
Nitrogen and crop rotation effects on fusarium crown rot in no-till spring wheat
12
Soilborne pathogens / Agents pathogènes telluriques Nitrogen and crop rotation effects on fusarium crown rot in no-till spring wheat Ryan A. Davis, David R. Huggins, R. James Cook, and Timothy C. Paulitz Abstract: Fusarium crown rot of wheat (Triticum aestivum), caused by Fusarium pseudograminearum and Fusarium culmorum, is a yield-limiting disease in the dryland wheat-production area of the intermountain Pacific Northwest and is exacerbated in water-stressed plants induced by overfertilizing with nitrogen (N). Plants with excess N deplete water from the soil profile more rapidly and become drought stressed prematurely. Traditionally a problem on winter wheat in summer fallow, this disease has become more important for spring wheat in continuous cropping areas managed for high grain protein levels. During 3 years with direct seeding (no till) near Pullman, Washington, we investigated whether a split application of N, with some applied the previous fall and some with planting, could limit the disease compared with all N applied in the spring and with no N as the check. We also investigated the influence of the previous (rotation) crop (winter and spring canola, Brassica rapa; barley, Hordeum vulgare; or peas, Pisum sativum) on disease, grain yield, grain protein concentration, and populations of Fusarium in the soil. Overall, the DNA concentration of F. culmorum was significantly greater than F. pseudograminearum, and F. culmorum was highest following spring barley. Disease severity and yield were consistently lower in the no-N treatments compared with the other N treatments. The split application reduced disease in only 1 of 3 years. The all-spring application resulted in higher grain protein in 2 of 3 years compared with the split application, but yield was not affected. The previous crop had small but significant effects on disease, but they were not consistent from year to year and often interacted with the N treatment. Grain protein was higher in wheat after pea in 2 of 3 years. In conclusion, splitting of N had little effect on fusarium crown rot, probably because the N level in both treatments was conducive for disease development. Even if not a host species, the previous crop had little effect on subsequent disease, probably because Fusarium persists for more than one season as chlamydospores and in crop residue in this dry summer climate. Key words: fusarium crown rot, no-till, Triticum aestivum, Fusarium pseudograminearum, F. culmorum, rotation crops, pea, barley, canola, nitrogen. 467 Résumé : La pourriture du collet chez le blé (Triticum aestivum), causée par Fusarium pseudograminearum et Fusarium culmorum, une maladie qui limite les rendements dans les régions où l’on produit du blé sur les terrains arides intramontagneux du Pacific Northwest états-unien, est aggravée chez les plants souffrant de stress hydrique engendré par la fertilization excessive à l’azote. Les plants contenant un excès d’azote épuisent le sol de son eau plus rapidement et sont prématurément sujets au stress causé par la sécheresse. Problème rencontré traditionnellement chez le blé d’hiver semé sur jachère d’été, cette maladie s’attaque de plus en plus au blé de printemps dans les régions où l’on pratique la culture continue et où l’on vise de hautes teneurs en protéines. Durant trois années, près de Pullman dans l’État de Washington, en appliquant la méthode de culture sans travail du sol (semis direct), nous avons voulu vérifier si une application fractionnée d’azote, dont une partie était appliquée à l’automne précédent et une autre lors du semis, limiterait l’incidence de la maladie, comparativement à une seule application printanière, le tout par rapport à une parcelle témoin non fertilizée. Nous avons également étudié l’effet des cultures (rotations) précédentes (canola d’hiver et de printemps, Brassica napus; orge, Hordeum vulgare; ou pois, Pisum sativum) sur la maladie, le rendement en grains, la concentration des protéines dans les grains et les populations de Fusarium dans le sol. Dans l’ensemble, la concentration de l’ADN du Fusarium culmorum était plus élevée postérieurement à la culture de l’orge de Can. J. Plant Pathol. 31: 456–467 (2009) 456 Accepted 1 August 2009. R.A. Davis. Department of Crop and Soil Sciences, Washington State University, Pullman, WA 99164-6430, USA. D.R. Huggins. USDA Agricultural Research Service, Land Management and Water Conservation Research Unit, Washington State University, Pullman, WA 99164-6421, USA. R.J. Cook. Department of Plant Pathology, Washington State University, Pullman, WA 99164-6430, USA. T.C. Paulitz. 1 USDA Agricultural Research Service, Root Disease and Biological Control Research Unit, Washington State University, Pullman, WA 99164-6430, USA. 1 Corresponding author (e-mail: [email protected]).
Transcript of Nitrogen and crop rotation effects on fusarium crown rot in no-till spring wheat

Soilborne pathogens / Agents pathogènes telluriques
Nitrogen and crop rotation effects on fusariumcrown rot in no-till spring wheat
Ryan A. Davis, David R. Huggins, R. James Cook, and Timothy C. Paulitz
Abstract: Fusarium crown rot of wheat (Triticum aestivum), caused by Fusarium pseudograminearum and Fusariumculmorum, is a yield-limiting disease in the dryland wheat-production area of the intermountain Pacific Northwest andis exacerbated in water-stressed plants induced by overfertilizing with nitrogen (N). Plants with excess N deplete waterfrom the soil profile more rapidly and become drought stressed prematurely. Traditionally a problem on winter wheatin summer fallow, this disease has become more important for spring wheat in continuous cropping areas managed forhigh grain protein levels. During 3 years with direct seeding (no till) near Pullman, Washington, we investigatedwhether a split application of N, with some applied the previous fall and some with planting, could limit the diseasecompared with all N applied in the spring and with no N as the check. We also investigated the influence of theprevious (rotation) crop (winter and spring canola, Brassica rapa; barley, Hordeum vulgare; or peas, Pisum sativum) ondisease, grain yield, grain protein concentration, and populations of Fusarium in the soil. Overall, the DNAconcentration of F. culmorum was significantly greater than F. pseudograminearum, and F. culmorum was highestfollowing spring barley. Disease severity and yield were consistently lower in the no-N treatments compared with theother N treatments. The split application reduced disease in only 1 of 3 years. The all-spring application resulted inhigher grain protein in 2 of 3 years compared with the split application, but yield was not affected. The previous crophad small but significant effects on disease, but they were not consistent from year to year and often interacted withthe N treatment. Grain protein was higher in wheat after pea in 2 of 3 years. In conclusion, splitting of N had littleeffect on fusarium crown rot, probably because the N level in both treatments was conducive for disease development.Even if not a host species, the previous crop had little effect on subsequent disease, probably because Fusariumpersists for more than one season as chlamydospores and in crop residue in this dry summer climate.
Key words: fusarium crown rot, no-till, Triticum aestivum, Fusarium pseudograminearum, F. culmorum, rotation crops,pea, barley, canola, nitrogen.
467Résumé : La pourriture du collet chez le blé (Triticum aestivum), causée par Fusarium pseudograminearum etFusarium culmorum, une maladie qui limite les rendements dans les régions où l’on produit du blé sur les terrainsarides intramontagneux du Pacific Northwest états-unien, est aggravée chez les plants souffrant de stress hydriqueengendré par la fertilization excessive à l’azote. Les plants contenant un excès d’azote épuisent le sol de son eau plusrapidement et sont prématurément sujets au stress causé par la sécheresse. Problème rencontré traditionnellement chezle blé d’hiver semé sur jachère d’été, cette maladie s’attaque de plus en plus au blé de printemps dans les régions oùl’on pratique la culture continue et où l’on vise de hautes teneurs en protéines. Durant trois années, près de Pullmandans l’État de Washington, en appliquant la méthode de culture sans travail du sol (semis direct), nous avons vouluvérifier si une application fractionnée d’azote, dont une partie était appliquée à l’automne précédent et une autre lorsdu semis, limiterait l’incidence de la maladie, comparativement à une seule application printanière, le tout par rapport àune parcelle témoin non fertilizée. Nous avons également étudié l’effet des cultures (rotations) précédentes (canolad’hiver et de printemps, Brassica napus; orge, Hordeum vulgare; ou pois, Pisum sativum) sur la maladie, le rendementen grains, la concentration des protéines dans les grains et les populations de Fusarium dans le sol. Dans l’ensemble,la concentration de l’ADN du Fusarium culmorum était plus élevée postérieurement à la culture de l’orge de
Can. J. Plant Pathol. 31: 456–467 (2009)
456
Accepted 1 August 2009.
R.A. Davis. Department of Crop and Soil Sciences, Washington State University, Pullman, WA 99164-6430, USA.D.R. Huggins. USDA Agricultural Research Service, Land Management and Water Conservation Research Unit, Washington StateUniversity, Pullman, WA 99164-6421, USA.R.J. Cook. Department of Plant Pathology, Washington State University, Pullman, WA 99164-6430, USA.T.C. Paulitz.1 USDA Agricultural Research Service, Root Disease and Biological Control Research Unit, Washington StateUniversity, Pullman, WA 99164-6430, USA.
1Corresponding author (e-mail: [email protected]).

printemps. La gravité de la maladie et le rendement étaient invariablement plus faibles dans la parcelle témoin sansazote que dans celles qui en avaient reçu. L’application fractionnée a permis de réduire l’incidence de la maladiedurant une année seulement sur trois. Comparativement à l’application fractionnée, l’application printanière a engendréune plus haute teneur en protéine deux années sur trois, mais n’a pas influencé le rendement. Les cultures précédentesont eu des effets minimes mais notables sur la maladie, cependant ils variaient d’une année à l’autre et interagissaientsouvent avec le traitement à l’azote. La teneur en protéines des grains chez le blé était plus élevée à la suite de laculture des pois, et ce, deux années sur trois. Pour conclure, l’application fractionnée d’azote a eu peu d’effets sur lapourriture du collet, probablement parce que les niveaux d’azote, lors des deux applications, étaient propices audéveloppement de la maladie. La culture précédente, même si elle n’était pas hôte, avait peu d’effets sur la maladie àvenir, probablement parce que, dans ces conditions estivales arides, Fusarium subsiste pour plus d’une saison sousforme de chlamydospores et dans les résidus de culture.
