
Occlusal plane canting: a treatment alternative using skeletal ...

Non-occlusal dental microwear variability in a sample ofMiddle and Late Pleistocene human populations from
Europe and the Near East
Alejandro Perez-Pereza*, Vanesa Espurza, Jose Marıa Bermudez de Castrob,Marie Antoinette de Lumleyc, Daniel Turbona
aSect. Anthropology, Department on Animal Biology, University of Barcelona, Av. Diagonal 645, 08028 Barcelona, SpainbC.S.I.C. Museo Nacional de Ciencias Naturales, c\ Jose Gutierrez Abascal 2, 28002 Madrid, Spain
cC.N.R.S. Lab. Paleontologie Humaine et Prehistoire, Universite de Provence, Place Victor Hugo, 13331 Marseille Cedex 3, France
Received 22 October 2001; accepted 31 January 2003
Abstract
Non-occlusal, buccal tooth microwear variability has been studied in 68 fossil humans from Europe and the NearEast. The microwear patterns observed suggest that a major shift in human dietary habits and food processingtechniques might have taken place in the transition from the Middle to the Late Pleistocene populations. Differences inmicrowear density, average length, and orientation of striations indicate that Middle Pleistocene humans had moreabrasive dietary habits than Late Pleistocene populations. Both dietary and cultural factors might be responsible for thedifferences observed. In addition, the Middle Paleolithic Neandertal specimens studied show a highly heterogeneouspattern of microwear when compared to the other samples considered, which is inconsistent with a hypothesis of allNeandertals having a strictly carnivorous diet. The high density of striations observed in the buccal surfaces of severalNeandertal teeth might be indicative of the inclusion of plant foods in their diet. The buccal microwear variabilityobserved in the Neandertals is compatible with an overall exploitation of both plant and meat foods on the basis of foodavailability. A preliminary analysis of the relationship between buccal microwear density and climatic conditionsprevailing in Europe during the Late Pleistocene has been attempted. Cold climatic conditions, as indicated by oxygenisotope stage data, seem to be responsible for higher densities of microwear features, whereas warmer periods couldcorrespond to a reduced pattern of scratch density. Such a relationship would be indicative of less abrasive dietaryhabits, perhaps more meat dependent, during warmer periods.� 2003 Elsevier Science Ltd. All rights reserved.
Keywords: Dietary habits; Buccal microwear; Teeth; Pleistocene; Neandertal
Introduction
The diets of most modern human hunter-gatherers include a high proportion of vegetable
* Corresponding author. Secc. Antropologia, Dpto.Biologia Animal, Fac. Biologia, Universitat de Barcelona,Avda. Diagonal 645, 08028 Barcelona, SPAIN
Journal of Human Evolution 44 (2003) 497–513
0047-2484/03/$ - see front matter � 2003 Elsevier Science Ltd. All rights reserved.doi:10.1016/S0047-2484(03)00030-7

materials. Up to eighty percent in weight has beendocumented for the Kalahari Desert Bushmen(Lee and DeVore, 1976) and for AustralianAborigines (Mulvaney and Golson, 1971). Strictlycarnivorous hunter-gatherer groups, such as theInuit or Lapps, are generally limited to highNorthern latitudes, though some carnivorousgroups, today extinct, were also present in Tierradel Fuego (Gusinde, 1989). Hunter-gatherers inmore temperate climates have traditionally hadmixed economies, and this might also be applicableto human populations since the emergence of earlyHomo. Meat consumption probably played amajor role in the adaptation of humans toopen savannah environments (Aiello, 1995), buthominid hunting or scavenging might have beenvirtually impossible without an effective gatheringadaptation (Wolpoff, 1999: p. 417). Even theaustralopithecines might have taken advantageof high-energy resources, including meat(Sponheimer and Lee-Thorp, 1999) and insects,such as termites (Backwell and Errico, 2001). InEurope, a significant adaptive shift to meat con-sumption in Middle Paleolithic Neandertals hasbeen identified based on amino acid residues andcarbon and nitrogen ratios analyses (Ball et al.,1987; Bocherens et al., 1991; 1999; Fizet et al.,1995). The Neandertals might have obtained pro-teins principally through hunting of terrestrialherbivores, rather than scavenging, and proteinintake from plants might have been insignificant(Richards et al., 2000). On the other hand, the dietof the Upper Paleolithic humans might haveshifted towards a greater exploitation of aquaticresources, with an increase in dietary breadth(Richards et al., 2001).
However, the strong climatic fluctuations inEurope during the Late Pleistocene, spanningmore than 100,000 years of Neandertal evolution,raise some questions about a hypothesis of uni-formity of the Neandertal’s diet. This is especiallyrelevant given the assumptions that Neandertalbehavior patterns were less advanced and, there-fore, less variable than those of Upper Paleolithicpeoples. Oxygen isotope records (Klein, 2000:p. 59) show significant fluctuations in oceanictemperatures during the Last Interglacial, oxygenisotope stage 5 (OIS 5; 127,000–71,000 years
BP—before present), as well as during the LastGlacial period, OIS 4 through OIS 2 (71,000–11,000 BP), which includes a long mild period,referred to as OIS 3 (57,000–24,000 BP). Suchfluctuations certainly altered the ecological con-ditions in Europe through time, and are likely tohave affected food availability and, hence, dietaryadaptations in human populations.
Cultural factors may also account for differ-ences in dietary habits and nutritional adaptations.The expansions of humans out of Africa roughlyparalleled the diversification of lithic assemblages.The Oldowan, Mode I Technology, documentedin many African sites, is similar to the assem-blages described for the sites of Dmanisi (Georgia,1.8 million years old—My) (Gabunia and Vekua,1995; Gabunia and Lordkipanidze, 2000), FuenteNueva 3 (Granada, Spain, 1.1 My) (Martınez-Navarro et al., 1997), and Gran Dolina TD6(Atapuerca, Spain, 0.78–0.86 My) (Carbonellet al., 1995; 1999a; Falgueres et al., 1999; 2001).Among other sites, the Acheulean, Mode IITechnology is present at Ubeidiya (Israel, 1.4 My)(Bar Yosef, 1994), and Trinchera Galerıa TG11(Atapuerca, 0.35 My) (Carbonell et al., 1999b).The Mousterian, Mode III assemblages, character-istic of Middle Paleolithic Neandertals, except forSaint Cesaire which was found in association withChatelperronian levels (Wolpoff, 1999: p. 689),were also made by modern humans at Qafzeh andSkhul (Israel, 0.11–0.09 My) (Bar Yosef, 1994;1995; Bar Yosef and Vandermeersch, 1993;Wolpoff, 1999: p. 595). The Levantine Mousterianhas been recently dated to 50–180,000 years BP byelectron-spin-resonance (ESR) (Grun, 1993) and50–270,000 years BP by Thermoluminescence(TL) (Mercier and Valladas, 1994). Finally, theAurignacian, Mode IV industries, brought in bythe Upper Paleolithic populations, included theuse of bone, ivory, and antler as raw materials fortool manufacture, and a set of new stone artifactssuch as long, thin blades, burins, and scrapers(Klein, 2000: pp. 524–529). This Upper Paleolithicindustry, which is certainly highly sophisticated,might have allowed for complex food processingtechniques and widened dietary resources, whereasLate Pleistocene assemblages, much more limitedin shapes and sizes, might have reflected functional
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513498

activities with less food processing and, perhaps,more abrasive diets. The abrasive potential ofedible materials might have decreased as a conse-quence of such technological evolution, though thehunter-gatherer economic pattern certainly per-sisted. Buccal microwear analysis applied toPleistocene human teeth from different timeperiods and geographical areas should provideinformation about these dietary modifications, aswell as of shifts in dietary habits due to climaticconditions and food availability.
