
A.F.R. Wollaston and the 'Utakwa River Mountain Papuan' skulls

H i , > r o r ~ ( l r i Biolugj. 1994. Vol. 8, pp. 209-234 Repr~nts availablc directly from thc publisher Photocopying ,?v;l~lahle by license unly
O 1994 Harwood Academlc Puhl~shers GmbH Pr~nted In Malays~a
NOTES ON SKULLS OF PLEISTOCENE SAIGA OF NORTHERN EURASIA
G. BARYSHNIKOV and A. TIKHONOV
Zoological Institute of Russiun Acudenl. of Scieni-es, Univel-sitetskaya emh.1. St Petershurg 199034. Russia
Eighteen fossil skulls of male Saiga from Northern Eurasia and 33 recent skulls from Kalmykia and Kazakhstan have been studied. Saiga from both the Khazarian Fauna of the Volga and Mammoth Fauna of Europe and Siberia are referred to Saiga horealis Tschersky, 1876. During the Pleistocene, S. horealis distribution extended from England in the west to Alaska in the east and is characterized by an elongated neurocranium, small frontal angle of the temporal bone from the plane of the frontal. and long nasal bones.
S. horealis was a typical representative of the "mammoth biome" in the Pleistocene periglacial steppes and cryogenic savannahs. Two subspecies are recognized: S. horealis horealis Tschersky (Eastern Sibera and Alaska); and S.b. prisca Nehring, 1891 (Europe, Urals and Western Siberia). At the end of the Pleistocene. when the mammoth disappeared, the range of S. borealis was reduced. Today they live only in West Mongolia (S. borealis i7zongolica Bannikov, 1946). S. tataricu rururic.u was widely distributed in the other territories of the steppe and semidesert zones of Eurasia. The arid landscapes of Transcaucasia and Kazakhstan were inhabited by Saiga with thinner legs and shorter nasal bones, such as S. tarat.ic.a binagodensis Alekperova, 1953, from the middle Pleistocene of Azerbaijan (Bynagady). Fossil skulls from the Ural River that are large, but with a short neurocranium are identified as Saiga sp. cf. S. rararica Linnaeus, 1766.
KEY WORDS: Saiga, Pleistocene, Eurasia, Suiga bot.eulis, morphometric characters.
I INTRODUCTION
Recent Saiga - Saiga tatarica Linnaeus, 1766 - inhabit steppes of the Lower Volga, Kazakhstan and Mongolia. Herds of this species migrate long distances and individuals are adapted to running in rugged country but not in deep snow cover. These biological features of the Saiga make it a good indicator of steppe conditions. Therefore, Quaternary remains of Saiga beyond the recent geographic range of the genus throughout Northern Eurasia from England in the west up to Chukotka in the east, to Alaska and the Northwest Territory, Canada, are of interest to researchers.
In North America, the Pleistocene Saiga has been described several times under different names: Saiga horealis Tschersky, 1876 (Yakutia); S. prisca Nehring, 1891 (Moravia); S. ricsei Frick, 1937 (Alaska); and S. binagadetzsis Alekperova, 1953 (Transcaucasus). Specimens refered to these taxa differ from the recent form in the structure of skull and dentition. The relationships among these extinct groups and their geographic distributions during the Pleistocene remain to be clarified. To elucidate these questions we have studied the available fossil and recent skulls from scientific collections is Leningrad, Moscow and Krakow.

210 G. BARYSHNIKOV AND A. TIKHONOV
MATERIAL AND METHODS
We studied collections of twelve adult male fossil Saiga reposited in the Zoologic Institute, Academy of Sciences, Leningrad, Russia (ZIAS: Volga (I), Ural River (2 Western Siberia (3), Middle and Eastern Siberia (5); others unknown). Two specimer were also examined from the Paleontological Institute, Academy of Sciences in Mosco\ Russia (Volga and Vilui River), and one from the Institute of Systematics ar Experimental Zoology, Polish Academy of Sciences, Krakow (Slupyanca Cave). A ca of a Saiga skull with homcores from Belgium (Leopold Canal) was also available fro] the Institut Royal des Sciences Naturelles de Belgique (N 16-1021). We also used dal on teeth and the postcranial skeleton from Palaeolithic sites of the Crimea, Urals, an from asphalt sands of the Apsheron Peninsula (collection of ZIAS). Modem comparativ material included 33 recent skulls of adult males from Kalmykia and Kazakhsta reposited in the ZIAS and the Zoological Museum, Moscow University. We also use data on fossil Saiga from Czersky (1876), Alekperova (1955), Vereshchagin (1959 Sher (1967), Alekseeva (1980), Harrington (198 l) , Lazarev and Tomskaya (1 987). an Kahlke (1990).
Since the fossil material is fragmentary and is represented by pieces of skulls wit horncores the measurements listed below were most useful (Figure 1). The T-test wa used to determine the significance of morphometric changes : p < 0.001 (+++); p < 0.0 (++); and p < 0.05 (+).
Upper neurocranium length: akrokranion - supraorbitale. Length behind horns: akrokranion - the hind surface of horncore bases. Cranial length: akrokranion - bregma. Diameter of the horncore base. Greatest breadth across the orbits: ectorbitale - ectorbitale. Width between foramen supraorbitale. Width between horns. Latero-medial diameter of the horncore base. Least breadth of the parietal. Greatest mastoid breadth: otion - otion. Greatest breadth of the occipital condyles. Height of occiput: akrokranion - basion. Frontal angle (in degrees).
MORPHOMETRIC CHARACTERISTICS OF THE SKULL OF SAIGA BOREALIS
Czersky (1876, p. 150) described from the Vilui River, Yakutia, an extinct species 01 Antilope (Saiga) borealis and assumed that the structural features of its skull indicatec sharp differences in climate between the Pleistocene distribution and the recent., The holotype of this species has been lost. To this new species, Czersky included the separate horncore from the Nizneudinskava Cave, Sayani (Czersky, 1876, p. 149). Later. Frick (1937, p. 546) described Saiga ricei from Alaska (F:AM 30495 from Lilian Creek, in the region of Fairbanks). Examining the fossil material from Eurasia, Vereshchagin (1959, p. 444) noted morphological differences between the East Siberian Saiga and the recent species, in particular the somewhat elongated neurocranium of the skull, the weak spread of the horns, and straightness of the horncores.
Sher (1967), who studied serial fossil material, concluded that Siberian and Alaskan Saiga were very similar and placed them in the same species - S. ricei. He thought

PLEISTOCENE SAIGA IN EURASIA
h 11 - e 10 w
.- 7 5 - 5 *
-6 a
Figure 1 Diagram of Saiga's skull measurements. Designation of measurements in text.