Mots-clés : pouriture fusarienne du colet, semis direct, Triticum aestivum, Fusarium pseudograminearum, F. culmorum,cultures d'assolement, pois, orge, canola, azote.
Davis et al.: Fusarium crown rot on wheat / effect of nitrogen andcrop rotationIntroduction
Fusarium crown rot, also know as dryland foot rot orfusarium root rot, is primarily caused by Fusariumpseudograminearum O’Donnell & Aoki (previously knownas Fusarium graminearum group I) and Fusariumculmorum (W.G. Sm.) Sacc. (Cook 1980; Smiley andPatterson 1996). The disease is commonly found in thedryland wheat-production areas of the Pacific Northwest(PNW) (Cook 1968, 1980, Paulitz et al. 2002), Australia(Backhouse et al. 2004; Wildermuth et al. 1997; Smiley etal. 2005a), and the Middle East (Bentley et al. 2006; Foulyet al. 1996). Barley (Hordeum vulgare L.) can also be ahost. In a survey of 13 fields in northeastern Oregon,fusarium crown rot caused a mean loss of 9.5% ($51/ha,based on a price of $3.85–$4.50/bushel; 1 bushel =0.035 m3), but losses were as high as 35% (1550 kg/ha) insome fields (Smiley et al. 2005b). These pathogens infectroots, subcrown internodes, and crowns and spread upwardinto one or more stem internodes when plants are waterstressed and exhibit low water potentials (Cook 1981). Un-der conditions favorable for severe disease, whiteheads ap-pear shortly before heads (spikes) mature, resulting in loweryields and shriveled-grain test weights (Wiese 1987). Underirrigated, rainy, or high-humidity conditions, these patho-gens can also infect the florets and cause head blight andscab, but this is very rare in the PNW (T.C. Paulitz, per-sonal observation). Surveys in the PNW during 1993–1994established that both pathogens often occurred in the samefield; however, F. pseudograminearum dominated the warm-est, driest parts of the region, whereas F. culmorum tendedto dominate in areas of slightly higher rainfall and coldersummer temperatures (Smiley and Patterson 1996).
Current management strategies for fusarium crown rotcontrol attempt to limit or avoid plant water stress by (i) de-laying fall planting of winter wheat (Triticum aestivum L.)in a wheat – summer fallow system, thereby limiting earlycrop growth and the premature consumption and depletionof water supplies prior to the linear phase of crop growththe following year, and (ii) avoiding excess N fertilizationrelative to the soil water supply, which promotes biomassproduction before anthesis but can lead to early depletion ofstored soil water and greater postanthesis water stress(Cook and Papendick 1972; Papendick and Cook 1974).
Two major changes in cropping systems have developedin the PNW within the last 10–15 years, with profound im-plications for the epidemiology of this disease: (i) an in-crease in the use of no-till practices and (ii) intensificationof cropping systems through greater use of spring cereals inplace of more traditional summer fallow in rotation withwinter wheat.
The no-till practice entails seeding the crop directly intothe undisturbed residue of the previous crop, typically withprecision placement of seed and fertilizer. Soil loss fromwind and water erosion is significantly reduced (Papendick2004) as are fuel, labor, and machine costs (Baker et al.2004). Conventional tillage in eastern Washington may re-quire seven or more tillage or cultivation operations in asingle season compared with one to three passes in no till toapply herbicides and achieve similar goals. Soil organicmatter often increases without tillage (Kennedy andSchillinger 2006; Purakayastha et al. 2008) as comparedwith conventional tillage and is coupled with improvementsin soil structure and water conservation. The no-till practiceleaves most of the Fusarium-infested stem and crown tissueat or near the soil surface where residue decomposition isoften slower, thereby leaving Fusarium inoculum sourcesideally positioned to infect the crowns of the followingcrop. The percentage of infection and severity of disease hasgenerally been greater without tillage compared with conven-tional seeding in New South Wales, Australia (Burgess et al.1993); Saskatchewan, Canada (Bailey et al. 2001); and north-eastern Oregon (Smiley and Patterson 1996).
Over the last 30 years, spring wheat production in thePNW has increased from formerly 5% to now 20% of thedryland wheat area. This increasing production base has re-sulted from (i) development of higher yielding spring wheatvarieties, (ii) increased cropping system intensificationthrough substitution of spring wheat for summer fallow inwinter wheat – fallow sequences (Cook 1980), and(iii) greater economic viability compared with other alterna-tive rotational crops such as pulses or oilseeds in areassuited to annual cropping. Concomitantly, many growers areplanting hard red (bread type) spring wheat varieties insteadof the soft white winter (pastry type) varieties that havebeen traditionally grown. Hard red wheat varieties withgrain protein levels of 12%–14% have historically com-manded a premium over soft white wheat with 9%–10%
Davis et al.: Fusarium crown rot on wheat / effect of nitrogen and crop rotation 457

grain protein; however, mean yields are typically lower(Baker et al. 2004; Janosky 1999). Nitrogen requirementsfor hard red are 50% greater than for soft white spring wheat(Koenig 2005), and the additional N applied to obtain grainprotein goals could also increase fusarium crown rot as hasbeen previously reported for winter wheat (Papendick andCook 1974).
Two management strategies may reduce or limit the se-verity of fusarium crown rot in hard red spring wheat man-aged with high N fertilizer inputs. Firstly, the Nrequirement could be split with up to 50% applied in thefall and the remainder supplied during spring planting, in-stead of supplying the total N requirement in the spring.Secondly, crop rotations could include a 1 year break to anonhost crop followed by spring wheat and then winterwheat (a 3 year rotation but with two wheat crops every3 years).
The agronomic advantage of the split fall–spring com-pared with all-spring application of N is that fall-applied Ncould be moved lower in the soil profile during the winterwhen most of the precipitation occurs and be favorably po-sitioned for crop uptake during heading and grain fill,whereas N added in the spring would be available in the up-per soil profile in support of tillering and jointing phases ofplant development (Cook and Veseth 1991; Huggins 1991).In contrast, supplying the entire N requirement in the springat planting could promote excessive (lush) vegetativegrowth and early use of stored soil water, thereby predis-posing the crop to more severe postanthesis water stress anddisease. Our hypothesis is that a change in the temporalavailability of N as achieved with a split fall–spring applica-tion might limit the development of fusarium crown rot.
Eastern Washington and adjacent northern Idaho andnortheastern Oregon are suited almost exclusively tocool-season crops, limiting the options for economically vi-able rotations. Rotation breaks to broadleaf crops arelargely limited to cool-season pulses and brassicas, whichare usually grown prior to winter wheat to provide thegreatest rotation benefits. We suggest a deviation from thisrotation strategy by following the break crop with springwheat and subsequently winter wheat, thereby increasingthe rotation benefits for reducing fusarium crown rot inspring wheat. Spring and winter barley could also be usedas cereal break crops between winter and spring wheat inthis 3 year rotation scenario. Although an 18 month breakbetween the winter and spring wheat would not be expectedto reduce the inoculum potential of F. culmorum because ofthe long half-life of the chlamydospores of this pathogen inPalouse silt loams (Inglis and Cook 1986), lower inoculumlevels could occur for the more aggressive but shorter livedF. pseudograminearum (Sitton and Cook 1981) and result inlower incidence of disease. For example, Smiley et al.(1996) showed that, for northeastern Oregon, fusariumcrown rot was more severe in annual wheat compared witha wheat–pea (Pisum satvium L.) rotation.
The objective of this research was to examine how croprotation and timing of N application could influencefusarium crown rot disease, inoculum of the pathogen, andgrain yield and protein.
Materials and methods
Site characteristics and cultural practicesThe study was carried out on the Washington State Uni-
versity Cook Agronomy Farm (CAF) dedicated to no-tillcropping systems and precision-farming research. The 57 haCAF is located 10 km northeast of Pullman, Washington, at117.08°W, 46.78°N (Fig. 1A). The CAF was established asone of the university’s research farms in 1999 and ranges inelevation from 771 to 823 m, typical of the rollingloess-derived hills of the Palouse region. Silt loam soils ofthe Palouse–Naff–Thatuna association are predominantacross the CAF. In 1999, a nonaligned systematic grid of369 georeferenced locations was established using a sur-vey-grade global positioning system over a 37 ha portion ofthe southwestern part of the CAF, where this study was sub-sequently conducted. The site was characterized by sam-pling the established georeferenced locations for variousbiophysical properties, including soil morphology, pH, or-ganic matter, weed seed bank, and soilborne pathogens. The37 ha site was direct seeded with ‘WPB926’ hard red springwheat in April 1999, harvested and then direct-seeded en-tirely to ‘Baronesse’ spring barley in April 2000. Followingharvest in 2000, the 37 ha field was divided into three main12.2 ha fields (fields A, B, and C) to establish multiple3 year crop rotation strategies (Fig. 1B). Fields A, B, and Cwere further subdivided into six strips of approximately2 ha each to initiate six alternative rotations consisting ofwinter wheat – X – spring wheat, where X represents six al-ternative crops, namely the winter and spring counterpartsof barley, pea, and canola (Brassica napus L.). The stripswere aligned to cross over and include the typical field vari-ations in terrain and soil. In the 2000–2001 season, field Awas direct-seeded to the six alternative crops; field B, uni-formly to hard red spring wheat; and field C, uniformly tohard red winter wheat. Thereafter, field A was planted tohard red spring wheat followed by hard red winter wheatand then back to the alternative crops; field B was plantedto hard red winter wheat, followed by the alternative cropsand then back to hard red spring wheat; and field C wasplanted to the six alternative crops followed by hard redspring wheat and then back to hard red winter wheat. Infor-mation on the crops and years planted is given in Table 1.
