
New observations on the skull of Archaeopteryx
11
RESEARCH PAPER New observations on the skull of Archaeopteryx Oliver W. M. Rauhut Received: 18 January 2013 / Accepted: 14 May 2013 / Published online: 8 June 2013 Ó Springer-Verlag Berlin Heidelberg 2013 Abstract Although skeletal remains of the iconic oldest known avialian Archaeopteryx have been known for almost 150 years, several aspects of the cranial anatomy of this taxon have remained enigmatic, mainly because of the strongly flattened and often fractured and incomplete nat- ure of available skull materials. New investigation of the skulls of the recently described, excellently preserved tenth (Thermopolis) and the seventh (Munich) specimens revealed several previously unrecognized characters and helps to resolve some problematic issues. Thus, the nasal of Archaeopteryx shows a lateral notch for the lacrimal, as is found in many other saurischian dinosaurs, the maxilla clearly participates in the margin of the external nares, and there seems to be a pneumatic foramen in the lacrimal, comparable to the lacrimal fenestra found in many non- avian theropods. In the braincase, Archaeopteryx shows pneumatic features reminiscent of non-avian theropods, including a ventral basisphenoid recess and an anterior tympanic recess that is laterally incised into the basisphe- noid/prootic. Most importantly, however, the postorbital process of the jugal shows a facet for the suture with the postorbital, thus resolving the question of whether Archaeopteryx had a closed postorbital bar. A new recon- struction of the skull of Archaeopteryx is presented, mak- ing the skull of this taxon even more theropod-like than previously recognized. Furthermore, the closed postorbital bar and the configuration of the bones of the skull roof cast serious doubt on claims that an avian-style cranial kinesis was present in this taxon. Keywords Archaeopteryx Á Upper Jurassic Á Avialae Á Cranial osteology Á Cranial kinesis Kurzfassung Obwohl Skelettreste des a ¨ltesten be- kannten Vogels Archaeopteryx seit nun 150 Jahren be- kannt sind, sind einige Aspekte der Scha ¨delanatomie dieses Taxon weiter ungewiss, vor allem da die meisten bekannten Scha ¨delreste stark komprimiert und meist zerbrochen und unvollsta ¨ndig sind. Neue Untersuchun- gen am Scha ¨del des ku ¨rzlich beschriebenen und her- vorragend erhaltenen 10. (Thermopolis) und des 7. (Mu ¨nchener) Exemplares zeigen einige bisher un- erkannte Merkmale der Scha ¨delanatomie und helfen, andere bisher umstrittene Fragen zu lo ¨sen. So zeigt das Nasale von Archaeopteryx einen lateralen Einschnitt fu ¨r die Sutur mit dem Lacrimale, wie er bei vielen Saurischia vorhanden ist, das Maxillare hat Anteil am Rand der externen Nares und das Lacrimale hat offenbar ein gro- ßes, pneumatisches Foramen, das in seiner Position dem Lacrimal-Fenster vieler basalerer Theropoden entspricht. Im Hirnscha ¨del zeigt Archaeopteryx Merkmale die an jene basalerer Theropoden erinnern, so etwa einen Re- zessus basisphenoidalis und einen anterioren tympani- schen Rezessus, der von lateral in das Basisphenoid und das Prooticum einschneidet. Insbesondere zeigt jedoch der Postorbital-Fortsatz des Jugale eine Facette fu ¨r die Sutur mit dem Postorbitale, was die Frage kla ¨rt, ob Archaeopteryx eine geschlossene Postorbital-Spange besass. Eine neue Rekonstruktion des Scha ¨dels von Archaeopteryx macht diesen noch Theropoden-a ¨hnlicher als bisher angenommen. Zudem stellt die geschlossene Postorbital-Spange und die Konfiguration der Knochen des Scha ¨deldaches die angenommene Vogel-a ¨hnliche Scha ¨del-Kinetik bei Archaeopteryx in Frage. O. W. M. Rauhut (&) Bayerische Staatssammlung fu ¨r Pala ¨ontologie und Geologie and Department of Earth and Environmental Sciences, LMU Munich, Richard-Wagner-Str. 10, 80333 Munich, Germany e-mail: [email protected] 123 Pala ¨ontol Z (2014) 88:211–221 DOI 10.1007/s12542-013-0186-0
-
Upload
lmu-munich -
Category
Documents
-
view
1 -
download
0
Transcript of New observations on the skull of Archaeopteryx

RESEARCH PAPER
New observations on the skull of Archaeopteryx
Oliver W. M. Rauhut
Received: 18 January 2013 / Accepted: 14 May 2013 / Published online: 8 June 2013
� Springer-Verlag Berlin Heidelberg 2013
Abstract Although skeletal remains of the iconic oldest
known avialian Archaeopteryx have been known for almost
150 years, several aspects of the cranial anatomy of this
taxon have remained enigmatic, mainly because of the
strongly flattened and often fractured and incomplete nat-
ure of available skull materials. New investigation of the
skulls of the recently described, excellently preserved tenth
(Thermopolis) and the seventh (Munich) specimens
revealed several previously unrecognized characters and
helps to resolve some problematic issues. Thus, the nasal of
Archaeopteryx shows a lateral notch for the lacrimal, as is
found in many other saurischian dinosaurs, the maxilla
clearly participates in the margin of the external nares, and
there seems to be a pneumatic foramen in the lacrimal,
comparable to the lacrimal fenestra found in many non-
avian theropods. In the braincase, Archaeopteryx shows
pneumatic features reminiscent of non-avian theropods,
including a ventral basisphenoid recess and an anterior
tympanic recess that is laterally incised into the basisphe-
noid/prootic. Most importantly, however, the postorbital
process of the jugal shows a facet for the suture with the
postorbital, thus resolving the question of whether
Archaeopteryx had a closed postorbital bar. A new recon-
struction of the skull of Archaeopteryx is presented, mak-
ing the skull of this taxon even more theropod-like than
previously recognized. Furthermore, the closed postorbital
bar and the configuration of the bones of the skull roof cast
serious doubt on claims that an avian-style cranial kinesis
was present in this taxon.
Keywords Archaeopteryx � Upper Jurassic � Avialae �Cranial osteology � Cranial kinesis
Kurzfassung Obwohl Skelettreste des altesten be-
kannten Vogels Archaeopteryx seit nun 150 Jahren be-
kannt sind, sind einige Aspekte der Schadelanatomie
dieses Taxon weiter ungewiss, vor allem da die meisten
bekannten Schadelreste stark komprimiert und meist
zerbrochen und unvollstandig sind. Neue Untersuchun-
gen am Schadel des kurzlich beschriebenen und her-
vorragend erhaltenen 10. (Thermopolis) und des 7.
(Munchener) Exemplares zeigen einige bisher un-
erkannte Merkmale der Schadelanatomie und helfen,
andere bisher umstrittene Fragen zu losen. So zeigt das
Nasale von Archaeopteryx einen lateralen Einschnitt fur
die Sutur mit dem Lacrimale, wie er bei vielen Saurischia
vorhanden ist, das Maxillare hat Anteil am Rand der
externen Nares und das Lacrimale hat offenbar ein gro-
ßes, pneumatisches Foramen, das in seiner Position dem
Lacrimal-Fenster vieler basalerer Theropoden entspricht.
