
Nitrogen reserve status affects the dynamics of nitrogen remobilization and mineral nitrogen uptake...
12
Printed from the CJO service for personal use only by... New Phytol. (1999), 142, 451–462 Nitrogen reserve status affects the dynamics of nitrogen remobilization and mineral nitrogen uptake during recovery of contrasting cultivars of Lolium perenne from defoliation S. LOUAHLIA " , J. H. MACDUFF # , A. OURRY " *, M. HUMPHREYS # J. BOUCAUD " " UA INRA 950 Physiologie et Biochimie Ve U ge U tales, Institut de Recherche en Biologie Applique U e, Universite U , 14032 Caen Cedex, France # Institute of Grassland and Environmental Research, Plas Gogerddan, Aberystwyth, Ceredigion, Dyfed SY23 3EB, UK Received 26 October 1998 ; accepted 15 March 1999 Nitrogen (N) allocated to leaf growth in forage grasses and legumes following severe defoliation is predominately mobilized from the remaining root and leaf sheath tissues, since both N uptake from the soil and N # fixation are severely down-regulated for several days. The hypothesis that a low N reserve status at the time of defoliation limits N remobilization and leaf regrowth was tested with contrasting cultivars of Lolium perenne (cvs Aberelan and Cariad) in flowing solution culture. Plants were grown under ‘ high ’ or ‘ low ’ (uptake of N decreased by 50 %) regimes of N supply for 10 d before a single severe defoliation. Labelling with "&N was used to assess the importance of N reserves, including putative vegetative storage proteins, relative to N translocated from concurrent uptake, as a source of leaf N during regrowth. Leaf regrowth, N uptake and N mobilization were all affected by previous N supply. Low plant N status at the time of defoliation increased regrowth dry weight of ‘ Aberelan ’ by 10 % and translocation of N absorbed from the medium by 23 %, while mobilization of N reserves was decreased by 56 %. On the contrary, regrowth dry weight of ‘ Cariad ’ was decreased by 23 %, and translocation of N absorbed by 21 % in low plant N status, compared with high plant N status. Concentrations of soluble protein in roots and remaining leaf sheaths decreased after defoliation in plants only under optimal N supply. Analysis of soluble proteins in sheath material by SDS–PAGE suggested that three polypeptides (55, 36.6 and 24 kDa) might function as vegetative storage proteins, although they were of low abundance in plants, subjected to monthly harvests, grown in controlled conditions and in the field. The apparent antagonism between uptake of NH % + or NO $ - by roots and mobilization of N reserves is discussed together with evidence for functional vegetative storage proteins in L. perenne. Key words : ammonium, defoliation, Poaceae, Lolium perenne, nitrate, vegetative storage proteins, fructans. Mechanical defoliation and grazing induce major modifications in the carbon (C) and N metabolism of grasses and forage legumes during their subsequent recovery (Richards, 1993 ; Volenec et al., 1996). For example, uptake of mineral N decreases rapidly and Abbreviations : DP, degree of polymerization ; FSC, flowing solution culture ; VSP, vegetative storage protein ; WSC, water soluble carbohydrate. *Author for correspondence (fax 00 33 2 56 53 60 ; e-mail ourry!ibba.unicaen.fr). acutely (Macduff & Jackson, 1992), influx of nitrate and ammonium decreases while their efflux from the roots to the external medium appears to increase. It is not understood how this is regulated and exactly which signals and transduction pathways are involved (Macduff & Jackson, 1992). It is generally accepted (Richards, 1993 ; Volenec et al., 1996) that adaptation to defoliation in many species involves a capacity for mobilization of N compounds stored in perennial tissue such as roots, stubble or crown, enabling N to be supplied to growing leaves despite the down-regulation of N
Transcript of Nitrogen reserve status affects the dynamics of nitrogen remobilization and mineral nitrogen uptake...

Printed from the CJO service for personal use only by...
New Phytol. (1999), 142, 451–462
Nitrogen reserve status affects the
dynamics of nitrogen remobilization and
mineral nitrogen uptake during recovery of
contrasting cultivars of Lolium perenne from
defoliation
S. LOUAHLIA " , J . H. MACDUFF # , A. OURRY "*, M. HUMPHREYS #
J. BOUCAUD"
"UA INRA 950 Physiologie et Biochimie VeU geU tales, Institut de Recherche en Biologie
AppliqueU e, UniversiteU , 14032 Caen Cedex, France
# Institute of Grassland and Environmental Research, Plas Gogerddan, Aberystwyth,
Ceredigion, Dyfed SY23 3EB, UK
Received 26 October 1998; accepted 15 March 1999
Nitrogen (N) allocated to leaf growth in forage grasses and legumes following severe defoliation is predominately
mobilized from the remaining root and leaf sheath tissues, since both N uptake from the soil and N#fixation are
severely down-regulated for several days. The hypothesis that a low N reserve status at the time of defoliation
limits N remobilization and leaf regrowth was tested with contrasting cultivars of Lolium perenne (cvs Aberelan
and Cariad) in flowing solution culture. Plants were grown under ‘high’ or ‘ low’ (uptake of N decreased by 50%)
regimes of N supply for 10 d before a single severe defoliation. Labelling with "&N was used to assess the
importance of N reserves, including putative vegetative storage proteins, relative to N translocated from
concurrent uptake, as a source of leaf N during regrowth. Leaf regrowth, N uptake and N mobilization were all
affected by previous N supply. Low plant N status at the time of defoliation increased regrowth dry weight of
‘Aberelan’ by 10% and translocation of N absorbed from the medium by 23%, while mobilization of N reserves
was decreased by 56%. On the contrary, regrowth dry weight of ‘Cariad’ was decreased by 23%, and
translocation of N absorbed by 21% in low plant N status, compared with high plant N status. Concentrations
of soluble protein in roots and remaining leaf sheaths decreased after defoliation in plants only under optimal N
supply. Analysis of soluble proteins in sheath material by SDS–PAGE suggested that three polypeptides (55, 36.6
and 24 kDa) might function as vegetative storage proteins, although they were of low abundance in plants,
subjected to monthly harvests, grown in controlled conditions and in the field. The apparent antagonism between
uptake of NH%
+or NO$
− by roots and mobilization of N reserves is discussed together with evidence for functional
vegetative storage proteins in L. perenne.
Key words: ammonium, defoliation, Poaceae, Lolium perenne, nitrate, vegetative storage proteins, fructans.
Mechanical defoliation and grazing induce major
modifications in the carbon (C) and N metabolism of
grasses and forage legumes during their subsequent
recovery (Richards, 1993; Volenec et al., 1996). For
example, uptake of mineral N decreases rapidly and
Abbreviations: DP, degree of polymerization; FSC, flowing
solution culture; VSP, vegetative storage protein; WSC, water
soluble carbohydrate.
*Author for correspondence (fax 00 33 2 56 53 60; e-mail
ourry!ibba.unicaen.fr).
acutely (Macduff & Jackson, 1992), influx of nitrate
and ammonium decreases while their efflux from the
roots to the external medium appears to increase. It
is not understood how this is regulated and exactly
which signals and transduction pathways are
involved (Macduff & Jackson, 1992).
It is generally accepted (Richards, 1993; Volenec
et al., 1996) that adaptation to defoliation in many
species involves a capacity for mobilization of N
compounds stored in perennial tissue such as roots,
stubble or crown, enabling N to be supplied to
growing leaves despite the down-regulation of N

Printed from the CJO service for personal use only by...