Spring wheat was direct-seeded (without tillage) with a5 m wide Great Plains drill equipped with leading flutedTurbo coulters spaced 25 cm apart that enabled deep place-ment (10 cm) of liquid N, P, and S followed by double-diskopeners also spaced 25 cm apart and positioned to placeseed directly above the fertilizer band. Starting in 2001,‘Hank’ hard red spring wheat was seeded at 134 kg/ha at adepth of 1 cm to 3 cm.
Weed control was accomplished with preplanting ap-plications of glyphosate for all crops in all years. A mix-ture of thifensulfuron-methyl and tribenuron-methyl wasused for broadleaf control in all wheat crops. In2001–2002, grassy weeds were controlled in winterwheat with tribenuron-methyl and tralkoxydim. Forspring wheat grassy weed control, clodinafop-propargyland tribenuron-methyl were used. For winter and springwheat in 2002–2003, grassy weeds were controlled withclodinafop-propargyl. In 2003–2004, grassy weeds were
458 Can. J. Plant Pathol. Vol. 31, 2009

controlled with mesosulfuron methyl and flucarbazonefor winter and spring wheat, respectively. In peas for allyears, grassy weeds were controlled with pendimethalinand imazethapyr. All canola crops were Roundup Ready,and glyphosate was used for weed control.
Nitrogen fertilizer treatmentsIn all years, the split fall–spring and all-spring applied N
treatments were located as side by side substrips in each ofthe six alternative crop sequence strips of either field A, B,or C. Strips were 5 m wide for the length of the field, whichvaried between 500 and 700 m. In addition, control plots(5 m × 20 m) with no applied N were established in se-
lected landscape positions within at least three of the sixstrips with previous crop of either canola, pea, or barley.No distinction was made in selection of control plots as towhether the previous crop was a winter or spring counter-part of the alternative crop. Instead, plot locations were se-lected to allow N treatment comparisons at locations withequivalent soil and terrain properties. The control plotswere used in conjunction with the neighboring N-timingtreatments for comparison of split fall–spring applied N,all-spring applied N, or no applied N. In 2001–2002, the Ntiming treatments with control plots were located on field Astrips with previous crops of spring canola, spring barley,and spring pea as these provided the most similar compari-
Davis et al.: Fusarium crown rot on wheat / effect of nitrogen and crop rotation 459
Cropa Planting datebSeeding rate(kg/ha)c
Fertilizer rate(kg/ha; N:P:K)
2000–2001Winter wheat SW ‘Madsen’ 20–24 Oct. 2000 112 196:22:17Spring wheat HR ‘Hank’ 24 Mar. 2001 112 157:22:35Winter barley ‘Hundred’ 19 Oct. 2000 100 157:22:17Spring barley ‘Baronesse’ 26 Mar. 2001 103 110:22:16Winter canola ‘Erica’ (‘Ryder’) 9, 26 Sept. 2000 (26 Mar. 2001) 6.2, 9.0 (1.1) 90:0:13 on
31 Oct. 2000Spring canola ‘Ryder’ 26 Mar. 2001 9.0 110:22:16Winter pea ‘Granger’ 28 Sept. 2000 112 NoneSpring pea ‘Karita’ 24 Apr. 2001 224 None2001–2002Winter wheat HR ‘Falcon’ 23 Oct. 2001 112 224:24:24Spring wheat HR ‘Hank’ 13–15 Apr. 2002 134 213:22:34Winter barley ‘Strider’ 26 Oct. 2001 112 179:22:22Spring barley ‘Baronesse’ 21 Apr. 2002 110 146:22:34Winter canola ‘Athena’ 25 Sept. 2001 9 62:28:28Spring canola ‘Hyola 357’ 21 Apr. 2002 11 146:22:28Winter pea ‘Nutrigreen’ 9 Oct. 2001 202 NoneSpring pea ‘Cruiser’ 2 June 2002 220 None2002–2003Winter wheat HR ‘Falcon’ 18 Oct. 2002 90 202:22:22Spring wheat HR ‘Hank’ 23 Apr. 2003 134 224:28:28Winter barley ‘Sun Star Pride’ 17 Oct. 2002 134 168:22:22Spring barley ‘Baronesse’ 5 May 2003 110 157:22:22Winter canola ‘Athena’ (‘Hyola 357’) 12 Sept. 2002 (14 Apr. 2003) 11 (11) 142:28:28Spring canola ‘Hyola 357’ 22 Apr. 2003 9 146:22:34Winter pea ‘Nutrigreen’ (‘Cruiser’) 10 Oct. 2002 (3 May 2003) 134 (246) None (none)Spring pea ‘Cruiser’ 6 May 2003 246 None2003–2004Winter wheat HR ‘Falcon’ 30 Sept. 2003 92 213:34:28Spring wheat SW cv. Hank’ 3 Apr. 2004 134 213:11:28Winter barley ‘Sun Star Pride’ 30 Sept. 2003 134 168:22:28Spring barley ‘Baronesse’ 9 Apr. 2004 112 123:11:18Spring canola ‘Dekalb 223’d 8 Apr. 2004 9 125:0:0Spring canola ‘Dekalb 223’ 12 Apr. 2004 7 123:11:28Winter pea ‘90105-9’ 29 Sept. 2003 170 NoneSpring pea ‘Cruiser’ 27 Apr. 2004 246 None
aWhen winter canola stands froze or did not survive the winter, plots were reseeded with the broadcasted spring canola cultivar given inparentheses.SW, soft white; HR, hard red.
bDates in parentheses are the dates that the spring canola cultivars were planted.cValues in parentheses are the seeding rates for the spring canola cultivars.d‘Dekalb 223’ is Roundup Ready.
Table 1. Summary of cropping inputs.
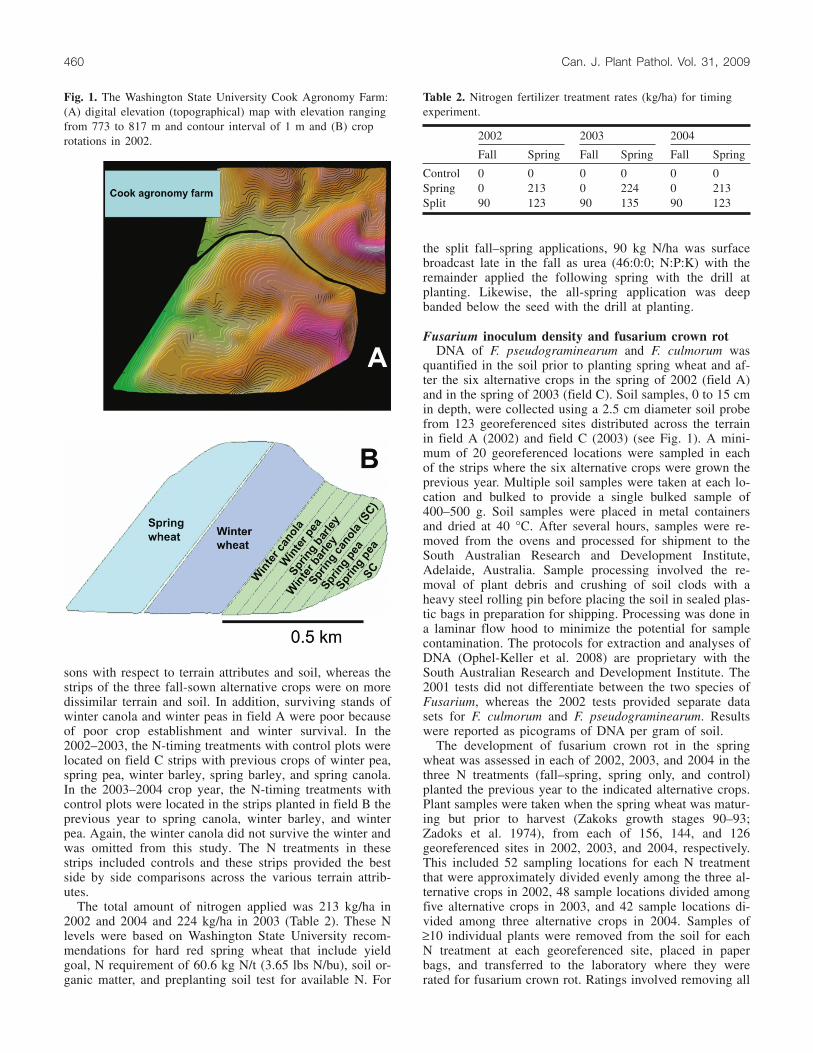
sons with respect to terrain attributes and soil, whereas thestrips of the three fall-sown alternative crops were on moredissimilar terrain and soil. In addition, surviving stands ofwinter canola and winter peas in field A were poor becauseof poor crop establishment and winter survival. In the2002–2003, the N-timing treatments with control plots werelocated on field C strips with previous crops of winter pea,spring pea, winter barley, spring barley, and spring canola.In the 2003–2004 crop year, the N-timing treatments withcontrol plots were located in the strips planted in field B theprevious year to spring canola, winter barley, and winterpea. Again, the winter canola did not survive the winter andwas omitted from this study. The N treatments in thesestrips included controls and these strips provided the bestside by side comparisons across the various terrain attrib-utes.
The total amount of nitrogen applied was 213 kg/ha in2002 and 2004 and 224 kg/ha in 2003 (Table 2). These Nlevels were based on Washington State University recom-mendations for hard red spring wheat that include yieldgoal, N requirement of 60.6 kg N/t (3.65 lbs N/bu), soil or-ganic matter, and preplanting soil test for available N. For
the split fall–spring applications, 90 kg N/ha was surfacebroadcast late in the fall as urea (46:0:0; N:P:K) with theremainder applied the following spring with the drill atplanting. Likewise, the all-spring application was deepbanded below the seed with the drill at planting.