Im Hirnschadel zeigt Archaeopteryx Merkmale die an
jene basalerer Theropoden erinnern, so etwa einen Re-
zessus basisphenoidalis und einen anterioren tympani-
schen Rezessus, der von lateral in das Basisphenoid und
das Prooticum einschneidet. Insbesondere zeigt jedoch
der Postorbital-Fortsatz des Jugale eine Facette fur die
Sutur mit dem Postorbitale, was die Frage klart, ob
Archaeopteryx eine geschlossene Postorbital-Spange
besass. Eine neue Rekonstruktion des Schadels von
Archaeopteryx macht diesen noch Theropoden-ahnlicher
als bisher angenommen. Zudem stellt die geschlossene
Postorbital-Spange und die Konfiguration der Knochen
des Schadeldaches die angenommene Vogel-ahnliche
Schadel-Kinetik bei Archaeopteryx in Frage.
O. W. M. Rauhut (&)
Bayerische Staatssammlung fur Palaontologie und Geologie
and Department of Earth and Environmental Sciences, LMU
Munich, Richard-Wagner-Str. 10, 80333 Munich, Germany
e-mail: [email protected]
123
Palaontol Z (2014) 88:211–221
DOI 10.1007/s12542-013-0186-0

Schlusselworter Archaeotperyx � Oberer Jura � Avialae �Schadel-Osteologie � Schadel-Kinetik
Introduction
Ever since the discovery of the first skeletal remains in
1861 (von Meyer 1861), just 2 years after the publication of
Darwin’s ‘‘Origin of Species’’ (Darwin 1859), the oldest
known ‘‘bird’’ Archaeopteryx has played a pivotal role in
the scientific discussion of the origin of birds (e.g., Huxley
1868; Heilmann 1926; Ostrom 1973, 1976). Thus, despite
the important discoveries of numerous bird-like non-avian
theropod dinosaurs and basal birds in the past decades in
China (see Xu and Norell 2006 and Zhou and Zhang 2006
for recent reviews) and recent claims that the taxon is not
placed on the immediate lineage leading toward birds (Xu
et al. 2011), Archaeopteryx can still be regarded as the
‘‘yardstick’’ of bird evolution, and new insights into the
anatomy and biology of this animal are published frequently
(e.g., Dominguez Alonso et al. 2004; Mayr et al. 2005, 2007;
Tischlinger 2005, 2009; Wellnhofer 2008; Erickson et al.
2009; Bergmann et al. 2010; Longrich et al. 2012).
For more than 100 years after the first discoveries, the
cranial structure of Archaeopteryx has been known only
from the very incomplete and/or strongly damaged remains
of the London and Berlin specimens (see Dames 1884;
Heilmann 1926; De Beer 1954). With the discovery of the
fifth (Eichstatt) specimen, a first well-preserved skull of
this animal became available, and study of this specimen
led to important new insights into the cranial anatomy of
the earliest known bird (Wellnhofer 1974). Further prepa-
ration of the skull of the London specimen and the dis-
covery of the Munich specimen provided new data,
especially on the structure of the braincase, palate, and
lower jaw of this taxon in the following 25 years (Whet-
stone 1983; Walker 1985; Wellnhofer 1993; Elzanowski
and Wellnhofer 1996). In the past decade, the application
of new techniques, such as computed tomography and new
methods in UV photography, have led to additional insights
into several anatomical details (Dominguez Alonso et al.
2004; Tischlinger and Unwin 2004; Tischlinger 2005).
Finally, the discovery of a new specimen with a beautifully
preserved skull and skeleton provided important new
information on many aspects of the anatomy of Archae-
opteryx (Mayr et al. 2005, 2007). Nevertheless, many
details of the skull anatomy of this taxon, especially the
configuration of the temporal region and the braincase,
have remained enigmatic.
The aim of this article is not to present a full description
of the cranial osteology of Archaeopteryx, but to help
clarify some details of its cranial anatomy and to provide a
new reconstruction of the skull. For more general accounts
of the osteology of this taxon, the reader is referred to the
excellent descriptions of Wellnhofer (1974, 2008), Elza-
nowski and Wellnhofer (1996), and Elzanowski (2002),
and to the numerous other contributions that have helped
clarify the cranial structure of this taxon (e.g., Dames 1884;
Heilmann 1926; Whetstone 1983; Walker 1985; Domin-
guez Alonso et al. 2004; Tischlinger 2005; Mayr et al.
2007).
For this study, the skulls of mainly two specimens were
studied in detail, the 7th (Munich) and the 10th (Ther-
mopolis) specimens. The Eichstatt, Berlin, Solnhofen, and
Daiting (8th) specimens were also examined first hand, as
were a large number of non-avian theropod specimens.
Comparative information for basal birds was mainly taken
from the literature.
The specimens were examined using a binocular
microscope and high-resolution UV photographs, gener-
ously provided by Helmut Tischlinger.
Institutional abbreviations
BSPG Bayerische Staatssammlung fur Palaontologie und
Geologie, Munich, Germany; IGM Institute of Geology,
Ulan Bataar, Mongoloia; MB Museum fur Naturkunde
Berlin, Germany; WDC Wyoming Dinosaur Center, Ther-
mopolis, Wyoming, USA.
Configuration of the skull roof
The 10th (Thermopolis) specimen (WDC-CSG-100) has
the dorsal skull roof preserved in exquisite detail (Fig. 1).
Many of the important characters have already been
reported by Mayr et al. (2007), so only a few details will be
pointed out here.
The premaxilla has a long anterior body, in which all
tooth positions are placed anterior to the external nares
(Fig. 1a). At the anterior end of the premaxilla, an
enlarged, anteriorly facing foramen is present, as in many
theropod dinosaurs (e.g., Rauhut et al. 2010). As mentioned
by Mayr et al. (2007: 101), another large foramen is present
anterodorsal to the anteriormost end of the external nares.
This foramen is connected to the anterodorsal margin of the
external nares by an elongate furrow and thus opens pos-
terolaterally (Figs. 1a, 5). Such a foramen is also present in
the Eichstatt specimen (pers. obs.), but not the London
(Wellnhofer 2008: Fig. 5.26), Solnhofen (pers. obs.), and
Berlin specimens (Tischlinger 2005: Fig. 10; Wellnhofer
2008: Fig. 5.48 b). The Thermopolis specimen further
confirms the reconstruction of the relation among the
premaxilla, maxilla, and nasal in Archaeopteryx by
Wellnhofer (1974, 2008). Thus, the dorsal nasal process of
212 O. W. M. Rauhut
123

the premaxilla is approximately 175 % of the length of the
main premaxillary body and almost reaches the posterior
end of the external nares. The ventral posterior process of
the premaxilla is much shorter than the dorsal process and
flanks the ventral margin of the nares for approximately
half of its length. However, it is slightly longer than the
premaxillary body, slender, and rod-like, and not as
abbreviated as illustrated by Elzanowski (2001). Although
the nasal has a slender anterior subnarial process, the latter
is clearly separated from the ventral posterior process of
the premaxilla so that the maxilla forms part of the ventral
margin of the nares, as argued by Wellnhofer (1974, 2008).