Although many fossil teeth generally show poorenamel preservation, post-mortem damage, notinformative of diet, can be clearly discriminatedfrom well-preserved enamel surfaces (Teaford,1988; King et al., 1999; Perez-Perez et al., 1999).Microwear patterns on preserved enamel dependlargely on the diet of a population (Grine, 1986;Teaford, 1994). Ecological factors related to diethave been shown to affect buccal microwear inmodern hunter-gatherer populations (Laluezaet al., 1996), and sex related differences in buccalmicrowear have also been shown (Perez-Perezet al., 1999). Thus, differences in diet, both withinand between populations, may be evidencedthrough buccal microwear analyses. Microwearpatterns depend not only on diet composition(Teaford and Lytle, 1996), but also on economicpattern, tool technology, and food processing tech-niques (Teaford, 1991; Perez-Perez et al., 1994).Although buccal microwear analyses of modernhuman, hunter-gatherer populations show a clearcorrelation between microwear feature density anddietary habits (Lalueza et al., 1996), buccal micro-wear patterns in fossil teeth from Sima de losHuesos (Perez-Perez et al., 1999) have raised sig-nificant questions about the appropriateness ofusing modern analogues in such studies. Micro-wear studies on ancient humans might requireancient models of enamel microwear for dietaryreconstruction. Therefore, a significant knowledgeof ante-mortem buccal microwear variabilitythrough time and space, as well as of post-mortemenamel damage, is required to be able to makedietary inferences on fossil human populations.The present paper examines the buccal microwearvariability of a sample of Middle and LatePleistocene human teeth from different geographic
areas. Both ante-mortem erosions and post-mortemmicrowear features are analyzed. The relationshipsof the buccal microwear pattern with prevailingclimatic conditions, stone tool technology andfood exploitation strategies are considered in thelight of the debate of the behavioral differ-ences between Neandertals and modern humans inEurope (Stringer and Davies, 2001).
Materials and methods
The sample
The sample studied consisted of teeth from 68fossil humans from Europe and the Near East,spanning from the Middle Pleistocene, OIS 21 tothe Upper Paleolithic and Mesolithic, OIS 1. Onlypremolar and molar teeth were studied, and asingle tooth, preferably the lower-left M1, waschosen as representative of each individual(Table 1). The older specimens studied includedthe teeth from Gran Dolina (Atapuerca, Spain;780,000 BP—OIS 21; Falgueres et al., 1999; 2001),of which only specimens TD6-7 and TD6–10showed well preserved enamel surfaces, Arago(Tautavel, France; 400–480,000 BP—OIS 13,Levels J-H; OIS 12, Levels G-D; OIS 11, Level C)and Sima de los Huesos, (Atapuerca, Spain;350,000 BP-OIS 9; Falgueres et al., 2001). Thesample also included teeth from the more recentspecimens of Montmaurin (129–198,000 BP—OIS6?), La Chaise Abri Suard (126–151,000 BP—OIS 6), La Chaise Abri Bourgeois-Delaunay(112–146,000 BP—OIS 6), Tabun 1 and 2(>127,000 BP—OIS 6, Mercier, 1994; Grun, 1993),Malarnaud 1 (122,000–129,000 BP—OIS 6–5),Jebel Qafzeh 9 (92.5–100,000 BP—OIS 5, BarYosef and Vand, 1993), Skhul 4 (80–120,000 BP—OIS 5, Akazawa et al., 1998),Macassargues (84–122,000 BP—OIS 5), La QuinaV (65,000 BP—OIS 4), Peyrard 2 and 3 (65–74,000 BP—OIS 4), Le Portel 1 (50–70,000 BP—OIS 4), Hortus (France; 59–71,000 BP—OIS 4, Lumley, 1973), Saint Cesaire(36,300 BP—OIS 3), Marillac (40–45,000 BP—OIS3), Amud 1 (41,000 BP—OIS 3; Delson et al.,2000), Gibraltar 2 (48,000 BP—OIS 3; Tillier,1982; Dean et al., 1986). El Salt 2(>40,000 BP—OIS 3), Banyoles (Spain,
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513 499

Table 1Fossil specimens analyzed. Only teeth with preserved enamel surfaces were included in the sample; post-mortem damaged enamelsurfaces were discarded. A single tooth was chosen to represent each individual. All enamel surface replicas and crown molds wereobtained from the original teeth curated at different institutions: British Museum of Natural History, London (Kabwe, Gibraltar2, Tabun 1); Musee de l’Homme, Paris (Montmaurin Le Niche 1—box 9, La Madeleine 1, Cro-Magnon 4, Rond du Barry,Veyrier-sous-Saleve, Abrie Pataud 26224—box 4, La Quina H5—box 26, Malarnaud 1—box 9); Universite de Bordeaux I,Bordeaux (Saint Cesaire, La Chaise Grotte Suard—left mandible fragment—and La Chaise Grotte Bourgeois—mandible fragmentand isolated teeth—, Marillac—isolated teeth); Rockefeller Museum, Jerusalem (Qafzeh 9, Skhul 4, Amud 1, Tabun 2); AlsiusPharmacy, Banyoles (Banyoles); Universidad Complutense, Madrid (Los Azules E2–3, Cuartamentero, El Salt 87); Universite deProvence, Marseille (Peyrard 2, Peyrard 3, Le Portel 1, Macassargues, Hortus—includes H- II, III, IV, V, VI, VIII, Xbis, XI);Musee de la Grotte de l’Arago, Tautavel (individuals 1, 2. 3, 4, 5, 6, 7, 8, 9, 11, 20, 21, 23; the numbers are not specimennumbers); Museo de Ciencias Naturales CSIC, Madrid (Gran Dolina TD6-7 and TD6–10, Sima de los Huesos—Perez-Perez et al.,1999). The modern hunter-gatherer samples (Lalueza et al., 1996) are shown for comparative purposes. The isotopic stage (OIS)was inferred from the dating (years before present, BP) reported by various authors (see material and methods section). The age ofeach specimen is included when available. The Symbol labeled column refers to the signs used in Fig. 6 for the identification of theisolated teeth and the samples. The total number of striations observed in each specimen (NT) and in the comparative samples(NT�s) are included for reference with regard to the Neandertals.
Specimen Tooth OIS Symbol Country Years BP Level NT��
Gran Dolina 10 M1 UR 21 d2 Spain 850,000 TD-6 179Gran Dolina 7 Pm3 UR 21 d1 Spain 850,000 TD-6 168Arago-21 (10-11 yo) M1 LR 13 R France 480–530,000 Level J 180Arago-1 (20–25 yo) M1 LR 12 R France 450–480,000 Level G 194Arago-2 (>40 yo) M1 LR 12 R France 450–480,000 Level G 218Arago-4 (18–25 yo) M2 UR 12 R France 450–480,000 Level G 145Arago-23 (11 yo) M2 LL 12 R France 450–480,000 Level G 90Arago-3 M2 UR 12 R France 440–450,000 Level F 132Arago-7 M1 UR 12 R France 440–450,000 Level F 147Arago-11 M1 LR 12 R France 440–450,000 Level F 260Arago-6 M2 LR 12 R France 400–410,000 Level D 207Arago-20 (12 yo) M1 LR 12 R France 400–410,000 Level D 208Arago-12 M1 LR 12 R France 450,000 ? 141Arago-25 Pm3 UL 12 R France 450,000 ? 112Arago-5 M1 LR 11 R France 370–400,000 Level C 145SH-001 M1 LL 9 T Spain 303–339,000 160SH-002 M1 LR 9 T Spain 303–339,000 201SH-003 Pm4 LL 9 T Spain 303–339,000 222SH-014 M1 LL 9 T Spain 303–339,000 198SH-075 M3 LL 9 T Spain 303–339,000 145SH-016 M1 UL 9 T Spain 303–339,000 194SH-556 M1 LL 9 T Spain 303–339,000 165SH-168 Pm4 LR 9 T Spain 303–339,000 148SH-300 M1 LR 9 T Spain 303–339,000 177SH-009 Pm4 LR 9 T Spain 303–339,000 194SH-286 M1 LL 9 T Spain 303–339,000 140SH-022 M1 LL 9 T Spain 303–339,000 191SH-279 Pm4 UR 9 T Spain 303–339,000 148SH-505 M2 LL 9 T Spain 303–339,000 255SH-888 M1 LL 9 T Spain 303–339,000 102SH-605 M1 LL 9 T Spain 303–339,000 171SH-607 M1 LL 9 T Spain 303–339,000 186SH-169 M3 LL 9 T Spain 303–339,000 272SH-792 M1 LL 9 T Spain 303–339,000 135Montmuarin 1 M1 LL 6? m France 129–198,000 mid M. Pleistocene 112La Chaise Abri Suard M2 LL 6 s France 126–151,000 Level 51 61La Chaise Bourgeois M2 LL 6 b France 112–146,000 Level 11 82Tabun 1 M1 UL 6 1 Israel >127,000 Level C or D? 27
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513500
45,000 BP—OIS 3; Julia and Bischoff, 1991),Cro-Magnon 4 (30,000 BP—OIS 3), Veyrier-sous-Saleve (12–35,000 BP—OIS 2), Abri Pataud(22,000 BP—OIS 2; Bricker and Mellars, 1987),
