springer-Verlag: New York.)
Carr, W. E. S., Gleeson, R. A., and Trapido-Rosenthal, H. G. (1990).The role of perireceptor events in chemosensory processes. Trendsin Neuroscience 13, 212–15.
Cate, H. S., and Derby, C. D. (2000). A chemo-mechanosensillum thatis ubiquitous on the Caribbean spiny lobster and other lobsterspecies. Society for Neuroscience Abstracts 30, 66. 16.
Cate, H. S., and Derby, C. D. (2001). Morphology and distribution ofsetae on the antennules of the Caribbean spiny lobster Panulirusargus reveal new types of bimodal chemo-mechanosensilla. Celland Tissue Research 304, 439–54.
Cromarty, S. I., and Derby, C. D. (1998). Inhibitory receptor bindingevents among the components of complex mixtures contribute tomixture suppression in responses of olfactory receptor neurons ofspiny lobsters. Journal of Comparative Physiology A 183, 699–707.
Daniel, P. C., and Derby, C. D. (1988). Behavioral olfactorydiscrimination of mixtures in the spiny lobster (Panulirus argus)based on a habituation paradigm. Chemical Senses 13, 385–95.
Daniel, P. C., Burgess, M. F., and Derby, C. D. (1996). Responses ofolfactory receptor neurons in the spiny lobster to binary mixturesare predictable using a noncompetitive model that incorporatesexcitatory and inhibitory transduction pathways. Journal ofComparative Physiology A 178, 523–36.
Derby , C . D. (1989) . Phys io logy of sensory neurons inmorphologically identified cuticular sensilla of crustaceans. In‘Functional Morphology of Feeding and Grooming in Crustacea’.(Eds B. E. Felgenhauer, L. Watling, and A. B. Thistle.) pp. 27–47.(A. A. Balkema: Rotterdam.)
Derby, C. D. (2000). Learning from spiny lobsters aboutchemosensory coding of mixtures. Physiology and Behavior 69,203–09.
Derby, C. D., and Atema, J. (1981). Selective improvement inresponses to prey odors by the lobster, Homarus americanus,following feeding experience. Journal of Chemical Ecology 7,1073–80.
Derby, C. D., and Atema, J. (1982). The function of chemo- andmechanoreceptors in lobster (Homarus americanus) feedingbehaviour. Journal of Experimental Biology 98, 317–27.
Derby, C. D., and Atema, J. (1988). Chemoreceptor cells in aquaticinvertebrates: peripheral filtering mechanisms in decapodcrustaceans. In ‘Sensory Biology of Aquatic Animals’. (Eds J.Atema, R. R. Fay, A. N. Popper, and W. N. Tavolga.) pp. 365–88.(Springer: New York.)
Derby, C. D., Girardot, M. -N., Daniel, P. C., and Fine-Levy, J. B.(1989). Olfactory discrimination of mixtures: behavioral,electrophysiological and theoretical studies using the spiny lobsterPanulirus argus. In ‘Perception of Complex Smells and Tastes’.(Eds D. G. Laing, W. S. Cain, R. L. McBride, and B. W. Ache.) pp.65–82. (Academic Press: Sydney.)
Devine, D. V., and Atema, J. (1982). Function of chemoreceptororgans in spatial orientation of the lobster, Homarus americanus:differences and overlap. Biological Bulletin (Woods Hole) 163,144–53.
Feinman, R. D., Abramson, C. I., and Forman, R. R. (1990). Classicalcondi t ion ing in the c rab . In ‘Front ie rs in Crus taceanNeurobiology’. (Eds K. Wiese, W. -D. Krenz, J. Tautz, H. Reichert,and B. Mulloney.) pp. 215–22. (Birkhäuser: Basel.)
Fine-Levy, J. B., and Derby, C. D. (1991). Effects of stimulus intensityand quality on discrimination of odorant mixtures by spiny lobstersin an associative learning paradigm. Physiology and Behavior 49,1163–68.
Fine-Levy, J. B., Girardot, M. -N., Derby, C. D., and Daniel, P. C.(1988). Differential associative conditioning and olfactorydiscrimination in the spiny lobster Panulirus argus. Behavioral andNeural Biology 49, 315–31.
Fine-Levy, J. B., Daniel, P. C., Girardot, M. -N., and Derby, C. D.(1989). Behavioral resolution of quality of odorant mixtures byspiny lobsters: differential aversive conditioning of olfactoryresponses. Chemical Senses 14, 503–24.
Girardot, M. -N., and Derby, C. D. (1988). Neural coding of quality ofcomplex olfactory stimuli in lobsters. Journal of Neurophysiology60, 303–24.
Girardot, M. -N., and Derby, C. D. (1990a). Independent componentsof the neural population response for discrimination of quality andintensity of chemical stimuli. Brain, Behavior and Evolution 35,129–45.