On the lateral side of the skull roof, the configuration of
the contacts among the nasal, lacrimal and maxilla largely
conforms to the situation found in basal theropods (Fig. 1b,
c). As in Zupaysaurus (Ezcurra 2007), Allosaurus (Madsen
1976), Sinraptor (Currie and Zhao 1994a), and other taxa,
but in contrast to the reconstructions of Elzanowski (2001)
and Wellnhofer (2008), the posterior end of the ascending
process of the maxilla is forked to receive a pointed
Fig. 1 Cranial anatomy of the 10th (Thermopolis) specimen of
Archaeopteryx (WDC-CSG-100). a Photograph of skull under
ultraviolet light, with indications of enlarged areas. b Enlargement
of lateral edge of left nasal, showing lateral process of the nasal.
c Enlargement of the contact between right maxilla, nasal, and
lacrimal. d Enlargement of the lacrimal fenestra. e Enlargement of the
ascending process of the jugal. en external nares, f frontal, fo foramina
in the premaxilla, j jugal, l lacrimal, lf lacrimal fenestra, lp lateral
process of nasal, m maxilla, mf maxillary fenestra, mu manual ungual,
n nasal, pa parietal, pm premaxilla, pof postorbital facet on the
ascending process of the jugal, pro premaxillary foramen, sc scleral
ring. Scale bar in a is 5 mm
Skull of Archaeopteryx 213
123

anterior process of the lacrimal (Fig. 1c). Furthermore, the
lateral margin of the nasal has a small posterolateral pro-
cess that embraced the anterior end of the dorsal surface of
the lacrimal (Fig. 1b), as is present in Syntarsus (Bristowe
and Raath 2004), Allosaurus (Madsen 1976), Sinraptor
(Currie and Zhao 1994a: Fig. 3a), and many basal saur-
opodomorphs (e.g., Yates 2003: Fig. 10b).
The Thermopolis specimen also helps to clarify some
aspects of the paranasal sinus system. Although Wellnhofer
(1974) identified both a maxillary fenestra and a large,
laterally facing promaxillary fenestra in the Eichstatt
specimen, Elzanowski (2001, 2002) argued that the dorsal
part of the ascending process of the maxilla represented a
nasal capsule, representing a rostral ethmoid ossification,
and thus reconstructed the skull without maxillary fenestra.
Wellnhofer (2008) again reconstructed the skull with both
a maxillary and promaxillary fenestra, but figured the two
considerably smaller than in his 1974 reconstruction. As
noted by Mayr et al. (2007) [see also Xu et al. 2011)], the
Thermopolis specimen confirms the interpretation of
Wellnhofer (1974), in that Archaeopteryx had a large,
semicircular maxillary fenestra and a more anteriorly
placed and smaller, laterally facing promaxillary foramen
(Fig. 1a). It is thus similar to Anchiornis (Hu et al. 2009)
and Compsognathus (BSPG AS 563), whereas in most
coelurosaurs, the premaxillary fenestra is concealed in the
lateral view (e.g., tyrannosaurids: Currie 2003; Carr and
Williamson 2004; ornithomimosaurs: Osmolska et al.
1972; Ji et al. 2003; oviraptorosaurs: Clark et al. 2002;
troodontids: Norell et al. 2009). In many dromaeosaurs, the
premaxillary fenestra is small and at least partially exposed
laterally (e.g., Ostrom 1969; Barsbold and Osmolska 1999;
Xu and Wu 2001; Burnham 2004; Norell et al. 2006), but it
is placed anteroventral to the maxillary fenestra and is
relatively smaller than in Archaeopteryx.
In the Thermopolis specimen, the lacrimal shows a large
recess in the posterodorsal corner laterally (Fig. 1d). The
recess is filled with matrix, so nothing can be said about its
medial extent, but it is in the same position as the lacrimal
fenestra in basal tetanurans such as Allosaurus (Madsen
1976) and Sinraptor (Currie and Zhao 1994a). In paravian
theropods, a lacrimal recess has otherwise only been men-
tioned, but not yet been described in detail, in Deinonychus
(Witmer 1997a).
One of the most significant new observations concerns
the configuration of the temporal region of Archaeopteryx.
In the Thermopolis specimen, no postorbital is preserved,
but the short dorsal process of the jugal is present and
shows a slightly depressed facet on the anterior side of its
dorsal part (Fig. 1e). This facet is slightly offset from
the ventral part of the anterior margin of the process and
slopes more strongly posterodorsally than the latter. A very
similar though not as conspicuous facet is also visible in
the medially exposed left jugal of the Munich specimen
(BSPG 1999 I 50). This facet fits the jugal facet for the
connection with the postorbital in non-avian theropods in
both its position and the details of its morphology (angu-
lation of the facet in relation to the ascending process of the
jugal; facet facing more laterally than medially) and thus
clearly indicates that a jugal-postorbital contact was pres-
ent in Archaeopteryx.
Finally, the area identified as the quadrate cotyle in the
squamosal of the Munich specimen by Elzanowski and
Wellnhofer (1996) most probably represents a slightly
depressed facet for the contact with the paroccipital process
of the braincase on a rather long posterior process (Fig. 2),
as is also present in dromaeosaurids (e.g., Ostrom 1969;
Barsbold and Osmolska 1999).
Braincase
The dorsal parts of the braincase of Archaeopteryx are
preserved in the London specimen and have been
described by de Beer (1954), Whetstone (1983), Walker
(1985) and Dominguez Alonso et al. (2004). The ventral
and lateral parts of the braincase are preserved in the
Munich specimen (BSPG 1999 I 50) and have been
described by Wellnhofer (1993) and Elzanowski and
Wellnhofer (1996). This specimen is redescribed here,
since detailed investigation of the braincase under both
normal and UV light led to some re-interpretations of
structures (Figs. 2, 3).
Elzanowski and Wellnhofer (1996) considered the cra-
nial base of the Munich specimen to be exposed in lateral
view. However, re-examination of the specimen indicates
that the basisphenoid and basioccipital are exposed in the
ventrolateral view (Fig. 3). Thus, the ventral side of both
bones and both left and right basal tubera and basipterygoid
processes are visible, though the left basipterygoid process
is broken and partially preserved on the counterslab
(Fig. 2). The occipital condyle is almost aligned with the
ventral surface of the basisphenoid and only slightly offset
dorsally, in contrast to most theropods, although this might
be partially due to preservation, as noted by Elzanowski
and Wellnhofer (1996). The posteroventral side of the
basioccipital anterior to the condyle is flat, with a very
shallow longitudinal groove extending along the midline
from the occipital condyle to the basioccipital-basi-
sphenoid suture. The basal tubera are small, placed far
laterally, and separated by a wide, U-shaped incision.