452 S. Louahlia et al.
uptake by the roots. Several studies (Phillips et al.,
1983; Thornton et al., 1993, 1994; Thornton &
Millard, 1996) have shown that different grass
species, including Lolium perenne, Festuca rubra,
Agrostis castellana, Poa trivialis, Bromus mollis are
able to remobilize internal N to sustain early
regrowth. In certain species such as Medicago sativa
(Ourry et al., 1994) or the annual grass Lolium
temulentum (Ourry et al., 1996) it has been argued
that the status of N reserves at the time of defoliation
may be a crucial determinant of regrowth rate. This
is based on a view that growth is severely decreased
by a shortage of organic N, delaying refoliation and
recovery of N#
fixation and}or N uptake. It follows
that large differences in forage regrowth yields might
result from relatively moderate initial differences in
availability of N reserves. However, this has not
been demonstrated convincingly for an agro-
nomically important and perennial species of grass
such as L. perenne. Thornton et al. (1993, 1994) and
Thornton & Millard (1996) have argued that reliance
on mobilized N during recovery from defoliation
tends to be higher in those grass species that are
abundant in low input, ‘extensive’ grassland farming
systems. As a consequence they suggested that a high
propensity for N storage in grasses is an adaptation
to environments poor in N.
Given that the free nitrate pool is only modest in
most plants, and functions as a short-lived storage
compartment (!24 h), it is organic forms of N such
as amino acids and proteins that constitute the main
N storage components within the plant (e.g. Millard,
1988). This is supported by the decline in soluble
protein concentrations observed during the first few
days following shoot removal in M. sativa
(Hendershot & Volenec, 1993), Trifolium repens
(Corre et al., 1996) and L. perenne (Ourry et al.,
1989). Staswick (1989, 1990, 1994) has proposed
several criteria by which the involvement of specific
proteins in N storage may be judged. Accordingly,
vegetative storage proteins (VSP, see Abbreviations
section) are supposed to be prominent (accounting
for at least 5% of total soluble proteins), readily
hydrolysed (since their degradation products are
used by growth of tissues other than those containing
VSP) and devoid of any catalytic activity.
Several polypeptides fulfilling these criteria have
been identified in T. repens and M. sativa, and have
been designated as functional VSPs. These are
relatively abundant proteins (Hendershot & Volenec,
1993; Corre et al., 1996) that are hydrolysed to a
greater extent than are the majority of soluble
proteins, and then accumulate during the later stages
of refoliation. As with many of the VSPs identified in
woody species (Ste!pien et al., 1994), it is notable that
these VSPs in forage legumes accumulate during
autumn and winter, and are then almost entirely
hydrolysed when growth resumes in spring
(Bouchard et al., 1998). Rather surprisingly, no
polypeptides conforming to the criteria for VSPs
have been identified in other forage legumes such
as Trifolium pratense or Melilotus officinalis
(Cunningham & Volenec, 1996; Li et al., 1996).
Furthermore, to our knowledge, there have been no
attempts to assess the presence or absence of VSPs in
grasses.
The aim of the present experimental work with
L. perenne was to resolve these uncertainties. The
specific objectives were, first, to determine whether
the N status of plants at the time of defoliation
determines regrowth rate of the shoot and the rate of
recovery in the uptake of mineral N supplied as
NO$
− plus NH%
+. As a corollary, the extent to which
rates of remobilization and uptake of these two forms
of N are inversely related during recovery from
defoliation should be forthcoming. To provide a
measure of intra-specific variation, two contrasting
varieties of perennial ryegrass were compared:
‘Aberelan’, known to have a high nitrogen use
efficiency (NUE) (Wilkins et al., 1997), and ‘Cariad’,
known to have a tendency to accumulate higher than
average concentrations of water soluble carbo-
hydrates (WSC) in its leaves and sheaths (Radojevic
et al., 1994). The second objective was to determine
whether specific polypeptides in L. perenne conform
to the criteria for VSPs, with N mobilization
triggered by defoliation or the onset of spring
growth.
A steady-state "&N-labelling technique in flowing
solution culture (FSC), which provided precisely
defined high and low regimes of N supply before
defoliation, was used to allow detailed resolution of
the dynamics of N mobilization and mineral N
uptake during a 21 d recovery period. A comp-
lementary plot experiment assessed the seasonal
variation in putative VSPs of L. perenne under field
conditions.
Plant culture
For the experiment in flowing solution culture
(FSC), seeds of L. perenne L., cvs Cariad and
Aberelan were imbibed for 48 h on tissue paper
saturated with 10 mol m−$ CaSO%. Each cultivar was
then sown separately into two culture units of an
FSC system, incorporating automatic control of
concentrations of NO$
−, NH%
+, K+ and H+ in
solution (Clement et al., 1974; Hatch et al., 1986).
Each culture unit contained 200 dm$ of recirculating
nutrient solution and 24 culture vessels, each with 10
plants. The FSC system was located in a glasshouse,
solution temperature was maintained at 20³0.5°Cand air temperature at 20³2 : 15³1°C day : night
(16.00–06.00 hours) throughout the experiment. The
plants were established under natural illumination
until day 14 after sowing. Supplementary light

Printed from the CJO service for personal use only by...
N reserves and regrowth 453
(08.00–16.00 hours) of 200 µmol m−# s−" PAR was
provided between days 14 and 25 by a single 400 W
SON-T lamp (Philips) suspended 1.5 m above the
surface of each culture unit. On day 25 after sowing
all natural light was excluded and illumination was
then provided (including during the treatment
period) by paired 400 W SON-T and HPI}T lamps
(Philips Lighting Ltd, Croydon, Surrey, UK) giving
500 µmol m−# s−" PAR at canopy height.
Until day 38 after sowing, the concentrations of
nutrients in solutions were maintained by regular
manual additions at (mmol m−$) : NO$
−, 250; NH%
+,
250; K+, 250; Mg#+, 100; Ca#+, 424; HPO%
−, 50;
SO%
#−, 550; Fe#+, 5.4; with micronutrients as given
by Clement et al. (1978). Thereafter, the automatic
nutrient control system measured the concentrations
of K+, NO$
− and NH%
+ in each culture unit every 27
min and maintained the treatment ‘set-point ’
concentrations of these ions (to be described later),
by automatic resupply of K#SO
%, (NH
%)#SO
%or
Ca(NO$) at rates equal to their depletion. Net uptake
of these ions was calculated on an hourly basis from
the amounts required to maintain the set-point
concentrations in the flowing solutions. All other
nutrients were resupplied automatically in fixed
ratios to the net uptake of NO$
− and NH%
+ (for one
mol of N, 0.645, 0.057, 0.045, 0.00075 and 0.522 mol
of, respectively, S, Mg, P, Fe and Ca were auto-
matically resupplied). Solution pH was automati-
cally maintained at 6.0³0.1 by delivery of H#SO
%or
Ca(OH)#.
Experimental treatments
In order to condition the N status of the plants at the
time of defoliation, treatments consisting of two
different rates of N supply (‘high N’ and ‘low N‘)
were imposed on day 38 after sowing and continued
for 10 d. The supply of N was defined in terms of
constant concentrations in the flowing nutrient
solutions. The aim was that the ‘ low N’ plants
should absorb approx. 50% of the total N absorbed
by ‘high N’ plants during this period. Based on
previous experience (Wild et al., 1987), the N supply
to ‘high N’ plants (one culture unit of each cultivar)
was maintained at 20 mmol m−$ NH%NO
$, whereas
the ‘ low N’ plants (one culture unit per cultivar)
received 5 mmol m−$ NH%NO
$. In reality the net
uptake of N over 10 d by ‘low N’ ‘Cariad’ was 58%
of the amount absorbed by ‘high N’ plants, whereas
the corresponding figure for ‘ low N’ plants of
‘Aberelan’ was 44%.