Girardot, M. -N., and Derby, C. D. (1990b). Peripheral mechanisms ofolfactory discrimination of complex mixtures by the spiny lobster:no cell types for mixtures but different contributions of the cells tothe across neuron patterns. Brain Research 513, 225–36.
Gleeson, R. A. (1991). Intrinsic factors mediating pheromonecommunication in the blue crab, Callinectes sapidus . In‘Crustacean Sexual Biology’. (Eds R. T. Bauer and J. W. Martin.)pp. 17–32. (Columbia Univ. Press: New York.)
Gleeson, R. A., Carr, W. E. S., and Trapido-Rosenthal, H. G. (1993).Morphological characteristics facilitating stimulus access andremoval in the olfactory organ of the spiny lobster, Panulirusargus: insight from the design. Chemical Senses 18, 67–75.
Gomez, G., Voigt, R., and Atema, J. (1999). Temporal resolution inolfaction. III: Flicker fusion and concentration-dependentsynchronization with stimulus pulse trains of antennularchemoreceptor cells in the American lobster. Journal ofComparative Physiology A 185, 427–36.
Grasso, F. W., Basil, J. A., and Atema, J. (1998a). Toward theconvergence: robot and lobster perspectives of tracking odors totheir source in the turbulent marine environment. In ‘Proceedings,1998 IEEE/RSJ International Conference on Intelligent Robots andSystems: Innovations in Theory, Practice and Applications,October 13–17, 1998, Victoria Conference Centre, Victoria, B. C.,Canada.’ pp. 259–64. (Institute of Electrical and ElectronicsEngineers: Piscataway, New Jersey, USA.)
Grasso, F. W., Basil, J. A., and Atema, J. (1998b). Roles of lateralantennule chemo- and mechano-sensation on the chemo-orientationin the American lobster (Homarus americanus). The FifthInternational Congress of Neuroethology (San Diego, CA; August23–28, 1998). Abstract #157.
Grasso, F. W., Consi, T. R., Mountain, D. C., and Atema, J. (1999).Biomimetic robot lobster performs chemo-orientation in turbulenceusing a pair of spatially separated sensors: progress and challenges.Journal of Robotics and Autonomous Systems 807, 1–17.
Grünert, U., and Ache, B. W. (1988). Ultrastructure of the aesthetasc(olfactory) sensilla of the spiny lobster, Panulirus argus. Cell andTissue Research 251, 95–103.
Hazlett, B. A. (1994a). Alarm responses in the crayfish Orconectesvirilis and Orconectes propinquus. Journal of Chemical Ecology20, 1525–35.
Hazlett, B. A. (1994b). Crayfish feeding responses to zebra musselsdepend on microorganisms and learning. Journal of ChemicalEcology 20, 2623–30.
Hazlett, B. A., and Schoolmaster, D. R. (1998). Responses of cambaridcrayfish to predator odor. Journal of Chemical Ecology 24, 1757–70.

Sensory basis of feeding behaviour, P. argus 1349
Hildebrand, J. G., and Shepherd, G. M. (1997). Mechanisms ofolfactory discrimination: converging evidence for commonprinciples across phyla. Annual Review of Neuroscience 20, 595–631.
Horner, A. J., Ngo, V., Steullet, P., Keller, T., Weissburg, M. J., andDerby, C. D. (2000). The role of different types of antennularsensilla in orientation by Caribbean spiny lobsters to a natural odorstimulus under controlled flow conditions. Chemical Senses 25,670 (abstract.)
Johnson, B. R., Voigt, R., Merrill, C. L., and Atema, J. (1992). Across-fiber patterns may contain a sensory code for stimulus intensity.Brain Research Bulletin 26, 327–31.
Karavanich, C., and Atema, J. (1998a). Individual recognition andmemory in lobster dominance. Animal Behaviour 56, 1553–60.
Karavanich, C., and Atema, J. (1998b). Olfactory recognition of urinesignals in dominance fights between male lobster, Homarusamericanus. Behaviour 135, 719–30.
Krasne, F. B. (1973). Learning in Crustacea. In ‘Invertebrate Learning.Vol. 2: Arthropods and Gastropod Mollusks’. (Eds W. C. Corning,J. A. Dyal, and A. O. D. Willows.) pp. 49–130. (Plenum Press:New York.)
Laverack, M. S. (1964). The antennular sense organs of Panulirusargus. Comparative Biochemistry and Physiology 13, 301–21.
Laverack, M. S. (1988a). The numbers of neurons in decapodCrustacea. Journal of Crustacean Biology 8, 1–11.
Laverack, M. S. (1988b). The diversity of chemoreceptors. In ‘SensoryBiology of Aquatic Animals’. (Eds J. Atema, R. R. Fay, A. N.Popper, and W. N. Tavolga.) pp. 287–312. (Springer-Verlag: NewYork.)