The ventral side of the basisphenoid is elongate, its
length between the basal tubera and the basipterygoid
processes being approximately 1.8 times the minimal
width. Posteriorly, the basisphenoid forms the anterior
half of the basal tubera. Immediately anterior to the
214 O. W. M. Rauhut
123
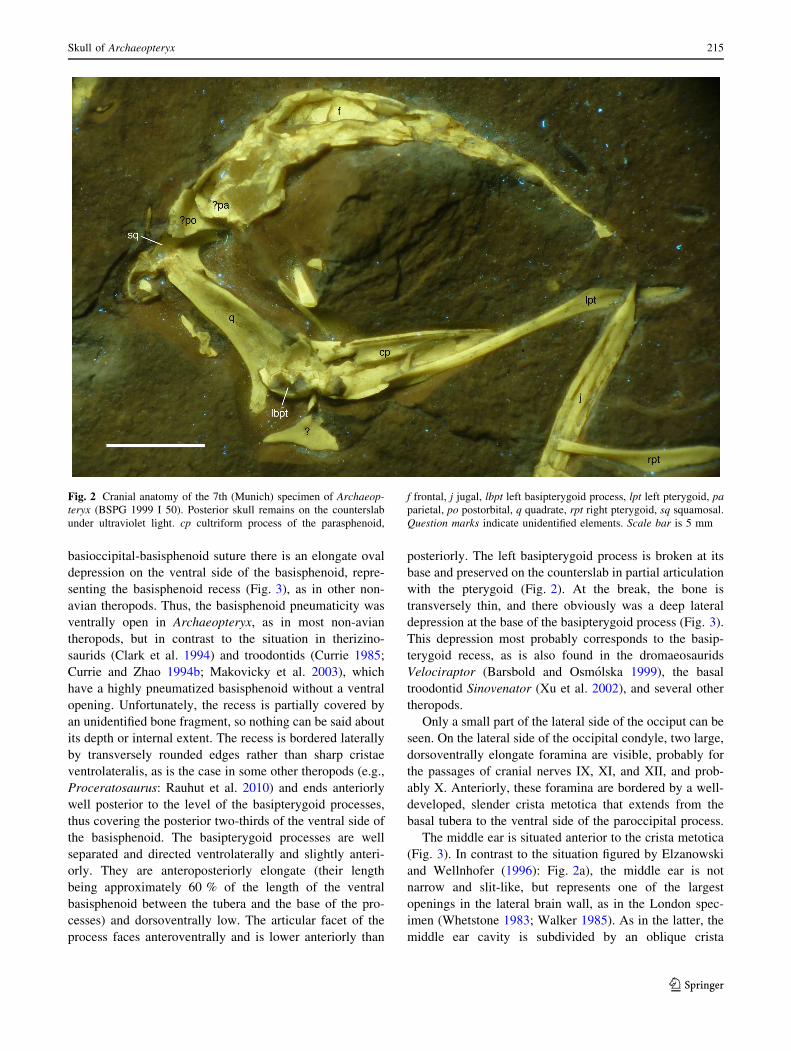
basioccipital-basisphenoid suture there is an elongate oval
depression on the ventral side of the basisphenoid, repre-
senting the basisphenoid recess (Fig. 3), as in other non-
avian theropods. Thus, the basisphenoid pneumaticity was
ventrally open in Archaeopteryx, as in most non-avian
theropods, but in contrast to the situation in therizino-
saurids (Clark et al. 1994) and troodontids (Currie 1985;
Currie and Zhao 1994b; Makovicky et al. 2003), which
have a highly pneumatized basisphenoid without a ventral
opening. Unfortunately, the recess is partially covered by
an unidentified bone fragment, so nothing can be said about
its depth or internal extent. The recess is bordered laterally
by transversely rounded edges rather than sharp cristae
ventrolateralis, as is the case in some other theropods (e.g.,
Proceratosaurus: Rauhut et al. 2010) and ends anteriorly
well posterior to the level of the basipterygoid processes,
thus covering the posterior two-thirds of the ventral side of
the basisphenoid. The basipterygoid processes are well
separated and directed ventrolaterally and slightly anteri-
orly. They are anteroposteriorly elongate (their length
being approximately 60 % of the length of the ventral
basisphenoid between the tubera and the base of the pro-
cesses) and dorsoventrally low. The articular facet of the
process faces anteroventrally and is lower anteriorly than
posteriorly. The left basipterygoid process is broken at its
base and preserved on the counterslab in partial articulation
with the pterygoid (Fig. 2). At the break, the bone is
transversely thin, and there obviously was a deep lateral
depression at the base of the basipterygoid process (Fig. 3).
This depression most probably corresponds to the basip-
terygoid recess, as is also found in the dromaeosaurids
Velociraptor (Barsbold and Osmolska 1999), the basal
troodontid Sinovenator (Xu et al. 2002), and several other
theropods.
Only a small part of the lateral side of the occiput can be
seen. On the lateral side of the occipital condyle, two large,
dorsoventrally elongate foramina are visible, probably for
the passages of cranial nerves IX, XI, and XII, and prob-
ably X. Anteriorly, these foramina are bordered by a well-
developed, slender crista metotica that extends from the
basal tubera to the ventral side of the paroccipital process.
The middle ear is situated anterior to the crista metotica
(Fig. 3). In contrast to the situation figured by Elzanowski
and Wellnhofer (1996): Fig. 2a), the middle ear is not
narrow and slit-like, but represents one of the largest
openings in the lateral brain wall, as in the London spec-
imen (Whetstone 1983; Walker 1985). As in the latter, the
middle ear cavity is subdivided by an oblique crista
Fig. 2 Cranial anatomy of the 7th (Munich) specimen of Archaeop-
teryx (BSPG 1999 I 50). Posterior skull remains on the counterslab
under ultraviolet light. cp cultriform process of the parasphenoid,
f frontal, j jugal, lbpt left basipterygoid process, lpt left pterygoid, pa
parietal, po postorbital, q quadrate, rpt right pterygoid, sq squamosal.
Question marks indicate unidentified elements. Scale bar is 5 mm
Skull of Archaeopteryx 215
123

interfenestralis (only the dorsal base of which is preserved;
Fig. 3) into the slightly smaller foramen ovale anterodor-
sally and the foramen pseudorotunda posteroventrally, thus
confirming the configuration of this region as reconstructed
by Walker ( 1985: Fig. 4). The left paroccipital process has
been sheared off and displaced dorsally, as noted by
Elzanowski and Wellnhofer (1996), so that its anteroventral
side is exposed (Fig. 3). As in most theropods, a shallow
stapedial groove extends along the anteroventral side of the
process distally. Proximally, a large, oval opening is present
at the base of this groove, as is also found in the base of the
paroccipital process of the London specimen (Whetstone
1983; Walker 1985); this opening corresponds to the
entrance of the posterior tympanic recess in other coeluro-
saurs (e.g., Clark et al. 1994; Norell et al. 2006).