Rond-du-Barry (17,100 BP—OIS 2; De Bayleand Heim, 1989), La Madeleine 1 (10,190 BP—OIS 1; Gambier et al., 2000), Los Azules (9–10,000 BP—OIS 1; Garralda, 1986) and
Table 1 (continued)
Specimen Tooth OIS Symbol Country Years BP Level NT��
Tabun 2 Pm4 LL 6 1 Israel >127,000 Level C 47Malarnaud 1 M1 LL 5? 2 France 122–129,000 68Qafzeh 9 M1 LL 5 q Israel 92.5–100,000 XVII-XXIV 73Skhul 4 Pm4 LL 5 u Israel 80–120,000 Level B 91Macassargues M2 LL 5 2 France 84–122,000 120La Quina 5 M1 LL 4 2 Israel 65,000 57Peyrard 2 M1 LR 4 2 France 65–74,000 142Peyrard 3 M1 LR 4 2 France 65–74,000 134Le Portel 1 Pm4 LL 4 2 France 50–70,000 126Hortus-II M1 LL 4 3 France 59–71,000 128Hortus-III M1 UR 4 3 France 59–71,000 104Hortus-IV M1 LR 4 3 France 59–71,000 104Hortus-V M1 LL 4 3 France 59–71,000 144Hortus-VI Pm3 LR 4 3 France 59–71,000 153Hortus-VIII M1 UR 4 3 France 59–71,000 103Hortus-Xbis Pm4 UL 4 3 France 59–71,000 122Hortus-XI M3 UR 4 3 France 59–71,000 130Saint Cesaire M1 UL 3 2 France 36,300 Chatelperronian 68Marillac M1 LL 3 2 France 40–45,000 102Amud 1 M1 LL 3 1 Israel 41,500 Level B 57Gibraltar 2 (4-5 yo) m1 LL 3 1 Great Britain 48,000 Mousterian 93El Salt 2 Pm- U- 3 1 Spain >40,000 Mousterian 114Banyoles M3 LL 3 1 Spain 45,000 145Cro-Magnon 4 Pm4 LR 3 U France 30,000 K Horizon 123Veyrier-sous-Saleve M1 UR 2 U France 12–35,000 110Abri Pataud M1 LL 2 U France 22,000 Bed 2 139Rond du Barry M1 UR 2 U France 17,100 96La Madeleine 1 M1 LL 1 U France 9–11,500 100Los Azules Pm4 UL 1 U Spain 9–10,000 Azilian 140Cuartamentero M2 U- 1 U Spain 6–8,000 Epipaleolithic 117
Summary by groupsArago (n = 13) 12 R France 423–478,000 H. heidelbergensis 167.6�48.0Sima de los Huesos (n = 19) 9 T Spain 303–339,000 H. heidelbergensis 179.2�41.4Middle Paleolithic (n = 13) 3–4–5 1,2 F, S, NE 30–120,000 Neandertal 88.9�37.1Hortus (n = 8) 4 3 France 59–71,000 Neandertal 123.5�19.0U. Pal. & Mesolithic (n = 7) 1–2 U France, Spain 6–30,000 modern H. sapiens 117.9�17.9
Modern comparative collection (Lalueza et al., 1996)Andamanese (n = 18) Andaman Tropical hunter-gatherer 74.8�20.5Veddhas (n = 9) Sri Lanka Tropical hunter-gatherer 74.2�28.0Tasmanian (n = 11) Tasmania Arid hunter-gatherer 73.4�29.0Bushmen (n = 15) Africa Arid hunter-gatherer 67.7�16.7Australian (n = 18) Australia Arid hunter-gatherer 66.2�11.0Vancouver Islanders (n = 17) Canada Carnivorous hunter gatherer 41.8�16.1Inuit (n = 20) Greenland Carnivorous hunter gatherer 38.4�18.3Fuegians (n = 20) Argentina Carnivorous hunter gatherer 32.0�13.5
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513 501

Cuartamentero (6–8,000 BP—OIS 1; Garralda,1982). The dating of each fossil specimen wasadopted from the cited literature as well as fromWolpoff (1999) and Delson et al. (2000). OxygenIsotopic Stages were ascribed using the OIS datinggiven by Klein (2000; p. 59).
Ante-mortem vs. post-mortem wear
Taphonomic processes have been shown tocause distinct post-mortem enamel damage (Kinget al., 1999). No association has been foundbetween the type of abrasive particles (phytoliths,sand, dust or ashes) and the striation pattern(Maas, 1991; Ungar, 1992; 1994a; 1994b), norbetween the width of the striations and theirorientation (Ungar, 1994). Experimental analyseswith abrasive agents, with distinct compositionsand particle sizes, have demonstrated that post-depositional processes produce microwear patternsthat are clearly distinct to those formed duringfood consumption and independent of size anddensity of the abrasive particles (Gordon andWalker, 1983; Maas, 1991). Post-depositionalerosive processes, both physical (sediment trans-portation) or chemical (acid erosion), tend to eraseante-mortem microwear features, rather than toincrease them (Teaford, 1988; King et al., 1999).However, the discrimination made by Teaford(1988) between post-mortem artifacts and ante-mortem enamel microwear in occlusal wear facets,might not be applicable to buccal microwearanalyses, for which a clear distinction between wellpreserved and unpreserved enamel still needs tobe done. Thus, detailed comparative analyses ofbuccal enamel surfaces versus interproximal wearfacets and buccal-to-occlusal surfaces were madeto further characterize well preserved enamel onthe buccal surface. The interproximal facets of allthe isolated teeth and the occlusal third of thebuccal surfaces of all the studied sample werescanned and SEM images were obtained at100� magnification. Interproximal facets, eithermesial or distal, could be observed in GranDolina ATD6-7 and ATD6–10, Saint Cesaire,Rond-du-Barry, Abri Pataud, Montmaurin 1 andon most specimens from Sima de los Huesos,Arago and Hortus. The interstitial facets of
premolar and molar teeth, seldom exposed tomicrowear during mastication (except for patho-logical conditions), are formed ante-mortemthrough tooth-to-tooth percussion causingenamel prism-plucking that results in a highlyprism-sized pitted surface lacking any type ofstriations (Puech, 1984; Walker, 1984; Teaford andRunestad, 1992). The absence of occlusal/buccal-like microwear on the interstitial facets may beconsidered as indicative of lack of post-depositional processes affecting dietary relatedmicrowear (Ungar and Teaford, 1996), sincetaphonomic processes are likely to affect in thesame manner all enamel surfaces in the isolatedteeth. Thus, a clear distinction between themwould be indicative of no post-mortem damage.Also, since a clear distinction has been described inthe frequency of striations versus pits between thebuccal and the occlusal surfaces of teeth (Ungarand Teaford, 1996), in the absence of post-mortemdamage buccal-to-nocclusal surfaces are expectedto show a transitional microwear pattern whenapproaching the occlusal surface of teeth.
Buccal microwear analysis
The buccal microwear pattern of each tooth wasderived by analysis of a Scanning Electron Micro-scope (SEM) micrograph of well preserved enamel(Figs. 1a,b). The studied surface covered an area ofexactly 0.56 mm2 on the medial third of the buccalsurface of the tooth, where no enamel defects,dental pathologies, or evidence of post-mortemdamage was observed. Post mortem damage wascharacterized as defined by Teaford (1988) andUngar and Teaford (1996), and from the post-mortem analyses made here. Replicas of buccalenamel surfaces were obtained with Triafol(Balzers Union) and with President Jet (Coltene)molding materials for full crown analyses, follow-ing standard procedures fully described elsewhere(Perez-Perez, 1990; Lalueza et al., 1993a; 1993b;Perez-Perez et al., 1994; 1999). All SEM micro-graphs were obtained at approximately 100�magnification, covering an area of 0.56 mm2 ofenamel on the medial third of the buccal side of thetooth crown. The side of each square SEM imagemeasures 748.331 µm. Image processing included a
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513502
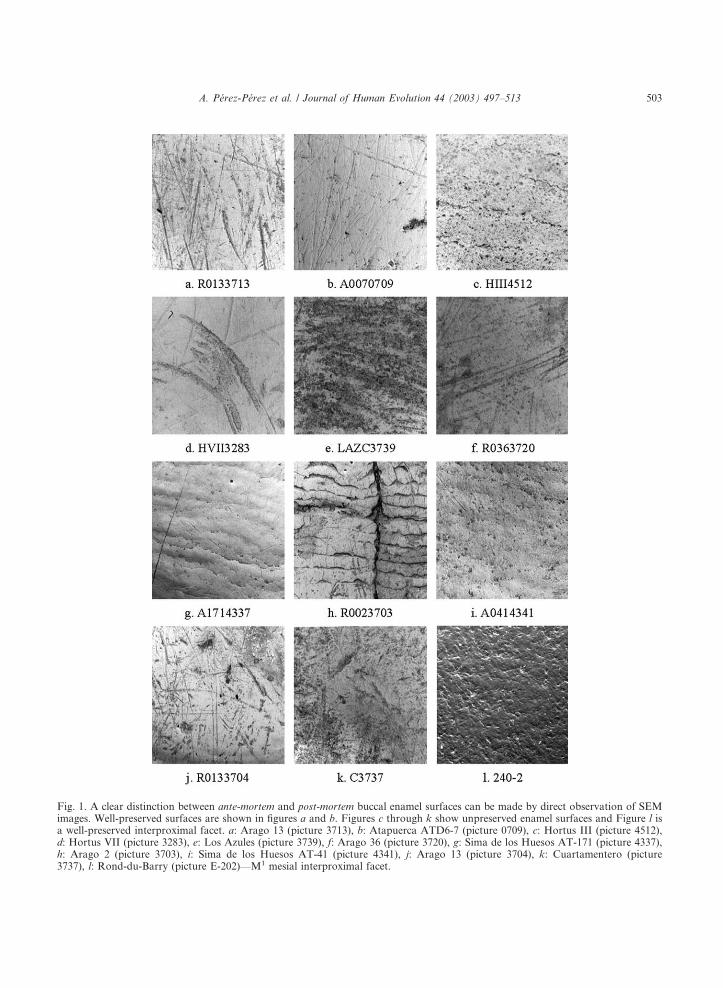
Fig. 1. A clear distinction between ante-mortem and post-mortem buccal enamel surfaces can be made by direct observation of SEMimages. Well-preserved surfaces are shown in figures a and b. Figures c through k show unpreserved enamel surfaces and Figure l isa well-preserved interproximal facet. a: Arago 13 (picture 3713), b: Atapuerca ATD6-7 (picture 0709), c: Hortus III (picture 4512),d: Hortus VII (picture 3283), e: Los Azules (picture 3739), f: Arago 36 (picture 3720), g: Sima de los Huesos AT-171 (picture 4337),h: Arago 2 (picture 3703), i: Sima de los Huesos AT-41 (picture 4341), j: Arago 13 (picture 3704), k: Cuartamentero (picture3737), l: Rond-du-Barry (picture E-202)—M1 mesial interproximal facet.