All plants were defoliated on day 48 after sowing,
referred to hereafter as day 0. At 1 h before
defoliation (17.00 hours) the automatic resupply of
N was terminated and the solution concentrations
allowed to deplete to !1 mmol m−$ NH%NO
$. Plants
were defoliated by cutting the shoot 4 cm above the
shoot-root junction. Automatic N supply to all four
culture units was immediately resumed at 20 mmol
m−$ "&NH%
"&NO$(5.00 atom % "&N), irrespective of
previous N supply, and continued over 14 d of
regrowth.
Plants were harvested at intervals during the
regrowth period: four culture vessels from each unit
on days 0 (6 h before defoliation), 2 and 4; then three
vessels on days 7, 11 and 14. On each occasion the
plants within a specific culture vessel were bulked
before dividing into roots, sheath, laminae inside the
sheath beneath the defoliation height (4 cm), and
regrowing leaves. Fresh weights were recorded,
after which fractions were freeze-dried, reweighed,
ground to a fine powder, and kept at ®80°C until
analysis.
Field experiment
Seasonal changes in VSP content of L. perenne were
measured in a plot experiment located at the INRA
experimental station in NW France. Three plots (8
m#) of L. perenne cv. Magella were established and
fertilized with 6 g m−# (as NH%NO
$) during March
1997. Following the resumption of growth in the
spring the plots were defoliated on three occasions
(22 Apr., 6 June, 10 July 1997). Whole plants were
harvested at monthly intervals between November
1996-July 1997. The leaf sheath fraction was freeze-
dried, ground to a fine powder and stored at ®80°Cuntil analysis.
Chemical analysis and calculation of N
remobilization
The determination of total N and "&N in plant
samples from the FSC experiment was performed by
a continuous flow isotope mass spectrometer (20–20,
Europa Scientific Ltd, Crewe, UK) linked to a C}N
analyser (Roboprep CN, Europa Scientific Ltd). All
mineral N absorbed from the flowing solutions
during regrowth was "&N labelled. Consequently,
patterns of net translocation of unlabelled N ("%N
absorbed before defoliation) between plant parts
were used to calculate N remobilization to regrowing
tissues. The N in regrowing leaves derived from the
mobilization of N reserves from roots and sheaths
was calculated by subtracting from total N
("%N"&N), first, the "&N content derived from
uptake of "&NH%
+ or "&NO$
− and, second, the initial
"%N content of laminae remaining after defoliation
(i.e. leaf tissue remaining inside the sheath between
the shoot-root junction and the point of defoliation).
Electrophoretic analysis of soluble proteins
In both experiments, the soluble proteins were
extracted from 100 mg of freeze-dried roots or
sheath at 4°C with 5 ml of 50 mM Tris buffer (pH
7.5) containing 2 mM phenylmethylsulphonyl

Printed from the CJO service for personal use only by...
454 S. Louahlia et al.
fluoride, 10 µM leupeptin, 1 mM EDTA and
0.1% (v}v) 2-mercaptoethanol. After centrifugation
(12 000 g, 10 min), using an aliquot of the supernatant
the soluble proteins were precipitated with 7.2%
(w}v) trichloroacetic acid and their content was
determined by the method of Lowry et al. (1951).
Bovine serum albumin (BSA) was used as the protein
standard. Nucleic acids in the remaining supernatant
were precipitated using protamine sulphate (1 mg
ml−") for 15 min. The nucleic acid pellet was
discarded after centrifugation (28000 g, 10 min).
Soluble proteins were separated into five sub-
samples, each receiving 0.015% (w}v) sodium
deoxycholate. After 10 min, the soluble proteins
were precipitated at 4°C with 7.2% (w}v) trichloro-
acetic acid. The protein pellet was rinsed with
acetone. For SDS–PAGE analysis, one pellet of
soluble protein was resuspended in 100 µl of the
buffer described by Leammli (1970), denatured for 5
min at 100°C, and centrifuged (12000 g, 3 min).
SDS-electrophoresis used a 15% duracryl running
gel with a stacking gel containing 5.5% acrylamide.
Two wells were used for loading proteins with
known molecular masses. The gels were run for 4 h
at a constant 500 V, and silver stained as described
by Lopez et al. (1991), then scanned and quan-
titatively analysed using a computerized image
analysis system (Millipore Bioimage).
For two-dimension gel electrophoresis, one pellet
of soluble protein was resuspended in the buffer
described by O’Farrell (1975) and used to run 2-D
SDS–PAGE gels, loaded with 124 µg of protein,
according to a modified procedure from O’Farrell
(1975) on an Investigator System (Millipore Corp.).
First-dimension isoelectric focusing was in a 4.1%
acrylamide tube gel containing 9.5 M urea, 2% (v}v)
Triton X100, 5 mM (3-((3-cholamidopropyl)-
dimethylammonio)-1-propanesulphonate), and 2%
(v}v) Millipore 2-D optimized carrier ampholytes
(pH 3–10). After 2 h of prefocusing to 1500 V with
current limited to 110 µA per tube, protein samples
(20 µl) were loaded on the basic end of the tube gel
and focused with 200 V per tube for 17.5 h. The gels
were removed and pre-equilibrated for 2 min in a
375 mM Tris-HCl buffer (pH 8.6), containing 3%
(w}v) sodium dodecyl sulphate and 50 mM dithio-
threitol. Separation of the isoelectrofocused proteins
in the second dimension was as for the one-
dimensional gels, using a 15% duracryl running gel,
run for 5 h at 500 V. Gels were subsequently silver
stained as described by Lopez et al. (1991).
The two-dimensional gels were analysed by
computerized image analysis (Millipore Bio Image
Millipore Corp., Ann Arbor, MI, USA). Gels were
scanned, and the individual staining intensities of
each polypeptide was then expressed as a percentage
of the total staining intensity of the gel. For each gel,
silver staining intensities of unknown proteins were
estimated using silver staining intensities of proteins
of known molecular weight and isoelectric point,
which were added to each gel in constant amounts of
proteins.
Analysis of soluble sugar
Soluble carbohydrates from freeze-dried samples
were extracted with 80% (v}v) ethanol (30 ml g−" d.
wt) for 1 h at 80°C. After centrifugation (12000 g, 10
min) the supernatant contained fructose, glucose,
sucrose and fructans of 4–7 degree of polymerization
(DP). Fructans of DP "7 were extracted by
resuspending the initial insoluble residue in water
for 1 h at 60°C. After centrifugation (12000 g, 10
min) the supernatants were pooled and evaporated to
dryness under vacuum. The dried extracts were
redissolved in water and filtered through a 0.45 µm
membrane. Aliquots of carbohydrate extracts were
passed through a column containing cation exchange
resin (DOWEX Hydrogen form, mesh 200–400,
Sigma Corp., St Louis, MO, USA) and anion
exchange resin (Amberlite IRA 416, formate form,
Serva, Germany) to remove charged compounds
(Smouter & Simpson, 1991). The columns were
eluted with water.
Samples were concentrated under vacuum, dis-
solved in water and aliquots were taken for analysis
by HPLC. Samples (20 µl) were first injected
through a cation exchange column (Sugar-PAK,
300¬6.5 mm, Millipore Waters, Milford, MA,
USA). The sugars were eluted at 85°C in water at a
flow rate of 0.5 ml min−" and detected with a
differential refractometer (156 Index Detector,
Beckman Instruments, San Ramon, CA, USA).
After separation, the sugars were quantified using
mannitol as an internal standard.
Regrowth, N uptake and N mobilization
There were marked differences between the two
cultivars in several aspects of their response to the
N-treatments. Dry weight of plants, tiller numbers,
and tissue concentrations of N and C immediately
after defoliation (day 0) are given in Table 1. ‘Low
N’ supply during the 10 d before defoliation did not
affect sheath or root dry weight significantly in cv.