212 G. BARYSHNIKOV AND A. TIKHONOV
that S . ricei differed from the holotype of S. borealis by a smaller orbital width (128 mm and 133.8 mm, respectively). However, this character however is variable, since the margins of the orbits frequently are broken. Orbital width is thus not diagnostic for the specimens we studied and does not separate S . horculis from S. tutat-icu. Proceeding from the assumption that all skulls of the Pleistocene Saiga from the basin of the Lena River (Vilui River included) are similar, we can retain the name S. hot.ealis for this form as the species having priority (Kahlke, 1975 p. 138) based on the following information.
Studying the fossil Saiga from Siberia, Sher (1967: 106) noted the following characteristic features: skull relatively narrow across orbits, homcores strongly bent backwards, and neurocranium elongated. The latter two characters confirm the differences between S , borealis and S. tatarica. The minimal values characterizing the length of the hind part of skull in the extinct species (measurments 1-3) on specimens from Siberia exceeds the mean indices in the recent S. tatarica (Table I). These forms differ in the index of relative skull width at the base of the homcores in relation to neurocranial length, which is much smaller in S. borealis. The angle of the frontal bones from the plane of temporal in S. horeulis is also significantly smaller. As a rule this angle does not exceed 32 degrees. In S . tataricu it is 33 degrees on the average.
The following parameters should be added to the above ones: the occiput in the fossil Saiga is broader (in relation to its height) and the nasal bones are longer. The nasals are preserved completely on only one specimen (Minusinsk, ZIAS N 15097, Figure 2) and are 67.4 mm in length. In recent S . taturic*~, the nasals range from 50.8 mm to 63.2 mm (x = 57.6, n = 32), and in S. mongolicu 48.3 mm to 56.8 mm (x = 52. I, n = 3).
Figure 2 Saiga borealis, from Aidorach River near Minusinsk ZIAS N 15097. A - front view, B - side view.

PLEISTOCENE SAIGA IN EURASIA

Table 1 (cont.)
Yakutia Alaska
Measurements River Vilui River Olenek River Olenek The delta of River Kolyma Lilian Creek Gold Hill (mm) (Sher, 1967) ZIAS NO7076 ZIAS NO7077 Lena ZIAS (Sher, 1967) near Livengood near Fairbanks 9
No 13636 (Sher, 1967) (Harington, 198 1) m
1. 99.0 96.7 89.1 94.7 97 G
28.3 30.0 (25.2)* 29.6 113 def. 116 def.
* Harington, 1981

PLEISTOCENE SAIGA IN EURASIA 215
The strong development of the posterior part of the skull and a weak bend between the frontal and temporal planes can be regarded as archaic characters which were more typical of extinct Siberian Saiga when compared to the recent species (Sher, 1967).
An examination of the fossil materials .shows that Saiga from Eastern Siberia differ from Saiga of West Siberia by a relatively more elongated hind part: the index of hind length (character 7) to width between horns (character 1) is from 98.5 to 101.7 (x = 101.2, n = 6). This index is much lower in the sample from Western and Middle Siberia (range is 104.6 to 110.0, x = 107.2, n = 4). Fossil Saiga from Alaska resemble that from Eastern Siberia (ratio is 95.9 for Lilian Creek specimens). Saiga from Yakutia also possess a lower occiput. This data allows us to assume that there are differences between the West Siberian and East Siberian fossil Saiga at the subspecies level.
The second form, S. borealis borealis, looks slightly more archaic because it has stronger differences from recent S. tatarica. One skull from the Lena River (ZIAS N 13636, Figure 3) preserves all the main characters of S. borealis, but is smaller in size (neurocranial length = 89.1 mm). However, it is still larger than skulls of the smallest recent Mongolian Saiga S.b. mongolica Bannikov, 1946, in which the neurocranial length ranges from 83 mm to 85.6 mm (x = 84.4, n = 3). It is interesting that the frontal- parietal index of S. mongolica is very small (range is 84.7 to 90.9, x = 87.7, n = 3), but is closer to S. borealis than to S. tatarica. The fossil skull from the Lena River has fused sutures and belongs to an adult male of S. borealis, which became smaller at the end of the Pleistocene. This results from the condition of isolation, noted by Vereshchagin (1959) and Harrington (1981). Reduction of size at the end of the Pleistocene also characterizes horses, bison, mountain sheep, and probably mammoths.
L-! 1 ! I c m
Figure 3 Saiga borealis, from the delta of Lena River. ZIAS N 13636. A - front view, B - side view.

216 G. BARYSHNIKOV AND A. TIKHONOV
EUROPEAN SAIGA BOREALIS
Pleistocene specimens of Saiga found in Europe from the Urals in the east to England in the west and southwards up to Crimean Peninsula are usually refered to S. tataric.~. However, the taxonomic position of fossil Saiga from Europe should be defined more precisely.
Volga River. - Two skulls of Saiga from the Volga River (Middle Pleistocene, Khazarian theriocomplex, Figure 4A) are of rather large size, with a long neurocranium (Table 2), a large frontal-parietal length index (103.0; 98.9), and also an angle of dip of the frontal plane in relation to that of the parietal bone that correspond to indices of S. borealis (Figure 6A). The Volga form is characterized by straight, relatively short and weakly spread, horncores (Vereshchagin, 1959). Perhaps this is a peculiar subspecies of S. borealis.
Bynagady, Baku. - Saiga binagadensis was described from the asphalt deposits of Bynagady near Baku (Alekperova, 1953, p. 70) which is of late Middle Pleistocene (Riss/Moscow) or last interglacial (Riss-Wurm/Mikulino) age. Vereshchagin (1959, p. 442) assumed that S. binagadensis Alekperova, 1953, from the eastern Transcaucasia is closely related to the Khazarian Saiga. This species is characterized by little spread between horns, but taller horncores than the Khazarian specimens with a more marked bend backwards in the sagittal plane. Thy Bynagady male skull measures 129 mm in width between the orbits, 101 mm between the bases of horncores on the outside, and 51 mm between the supraorbital apertures. The nasal bones are short and narrow, with a length of 48 mm (Alekperova, 1955, p. 20). Lower jaws and limb bones of S. binagadensis (ZIAS N 24406) are much smaller than in S. borealis.
In skull proportions the Bynagady Saiga is close to S. taturica, and is refered here to the subspecies, S. tatarica binagadensis. This systematic view suggests that the divergence of the phylogenetic lines of S. tatarica (south steppe and semideserts) and S. borealis (north steppe) began in the Middle Pleistocene.
Figure 3 The skulls of saiga, front view. A - Sni,ycr borealis ssp., Luchka near Saratov, Volga River, Khosar fauna. ZIAS N 1084. B -- Saigu cf. lalarica, Ural River. ZIAS N 24201-10.

PLElSTOCENE SAlGA 1N EURASIA 2 17
Table 2 The size of fossil saiga skulls from Europe
Measurements S . borealis S. cf. borealis S . c:f. tu/uric,c~
(nun) Middle pleistocene Late pleistocene The end of pleistocene
River Volga Belgium Poland River Ural
Luchka Tungus cast Slupianca ZIAS Zl AS ZIAS ZIAS"' PIASb' ZIAS NO5 IASE" N024201-10 N02420lL8 N024201-29 NO1084 N0131-277 2.206138
a) - ZIAS - Zoological Institute Academy of Sciences b) - PIAS - Palaeontological Institute Academy of Sciences C) - IASE - Institute of Animal Systematics and Evolution, Krakow, Poland
Figure 6 The skulls of saiga, side view. A - Saiga borealis ssp., Luchka near Saratov, Volga River. B - S a i ~ a cf. tcrtcrric,cr, Ural River. '

