
Computational Identification and Characterization of Novel Genes from Legumes
Upload
independentCategory
view
0download
0
Molecular Microbiology (2005)
57
(5), 1304–1317 doi:10.1111/j.1365-2958.2005.04768.x
© 2005 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd, 2005
? 2005
57
513041317
Original Article
NopP of Rhizobium sp. NGR234P. Skorpil
et al.
Accepted 9 June, 2005. *For correspondence. [email protected]; Tel. (
+
41) 22 379 3108; Fax(
+
41) 22 379 3009. Present address:
†
Universidad Central delCaribe, School of Medicine, Department of Microbiology and Immu-nology, Call Box 60-327, Bayamon, Puerto Rico 00960-6032;
‡
Department of Biology, Massachusetts Institute of Technology, Mas-sachusetts Avenue, Cambridge, MA 02139-4307, USA.
NopP, a phosphorylated effector of
Rhizobium
sp. strain NGR234, is a major determinant of nodulation of the tropical legumes
Flemingia congesta
and
Tephrosia vogelii
Peter Skorpil, Maged M. Saad, Nawal M. Boukli,
†
Hajime Kobayashi,
‡
Florencia Ares-Orpel, William J. Broughton* and William J. Deakin
Laboratoire de Biologie Moléculaire des Plantes Supérieures (LBMPS), Sciences III, Université de Genève, 30 quai Ernest-Ansermet, 1211, Genève 4, Switzerland.
Summary
Rhizobium
sp. NGR234 nodulates many plants, someof which react to proteins secreted via a type threesecretion system (T3SS) in a positive- (
Flemingiacongesta
,
Tephrosia vogelii
) or negative- (
Crotalariajuncea
,
Pachyrhizus tuberosus
) manner. T3SSs aredevices that Gram-negative bacteria use to injecteffector proteins into the cytoplasm of eukaryoticcells. The only two rhizobial T3SS effector proteinscharacterized to date are NopL and NopP of NGR234.NopL can be phosphorylated by plant kinases and weshow this to be true for NopP as well. Mutation of
nopP
leads to a dramatic reduction in nodule num-bers on
F. congesta
and
T. vogelii
. Concomitant muta-tion of
nopL
and
nopP
further diminishes nodulationcapacity to levels that, on
T. vogelii
, are lower thanthose produced by the T3SS null mutant NGR
WWWW
rhcN
.We also show that the T3SS of NGR234 secretes atleast one additional effector, which remains to beidentified. In other words, NGR234 secretes a cocktailof effectors, some of which have positive effects onnodulation of certain plants while others are per-ceived negatively and block nodulation. NopL andNopP are two components of this mix that extend theability of NGR234 to nodulate certain legumes.
Introduction
Rhizobia, soil-dwelling
a
- and
b
-proteo-bacteria, inducethe formation of nodules on many leguminous plantsincluding soybeans (
Glycine max
), common beans(
Phaseolus vulgaris
), peas (
Pisum sativum
), alfalfa (
Med-icago sativa
), etc. Some rhizobia even nodulate the non-legume
Parasponia andersonii
(
Ulmaceae
, the elm family)(Trinick and Galbraith, 1980; Pueppke and Broughton,1999). During nodule development, rhizobia are trans-formed into bacteroids that reduce atmospheric nitrogento ammonia, which is transferred to the plant in the formof amino acids. In turn, the plant provides energy ascarbohydrates to the bacteroids. Invading rhizobia have toovercome a number of obstacles. This is analogous to alock and key system: for every symbiotic lock, the legumeor rhizobia must have the appropriate key (i.e. molecularsignal). One set of keys are plant-produced phenolicsubstances that induce the synthesis of bacterial lipo-chito-oligosaccharides called Nod factors, which open thelegume door to the bacteria. Once within the root hair,other rhizobial keys are needed including surface polysac-charides and nodulation outer proteins (Nops) secretedby a type three secretion system (T3SS) (Broughton
et al
.,2000).
Type three secretion systems are well known from ani-mal and plant systems, but after their discovery in
Rhizo-bium
sp. NGR234 (hereafter NGR234) (Freiberg
et al
.,1997; Viprey
et al
., 1998) they have been found in a grow-ing number of other rhizobia including
Mesorhizobium loti
MAFF303099 (Kaneko
et al
., 2000a,b; Hubber
et al
.,2004),
Bradyrhizobium japonicum
USDA110 (Göttfert
et al
., 2001; Kaneko
et al
., 2002a,b) as well as
Rhizobiumfredii
strains USDA257 and HH103 (Krishnan
et al
., 2003;Vinardell
et al
., 2004). T3SSs are devices that deliver‘effector’ proteins directly into eukaryotic cells or secreteHrp proteins into the extracellular milieu (Perino
et al
.,1999; Rossier
et al
., 1999). The core secretion machineconsists of a minimal set of 10 conserved gene productsthat span both the inner- and outer-bacterial membrane(Hueck, 1998). A pilus establishes the link with a poreformed by the T3SS in the host cell’s plasma membrane(He
et al
., 2004). Effector proteins are then secreted

NopP of
Rhizobium
sp. NGR234
1305
© 2005 Blackwell Publishing Ltd,
Molecular Microbiology
,
57
, 1304–1317
through an extended pilus without prior proteolytic cleav-age (Ghosh, 2004). A number of animal and plant patho-gens inject effector proteins directly into their host cells(Büttner and Bonas, 2003), as has been clearly shown foreffectors of
Yersinia
spp. (Cornelis, 2002a) and
Ralstoniasolanacearum
(Cunnac
et al
. 2004). Plant pathogens,such as
Pseudomonas
species,
Ralstonia
and
Xanthomo-nas
, secrete an array of effectors via their T3SS into theplant host cell (White
et al
., 2000). Recognition of theseeffectors by the plant may then lead to a hypersensitivereaction (HR), cell death and arrested infection (Büttnerand Bonas, 2003). Mounting evidence suggests that thisis also the case for some rhizobia, including NGR234(Marie
et al
., 2003). However, translocation of rhizobialT3SS effectors into plant host cells still awaits formalproof.
NGR234 secretes at least six proteins in a T3SS-dependent manner: NopA, NopB, NopC, NopL, NopP andNopX (Marie
et al
., 2003; Ausmees
et al
., 2004; Deakin
et al
., 2005; Saad
et al
., 2005). So far, only NopL andNopP have been shown to be effectors (Bartsev
et al
.,2003; 2004; Ausmees
et al
., 2004; this work), but it islikely that more will be identified. Complete absence ofsecreted proteins has various effects on the nodulation oflegumes, ranging from highly beneficial (
Crotalariajuncea
,
Pachyrhizus tuberosus
), to very weak (
Lotusjaponicus
,
Vigna unguiculata
), to deleterious (
Flemingiacongesta
,
Tephrosia vogelii
) (Marie
et al
., 2001). Yet it isnot known how the Nops affect nodule development. Weexpect, as is the case for phytopathogenic bacteria, thathost plants must respond to the cocktail of secreted effec-tors in different ways. Perhaps
C. juncea
and
P. tuberosus
recognize some components of the effector mix as aviru-lence factors and mount a strong defence reaction againstNGR234, whereas
F. congesta
and
T. vogelii
lack thecorresponding resistance genes, so allowing the effectorsto fulfil their function. The T3SS of NGR234 undoubtedlyhelps modulate the extremely broad host range ofNGR234 (that includes
>
112 genera of legumes, and thenon-legume
P. andersonii
– Pueppke and Broughton,1999), both extending (e.g.
F. congesta
) and restricting(e.g.
C. juncea
and
P. tuberosus
) the number of plants thatare effectively nodulated. Interestingly, functional T3SSsalso help modulate the host range of a number of animaland plant pathogens (e.g. Hueck, 1998; Büttner andBonas, 2003; Mattoo
et al
., 2004).In plant and animal pathogens, T3SS effectors subvert
the host’s metabolism and defence reactions by actingeither on the cytoskeleton or on intracellular signallingcascades (Cornelis, 2000). In NGR234, NopL is a targetfor the mitogen-activated protein (MAP) kinases of
Nicoti-ana tabacum
and
L. japonicus
(Bartsev
et al.
, 2003;2004). Phosphorylation of NopL
in vitro
is inhibited by aMAP kinase kinase inhibitor (Bartsev
et al.
, 2003). MAP
kinases are often involved in recognition processes ofpathogens and are known to activate plant defence genes(Jonak
et al.
, 2002). Furthermore, T3SS-dependent effec-tors of
Yersinia
spp. inhibit various signalling cascades ineukaryotic cells by interfering with MAP kinase pathways(Cornelis, 2002b; Orth, 2002; Yoon
et al.
, 2003). Receivedwisdom thus suggests that a spectrum of effectors mod-ulate host responses to pathogens, and naturally weasked whether the situation in a symbiotic bacterium wassimilar. Accordingly, we investigated whether other effec-tor proteins exist in NGR234 and if so whether they mightmodulate the activity of signalling cascades in roots of
C.juncea
,
F. congesta
,
P. tuberosus
,
T. vogelii
and
V. unguic-ulata
. Our results show that NopP, like NopL, is phospho-rylated by the plant and is a major determinant ofnodulation of the tropical legumes
F. congesta
and
T. vogelii
.