Cariad, although the root dry weight of cv. Aberelan
was increased by 25%. The number of tillers per
plant was also unaffected by the regime of N
pretreatment, but was higher for ‘Cariad’ than
‘Aberelan’, accounting for the cultivar differences in
root and sheath dry weight.
As anticipated, ‘ low N’ pretreatment decreased
the total N and soluble protein concentrations in the
plants by the time of defoliation, except for soluble
protein concentrations in the roots of cv. Aberelan.
By contrast, the total concentration of WSC was
Printed from the CJO service for personal use only by...
N reserves and regrowth 455
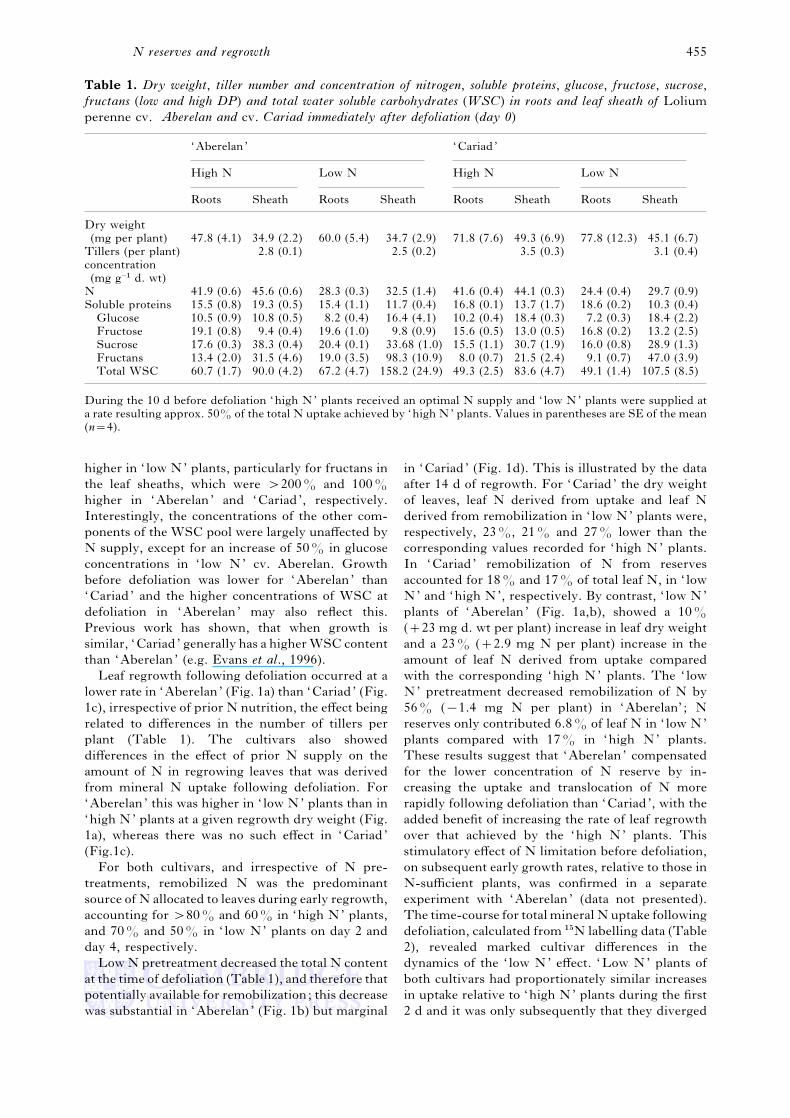
Table 1. Dry weight, tiller number and concentration of nitrogen, soluble proteins, glucose, fructose, sucrose,
fructans (low and high DP) and total water soluble carbohydrates (WSC) in roots and leaf sheath of Lolium
perenne cv. Aberelan and cv. Cariad immediately after defoliation (day 0)
‘Aberelan’ ‘Cariad’
High N Low N High N Low N
Roots Sheath Roots Sheath Roots Sheath Roots Sheath
Dry weight
(mg per plant) 47.8 (4.1) 34.9 (2.2) 60.0 (5.4) 34.7 (2.9) 71.8 (7.6) 49.3 (6.9) 77.8 (12.3) 45.1 (6.7)
Tillers (per plant) 2.8 (0.1) 2.5 (0.2) 3.5 (0.3) 3.1 (0.4)
concentration
(mg g−" d. wt)
N 41.9 (0.6) 45.6 (0.6) 28.3 (0.3) 32.5 (1.4) 41.6 (0.4) 44.1 (0.3) 24.4 (0.4) 29.7 (0.9)
Soluble proteins 15.5 (0.8) 19.3 (0.5) 15.4 (1.1) 11.7 (0.4) 16.8 (0.1) 13.7 (1.7) 18.6 (0.2) 10.3 (0.4)
Glucose 10.5 (0.9) 10.8 (0.5) 8.2 (0.4) 16.4 (4.1) 10.2 (0.4) 18.4 (0.3) 7.2 (0.3) 18.4 (2.2)
Fructose 19.1 (0.8) 9.4 (0.4) 19.6 (1.0) 9.8 (0.9) 15.6 (0.5) 13.0 (0.5) 16.8 (0.2) 13.2 (2.5)
Sucrose 17.6 (0.3) 38.3 (0.4) 20.4 (0.1) 33.68 (1.0) 15.5 (1.1) 30.7 (1.9) 16.0 (0.8) 28.9 (1.3)
Fructans 13.4 (2.0) 31.5 (4.6) 19.0 (3.5) 98.3 (10.9) 8.0 (0.7) 21.5 (2.4) 9.1 (0.7) 47.0 (3.9)
Total WSC 60.7 (1.7) 90.0 (4.2) 67.2 (4.7) 158.2 (24.9) 49.3 (2.5) 83.6 (4.7) 49.1 (1.4) 107.5 (8.5)
During the 10 d before defoliation ‘high N’ plants received an optimal N supply and ‘ low N’ plants were supplied at
a rate resulting approx. 50% of the total N uptake achieved by ‘high N’ plants. Values in parentheses are SE of the mean
(n¯4).
higher in ‘ low N’ plants, particularly for fructans in
the leaf sheaths, which were "200% and 100%
higher in ‘Aberelan’ and ‘Cariad’, respectively.
Interestingly, the concentrations of the other com-
ponents of the WSC pool were largely unaffected by
N supply, except for an increase of 50% in glucose
concentrations in ‘ low N’ cv. Aberelan. Growth
before defoliation was lower for ‘Aberelan’ than
‘Cariad’ and the higher concentrations of WSC at
defoliation in ‘Aberelan’ may also reflect this.
Previous work has shown, that when growth is
similar, ‘Cariad’ generally has a higher WSC content
than ‘Aberelan’ (e.g. Evans et al., 1996).
Leaf regrowth following defoliation occurred at a
lower rate in ‘Aberelan’ (Fig. 1a) than ‘Cariad’ (Fig.
1c), irrespective of prior N nutrition, the effect being
related to differences in the number of tillers per
plant (Table 1). The cultivars also showed
differences in the effect of prior N supply on the
amount of N in regrowing leaves that was derived
from mineral N uptake following defoliation. For
‘Aberelan’ this was higher in ‘ low N’ plants than in
‘high N’ plants at a given regrowth dry weight (Fig.
1a), whereas there was no such effect in ‘Cariad’
(Fig.1c).
For both cultivars, and irrespective of N pre-
treatments, remobilized N was the predominant
source of N allocated to leaves during early regrowth,
accounting for "80% and 60% in ‘high N’ plants,
and 70% and 50% in ‘low N’ plants on day 2 and
day 4, respectively.