218 G. BARYSHNIKOV AND A. TIKHONOV
Moravia, Czechoslovakia. - Saiga prisca was described by A. Nehring (1 89 1, p. 13 1) from the Late Pleistocene deposits of Sipka-Hohle in Moravia, Czechoslovakia. The primary diagnostic difference of this species is the presence of an alveolus in front of PI3 on the jaw of the adult individual. As is known, adults of recent Saiga sometimes preserve the dPl2. The size of the alveolus in recent Saiga fully coincides with the size of the alveolus in front of PI3 in Moravian specimen. It is clear that this tooth anomaly cannot be used as a taxonomic character.
Among 109 adult skulls of S. tatarica, dP/2 is present in front of PI3 on eight specimens and in three others a single rooted needle-like tooth (reduced dP2?) is present. The one skull of S. mongolica has a PI2 (Baryshnikov et al., 1990, p. I 1). Length of the tooth row in the holotype of S. prisca equals that of Saiga from Pleistocene deposits of the Crimea, Urals and southern regions of Western Siberia. All are larger in the length of the tooth row than in S. tatarica. The length of PI3 to MI3 (when MI3 is strongly worn) is 74 mm in S. prisca. This length is 71.2 mm in the specimen from grotto Kiik-Koba in Crimea, 75.7 mm in the specimen from Stolbovoi Grotto in the Urals, and 75 mm in the material from Krasnyi Yar on the Ob River. In the recent species the length of Pl3-MI3 is less than 71 mm. Thus the form from the Pleistocene of Moravia resembles the northern Saiga of Eastern Europe and Siberia in length of the P3-M3. The name S. prisca in our opinion should be synonymized with S. borealis and reduced to subspecific status. In which case the specimen previously called S. prisca should be considered the subspecies S. borealis prisca from Western Eurasia up to east to the Yenisei River.
Belgium. - A study of a cast of fossil Saiga from Belgium (ZIAS 5, Leopold Canal) shows that the neurocranium length and the frontal-parietal angle more closely resemble S. borealis than S. tatarica (Table 2). The nasal bones are partly broken (the remaining part is 49.5 mm) but their total length was apparently 65 mm, and as such the were somewhat longer than in the recent S. tatarica. One can assume that S. borealis is a typical representative of the Mammoth Fauna, living not only in the Late Pleistocene of Siberia, but also in coeval northern Eurasia west of the Atlantic coast. This assump- tion appears to be confirmed by findings of the large form of Saiga in Germany (Pahren), France (Grotte de Fees), and England (Pratt, 1977; Currant, 1986; Kahlke, 1 990).
Poland. - One skull from Poland was found in the Slupyanca Cave, Ojcew near .
Krakow without accurate geological age. The posterior part is preserved completely as are the frontal bones. Both horncores are present, but the upper third of one is missing. The nasal and maxillary bones are broken. The frontal-parietal angle is 26 degrees. This is similar to minimal value of S. tatarica and is common to S. borealis. Neurocranial length and width between the outer margins of the horncore bases (measurments I and 7 ) are near the upper limit of corresponding values for S. tatarica but well within the range of S. borealis. In the scatter diagram plotting absolute values of neurocranial length against width between horncores (Figure 5), the skull from Slupyanca Cave is intermediate between S. borealis and S. tatarica but is closer to the values for S. tatarica. Therefore we consider this animal as Saiga sp., cf. S . borealis.
A second fossil Saiga from Maszyce cave, Ojcew near Krakow, of Magdalenian age is related to the one from Slupyanca Cave (Kiernik, 1912). The measurements of this specimen are 129 mm width between orbits, 70.5 mm with of neurocranium behind the horncores, 45.5 mm width between condyles. The length of PI3 to MI3 in the Maszyce Cave jaw is 74 mm.
Ural River. - Specimens collected from the sand-banks of the Ural River, may be dated at the end of the Pleistocene by the mineralization of bones (Figure 4B). The sizes of the skulls are within the range for recent S. tatarica. The posterior part is

PLEISTOCENE SAIGA IN EURASIA
Figure 5 Correlation between upper neurocranial length (A) and breadth between horncores (B) in the skulls of fossil and recent saiga. 1 - S. borealis, 2 - S. cf. borealis, Slupianca, 3 - S. cf. rararica, Ural River, 4 - S. rararica, recent.
relatively weakly elongated (index of neurocranial length is 109.5 to 112.1), and the frontal angle is large at 34 degrees (Figure 6B). Ratios of neurocranial length and the interorbital width for the Ural River specimens are more similar to S. tatarica than S. borealis (Figure 5). We think that this form is closely related to S. tatarica and could be defined as Saiga sp., cf. S. tatarica. This finding shows that Saiga of recent type inhabited areas further south in Kazakhstan at the same time as S , borealis.
DISCUSSION
Taxonomy
Usually the Pleistocene Saiga is defined as S. tatarica (Vereshchagin, 1959; Hanington, 1981) or as a subspecies S. tatarica borealis (Bannikov, 1963, Kahlke, 1975, 1990, Baryshnikov, 1981). Our original researches of the material shows that S. borealis (including S. prisca, S. ricei) is an independent species. Thus, we confirm Sher's (1967) opinion of the distinct status of the northern Saiga.
Morphometric comparisons of the indices of the skulls of S. borealis (including the skull from Slupyanca Cave) and recent S. tatarica demonstrate a number of differences between these species with a high degree of significance (Table 3). Particularly significant
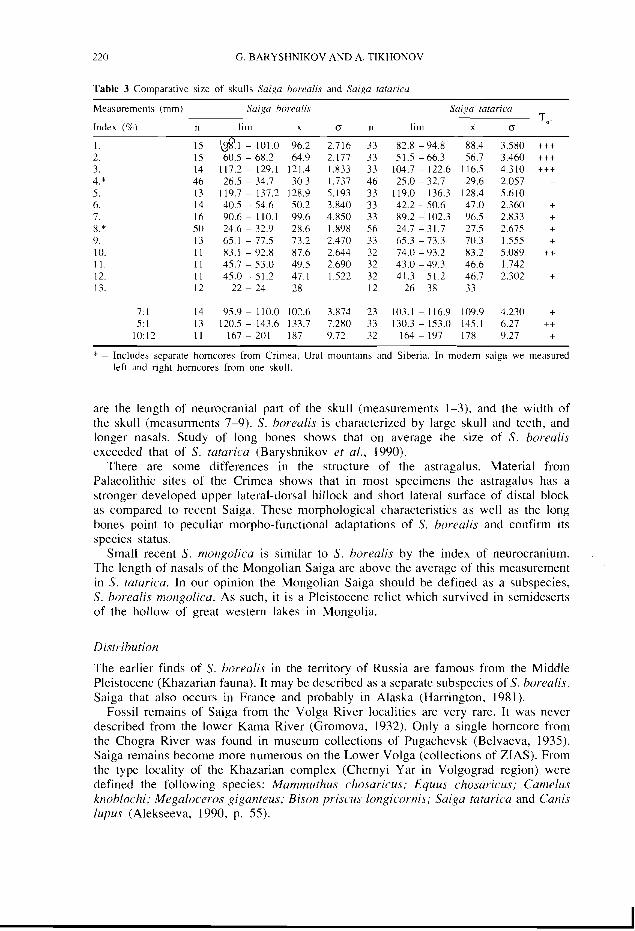
220 G. BARYSHNIKOV AND A. TIKHONOV
Table 3 Comparative size of skulls Saiga borealis and Saiga tatarica
Measurements (rnrn) Saiga borealis Saiga tatarica
Index (%) - -
n lim x (5 n lim x (5 T,,.
7: 1 14 95.9 - 110.0 10.7.6 3.874 23 103.1 - 116.9 109.9 4.230 + 5: 1 13 120.5 - 143.6 133.7 7.280 33 130.3 - 153.0 145.1 6.27 ++
10:12 1 1 1 6 7 2 0 1 187 9.72 32 164-197 178 9.27 + * - Includes separate horncores from Crlrnea. Ural mountains and Siberia. In modem saiga we measured
left and right horncores from one skull.
are the length of neurocranial part of the skull (measurements 1-3). and the width of the skull (measurments 7-9). S. borealis is characterized by large skull and teeth, and longer nasals. Study of long bones shows that on average ihe size of S. borealis exceeded that of S. tatarica (Baryshnikov et al., 1990).
There are some differences in the structure of the astragalus. Material from Palaeolithic sites of the Crimea shows that in most specimens the astragalus has a stronger developed upper lateral-dorsal hillock and short lateral surface of distal block as compared to recent Saiga. These morphological characteristics as well as the long bones point to peculiar morpho-functional adaptations of S. borealis and confirm its species status.
Small recent S. mongolica is similar to S. borealis by the index of neurocranium. -
The length of nasals of the Mongolian Saiga are above the average of this measurement in S. tuturic~u. In our opinion the Mongolian Saiga should be defined as a subspecies, S. borealis mongolica. As such, it is a Pleistocene relict which survived in semideserts of the hollow of great western lakes in Mongolia.
Distribution
The earlier finds of S. borealis in the territory of Russia are famous from the Middle Pleistocene (Khazarian fauna). It may be described as a separate subspecies of S . borealis. Saiga that also occurs in France and probably in Alaska (Harrington, 198 1 ).
Fossil remains of Saiga from the Volga River localities are very rare. It was never described from the lower Kama River (Gromova, 1932). Only a single horncore from the Chogra River was found in museum collections of Pugachevsk (Belvaeva, 1935). Saiga remains become more numerous on the Lower Volga (collections of ZIAS). From the type locality of the Khazarian complex (Chernyi Yar in Volgograd region) were defined the following species: Mummuthus chosuricus; Equus chosaricus; Camelus knoblochi; Meguloceros giganteus; Bison priscus 1ongicor.nis; Suiga tatarica and Canis lupus (Alekseeva, 1990, p. 55).