Fusarium inoculum density and fusarium crown rotDNA of F. pseudograminearum and F. culmorum was
quantified in the soil prior to planting spring wheat and af-ter the six alternative crops in the spring of 2002 (field A)and in the spring of 2003 (field C). Soil samples, 0 to 15 cmin depth, were collected using a 2.5 cm diameter soil probefrom 123 georeferenced sites distributed across the terrainin field A (2002) and field C (2003) (see Fig. 1). A mini-mum of 20 georeferenced locations were sampled in eachof the strips where the six alternative crops were grown theprevious year. Multiple soil samples were taken at each lo-cation and bulked to provide a single bulked sample of400–500 g. Soil samples were placed in metal containersand dried at 40 °C. After several hours, samples were re-moved from the ovens and processed for shipment to theSouth Australian Research and Development Institute,Adelaide, Australia. Sample processing involved the re-moval of plant debris and crushing of soil clods with aheavy steel rolling pin before placing the soil in sealed plas-tic bags in preparation for shipping. Processing was done ina laminar flow hood to minimize the potential for samplecontamination. The protocols for extraction and analyses ofDNA (Ophel-Keller et al. 2008) are proprietary with theSouth Australian Research and Development Institute. The2001 tests did not differentiate between the two species ofFusarium, whereas the 2002 tests provided separate datasets for F. culmorum and F. pseudograminearum. Resultswere reported as picograms of DNA per gram of soil.
The development of fusarium crown rot in the springwheat was assessed in each of 2002, 2003, and 2004 in thethree N treatments (fall–spring, spring only, and control)planted the previous year to the indicated alternative crops.Plant samples were taken when the spring wheat was matur-ing but prior to harvest (Zakoks growth stages 90–93;Zadoks et al. 1974), from each of 156, 144, and 126georeferenced sites in 2002, 2003, and 2004, respectively.This included 52 sampling locations for each N treatmentthat were approximately divided evenly among the three al-ternative crops in 2002, 48 sample locations divided amongfive alternative crops in 2003, and 42 sample locations di-vided among three alternative crops in 2004. Samples of≥10 individual plants were removed from the soil for eachN treatment at each georeferenced site, placed in paperbags, and transferred to the laboratory where they wererated for fusarium crown rot. Ratings involved removing all
460 Can. J. Plant Pathol. Vol. 31, 2009
Fig. 1. The Washington State University Cook Agronomy Farm:(A) digital elevation (topographical) map with elevation rangingfrom 773 to 817 m and contour interval of 1 m and (B) croprotations in 2002. 2002 2003 2004
Fall Spring Fall Spring Fall Spring
Control 0 0 0 0 0 0Spring 0 213 0 224 0 213Split 90 123 90 135 90 123
Table 2. Nitrogen fertilizer treatment rates (kg/ha) for timingexperiment.
tillers and then stripping away the outer leaf sheaths of themain stem to expose the lower internodes. Once removed,the extent of disease progress up the main stem was deter-mined based on the number of discolored (chocolate brown)internodes (Cook 1968). Stems were placed into one of fourdisease severity categories. These four categories were asfollows: nil, no sign of fusarium crown rot, no discolorationpresent; slight, slight discoloration of the lowest internode;moderate, discoloration of internodes that extend two orthree nodes up the stem; and heavy, dark discoloration ofinternodes that extend three or more nodes up the stem. Fornonparametric analyses, these classes were assigned as 0, 1,2, and 3, respectively. Each sample had 10 stems rated forcrown rot severity. In 2002, only the 0–3 rating system wasused. In 2003 and 2004, fusarium crown rot was quantifiedboth with the rating system and by counting the number ofinfected (discolored) internodes on each plant.
Grain yield and proteinFor yield determinations, a 2 m × 2 m section within
each georeferenced site within each fertilizer treatment ofeach alternative-crop site was hand harvested, and the grainwas threshed and weighed. The percentage of grain N wasdetermined using a Leco analyzer (Leco Corp., St. Joseph,Mich.) and multiplied by 5.7 to calculate grain protein con-tent.
Statistical analysisAll data were analyzed as a factorial analysis of variance
(ANOVA), with N treatment and alternative crop as mainfactors using the SAS JMP software (SAS Institute Inc.,Cary, N.C.). Fusarium crown rot severity ratings, whichwere ordinal data, were analyzed as nonparametric data us-ing Kruskal–Wallis rank test to determine significant differ-ences. For all other continuous data, mean separation wasperformed using Tukey’s honestly significant difference test.
Results
Fusarium inoculum densityIn 2002, DNA concentrations for the two Fusarium species
ranged from 7 to 26 pg DNA/g soil, with no significant differ-ences among the six rotation strips (data not shown). In 2003,the test distinguished between the two Fusarium spp., andoverall concentrations of DNA of F. culmorum were higherthan F. pseudograminearum (Table 3; P < 0.001). The DNAconcentrations representing F. pseudograminearum were notsignificantly different among the rotation strips, and meansranged from 4 to 107 pg DNA/g soil. In contrast, the highestconcentration of inoculum of F. culmorum based on the DNAtest occurred following spring barley, which was higher thanthe other rotation crops except the winter canola strip that hadbeen replanted with spring canola because of winterkill.
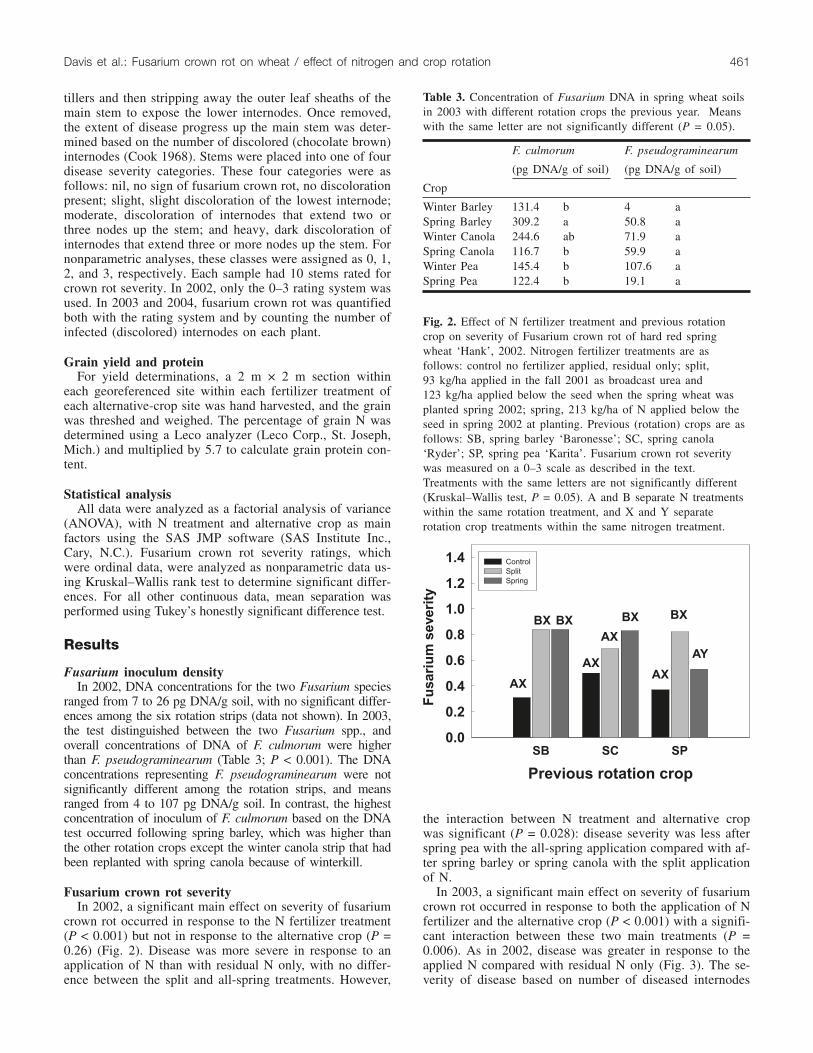
Fusarium crown rot severityIn 2002, a significant main effect on severity of fusarium
crown rot occurred in response to the N fertilizer treatment(P < 0.001) but not in response to the alternative crop (P =0.26) (Fig. 2). Disease was more severe in response to anapplication of N than with residual N only, with no differ-ence between the split and all-spring treatments. However,
the interaction between N treatment and alternative cropwas significant (P = 0.028): disease severity was less afterspring pea with the all-spring application compared with af-ter spring barley or spring canola with the split applicationof N.
In 2003, a significant main effect on severity of fusariumcrown rot occurred in response to both the application of Nfertilizer and the alternative crop (P < 0.001) with a signifi-cant interaction between these two main treatments (P =0.006). As in 2002, disease was greater in response to theapplied N compared with residual N only (Fig. 3). The se-verity of disease based on number of diseased internodes
Davis et al.: Fusarium crown rot on wheat / effect of nitrogen and crop rotation 461
F. culmorum F. pseudograminearum
(pg DNA/g of soil) (pg DNA/g of soil)
Crop
Winter Barley 131.4 b 4 aSpring Barley 309.2 a 50.8 aWinter Canola 244.6 ab 71.9 aSpring Canola 116.7 b 59.9 aWinter Pea 145.4 b 107.6 aSpring Pea 122.4 b 19.1 a
Table 3. Concentration of Fusarium DNA in spring wheat soilsin 2003 with different rotation crops the previous year. Meanswith the same letter are not significantly different (P = 0.05).