The prootic of the Munich specimen has been well
described and figured by Elzanowski and Wellnhofer
(1996). However, the separate bone identified as the broken
prootic wing by these authors most probably rather repre-
sents the laterosphenoid (Fig. 3), as originally suggested by
Wellnhofer (1993). Thus, the prootic pendant (Sampson
and Witmer 2007; ‘‘prootic wing’’ of Elzanowski and
Wellnhofer 1996) is considerably smaller than recon-
structed by Elzanowski and Wellnhofer (1996) and
directed ventrally (Figs. 3, 4), as in other theropods, rather
than anteroventrally. The laterosphenoid is a subquadran-
gular element with a stout lateral process anterodorsally for
Fig. 3 Cranial anatomy of the 7th (Munich) specimen of Archaeop-
teryx (BSPG 1999 I 50). Posterior skull elements and braincase on the
main slab. a Photograph under ultraviolet light. b Interpretative
drawing. at anterior tympanic recess, boc basioccipital, bpt basip-
terygoid process, bptr basipterygoid recess, bs basisphenoid, bsr
basisphenoid recess, bt basal tubera, cif crista interfenestralis, cm
crista metotica, dtr dorsal tympanic recess, ec ectopterygoid fragment,
epi epipterygoid, f frontal, fo fenestra ovale, fp fenestra pseudoro-
tunda, ls laterosphenoid, oc occipital condyle, pa parietal, pap
paroccipital process, pro prootic, pt pterygoid, ptr posterior tympanic
recess, qw quadrate wing of the pterygoid, sa surangular, sg stapedial
groove. Roman numerals denote cranial nerves; question marks
indicate unidentified elements. Scale bar is 5 mm
Fig. 4 Schematic reconstruction of the braincase of Archaeopteryx,
based on the 7th specimen. Note that most sutures are conjectural,
since the elements are partially disarticulated and damaged, so their
courses should not be used for phylogenetic coding. Abbreviations as
in Figs. 2 and 3; op opisthotic, pp preotic pendant
216 O. W. M. Rauhut
123

the contact with the frontal and, possibly, the postorbital, as
in other theropods. However, the element has been flat-
tened into the bedding plane, so that the orientation of this
process is not as obvious, but the lateral convexity of the
main laterosphenoid body and the concavity at the base of
the lateral process are still visible (Fig. 3a). In the London
specimen, only the lateral process of the laterosphenoid
seems to be preserved (Whetstone 1983). The laterosphe-
noid is not pierced by any foramina in its lateral part,
indicating that the exit for the ophthalmic branch of the
trigeminal nerve was not separate from the maxillary and
mandibular branches, as is the case in Troodon (Currie and
Zhao 1994b).
Thus, the trigeminal foramen in Archaeopteryx seems to
be somewhat smaller than reconstructed by Walker (1985)
and Elzanowski and Wellnhofer (1996). The laterosphe-
noid in the Munich specimen is displaced ventrally, and the
margin that probably contacted the prootic is partially
covered by unidentified bone fragments, but the incision
for the trigeminal opening in this bone, if present, seems to
have been rather small. Whetstone (1983) figured a small
incision in the posterodorsal rim of this opening and ten-
tatively identified it as the trigeminal foramen. The same
feature was refigured, but not discussed by Elzanowski and
Wellnhofer ( 1996: 5B), but Walker (1985) argued that the
posterior margin of the trigeminal foramen was incomplete
and identified this incision as part of a pneumatic recess. In
many saurischians, the exit of the mid-cerebral vein is
separated from the exit of the trigeminal nerve and is
developed as a small foramen in the dorsal margin of the
trigeminal opening or somewhat more dorsally (Rauhut
2003). Interestingly, this opening is positioned on the
prootic-latereosphenoid suture in some taxa (e.g., Drom-
aeosaurus: Currie 1995) or entirely in the laterosphenoid
(e.g., Troodon: Rauhut 2003), but it is entirely within the
prootic, at the posterodorsal rim of the trigeminal foramen,
and thus in a very similar position to this incision in
Plateosaurus (MB.R. 5586.1). Thus, this incision might
represent the exit of the mid-cerebral vein. In the Munich
specimen, there is a posterior rim around the trigeminal
foramen (Fig. 3), as figured for the London specimen by
Walker ( 1985: Fig. 4), and there seems to be a matrix-
filled depression or opening in the elevated posterodorsal
margin of this depression. This depression or opening is in
the same position as the incision figured by Whetstone
(1983) in the London specimen, thus supporting Walker’s
interpretation that the posterior rim of the trigeminal
foramen is broken in this specimen, and the opening might
represent a small separate foramen rather than an incision
branching off the trigeminal foramen.
Below the opening for the seventh cranial nerve, a large
depression is present on the ventral side of the prootic, as in
the London specimen (Whetstone 1983; Walker 1985).
Wellnhofer (1993) identified this matrix-filled structure as
the fenestra ovalis, but Walker (1985) argued that pneu-
matic openings were located within this depression.
Indeed, the depression is in exactly the position covered by
the anterior tympanic recess in non-avian theropods (e.g.,
Raath 1985; Currie and Zhao 1994b; Witmer 1997b;
Makovicky et al. 2003; Rauhut 2004; Paulina-Carabajal
and Currie 2012) and thus most probably represents this
structure. This recess is partially covered by a broken,
plate-like bone, which probably represents remains of the
quadrate wing of the pterygoid. At the level of the anterior
end of the prootic, a slender, triangular bone sits laterally
on this bony plate (Fig. 3). This bone tapers dorsally and
reaches approximately the level of the dorsal margin of the
trigeminal foramen. In position and shape, it is similar to
the epipterygoid in other theropods (e.g., Clark et al. 2002;
Eddy and Clarke 2011; Brusatte et al. 2012a) and might
thus represent this element.
Discussion
The observations presented here help to clarify some
details of the cranial anatomy of Archaeopteryx that might
be of interest for both functional and phylogenetic studies.
The configuration of the dorsal skull roof, especially the
detailed sutures between the nasal, lacrimal, and maxilla,
corresponds well to the situation in basal tetanuran thero-
pods in which the anterior process of the lacrimal slots into
the forked posterior end of the ascending process of the
maxilla and the dorsal surface of the lacrimal is braced
anteriorly by a posterolaterally directed lateral process of
the nasal (e.g., Allosaurus: Madsen 1976; Sinraptor: Currie
and Zhao 1994a). Interestingly, most reconstructions of
skulls of paravian theropods show the anterior end of the
lacrimal overlapping the posterior end of the ascending
process of the maxilla dorsally and lack a lateral process of
the nasal (e.g., Barsbold and Osmolska 1999; Xu and Wu
2001; Burnham 2004; Xu et al. 2011). More detailed
studies of the cranial sutures in paravian theropods are
necessary to decide whether this indicates a remarkable
retention of the plesiomorphic condition in Archaeopteryx
or if this might simply reflect lack of detail in available
reconstructions; the fact that the skull of the ‘‘fighting
dinosaur’’ specimen of Velociraptor (IGM 100/25) seems
to show a slender anterior end of the lacrimal that slots into
the forked posterior end of the ascending process of the
maxilla (Barsbold and Osmolska 1999: Fig. 1a; Norell
et al. 2006: Fig. 6b) indicates that the latter might at least
account for some of this apparent variation. However, the
dorsal view of the same specimen does not show any
evidence of a lateral process of the nasal (Barsbold and
Osmolska 1999: Fig. 2b).
Skull of Archaeopteryx 217
123

One of the most significant findings is that of an articular
facet for the postorbital on the ascending process of the
jugal (Fig. 1e), presenting evidence for a closed postorbital
bar, as originally hypothesized by Wellnhofer (1974) and
Elzanowski and Wellnhofer (1996) on the basis of the
presence of a postorbital process of the jugal. Several
previous analyses of the available skull material of
Archaeopteryx failed to recognize a postorbital, resulting in
the hypothesis that this element was absent and the orbit
open posteriorly, as in recent birds, by Buhler (1985),
Martin (1991) and Elzanowski (2001, 2002). This alleged
absence of a postorbital bar was one of the key arguments
that led Buhler (1985) to propose a bird-like cranial kinesis
in Archaeopteryx. Even if a complete absence of a post-
orbital was not assumed and after UV analyses by Tisch-
linger (2005) unequivocally showed a postorbital to be
present in this taxon, many authors retained an open
postorbital bar in their reconstruction to allow for cranial
kinesis (e.g., Chiappe 2007; Wellnhofer 2008). Wellnhofer
(2008: Fig. 6.3) proposed that, in a prokinetic movement,
the nasal might move over the frontals in Archaeopteryx.