PLEISTOCENE SAIGA IN EURASIA 22 1
In the late Pleistocene (Wurm~Waldayi) the fossil remains of Saiga become more numerous. It inhabited arctic and boreal zones from the British Isles on the west up to northwestern Canada on the east (Figure 7). We can separate two subspecies from this vast area: S. borealis prisca Nehring (Europe, Ural, West Siberia up to the Yeniseyi River) and S. borealis borealis Tchersky (East Siberia, Alaska).
The remains of Saiga from the Pleistocene of West Europe and Alaska are small in numbers and as a rule without exact geological age, so it is not easy to retrace the dyliamics of range and density of Saiga in the Late Pleistocene in the territory of Russia. However, Saiga lived in Western Europe to the upper Magdalenian (Harrington, 1981; Currant, 1986).
In early Wurm (Waldayi) the remains of Saiga in Russia are noted mainly from southern regions. Particularly, they were numerous in the Crimea and dominated Mousterian sites. The accompanying fauna in the Crimea includes the following species: Mammuthus primigenius; Coelodonta antiquitalis; Equus caballus; Equus hydruntinus; Cerllus elaphus; Megaloceros giganteus; Rangifer tarandus; Bison priscus; Canis lupus; Vulpes lulpes; Vulpes corsac; Alopex lagopus; Ursus spelaeus crimaeus; Crocuta spelaea; and Panthera spelaea. Saiga also inhabitated the north, penetrating Eastern Siberia. The skull of Saiga from the Balyktiach River was refered to deposits of the Karginskyi level (Lazarev Tomskaya, 1987, pp. 1 14-129). Harrington (198 1) supposed that dispersal of Saiga was in waves, beginning during the Middle Pleistocene (Illinoian). The radiocarbon date of a horncore from Usuktuk River, 37,000 f 990 BP (GSC-3050), shows that Saiga penetrated to northern Alaska in the Middle Wisconsin (Harrington, 1981, p. 225).
In the late Wurm (Waldayi) the remains of Saiga are known from the Crimea, Russian Plane, Ural, Altayi, and East Siberia. They are small in number in most areas, but are very numerous in the Crimea. It is remarkable that the fossil remains of Saiga are never found in the Baikal region (Ermolova, 1987; Ovodov, 1987). The report of Saiga remains from the Upper Palaeolithic site of Suchotino near Chita are erroneous (Kasparov, 1988, p. 99).
The northern Saiga S. borealis in the Late Wiirm is an index element of the Mammoth Fauna. Its remains have been found together with Mammuthus primigenius; Coelodonta antiquitatis; Equus latipes; Equus lenensis; Alces alces; Rangifer tarandus;
. Bison priscus; Ovibos moschatus; Canis lupus; Gulo gulo; Crocuta spelaea and Panthera spelaea.
Further south in the Transcaucasus, Kazakhstan and Middle Asia, S. tatarica apparently existed in arid steppe and semidesert areas. S. t. binagadensis had small metacarpal and metatarsal bones that are relatively narrow in the distal part. Probably the Apscheron Saiga inhabited hot dry steppes. Other animals thought to indicate warm climate are found together with Saiga in the Bynagady site, such as Equus caballus; Equus hydruntinus; Rhinoceros binagadensis; Sus apscheronicus; Cervus elaphus; Megaceros euryceros; Bos mastanzadei; Canis lupus; Vulpes corsac; Vulpes vulpes; Ursus arctos; Vormela peregusna; Meles meles; Panthera spelaea and Acinonvx jubatus (Vereshchagin, 1959, p. 141).
Pleistocene Saiga are rare in Kazakhstan. Remains of Saiga sp., cf., S. tatarica are found on beaches of the Ural River together with bones of Mammuthus primigenius, Rhinoceros tichorhinus, Equus caballus, Rangifer tarandus, Alces alces, Cervus elapphus, and Bos primigenius (Vereshchagin, Gromov, 1952). Such a grouping of large animals is characteristic of the southern zone of the Mammoth Fauna. A transition to the fauna of the Kazakhstan steppes is observed in the composition of the rodent fauna: Citellus pygmaeus; Allactaga jaculus; Cricetus cricetus; Meriones tarnariscinus; Lagurus lufeus; Lagurus lagurus and Ellobius talpinus. Here in the region of the Ural

Figure 7 The Pleistocene finds of Saiga. Triangles - fossil skulls : A - Leopold Canal (Belgium), B - Slupianca Cave (Poland), C - Tungus (Volga River near delta of Cheremshan River), D - Luchka (near Saratov), E - Ural River, F - Tura River, G - Uschaika River (near Tomsk) H - Kuzbass, I - Aidorach River, (near Minusinsk), J - Yenisei River (near Krasnoyarsk). K - Vilui River, L - Olenek River, M - Delta of Lena River, N - Kolyma River, 0 - Lillian Creek (near Livengood), P - Gold Hill (near Fairbanks). Circles - Localities of the Pleistocene remains of saiga (numbers near circles are similar to number of the localities from appendix).