Low N pretreatment decreased the total N content
at the time of defoliation (Table 1), and therefore that
potentially available for remobilization; this decrease
was substantial in ‘Aberelan’ (Fig. 1b) but marginal
in ‘Cariad’ (Fig. 1d). This is illustrated by the data
after 14 d of regrowth. For ‘Cariad’ the dry weight
of leaves, leaf N derived from uptake and leaf N
derived from remobilization in ‘ low N’ plants were,
respectively, 23%, 21% and 27% lower than the
corresponding values recorded for ‘high N’ plants.
In ‘Cariad’ remobilization of N from reserves
accounted for 18% and 17% of total leaf N, in ‘ low
N’ and ‘high N’, respectively. By contrast, ‘ low N’
plants of ‘Aberelan’ (Fig. 1a,b), showed a 10%
(23 mg d. wt per plant) increase in leaf dry weight
and a 23% (2.9 mg N per plant) increase in the
amount of leaf N derived from uptake compared
with the corresponding ‘high N’ plants. The ‘low
N’ pretreatment decreased remobilization of N by
56% (®1.4 mg N per plant) in ‘Aberelan’ ; N
reserves only contributed 6.8% of leaf N in ‘low N’
plants compared with 17% in ‘high N’ plants.
These results suggest that ‘Aberelan’ compensated
for the lower concentration of N reserve by in-
creasing the uptake and translocation of N more
rapidly following defoliation than ‘Cariad’, with the
added benefit of increasing the rate of leaf regrowth
over that achieved by the ‘high N’ plants. This
stimulatory effect of N limitation before defoliation,
on subsequent early growth rates, relative to those in
N-sufficient plants, was confirmed in a separate
experiment with ‘Aberelan’ (data not presented).
The time-course for total mineral N uptake following
defoliation, calculated from "&N labelling data (Table
2), revealed marked cultivar differences in the
dynamics of the ‘ low N’ effect. ‘Low N’ plants of
both cultivars had proportionately similar increases
in uptake relative to ‘high N’ plants during the first
2 d and it was only subsequently that they diverged

Printed from the CJO service for personal use only by...
456 S. Louahlia et al.
20
15
10
5
00 100 200 300 400
Regrowth dry weight (mg per plant)
Lab
elle
d N
fro
m u
pta
ke (
mg
per
pla
nt)
0 100 200 300 400
20
15
10
5
0
0500
1
2
3
4
5
0
1
2
3
Un
lab
elle
d N
fro
m m
ob
iliza
tio
n (
mg
per
pla
nt)
0 10050 150 200 250 0 10050 150 200 250
(a) (b)
(c) (d)
Fig. 1. Effect of N supply before defoliation of Lolium perenne ‘Aberelan’ (a,b) and ‘Cariad’ (c,d) on the source
of N allocated to regrowing leaves plotted as a function of leaf dry weight. Successive points refer to days 0,
2, 4, 7, 11 and 14 after defoliation. N from uptake of NH%NO
$after defoliation (a,c) was labelled with "&N; N
remobilized from roots and sheath tissue (b,d) was unlabelled. During the 10 d before defoliation ‘high N’
plants (filled squares) received an optimal N supply whereas ‘ low N’ plants (open squares) were supplied at
a rate resulting in approx. 50% of the total N uptake achieved by ‘high N’ plants. Vertical and horizontal bars
indicate³SE of the mean when larger than the symbol, n¯4 for the first three harvests and n¯3 for the
remaining harvests. Note the different scales on the horizontal axis for a,b and for c,d.
200
150
100
50
00 2 4 6 8
Days of regrowth
Cu
mu
late
d µ
mo
l nit
rate
or
amm
on
ium
per
pla
nt
10 12
200
150
100
50
0
500
750
1000
1250
250
1000
1250
1500
(a) (b)
(c) (d)
14 0 2 4 6 8 10 12 14
250
250
300
750
500
0
200
400
600
0
200
400
600
800
Fig. 2. Effect of previous N supply on cumulative net uptake of NO$
− (a,c) and NH%
+ (b,d) from 20 mmol m−$
NH%NO
$by Lolium perenne ‘Aberelan’ (a,b) and ‘Cariad’ (c,d) during 14 d of regrowth following defoliation.
During the 10 d before defoliation ‘high N’ plants (filled squares) received an optimal N supply whereas ‘ low
N’ plants (open squares) were supplied at a rate resulting in approx. 50% of the total N uptake achieved by
‘high N’ plants. Uptake was calculated from the amount of each N form delivered to flowing nutrient solutions
in order to maintain constant concentrations, and was highly correlated (r !0.99) with values obtained by "&N
labelling of the solutions. Note that the numbers in the centre apply both to the right hand axis of a and c as
well as to the left hand axis of b and d.

Printed from the CJO service for personal use only by...
N reserves and regrowth 457
Table 2. Total uptake of N from 20 mmol m−$ NH%NO
$by Lolium perenne cv. Aberelan and cv. Cariad during
14 d of regrowth after defoliation, calculated from "&N labelling data
Cumulative N uptake (mg N g−" root d. wt)
‘Aberelan’ ‘Cariad’
Days after
defoliation High N Low N % high N High N Low N % high N
2 15.6 (0.6) 26.3 (1.2) 69 12.6 (0.4) 21.26 (0.9) 69
4 36.4 (3.7) 46.6 (1.8) 28 36.9 (5.4) 38.0 (1.4) 0
7 96.0 (1.6) 97.6 (3.2) 0 100.2 (6.7) 86.9 (1.4) ®13
11 180.8 (5.8) 187.5 (3.5) 0 198.5 (5.0) 169.6 (5.9) ®14
14 212.2 (3.9) 212.4 (5.7) 0 225.0 (4.9) 200.2 (8.3) ®11
During the 10 d before defoliation,‘high N’ plants received an optimal N supply and ‘ low N’ plants were supplied at
a rate resulting in approx. 50% of the total N uptake achieved by ‘high N’ plants. Values in parentheses are SE of the
mean (n¯4 for the first three harvests and n¯3 for the remaining harvests). Uptake by ‘ low N’ plants is also expressed
as a proportion of uptake by ‘high N’ plants.
20
10
50 3 6 9
Days of regrowth
So
lub
le p
rote
ins
(mg
g–1
d. w
t)
12
20
15
(a) (b)
(c) (d)
30
25
10
0 3 6 9 12
15
25
Fig. 3. Changes in concentrations of soluble protein in sheath (a,c) and roots (b,d) of Lolium perenne ‘Aberelan’
(a,b) and ‘Cariad’ (c,d) during 14 d of regrowth after defoliation. During the 10 d before defoliation ‘high N’
plants (filled squares) received an optimal N supply whereas ‘ low N’ plants (open squares) were supplied at
a rate resulting in approx. 50% of the total N uptake achieved by ‘high N’ plants. Vertical and horizontal bars
indicate³SE of the mean when larger than the symbol, n¯4 for the first three harvests and n¯3 for the
remaining harvests.
in response. In terms of the net effect over 14 d, ‘ low
N’ pretreatment decreased total N uptake by 11% in
‘Cariad’ compared with ‘high N’ plants, but had no
effect on ‘Aberelan’.
The extent to which ‘low N’ treatment before
defoliation affected the subsequent uptake of NH%
+
and NO$
−, from the 20 mmol m−$ NH%NO
$supply,
varied with cultivar and time (Fig. 2). For example,
during the early phase of regrowth (0–4 d) of
‘Aberelan’, ‘ low N’ showed increased uptake of
both NH%
+ and NO$
− relative to ‘high N’ plants
(Fig. 2a,b), whereas ‘ low N’ ‘Cariad’ increased only
NO$
− uptake (Fig. 2c). Further, the relative en-
hancement in NO$
− uptake by ‘low N’ compared
with ‘high N’ plants persisted over 14 d in ‘Cariad’,
but was reversed after day 4 in ‘Aberelan’, although
the uptake of NH%
+ remained higher by ‘low N’
plants of both cultivars. The total uptake of NH%
+
over the experimental period was approx. twofold
higher than that of NO$
−, irrespective of cultivar and
N treatment before defoliation.