Fig. 2. Effect of N fertilizer treatment and previous rotationcrop on severity of Fusarium crown rot of hard red springwheat ‘Hank’, 2002. Nitrogen fertilizer treatments are asfollows: control no fertilizer applied, residual only; split,93 kg/ha applied in the fall 2001 as broadcast urea and123 kg/ha applied below the seed when the spring wheat wasplanted spring 2002; spring, 213 kg/ha of N applied below theseed in spring 2002 at planting. Previous (rotation) crops are asfollows: SB, spring barley ‘Baronesse’; SC, spring canola‘Ryder’; SP, spring pea ‘Karita’. Fusarium crown rot severitywas measured on a 0–3 scale as described in the text.Treatments with the same letters are not significantly different(Kruskal–Wallis test, P = 0.05). A and B separate N treatmentswithin the same rotation treatment, and X and Y separaterotation crop treatments within the same nitrogen treatment.
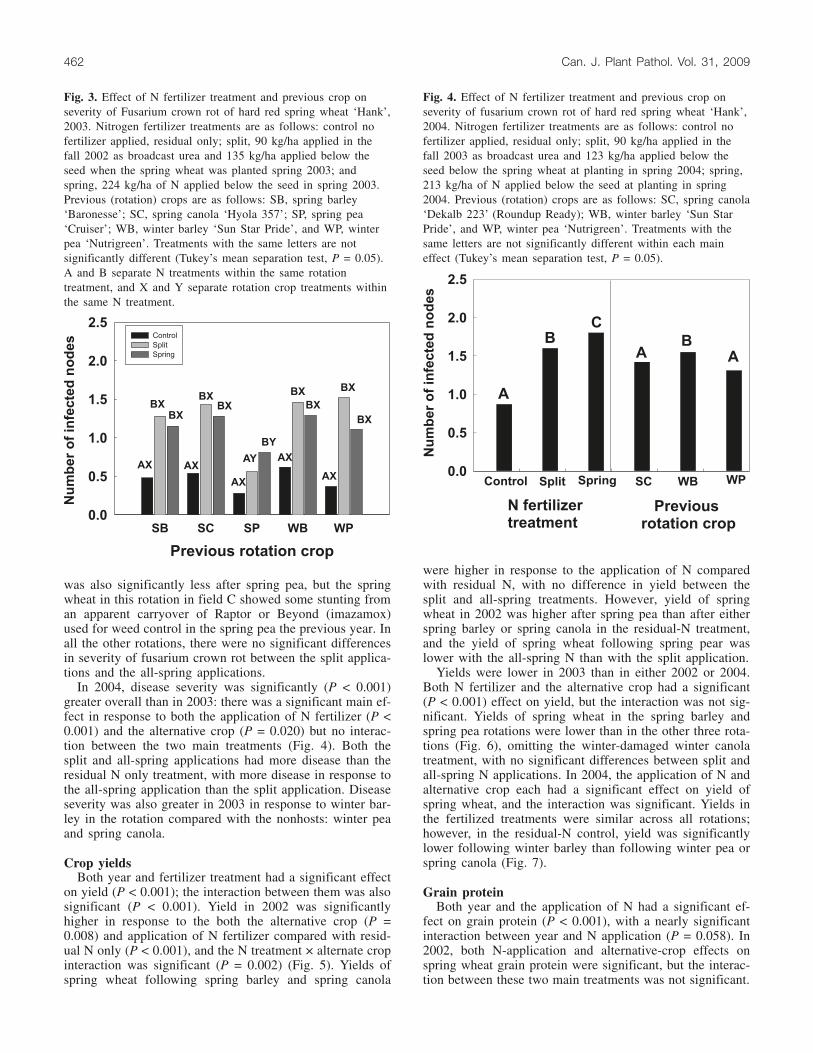
was also significantly less after spring pea, but the springwheat in this rotation in field C showed some stunting froman apparent carryover of Raptor or Beyond (imazamox)used for weed control in the spring pea the previous year. Inall the other rotations, there were no significant differencesin severity of fusarium crown rot between the split applica-tions and the all-spring applications.
In 2004, disease severity was significantly (P < 0.001)greater overall than in 2003: there was a significant main ef-fect in response to both the application of N fertilizer (P <0.001) and the alternative crop (P = 0.020) but no interac-tion between the two main treatments (Fig. 4). Both thesplit and all-spring applications had more disease than theresidual N only treatment, with more disease in response tothe all-spring application than the split application. Diseaseseverity was also greater in 2003 in response to winter bar-ley in the rotation compared with the nonhosts: winter peaand spring canola.
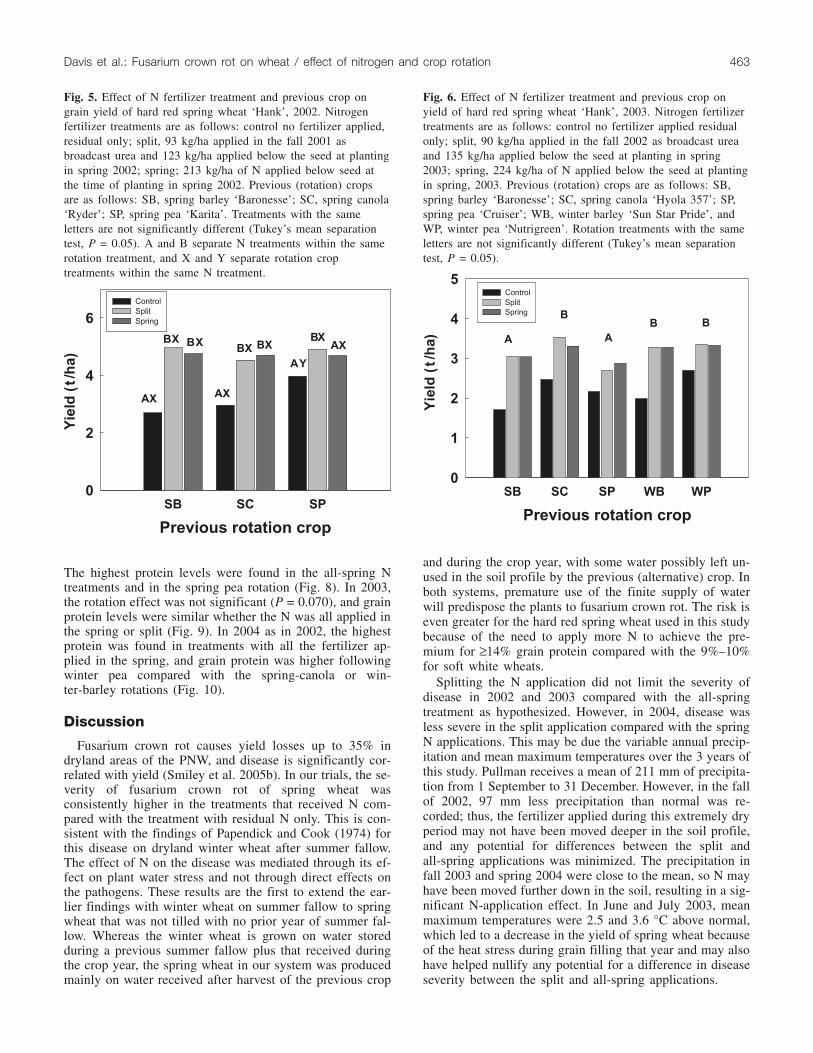
Crop yieldsBoth year and fertilizer treatment had a significant effect
on yield (P < 0.001); the interaction between them was alsosignificant (P < 0.001). Yield in 2002 was significantlyhigher in response to the both the alternative crop (P =0.008) and application of N fertilizer compared with resid-ual N only (P < 0.001), and the N treatment × alternate cropinteraction was significant (P = 0.002) (Fig. 5). Yields ofspring wheat following spring barley and spring canola
were higher in response to the application of N comparedwith residual N, with no difference in yield between thesplit and all-spring treatments. However, yield of springwheat in 2002 was higher after spring pea than after eitherspring barley or spring canola in the residual-N treatment,and the yield of spring wheat following spring pear waslower with the all-spring N than with the split application.
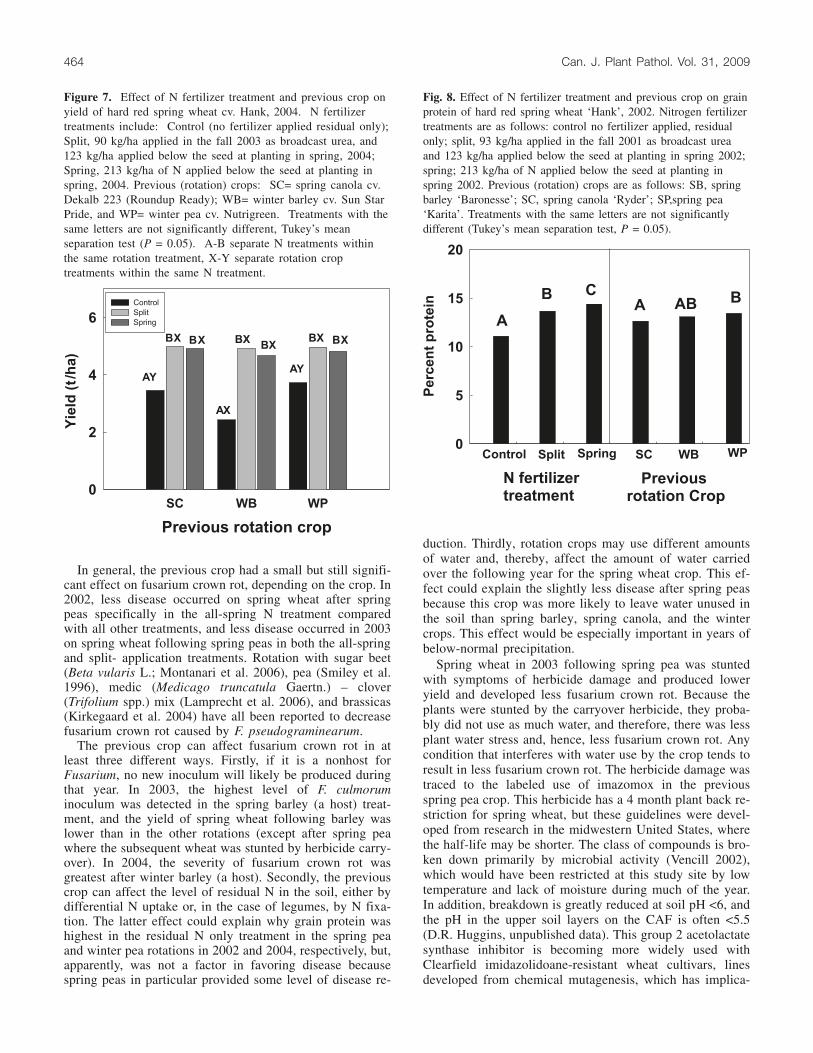
Yields were lower in 2003 than in either 2002 or 2004.Both N fertilizer and the alternative crop had a significant(P < 0.001) effect on yield, but the interaction was not sig-nificant. Yields of spring wheat in the spring barley andspring pea rotations were lower than in the other three rota-tions (Fig. 6), omitting the winter-damaged winter canolatreatment, with no significant differences between split andall-spring N applications. In 2004, the application of N andalternative crop each had a significant effect on yield ofspring wheat, and the interaction was significant. Yields inthe fertilized treatments were similar across all rotations;however, in the residual-N control, yield was significantlylower following winter barley than following winter pea orspring canola (Fig. 7).
