Neuroscience 2013 Syllabus v13.1.0 - Department of ...
138
Medical Neuroscience Syllabus - Block 1 Spring 2013 Version 13.1.0 Offered and Coordinated by the Department of Neurobiology and Anatomy The University of Texas Health Science Center at Houston Contents © 2000-Present University of Texas Health Science Center at Houston. All Rights Reserved. Unauthorized use of contents subject to civil and/or criminal prosecution. To access Adobe Acrobat PDF versions of the course syllabus as well as other course information, visit the official course web address: http://nba.uth.tmc.edu/courses/neuroscience/
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of Neuroscience 2013 Syllabus v13.1.0 - Department of ...

Gross Anatomy Fall 2008 Syllabus – Block I
Offered and Coordinated by the Department of Neurobiology and AnatomyThe University of Texas Health Science Center at Houston
Contents © 2000-Present University of Texas Health Science Center at Houston. All Rights Reserved. Unauthorized use of contents subject to civil and/or criminal prosecution.
All Netter images © ICON Learning Systems. Used by permission. Radiographic images are from www.webanatomy.com by Stephen P. Raskin, M.D. and Raymond J. Walsh, Ph.D.,
The George Washington University Medical Center. Copyright © 1996 Scholar Educational Systems, Inc.
To access Adobe Acrobat PDF versions of the course syllabus as well as other course information, visit the official course website:
http://nba.uth.tmc.edu/courses/gross/
Medical Neuroscience
Syllabus - Block 1Spring 2013
Version 13.1.0
Offered and Coordinated by the Department of Neurobiology and AnatomyThe University of Texas Health Science Center at Houston
Contents © 2000-Present University of Texas Health Science Center at Houston. All Rights Reserved. Unauthorized use of contents subject to civil and/or criminal prosecution.
To access Adobe Acrobat PDF versions of the course syllabus as well as othercourse information, visit the official course web address:http://nba.uth.tmc.edu/courses/neuroscience/

Medical Neuroscience 2013 : Block 1
Page 1Table of Contents
Table of ConTenTs
Introduction to Neuroscience ................................................................................................................ 4Some Major Diseases of the Nervous System........................................................................................ 5Introduction to Neurons and Neural Networks ...................................................................................... 5The Neuron ............................................................................................................................................. 6Resting Potentials and Action Potentials ................................................................................................ 7Neuronal Networks............................................................................................................................... 11Summary .............................................................................................................................................. 19
Synaptic Formation/Development and Regeneration ......................................................................... 20Introduction .......................................................................................................................................... 21Axon Pathfinding ................................................................................................................................. 22Multiple Guidance Mechanisms Direct Growing Axons Simultaneously ........................................... 25Presynaptic and Postsynaptic Differentiation ....................................................................................... 25Synapse Elimination ............................................................................................................................. 26Growth Factors ..................................................................................................................................... 27Trophic Factors are Important for Development and Injury Repair ..................................................... 28Neurodegeneration ............................................................................................................................... 28
Organization of Cell Types ................................................................................................................. 30Introduction to Neurons and Glial Cells............................................................................................... 31Cell Soma ............................................................................................................................................. 31Dendrites .............................................................................................................................................. 32Initial Segment and Axon Hillock ....................................................................................................... 32Axon .................................................................................................................................................... 33Nerve Endings ..................................................................................................................................... 33Cellular Elements at the Typical Nerve Terminal ............................................................................... 33Structural Variations ............................................................................................................................. 34Naming Neurons................................................................................................................................... 35Glial Cells and Function ....................................................................................................................... 35Macroglia ............................................................................................................................................. 36Protoplasmic and Fibrous Astrocytes ................................................................................................... 36Oligodendroglia .................................................................................................................................... 37Ependyma ............................................................................................................................................. 37Microglia .............................................................................................................................................. 38
Resting Potentials and Action Potentials ............................................................................................ 39Features of Action Potentials ................................................................................................................ 40Intracellular Recordings from Neurons ................................................................................................ 41Components of the Action Potentials ................................................................................................... 44Ionic Mechanisms of Resting Potentials .............................................................................................. 44Goldman-Hodgkin and Katz (GHK) Equation ..................................................................................... 45Membrane Potential Laboratory ........................................................................................................... 46
Ionic Mechanisms of Action Potentials .............................................................................................. 47Voltage-Dependent Conductances ........................................................................................................ 49Sequence of Conductance Changes Underlying the Nerve Action Potential ....................................... 50Na+ Inactivation .................................................................................................................................... 50Voltage-Dependent K+ Conductance .................................................................................................... 50Pharmacology of the Voltage-dependent Membrane Channels ............................................................ 51Pumps and Leaks .................................................................................................................................. 52Types of Membrane Channels .............................................................................................................. 53Channelopaties ..................................................................................................................................... 53Absolute and Relative Refractory Periods ........................................................................................... 54Action Potential Laboratory ................................................................................................................. 54