Results
NopP is phosphorylated by microsomes of legume roots
As NopL is phosphorylated by MAP kinases (Bartsevet al., 2003), it was reasonable to investigate whetherNopP was also phosphorylated by plant kinases. InPhaseoloid legumes, the root hairs that are most suscep-tible to bacterial invasion are those of intermediate age(Reli
et al
., 1994; Gehring
et al.
, 1997) that correspondto zone II in the
Trifolieae
and
Vicieae
(Heidstra
et al.
,1994; Gage 2004). In an extension of these findings, thesimplest way of enriching root preparations in susceptibleroot hairs is to grow seedlings on an agar surface. Follow-ing the protocols of Krause and Broughton (1992) andKrause
et al.
(1994), 2-day-old seedlings of
P. tuberosus
,
T. vogelii
and
V. unguiculata
were sprayed with suspen-sions of NGR234 or NGR
W
rhcN
(a NGR234 derivativewith a non-functional T3SS). One, 2 or 4 days later (allow-ing time for T3SS induction), the roots were separatedfrom the cotyledons, and plunged into liquid nitrogen.Then, the frozen roots were separated into total solubleproteins, the microsomal fraction and the plasma mem-brane fraction as described in
Experimental procedures
.When these fractions were phosphorylated
in vitro
usingthe
33
P procedure, no significant alterations of the phos-phorylation patterns were found as a result of treating theroots with either NGR234 or NGR
W
rhcN
(data not shown).In another attempt to check whether phosphorylationplays a symbiotic role, NGR234 and its derivatives weregrown in liquid cultures in the presence of apigenin andthe secreted proteins harvested from each culture super-natant. In this way, a highly concentrated solution ofsecreted proteins was obtained. Then, we mixedmicrosomes of roots from seedlings of
C. juncea, F. con-gesta, P. tuberosus, T. vogelii and V. unguiculata, with
c¢

1306 P. Skorpil et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1304–1317
secreted proteins of either NGR234 or NGRWrhcN(Fig. 1). As controls, microsomes were also mixed withsecreted proteins of non-induced cultures of NGR234(Fig. 1, lanes 3) and either microsomes or secreted pro-teins were phosphorylated alone (shown for F. congesta;Fig. 1, lanes 1 and 2). Using these techniques, markedchanges in phosphorylation patterns were observed andat least three proteins were phosphorylated in a T3SS-dependent manner (bands phosphorylated only in mix-tures containing secreted proteins of NGR234, not withthose of NGRWrhcN, as indicated by dots in Fig. 1).Secreted proteins alone were not phosphorylated, sug-gesting that the T3SS of NGR234 does not secretekinases, or if it does, they cannot phosphorylate any ofthe other secreted proteins. Similar experiments per-formed with the plasma membrane fraction as well as thetotal soluble protein fractions gave comparable results, butas the strongest bands were seen with microsomes, onlythese data are shown here. Intriguingly, although the fiveplants tested respond to the T3SS in radically differentways, all phosphorylate the same three proteins.
To identify the phosphorylated proteins, the 33P reactionmixtures were subjected to two-dimensional (2D) gel anal-ysis. Identical mixtures, but without ATP, were analysed in
parallel. The spots of interest were excised from the non-radioactive gels, digested with trypsin and analysed usingmatrix-assisted laser desorption/ionization-time of flight(MALDI-TOF) procedures (Fig. 2). Two of the three T3SS-dependent phosphorylated proteins were identified asNopL and NopP respectively. The third protein could beeither of plant or of bacterial origin, but was not investi-gated further. Phosphorylation of NopL was expected(Bartsev et al., 2003; 2004). Phosphorylation of NopP isshown here for the first time, and we have extended theseobservations to a number of other legumes.
The N-terminus of NopP is phosphorylated and is sufficient for secretion
We repeated the phosphorylation analyses usingmicrosomes and mixtures of proteins secreted by a nopLmutant (NGRWnopL) and a nopP mutant (NGRDnopP).As expected, the band that corresponds to NopL was notpresent when induced extracts from NGRWnopL wereused (Fig. 3, lane 4). Similarly, the band corresponding toNopP was also missing in the nopP mutant (Fig. 3, lane5), but surprisingly, a new, low-molecular-weight bandappeared (marked by dots in Fig. 3, lanes 5 and 6). Aus-
Fig. 1. Phosphorylation of T3SS proteins by microsomes of roots from five tropical legumes. Five micrograms of microsomes of roots of Crotalaria juncea, Flemingia congesta, Pachyrhizus tuberosus, Tephrosia vogelii and Vigna unguiculata were mixed in phosphorylation reactions with 1.5 mg of secreted proteins of a non-induced culture of NGR234 (lanes 3) or with secreted proteins of induced cultures of NGR234 (lanes 4) or NGRWrhcN (lanes 5). The phosphorylation reaction was started by adding [g-33P]-ATP. The samples were separated on 15% SDS-PAGE and radiolabelled proteins were visualized on autoradiograms. As a negative control, the phosphorylation reaction was performed with secreted proteins of an induced culture of NGR234 only (lane 1), or with microsomes only (lane 2), as exemplified for F. congesta. At least three proteins were phosphorylated in a T3SS-dependent manner, two of which were later identified as NopL and NopP (shown by dots). Note that in each lane 5 there is another phosphorylated protein of almost the same size as NopL. Mr, relative molecular mass in kDa.
1 2 3 4 5 3 4 5 3 4 5 3 4 5 3 4 5
NopL
NopP
?
Microsomes of: F. congesta T. vogelii V. unguiculata P. tuberosus C. juncea
100
75 50
37
20
25
15
10
Mr
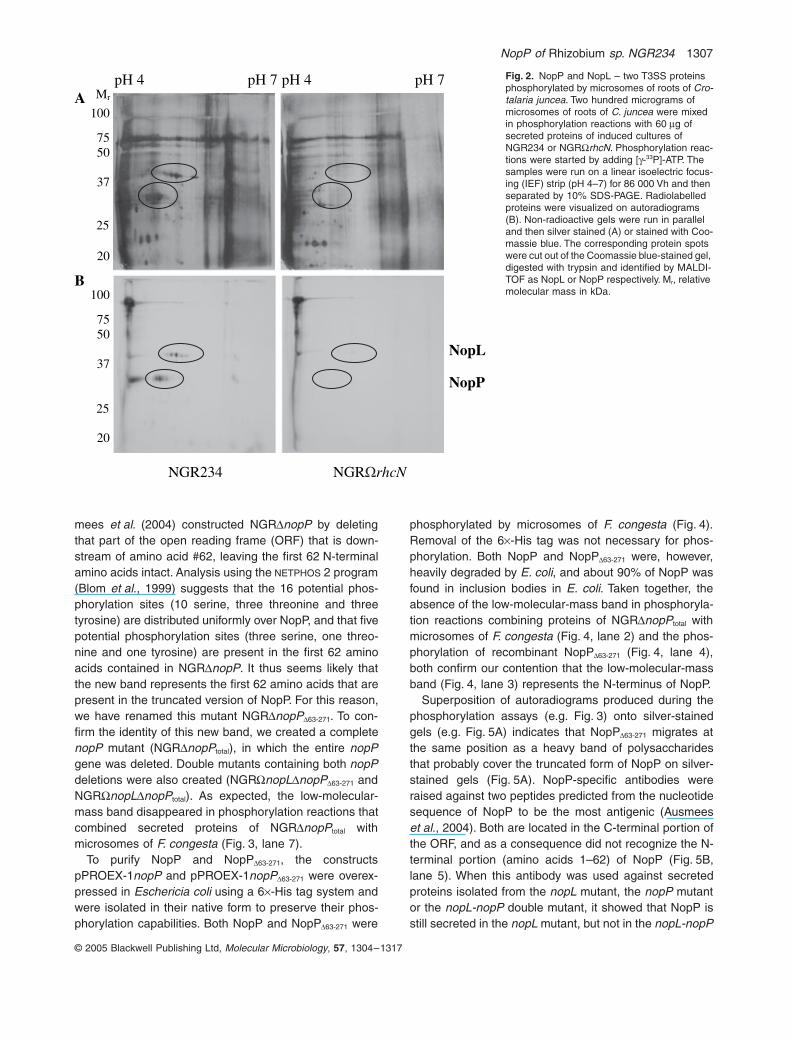
NopP of Rhizobium sp. NGR234 1307
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1304–1317
mees et al. (2004) constructed NGRDnopP by deletingthat part of the open reading frame (ORF) that is down-stream of amino acid #62, leaving the first 62 N-terminalamino acids intact. Analysis using the NETPHOS 2 program(Blom et al., 1999) suggests that the 16 potential phos-phorylation sites (10 serine, three threonine and threetyrosine) are distributed uniformly over NopP, and that fivepotential phosphorylation sites (three serine, one threo-nine and one tyrosine) are present in the first 62 aminoacids contained in NGRDnopP. It thus seems likely thatthe new band represents the first 62 amino acids that arepresent in the truncated version of NopP. For this reason,we have renamed this mutant NGRDnopPD63-271. To con-firm the identity of this new band, we created a completenopP mutant (NGRDnopPtotal), in which the entire nopPgene was deleted. Double mutants containing both nopPdeletions were also created (NGRWnopLDnopPD63-271 andNGRWnopLDnopPtotal). As expected, the low-molecular-mass band disappeared in phosphorylation reactions thatcombined secreted proteins of NGRDnopPtotal withmicrosomes of F. congesta (Fig. 3, lane 7).