Grain proteinBoth year and the application of N had a significant ef-
fect on grain protein (P < 0.001), with a nearly significantinteraction between year and N application (P = 0.058). In2002, both N-application and alternative-crop effects onspring wheat grain protein were significant, but the interac-tion between these two main treatments was not significant.
462 Can. J. Plant Pathol. Vol. 31, 2009
Fig. 3. Effect of N fertilizer treatment and previous crop onseverity of Fusarium crown rot of hard red spring wheat ‘Hank’,2003. Nitrogen fertilizer treatments are as follows: control nofertilizer applied, residual only; split, 90 kg/ha applied in thefall 2002 as broadcast urea and 135 kg/ha applied below theseed when the spring wheat was planted spring 2003; andspring, 224 kg/ha of N applied below the seed in spring 2003.Previous (rotation) crops are as follows: SB, spring barley‘Baronesse’; SC, spring canola ‘Hyola 357’; SP, spring pea‘Cruiser’; WB, winter barley ‘Sun Star Pride’, and WP, winterpea ‘Nutrigreen’. Treatments with the same letters are notsignificantly different (Tukey’s mean separation test, P = 0.05).A and B separate N treatments within the same rotationtreatment, and X and Y separate rotation crop treatments withinthe same N treatment.
Fig. 4. Effect of N fertilizer treatment and previous crop onseverity of fusarium crown rot of hard red spring wheat ‘Hank’,2004. Nitrogen fertilizer treatments are as follows: control nofertilizer applied, residual only; split, 90 kg/ha applied in thefall 2003 as broadcast urea and 123 kg/ha applied below theseed below the spring wheat at planting in spring 2004; spring,213 kg/ha of N applied below the seed at planting in spring2004. Previous (rotation) crops are as follows: SC, spring canola‘Dekalb 223’ (Roundup Ready); WB, winter barley ‘Sun StarPride’, and WP, winter pea ‘Nutrigreen’. Treatments with thesame letters are not significantly different within each maineffect (Tukey’s mean separation test, P = 0.05).
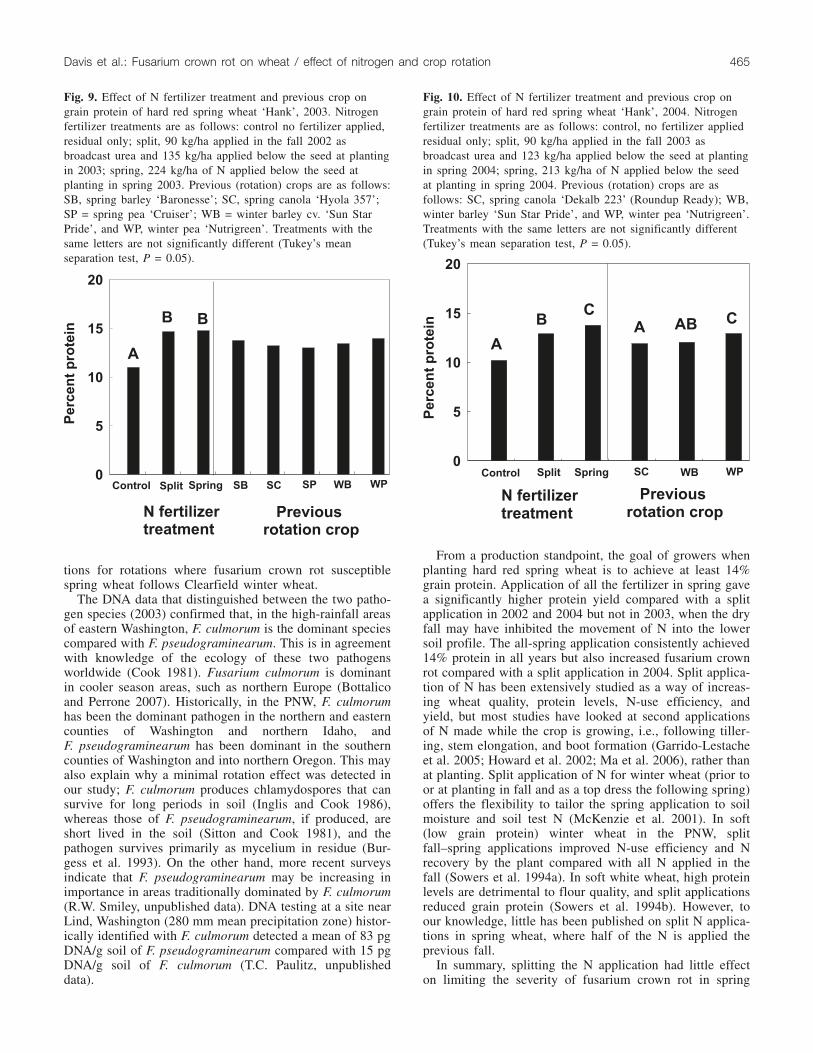
The highest protein levels were found in the all-spring Ntreatments and in the spring pea rotation (Fig. 8). In 2003,the rotation effect was not significant (P = 0.070), and grainprotein levels were similar whether the N was all applied inthe spring or split (Fig. 9). In 2004 as in 2002, the highestprotein was found in treatments with all the fertilizer ap-plied in the spring, and grain protein was higher followingwinter pea compared with the spring-canola or win-ter-barley rotations (Fig. 10).
Discussion
Fusarium crown rot causes yield losses up to 35% indryland areas of the PNW, and disease is significantly cor-related with yield (Smiley et al. 2005b). In our trials, the se-verity of fusarium crown rot of spring wheat wasconsistently higher in the treatments that received N com-pared with the treatment with residual N only. This is con-sistent with the findings of Papendick and Cook (1974) forthis disease on dryland winter wheat after summer fallow.The effect of N on the disease was mediated through its ef-fect on plant water stress and not through direct effects onthe pathogens. These results are the first to extend the ear-lier findings with winter wheat on summer fallow to springwheat that was not tilled with no prior year of summer fal-low. Whereas the winter wheat is grown on water storedduring a previous summer fallow plus that received duringthe crop year, the spring wheat in our system was producedmainly on water received after harvest of the previous crop
and during the crop year, with some water possibly left un-used in the soil profile by the previous (alternative) crop. Inboth systems, premature use of the finite supply of waterwill predispose the plants to fusarium crown rot. The risk iseven greater for the hard red spring wheat used in this studybecause of the need to apply more N to achieve the pre-mium for ≥14% grain protein compared with the 9%–10%for soft white wheats.
Splitting the N application did not limit the severity ofdisease in 2002 and 2003 compared with the all-springtreatment as hypothesized. However, in 2004, disease wasless severe in the split application compared with the springN applications. This may be due the variable annual precip-itation and mean maximum temperatures over the 3 years ofthis study. Pullman receives a mean of 211 mm of precipita-tion from 1 September to 31 December. However, in the fallof 2002, 97 mm less precipitation than normal was re-corded; thus, the fertilizer applied during this extremely dryperiod may not have been moved deeper in the soil profile,and any potential for differences between the split andall-spring applications was minimized. The precipitation infall 2003 and spring 2004 were close to the mean, so N mayhave been moved further down in the soil, resulting in a sig-nificant N-application effect. In June and July 2003, meanmaximum temperatures were 2.5 and 3.6 °C above normal,which led to a decrease in the yield of spring wheat becauseof the heat stress during grain filling that year and may alsohave helped nullify any potential for a difference in diseaseseverity between the split and all-spring applications.
Davis et al.: Fusarium crown rot on wheat / effect of nitrogen and crop rotation 463
Fig. 5. Effect of N fertilizer treatment and previous crop ongrain yield of hard red spring wheat ‘Hank’, 2002. Nitrogenfertilizer treatments are as follows: control no fertilizer applied,residual only; split, 93 kg/ha applied in the fall 2001 asbroadcast urea and 123 kg/ha applied below the seed at plantingin spring 2002; spring; 213 kg/ha of N applied below seed atthe time of planting in spring 2002. Previous (rotation) cropsare as follows: SB, spring barley ‘Baronesse’; SC, spring canola‘Ryder’; SP, spring pea ‘Karita’. Treatments with the sameletters are not significantly different (Tukey’s mean separationtest, P = 0.05). A and B separate N treatments within the samerotation treatment, and X and Y separate rotation croptreatments within the same N treatment.