Livermore, B. A., Hutson, M., Ngo, V., Hadjisimos, R., and Derby, C.D. (1997). Elemental and configural learning and the perception ofodorant mixtures by the spiny lobster Panulirus argus. Physiologyand Behavior 62, 169–74.
Lynn, W. H., Meyer, E. A., Peppiatt, C. E., and Derby, C. D. (1994).Perception of odor mixtures by the spiny lobster Panulirus argus.Chemical Senses 19, 331–47.
McClintock, T. S., Xu, F., Quintero, J., Gress, A. M., and Landers, T.M. (1997). Molecular cloning of a lobster Gα-q protein expressedin neurons of olfactory organ and brain. Journal of Neurochemistry68, 2248–54.
Marschall, H. -P., and Ache, B. W. (1989). Response dynamics oflobster olfactory neurons during simulated natural sampling.Chemical Senses 14, 725 (abstract.)
Maynard, D. M. (1966). Integration in crustacean ganglia. Symposiumof the Society for Experimental Zoology 20, 111–49.
Mellon, D., Jr., and Alones, V. (1993). Cellular organization andgrowth-related plasticity of the crayfish olfactory midbrain.Microscopy Research and Technique 24, 231–59.
Merrill, C. L., Voigt, R., and Atema, J. (1994). Reliability ofchemoreceptor cell response. I. Intensity coding by pattern andresponse magnitude with a comparison of analytical methods.Journal of Comparative Physiology A 175, 95–105.
Moore, P., and Atema, J. (1988). A model of a temporal filter inchemoreception to extract directional information from a turbulentodor plume. Biological Bulletin (Woods Hole) 174, 355–63.
Moore, P. A., and Grills, J. L. (1999). Chemical orientation to food bythe crayfish Orconectes rusticus: influence of hydrodynamics.Animal Behaviour 58, 953–963.
Moore, P. A., Scholz, N., and Atema, J. (1991). Chemical orientationof lobster, Homarus americanus, in turbulent odor plumes. Journalof Chemical Ecology 17, 1293–307.
Munger, S. D., Gleeson, R. A., Aldrich, H. C., Rust, N. C., Ache, B.W., and Greenberg, R. M. (2000). Characterization of aphosphoinositide-mediated odor transduction pathway revealsplasma membrane localization of an inositol 1,4,5-trisphosphate
receptor in lobster olfactory receptor neurons. Journal ofBiological Chemistry 275, 20450–57.
Olson, K. S., and Derby, C. D. (1995). Inhibition of taurine and 5’AMP olfactory receptor sites of the spiny lobster Panulirus argusby odorant compounds and mixtures. Journal of ComparativePhysiology A 176, 527–40.
Pereyra, P., Saraco, M., and Maldonado, H. (1999). Decreasedresponse or alternative defensive strategies in escape: two differenttypes of long-term memory in the crab Chasmagnathus. Journal ofComparative Physiology A 184, 301–10.
Ratchford, S. G., and Eggleston, D. B. (1998). Size- and scale-dependent chemical attraction contribute to an ontogenetic shift insociality. Animal Behaviour 56, 1027–34.
Ratchford, S. G., and Eggleston, D. B. (2000). Temporal shift in thepresence of a chemical cue contributes to a diel shift in sociality.Animal Behaviour 59, 793–99.
Reeder, P. B., and Ache, B. W. (1980). Chemotaxis in the Floridaspiny lobster, Panulirus argus. Animal Behaviour 28, 831–39.
Rescorla, R. A., Grau, J. W., and Durlach, P. J. (1985). Analysis of theunique cue in configural discrimination. Journal of ExperimentalPsychology 11, 356–66.
Rudy, J. W., and Sutherland, R. J. (1992). Configural and elementalassociations and the memory of coherence problem. Journal ofComparative Neuroscience 4, 208–16.
Sandeman, D. (1990). Structural and functional levels in theorganization of decapod crustacean brains. In ‘Frontiers inCrustacean Neurobiology’. (Eds K. Wiese, W. -D. Krenz, J. Tautz,H. Reichert, and B. Mulloney.) pp. 223–39. (Birkhäuser: Basel.)
Sandeman, D., Sandeman, R., Derby, C., and Schmidt, M. (1992).Morphology of the brain of crayfish, crabs, and spiny lobsters: acommon nomenclature for homologous structures. BiologicalBulletin (Woods Hole) 183, 304–26.
Sandeman, D. C. (1989). Physical properties, sensory receptors andtactile reflexes of the antenna of the Australian freshwater crayfishCherax destructor. Journal of Experimental Biology 141, 197–217.