To purify NopP and NopPD63-271, the constructspPROEX-1nopP and pPROEX-1nopPD63-271 were overex-pressed in Eschericia coli using a 6¥-His tag system andwere isolated in their native form to preserve their phos-phorylation capabilities. Both NopP and NopPD63-271 were
phosphorylated by microsomes of F. congesta (Fig. 4).Removal of the 6¥-His tag was not necessary for phos-phorylation. Both NopP and NopPD63-271 were, however,heavily degraded by E. coli, and about 90% of NopP wasfound in inclusion bodies in E. coli. Taken together, theabsence of the low-molecular-mass band in phosphoryla-tion reactions combining proteins of NGRDnopPtotal withmicrosomes of F. congesta (Fig. 4, lane 2) and the phos-phorylation of recombinant NopPD63-271 (Fig. 4, lane 4),both confirm our contention that the low-molecular-massband (Fig. 4, lane 3) represents the N-terminus of NopP.
Superposition of autoradiograms produced during thephosphorylation assays (e.g. Fig. 3) onto silver-stainedgels (e.g. Fig. 5A) indicates that NopPD63-271 migrates atthe same position as a heavy band of polysaccharidesthat probably cover the truncated form of NopP on silver-stained gels (Fig. 5A). NopP-specific antibodies wereraised against two peptides predicted from the nucleotidesequence of NopP to be the most antigenic (Ausmeeset al., 2004). Both are located in the C-terminal portion ofthe ORF, and as a consequence did not recognize the N-terminal portion (amino acids 1–62) of NopP (Fig. 5B,lane 5). When this antibody was used against secretedproteins isolated from the nopL mutant, the nopP mutantor the nopL-nopP double mutant, it showed that NopP isstill secreted in the nopL mutant, but not in the nopL-nopP
pH 4 pH 7 A
B
pH 4 pH 7
NGR234 NGR rhcN
NopL
NopP
100
75 50
37
20
25
Mr
100
75 50
37
20
25
Fig. 2. NopP and NopL – two T3SS proteins phosphorylated by microsomes of roots of Cro-talaria juncea. Two hundred micrograms of microsomes of roots of C. juncea were mixed in phosphorylation reactions with 60 mg of secreted proteins of induced cultures of NGR234 or NGRWrhcN. Phosphorylation reac-tions were started by adding [g-33P]-ATP. The samples were run on a linear isoelectric focus-ing (IEF) strip (pH 4–7) for 86 000 Vh and then separated by 10% SDS-PAGE. Radiolabelled proteins were visualized on autoradiograms (B). Non-radioactive gels were run in parallel and then silver stained (A) or stained with Coo-massie blue. The corresponding protein spots were cut out of the Coomassie blue-stained gel, digested with trypsin and identified by MALDI-TOF as NopL or NopP respectively. Mr, relative molecular mass in kDa.

1308 P. Skorpil et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1304–1317
double mutant (cf. Fig. 5B, lanes 4–8). This suggests thatmutation of nopL or nopP had no generalized effect onT3SS-dependent secretion, and confirms that they are notpart of the secretion machinery. It is also clear that thefirst 62 amino acids of NopP contain both the signal forT3SS-dependent secretion and sites for phosphorylationby plant kinases.
Inhibition of phosphorylation of NopP
As phosphorylation of NopL is inhibited by PD98059, aMAP kinase kinase inhibitor (Bartsev et al., 2003), wetested the effects of a number of kinase inhibitors on thephosphorylation of NopP (Fig. 6). Bis-indolylmaleimide I(a protein kinase C inhibitor), KN62 (a CaM kinase IIinhibitor) and PD98059 did not significantly alter thephosphorylation of NopP. Genistein, a general tyrosinekinase inhibitor, and K252a, a general serine/threoninekinase inhibitor, significantly reduced the phosphoryla-tion of NopP. A combination of both general kinaseinhibitors further reduced the phosphorylation of NopP(Fig. 6). Phosphorylation of NopL cannot be assessed inthis assay, because there is another phosphorylatedprotein of almost the same size as NopL (Fig. 1). Asexpected, the general kinase inhibitors affected phos-phorylation of all proteins. Unfortunately, these resultsdo not pinpoint a specific kinase family that phosphory-lates NopP.
1 2 3 4 5 6
NopLNopP
NopPD63-271
NopPD63-271
and its degradation
products
NopP
Secreted proteins of NGR234
and its derivatives
Overexpressed, 6x-His-Tag purified proteins of E. coli
Degradation products of NopP
100 75 50
20
37
25
15
10
MrFig. 4. Microsomes of Flemingia congesta phosphorylate recombinant NopP and NopPD63-271. Five micrograms of microsomes of roots of F. congesta were mixed in phosphory-lation reactions with 1.5 mg of secreted proteins of induced cultures of NGR234 (lane 1), NGRD-nopPtotal (lane 2) or NGRDnopPD63-271 (lane 3). Microsomes were also added to phosphoryla-tion reactions containing 5 mg of recombinant NopPD63-271 (lane 4) or recombinant NopPtotal (lane 5) purified from E. coli pPROEX-1nopPD63-271 or pPROEX-1nopP respectively. These recombinant proteins carry a 6¥-His tag. The corresponding fraction puri-fied from E. coli pPROEX-1 (empty vector) was used as a control (lane 6). Phosphorylation reactions were initiated by adding [g-33P]-ATP. The samples were separated by 15% SDS-PAGE and radiolabelled proteins were visual-ized on autoradiograms. Microsomes phospho-rylated recombinant NopP (lane 5), as well as its diverse degradation products (recombinant NopP is not stable in E. coli). Microsomes also phosphorylated recombinant NopPD63-271 (lane 4). Recombinant NopPD63-271 was also partially degraded by E. coli. Potential phosphorylation sites are uniformly distributed all over NopP and are functional targets for kinases, even in trun-cated versions of NopP. Mr, relative molecular mass in kDa.
Fig. 3. Phosphorylation profiles of various NGR234 derivatives. Five micrograms of microsomes of roots of Flemingia congesta were mixed in phosphorylation reactions with 1.5 mg of secreted proteins of non-induced cultures of NGR234 (lane 1), or with secreted proteins of induced cultures of NGR234 (lane 2), NGRWrhcN (lane 3), NGR-WnopL (lane 4), NGRDnopPD63-271 (lane 5), NGRWnopLDnopPD63-271 (lane 6), NGRDnopPtotal (lane 7) or NGRWnopLDnopPtotal (lane 8). Phosphorylation reactions were initiated by adding [g-33P]-ATP. After-wards, the samples were separated by 15% SDS-PAGE and radiola-belled proteins were visualized on autoradiograms. Two new, low-molecular-weight phosphorylated bands appeared only in lane 5 (NGRDnopPD63-271) and lane 6 (NGRWnopLDnopPD63-271). Mr, relative molecular mass in kDA.
1 2 3 4 5 6 7 8
NopL
NopP
NopP 63-271
100
75 50
20
37
25
15
10
Mr

NopP of Rhizobium sp. NGR234 1309
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1304–1317
NopP is the main T3SS determinant of nodulation of the tropical legumes F. congesta and T. vogelii
Given that NopP is a secreted effector protein capableof being phosphorylated by plant kinases, we askedwhether its presence or absence has any effect on nod-ulation of legumes known to respond to T3SS mutants.Proteins secreted via the T3SS of NGR234 (includingNopL) are crucial to nodulation of F. congesta (Marieet al., 2003). The data in Table 1 show that NopP iseven more important than NopL in nodulation of thisplant. Mutation of nopP led to a much stronger reduc-tion in nodule numbers than were obtained with thenopL mutant, suggesting that NopP is the major T3SSdeterminant for nodulation of F. congesta. The reductionin nodule numbers was even greater with the nopL-nopP double mutant, although in this case, morenodules were still formed than with NGRWrhcN. Nosignificant differences in the nodulation of any of thelegumes used in this study were seen following inocula-tion with NGRDnopPD63-271 or NGRDnopPtotal, norbetween inoculations with NGRWnopLDnopPD63-271 orNGRWnopLDnopPtotal (Table 1).
Absence of NopP led to a 50% reduction in the num-ber of nodules formed on T. vogelii (Table 1). Interest-ingly, mutation of both nopL and nopP led to a further50% reduction in nodule number, with the result that only33% as many nodules were formed in comparison withplants inoculated with NGRWrhcN. This observation isimportant, for it suggests that while both NopL and NopPare needed for optimal nodulation of T. vogelii, their pres-ence masks the impact of a negative effector (seebelow).