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513 503

high pass (50 pixel) filter and automatic leveladjustment in Photoshop 5.0 (Adobe�). Striationmeasurements were made with SigmaScan (JandelScientific�). Within each analyzed surface micro-wear measurements included the number (N),average length (X) and standard deviation of thelength (S) of all observed striations (T), as well asby categories of orientation in 45-degree intervals:vertical (V), horizontal (H), mesio-occlusal todisto-cervical (MD), and disto-occlusal to mesio-cervical (DM). Thus, a total of 15 variables (N, Xand S for T, V, H, MD and DM orientations) werederived for the sample studied, as well as for somecomparative modern hunter-gatherer populations(Lalueza et al., 1996). Within each fossil groupconsidered (Arago, Sima de los Huesos, Hortus,Middle Paleolithic, and Upper Paleolithic) all thevariables studied passed Kolmogorov-SmirnovNormality tests.
A multivariate Principal Component (PC)analysis (SPSS PC, v. 9) was carried out with mostspecimens (n = 61), except the isolated teeth fromGran Dolina (TD6-7, TD6-10), La Chaise AbriSuard and Bourgeois-Delaunay, Montmaurin,Skhul 4, and Jebel Qafzeh 9, for which PC valueswere computed using the functions derived fromthe analysis. The first three PC factors extractedexplained 70.6% of the total variance (44.5%,16.2%, and 9.9% respectively). Significant differ-ences (ANOVA) between the chronologicallydefined groups (Lower Paleolithic—LP, MiddlePaleolithic—MP, and Upper Paleolithic—UP)were observed for the first (F = 5.872, P = 0.005)and the second (F = 17.883, P = 0.000) PrincipalComponents, but not for the third (F = 0.926,P = 0.402). A Bonferroni post-hoc test showed thatdifferences in buccal microwear were only obtainedin the comparisons of LP-UP for PC1 (P = 0.006),LP-MP for PC2 (P<= 0.001), and MP-UP for PC2
(P = 0.033).A Canonical Analysis of Populations (CANP),
including a Multivariate Analysis of Variance(IANOVER), was applied to the PC factors usingthe canp module of the non-standard MultiCua�
statistical package (Arenas et al., 1991). Thisanalysis is useful to graphically represent sampleswith maximum power of discrimination based onthe Mahalanobis distance, and allows single indi-
viduals to be included in the plot along with theother groups but excluding them from the calcu-lation of the matrix of dispersion of the means foreach group (Arenas et al., 1991). Six groups,geographically and chronologically defined(R—Arago, T—Sima de los Huesos, U—UpperPaleolithic and Mesolithic, 1—Middle Paleolithicfrom the Near East and Spain, 2—Middle Paleo-lithic from France, and 3—Hortus), entered theCANP analysis in order to maximize both chrono-logical and geographical differences amongsamples, and seven isolated specimens (mt, cs, cb,sk, qz9, gd7, gd10) were plotted alongside thegroups defined. Only the three main PC factorsentered the CANP analysis for maximum sim-plicity in the interpretation of variables and theircorrelations. However, the inclusion of a largernumber of factors did not significantly increase thediscrimination power. A significant MANOVAvalue was obtained (F = 2.7315; F� = 1.4765,P<0.05; no post-hoc test is available within theCANP package) reflecting clear inter-populationdifferences in their buccal microwear patterns. Thecanonical variables (CV1, CV2, CV3), derived fromthe canonical transformation, explained 58.4%,30.5% and 11.1% of the variance represented bythe three original principal components thatentered the analysis. CV1 was mainly correlatedwith PC3 (R = 0.78), CV2 with PC3 (R = 0.76) andPC1 (R = 0.63), and CV3 with PC2 (R = 0.85).Table 2 shows the correlations among the PCfactors, the CV functions and the original vari-ables. CV1 mainly reflects differences in the lengthof the horizontal striations (XH); CV2 greatlydepends on the length of all striations; and CV3
reflects differences in the overall number ofstriations.
Results and discussion
Analysis of enamel surfaces
Well preserved enamel on the buccal surface ofteeth always showed a distinct glow and a clearmicrowear pattern (Figs. 1a,b) consisting ofstriations of various lengths with a main occlusal-to-cervical orientation. Unpreserved patches ofenamel were also observed on the buccal surface of
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513504
the analyzed teeth, always showing either enamelprisms exposure (Fig. 1c), an intense pattern ofparallel striations (Figs. 1d–f), perykimata lines(Figs. 1g,h), a generalized pitted surface (Fig. 1i) oran intense combination of pitting and striations(Figs. 1j,k). These unpreserved surfaces could beclearly discriminated, not only from well preservedones (Figs. 1a, 1b), but also from the interstitialwear facets (Fig. 1l), that always showed a highlypitted topography (Figs. 2a–e), without the char-acteristic occlusal/buccal microwear patternpresent on well-preserved surfaces. Many of theinterstitial facets also showed subvertical grooves(Fig. 3), a frequent trait in Neandertal teeth,probably caused by strong tooth-to-tooth chewingforces (Villa and Giacobini, 1995). Finally, wellpreserved buccal-to-occlusal transitional enamelsurfaces tended to show an increased number ofvertical striations and pits when approaching theocclusal rim of the tooth crown (Figs. 4a,b), and
lacked the characteristic prism pitting of the inter-stitial wear facets. A systematic selection of well-preserved enamel surfaces for dietary analysiscould, thus, be done since on the medial third ofthe buccal surface of teeth well preserved enamelcould be easily discriminated from non-dietarypost-mortem affected enamel surfaces.
Dietary habits of fossil groups
There seems to be a mismatch between thebuccal microwear patterns of the fossil samplesstudied and those of the comparative modernhuman populations. Modern hunter-gatherersfrom arid environments, such as AustralianAborigines or Bushmen, show a less abrasivemicrowear pattern (Fig. 5) than that of the UpperPaleolithic humans, although no significant differ-ences in diet composition between them should beexpected. Furthermore, great divergences in themicrowear patterns are evident between the strictlycarnivorous modern groups (Inuit and Fuegian)and the Neandertals, despite the fact that eachmight have relied intensely on hunting strategiesfor food acquisition. Thus, modern models ofmicrowear might not be representative of pastpopulations’ diet.