Propagation of Action Potentials ........................................................................................................ 55Changes in the Spatial Distribution of Charge .................................................................................... 56Determinants of Propagation Velocity ................................................................................................. 56Propagation in Myelinated Fibers ........................................................................................................ 59

Medical Neuroscience 2013 : Block 1
Page 2Table of Contents
Synaptic Transmission at Skeletal Neuromuscular Junction and Mechanisms of Neurotransmitter Release ................................................................................................................................................ 60
Anatomy of the Neuromuscular Junction ............................................................................................. 61Physiology of Synaptic Transmission at the Neuromuscular Junction ................................................ 62Propagation of the EPP ......................................................................................................................... 63Overview of the Sequence of Events Underlying the EPP ................................................................... 64Role of AChE ....................................................................................................................................... 64Myasthenia Gravis ................................................................................................................................ 64Iontophoresis of ACh ........................................................................................................................... 65Ionic Mechanisms of the EPP............................................................................................................... 66Calcium Hypotheses for Chemical Synaptic Transmission ................................................................. 67Quantal Nature of Transmitter Release ................................................................................................ 67
Synaptic Transmission in the Central Nervous System/Synaptic Plasticity ....................................... 70Synaptic Transmission in a Simple Reflex Circuit ............................................................................... 71Ionic Mechanisms of IPSPs .................................................................................................................. 72Synaptic Potentials ............................................................................................................................... 72Differences between the EPSP at the Skeletal Neuromuscular Junction and EPSPs in the CNS ........ 72Temporal and Spatial Summation ........................................................................................................ 73IPSPs .................................................................................................................................................... 74Ionic Mechanisms for IPSPs ................................................................................................................ 74Transmitter Substance of the Spinal Inhibitory Neuron ....................................................................... 74Metabotropic Synaptic Responses........................................................................................................ 74Neurotoxins .......................................................................................................................................... 76Summary .............................................................................................................................................. 81
Transport and Molecular Mechanisms of Secretion ........................................................................... 82Introduction .......................................................................................................................................... 83Synthesis of Vesicles and Proteins ....................................................................................................... 84Perinuclear Cisternae and Ribosomal Protein Synthesis ...................................................................... 84Rough Endoplasmic Reticulum ............................................................................................................ 85Golgi ..................................................................................................................................................... 85Anterograde Transport of Vesicles ....................................................................................................... 86Secretory Mechanism ........................................................................................................................... 86Vesicle Recapture ................................................................................................................................. 88Retrograde Axoplasmic Transport ........................................................................................................ 88
Acetylcholine Neurotransmission ....................................................................................................... 90Introduction ......................................................................................................................................... 91Acetylcholine in the Autonomic Nervous System ............................................................................... 91ACh in the Peripheral Nervous System ................................................................................................ 91ACh in the Central Nervous System .................................................................................................... 92Introduction to the Cell Biology of the Cholinergic Synapse ............................................................. 92Synthesis of ACh .................................................................................................................................. 93Storage of ACh ..................................................................................................................................... 93Release of ACh .................................................................................................................................... 94ACh Receptors ..................................................................................................................................... 94The Nicotinic Receptor is an Ion Channel .......................................................................................... 95The Muscarinic Receptor is coupled to G-Proteins ............................................................................. 96Physiology ............................................................................................................................................ 98Behavior ............................................................................................................................................... 98Clinical ................................................................................................................................................ 98Cholinergic Pharmacological Agents .................................................................................................. 99
Biogenic Amine Neurotransmitters ................................................................................................... 100Introduction ........................................................................................................................................ 101Anatomy of Catecholamines .............................................................................................................. 101Cell Biology ....................................................................................................................................... 102Storage of Monoamines ..................................................................................................................... 104Release of Monoamines ..................................................................................................................... 104