In contrast to the above, a functional T3SS inhibitsnodulation of C. juncea and P. tuberosus (Viprey et al.,1998; Marie et al., 2003). On both plants, none of the
mutants (nopL, nopP or nopL-nopP) permitted effectivenodulation, a phenotype that was confined to inoculationwith the T3SS null mutant NGRWrhcN. Although NGR-WrhcN led to the formation of a great number of func-tional nodules on C. juncea, inoculation with NGR234 orany of the other mutants used in this study led to the
Fig. 6. Effects of diverse inhibitors on the in vitro phosphorylation of NopP by kinases of plant roots. Microsomes (0.5 mg) extracted from the roots of Flemingia congesta were mixed in phosphorylation reac-tions with 1.5 mg of secreted proteins of an induced culture of NGR234. The phosphorylation reaction was started by adding [g-33P]-ATP. The samples were separated by 15% SDS-PAGE and radiola-belled proteins were visualized on autoradiograms. Control reactions contained 16% DMSO (lane 1). Genistein, a general tyrosine kinase inhibitor (lane 2), and K252a, a general serine/threonine kinase inhib-itor (lane 3), inhibit significantly the phosphorylation of NopP by plant root kinases. If both general kinase inhibitors were used together, phosphorylation was further reduced (lane 4). Bis-indolylmaleimide I (a protein C kinase inhibitor, lane 5), KN62 (a CaM kinase II inhibitor, lane 6) and PD98059 (a MAP kinase kinase inhibitor, lane 7) did not significantly alter the phosphorylation of NopP. Mr, relative molecular mass in kDa.
100 75 50
20
Mr
15
10
37
25
1 2 3 4 5 6 7
NopP
100
A B
75 50
20
37
25
15
10
Mr
1 2 3 4 5 6 7 8 1 2 3 4 5 6 7 8
NopX
NopL
NopP
NopA
Silver-stained gel Western blots
Fig. 5. Profiles of secreted proteins of various NGR234 derivatives.A. Silver-stained gel of secreted proteins of a non-induced culture of NGR234 (lane 1), secreted proteins of induced cultures of NGR234 (lane 2), NGRWrhcN (lane 3), NGR-WnopL (lane 4), NGRDnopPD63-271 (lane 5), NGRWnopLDnopPD63-271 (lane 6), NGRDnopPtotal (lane 7) and NGRWnopLDnopPtotal (lane 8).B. Corresponding Western blots. Note that the antibody against NopP was raised against pep-tides from the C-terminal end of the protein. It did not recognize NopPD63-271, in which only the N-terminus of NopP remains. The samples were separated by 15% SDS-PAGE. Mr, relative molecular mass in kDa.

1310 P. Skorpil et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1304–1317
formation of many pseudonodules (Fig. 7). In contrast, P.tuberosus makes almost no pseudonodules. Again, thesedata suggest that a (or more than one) negative effector
must exist, and only in its absence (i.e. in the T3SS nullmutant) can functional nodules form on C. juncea andP. tuberosus.
Fig. 7. Nodulation of Tephrosia vogelii and Crotalaria juncea by NGR234 and its derivatives. Only the aerial portions of the plants are shown. Plants were harvested 7 weeks post inoculation. Size of plants is representative of the number of nitrogen-fixing nodules formed. Comparison between plants inoculated with NGRWnopLDnopPtotal or NGRWrhcN highlights the existence of at least one negative T3SS effector of NGR234.
NGR rhcN NGR234 NGR rhcN NGR234
NGR nopL nopPtotal NGR nopL nopPtotal
Tephrosia vogelii
Crotalaria juncea
Table 1. Nodulation of various tropical legumes by NGR234 and its derivatives.
Tephrosia vogelii Flemingia congesta Pachyrhizus tuberosus
Control 0 0 0NGR234 27 (± 3) 49 (± 8) 0.4 (± 0.3)NGRWrhcN 17 (± 2.5) 3 (± 1) 41 (± 8)NGRWnopL 25 (± 4) 33 (± 7) 0.5 (± 0.4)NGRDnopPD63-271 13 (± 3) 22 (± 4) 0.4 (± 0.4)NGRDnopPtotal 13 (± 2) 18 (± 4) 0.3 (± 0.4)NGRWnopLDnopPD63-271 8 (± 3) 7 (± 2) 0.2 (± 0.3)NGRWnopLDnopPtotal 6 (± 2) 5 (± 2) 0.4 (± 0.4)
Plants were harvested 7 weeks post inoculation and the number of nitrogen-fixing nodules counted. Standard errors of the means are indicated.Nodulation tests were performed at least twice, using about 15 plants in each treatment.

NopP of Rhizobium sp. NGR234 1311
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1304–1317
Discussion
Although Nod factors allow rhizobia to enter the tips ofroot hairs of Phaseoloid legumes, other bacterial sub-stances are required for infection thread and nodule devel-opment (Broughton et al., 2000; Perret et al., 2000; Mathiset al., 2005). Included among these are proteins secretedvia the T3SS. To test whether these proteins affect infec-tion thread development, we inoculated P. tuberosus, T.vogelii and V. unguiculata, with NGR234 as well as NGR-WrhcN marked with GUS. Histological observation ofGUS-stained roots in a quantitative assay failed to pin-point differences in infection thread growth when either ofthe two bacterial strains was used to infect any of theseplants (M. Crèvecœur, M. Saad, P. Skorpil, unpubl. data).Thus we began to investigate whether the effector pro-teins bring about differential responses within legume rootcells. NopL is the best studied of these effectors and is adeterminant for nodulation by NGR234 of F. congesta(Marie et al., 2003). Possibly, NopL favours nodulation byinterfering with MAP kinase signalling pathways and inthis way, downregulates the host’s defence reactions(Bartsev et al., 2004). Obviously, this poses the questionof whether other effectors behave in a similar way. Toanswer this, we designed procedures that allow screeningboth bacteria and plants for protein–protein interactionsinvolved in phosphorylation. We modified an in vitro sys-tem that had been used to show that cytoplasmic proteinsof leaves of L. japonicus and tobacco could phosphorylateNopL (Bartsev et al., 2003). Initially plasma membranesof roots were targeted as they are first to be exposed toexternal signals. However, isolation of plasma membranesis time-consuming, and preliminary experiments showedthat the total soluble fraction, the microsome fraction andthe plasma membrane fraction all responded to secretedproteins. Sensitivity to phosphoryation seemed highest inthe microsomal fraction, which was used in subsequentexperiments. Surprisingly, the same three bands werephosphorylated in microsomal extracts of plants thatrespond in positive and negative ways to a functionalT3SS (Fig. 1). One implication of the observation that allthe legumes tested phosphorylate the same three pro-teins could be that there are different downstream targetsites for NopL and NopP depending on the legume. Alter-natively NopL and NopP could perform the same basicfunction in each legume into which they are translocated,but on legumes such as C. juncea or P. tuberosus thereaction provoked by other (negative) effectors is toostrong to be overcome by the (positive) actions of NopLand NopP.
Proteomic methods successfully identified NopP as thesecond rhizobial effector protein to be a target of kinases(Fig. 2). Phosphorylation of NopP by kinases of legumeroots was partially blocked by general inhibitors of tyrosine
and serine/threonine kinases, but not by a MAP kinasekinase inhibitor previously shown to inhibit phosphoryla-tion of NopL (Fig. 6). Our working hypothesis concerningNopL, based upon the characterization of its in vitro phos-phorylation and the fact that transgenic plants expressingnopL show reduced defence responses when challengedby viral pathogens, is that NopL may interfere with signal-ling cascades to thwart activation of inducible plantdefence genes (Bartsev et al., 2004). It is too preliminaryto say whether NopP may act in a similar way. Could NopLand NopP compete with the native targets for phosphory-lation by eukaryotic kinases? Generally this is not thestrategy used when effector proteins block signalling path-ways. Rather, the effectors act as phosphatases or pro-teases to downregulate host responses (Chang et al.,2004). The only bacterial T3SS effector protein known tobe phosphorylated by eukaryotic cells is Tir of entero-pathogenic E. coli (EPEC). This protein is translocated tothe plasma membrane of host cells where it acts as areceptor for EPEC binding (Kenny et al., 1997). Tir isphosphorylated by different eukaryotic kinases at serine,threonine and tyrosine residues (Vallance and Finlay,2000). A secondary function of Tir is to trigger signallingcascades to alter actin polymerization resulting in cytosk-eletal re-arrangements of the eukaryotic cell (Campelloneand Leong, 2003). Phosphorylation of a specific tyrosineresidue of Tir is critical for this to occur. Alterations of theeukaryotic cytoskeleton are often consequences of effec-tor proteins secreted by bacterial T3SSs and can beimportant for the internalization of certain pathogens,such as Salmonella typhimurium (Fu and Galán, 1999).An internalization of rhizobia into plant cells occurs duringthe formation of nodules, when they are released frominfection threads into root cortical cells. Indeed, re-orga-nization of microtubular cytoskeletal proteins occurs in theroot inner cortical cells where the nodule eventually forms(Timmers et al., 1999). It is thus possible that the phos-phorylation of NopL and NopP may be necessary foralterations to cytoskeletal proteins. This could also explainwhy all the legumes tested phosphorylate NopL and NopPas it is a common requirement for NGR234 entry. How-ever, if one or more negative effector proteins are recog-nized by a particular legume (e.g. C. juncea or P.tuberosus), it seems likely that the initiation of a putativedefence response by the legume will block entry of therhizobia.