Fig. 6 shows the 85% confidence regions of theCanonical Representation of the three PrincipalComponents analyzed for the six fossil samplesconsidered, along with the isolated specimens thatdid not enter the analysis. Within the fossilsamples, a clear separation appears between theMiddle and Late Pleistocene groups, the oldersamples having more, though shorter, striations.The Middle Pleistocene groups (Sima de losHuesos and Arago) show a distinct microwearpattern, with the largest density of striations, fol-lowed by the Neandertals from Hortus, the UpperPaleolithic modern humans and the Middle Paleo-lithic Neandertals. Differences in food processingtechniques and tool technologies might accountfor the highly abrasive diet of the Middle Pleisto-cene populations. The Middle and Upper Paleo-lithic samples overlap somewhat in their variation,and the Middle Paleolithic group clearly differsfrom Hortus, which shows a microwear patterncharacterized by a large number of relatively short
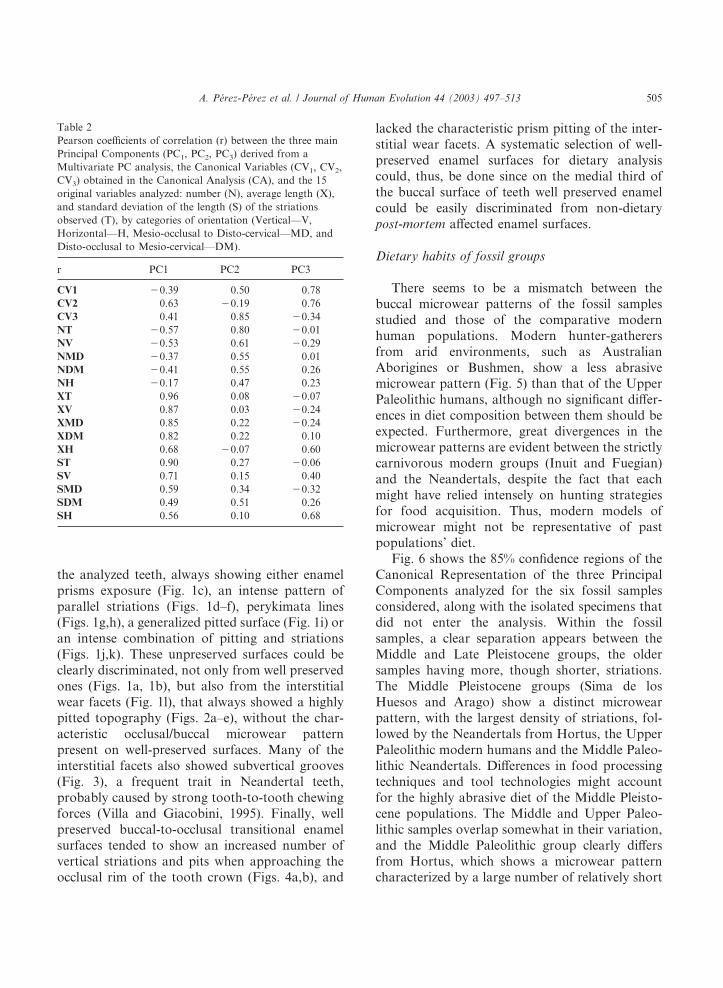
Table 2Pearson coefficients of correlation (r) between the three mainPrincipal Components (PC1, PC2, PC3) derived from aMultivariate PC analysis, the Canonical Variables (CV1, CV2,CV3) obtained in the Canonical Analysis (CA), and the 15original variables analyzed: number (N), average length (X),and standard deviation of the length (S) of the striationsobserved (T), by categories of orientation (Vertical—V,Horizontal—H, Mesio-occlusal to Disto-cervical—MD, andDisto-occlusal to Mesio-cervical—DM).
r PC1 PC2 PC3
CV1 �0.39 0.50 0.78CV2 0.63 �0.19 0.76CV3 0.41 0.85 �0.34NT �0.57 0.80 �0.01NV �0.53 0.61 �0.29NMD �0.37 0.55 0.01NDM �0.41 0.55 0.26NH �0.17 0.47 0.23XT 0.96 0.08 �0.07XV 0.87 0.03 �0.24XMD 0.85 0.22 �0.24XDM 0.82 0.22 0.10XH 0.68 �0.07 0.60ST 0.90 0.27 �0.06SV 0.71 0.15 0.40SMD 0.59 0.34 �0.32SDM 0.49 0.51 0.26SH 0.56 0.10 0.68
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513 505

striations. The specificity of Hortus might berelated to the cold climatic conditions of OIS 4.However, Sima de los Huesos, dated to the warmOIS 9 period, also shows a highly abraded micro-wear pattern, similar to that of Arago, wheremost individuals lived during the cold OIS 12period. If cold climates were related to an exclusivedependence on meat for subsistence, a reduceddensity of striations would be expected for themore carnivorous groups, but this is not the case.It is possible that cold climates forced an increaseddegree of hard plant food intake, perhaps on a
seasonal basis, thereby taking advantage of bothplant and meat foods. Alternatively, similar tooltechnologies may cause comparative microwearpatterns, independently of climatic conditions.
Certainly, buccal microwear patterns seemto depend on lithic technologies. TechnologyModes I and II correspond to the more abradedmicrowear patterns observed, whereas technologyModes III and IV show less microwear features.The similarities in the microwear patterns ofArago and Sima de los Huesos might be indicativeof similar technologies for food acquisition and
Fig. 2. Well-preserved interproximal facets. No ante-mortem formed striations are observed. All surfaces consist of highly pittedsurfaces formed by tooth-to-tooth percussion during mastication of food. a: V202i—Rond-du-Barry, M1 UL distal facet, b:V024901—Sima de los Huesos AT-576, c: V201i—Rond du Barry Pm4 LL proximal facet, d: 252-1—Sima de los Huesos AT-946interproximal facet at 15.3�, e: same as d at 102�.
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513506

processing of Middle Pleistocene populations,which contributed to the highly abrasive nature oftheir diets. These diets probably included signifi-cant amounts of highly abrasive food resources(such as seeds, nuts, bulbs or roots). On the otherhand, Late Pleistocene Neandertals and modernhumans might have either relied more intensivelyon less abrasive foodstuffs (such as meat or fruits)or more efficiently processed them (i.e. the use offire for cooking) than the Middle Pleistocenepopulations, since the former show fewer striations(Fig. 7). As for the isolated teeth, the microwearpatterns of Gran Dolina resemble those of theMiddle Pleistocene samples, whereas the micro-wear of the teeth from La Chaise and the Skhuland Qafzeh modern humans is similar to that ofthe Middle Paleolithic Neandertals.