Soluble protein and profiles after SDS–PAGE
In both cultivars, ‘ low N’ pretreatment decreased
the concentrations of soluble proteins present at the
time of defoliation in sheath tissues (Fig. 3a,c) but
had hardly any effect in roots (Fig. 3b,d). The

Printed from the CJO service for personal use only by...
458 S. Louahlia et al.
45
kDa
31
21.5
14.5
D0 D2 D4 D7 D11 D14
(a)
55 kDa
36.6 kDa
24 kDa
45
kDa
31
21.5
14.5
D0 D2 D4 D7 D11 D14
(b)
Fig. 4. The effect of ‘high N’ (a) or ‘ low N’ (b) pretreatment before defoliation of Lolium perenne ‘Aberelan’
on subsequent changes in SDS–PAGE profiles of soluble proteins extracted from sheath tissue on days 0, 2,
4, 7, 11 and 14. Each well was loaded with soluble proteins extracted from 1.8 mg tissue d. wt, and the position
of molecular weight markers is indicated on each side. Arrows indicate polypeptides of 55, 36.6 and 24 kDa
affected by defoliation.
43
kDa
30
20.1
14.1
(a)
43
kDa
30
20.1
14.1
(b)
4.8 6.7IEF
MW4.8 6.7
IEF
MW
Fig. 5. Two-dimensional gel electrophoresis of soluble proteins extracted from ‘high N’ sheath tissue of Loliumperenne ‘Aberelan’, on day 0 immediately after defoliation (a) and 4 d later (b). Each gel was loaded with 124
µg of soluble protein and proteins of 55, 36.6 and 24 kDa affected by defoliation are indicated by arrows. The
position of isolectric points and molecular weight markers are indicated on the side of each gel.
relative insensitivity of roots to ‘ low N’ pretreatment
in this respect extended to trends in soluble protein
concentrations over the subsequent 14 d. These were
of broadly similar direction and magnitude in both
‘low N’ and ‘high N’ plants (Fig. 3). By contrast,
the concentrations of soluble protein in sheaths of
‘ low N’ ‘Aberelan’ and ‘Cariad’ increased pro-
gressively over the 14 d recovery period, whereas in

Printed from the CJO service for personal use only by...
N reserves and regrowth 459
25
So
lub
le p
rote
ins
(mg
g–1
d. w
t)
10
5
0
15
20
08 N
ov.
06 D
ec.
13 J
an.
20 F
eb.
20 M
ar.
22 A
pr.
22 M
ay
06 J
un
e
10 J
uly
1996–97
Mo
nth
ly m
ean
air
tem
per
atu
re (
°C)
Fig. 6. Seasonal changes in the concentration of total
soluble proteins (open squares) extracted from leaf sheaths
of Lolium perenne ‘Magella ’ grown under field conditions
and the monthly mean air temperature (filled circles).
Arrows indicate harvests. Vertical bars indicate³SE of the
mean for n¯3, when larger than the symbol.
‘high N’ plants they decreased until day 4, indicating
net hydrolysis, and then increased, the overall trend
being similar to for ‘Aberelan’ and ‘Cariad’.
The net hydrolysis of soluble protein in the
‘reserve’ tissues of ‘high N’ plants over the first 4 d
following defoliation was substantial. Concentra-
tions decreased by 32% in leaf sheath (Fig. 3a) and
by 15% in root tissue (Fig. 3b) of ‘Aberelan’ and by
47% in the sheath (Fig. 3c) and 25% in roots (Fig.
3d) of ‘Cariad’. The SDS–PAGE profiles of soluble
protein extracts of leaf sheath tissue from ‘Aberelan’
at different times after defoliation (Fig. 4) showed
that defoliation induced specific hydrolysis of several
polypeptides. In ‘high N’ plants (Fig. 4a) poly-
peptides with molecular weights of 55, 36.6 and 24
kDa, were evident at the time of defoliation and then
faded from the SDS–PAGE profile over the next 2–4
d, before reappearing and accumulating after day 7.
The same three polypeptides were present in lower
abundance in the tissues of ‘ low N’ ‘Aberelan’ on
the day of defoliation (Fig. 4b) and accumulated
between days 7 and 11 to concentrations above those
recorded on day 0.
Whilst the pattern of net hydrolysis followed by a
net accumulation of these particular polypeptides in
‘high N’ plants, together with their relative initial
scarcity in ‘ low N’ plants, suggested that they were
candidate VSPs, their silver staining intensities in
SDS–PAGE were low. Analysis of two-dimensional
gel profiles of soluble proteins extracted from sheath
tissue on day 0 (Fig. 5a) and day 4 after defoliation
(Fig. 5b) confirmed that these polypeptides were not
prominent in plants grown in FSC.
Under field conditions the concentrations of
soluble protein in the leaf sheath fraction (Fig. 6) in-
creased during autumn and winter, reaching a maxi-
mum during February and thereafter decreasing
31
14.5
21.5
08 N
ov.
06 D
ec.
13 J
an.
20 F
eb.
20 M
ar.
22 A
pr.
22 M
ay
10 J
uly
1996–97
45
kDa
55 kDa
36.6 kDa
24 kDa
Fig. 7. Seasonal changes in SDS–PAGE profile of soluble
proteins extracted from leaf sheaths of Lolium perenne‘Magella ’, grown under field conditions. Each well was
loaded with 10.23 µg of soluble protein, and the position of
molecular weight markers is indicated on each side. Arrows
indicate polypeptides of 55, 36.6, 24 kDa affected by
defoliation.
during Spring as growth resumed. The trend in the
concentrations of soluble protein during the year was
inversely related to the trend in monthly mean air
temperatures (Fig. 6), with a linear correlation
coefficient of determination of 0.88.
The 82% decrease in protein content of sheath
tissue between February and July, suggested a high
rate of protein hydrolysis. However, as the sheath
fraction was a composite of tissues with very different
ages, it is probable that much of this decline in
protein content was associated with senescence. The
presence of the same 55, 36.6 and 24 kDa poly-
peptides in plants grown in FSC was confirmed by
SDS–PAGE of proteins extracted from the field-
grown material (Fig. 7), but the profiles did not
provide convincing evidence of any seasonal pattern
in abundance.
Plant N status and the source of N for leaf growth
after defoliation
For numerous forage species (Ourry et al., 1989;
Richards, 1993; Thornton et al., 1993, 1994;
Thornton & Millard, 1996; Volenec et al., 1996),
mobilization of N compounds from tissues remain-
ing after severe defoliation plays an important role in
providing N for subsequent leaf growth. Whilst the

Printed from the CJO service for personal use only by...
460 S. Louahlia et al.
present results confirm this to be true for L. perenne,
more significantly they demonstrate for this species
that plant N status at the time of defoliation, which
is indicative of the potential availability of N reserves
for mobilization, is an important determinant of the
subsequent magnitude and dynamics of N
mobilization into the regrowing leaves. Further, they
point to the existence of substantial intra-specific
genotypic variation in the interactions between N
status, N mobilization and the rate of leaf growth
following defoliation. Decreased remobilization of N
in ‘low N’ plants was more acute in ‘Aberelan’ than
in ‘Cariad’ (Fig. 1). It is unlikely that this particular
difference was associated with variation in the
concentrations of tissue soluble carbohydrate be-
tween the two cultivars. With the notable exception
of fructans, there were only modest variations under
the present experimental conditions.