The presence of a theropod-like articular facet on the
ascending process of the jugal for the postorbital calls the
interpretation of a prokinetic skull in Archaeopteryx into
question. According to this hypothesis, the streptostylic
quadrate would move the anterior part of the skull via the
jugal bar (see Buhler 1985; Wellnhofer 2008). However, a
closed postorbital bar would severely hamper any antero-
posterior movement of the jugal and thus the mechanism
by which the snout is allegedly raised. Furthermore, the
sutures between the jugal and postorbital, and, as far as can
be said, between the nasal and frontal (based on the
Thermopolis and Eichstatt specimens) correspond well to
the situation in other theropods, and there is no indication
for either a flexible overlap between the nasal and frontal
nor for a flexible bending zone within the nasal (as
hypothesized by Buhler, 1985). Thus, the situation is
comparable to that in non-avian theropods, for which
Holliday and Witmer (2008) concluded that there is little
evidence for any sophisticated kinesis. These findings are
also in agreement with the skull configuration reported in
the more advanced basal avialian Confuciusornis, which
also has a closed postorbital bar (Peters and Ji 1998; Chi-
appe et al. 1999; Hou et al. 1999), and the possibility that at
least some Enantiornithes still retained a postorbital-jugal
contact (Wang et al. 2010). This indicates that the opening
of the postorbital bar and the development of the cranial
kinesis typical for modern birds happened later in avialian
evolution.
The new interpretation of the braincase of Archaeop-
teryx indicates that this structure is closely comparable to
that in other non-avian maniraptorans (Fig. 4). The
description of the middle ear cavity largely confirms the
interpretation of Walker (1985) and shows close similarity
to deinonychosaurs, such as Byronosaurus (Makovicky
et al. 2003). The lateral braincase wall is closely compa-
rable to that of dromaeosaurids, in both the position of
nerve foramina and the presence and position of an anterior
tympanic recess and a basipterygoid recess (e.g., Barsbold
and Osmolska 1999). The braincase differs from that of
troodontids in that the basisphenoid recess is ventrally open
and the exit for the trigeminal nerve is not subdivided,
Fig. 5 Revised reconstruction of the skull of Archaeopteryx, mainly
based on the Eichstatt and Thermopolis specimens. A angular, aof
antorbital fenestra, ar articular, d dentary, en external nares, f frontal,
fao fossa antorbitalis, fo foramen, itf infratemporal fenestra, j jugal,
l lacrimal, lf lacrimal fenestra, m maxilla, mf maxillary fenestra,
n nasal, o orbit, pa parietal, pap paroccipital process, pm premaxilla,
po postorbital, pro premaxillary foramen quadrate, qj quadratojugal,
sa surangular, sq squamosal. Modified from Rauhut (2003). Scale bar
is 10 mm
218 O. W. M. Rauhut
123

indicating that the ‘‘inflated’’ basisphenoid without a ven-
tral recess and subdivided trigeminal foramen in birds
arose independently from the situation in that clade. With
the presence of anterior, posterior, and dorsal tympanic
recesses, a basisphenoid recess and basipterygoid recesses,
basicranial pneumaticity is also closely comparable to that
in many coelurosaurs (Witmer 1997b).
Conclusions
Detailed re-examinations of the skulls of two specimens of
Archaeopteryx show that the skull of this primeval bird is
more theropod-like than previously recognized (Fig. 5).
The detailed sutures between the nasal, lacrimal and
maxilla are closely comparable to those in more basal
theropods. These findings contrast with the reconstructions
of the skulls of other advanced coelurosaurians, which,
however, might be due to the lack of detail in at least some
available reconstructions. Given that the exact position and
morphology of cranial sutures is important for interpreta-
tions of cranial function (e.g., Weishampel 1984; Rayfield
2005), landmark-based analyses of cranial shape (e.g.,
Brusatte et al. 2012b; Foth and Rauhut (2013), and phy-
logenetic analyses, more detailed anatomical studies of
cranial sutures in coelurosaurs and basal birds and inves-
tigations of the evolution of these structures are needed.
An articular facet on the ascending process of the jugal
is closely comparable to the postorbital articulation in non-
avian theropods, indicating a closed postorbital bar.
Together with the lack of indications for a preorbital
bending zone in the dorsal skull roof, this makes the
presence of a bird-like prokinetic skull in Archaeopteryx
unlikely.
Cranial pneumaticity in Archaeopteryx conforms to that
seen in non-avian coelurosaurs. This applies to both para-
nasal pneumaticity, with the development of a maxillary
fenestra, premaxillary foramen, and lacrimal fenestra (see
Witmer 1997a), and basicranial pneumaticity. The latter
includes three tympanic recesses (anterior, posterior and
dorsal), basipterygoid recesses, and a basisphenoid recess.
All of these pneumatic features are in the same position and
show the same development as in many non-avian coel-
urosaurs, such as dromaeosaurids.
Acknowledgments This article resulted from the Archaeopteryx
event during the Munich Mineralientage in 2009, during which six of
the original Archaeopteryx specimens were gathered together. Special
thanks are therefore due to Christoph Keilmann, who made this event
possible, and the institutions and private individuals who made their
specimens of the Urvogel available during the event. Very special
thanks are furthermore due to Burkhard Pohl for the loan of the
Thermopolis specimen after the event and to Helmut Tischlinger for
UV photography. The article benefited from discussions with Chris-
tian Foth, Xu Xing, and Adriana Lopez-Arbarello, and from financial
support by the Volkswagen Foundation under grant AZ I/84 640.
Gerald Mayr is thanked for a critical review of the manuscript.
References
Barsbold, R., and H. Osmolska. 1999. The skull of Velociraptor
(Theropoda) from the Late Cretaceous of Mongolia. Acta
Palaeontologica Polonica 44(2): 189–219.
Bergmann, U., R.W. Morton, P.L. Manning, W.I. Sellers, S. Farrar,
K.G. Huntley, R.A. Wogelius, and P. Larson. 2010. Archaeop-
teryx feathers and bone chemistry fully revealed via synchrotron
imaging. Proceedings of the National Academy of Sciences
107(20): 9060–9065.
Bristowe, A., and M.A. Raath. 2004. A juvenile coelophysoid skull
from the Early Jurassic of Zimbabwe, and the synonymy of
Coelophysis and Syntarsus. Palaeontologia Africana 40: 31–41.
Brusatte, S.L., T.D. Carr, and M.A. Norell. 2012a. The osteology of
Alioramus, a gracile and long-snouted tyrannosaurid (Dinosau-
ria: Theropoda) from the Late Cretaceous of Mongolia. Bulletin
of the American Museum of Natural History 366: 1–197.
Brusatte, S.L., S. Manabu, S. Montanari, and W.E.H. Harcourt Smith.
2012b. The evolution of cranial form and function in theropod
dinosaurs: insights from geometric morphometrics. Journal of
Evolutionary Biology 25: 365–377.