PLEISTOCENE SAIGA IN EURASIA 223
steppes, S. borealis and S. tatarica could have been sympatric. The remains of Saiga are found in the upper palaeolithic site of Ushbas Cave in southern Kazakhstan (Dzhambul region) together with Rhinoceros sp, Bison sp, (Alpysbaev, 1962). There are no fossil remains of Saiga in Middle Asia. The definition of Saiga from Samarkandskaya (upper Palaeolithic) are erroneous (Dzhurakulov et al., 1980).
Paleoecology
It is commonly held that species represented by fossils have similar habitat require- ments to those of their closest recent species. S. tatarica today lives in dry semideserts with low sparse vegetation and thin snow cover. It forms large herds which migrate widely. The main food includes the following: Kochia prostata; Artemisia; Ephedra; and Anabasia (Bannikov, 1963: Zhirnov, 1982).
S. mongolica inhabits lowlands of the great western lakes of Mongolia in arid conditions. Small herds of these animals do not migrate long distances. It feeds on: Stipa gobica, Allium, and Anabasia. Artemisia is not a common food of the mongolian saiga.
S. borealis inhabitated vast grassy planes together with other herbivorous animals - Mammuthus, Coelodonta, Equus, and Bison. In addition it ranged with Rangifel- and Ovibos on the north. The above species of proboscideans and ungulates are thought to have lived in large herds in grasslands of the periglacial steppes, tundra-steppes and arctic cryogenic savannah. Winter is assumed to have been very cold with little snow, and the summers were probably short and dry because morphology of northern Saiga suggests they were best adapted to open country that lacked deep snow.
We can suggest that the morphological similarity between S. borealis and S. mongolica is closely related to their ecological similarity. Investigation of fossil remains of Saiga from the Palaeolithic of the Crimea shows that crimean Saiga inhabitated there all year around and bred (Baryshnikov et al., 1990). Probably their food included cereals and not Kochia and Artemisia, as in S. tatarica. Confirmation of this lies in the proportions of lower molars MI and M3 of Saiga from the Palaeolithic of the Crimea (Baryshnikov et al., 1990).
Thus the wise distribution of Pleistocene Saiga in Eurasia, can be explained by . corresponding distribution in the Pleistocene tundra-steppe or cryogenic savannah, in
areas which now are tundra and taiga zonal landscapes. Along with the mammoth, woolly rhinoceros, reindeer and musk-ox, the northern S. borealis was a natural part of ecological structure of the mammoth biome.
General Appearance
S. borealis was a rather large antelope. Adult males had a height in shoulders 140-150 cm and weight near 65-70 kg. They had a massive head with a large horns. The skull was characterized by elongated posterior part and nasals. In comparison with S. tatarica, by the frontal-parietal angle, S. borealis had a small hump-nose profile. By this feature it resembles the recent S. mongolic-a. Firstly this was noted by Bannikov (1963, p. 282). Our suggestion about small "hump-nose" profile of the Pleistocene Saiga are confirmed by the Palaeolithic painting of the Saiga's head from French caves (Bequen et al. 1986, Figure 5)

PLEISTOCENE SAIGA IN EURASIA 223
steppes, S. borealis and S. tatarica could have been sympatric. The remains of Saiga are found in the upper palaeolithic site of Ushbas Cave in southern Kazakhstan (Dzhambul region) together with Rhinoceros sp, Bison sp, (Alpysbaev, 1962). There are no fossil remains of Saiga in Middle Asia. The definition of Saiga from Samarkandskaya (upper Palaeolithic) are erroneous (Dzhurakulov et al., 1980).
Paleoecology
It is commonly held that species represented by fossils have similar habitat require- ments to those of their closest recent species. S. tatarica today lives in dry semideserts with low sparse vegetation and thin snow cover. It forms large herds which migrate widely. The main food includes the following: Kochia prostata; Artemisia; Ephedr-a; and Anabasia (Bannikov, 1963: Zhirnov, 1982).
S . mongolica inhabits lowlands of the great western lakes of Mongolia in arid conditions. Small herds of these animals do not migrate long distances. It feeds on: Stipa gobica, Allium, and Anabasia. Arteniisia is not a common food of the mongolian saiga.
S. horealis inhabitated vast grassy planes together with other herbivorous animals - Mammuthus, Coelodonta, Equus, and Bison. In addition it ranged with Rangifer and Ovibos on the north. The above species of proboscideans and ungulates are thought to have lived in large herds in grasslands of the periglacial steppes, tundra-steppes and arctic cryogenic savannah. Winter is assumed to have been very cold with little snow, and the summers were probably short and dry because morphology of northern Saiga suggests they were best adapted to open country that lacked deep snow.
We can suggest that the morphological similarity between S. horealis and S . mongolica is closely related to their ecological similarity. Investigation of fossil remains of Saiga from the Palaeolithic of the Crimea shows that crimean Saiga inhabitated there all year around and bred (Baryshnikov et al., 1990). Probably their food included cereals and not Kochia and Artemisia, as in S. tatarica. Confirmation of this lies in the proportions of lower molars M1 and M3 of Saiga from the Palaeolithic of the Crimea (Baryshnikov et al., 1990).
Thus the wise distribution of Pleistocene Saiga in Eurasia, can be explained by - corresponding distribution in the Pleistocene tundra-steppe or cryogenic savannah, in
areas which now are tundra and taiga zonal landscapes. Along with the mammoth, woolly rhinoceros, reindeer and musk-ox, the northern S. horealis was a natural part of ecological structure of the mammoth biome.
General Appearance
S. borealis was a rather large antelope. Adult males had a height in shoulders 140-150 cm and weight near 65-70 kg. They had a massive head with a large horns. The skull was characterized by elongated posterior part and nasals. In comparison wi,th S. tatarica, by the frontal-parietal angle, S. horealis had a small hump-nose profile. By this feature it resembles the recent S. mongolica. Firstly this was noted by Bannikov (1963, p. 282). Our suggestion about small "hump-nose" profile of the Pleistocene Saiga are confirmed by the Palaeolithic painting of the Saiga's head from French caves (Bequen et al. 1986, Figure 5)