Although NopP is a known secreted protein (Ausmeeset al., 2004), the first 62 N-terminal amino acids were leftuntouched during construction of the original nopP mutant(NGRDnopPD63-271). As a consequence, a truncated NopPwas still secreted and phosphorylated by plant kinases(Fig. 4). Obviously this means that the signal(s) for secre-tion are contained in the 62 N-terminal amino acids, whichmust also harbour phosphorylation sites. Given this infor-

1312 P. Skorpil et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1304–1317
mation, it is possible that the truncated version of NopPmight also affect the nodulation processes. Therefore, toassess the exact function of NopP, we created a true nullmutant of nopP. When NGRDnopPtotal was assayed forphosphorylation, bands corresponding to neither the full-length NopP nor the truncated N-terminal fragment weredetected. When the nodulation ability of NGRDnopPtotal
and NGRDnopPD63-271 was compared, no significant differ-ences were observed between the two nopP mutants,suggesting that the truncated version of NopP has littleeffect on nodulation. It is possible that although the NopPfragment can be secreted (and phosphorylated) it may notbe translocated into plant cells in vivo. The exact role ofthe NopP fragment remains to be investigated, especiallyas in earlier studies (Ausmees et al., 2004) theNGRDnopPD63-271 mutant was shown to slightly improvenodulation of F. congesta, relative to NGR234. We wereunable to repeat these earlier nodulation tests. Never-theless, it is possible that under certain conditions, thesecreted NopP fragment may be recognized as a patho-gen-associated molecular pattern (PAMP) that provokesthe plant to react against the bacteria.
Comparison of the nodulation phenotypes of the nopLand nopP mutants on different plants provided significantinsights into effector biology. Although inoculation of T.vogelii with NGRWnopL consistently led to a slight reduc-tion in nodule numbers compared with those obtained withNGR234, the symbiotic importance of NopL was onlyunmasked by using double mutants. Secretion of NopP inNGRWnopL (or NopL in NGRDnopPtotal) still allowednodulation and suggests that NopL and NopP are at leastpartially redundant. Differences in nodule numbersobserved after inoculating the plants with the nopP mutantand nopL-nopP double mutant suggest that NopL is ben-eficial to nodulation. However, on P. tuberosus and C.juncea, only NGRWrhcN is able to induce the formation ofnitrogen-fixing nodules, suggesting that other effectorsinhibit nodulation.
Inoculation of F. congesta and T. vogelii with the nopLmutant reduced nodulation. In turn, plants exposed to thenopP mutant had even fewer nodules (Table 1). Thisadditive phenotype suggests that NopL and NopP acttogether. These data also suggest that NopP is the mainT3SS determinant for nodulation of F. congesta and T.vogelii. Inoculation of F. congesta with the nopL-nopPdouble mutant led to the formation of more nodules thaninoculation with NGRWrhcN. This leaves open the possi-bility that another, as yet unidentified effector plays a pos-itive role in the nodulation of NGR234 of F. congesta. Moreinterestingly, the number of nodules observed on T. vogeliiplants infected with the double nopL-nopP mutant wasonly one-third of that obtained with NGRWrhcN, the strainthat is completely blocked in protein secretion. This isprobably the strongest evidence obtained so far that
another, as yet unidentified effector inhibits nodulation.Perhaps this (or these) effector(s) would be recognized byplants like C. juncea and P. tuberosus as avirulence fac-tors, and induce strong defence reactions (Fig. 7). Obvi-ously, neither NopL nor NopP is able to suppress thisadverse reaction.
Potential candidates for additional effectors are proteinscorresponding to ORFs y4fR, y4lO and y4zC (Marie et al.,2004). y4lO and y4zC are homologous to the Yersinaouter proteins YopJ and YopT respectively, whereas y4fRis homologous to YopM (Marie et al., 2004). YopJ andYopT are two cysteine proteases that interfere witheukaryotic cell signalling pathways (Orth, 2002; Innes,2003). YopJ does this by inhibiting MAP kinase signallingpathways (Yoon et al., 2003). The Yersinia spp. T3SSeffector YopM is targeted to cell nuclei, where it probablyinterferes with the cell cycle, cell growth and organizationof the cytoskeleton (Sauvonnet et al., 2002; Benabdillahet al., 2004). Another possible effector protein is the third,low-molecular-mass protein that is phosphorylated in aT3SS-dependent manner by root kinases (Fig. 1). Thisprotein is too small to be a product of the ORFs y4lO,y4zC or y4fR of NGR234, and although we were able tooptimize the 2D PAGE system to obtain the best resolutionof NopL and NopP (Fig. 2), we have not succeeded withthis small spot.
The well-characterized effector proteins secreted by theT3SSs of pathogenic bacteria mount a co-ordinatedattack on the signalling pathways of eukaryotic cells. Inthis work we show that two Rhizobium-specific effectorproteins (NopL and NopP) also act together, possibly asa consequence of their phosphorylation, in leguminousplants to improve nodulation efficiency. Exactly which sig-nalling events are altered (and the resulting effects) isdifficult to determine in legumes such as F. congesta andT. vogelli that have not been subject to molecular charac-terization. Our future aims are to characterize the rhizobialhomologues of the Yops, to see whether they have similarroles in legumes and to study whether (and then how) therhizobial-specific effectors interact with the ‘classical’effectors to either facilitate or reduce nodulation. Onestrategy that is being used is to express the effector genesin a genetically tractable legume to see whether theseputative rhizobial effectors function in the same way astheir pathogenic counterparts.
Experimental procedures
Bacterial growth conditions
Bacterial strains and plasmids used are listed in Table 2.Rhizobium strains were grown at 27∞C on tryptone yeast (TY)medium (Beringer, 1974), or on rhizobial minimal medium [ing l-1: K2HPO4, 2.05; KH2PO4, 1.45; NaCl, 0.15; NH4NO3, 0.5;MgSO4·7 H2O, 0.5; CaCl2·2 H2O, 0.025; C4H4O4Na2·6 H2O
NopP of Rhizobium sp. NGR234 1313
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1304–1317
(sodium succinate), 3.24; FeC6H6O7·2 H2O (ferric(III) citrate),3.35 mg l-1; micronutrient solution (Flow Laboratories, Irvine,Scotland, UK), 1000¥ diluted; Gamborgs’s vitamin solution(Sigma Chemical, St Louis, MO, USA), 1000¥ diluted], mod-ified after Broughton et al. (1986) and Marie et al. (2004). E.coli strains were grown in Luria–Bertani medium at 37∞C(Sambrook et al., 1989). The media were supplemented withantibiotics as described in Marie et al. (2004), except forgentamicin, which was used at 10 mg ml-1 for E. coli and5 mg ml-1 for Rhizobium. When indicated, Rhizobium cultureswere induced with apigenin at a final concentration of 10-6 M.
Construction of NGR234DnopPtotal, NGRWnopLDnopPD63-271 and NGRWnopLDnopPtotal
Standard molecular techniques were performed as describedin Sambrook et al. (1989). pJQyPSalI that carries the nopPlocus cloned into the SalI site of pJQ200SK (Quandt andHynes, 1993) was the source of nopP. The 3 kb SalI fragmentof pJQyPSalI was cloned into the SalI site of a modifiedpBlueScript II KS+ vector (Stratagene, La Jolla, CA, USA)lacking its EcoRV, HindIII and ClaI sites, called pKSyPSalI.Two fragments of pKSyPSalI were cloned separately into
pBlueScript II KS+ – a 190 bp EcoRI fragment upstream anda 1.75 kb Xho1 fragment downstream of nopP (giving thepKSnopP deletion). Then, a spectinomycin-resistant (Spr)interposon (Fellay et al., 1987) was cloned into the HindIIIsite of the pKSnopP deletion which thus replaced positions517 954–519 108 of pNGR234a (Freiberg et al., 1997) con-taining the nopP gene and its putative promoter. The 4 kbPstI–KpnI fragment containing the complete construct wastransferred to pJQ200SK, thus creating pJQDnopPtotal. Thisconstruct was mobilized into NGR234 by tri-parental matingsusing the helper plasmid pRK2013 (Figurski and Helinski,1979). Rhizobial DNA was prepared as described in Chenand Kuo (1993). Putative NGR234DnopPtotal mutants wereconfirmed by Southern blotting using standard methods(Sambrook et al., 1989). The double mutantsNGRWnopLDnopPD63-271 and NGRWnopLDnopPtotal wereobtained by the introduction of a mutated nopL gene (carriedon pJQnopLWKm) into NGRDnopPD63-271 and NGRDnopPtotal
respectively.
Purification and analysis of secreted proteins
Secreted proteins were purified and analysed as described
Table 2. Bacterial strains and plasmids used in this study.