Low N status at the time of defoliation lessened the
extent to which mineral N uptake was subsequently
down-regulated (Fig. 2). This is consistent with
previous observations for L. perenne supplied solely
with NO$
− (Macduff et al., 1989). However, the
present results suggest that the extent of this N
status-related attenuation differs for the NH%
+ and
NO$
− uptake systems, and may also vary between
cultivars. Relative to the ‘high N’ plants, ‘ low N’
‘Aberelan’ exhibited significant NH%
+ and NO$
−
uptake during the first 4 d of recovery, whereas only
NO$
− uptake continued in ‘ low N’ ‘Cariad’ (Fig. 2).
The basis for these differences is unclear, but it is
tempting to regard the ability of ‘ low N’ ‘Aberelan’
to sustain NH%
+ uptake as having a direct bearing on
the higher dry weight of leaves of ‘ low N’ compared
with ‘high N’ plants of this cultivar (Fig. 1). The
capability to sustain uptake of NH%
+ following
defoliation presumably compensated the decrease in
N reserves resulting from ‘low N’ pretreatment
(Table 1).
The ability of L. perenne ‘Aberelan’, to com-
pensate for decreased availability of remobilizable N
by enhancing the rate of recovery in mineral N
uptake (Table 2), contrasts with that reported for L.
temulentum (Ourry et al., 1996) and M. sativa (Ourry
et al., 1994; Avice et al., 1997). In the latter species,
no such compensation was evident and the greater
delay in recovery of N uptake and N#
fixation may
account for their relatively slower refoliation, and
lower dry weight yields compared with L. perenne.
In this context it may be significant that L.
temulentum is an annual species, of little agronomic
significance and regarded as adapted to conditions of
relatively low N availability. On the other hand,
cultivars of L. perenne, such as those used in the
present study are usually the result of performance
selection under a relatively high plane of N nutrition.
These inter-specific differences in the ability to use
alternative sources of N following defoliation, would
appear to be consistent with the view that N storage
by grasses is a primary adaptation to environmental
conditions of low or fluctuating availability of N
(Thornton et al., 1993, 1994; Thornton & Millard,
1996); with N reserves constituting a buffer system
against periods of prolonged N starvation.
Are mobilization of N reserves and uptake of mineral
N coregulated following defoliation?
The generally inverse relationship between rates of
remobilization of stored N and uptake of mineral N
following defoliation stress, illustrated by the present
data, prompts several questions concerning how
these changes in internal N fluxes might be regu-
lated. Of particular interest is whether or not N
mobilization and N uptake are subject to a common
set of regulatory signals and, if so, how these up-
regulate mobilization almost coincidentally with the
down-regulation of N uptake following defoliation.
A definitive answer to these questions requires far
more detailed study of N dynamics during the first
24 h after defoliation than is reported here. However,
there is sufficient relevant information to suggest the
following crude ‘supply and demand’-based model
may apply at the whole plant level. Severe defoliation
has the immediate effect of removing a large
proportion of the plant’s ‘sink capacity’ for N
assimilates. A sudden perturbation in ‘demand’
would be expected to cause a rapid increase in
concentrations of amino acids in the root system,
and}or in the flux of amino acids cycling through the
remaining vascular system. Given that NH%
+ and
NO$
−uptake are considered to be subject to various
forms of negative feedback regulation, by amino
acids amongst other effectors (Ourry et al., 1997),
this would explain the very rapid down-regulation of
N uptake following defoliation (Macduff & Jackson,
1992). The down-regulation of uptake may then be
reinforced and sustained by the amino acid flux from
hydrolysis of reserve proteins in the roots and sheath
tissue, until such time as the net consumption rate of
amino acids for protein synthesis in new leaf tissue
exceeds their mobilization-related production in the
source tissues.
According to this scheme, the attenuation, or
absence, of down-regulation of N uptake in ‘ low N’
status plants would occur initially because there is a
lower (or zero) increase in amino acid concentrations
following removal of ‘sink’ tissues. This would be a
consequence of the prevailing shortfall in the uptake
of N relative to assimilatory N demand for tissue
synthesis. Subsequently, as a consequence of the
smaller N reserve pool in ‘ low N’ plants, there
would be a lower rate of amino acid production from
protein hydrolysis and other N mobilization pro-
cesses and, more speculatively, a much shorter
residence time for these amino acids at sites of
regulatory action on the N uptake systems. This
model could also explain the apparent over-com-

Printed from the CJO service for personal use only by...
N reserves and regrowth 461
pensation observed in ‘ low N’ ‘Aberelan’ (Fig. 1).
However, it does not account for the dramatic up-
regulation of N mobilization from source tissues
following defoliation, the exact timing of which in
relation to the initiation of the decline in mineral N
uptake remains uncertain. Intuitively it is unlikely
that N remobilization is stimulated directly by an
‘amino acid surge’ in the vascular system of the
plant following defoliation, as anticipated in the
model outlined: other signalling systems uncon-
nected with plant N status are more probable in our
opinion (e.g. aspects of soluble carbohydrate status).
Moreover, there are grounds for expecting the
opposite effect. For example, a temporary increase
in plant N status, denoted by increased vascular
amino acid concentrations, might down-regulate N
mobilization if there is no immediate requirement
for the resulting amino acids associated with tissue
synthesis. It is significant that apparent ‘antagonism’
between N uptake and mobilization of N stores is a
widespread phenomenon in the plant kingdom
(Imsande & Touraine, 1994), and is of particular
significance at certain ontogenetic stages, such as
seed setting.
There are clearly various other hypothetical
explanations for aspects of the N dynamics observed
following defoliation of grasses. The down-regu-
lation of N uptake per se might trigger the
mobilization of N from reserves: the plant in effect
mistaking the ensuing post-defoliation stress for a
shortfall in external N supply that requires tem-
porary buffering from N reserves. Another possible
explanation for the continuation of mineral N uptake
by ‘low N’ plants, at least in the short-term following
defoliation, invokes the ‘N demand’ constituted by
replenishment of the vacuolar nitrate pool, pre-
viously depleted during ‘low N’ pretreatment
(Macduff et al., 1989). However, it is less straight-
forward to explain continued uptake of NH%
+ by this
mechanism.
Are VSPs involved in the storage and remobilization
of N in perennial ryegrass?
VSPs have been characterized in two species of
forage legumes: M. sativa (Hendershot & Volenec,
1993) and T. repens (Corre et al., 1996). In both
species they account for at least 20% of total soluble
protein, in taproots and stolons. They have at least
two functions, which are usually separated in time:
the provision of reduced N for early regrowth
following defoliation, and also for spring growth. As
far as we are aware, functional VSPs have not been
reported in grasses such as L. perenne. The three
polypeptides (of 55, 36.6 and 24 kDa) identified in
the present study (Figs. 4, 5) satisfied some of the
criteria for classification as VSPs. They were nearly
fully hydrolysed during early regrowth and accumu-
lated in tissue during autumn and winter. Moreover,
under conditions of ‘ low N’ supply (Fig. 4b) they
did not accumulate to the same extent as in ‘high N’
plants (Fig. 4a), suggesting that their synthetic rate is
somehow coupled with plant N status. Taken
together, these characteristics might suggest that
they have a functional role as VSPs. However, they
were not found to be prominent polypeptides
(accounting for !8% of total silver staining in-
tensity), and accordingly do not satisfy all of the
criteria for VSP status as prescribed by Staswick
(1989, 1990, 1994). It is possible, but unlikely, that
these polypeptides belong to the same hetero-
polymer, in which case their VSP status would be
less doubtful. Similar conclusions were reached by
other workers searching for VSPs in M. officinalis
and T. pratense (Cunningham & Volenec, 1996; Li et
al., 1996).
This work was partly funded through an INRA}BBSRC
grant to A. O. The authors thank Mrs Micheline Meyer
(ATOS, University of Caen, France) for her invaluable
help with electrophoresis, Dr Terence N. Ledger for
valuable discussions and improvement of the manuscript
and Mr D. Perrin for his efficient collaboration for field
work.