G. BARYSHNIKOV AND A. TIKHONOV
Acknowledgements
W e are especially grateful to Dr. A. Nadachowski (Institute of Animal Systomatics and Evolution, Krakow, Poland) for permission to investigate the fossil skull of Saiga from Poland, and to Dr. P. Sartonaer (Institut Royal des Sciences Naturelles d e Belgique, Bruxelles) for information about Saiga material from Belgium. The help of A.K. Kasparov and O.R. Potapova (Zoological Institute of Russian Academy of Sciences, Leningrad) is gratefully acknowledged.
The paper's improvement was kindly made by the editing of Drs. A. Barnosky (University of California, Berkely, USA), and E. Anderson, T . Hardy, and R. Stucky (Denver Museum of Natural History, USA). W e are also thankful to Carol Carpenter for typing the edited manuscript.
Drawings were made by G.I. Baranova, and photos by G.A. Apanovich.
References
Abramova, Z.V. (1979) Palaeolithic of Yenisei River: Kokorevskaya Culture, Novosibirsk, Nauka, p. 199 (in Russian)
Alekperova, N.A. (1955) The fossil Saiga from Bynagady, Trudy Eststvenno-istoricheskogo museya AN Azerb, SSR, Baku, 10, 1&64 (in Russian)
Alexeeva, L.I. (1990) The theriofauna of Upper Pleistocene of Eastern Europe (large mammals). Trudy Geologicheskogo Instituta AN SSSR, Moscow, 455, 109 p. (in Russian)
Alpysbaev, Kh. A. (1962) The finds of stone age sites in the Karatau Ridge. Trudy Instituta istorii, archeologii i ethnografii AN Kaz SSR, Alma-Ata, 14, 12-37 (in Russian)
Bader, O.N. and Bader, N.O. (1979) Volchii grot, some results of its studying In: Kolossov Yu.G .... ed. Issledovanie paleolita v Krymu (1879-1979). Kiev. Naukova Dumka, 15-33 (in Russian)
Bannikov, A.G. (1954) Mammals of Mongolian People's Republic. Trudy Mongolskoi komissii, Moscow, 53, 669 p. (in Russian)
Bannikov, A.G. (1963) Die Saiga-Antelope (Saiga tatarica). Wittenberg, A. Ziemsen Verlag. 143p Baryshnikov, G.F. (1981) Order Artiodactyla Owen. 1848. In: Gromov, I.M. and Baranova,
G.I., ed. Katalog mlekopitaiuschich SSSR, pliocen-sovremennost. Leningrad, Nauka, 343408 (in Russian)
Baryshnikov, G.F., Kasparov, A.K. and Tikhonov, A.N. (1990) Saiga paleolita Kryma. Trudy Zoologicheskogo Instituta AN SSSR. Leningrad. 212. 3 4 8 . (in Russian)
BCgouen, R., Clottes, J., Giraud, J-P. and Rouzard, F. (1986) Le propulseur au saiga d' Enlene. Bulletin de la SociCte Przhistorique. Arikge-Pyrenkes. 41, 11-22
Bibikova, V.I. and Starkin, A.V. (1989) Description of the osteological matherial from the Late Palaeolithic site Anetovka 2. In: Stanko, V.N., Grigorieva, G.V., Shvaiko, T.N., Pozdnepaleoliticheskoe poselenie Anetovka 2. Prilozhenie., Kiev. Naukova Dumka, 127-131 (in Russian)
Chersky, I.D. (1876) Antilope (Saiga) borealis n. spec. fossilis. Izvestia Sibirskogo otdela Imperatorskogo Geographicheskogo Obschestwa. St. Petersburg, 7, 4-5, 145-151 (in Russian)
Currant, A.P. (1986) The Late glacial mammal fauna of Gough's Cave. Cheddar Somerset. Proceedings of University of Bristol Speleological Society, Bristol, 17, 3, 286-304
David, A.I. (1980) The theriofauna of Pleistocene of Moldavia. Kishinev, Shtilintsa, 186 p. (in Russian)
Dzurakulov, M.D., Kholushkin, Yu. P., Kholushkina, V.A. and Batyrov, B.H. (1980) Samarkandskaya site with comment of its position within the Late Palaeolithic of Middle Asia. In: Larichev, V.E., ed.. Paleolit Srednei i Vostochnoi Asii. Novosibirsk. Nauka, 51-95 (in Russian)
Ermolova, N.M. (1978) The theriofauna of Angara Valley in Late Anthropogene. Novosibirsk. Nauka, 223 p. (in Russian)

PLEISTOCENE SAIGA IN EURASIA 225
Frick, Ch. (1937) Homed ruminants of North America. Bulletin of American Museum of Natural History, 69, 1-669
Harrington, C.R. (1981) Pleistocene saiga antilopes in North America and their paleo- environmental implications. In: Mahaney W.C., ed., Quaternary paleoclimate. Norwich. University of East Anglia. 193-225
Hoffecker, J.F., Baryshnikov, G.F. and Potapova, O.R. (1989) Mousterian bison hunters of the northern Caucasus: Analysis of faunal remains from Ilskaya 1. Current Research in the Pleistocene., 6, 69-72
Kahlke, H.D. (1975) Der Saiga-Fund von Bottrop/Westfalen. Quartar, 26, 135-146 Kahlke, R.D. (1990) Der Saiga-Fund von Pahren. Ein Beitrag zur Kenntniss der Palaarktischen
Verbreitungsgeschichte der Gattung Saiga Gray 1843 unter besonderer Beriicksichtigung des Gebeites der DDR. Eiszeitalter und Gegenwart. Hannover, 40, 2G37
Kasparov, A.K. (1986) Mammal remains from the Late Palaeolithic site Sukhotino 1 in Transbaikalia. Trudy Zoologicheskogo Instituta AN SSSR. 149, 98-106 (in Russian)
Kiernik, E. (1912) Materyaly do paleozoologii dyluwialnych ssakow Ziem Polskich. CzeSc' 3. Szczgtki suhaka (Antilope saiga) z jaskini Maszyckiei kolo Ojcowa. Rozprawy wydzialy matematychno-pryyrodnichego Akademii Umiejetnosci. Krakow, ser, 3, 12, 399434
Kolossov, Yu. G. (1983) The Mousterian sites in Belogorsk Region (the problems of the Crimean Late Palaeolithic periodization). Kiev. Naukova Dumka, 208 p. (in Russian)
Kolossov, Yu. G. (1986) Akkaiskaya Mousterian Culture. Kiev. Naukova Dumka, 224 p. (in Russian)
Kosintsev, P.A. and Borodin, A.V. (1990) The theriofauna of eastern slope of Northern Ural in the Late Pleistocene and Holocene. Trudy Zoologicheskogo Instituta AN SSSR, 212, 12G134 (in Russian)
Kuzmina, I.E. (1971) Forming of theriofauna of Northern Ural in late anthropogene. Trudy Zoologicheskogo Instituta AN SSSR, Leningrad, 49, 44-122 (in Russian)
Kuzmina, I.E. (1975) Some data on the Late Pleistocene mammals from the Middle Urals. Bulletin Komissii po izucheniu chetvertichnogo perioda, 43, 63-77 (in Russian)
Lazarev, P.A. and Tomskaya, A.I. (1987) Mammals and biostratigraphy of late cainozoic of Northern Yakutia, Yakutsk, Izdatelstwo Yakutskogo filiala SO AN SSSR, 172 p. (in Russian)
Nehring, A., (1891) Diluviale Reste von Cuon, Ovis, Saiga, Ibex und Rupicapra aus Mahren. Neues Jiihrbuch fur Mineralogie, Geologie und Palaontologie, Stuttgart, 2, 107-155
Ovodov, N.D. (1987) Fauna of the Palaeolithic sites Tolbaga and Vawarina Gora in Western Transbaikalia. In: Resanov I., Basarova L. Khamsina E. ed. Prirodnaya sreda i drevnii chelovek v pozdnem anthropogene. Ulan-Ude. Geologicheskii Institut, 122-140 (in Russian)
Petrin, V.T. and Smirnov N.G. (1975) The palaeolithic site at Shikaevka on the right bank of Tobol River. Voprosy archeologii Urala, 13, 75-85 (in Russian)
Pidoplichko, I.G. (1956) Matherials for studying of fossil faunas of the Ukraine, Vol. 2. Kiev, AN USSR, 236 p. (in Ukrainen).
Prat, F. (1966) Deuxieme partie. Chapter 9. Les antilopes, Atlas de prehistoire, Faunes at flores prehistoriques de 1'Europe occidentale, 3, 323-336, Paris
Sher, A.V. (1967) The fossil saiga on the North of Eastern Siberia and Alaska. Bulletin Komissii po izucheniu chetvertichnogo perioda, Moscow, 33, 97-1 12 (in Russian)
Sher, A.V. (1971) Mammals and stratigraphy of the Pleistocene of far north-east of the USSR and Northern America, Moscow, Nauka, 312 p (in Russian)
Vereshchagin, N.K. (1959) Mammals of Caucasus. History of forming of the fauna. Moscow- Leningrad, Izdatelstwo AN SSSR, 704 (in Russian)
Vereshchagin, N.K. and Baryshnikov, G.F. (1980) Mammals of northern Crimea foothills in the Palaeolithic (on the base of food remains from the caves Chokurcha, Staroselye and Mamat- Koba. Trudy Zoologicheskogo Instituta AN SSSR, Leningrad, 93, 2 6 4 9 (in Russian)
Vereshchagin, N.K. and Gromov, I.M. (1952) History of fauna of vertebrates from the region of lower part of River Ural. Trudy zoologicheskogo Instituta AN SSSR, Leningrad, 9, 1226-1 269 (in Russian)
Vereshchagin, N.K. and Kolbutov, A.D. (1957) Animal remains from the mousterian site at Stalingrad and stratigraphical position of the Paleolithic stratum. Trudy Zoologicheskogo Instituta AN SSSR, Leningrad, 22, 75-1 11 (in Russian)