Medical Neuroscience 2013 : Block 1
Page 3Table of Contents
Properties of Monoamine Receptors .................................................................................................. 104NE and E Peripheral Receptors .......................................................................................................... 105CNS NE and DA Receptors................................................................................................................ 105Inactivation of MA Neurotransmitters by Reuptake and Metabolism ............................................... 106Reuptake of MA Neurotransmitters ................................................................................................... 106Dopamine - Physiological and Behavioral Actions ............................................................................ 107Norepinephrine - Physiological and Behavioral Actions ................................................................... 108Serotonin - Physiological and Behavioral Actions ............................................................................. 108Dopamine - Clinical Importance and Pharmacology ......................................................................... 109Norepinephrine - Clinical Importance and Pharmacology ................................................................. 109Serotonin - Clinical Importance and Pharmacology .......................................................................... 109Other Clinical Uses of 5-HT Drugs .................................................................................................... 110
Amino Acid Neurotransmitters ......................................................................................................... 111Introduction and Review .................................................................................................................... 112Spatial and Morphological Distinctions between Excitatory and Inhibitory Inputs .......................... 112Structure of Amino Acid Transmitters ............................................................................................... 113Biosynthesis of Amino Acid Neurotransmitters ................................................................................. 114Glutamate and Aspartate .................................................................................................................... 114Glycine ............................................................................................................................................... 114GABA ................................................................................................................................................. 115Ca2+-Dependent Release ..................................................................................................................... 115Receptors for Amino Acid Neurotransmitters .................................................................................... 116Glutamate Receptors .......................................................................................................................... 116Receptors-GABAA and Glycine ......................................................................................................... 118GABA Receptors ................................................................................................................................ 118Characteristics of GABAA Receptor................................................................................................... 118G-protein Coupled Glutamate and GABAB Receptors....................................................................... 119Clinical Manifestations of Altered Glutamate Levels ........................................................................ 120Diseases Associated with GABA ....................................................................................................... 120
Neuropeptides and Nitric Oxide ....................................................................................................... 121Introduction ........................................................................................................................................ 122Neuropeptides ..................................................................................................................................... 122Classification of Peptides by Families ............................................................................................... 123Biosynthesis and Regulation .............................................................................................................. 123Multiple Mechanisms are Utilized to Produce the Diversity of Neuropeptides................................. 124Release................................................................................................................................................ 125Termination of Action ........................................................................................................................ 126Receptors are all G-protein Linked .................................................................................................... 126Nitric Oxide (NO) .............................................................................................................................. 126Characteristics of NO ......................................................................................................................... 127Summary of NO’s Properties ............................................................................................................. 127“Receptors” for NO ............................................................................................................................ 128Synthesis by Nitric Oxide Synthase (NOS) and Release ................................................................... 128Biological Effects of NO .................................................................................................................... 129Vasodilator .......................................................................................................................................... 129Neuromodulator.................................................................................................................................. 130Toxicity ............................................................................................................................................... 130
Principles of Structural Neuroimaging ............................................................................................. 131Introduction to Neuroimaging ............................................................................................................ 132X-Ray Computed Tomography (CT) ................................................................................................. 132Magnetic Resonance Imaging (MRI) ................................................................................................ 132Positron Emission Tomography (PET) and Single Photon Emission Computed Tomography (SPECT) 134Functional Neuroimaging ................................................................................................................... 135Examples of Images From Each Modality ......................................................................................... 135

Medical Neuroscience 2013 : Block 1
Page 4Introduction to Neuroscience
Introduction to NeuroscienceLecturer: John H. Byrne, Ph.D.
January 7, 2013 | 8:00 AM
assignmenT
I. Importance of Neuroscience in MedicineII. Major Diseases of the Nervous System
III. Introduction to Neurons and Neural NetworksA. The Neuron
1. Resting Potentials and Action Potentials2. Synaptic Potentials and Synaptic Integration
B. Neuronal Networks1. Micronetwork Motifs2. Feedforward Excitation and Feedforward Inhibition3. Convergence and Divergence4. Lateral Inhibition5. Feedback/Recurrent Inhibition6. Feedback/Recurrent Excitation
C. Summary
major objeCTives
1. Know the major diseases of the nervous system and their mechanisms2. Know the basic morphological and electrophysiological properties of neurons3. Know the common microcircuit motifs and their functions
required reading
● Byrne, J.H. Introduction to Neuroscience, Lecture Supplement Volume I.; or ● Byrne, J.H. Introduction to Neurons and Neural Networks, Neuroscience Online:
http://nba.uth.tmc.edu/neuroscience/s1/introduction.html
suggesTed reading
● Byrne, J. H., Roberts, J. L., From Molecules to Networks: An Introduction to Cellular and Molecular Neuroscience, 2nd Edition, Elsevier, Amsterdam, 2009, Chapter 19.