Neandertal’s diet
The high intragroup variation in the number ofstriations observed in the Neandertal specimensstudied, including Hortus, does not sustain ahypothesis of a homogeneous and strictly car-nivorous diet, with no consumption whatsoever ofplant foods, at least for the specimens studied. Ofcourse, this does not contradict the hypothesis thatprotein intake mostly came from hunting activities
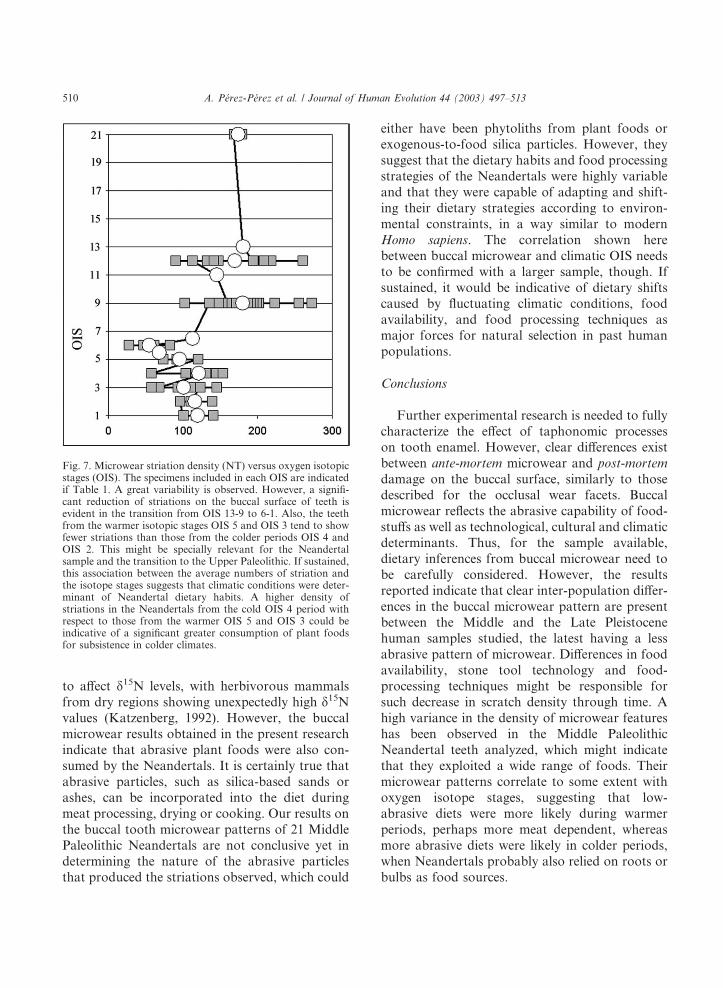
in the Neandetals (Richards et al., 2000). Asopposed to what is observed in the Lower Paleo-lithic groups, where technology was probably themain factor affecting buccal tooth microwear, dif-ferences in microwear patterns within the MiddlePaleolithic sample seem to reflect to a certainextent climatic fluctuations through time, ratherthan geographic dispersion. Within the Late Pleis-tocene sample, Montmaurin 1, La Chaise, Tabun 1and 2 are dated to the cold OIS 6; Malarnaud,Macassargues, Qafzeh 9 and Skhul 4 are dated tothe warm OIS 5; La Quina 5, Peyrard 2 and 3, LePortel 1, and Hortus (n = 8) are dated to the coldOIS 4; Saint Cesaire, Marillac, Amud 1, Gibraltar2, El Salt 2, Banyoles and Cro-Magnon 4 belongto the warm OIS 3 period; Veyrier-sous-Saleve,Abri Pataud and Rond du Barry correspond toOIS 2; and La Madeleine 1, Los Azules andCuartamentero are included in the OIS 1. Thisclassification depends not only on the accuracy ofthe dating of each specimen, but also on theprevailing local climatic conditions, that may varygeographically within a specific OIS. Despite this,considering the available data, it seems that themicrowear pattern of the specimens belonging tothe warmer OIS tend to show fewer striations thanthose from colder OIS, specially for the span fromOIS 2 to 5 (cold OIS 2, NT = 115.0, n = 3; warmOIS 3, NT = 100.3, n = 7; cold OIS 4, NT = 120.6,n = 12; warm OIS 5, NT = 94.7, n = 3). Thesample sizes for each OIS are small yet to drawsignificant conclusions. However, from Fig. 7 itcan be hypothesized that cold climatic conditionsmight cause an increased number of striations (i.e.a more abrasive diet during OIS 2 and 4) whereaswarmer periods might allow for less abrasivedietary habits during OIS 3 and 5, perhaps moremeat dependent. A carnivorous diet has alreadybeen suggested by one of us (A.P.P.), using thedensity of striations (NT), for Tabun 1 (NT = 27),Tabun 2 (NT = 47), Amud 1 (NT = 57), and LaQuina 5 (NT = 57), and also for Gibraltar 2(NT = 93), based on microwear analyses (Laluezaet al., 1993a; 1996). These Neandertal specimens,along with Malarnaud (NT = 68) and SaintCesaire (NT = 68), are the ones that show thelowest total number of striations (Table 1). Carbonand nitrogen isotope values have suggested a
Fig. 3. Interproximal facet of Sima de los Huesos AT-970, at14.4�. Three subvertical grooves are present and are indicativeof strong chewing forces and tooth-to-tooth micropercussion.
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513 507

highly carnivorous diet also for Marillac (Fizetet al., 1995), which shows 102 striations. From thebuccal microwear results, Malarnaud, SaintCesaire, and tentatively also Marillac, might beincluded in the “more carnivorous” category.However, El Salt (NT = 114), Macassargues(NT = 120), and Hortus (NT =123.5�19.0, n = 8)have significantly more striations, though stillfewer than the samples from the OIS 4 period(where NT ranges from 57 to 153). The microwearpattern of the Neandertals reflects a continuum ofstriation densities which is consistent with wide-
ranging, highly adaptive, and climatic-dependentdietary habits.
Although meat foods might have been prepon-derant in Neandertal’s diet, particularly duringwarmer periods, a more abrasive diet, moreheavily relying on hard foods, such as roots orbulbs, might have gained in importance duringcolder periods. The high �15N levels reported forthe Neandertals from Vindija (10.1‰ and 10.8‰),Marillac (9.3‰ and 11.6‰) and Scladina (10.9‰)indicate a high consumption of proteins fromanimal sources through hunting strategies. Marine
Fig. 4. Composition of SEM images of the buccal-to-occlusal enamel to show the transition from buccal to occlusal well-preservedsurfaces. A clear transitional pattern from striation to pits is evident. a: V202—Rond du Barry M1 UR, b: V208—Mountmaurin M1LL.
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513508

foods may also cause increased �15N levels inhumans (Walker and DeNiro, 1986). Marinevertebrates show �15N levels that can be 6‰ to 8‰higher than those in terrestrial vertebrates at
similar trophic levels (Schoeninger et al., 1983;Walker and DeNiro, 1986; Katzenberg, 1992;Schoeninger, 1995). Climatic conditions (Heatonet al., 1986; Schwarcz et al., 1999) and metabolicfactors in water-conserving mammals (Ambroseand DeNiro, 1986; 1987) have also been reported
Fig. 5. Between-population variability of the total striationnumber (NT) and the average length of all striations (XT). Theshaded boxes show the confidence intervals of the mean numberof striations (NT�eNT—error of the mean), and the lines acrossthe box-plots show the 68% confidence interval around themean (XNT�SNT), for all the groups compared, either fossil ormodern. The superimposed points show the average length ofthe striations (XT) for each group and the curve is the quadraticregression curve for this variable. The groups are sorted fromtop to bottom by increasing total number of striations (NT).The average length of the striations decreases with increasingnumber of striations due to fragmentation of striations inhighly abraded surfaces. Data of modern hunter-gathererscome from Lalueza et al., (1996), and that of Atapuerca is fromSima de los Huesos (Perez-Perez et al., 1999). The MiddlePaleolithic sample includes the Neandertal teeth and the UpperPaleolithic one includes the modern Homo sapiens teeth (Table1). The Neandertal teeth from Hortus are considered separatelyfrom the Middle Paleolithic group because they clearly differ intheir buccal microwear patterns.
Fig. 6. Two-dimensional canonical representation of the buccaltooth microwear pattern of all fossil groups studied. The axesshown are the first two Canonical Variables (CV1 and CV2)derived from the main 3 Principal Components (PC) extractedfrom the 15 original variables considered. All 15 variablesentered the PC analysis and the first 3 PC factors derived wereused in a Canonical Analysis. The 3 PC factors used explain70.6% of the total variance of the 15 original variables (NT,XT, ST, NV, XV, SV, NH, XH, SH, NMD, XMD, SMD,NDM, XDM, SDM). All the variables passed Kolmogorov-Smirnov Normality tests within each group considered. Thecircles include 85% confidence regions of the within-groupvariability, and explain 88.9% of the total variance of the 3 PCfactors used in the Canonical Analysis. The upper case letters(R, T, U) and the numbers (1, 2, 3) in the plot are indicative ofthe samples that entered the Canonical Analysis: R—Arago;T—Sima de los Huesos; U—Upper Paleolithic and Mesolithic;1—Middle Paleolithic from the Near East and Spain;2—Middle Paleolithic from France; and 3—Middle PaleolithicHortus (Table 1). The groups were defined in order to maximizeboth chronological and geographical differences amongsamples. The lower case letters show the relative position withinthe Canonical Analysis plot of the isolated specimens:m—Montmaurin 1; k—Kabwe; s—La Chaise Suard; b—LaChaise Bourgeois; u—Skhul 4; q—Qafzeh 9; d1—Gran DolinaTD6-7; d2—Gran Dolina TD6—10 (Table 1). The betweenpopulation variability observed reflects differences in the micro-wear pattern. The x-axis (CV1) is mainly correlated with theaverage density of striations, and the y-axis is mainly indicativeof differences in the average length of the striations (Table 2).
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513 509
to affect �15N levels, with herbivorous mammalsfrom dry regions showing unexpectedly high �15Nvalues (Katzenberg, 1992). However, the buccalmicrowear results obtained in the present researchindicate that abrasive plant foods were also con-sumed by the Neandertals. It is certainly true thatabrasive particles, such as silica-based sands orashes, can be incorporated into the diet duringmeat processing, drying or cooking. Our results onthe buccal tooth microwear patterns of 21 MiddlePaleolithic Neandertals are not conclusive yet indetermining the nature of the abrasive particlesthat produced the striations observed, which could
either have been phytoliths from plant foods orexogenous-to-food silica particles. However, theysuggest that the dietary habits and food processingstrategies of the Neandertals were highly variableand that they were capable of adapting and shift-ing their dietary strategies according to environ-mental constraints, in a way similar to modernHomo sapiens. The correlation shown herebetween buccal microwear and climatic OIS needsto be confirmed with a larger sample, though. Ifsustained, it would be indicative of dietary shiftscaused by fluctuating climatic conditions, foodavailability, and food processing techniques asmajor forces for natural selection in past humanpopulations.