Strain or plasmid Relevant characteristics Source
Escherichia coliE. coli strain DH5a supE44, DlacU169(F80 lacZDM15), hsdR17, recA1, gyrA96, thi-1, relA1 Invitrogen, Carlsbad, CA, USA
Rhizobium strainsNGR234 Rifr derivative of the broad host range bacterium isolated from nodules
of Lablab purpureusStanley et al. (1988)
NGRWrhcN NGR234 derivative containing an W cassette inserted into the EcoRI siteof rhcN, Rifr, Spr
Viprey et al. (1998)
NGRWnopL NGR234 derivative containing an W cassette inserted into the EcoRV siteof nopL, Rifr, Kmr
Marie et al (2003)
NGRDnopPD63-271 NGR234 derivative in which the C-terminus of nopP downstream from the first XhoI site was replaced by an W cassette, Rifr, Spr
Ausmees et al. (2004)
NGRWnopLDnopPD63-271 NGRDnopPD63-271 derivative containing an W cassette inserted into theEcoRV site of nopL, Rifr, Kmr, Spr
This study
NGRDnopPtotal NGR234 derivative in which the entire nopP was replaced by an Wcassette, Rifr, Spr
This study
NGRWnopLDnopPtotal NGRDnopPtotal derivative containing an W cassette inserted into the EcoRVsite of nopL, Rifr, Kmr, Spr
This study
PlasmidspBluescript II KS+ High-copy-number ColE1-based phagemid, Ampr Stratagene, La Jolla, CA, USApJQ200SK Suicide vector used for directed mutagenesis in Rhizobium, Ger Quandt and Hynes (1993)pRK2013 Helper plasmid necessary to transfer pJQ200SK into Rhizobium, Kmr Figurski and Helinski (1979)pJQnopLWKm pJQ200SK derivative carrying a construct in which an W cassette was
inserted into the EcoRV site of nopL, Ger, KmrMarie et al. (2003)
pJQyPSalI pJQ200SK derivative carrying a 3 kb fragment containing nopP, Ger This studypKSyPSalI pBluescript II MS+ derivative without EcoRV, HindIII and ClaI sites carrying
a 3 kb fragment containing nopP, AmprThis study
pKSDnopPtotal pBluescript II KS+ derivative carrying a construct in which nopP wasreplaced by an W cassette, Ampr, Spr
This study
pJQDnopPtotal pJQ200SK derivative carrying a construct in which nopP was replaced byan W cassette, Ger, Spr
This study
pBDG88 pBluescript II KS+ derivative carrying nopP in its SmaI site, Ampr This studypPROEX-1 E. coli protein expression system that adds an in-frame 6¥-His tag to the
overexpressed protein, AmprInvitrogen, Carlsbad, CA, USA
pPROEX-1nopP pPROEX-1 derivative carrying nopP, Ampr This studypPROEX-1nopPD63-271 pPROEX-1nopP derivative carrying the first 62 N-terminal amino acids of
NopP, AmprThis study
Amp, ampicillin; Ge, gentamicin; Km, kanamycin; Sp, spectinomycin; Rif, Rifampicin; superscript r, resistant.

1314 P. Skorpil et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1304–1317
in Marie et al. (2003), except that the induced bacterial cul-ture was grown to a final optical density at 600 nm of at least1.5. Secreted proteins of a 400 ml culture were condensedinto a final volume of 200 ml, which resulted in a typicalprotein concentration of 0.5 mg ml-1. Proteins were quantifiedusing a Bradford-based reagent (Bio-Rad, Hercules, CA,USA) or using a modified Lowry method (Peterson, 1977).Proteins were separated by SDS-PAGE (Laemmli, 1970) andeither silver stained (Ausubel et al., 1991) or stained by Coo-massie blue (Candiano et al., 2004). The IPGphor system(Amersham Biosciences, Little Chalfont, Buckinghamshire,UK) was used for the first dimension of 2D gels, accordingto the manufacturer’s protocols.
Plant assays
Suppliers of seeds are the same as those listed in Pueppkeand Broughton (1999). Surface sterilization of plant seeds: V.unguiculata: 70% (v/v) ethanol, 10 min and 5% (v/v) H2O2,10 min. P. tuberosus: concentrated sulphuric acid, 10 minand a bleach-mix [20 times diluted household bleach, 2% (v/v) Tween 20 and 0.1% (w/v) SDS], 20 min. T. vogelii: concen-trated sulphuric acid, 5 min, 0.1% (v/v) Tween 20 duringsonication, 5 min and 5% (v/v) H2O2, 10 min. C. juncea:bleach-mix, 30 min and 70% (v/v) ethanol, 10 min. F. con-gesta: concentrated sulphuric acid, 10 min, 0.1% (v/v) Tween20 during sonication, 10 min, 70% (v/v) ethanol, 10 min and5% (v/v) H2O2, 10 min. For germination, seeds were placedon B and D medium + agar [1 mM CaCl2; 0.5 mM KH2PO4;0.5 mM K2HPO4; 12 mM FeC6H6O7·2 H2O (ferric(III) citrate);micronutrient solution (Flow Laboratories, Irvine, Scotland,UK), 2000¥ diluted; pH 7.5; +1.5% (w/v) agar], modified afterBroughton and Dilworth (1971). Nodulation tests were per-formed in Magenta jars (Lewin et al., 1990). One plant wasgrown per Magenta jar. The vermiculite (Vermex AG, Rein-ach, Switzerland) was covered with sterilized white plasticbeads (Blein Meyer SA, Bulle, Switzerland). The lowerassembly of MagentaTM jars was filled with B and D nutrientsolution, except for P. tuberosus, where B and D was supple-mented with 1 mM KNO3. Plants were grown at 28∞C duringa 15 h day including a 1.5 h dawn and a 1.5 h dusk phase.Maximal light intensity was 100 mE m-2 s-1. Night time tem-perature was 18∞C. Plants were inoculated with 107 bacteriaand harvested 7 weeks post inoculation.
Isolation of microsomes, plasma membranes and total soluble proteins of roots
Plant seedlings were germinated in the dark and their rootswere harvested after snap freezing in liquid nitrogen. Rootswere homogenized for 1 min in 10¥ (w/v) membrane extrac-tion buffer [0.5 M sorbitol; 10 mM EGTA; 2.5 mM potassiummetabisulphite; 4 mM salicylhydroxamic acid; 5% (w/v) cross-linked polyvinylpyrrolidone; 0.5% (w/v) serum bovine albu-min; 50 mM MOPS; pH 7.6]. The homogenate was passedthrough cheese cloth and centrifuged for 20 min at 13 000 gand 4∞C. The supernatant was centrifuged for 50 min at45 000 g and 4∞C. The pellet containing the microsomes wasresuspended in 0.5 M sorbitol dissolved in 3 mM Tris-maleatebuffer (pH 7.3) to give protein concentrations of ~5 mg ml-1.
For purification of plasma membranes, microsomes wereloaded onto a polymer two-phase system (Albertsson, 1970;1982; Yoshida et al., 1983) buffered with 0.5 M sorbitol dis-solved in 10 mM potassium phosphate (pH 7.6). Optimalpolymer concentrations were 6% for C. juncea, 5.9% for V.unguiculata, F. congesta and P. tuberosus and 5.8% for T.vogelii. The two-phase system was thoroughly mixed byinversion and centrifuged for 90 min at 3000 g and 4∞C. Theplasma membrane-containing phase was diluted with 0.5 Msorbitol dissolved in 15 mM Tris-maleate buffer (pH 7.3) andcentrifuged for 50 min at 45 000 g and 4∞C. Plasma mem-branes were resuspended in 0.5 M sorbitol dissolved in3 mM Tris-maleate buffer (pH 7.3) to give protein concentra-tions of ~1 mg ml-1. For isolation of the total soluble proteinfraction, roots were ground for 1 min at 4∞C in 10¥ (w/v)extraction buffer [10 mM EGTA; 10 mM potassium metabisul-phite; 8 mM salicylhydroxamic acid; 30 mM NaCl; 10 mMDDT; 1 mM AEBSF; 0.1% (w/v) cross-linked polyvinylpyrroli-done; 20 mM MES-KOH; pH 6.5]. The homogenate wascentrifuged twice, each for 15 min at 12 000 g and 4∞C anddesalted using 30 kDa Microcon columns (Millipore, Bedford,MA, USA). The remaining liquid was diluted with 4 mMAEBSF dissolved in 3 mM Tris-maleate buffer (pH 7.3) to giveprotein concentrations of ~5 mg ml-1.
Phosphorylation assays
Phosphorylation reactions were carried out in phosphoryla-tion buffer [20 mM MES; 10% (v/v) glycerol; 5 mM MgCl2;1 mM CaCl2; pH 6.5 adjusted with NaOH; 1 mM MnSO4;0.4% (w/v) detergent C12E8; 5 mM b-mercaptoethanol].Reactions were optimal at 32∞C and for 2 min. They werestarted by adding 10 mCi [g-33P]-ATP and stopped by addingsample loading buffer for SDS-PAGE. X-ray-sensitive filmwas exposed at -70∞C to the dried gels. The kinase inhibitorsPD98059, KN62, K252a and Bisindolylmaleimide I (EMD Bio-sciences, San Diego, CA, USA) were dissolved in DMSO.The control reaction contained 16% (v/v) DMSO. Concentra-tions of inhibitors used in the assays were (values in bracketsif the inhibitors were used in combination): genistein (AcrosOrganics, Geel, Belgium), 3 (1.5) mM; K252a, 35 (17.5) mM;bis-indolylmaleimide I, 100 mM; KN62, 230 mM; PD98059,760 mM.