226 G. BARYSHNIKOV AND A. TIKHONOV
Vereshchagin, N.K. and Kuzmina, I.E. (1977) Mammal remains from the Palaeolithic sites on Don and Upper Desna Rivers. Trudy Zoologicheskogo Instituta AN SSSR, Leningrad, 72, 77-1 10 (in Russian)
Zeitlin, S.M. (1979) Geology of the Paleolithic of the Northem Asia. Moscow, Nauka, 287 p. (in Russian)
Zhimov, L.V. (1982) Gone back to life. Ecology, preservation and utilisation of saigas, Moscow: Lesnaya promyshlennost, 224 p. (in Russian)
APPENDIX
Fossil remains of in the Pleistocene assamblages with Proboscidea. Perissodactyla and Artiodactyla in the former USSR1),*'
Middle Pleistocene 1. Bynagady, Baku, Transcaucasia Riss or Riss-Wurm (Moskow or Mikulino)
(Vereshchagin, 1959, p. 141)
Equus caballus ssp. Equus aff. hydruntinus Rhinoceros binagadensis Sus apscheronicus Cervus elaphus binagadensis Megaceros cf. euryceros Bos mastanzadei Saiga tatarica binagadensis Ovis cf. ammon
2. Ural River, Sandy banks Pleistocene (mixed material) (Vereshchagin, Gromov. p. 1244)
Elephas cf. primigenius Equus caballus Equus cf. hydruntinus Equus hemionus Rhinoceros cf. tichorinus Elasmotherium sibiricum Camelus knoblochi Cervus elaphus Megaceros euryceros Alces alces Rangifer tarandus Bos primigenius Bison priscus Saiga tatarica
'I According to Alpine and East-European stratigraphical scales '' Number of individuals in localities NNO1,10 in other cases the number of remains.

PLEISTOCENE SAIGA IN EURASIA
Upper Pleistocene, Lower Wiirm (Lower Waldai)
3. Buzduzhany 1 Grotto Rakovets River, Moldova Mousterien (David, 1980, p. 38)
Mammuthus primigenius Equus latipes Equus (Asinus) hydruntinus Coelodonta antiquitatis Cervus elaphus Megaceros euryceros Rangifer tarandus Cervinae Bison priscus Saiga tatarica
4. Iliinka Grotto Odessa, Ukraine Mousterien (Pidoplichko, 1956, p. 99)
Equus equus Rhinoceros antiquitatis Cervus elaphus Capreolus capreolus Bison priscus Saiga tatarica
5. Staroselie Bakhchisarai, the Crimea Mousterien (Vereshchagin, Baryshnikov, 1980, P 39)
Mammuthus primigenius Equus caballus Equus hydruntinus Coelodonta antiquitatis Cervus cf. elaphus Capreolus cf. capreolus Megaceros giganteus
. Rangifer tarandus Bison priscus Saiga tatarica
6. Shaitan-Koba Grotto Bodrak River, the Crimea Mousterien (Gromova, Gromov, 1937, p. 92)
Elephas sp. (cf. primigenius, trogontherii) Equus (Equus) caballus foss Equus (Asinus) sp. Cervus elaphus Bos sp.? (cf. Bison priscus) Saiga tatarica
7. Chokurcha Grotto Simpheropol, the Crimea Mousterien (Vereshchagin, Baryshnikov, 1980. p. 32)
Mammuthus primigenius Equus caballus

228 G. BARYSHNIKOV AND A. TIKHONOV
Equus hydruntinus Coelodonta antiquitatis Cervus elaphus Megaceros giganteus Rangifer tarandus Bison priscus Bison aut Bos Saiga tatarica
8. Kosh-Koba Grotto Zuya River, the Crimea Mousterien (Gromova, Gromov, 1937, P. 92)
Elephas sp.? (cf. primigenius, trogontherii) Equus (Equus) caballus foss Equus (Asinus) sp.? Rhinoceros antiquitatis Sus scrofa Cervus elaphus Cervus euryceros Rangifer tarandus Bos sp.? (cf. Bison priscus) Saiga tatarica
9. Kiik-Koba Zuya River, the Crimea Mousterien (Gromova, Gromov, 1937. p. 92)
Elephas sp.? (cf. primigenius trogontherii) Equus (Equus) caballus foss Equus (Asinus) sp.? Rhinoceros antiqiutatis Sus scrofa Cervus elaphus Cervus euryceros Rangifer tarandus Bos sp.? (cf. Bison priscus) Saiga tatarica Ovis sp.? (cf. argaloides)
10. Volchii Grotto Beshterek River, the Crimea Mousterien (Bader, Bader, 1979, p. 25)
Mammuthus primigenius Equus caballus Equus hydruntinus Coelodonta antiquitatis Sus scrofa Cervus elaphus Megaceros giganteus Rangifer tarandus Bison priscus Saiga tatarica Ovis sp.

PLEISTOCENE SAIGA IN EURASIA 229
1 1. Zaskalnaya 5 Grotto Biuk-Karasu River, the Crimea Mousterien (Kolossov, 1983, p. 28)
Mammuthus primigenius Equus caballus Cervus elaphus Megaceros giganteus Bison priscus Saiga tatarica
12. Zaskalnaya 6 Grotto Biuk-Karasu River, the Crimea Mousterien (Kolossov, 1986, p. 11)
Mammuthus primigenius Equus caballus Equus hydruntinus Coelodonta antiquitatis Cervus elaphus Megaceros giganteus Rangifer tarandus Bison priscus Saiga tatarica
13. Prolom 2 Kuchuk-Karasu River, the Crimea Mousterien (Kolossov, 1986. p. 1 1)
Mammuthus primigenius Equus cf. latipes Equus (Hydruntinus) hydruntinus Coelodonta antiquitatis Sus scrofa Cervus elaphus Megaceros giganteus Rangifer tarandus Bison priscus Saiga cf. borealis Capra aut Ovis . 14. Adzhi-Koba Cave (stratum 3) Karabi-Yaila, the Crimea Mousterien. (Gromova,
Gromov, 1937 p. 92)
Equus (Equus) caballus Equus (Asinus) sp.? Rhinoceros antiquitatis Cervus elaphus Bos sp.? (cf. Bison priscus) Saiga tatarica Ovis sp.? (cf. ammon)
15. Marnat-Koba Cave Karabi-Yaila, the Crimea Mousterien? (Vereshchagin, Baryshnikov, 1980, p. 40)
Equus caballus Equus hydruntinus