Conclusions
Further experimental research is needed to fullycharacterize the effect of taphonomic processeson tooth enamel. However, clear differences existbetween ante-mortem microwear and post-mortemdamage on the buccal surface, similarly to thosedescribed for the occlusal wear facets. Buccalmicrowear reflects the abrasive capability of food-stuffs as well as technological, cultural and climaticdeterminants. Thus, for the sample available,dietary inferences from buccal microwear need tobe carefully considered. However, the resultsreported indicate that clear inter-population differ-ences in the buccal microwear pattern are presentbetween the Middle and the Late Pleistocenehuman samples studied, the latest having a lessabrasive pattern of microwear. Differences in foodavailability, stone tool technology and food-processing techniques might be responsible forsuch decrease in scratch density through time. Ahigh variance in the density of microwear featureshas been observed in the Middle PaleolithicNeandertal teeth analyzed, which might indicatethat they exploited a wide range of foods. Theirmicrowear patterns correlate to some extent withoxygen isotope stages, suggesting that low-abrasive diets were more likely during warmerperiods, perhaps more meat dependent, whereasmore abrasive diets were likely in colder periods,when Neandertals probably also relied on roots orbulbs as food sources.
Fig. 7. Microwear striation density (NT) versus oxygen isotopicstages (OIS). The specimens included in each OIS are indicatedif Table 1. A great variability is observed. However, a signifi-cant reduction of striations on the buccal surface of teeth isevident in the transition from OIS 13-9 to 6-1. Also, the teethfrom the warmer isotopic stages OIS 5 and OIS 3 tend to showfewer striations than those from the colder periods OIS 4 andOIS 2. This might be specially relevant for the Neandertalsample and the transition to the Upper Paleolithic. If sustained,this association between the average numbers of striation andthe isotope stages suggests that climatic conditions were deter-minant of Neandertal dietary habits. A higher density ofstriations in the Neandertals from the cold OIS 4 period withrespect to those from the warmer OIS 5 and OIS 3 could beindicative of a significant greater consumption of plant foodsfor subsistence in colder climates.
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513510

Acknowledgements
We thank M.D. Garralda, B. Vandermeersch,J.L. Heim, H. de Lumley, and the RockefellerMuseum in Jerusalem, for providing access to theircollections and for their kindness. Three anony-mous reviewers greatly contributed to the finalversion of this paper. This research was funded bythe DGCyT BMC2000-0538. SEM micrographswere obtained at the Serveis Cientificotecnics ofthe University of Barcelona.
References
Aiello, L.C., 1995. The expensive-tissue hypothesis: the brainand digestive system in human and Primate evolution. Curr.Anthropol. 36, 199–221.
Akazawa, T., Aoki, K., Bar-Yosef, O., 1998. Neandertals andmodern humans in Western Asia. Plenum, New York.
Ambrose, S.H., DeNiro, M.J., 1986. Reconstruction of Africanhuman diet using bone collagen carbon and nitrogenisotope ratios. Nature 319, 321–324.
Ambrose, S.H., DeNiro, M.J., 1987. Bone nitrogen isotopecomposition and diet. Nature 325, 201.
Arenas, C., Cuadras, C.M., Fortiana, J., 1991. MultiCua.Paquete no standard de Analisis Multivariante. Publ. 4,Departament d’Estadıstica, Universitat de Barcelona,Barcelona.
Backwell, L.R., Errico, F., 2001. Evidence of termite foragingby Swartkrans early Hominids. Proc. Natl. Acad. Sci. 98,1358–1363.
Ball, G., Saint Martin, G., Beraud-Colomb, E., 1987. A sys-tematic study of the amino acid composition and D/L ratiosin fossil bones from two French Neandertalian sites. Hum.Evol. 2, 289–296.
Bar Yosef, O., 1994. The Lower Paleolithic of the Near East. J.World Prehist. 8, 211–265.
Bar Yosef, O., 1995. The Lower and Middle Paleolithic in theMediterranean Levant; chronology and cultural entities, In:H. Ullrich, (ed.), Man and Environment in the Paleolithic.E.R.A.U.L. 62: 247–263.
Bar Yosef, O., Vandermeersch, B., 1993. Modern humans inthe Levant. Sci. Am. 267, 94–100.
Bocherens, H., Fizet, M., Mariott, A., Lange-Badre, B.,Vandermeersch, B., Borel, J.P., Bellon, G., 1991. Isotopicbiogeochemistry (13C, 15N) of fossil vertebrae collagen:application to the study of past food web includingNeandertal man. J. Hum. Evol. 20, 481–492.
Bocherens, H., Billiou, D., Mariotti, A., Patou-Mathis, M.,Otte, M., Bonjean, D., Toussaint, M., 1999. Peleoenviron-mental and paleodietary implications of isotopic biogeo-chemistry of Last Interglacial Neandertal and mammalbones in Scladina Cave (Belgium). J. Archaeol. Sci. 26,599–607.
Bricker, H., Mellars, P., 1999. Datations 14C de l’Abri Pataud(Les Eyzies, Dordogne) par le procede “accelerateur-spectrometer de masse”. L’Antropologie 91, 227–234.
Carbonell, E., Bermudez de Castro, J.M., Arsuaga, J.L., Dıez,J.C., Rosas, A., Cuenca-Bescos, G., Sala, R., Mosquera,M., Rodrıguez, X.P., 1995. Lower Pleistocene Hominidsand artefacts from Atapuerca-TD6 (Spain). Science 269,826–830.
Carbonell, E., Garcıa-Anton, M.D., Mallol, C., Mosquera, M.,Olle, A., Rodrıguez, X.P., Sahnouni, M., Sala, R., Verges,J.M., 1999a. The TD6 level lithic industry from GranDolina, Atapuerca (Burgos, Spain): production and use. J.Hum. Evol. 37, 653–693.
Carbonell, E., Marquez, B., Mosquera, M., Olle, A.,Rodrıguez, X.P., Sala, R., Verges, J.M., 1999b. El modo IIen Galerıa. Analisis de la industria lıtica y sus procesostecnicos, in: Carbonell, E., Rosas, A., Dıez, J.C. (Eds.),Atapuerca: ocupaciones humanas y Paleoecologıa delyacimiento de Galerıa. Junta de Castilla y Leon, Valladolid.
De Bayle des Hermens, R., Heim, J.L., 1989. Decouverte d’uncrane humain dans une sepulture secondaire deMagdalenien I de la grotte de Rond-du-Barry, Polignac,Haute-Loire. Paleontology Humaine—C.R. Acad. Sci.Paris 309, 1349–1352.
Dean, M.C., Stringer, C.B., Bromage, T.G., 1986. Age at deathof the Neanderthal child from Devil’s Tower, Gibraltar, andthe implications for studies of general growth and devel-opment in Neanderthals. Am. J. Phys. Anthropol. 70,301–309.
Delson, E., Tattersall, I., Van Couvering, J.A., Brooks, A.S.(Eds.), 2000. Encyclopedia of human evolution andprehistory. Garland Pub. Inc, New York.
Falgueres, C., Bahainm, J.-J., Yokoyama, Y., Arsuaga, J.L.,Bermudez de Castro, J.M., Carbonell, E., Bischoff, J.L.,Dolo, J.M., 1999. Earliest humans in Europe: the age ofTD6 Gran Dolina, Atapuerca, Spain. J. Hum. Evol. 37,343–352.
Falgueres, C., Bahain, J.-J., Yokoyama, Y., Bischoff, J.L.,Arsuaga, J.L., Bermudez de Castro, J.M., Carbonell, E.,Dolo, J.M., 2001. Datation par RPE et U-TH des sitespleistocenes d’Atapuerca: Sima de los Huesos, TrincheraDolina et Trinchera Galerıa. bilan geochronologique.L’Anthropologie 105, 71–81.
Fizet, M., Mariotti, A., Bocherens, H., Lange-Badre, G.,Vandermeersch, B., Borel, J., Bellon, G., 1995. Effect of dietphysiology and climate on carbon and nitrogen stableisotopes of collagen in Late Pleistocene anthropic paleo-echosystem (France, Charente, Marillac). J. Archaeol. Sci.22, 67–79.
Gabunia, L., Lordkipanidze, D., 2000. The environmentalcontexts of early human occupation of Georgia(Transcaucasia). J. Hum. Evol. 38, 785–802.
Gabunia, L., Vekua, A., 1995. A Plio-Pleistocene hominidfrom Dmanisi, east Georgia, Caucasus. Nature 373,509–512.