Overexpression of NopP and NopPD63-271
Vent polymerase (New England Biolabs, Beverly, MA, USA)and primers matched to the 5¢- and 3¢-ends of nopP (5¢-CGGAT ATC ATG TAC GGT CGA ATT GAT AGC-3¢ and 5¢-CATGAA GTC ATC TTC-3¢) were used in polymerase chain reac-tions (PCR). The PCR products were cloned into the SmaIsite of pBluescript KS+ creating pBDG88 and sequenced toverify the accuracy of the PCR. A DNA fragment containingnopP was excised from pBDG88 using an EcoRV site engi-neered into the 5¢ primer (underlined) and an XbaI site fromthe MCS of KS+ at the 3¢-end of nopP. This fragment wascloned into the EheI and XbaI sites of pPROEX-1 (Invitrogen,Carlsbad, CA, USA), giving rise to pPROEX-1nopP, whichadded an in-frame 6¥-His tag to the N-terminus of NopP. Tooverexpress the N-terminus of NopP, which is still present in

NopP of Rhizobium sp. NGR234 1315
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1304–1317
NGRDnopPD63-271, pPROEX-1nopP was digested with XhoIand subsequently re-ligated. In this way, pPROEX-1nopPD63-271 carrying an N-terminal in-frame 6¥-His tag wasobtained. E. coli cultures were induced with 0.6 mM isopro-pyl-b-D-thiogalactopyranoside for 2 h. The cell pellet waslysed with BugBuster HT (EMD Biosciences, San Diego, CA,USA) and 6¥-His-tagged proteins purified using TALON MetalAffinity Resin (BD Biosciences Clontech, Palo Alto, CA, USA)according to the manufacturer’s instructions. Proteins werepurified in native conformation.
Acknowledgements
We are grateful to Yin-Yin Aung for help with this work. Wethank Dora Gerber for general support and Christian Stae-helin, Olivier Schumpp, Antoine Le Quéré and ChristopheDunand for many interesting discussions. Thanks, too, to NickBrewin and Claude Penel for critical reading of the article.Financial assistance was provided by the Département del’Instruction Publique du Canton de Genève, by the Universityof Geneva and by the Fonds National de la RechercheScientifique (Projects 31-63893.00 and 3100AO-104097/1).
References
Albertsson, P.-A. (1970) Partition of cell particles and mac-romolecules in polymer two-phase systems. Adv ProteinChem 24: 309–341.
Albertsson, P.-A., Andersson, B., Larsson, C., and Akerlund,H.-E. (1982) Phase partition – a method for purification andanalysis of cell organelles and membrane vesicles. Meth-ods Biochem Anal 28: 115–150.
Ausmees, N., Kobayashi, H., Deakin, W.J., Marie, C.,Krishnan, H.B., Broughton, W.J., and Perret, X. (2004)Characterization of NopP, a type III secreted effector ofRhizobium sp. strain NGR234. J Bacteriol 186: 4774–4780.
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D.,Seidman, J.G., Smith, J.A., and Struhl, K. (1991) CurrentProtocols in Molecular Biology. New York, USA: John Wiley& Sons.
Bartsev, A.V., Boukli, N.M., Deakin, W.J., Staehelin, C., andBroughton, W.J. (2003) Purification and phosphorylation ofthe effector protein NopL from Rhizobium sp. NGR234.FEBS Lett 554: 271–274.
Bartsev, A.V., Deakin, W.J., Boukli, N.M., McAlvin, C.B.,Stacey, G., Malnoë, P., et al. (2004) NopL, an effectorprotein of Rhizobium sp. NGR234, thwarts activation ofplant defense reactions. Plant Physiol 134: 871–879.
Benabdillah, R., Mota, L.J., Lützelschwab, S., Demoinet, E.,and Cornelis, G.R. (2004) Identification of a nuclear target-ing signal in YopM from Yersinia spp. Microb Pathog 36:247–261.
Beringer, J.E. (1974) R factor transfer in Rhizobium legumi-nosarum. J Gen Microbiol 84: 188–198.
Blom, N., Gammeltoft, S., and Brunak, S. (1999) Sequenceand structure-based prediction of eukaryotic proteinphosphorylation sites. J Mol Biol 294: 1351–1362.
Broughton, W.J., and Dilworth, M.J. (1971) Control of leghae-moglobin synthesis in snake beans. Biochem J 125: 1075–1080.
Broughton, W.J., Wong, C.-H., Lewin, A., Samrey, U., Myint,H., Meyer z.A., H., et al. (1986) Identification of Rhizobiumplasmid sequences involved in recognition of Psophocar-pus, Vigna, and other legumes. J Cell Biol 102: 1173–1182.
Broughton, W.J., Jabbouri, S., and Perret, X. (2000) Keys tosymbiotic harmony. J Bacteriol 182: 5641–5652.
Büttner, D., and Bonas, U. (2003) Common infection strate-gies of plant and animal pathogenic bacteria. Curr OpinPlant Biol 6: 312–319.
Campellone, K.G., and Leong, J.M. (2003) Tails of two Tirs:actin pedestal formation by enteropathogenic E. coli andenterohemorrhagic E. coli O157:H7. Curr Opin Microbiol6: 82–90.
Candiano, G., Bruschi, M., Musante, L., Santucci, L., Ghig-geri, G.M., Carnemolla, B., et al. (2004) Blue silver: a verysensitive colloidal Coomassie G-250 staining for proteomeanalysis. Electrophoresis 25: 1327–1333.
Chang, J.H., Goel, A.K., Grant, S.R., and Dangl, F.L. (2004)Wake of the flood: ascribing functions to the wave of typeIII effector proteins of phytopathogenic bacteria. Curr OpinMicrobiol 7: 11–18.
Chen, W.-p., and Kuo, T.-T. (1993) A simple and rapidmethod for the preparation of gram-negative bacterialgenomic DNA. Nucleic Acids Res 21: 2260.
Cornelis, G.R. (2000) Type III secretion: a bacterial devicefor close combat with cells of their eukaryotic host. PhilosTrans R Soc Lond B Biol Sci 355: 681–693.
Cornelis, G.R. (2002a) The Yersinia Ysc-Yop ‘type III’ weap-onry. Nat Rev Mol Cell Biol 3: 742–752.
Cornelis, G.R. (2002b) Yersinia type III secretion: send in theeffectors. J Cell Biol 158: 401–408.
Cunnac, S., Occhialini, A., Barberis, P., Boucher, C., andGenin, S. (2004) Inventory and functional analysis of thelarge Hrp regulon in Ralstonia solanacearum: identificationof novel effector proteins translocated to plant host cellsthrough the type III secretion system. Mol Microbiol 53:115–128.
Deakin, W.J., Marie, C., Saad, M.M., Krishnan, H.B., andBroughton, W.J. (2005) NopA is associated with cell sur-face appendages produced by the type III secretion systemof Rhizobium sp. strain NGR234. Mol Plant Microbe Inter-act 18: 499–507.
Fellay, R., Frey, J., and Krisch, H. (1987) Interposonmutagenesis of soil and water bacteria: a family of DNAfragments designed for in vitro insertional mutagenesis ofGram-negative bacteria. Gene 52: 147–154.
Figurski, D.H., and Helinski, D.R. (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans. Proc Natl Acad SciUSA 76: 1648–1652.
Freiberg, C., Fellay, R., Bairoch, A., Broughton, W.J.,Rosenthal, A., and Perret, X. (1997) Molecular basis ofsymbiosis between Rhizobium and legumes. Nature 387:394–401.
Fu, Y., and Galán, J.E. (1999) A Salmonella protein antago-nizes Rac-1 and Cdc42 to mediate host-cell recovery afterbacterial invasion. Nature 401: 293–297.
Gage, D.J. (2004) Infection and invasion of roots by symbi-otic, nitrogen-fixing rhizobia during nodulation of temperatelegumes. Microbiol Mol Biol Rev 68: 280–300.

1316 P. Skorpil et al.
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1304–1317
Gehring, C.A., Irving, H.R., Kabbara, A.A., Parish, R.W.,Boukli, N.M., and Broughton, W.J. (1997) Rapid, plateau-like increases in intracelluar free calcium are associatedwith Nod-factor-induced root-hair-deformation. Mol PlantMicrobe Interact 10: 791–802.
Ghosh, P. (2004) Process of protein transport by the typeIII secretion system. Microbiol Mol Biol Rev 68: 771–795.
Göttfert, M., Röthlisberger, S., Kündig, C., Beck, C., Marty,R., and Hennecke, H. (2001) Potential symbiosis-specificgenes uncovered by sequencing a 410-kilobase DNAregion of the Bradyrhizobium japonicum chromosome. JBacteriol 183: 1405–1412.
He, S.Y., Nomura, K., and Whittam, T.S. (2004) Type IIIprotein secretion mechanism in mammalian and plantpathogens. Biochim Biophys Acta 1694: 181–206.
Heidstra, R., Geurts, R., Franssen, H., Spaink, H.P., vanKammen, A., and Bisseling, T. (1994) Root hair deforma-tion activity of nodulation factors and their fate on Viciasativa. Plant Physiol 105: 787–797.
Hubber, A., Vergunst, A.C., Sullivan, J.T., Hooykaas, P.J.,and Ronson, C.W. (2004) Symbiotic phenotypes and trans-located effector proteins of the Mesorhizobium loti strainR7A VirB/D4 type IV secretion system. Mol Microbiol 54:561–574.
Hueck, C.J. (1998) Type III protein secretion systems in bac-terial pathogens of animals and plants. Microbiol Mol BiolRev 62: 379–433.