230 G. BARYSHNIKOV AND A. TIKHONOV
Coelodonta antiquitalis Cervus elaphus Capreolus capreolus Rangifer tarandus Saiga tatarica
16. Ilskaya 1 I1 River. Northern Caucasus Mousterien (Hoffecker et al., 1989, p. 70)
Mammuthus cf. chosaricus Equus (Equus) cf. mosbachensis Equus (Hydruntinus) hydruntinus Cervus elaphus Megaceros giganteus Bison priscus Saiga tatarica
17. Sukhaya Mechetka Volgograd, Volga Rover Mousterien (Vereshchagin, Kolbutov, 1957, p. 84)
Elephas primigenius Equus caballus Cervus elaphus Rangifer tarandus Bison priscus Saiga tatarica
Upper Wurm (Upper Waldai)
18. Starye Duruitory Grotto (stratum 2) Duruitory, Moldova Upper Palaeolithic (David, 1980, p. 26-27)
Mammuthus primigenius Equus latipes Equus sp. Coelodonta antiquitatis Sus scrofa Cervus elaphus Capreolus capreolus Megaceros giganteus Alces alces Rangifer tarandus Bison priscus Saiga tatarica
19. Anetovka 2 Yuzhnyi Bug River, Ukraine Upper Palaeolithic: 18040+150 (LE-2424) (Bibikova, Starkin, 1989, p. 129)
Equus latipes Cervus elaphus Rangifer tarandus Bison priscus Saiga tatarica

PLEISTOCENE SAIGA IN EURASIA 23 1
20. Siuren 1 Grotto Belbek River, the Crimea Upper Palaeolithic (Gromova, Gromov, 1937, p. 92)
Equus (Equus) caballus foss. Sus scrofa Cervus elaphus Cervus euryceros Rangifer tarandus Bos sp.? (cf. Bison primigenius) Saiga tatarica
21. Adzhi-Koba Cave (stratum 2) Karabi-Yaila, the Crimea Upper Palaeolithic (Gromova, Gromov, 1937, p. 92)
Equus (Equus) caballus foss. Equus (Asinus) sp.? Cervus elaphus Rangifer tarandus Bos sp.? (cf. Bison priscus) Saiga tatarica Capra (Ibex) sp.? (aff. sibirica) Ovis sp.? (cf. ammon) Ovis sp.? (cf. argaloides)
22. Kostenki 1 (stratum 3) Kostenki, Don River Upper Palaeolithic: stratum I - 21 300+400 (GIN-2534); 22300+230 (GIN-1 870); 23000+500 (GIN-2528); 24 100+500 (GIN-2524). (Vereshchagin, Kuzmina, 1977, p. i OOj
Mammuthus primigenius Equus caballus latipes Rangifer tarandus Saiga tatarica
. 23. Kostenki 4 (Vereshchagin, Kuzmina. 1977, p. 102)
Mammuthus primigenius Equus caballus latipes Coelodonta antiquitatis Sus scrofa Cervus elaphus Rangifer tarandus Bos aut Bison Saiga tatarica
24. Kostenki 8 (stratum 2) 27700+750 (GIN-10509) (Vereshchagin, Kuzmina, 1977, p. 104)
Mammuthus primigenius Equus caballus latipes Coelodonta antiquitatis Cervus elaphus Megaceros giganteus Rangifer tarandus

G. BARYSHNIKOV AND A. TIKHONOV
Bos aut Bison Saiga tatarica
25. Kostenki 14 (stratum 2) 19300+200 (LE-1400): 26400+660, 28200+700 (LU-59) (Vereshchagin, Kuzmina, 1977, p. 107)
Mammuthus primigenius Equus caballus latipes Coelodonta antiquitatis Cervus elaphus Rangifer tarandus Bos aut Bison Saiga tatarica
26. Bliznetsov Grotto Chanva River, Middle Urals Upper Palaeolithic (Kuzmina, 1975, p. 64)
Mammuthus primigenius Equus caballus latipes Coelodonta antisuitatis Alces alces Rangifer tarandus Bison priscus Saiga tatarica Ovibos moschatus
27. Stolbovoi Grotto Usva River, Middle Urals Upper Palaeolithic (Kuzmina, 1975, p. 64)
Equus caballus uralensis Coelodonta antiquitatis Rangifer tarandus Bison priscus Saiga tatarica
28. Medvezhya Cave Pechora River, North Urals Upper Palaeolithic: 17980+200 (LE-233) (Kuzmina, 1971, p. 56)
Mammuthus primigenius Equus caballus Coelodonta antiquitatis Capreolus capreolus Alces alces Rangifer tarandus Bison priscus Saiga tatarica Ovibos moschatus
29. Shaitan Grotto Ivdel River, Middle Urals Upper palaeolithic (Kosintsev, Borodin. 1990, p. 123)
Mammuthus primigenius Equus cf. uralensis

PLEISTOCENE SAIGA IN EURASIA 233
Coelodonta antiquitatis Cervus elaphus Rangifer tarandus Bison priscus Saiga tatarica Ovibos moschatus
30. Shikaevka 2 Shikaevka, Tobolo River Upper Palaeolithic (Petrin, Smirnov, 1975, p. 84-85)
Mammuthus primigenius Rangifer tarandus? Saiga tatarica
3 1. Novoselovo 7 Popovka River, Yenisei River Upper Palaeolithic (Abramova, 1979, p. 151)
Equus caballus Rangifer tarandus Bison priscus Saiga tatarica
32. Tashtyk 1 (stratum 3) Tashtyk River. Yenisei River Upper Palaeolithic: stratum I - 12180+120 (LE-771) (Zeitlin, 1979, p. 98)
Equus caballus Cervus elaphus Rangifer tarandus Bison aut Bos Saiga tatarica
33. Kokorevo 2 Yenisei River Upper Palaeolithic: 13330+100 (GIN-90) (Zeitlin, 1979, p. 112)
Mammuthus primigenius Equus caballus . Cervus elaphus Alces alces Rangifer tarandus Bison priscus Saiga tatarica Ovis ammon
34. Aphontova Gora 2 Krasnoyarsk. Yenisei River Upper Palaeolithic: 20900+300 (GIN-1 17) (Gromov, 1948, p. 322)
Elephas primigenius Equus (Equus) sp. Equus hemionus Cervus elaphus Capreolus pygargus Rangifer tarandus Bos aut Bison

G. BARYSHNIKOV AND A. TIKHONOV
Bos sp. Saiga tatarica Capra sibirica Capra sp. Ovis ammon
35. Aleshkinskaya Zaimka Kolyma River 14980+ 100 (MAG-470): 15000+200 (MAG-468) (Sher, 1971, p. 95)
Mammuthus primigenius Equus caballus subsp. Rangifer tarandus Bison priscus subsp. Saiga ricei
36. Northern Yakutia Bolshoy Liachovsky Island, Delta of Lena River (Chersky, 1891 p. 68-69)
Elephas primigenius Equus caballus Rhinoceros tichorhinus Cervus canadensis Alces palmatus Rangifer tarandus Bison priscus Colus saiga Ovibos moschatus Ovis nivicola
Copyright © 2022 FDOKUMEN




