Gambier, D., Valladas, H., Tisnerat-Laborde, N., Arnold, M.,Bresson, F., 2000. Datation de vestiges humaines presumes
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513 511

du Paleolithique Superieur par la methode du Carbone 14en spectrometrie de masse par accelerateur. Paleo 12,201–212.
Garralda, M.D., 1982. El craneo asturiense de Cuartamentero(Llanes, Oviedo). Kobie 12.
Garralda, M.D., 1986. The Azilian man from Los AzulesCave I (Cangas de Onıs, Oviedo, Sapin). Hum. Evol. 1,431–448.
Gordon, K.D., Walker, A., 1983. Taphonomy of dental micro-wear: can fossil microwear be studied productively? Am. J.Phys. Anthropol.. 60, 200.
Grine, F.E., 1986. Dental evidence for dietary differences inAustralopithecus and Paranthropus: a quantitative analysisof permanent molar microwear. J. Hum. Evol. 15,783–822.
Grun, R., 1993. Electron spin resonance dating inPaleoanthropology. Evol. Anthropol. 2, 172–181.
Gusinde, M., 1989. Los Indios de Tierra del Fuego. Antro-pologıa Fısica. Centro Argentino de Etnologıa Americana,Buenos Aires. Tomo IV, Vols. I and, II, ISBN950-9252-02-6.
Heaton, T.H.E., Vogel, J.C., Chevallaire, G., Collett, G., 1986.Climatic influence on the isotopic composition of bonenitrogen. Nature 322, 822–823.
Julia, R., Bischoff, J.L., 1991. Radiometric dating ofQuaternary deposits and the Hominid mandible of LakeBanyolas, Spain. J. Archaeol. Sci. 18, 707–722.
Katzenberg, M.A. 1992. Skeletal biology of past people:research methods. Wiley, New York, pp. 105–119.
King, T., Andrews, P., Boz, B., 1999. The effect of taphonomicprocesses on dental microwear. Am. J. Phys. Anthropol.108, 359–373.
Klein, R.G., 2000. The human career. Human biologicaland cultural origins. The Univ. of Chicago Press, Chicago.p. 59.
Lalueza, C., Perez-Perez, A., 1993a. The diet of the Neandertalchild Gibraltar 2 (Devil’s Tower) through the study of thevestibular striation pattern. J. Hum. Evol. 24, 29–41.
Lalueza, C., Perez-Perez, A., Turbon, D., 1993b. Microscopicstudy of the Banyoles mandible (Girona, Spain): diet,cultural activity and tooth-pick use. J. Hum. Evol. 24,281–300.
Lalueza, C., Perez-Perez, A., Turbon, D., 1996. Dietary infer-ences through buccal microwear analysis of Middle andLate Pleistocene human fossils. Am. J. Phys. Anthropol.100, 367–387.
Lee, R.B., DeVore, I., 1976. Kalahari hunter-gatherers: studiesof !Kung San and their neighbours. University Press,Harvard.
de Lumley, M.A., 1973. Anteneandertaliens et Neandertaliensdu basin mediterraneen occidental europeen. Universite deProvence, Marseille.
Martınez Navarro, B., Turq, A., Agustı Ballester, J., Oms, O.,1997. Fuente Nueva-3 (Orce, Granada, Spain) and thefirst human occupation in Europe. J. Hum. Evol. 33,611–620.
Maas, M.C., 1991. Enamel structure and microwear: an experi-mental study of the response of enamel to shearing force.Am. J. Phys. Anthropol. 85, 31–50.
Mercier, N., Valladas, H., 1994. Thermoluminescence dates forthe Paleolithic Levant, in: Bar Yosef, O., Kra, R.S. (Eds.),Late Quaternary Chronology and Paleoclimates of EasternMediterranean. American School of Prehistoric Research,Cambridge, pp. 13–20.
Mulvaney, D.J., Golson, J. (Eds.), 1971. Aboriginal man andenvironment in Australia. Australian National UniversityPress, Canberra.
Perez-Perez, A., 1990. Evolucion de la dieta en Cataluna yBaleares desde el Paleolıtico hasta la Edad Media a partir derestos esqueleticos. PhD Dissertation Series, University ofBarcelona, Barcelona.
Perez-Perez, A., Lalueza, C., Turbon, D., 1994. Intraindividualand intragoup variability of of buccal tooth striationpattern. Am. J. Phys. Anthropol. 94, 175–187.
Perez-Perez, A., Bermudez de Castro, J.M., Arsuaga, J.L.,1999. Nonocclusal dental microwear analysis of 300,000-year-old Homo heidelbergensis teeth from Sima de losHuesos (Sierra de Atapuerca, Spain). Am. J. Phys.Anthropol. 108, 433–457.
Puech, P.-F., 1984. Acidic food choice in Homo habilis atOlduvai. Curr. Anthropol. 25, 349–350.
Richards, M.P., Petitt, P.B., Trinkaus, E., Smith, F.H.,Paunovic, M., Karavani, I., 2000. Neandertal diet at Vindijaand Neandertal predation: the evidence from stableisotopes. Proc. Natl. Acad. Sci. U.S.A. 97, 7663–7666.
Richards, M.P., Pettit, P.B., Stiner, M.C., Trinkaus, E., 2001.Stable isotope evidence for increasing dietary breadth in theEuropean mid-Upper Paleolithic. Proc. Nat. Acad. Sci. 98,6528–6532.
Schoeninger, M.J., 1995. Stable isotope studies in humanevolution. Evol. Anthropol. 4, 83–87.
Schoeninger, M.J., DeNiro, M.J., Tauber, H., 1983. Stablenitrogen isotope ratios of bone collagen reflect marine andterrestrial components of prehistoric human diet. Science220, 1381–1383.
Schwarcz, H.P., Dupras, T.L., Fairgrieve, S.I., 1999. 15Nenrichment in the Sahara: in search of global relationship.J. Archaeol. Sci. 26, 629–636.
Sponheimer, M., Lee-Thorp, J.A., 1999. Isotopic evidence forthe diet of an early hominid, Australopithecus africanus.Science 283, 368–370.
Stringer, C., Davies, W., 2001. Those elusive Neandertals.Nature 413, 791–792.
Teaford, M.F., 1988. Scanning electron microscopy diagnosisof wear pattern versus artefacts. Scanning Microscopy 2,1167–1175.
Teaford, M.F., 1991. Dental microwear: what can it tell usabout diet and dental function, in: Kelley, M.A., Larsen,C.S. (Eds.), Advances in Dental Anthropology. Alan R.Liss, New York, pp. 341–356.
Teaford, M.F., 1994. Dental microwear and dental function.Evol. Anthropol. 3, 17–30.
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513512

Teaford, M.F., Lytle, J.D., 1996. Brief communication: diet-induced changes in rates of human tooth microwear: a casestudy involving stone-ground maize. Am. J. Phys.Anthropol. 100, 143–147.
Teaford, M.F., Runestad, J.A., 1992. Dental microwear and diet inVenezuelan Primates. Am. J. Phys. Anthropol. 88, 347–364.
Tillier, A.M., 1982. Les enfants neandertaliens de Devil’sTower (Gibraltar). Z. Morpho. Anthropol. 73, 125–148.
Ungar, P.S., 1992. Feeding behavior and dental microwear inSumatran anthropoids. Am. J. Phys. Anthropol. Sup. 14, 165.
Ungar, P.S., 1994a. Incisor microwear in Sumatran anthropoidPrimates. Am. J. Phys. Anthropol. 94, 339–363.
Ungar, P.S., 1994b. Patterns of ingestive behavior and anteriortooth use differences in sympatric anthropoid Primates.Am. J. Phys. Anthropol. 95, 197–219.
Ungar, P.S., Teaford, M.F., 1996. Preliminary examination ofnon-occlusal dental microwear in Anthropoids: implicationsfor the study of fossil Primates. Am. J. Phys. Anthropol.100, 101–113.
Villa, G., Giacobini, G., 1995. Subvertical grooves of inter-proximal facets in Neandertal posterior teeth. Am. J. Phys.Anthropol. 96, 51–62.
Walker, A.C., 1984. Mechanisms of honing in the male babooncanine. Am. J. Phys. Anthropol. 65, 47–60.
Walker, P.L., DeNiro, M.J., 1986. Stable Nitrogen and Carbonratios in bone collagen as indices of prehistory dietarydependence on marine and terrestrial resourcesin SouthernCalifornia. Am. J. Phys. Anthropol. 71, 51–61.
Wolpoff, M.H., 1999. Paleoanthropology. McGraw-Hill,Boston.
A. Perez-Perez et al. / Journal of Human Evolution 44 (2003) 497–513 513
Copyright © 2022 FDOKUMEN




