Schmidt, M., and Ache, B. W. (1996a). Processing of antennular inputin the brain of the spiny lobster, Panulirus argus. I. Non-olfactorychemosensory and mechanosensory pathway of the lateral andmedian antennular neuropils. Journal of Comparative Physiology A178, 579–604.
Schmidt, M., and Ache, B. W. (1996b). Processing of antennular inputin the brain of the spiny lobster, Panulirus argus. II. The olfactorypathway. Journal of Comparative Physiology A 178, 605–28.
Smith, B. H. (1996). The role of attention in learning about odorants.Biological Bulletin (Woods Hole) 191, 76–83.
Steullet, P., Flavus, T., Radman, D., Hamidani, G., Zhou, M., Dudar,O., Hill, R., and Derby, C. D. (1999). The aesthetasc-olfactory lobepathway of spiny lobsters is not necessary for odor-activatedsearching behavior, odor-associate learning, and discrimination ofcomplex odors. Chemical Senses 24, 613 (abstract.)
Steullet, P., Cate, H. S., Michel, W. C., and Derby, C. D. (2000a).Functional units of a compound nose: aesthetasc sensilla housesimilar populations of olfactory receptor neurons on the crustaceanantennule. Journal of Comparative Neurology 418, 270–80.
Steullet, P., Kruetzfeldt, D. R., Hamidani, G., Flavus, T., and Derby, C.D. (2000b). Functional overlap of two antennular chemosensorypathways in food odor discrimination behavior of spiny lobsters.Chemical Senses 25, 671 (abstract.)
Tautz, J., and Sandeman, D. C. (1980). The detection of waterbornevibration by sensory hairs on the chelae of the crayfish. Journal ofExperimental Biology 88, 351–56.
Weissburg, M. J. (1997). Chemo- and mechanosensory orientation bycrustaceans in laminar and turbulent flows: from odor trails tovortex streets. In ‘Orientation and Communication in Arthropods’.(Ed. M. Lehrer.) pp. 215–46. (Birkhäuser: Basel.)

1350 Charles D. Derby et al.
Weissburg, M. J . (2000). The f luid dynamical context ofchemosensory behavior. Biological Bulletin (Woods Hole) 198,188–202.
Weissburg, M. J., and Zimmer-Faust, R. K. (1993). Life and death inmoving fluids: hydrodynamic effects on chemosensory-mediatedpredation. Ecology 74, 1428–43.
Weissburg, M. J., and Zimmer-Faust, R. K. (1994). Odor plumes andhow blue crabs use them in finding prey. Journal of ExperimentalBiology 197, 349–75.
Wight, K., Francis, L., and Eldridge, D. (1990). Food aversion learningby the hermit crab Pagurus granosimanus. Biological Bulletin(Woods Hole) 178, 205–09.
Winn, H. E., and Olla, B. L., Eds (1972). ‘Behavior of MarineAnimals. Vol. 1: Invertebrates.’ (Plenum Press: New York.)
Xu, F., Hollins, B., Landers, T. M., and McClintock, T. S. (1998).Molecular cloning of a lobster Gβ subunit and Gβ expression inolfactory receptor neuron dendrites and brain neuropil. Journal ofNeurobiology 36, 525–36.
Zhainazarov, A. B. , and Ache, B. W. (1999) . Effec ts ofphosphatidylinositol 4,5-bisphosphate and phosphatidylinositol 4-
phosphate on a Na+-gated nonselective cation channel. Journal ofNeuroscience 19, 2929–37.
Zimmer, R. K., and Butman, C. A. (2000). Chemical signalingprocesses in the marine environment. Biological Bulletin (WoodsHole) 198, 168–87.
Zimmer-Faust, R. K. (1987). Gregariousness and sociality in spinylobsters: implications for den habituation. Journal of ExperimentalMarine Biological and Ecology 105, 57–71.
Zimmer-Faust, R. K., Finelli, C. M., Pentcheff, N. D., and Wethey, D.S. (1995). Odor plumes and animal navigation in turbulent waterflow: a field study. Biological Bulletin (Woods Hole) 188, 111–16.
Zimmer-Faust, R. K., O’Neill, P. B., and Schar, D. W. (1996). Therelationship between predator activity state and sensitivity to preyodor. Biological Bulletin (Woods Hole) 190, 82–87.
Zulandt Schneider, R. A., and Moore, P. A. (1999). Recognition ofdominance status through chemoreception in the red swampcrayfish, Procambarus clarkii. Journal of Chemical Ecology 25,781–94.
Zulandt Schneider, R. A., and Moore, P. A. (2000). Urine as a sourceof conspecific disturbance signals in the crayfish Procambarusclarkii. Journal of Experimental Biology 203, 765–71.
http://www.publish.csiro.au/journals/mfr




