Medical Neuroscience 2013 : Block 1
Page 5Introduction to Neuroscience
The three pounds of jelly-like material found within our skulls is the most complex machine on Earth and perhaps the universe. Its phenomenal features would not be possible without the hundreds of billions of neurons that make it up, and, importantly, the connections between those neurons. Fortunately, much is known about the properties of individual neurons and simple neuronal networks, and aspects of complex neuronal networks are beginning to be unraveled. This chapter will begin with a discussion of the neuron, the elementary node or element of the brain, and then move to a discussion of the ways in which individual neurons communicate with each other. What makes the nervous system such a fantastic device and distinguishes the brain from other organs of the body is not that it has 100 billion neurons, but that nerve cells are capable of communicating with each other in such a highly structured manner as to form neuronal networks. To understand neural networks, it is necessary to understand the ways in which one neuron communicates with another through synaptic connections and the process called synaptic transmission. Synaptic transmission comes in two basic flavors: excitation and inhibition. Just a few interconnected neurons (a microcircuit) can perform sophisticated tasks such as mediate reflexes, process sensory information, generate locomotion and mediate learning and memory. More complex networks (macrocircuits) consist of multiple imbedded microcircuits. Macrocircuits mediate higher brain functions such as object recognition and cognition. So, multiple levels of networks are ubiquitous in the nervous system. Networks are also prevalent within neurons. These nanocircuits constitute the underlying biochemical machinery for mediating key neuronal properties such as learning and memory and the genesis of neuronal rhythmicity.
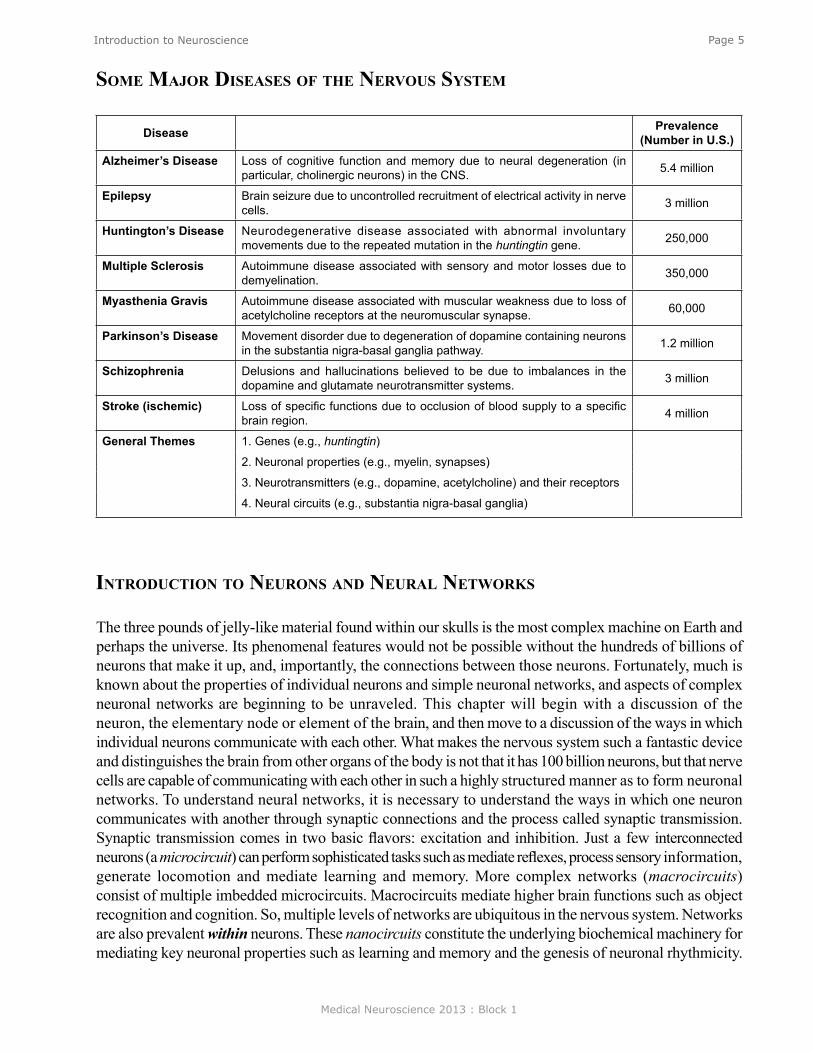
Disease Prevalence (Number in U.S.)