Avice JC, Ourry A, Lemaire G, Volenec JJ, Boucaud J. 1997.Root proteins and VSP are key organic nutrients for alfalfa
shoot regrowth. Crop Science 37 : 1187–1193.
Bouchart V, Macduff JH, Ourry A, Svenning MM, Gay AP,Simon JC, Boucaud J. 1998. Seasonal pattern of accumulation
and effects of low temperatures on storage compounds in
Trifolium repens. Physiologia Plantarum 104 : 65–74.
Clement CR, Hopper MJ, Canaway RJ, Jones LPH. 1974. A
system for measuring the uptake of ions by plants from
solutions of controlled composition. Journal of Experimental
Botany 25 : 81–99.
Clement CR, Hopper MJ, Jones LHP. 1978. The uptake of
nitrate by Lolium perenne L. from flowing nutrient solution. I.
Effect of NO$
− concentration. Journal of Experimental Botany
37 : 589–596.
Corre N, Bouchart V, Ourry A, Boucaud J. 1996. Mobilization
of nitrogen reserves during regrowth of defoliated Trifolium
repens L. and identification of potential vegetative storage
proteins. Journal of Experimental Botany 301 : 1111–1118.
Cunningham SM, Volenec JJ. 1996. Purification and charac-
terization of alfalfa taproot vegetative storage proteins. Journal
of Plant Physiology 147 : 625–632.
Evans DR, Humphreys MO, Williams TA. 1996. Forage yield
and quality interactions between white clover and contrasting
ryegrass varieties in grazed swards. Journal of Agricultural
Science, Cambridge 126 : 295–299.
Hatch DJ, Hopper MJH, Dhanoa MS. 1986. Measurement of
ammonium ions in flowing solution culture and diurnal
variation in uptake in Lolium perenne. Journal of Experimental
Botany 37 : 589–596.
Hatch DJ, Macduff JH. 1991. Concurrent rates of N#
fixation,
nitrate and ammonium uptake by white clover in response to
regrowth at different root temperatures. Annals of Botany 67 :
265–274.
Hendershot KL, Volenec JJ. 1993. Taproot nitrogen accumu-
lation and use in overwintering alfalfa (Medicago sativa L.).
Journal of Plant Physiology 141 : 68–74.
Imsande J, Touraine B. 1994. N demand and the regulation of
nitrate uptake. Plant Physiology 105 : 3–7.

Printed from the CJO service for personal use only by...
462 S. Louahlia et al.
Laemmli UK. 1970. Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227 : 680–685.
Li R, Volenec JJ, Joern BC, Cunningham SM. 1996. Seasonal
changes in nonstructural carbohydrates, protein, and macro-
nutrients in roots of alfalfa, redclover, sweetclover and birdfoot
trefoil. Crop Science 36 : 617–623.
Lopez MF, Patton WF, Utterback BL, Chung-Welch N,
Barry P, Skea WM, Cambria RP. 1991. Effect of various
detergents on protein migration in the second dimension of two
dimension gels. Annals of Biochemistry 199 : 35–44.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. 1951.
Protein measurement with the folin phenol reagent. Journal of
Biological Chemistry 193 : 265–275.
Macduff JH, Jackson SB. 1992. Influx and efflux of nitrate and
ammonium in Italian ryegrass and white clover roots : com-
parisons between effects of darkness and defoliation. Journal of
Experimental Botany 249 : 525–535.
Macduff JH, Jarvis SC, Mosquera A. 1989. Nitrate nutrition of
grasses from steady-state supplies in flowing solution culture
following nitrate deprivation and}or defoliation. II. Assimi-
lation of NO$
− and short-term effects on NO$
− uptake. Journal
of Experimental Botany 40 : 977–984.
Millard P. 1988. The accumulation and storage of nitrogen by
herbaceous plants. Plant, Cell and Environment 11 : 1–8.
O’Farrell PH. 1975. High resolution two-dimensional electro-
phoresis of proteins. Journal of Biological Chemistry 250 :
4007–4021.
Ourry A, Bigot J, Boucaud J. 1989. Protein mobilization from
stubble and roots and proteolytic activities during post-clipping
re-growth of perennial ryegrass. Journal of Plant Physiology
134 : 298–303.
Ourry A, Kim TH, Boucaud J. 1994. Nitrogen reserve
mobilization during regrowth of Medicago sativa L.:
relationships between their availability and regrowth yield.
Plant Physiology 105 : 831–837.
Ourry A, Gordon AJ, Macduff JH. 1997. Nitrogen uptake and
assimilation in roots and root nodules. In: Foyer CH, Quick
WP, eds. A molecular approach to primary metabolism in higher
plants. London, UK: Taylor and Francis, 237–253.
Ourry A, Macduff JH, Ougham HJ. 1996. The relationship
between mobilization of N reserves and changes in translatable
messages following defoliation in Lolium temulentum L. and
Lolium perenne L. Journal of Experimental Botany 299 : 739–747.
Phillips DA, Center DM, Jones MB. 1983. Nitrogen turnover
and assimilation during regrowth in Trifolium subterraneum L.
and Bromus mollis L. Plant Physiology 71 : 472–476.
Radojevic I, Simpson RJ, St John JA, Humphreys MO. 1994.Chemical composition and in vitro digestibility of lines of
Lolium perenne selected for high concentrations of water-soluble
carbohydrate. Australian Journal of Agriculture Research 45901–12.
Richards JH. 1993. Physiology of plants recovering from
defoliation. Proceedings of the XVII International Grassland.
Congress, Palmerston, New Zealand, 85–94.
Smouter H, Simpson RJ. 1991. Fructan metabolism in leaves of
Lolium rigidum Gaudin. I. Synthesis of fructan. New Phytologist
119 : 509–516.
Staswick PE. 1989. Developmental regulation and the influence
of plant sinks on vegetative storage protein gene expression in
soybean leaves. Plant Physiology 89 : 309–315.
Staswick PE. 1990. Novel regulation of vegetative storage protein
genes. The Plant Cell 2 : 1–6.
Staswick PE. 1994. Storage proteins of vegetative plant tissues.
Annual Review of Plant Physiology and Plant Molecular Biology
45 : 303–322.
Ste! pien V, Sauter JJ, Martin F. 1994. Vegetative storage
proteins in woody plants. Plant Physiology and Biochemistry 32 :
185–192.
Thornton B, Millard P. 1996. Effects of severity of defoliation on
root functioning in grasses. Journal of Range Management 49 :
443–447.
Thornton B, Millard P, Duff EI. 1994. Effects of nitrogen supply
on the source of nitrogen used for regrowth of laminae after
defoliation of four grass species. New Phytologist 128 : 615–620.
Thornton B, Millard P, Duff EI, Buckland ST. 1993. The
relative contribution of remobilization and root uptake in
supplying nitrogen after defoliation for regrowth of laminae in
four grass species. New Phytologist 124 : 689–694.
Volenec JJ, Ourry A, Joern BC. 1996. A role for nitrogen
reserves in forage regrowth and stress tolerance. Physiologia
Plantarum 97 : 185–193.
Wild A, Jones LHP, Macduff JH. 1987. Uptake of mineral
nutrients and crop growth: the use of flowing nutrient solutions.
Advances in Agronomy 41 : 171–219.
Wilkins PW, Macduff JH, Raistrick N, Collison M. 1997.Varietal differences in perennial ryegrass for nitrogen use
efficiency in leaf growth following defoliation: performance in
flowing solution culture and its relationship to yield under
simulated grazing in the field. Euphytica 98 : 109–119.




