Buhler, P. 1985. On the morphology of the skull of Archaeopteryx. In
The beginning of birds, ed. M.K. Hecht, J.H. Ostrom, G. Viohl,
and P. Wellnhofer, 135–140. Eichstatt: Freunde des Jura
Museums.
Burnham, D.A. 2004. New information on Bambiraptor feinbergi
(Theropoda: Dromaeosauridae) from the Late Cretaceous of
Montana. In Feathered dragons. Studies on the transition from
dinosaurs to birds, ed. P.J. Currie, E.B. Koppelhus, M.A.
Shugar, and J.L. Wright, 67–111. Bloomington & Indianapolis:
Indiana University Press.
Carr, T.D., and T.E. Williamson. 2004. Diversity of late Maastrich-
tian Tyrannosauridae (Dinosauria: Theropoda) from western
North America. Zoological Journal of the Linnean Society 142:
479–523.
Chiappe, L.M. 2007. Glorified dinosaurs: the origin and early
evolution of birds. London: Wiley-Liss.
Chiappe, L.M., S.-A. Ji, Q. Ji, and M.A. Norell. 1999. Anatomy and
systematics of the Confucuisornithidae (Theropoda: Aves) from
the Late Mesozoic of northeastern China. Bulletin of the
American Museum of Natural History 242: 1–89.
Clark, J.M., M.A. Norell, and T. Rowe. 2002. Cranial anatomy of
Citipati osmolskae (Theropoda, Oviraptorosauria), and a rein-
terpretation of the holotype of Oviraptor philoceratops. Amer-
ican Museum Novitates 3364: 1–24.
Clark, J.M., A. Perle, and M.A. Norell. 1994. The skull of
Erlicosaurus andrewsi, a Late Cretaceous ‘‘segnosaur’’ (Thero-
poda: Therizinosauridae) from Mongolia. American Museum
Novitates 3115: 1–39.
Currie, P.J. 1985. Cranial anatomy of Stenonychosaurus inequalis
(Saurischia, Theropoda) and its bearing on the origin of birds.
Canadian Journal of Earth Sciences 22: 1643–1658.
Currie, P.J. 1995. New information on the anatomy and relationships
of Dromaeosaurus albertensis (Dinosauria: Theropoda). Journal
of Vertebrate Paleontology 15(3): 576–591.
Currie, P.J. 2003. Cranial anatomy of tyrannosaurid dinosaurs from
the Late Cretaceous of Alberta. Canada. Acta Palaeontologica
Polonica 48(2): 191–226.
Currie, P.J., and X.-J. Zhao. 1994a. A new carnosaur (Dinosauria,
Theropoda) from the Jurassic of Xinjiang, People’s Republic of
China. Canadian Journal of Earth Sciences 30: 2037–2081.
Skull of Archaeopteryx 219
123

Currie, P.J., and X.-J. Zhao. 1994b. A new troodontid (Dinosauria,
Theropoda) braincase from the Dinosaur Park Formation
(Campanian) of Alberta. Canadian Journal of Earth Sciences
30: 2231–2247.
Dames, W. 1884. Ueber Archaeopteryx. Palaeontologische Abhandl-
ungen 2(3): 119–196.
Darwin, C. 1859. On the origin of species by means of natural
selection. London: Murrey.
De Beer, G. 1954. Archaeopteryx lithographica. A study based on the
British Museum specimen. London: British Museum (Natural
History).
Dominguez Alonso, P., A.C. Milner, R.A. Ketcham, M.J. Cookson,
and T.B. Rowe. 2004. The avian nature of the brain and inner ear
of Archaeopteryx. Nature 430: 666–669.
Eddy, D.R., and J.A. Clarke. 2011. New information on the cranial
anatomy of Acrocanthosaurus atokensis and its implications for
the phylogeny of Allosauroidea (Dinosauria: Theropoda). PLoS
ONE 6(3): e17932.
Elzanowski, A. 2001. A novel reconstruction of the skull of
Archaeopteryx. Netherlands Journal of Zoology 51(2): 207–215.
Elzanowski, A. 2002. Archaeopterygidae (Upper Jurassic of Ger-
many). In Mesozoic birds. Above the heads of dinosaurs, ed.
L.M. Chiappe, and L.M. Witmer, 129. Berkeley: University of
California Press.
Elzanowski, A., and P. Wellnhofer. 1996. Cranial morphology of
Archaeopteryx: evidence from the seventh skeleton. Journal of
Vertebrate Paleontology 16(1): 81–94.
Erickson, G.M., O.W.M. Rauhut, Z. Zhou, A.H. Turner, B.D. Inouye,
D. Hu, and M.A. Norell. 2009. Was dinosaurian physiology
inherited by birds? Reconciling slow growth in Archaeopteryx.
PLoS ONE 4(10): 1–9.
Ezcurra, M.D. 2007. The cranial anatomy of the coelophysoid
theropod Zupaysaurus rougieri from the Upper Triassic of
Argentina. Historical Biology 19(2): 185–202.
Foth, C., and O.W.M. Rauhut. 2013. Macroevolutionary and
morphofunctional patterns in theropod skulls: a morphometric
approach. Acta Palaeontologica Polonica 58(1): 1–16.
Heilmann, G. 1926. The origin of birds. London: Witherby.
Holliday, C.M., and L.M. Witmer. 2008. Cranial kinesis in dinosaurs:
intracranial joints, protractor muscles, and their significance for
cranial evolution and function in diapsids. Journal of Vertebrate
Paleontology 28(4): 1073–1088.
Hou, L., L.D. Martin, Z. Zhou, A. Feduccia, and F. Zhang. 1999. A
diapsid skull in a new species of the primitive bird Confuciu-
sornis. Nature 399: 679–682.
Hu, D., L. Hou, L. Zhang, and X. Xu. 2009. A pre-Archaeopteryx
troodontid theropod from China with long feathers on the
metatarsus. Nature 461: 640–643.
Huxley, T.H. 1868. On the animals which are most nearly interme-
diate between birds and reptiles. Annals and Magazin of Natural
History 4: 66–75.
Ji, Q., M.A. Norell, P.J. Makovicky, K.-Q. Gao, S. Ji, and C.-X. Yuan.
2003. An early ostrich dinosaur and implications for ornithomi-
mosaur phylogeny. American Museum Novitates 3420: 1–12.
Longrich, N.R., J. Vinther, Q. Meng, Q. Li, and A.P. Russell. 2012.
Primitive wing feather arrangement in Archaeopteryx litho-
graphica and Anchiornis huxleyi. Current Biology 22:
2262–2267.
Madsen, J.H. 1976. Allosaurus fragilis: a revised osteology. Utah
Geological and Mineralogical Survey Bulletin 109: 3–163.
Makovicky, P.J., M.A. Norell, J.M. Clark, and T. Rowe. 2003.
Osteology and relationships of Byronosaurus jaffei (Theropoda:
troodontidae). American Museum Novitates 3402: 1–32.
Martin, L.D. 1991. Mesozoic birds and the origin of birds. In Origins
of higher groups of tetrapods, ed. H.-P. Schultze, and L. Trueb,
485–540. Ithaka and London: Comstock Publishing Associate.
Mayr, G., B. Pohl, S. Hartman, and D.S. Peters. 2007. The tenth
skeletal specimen of Archaeopteryx. Zoological Journal of the
Linnean Society 149: 97–116.