Alzheimer’s Disease Loss of cognitive function and memory due to neural degeneration (in particular, cholinergic neurons) in the CNS. 5.4 million
Epilepsy Brain seizure due to uncontrolled recruitment of electrical activity in nerve cells. 3 million
Huntington’s Disease Neurodegenerative disease associated with abnormal involuntary movements due to the repeated mutation in the huntingtin gene. 250,000
Multiple Sclerosis Autoimmune disease associated with sensory and motor losses due to demyelination. 350,000
Myasthenia Gravis Autoimmune disease associated with muscular weakness due to loss of acetylcholine receptors at the neuromuscular synapse. 60,000
Parkinson’s Disease Movement disorder due to degeneration of dopamine containing neurons in the substantia nigra-basal ganglia pathway. 1.2 million
Schizophrenia Delusions and hallucinations believed to be due to imbalances in the dopamine and glutamate neurotransmitter systems. 3 million
Stroke (ischemic) Loss of specific functions due to occlusion of blood supply to a specific brain region. 4 million
General Themes 1. Genes (e.g., huntingtin)
2. Neuronal properties (e.g., myelin, synapses)
3. Neurotransmitters (e.g., dopamine, acetylcholine) and their receptors
4. Neural circuits (e.g., substantia nigra-basal ganglia)
some major diseases of The nervous sysTem
inTroduCTion To neurons and neural neTworks

Medical Neuroscience 2013 : Block 1
Page 6Introduction to Neuroscience
The neuron
Basic Morphological Features oF NeuroNsThe 100 billion neurons in the brain share a number of common features (Figure 1). Neurons are different from most other cells in the body in that they are polarized and have distinct morphological regions, each with specific functions. Dendrites are the region where one neuron receives connections from other neurons. The cell body or soma contains the nucleus and the other organelles necessary for cellular function. The axon is a key component of nerve cells over which information is transmitted from one part of the neuron (e.g., the cell body) to the terminal regions of the neuron. Axons can be rather long extending up to a meter or so in some human sensory and motor nerve cells. The synapse is the terminal region of the axon and it is here where one neuron forms a connection with another and conveys information through the process of synaptic transmission (see Neuroscience Online for a colored version). The aqua-colored neuron in Figure 1 is referred to as the postsynaptic neuron. The tan-colored terminal to the left is consequently referred to as the presynaptic neuron. One neuron can receive contacts from many different neurons. Figure 1 shows an example of three presynaptic neurons contacting the one tan-colored postsynaptic neuron, but it has been estimated that one neuron can receive contacts from up to 10,000 other cells. Consequently, the potential complexity of the networks is vast. Similarly, any one neuron can contact up to 10,000 postsynaptic cells. (Note that the tan-colored neuron that was presynaptic to the aqua-colored neuron is postsynaptic to the pink, green, and blue neurons. So most “presynaptic” neurons are “postsynaptic” to some other neuron(s).
Figure 1 also shows an expanded view of the synapse. Note that the presynaptic cell is not directly connected to the postsynaptic cell. The two are separated by a gap known as the synaptic cleft. Therefore, to communicate with the postsynaptic cell, the presynaptic neuron needs to release a chemical messenger. That messenger is found within the neurotransmitter-containing vesicles (the blue dots represent the neurotransmitter). An action potential that invades the presynaptic terminal causes these vesicles to fuse with the inner surface of the presynaptic membrane and release their contents th rough a p rocess called exocytosis. The released transmitter diffuses across the gap between the pre- and the postsynaptic cell and very rapidly reaches the postsynaptic side of the synapse where it binds to specialized receptors that “recognize” the t r a n s m i t t e r . T h e binding to the receptors leads to a change in the permeability of ion channels in the membrane and in turn Figure 1
Medical Neuroscience 2013 : Block 1
Page 7Introduction to Neuroscience
a change in the membrane potential of the postsynaptic neuron known as a postsynaptic synaptic potential (PSP). So signaling among neurons is associated with changes in the electrical properties of neurons. To understand neurons and neuronal circuits, it is necessary to understand the electrical properties of nerve cells.