Innes, R. (2003) New effects of type III effectors. Mol Micro-biol 50: 363–365.
Jonak, C., Ökrész, L., Bögre, L., and Hirt, H. (2002) Com-plexity, cross talk and integration of plant MAP kinase sig-nalling. Curr Opin Plant Biol 5: 415–424.
Kaneko, T., Nakamura, Y., Sato, S., Asamizu, E., Kato, T.,Sasamoto, S., et al. (2000a) Complete genome structureof the nitrogen-fixing symbiotic bacterium Mesorhizobiumloti. DNA Res 7: 331–338.
Kaneko, T., Nakamura, Y., Sato, S., Asamizu, E., Kato, T.,Sasamoto, S., et al. (2000b) Complete genome structureof the nitrogen-fixing symbiotic bacterium Mesorhizobiumloti (supplement). DNA Res 7: 381–406.
Kaneko, T., Nakamura, Y., Sato, S., Minamisawa, K.,Uchiumi, T., Sasamoto, S., et al. (2002a) Completegenomic sequence of nitrogen-fixing symbiotic bacteriumBradyrhizobium japonicum USDA110. DNA Res 9: 189–197.
Kaneko, T., Nakamura, Y., Sato, S., Minamisawa, K.,Uchiumi, T., Sasamoto, S., et al. (2002b) Completegenomic sequence of nitrogen-fixing symbiotic bacteriumBradyrhizobium japonicum USDA110 (supplement). DNARes 9: 225–256.
Kenny, B., DeVinney, R., Stein, M., Reinscheid, D.J., Frey,E.A., and Finlay, B.B. (1997) Enteropathogenic E. coli(EPEC) transfers its receptor for intimate adherence intomammalian cells. Cell 91: 511–520.
Krause, A., and Broughton, W.J. (1992) Proteins associatedwith root-hair deformation and nodule initiation in Vignaunguiculata. Mol Plant Microbe Interact 5: 96–103.
Krause, A., Sigrist, C.J.A., Dehning, I., Sommer, H., andBroughton, W.J. (1994) Accumulation of transcripts encod-ing a lipid transfer-like protein during deformation of
nodulation-competent Vigna unguiculata root hairs. MolPlant Microbe Interact 7: 411–418.
Krishnan, H.B., Lorio, J., Kim, W.S., Jiang, G., Kim, K.Y.,DeBoer, M., and Pueppke, S.G. (2003) Extracellular pro-teins involved in soybean cultivar-specific nodulation areassociated with pilus-like surface appendages and exportedby a type III protein secretion system in Sinorhizobium frediiUSDA257. Mol Plant Microbe Interact 16: 617–625.
Laemmli, U.K. (1970) Cleavage of structural proteins duringthe assembly of the head of bacteriophage T4. Nature 227:680–685.
Lewin, A., Cervantes, E., Wong, C.-H., and Broughton, W.J.(1990) nodSU, two new nod genes of the broad host rangeRhizobium strain NGR234 encode host-specific nodulationof the tropical tree Leucaena leucocephala. Mol PlantMicrobe Interact 3: 317–326.
Marie, C., Broughton, W.J., and Deakin, W.J. (2001) Rhizo-bium type III secretion systems: legume charmers oralarmers? Curr Opin Plant Biol 4: 336–342.
Marie, C., Deakin, W.J., Viprey, V., Kopcinska, J.,Golinowski, W., Krishnan, H.B., et al. (2003) Characteriza-tion of Nops, nodulation outer proteins, secreted via thetype III secretion system of NGR234. Mol Plant MicrobeInteract 16: 743–751.
Marie, C., Deakin, W.J., Ojanen-Reuhs, T., Diallo, E., Reuhs,B.L., Broughton, W.J., and Perret, X. (2004) TtsI, a keyregulator of Rhizobium species NGR234, is required fortype III-dependent protein secretion and synthesis ofrhamnose-rich polysaccharides. Mol Plant Microbe Interact17: 958–966.
Mathis, R., Van Gijsegem, F., De Rycke, R., D’Haeze, W.,Van Maelsaeke, E., Anthonio, E., et al. (2005) Lipopolysac-charides as a communication signal for progression oflegume endosymbiosis. Proc Natl Acad Sci USA 102:2655–2660.
Mattoo, S., Yuk, M.H., Huang, L.L., and Miller, J.F. (2004)Regulation of type III secretion in Bordetella. Mol Microbiol52: 1201–1214.
Orth, K. (2002) Function of the Yersinia effector YopJ. CurrOpin Microbiol 5: 38–43.
Perino, C., Gaudriault, S., Vian, B., and Barny, M.A. (1999)Visualisation of harpin secretion in planta during infectionof apple seedlings by Erwinia amylovora. Cell Micorbiol 1:131–141.
Perret, X., Staehelin, C., and Broughton, W.J. (2000) Molec-ular basis of symbiotic promiscuity. Microbiol Mol Biol Rev64: 180–201.
Peterson, G.L. (1977) A simplification of the protein assaymethod of Lowry et al. which is more generally applicable.Anal Biochem 83: 346–356.
Pueppke, S.G., and Broughton, W.J. (1999) Rhizobium sp.strain NGR234 and R. fredii USDA257 share exceptionallybroad, nested host ranges. Mol Plant Microbe Interact 12:293–318.
Quandt, J., and Hynes, M.F. (1993) Versatile suicide vectorswhich allow direct selection for gene replacement in Gram-negative bacteria. Gene 127: 15–21.
Reli , B., Perret, X., Estrada-García, M.T., Kopcinska, J.,Golinowski, W., Krishnan, H.B., et al. (1994) Nod factorsof Rhizobium are a key to the legume door. Mol Microbiol13: 171–178.
c¢

NopP of Rhizobium sp. NGR234 1317
© 2005 Blackwell Publishing Ltd, Molecular Microbiology, 57, 1304–1317
Rossier, O., Wengelnik, K., Hahn, K., and Bonas, U. (1999)The Xanthomonas Hrp type III system secretes proteinsfrom plant and mammalian bacterial pathogens. Proc NatlAcad Sci USA 96: 9368–9373.
Saad, M.M., Kobayashi, H., Marie, C., Brown, I.R., Mansfield,J.W., Broughton, W.J., and Deakin, W.J. (2005) NopB, atype III secreted protein of Rhizobium sp. strain NGR234,is associated with pilus-like surface appendages. J Bacte-riol 187: 1173–1181.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecu-lar Cloning: A Laboratory Manual, 2nd edn. Cold SpringHarbour, NY: Cold Spring Harbor Laboratory Press.
Sauvonnet, N., Pradet-Balade, B., Garcia-Sanz, J.A., andCornelis, G.R. (2002) Regulation of mRNA expression inmacrophages after Yersinia enterocolitica infection. Role ofdifferent Yop effectors. J Biol Chem 277: 25133–25142.
Stanley, J., Dowling, D.N., and Broughton, W.J. (1988) Clon-ing of hemA from Rhizobium sp. NGR234 and symbioticphenotype of a gene-directed mutant in diverse legumegenera. Mol Gen Genet 215: 32–37.
Timmers, A.C.J., Auriac, M.-C., and Truchet, G. (1999)Refined analysis of early symbiotic steps of the Rhizo-bium–Medicago interaction in relationship with microtubu-lar cytoskeleton rearrangements. Development 126: 3617–3628.
Trinick, M.J., and Galbraith, J. (1980) The Rhizobium require-
ments of the non-legume Parasponia in relationship to thecross-inoculation concept of legumes. New Phytol 86: 17–26.
Vallance, B.A., and Finlay, B.B. (2000) Exploitation of hostcells by enteropathogenic Escherichia coli. Proc Natl AcadSci USA 97: 8799–8806.
Vinardell, J.M., Ollero, F.J., Hidalgo, Á., López-Baena, F.J.,Medina, C., Ivanov-Vangelov, K., et al. (2004) NolR regu-lates diverse symbiotic signals of Sinorhizobium frediiHH103. Mol Plant Microbe Interact 17: 676–685.
Viprey, V., Del Greco, A., Golinowski, W., Broughton, W.J.,and Perret, X. (1998) Symbiotic implications of type IIIprotein secretion machinery in Rhizobium. Mol Microbiol28: 1381–1389.
White, F.F., Yang, B., and Johnson, L.B. (2000) Prospectsfor understanding avirulence gene function. Curr OpinPlant Biol 3: 291–298.
Yoon, S., Liu, Z., Eyobo, Y., and Orth, K. (2003) Yersiniaeffector YopJ inhibits yeast MAPK signaling pathways byan evolutionarily conserved mechanism. J Biol Chem 278:2131–2135.
Yoshida, S., Uemura, M., Niki, T., Sakai, A., and Gusta, L.V.(1983) Partition of membrane particles in aqueous two-polymer phase systems and its practical use for purificationof plasma membranes from plants. Plant Physiol 72: 105–114.
Copyright © 2022 FDOKUMEN









![Comparison between the reference [i]Rhizobium - Archive ...](https://static.fdokumen.com/doc/165x107/63134e0faca2b42b580d2177/comparison-between-the-reference-irhizobium-archive-.jpg)