Fig. 6. Effect of N fertilizer treatment and previous crop onyield of hard red spring wheat ‘Hank’, 2003. Nitrogen fertilizertreatments are as follows: control no fertilizer applied residualonly; split, 90 kg/ha applied in the fall 2002 as broadcast ureaand 135 kg/ha applied below the seed at planting in spring2003; spring, 224 kg/ha of N applied below the seed at plantingin spring, 2003. Previous (rotation) crops are as follows: SB,spring barley ‘Baronesse’; SC, spring canola ‘Hyola 357’; SP,spring pea ‘Cruiser’; WB, winter barley ‘Sun Star Pride’, andWP, winter pea ‘Nutrigreen’. Rotation treatments with the sameletters are not significantly different (Tukey’s mean separationtest, P = 0.05).
In general, the previous crop had a small but still signifi-cant effect on fusarium crown rot, depending on the crop. In2002, less disease occurred on spring wheat after springpeas specifically in the all-spring N treatment comparedwith all other treatments, and less disease occurred in 2003on spring wheat following spring peas in both the all-springand split- application treatments. Rotation with sugar beet(Beta vularis L.; Montanari et al. 2006), pea (Smiley et al.1996), medic (Medicago truncatula Gaertn.) – clover(Trifolium spp.) mix (Lamprecht et al. 2006), and brassicas(Kirkegaard et al. 2004) have all been reported to decreasefusarium crown rot caused by F. pseudograminearum.
The previous crop can affect fusarium crown rot in atleast three different ways. Firstly, if it is a nonhost forFusarium, no new inoculum will likely be produced duringthat year. In 2003, the highest level of F. culmoruminoculum was detected in the spring barley (a host) treat-ment, and the yield of spring wheat following barley waslower than in the other rotations (except after spring peawhere the subsequent wheat was stunted by herbicide carry-over). In 2004, the severity of fusarium crown rot wasgreatest after winter barley (a host). Secondly, the previouscrop can affect the level of residual N in the soil, either bydifferential N uptake or, in the case of legumes, by N fixa-tion. The latter effect could explain why grain protein washighest in the residual N only treatment in the spring peaand winter pea rotations in 2002 and 2004, respectively, but,apparently, was not a factor in favoring disease becausespring peas in particular provided some level of disease re-
duction. Thirdly, rotation crops may use different amountsof water and, thereby, affect the amount of water carriedover the following year for the spring wheat crop. This ef-fect could explain the slightly less disease after spring peasbecause this crop was more likely to leave water unused inthe soil than spring barley, spring canola, and the wintercrops. This effect would be especially important in years ofbelow-normal precipitation.
Spring wheat in 2003 following spring pea was stuntedwith symptoms of herbicide damage and produced loweryield and developed less fusarium crown rot. Because theplants were stunted by the carryover herbicide, they proba-bly did not use as much water, and therefore, there was lessplant water stress and, hence, less fusarium crown rot. Anycondition that interferes with water use by the crop tends toresult in less fusarium crown rot. The herbicide damage wastraced to the labeled use of imazomox in the previousspring pea crop. This herbicide has a 4 month plant back re-striction for spring wheat, but these guidelines were devel-oped from research in the midwestern United States, wherethe half-life may be shorter. The class of compounds is bro-ken down primarily by microbial activity (Vencill 2002),which would have been restricted at this study site by lowtemperature and lack of moisture during much of the year.In addition, breakdown is greatly reduced at soil pH <6, andthe pH in the upper soil layers on the CAF is often <5.5(D.R. Huggins, unpublished data). This group 2 acetolactatesynthase inhibitor is becoming more widely used withClearfield imidazolidoane-resistant wheat cultivars, linesdeveloped from chemical mutagenesis, which has implica-
464 Can. J. Plant Pathol. Vol. 31, 2009
Fig. 8. Effect of N fertilizer treatment and previous crop on grainprotein of hard red spring wheat ‘Hank’, 2002. Nitrogen fertilizertreatments are as follows: control no fertilizer applied, residualonly; split, 93 kg/ha applied in the fall 2001 as broadcast ureaand 123 kg/ha applied below the seed at planting in spring 2002;spring; 213 kg/ha of N applied below the seed at planting inspring 2002. Previous (rotation) crops are as follows: SB, springbarley ‘Baronesse’; SC, spring canola ‘Ryder’; SP,spring pea‘Karita’. Treatments with the same letters are not significantlydifferent (Tukey’s mean separation test, P = 0.05).
Figure 7. Effect of N fertilizer treatment and previous crop onyield of hard red spring wheat cv. Hank, 2004. N fertilizertreatments include: Control (no fertilizer applied residual only);Split, 90 kg/ha applied in the fall 2003 as broadcast urea, and123 kg/ha applied below the seed at planting in spring, 2004;Spring, 213 kg/ha of N applied below the seed at planting inspring, 2004. Previous (rotation) crops: SC= spring canola cv.Dekalb 223 (Roundup Ready); WB= winter barley cv. Sun StarPride, and WP= winter pea cv. Nutrigreen. Treatments with thesame letters are not significantly different, Tukey’s meanseparation test (P = 0.05). A-B separate N treatments withinthe same rotation treatment, X-Y separate rotation croptreatments within the same N treatment.
tions for rotations where fusarium crown rot susceptiblespring wheat follows Clearfield winter wheat.
The DNA data that distinguished between the two patho-gen species (2003) confirmed that, in the high-rainfall areasof eastern Washington, F. culmorum is the dominant speciescompared with F. pseudograminearum. This is in agreementwith knowledge of the ecology of these two pathogensworldwide (Cook 1981). Fusarium culmorum is dominantin cooler season areas, such as northern Europe (Bottalicoand Perrone 2007). Historically, in the PNW, F. culmorumhas been the dominant pathogen in the northern and easterncounties of Washington and northern Idaho, andF. pseudograminearum has been dominant in the southerncounties of Washington and into northern Oregon. This mayalso explain why a minimal rotation effect was detected inour study; F. culmorum produces chlamydospores that cansurvive for long periods in soil (Inglis and Cook 1986),whereas those of F. pseudograminearum, if produced, areshort lived in the soil (Sitton and Cook 1981), and thepathogen survives primarily as mycelium in residue (Bur-gess et al. 1993). On the other hand, more recent surveysindicate that F. pseudograminearum may be increasing inimportance in areas traditionally dominated by F. culmorum(R.W. Smiley, unpublished data). DNA testing at a site nearLind, Washington (280 mm mean precipitation zone) histor-ically identified with F. culmorum detected a mean of 83 pgDNA/g soil of F. pseudograminearum compared with 15 pgDNA/g soil of F. culmorum (T.C. Paulitz, unpublisheddata).
From a production standpoint, the goal of growers whenplanting hard red spring wheat is to achieve at least 14%grain protein. Application of all the fertilizer in spring gavea significantly higher protein yield compared with a splitapplication in 2002 and 2004 but not in 2003, when the dryfall may have inhibited the movement of N into the lowersoil profile. The all-spring application consistently achieved14% protein in all years but also increased fusarium crownrot compared with a split application in 2004. Split applica-tion of N has been extensively studied as a way of increas-ing wheat quality, protein levels, N-use efficiency, andyield, but most studies have looked at second applicationsof N made while the crop is growing, i.e., following tiller-ing, stem elongation, and boot formation (Garrido-Lestacheet al. 2005; Howard et al. 2002; Ma et al. 2006), rather thanat planting. Split application of N for winter wheat (prior toor at planting in fall and as a top dress the following spring)offers the flexibility to tailor the spring application to soilmoisture and soil test N (McKenzie et al. 2001). In soft(low grain protein) winter wheat in the PNW, splitfall–spring applications improved N-use efficiency and Nrecovery by the plant compared with all N applied in thefall (Sowers et al. 1994a). In soft white wheat, high proteinlevels are detrimental to flour quality, and split applicationsreduced grain protein (Sowers et al. 1994b). However, toour knowledge, little has been published on split N applica-tions in spring wheat, where half of the N is applied theprevious fall.
In summary, splitting the N application had little effecton limiting the severity of fusarium crown rot in spring
Davis et al.: Fusarium crown rot on wheat / effect of nitrogen and crop rotation 465
Fig. 9. Effect of N fertilizer treatment and previous crop ongrain protein of hard red spring wheat ‘Hank’, 2003. Nitrogenfertilizer treatments are as follows: control no fertilizer applied,residual only; split, 90 kg/ha applied in the fall 2002 asbroadcast urea and 135 kg/ha applied below the seed at plantingin 2003; spring, 224 kg/ha of N applied below the seed atplanting in spring 2003. Previous (rotation) crops are as follows:SB, spring barley ‘Baronesse’; SC, spring canola ‘Hyola 357’;SP = spring pea ‘Cruiser’; WB = winter barley cv. ‘Sun StarPride’, and WP, winter pea ‘Nutrigreen’. Treatments with thesame letters are not significantly different (Tukey’s meanseparation test, P = 0.05).
Fig. 10. Effect of N fertilizer treatment and previous crop ongrain protein of hard red spring wheat ‘Hank’, 2004. Nitrogenfertilizer treatments are as follows: control, no fertilizer appliedresidual only; split, 90 kg/ha applied in the fall 2003 asbroadcast urea and 123 kg/ha applied below the seed at plantingin spring 2004; spring, 213 kg/ha of N applied below the seedat planting in spring 2004. Previous (rotation) crops are asfollows: SC, spring canola ‘Dekalb 223’ (Roundup Ready); WB,winter barley ‘Sun Star Pride’, and WP, winter pea ‘Nutrigreen’.Treatments with the same letters are not significantly different(Tukey’s mean separation test, P = 0.05).