Mayr, G., B. Pohl, and D.S. Peters. 2005. A well-preserved
Archaeopteryx specimen with theropod features. Science 310:
1483–1486.
Norell, M.A., J.M. Clark, A.H. Turner, P.J. Makovicky, R. Barsbold,
and T. Rowe. 2006. A new dromaeosaurid theropod from Ukhaa
Tolgod (Omnogov, Mongolia). American Museum Novitates
3545: 1–51.
Norell, M.A., P.J. Makovicky, G.S. Bever, A.M. Balanoff, J.M. Clark,
R. Barsbold, and T. Rowe. 2009. A review of the Mongolian
Cretaceous dinosaur Saurornithoides (Troodontidae: Thero-
poda). American Museum Novitates 3654: 1–63.
Osmolska, H., E. Roniewicz, and R. Barsbold. 1972. A new dinosaur,
Gallimimus bullatus n. gen., n. sp. (Ornithomimidae) from the Upper
Cretaceous of Mongolia. Palaeontologia Polonica 27: 103–143.
Ostrom, J.H. 1969. Osteology of Deinonychus antirrhopus, an
unusual theropod from the Lower Cretaceous of Montana.
Bulletin of the Peabody Museum of Natural History 30: 1–165.
Ostrom, J.H. 1973. The ancestry of birds. Nature 242: 136.
Ostrom, J.H. 1976. Archaeopteryx and the origin of birds. Biological
Journal of the Linnean Society 8: 91–182.
Paulina-Carabajal, A., and P.J. Currie. 2012. New information on the
braincase of Sinraptor dongi (Theropoda: allosauroidea): eth-
moidal region, endocranial anatomy, and pneumaticity. Verte-
brata Palasiatica 50(2): 85–101.
Peters, S., and Q. Ji. 1998. The diapsid temporal construction of the
Chinese fossil bird Confuciusornis. Senckenbergiana Lethaea
78: 155–158.
Raath, M.A. 1985. The theropod Syntarsus and its bearing on the
origin of birds. In The beginning of birds, ed. M.K. Hecht, J.H.
Ostrom, G. Viohl, and P. Wellnhofer, 219–227. Eichstatt:
Freunde des Jura Museums.
Rauhut, O.W.M. 2003. The interrelationships and evolution of basal
theropod dinosaurs. Special Papers in Palaeontology 69: 1–213.
Rauhut, O.W.M. 2004. Braincase structure of the Middle Jurassic
theropod dinosaur Piatnitzkysaurus. Canadian Journal of Earth
Sciences 41(9): 1109–1122.
Rauhut, O.W.M., A.C. Milner, and S.C. Moore-Fay. 2010. Cranial
osteology and phylogenetic position of the theropod dinosaur
Proceratosaurus bradleyi (Woodward, 1910) from the Middle
Jurassic of England. Zoological Journal of the Linnean Society
158: 155–195.
Rayfield, E.J. 2005. Using Finite-Element Analysis to investigate
suture morphology: a case study using large carnivorous
dinosaurs. Anatomical Record A 283A: 349–365.
Sampson, S.D., and L.M. Witmer. 2007. Craniofacial anatomy of
Majungsaurus crenatissimus (Theropoda: abelisauridae) from
the Late Cretaceous of Madagascar. Society of Vertebrate
Paleontology, Memoir 8: 32–102.
Tischlinger, H. 2005. Neue Informationen zum Berliner Exemplar
von Archaeopteryx lithographica H. v. Meyer 1861. Archaeop-
teryx 23: 33–50.
Tischlinger, H. 2009. Der achte Archaeopteryx–das Daitinger Exem-
plar. Archaeopteryx 27: 1–20.
Tischlinger, H., and D.M. Unwin. 2004. UV-Untersuchungen des
Berliner Exemplars von Archaeopteryx lithographica H. v.
Meyer 1861 und der isolierten Archaeopteryx-Feder. Archaeop-
teryx 22: 17–50.
von Meyer, H. 1861. Archaeopteryx lithographica und Pterodactylus.
Neues Jahrbuch fuer Mineralogie, Geognosie, Geologie und
Petrefaktenkunde 1861: 678–679.
Walker, A. 1985. The braincase of Archaeopteryx. In The beginning
of birds, ed. M.K. Hecht, J.H. Ostrom, G. Viohl, and P.
Wellnhofer, 123–134. Eichstatt: Freunde des Jura Museums.
220 O. W. M. Rauhut
123

Wang, X., J.K. O’Connor, B. Zhao, L.M. Chiappe, C. Gao, and X.
Cheng. 2010. New species of Enantiornithes (Aves: ornithotho-
races) from the Qiaotou Formation in northern Hebei, China.
Acta Geologica Sinica 84: 247–256.
Weishampel, D.B. 1984. Evolution of jaw mechanisms in ornithopod
dinosaurs. Advancements in Anatomy, Embryology and Cell
Biology 87: 1–110.
Wellnhofer, P. 1974. Das funfte Skelettexemplar von Archaeopteryx.
Palaeontographica A 147(4–6): 169–216.
Wellnhofer, P. 1993. Das siebte Exemplar von Archaeopteryx aus den
Solnhofener Schichten. Archaeopteryx 11: 1–47.
Wellnhofer, P. 2008. Archaeopteryx. Der Urvogel von Solnhofen.
Munich: Dr. Friedrich Pfeil.
Whetstone, K.N. 1983. Braincase of Mesozoic birds: I. New
preparation of the ‘‘London’’ Archaeopteryx. Journal of Verte-
brate Paleontology 2(4): 439–452.
Witmer, L.M. 1997a. The evolution of the antorbital cavity of
archosaurs: a study in soft-tissue reconstruction in the fossil
record with an analysis of the function of pneumaticity. Society
of Vertebrate Paleontology, Memoir 3: 1–73.
Witmer, L.M. 1997b. Craniofacial air sinus systems. In Encyclopedia
of dinosaurs, ed. P.J. Currie, and K. Padian, 151–159. San
Diego: Academic Press.
Xu, X., and M.A. Norell. 2006. Non-avian dinosaur fossils from the
Lower Cretaceous Jehol Group of western Liaoning, China.
Geological Journal 41: 419–437.
Xu, X., M.A. Norell, X.-L. Wang, P.J. Makovicky, and X.-C. Wu.
2002. A basal troodontid from the early Cretaceous of China.
Nature 415: 780–784.
Xu, X., and X.-C. Wu. 2001. Cranial morphology of Sinornithosaurus
millenii Xue et al. 1999 (Dinosauria: Theropoda: Dromaeosau-
ridae) from the Yixian Formation of Liaoning, China. Canadian
Jorunal of Earth Sciences 38: 1739–1752.
Xu, X., H.-L. You, K. Du, and F. Han. 2011. An Archaeopteryx-like
theropod from China and the origin of Avialae. Nature 475: 465–470.
Yates, A.M. 2003. The species taxonomy of the sauropodomorph
dinosaurs from the Lowenstein Formation (Norian, Late Trias-
sic) of Germany. Palaeontology 46(2): 317–337.
Zhou, Z., and F. Zhang. 2006. Mesozoic birds of China: A synoptic
review. Vertebrata Palasiatica 44(1): 74–98.
Skull of Archaeopteryx 221
123




