wheat, possibly because both treatments resulted in highlevels of N needed to meet the goal of 14% grain protein.Rotation with nonhosts and barley had minor effects, proba-bly because of the ability of the Fusarium spp. to carry overat high inoculum potentials during a 1 year break to anonhost crop, especially in a direct-seed system where lev-els of infested residue on and within the top few centime-ters of soil are high and the rate of residue decomposition isslow.
Acknowldegements
We thank Ron Sloot, Shawn Wetterau, and DaveUberuaga for assistance with the field plots and databasemanagement. This research is partially supported by a grantfrom the Washington Wheat Commission and the R. JamesCook Endowed Chair in Wheat Research.
References
Backhouse, D., Abubakar, A.A., Burgess, L.W., Dennis, J.I.,Hollaway, G.J., Wildermuth, G.B., Wallwork, H., andHenry, F.J. 2004. Survey of Fusarium species associated withcrown rot of wheat and barley in eastern Australia. Australas.Plant Pathol. 33: 255–261.
Bailey, K.L., Gossen, B.D., Lafond, G.P., Watson, P.R., andDerksen, D.A. 2001. Effect of tillage and crop rotation on rootand foliar diseases of wheat and pea in Saskatchewan from1991–1998: univariate and multivariate analyses. Can. J. PlantSci. 81: 789–803.
Baker, C.J., Saxton, K.E., and Ritchie, W.R. 1996. No-tillageseeding: science and practice. CAB International, Wallingford,UK.
Baker, D.A., Young, D.L., Huggins, D.R., and Pan, W.L. 2004.Economically optimal nitrogen fertilization for yield and pro-tein in hard red spring wheat. Agron. J. 96:116–123.
Bentley, A.R., Tunali, B., Nicol, J.M., Burgess, L.W., andSummerell, B.A. 2006. A survey of Fusarium species associ-ated with wheat and grass stem bases in northern Turkey.Sydowia, 58: 163–177.
Bottalico, A., and Perrone, G. 2007. Toxigenic Fusarium speciesand mycotoxins associated with head blight in small-grain cere-als in Europe. Eur. J. Plant Pathol. 108: 611–624.
Burgess, L.W., Backhouse, D., Summerell, B.A., Pattison, A.B.,Klein, T.A., Esdaile, R.J., and Ticehurst, G. 1993. Long-termeffects of stubble management on the incidence of infection ofwheat by Fusarium graminearum Schw. Group 1. Australas. J.Exp. Agric. 33: 451–456.
Cook, R.J. 1968. Fusarium root and foot rot of cereals in the Pa-cific Northwest. Phytopathology, 58: 127–131.
Cook, R.J. 1980. Fusarium foot rot of wheat and its control in thePacific Northwest. Plant Dis. 64: 1061–1066.
Cook, R.J. 1981. Fusarium diseases of wheat and other smallgrains in North America. In Fusarium diseases, biology andtaxonomy. Edited by P.E. Nelson, T.A. Toussoun, and R.J.Cook. Pennsylvania State University Press, University Park, Pa.pp. 39–52.
Cook, R.J., and Papendick, R.I. 1972. Influence of water poten-tial soils and plants on root disease. Annu. Rev. Phytopathol.10: 349–374.
Cook, R.J., and Veseth, R.J. 1991. Wheat health management.American Phytopathological Society, St. Paul, Minn.
Douglas, J.T., and Goss, M.J. 1982. Stability and organic mattercontent of surface soil aggregates under different methods ofcultivation and in grassland. Soil Tillage Res. 2: 155–175.
Fouly, H.M., Pedersen, W.L., Wilkinson, H.T., and AbdEl-Kader, M.M. 1996. Wheat root rotting fungi in the “old”and “new” agriculture lands of Egypt. Plant Dis. 80:1298–1300.
Garrido-Lestache, E., Lopez-Bellido, R.J., andLopez-Bellido, L. 2005. Durum wheat quality under Mediter-ranean conditions as affected by N rate, timing and splitting, Nform and S fertilization. Eur. J. Agron. 23: 265–278.
Howard, D.D., Newman, M.A., Essington, M.E., and Percell,W.M. 2002. Nitrogen fertilization of conservation-tilled wheat.II. Timing of application of two nitrogen sources. J. Plant Nutr.25: 1329–1339.
Huggins, D.R. 1991. Redesigning no-tillage cropping systems: al-ternatives for increasing productivity and nitrogen use effi-ciency. PhD dissertation, Washington State University, Pullman,Wash.
Inglis, D.A., and Cook, R.J. 1986. Persistence ofchlamydospores of Fusarium culmorum in wheat field soils ofeastern Washington. Phytopathology, 76: 1205–1208.
Janosky, J.S. 1999. An economic analysis of conservation tillagecropping systems in eastern Washington. M.A. thesis, Depart-ment of Agricutural Economics, Washington State University,Pullman, Wash.
Kennedy, A.C., and Schillinger, W.F. 2006. Soil quality and wa-ter intake in traditional-till vs. no-till paired farms in Washing-ton’s Palouse region. Soil Sci. Soc. Am. J. 70: 940–949.
Kirkegaard, J.A., Simpfendorfer, S., Holland, J., Bambach, R.,Moore, K.J., and Rebetzke, G.J. 2004. Effect of previouscrops on crown rot and yield of durum and bread wheat innorthern NSW. Aust. J. Agric. Res. 55: 321–334.
Koenig, R.T. 2005. Dryland winter wheat, eastern Washington nu-trient management guide. Washington State University, Pull-man, Wash. Wash. State Univ. Ext. Guide EB1987.
Lamprecht, S.C., Marasas, W.F.O., Hardy, M.B., and Calitz,F.J. 2006. Effect of crop rotation on crown rot and the inci-dence of Fusarium pseudograminearum in wheat in the West-ern Cape, South Africa. Australas. Plant Pathol. 35: 419–426.
Ma, B.L., Subedi, K.D., and Dwyer, L.M. 2006. Timing andmethod of 15nitrogen-labeled fertilizer application on grain pro-tein and nitrogen use efficiency of spring wheat. J. Plant Nutr.29: 469–483.
McKenzie, R.H., Middleton, A.B., and Zhang, M. 2001. Opti-mal time and placement of nitrogen fertilizer with direct andconventionally seeded winter wheat. Can. J. Soil Sci. 81:613–621.
Montanari, A., Innocenti, G., and Toderi, G. 2006. Effects ofcultural management on the foot and root disease complex ofdurum wheat. J. Plant Pathol. 88: 149–156.
Ophel-Keller, K., McKay, A., Hartley, D., Herdina, and Curran,J. 2008. Development of a routine DNA-based testing service forsoilborne diseases in Australia. Australas. Plant Pathol. 37:243–253
Papendick, R.I. 2004. Farming with the wind II: wind erosion andair quality control on the Columbia Plateau and Columbia Basin.University Publishing, Washington State University, Pullman,Wash.
Papendick, R.I., and Cook, R.J. 1974. Plant water stress and devel-opment of Fusarium (Roseum) foot rot in wheat subjected to dif-ferent cultural practices. Phytopathology, 64: 358–363.
466 Can. J. Plant Pathol. Vol. 31, 2009

Paulitz, T.C., Smiley, R.W., and Cook, R.J. 2002. Insights intothe prevalence and management of soilborne cereal pathogensunder direct seeding in the Pacific Northwest, U.S.A. Can. J.Plant Pathol. 24: 416-428.
Purakayastha, T.J., Huggins, D.R., and Smith, J.L. 2008. Carbonsequestration in native prairie, perennial grass, no-till and culti-vated Palouse silt loam. Soil Sci. Soc. Am. J. 72:534–540.
Sitton, J.W., and Cook, R.J. 1981. Comparative morphology andsurvival of chlamydospores of Fusarium roseum ‘Culmorum’and ‘Graminearum’. Phytopathology, 71: 85–90.
Smiley, R.W., and Patterson, L.M. 1996. Pathogenic fungi associ-ated with Fusarium foot rot of winter wheat in the semiarid Pa-cific Northwest. Plant Dis. 80: 994–949.
Smiley, R.W., Collins, H.P., and Rasmussen, P.E. 1996. Diseasesof wheat in long-term agronomic experiments at Pendleton, Ore-gon. Plant Dis. 80: 813–820.
Smiley, R.W., Gourlie, J.A., Easley, S.A., and Patterson, L.M.2005a. Pathogenicity of fungi associated with the wheat crownrot complex in Oregon and Washington. Plant Dis. 89: 949–957.
Smiley, R.W., Gourlie, J.A., Easley, S.A., Patterson, L.M., andWhittaker, R.G. 2005b. Crop damage estimates for crown rot ofwheat and barley in the Pacific Northwest. Plant Dis. 89:595–604.
Sowers, K.E., Pan, W.L., Miller, B.C., and Smith, J.L. 1994a.Nitrogen use efficiency of split nitrogen applications in softwhite winter wheat. Agron. J. 86: 942–948.
Sowers, K.E., Miller, B.C., and Pan, W.L. 1994b. Optimizingyield and grain protein in soft white winter wheat with split ni-trogen applications. Agron. J. 86: 1020–1025.
Vencill, W.K. (Editor). 2002. Herbicide handbook. 8th ed. WeedScience Society of America, Lawrence, Kan.
Wiese, M.V. (Editor). 1987. Compendium of wheat diseases. 2nded. American Phytopathological Society Press, St. Paul, Minn.
Wildermuth, G.B., Thomas, G.A., Radford, B.J., McNamara,R.B., and Kelly, A. 1997. Crown rot and common root rot inwheat grown under different tillage and stubble treatments insouthern Queensland, Australia. Soil Tillage Res. 44: 211–224.
Zadoks, J.C., Chang, T.T., and Konzak, C.F. 1974. A decimalcode for the growth stages of cereals. Weed Res. 14: 415–421.
Davis et al.: Fusarium crown rot on wheat / effect of nitrogen and crop rotation 467




