resTing PoTenTials and aCTion PoTenTials
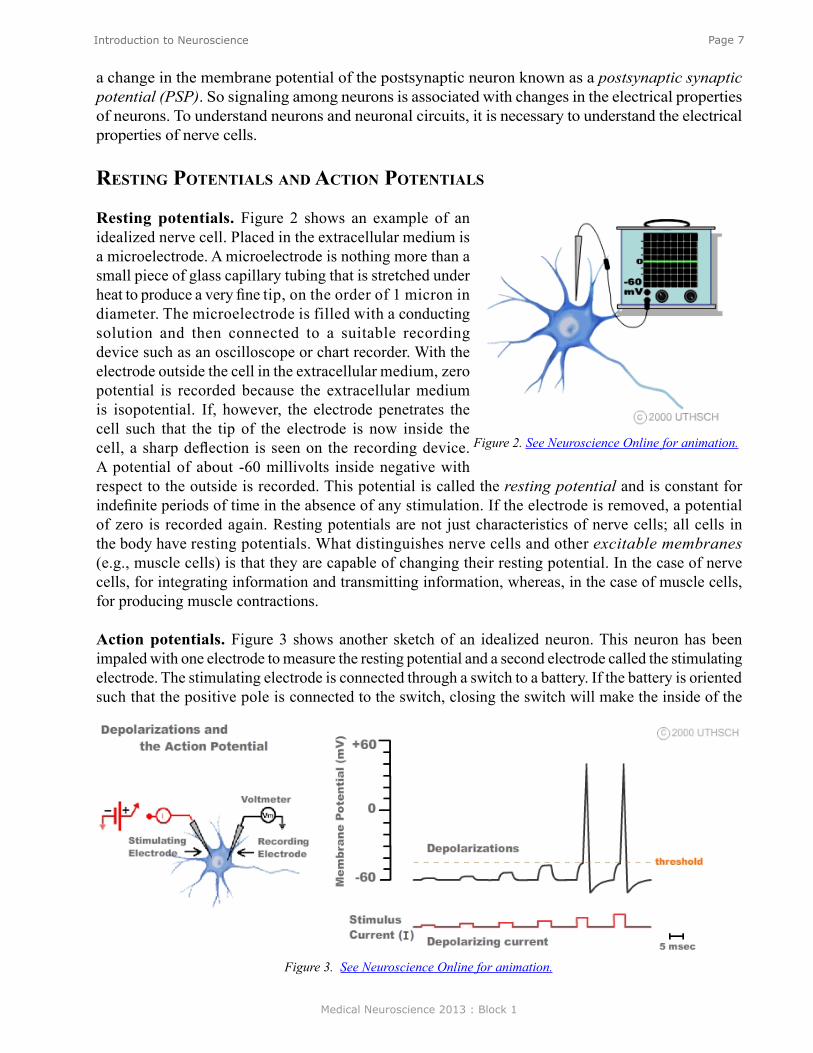
Resting potentials. Figure 2 shows an example of an idealized nerve cell. Placed in the extracellular medium is a microelectrode. A microelectrode is nothing more than a small piece of glass capillary tubing that is stretched under heat to produce a very fine tip, on the order of 1 micron in diameter. The microelectrode is filled with a conducting solution and then connected to a suitable recording device such as an oscilloscope or chart recorder. With the electrode outside the cell in the extracellular medium, zero potential is recorded because the extracellular medium is isopotential. If, however, the electrode penetrates the cell such that the tip of the electrode is now inside the cell, a sharp deflection is seen on the recording device. A potential of about -60 millivolts inside negative with respect to the outside is recorded. This potential is called the resting potential and is constant for indefinite periods of time in the absence of any stimulation. If the electrode is removed, a potential of zero is recorded again. Resting potentials are not just characteristics of nerve cells; all cells in the body have resting potentials. What distinguishes nerve cells and other excitable membranes (e.g., muscle cells) is that they are capable of changing their resting potential. In the case of nerve cells, for integrating information and transmitting information, whereas, in the case of muscle cells, for producing muscle contractions.
Action potentials. Figure 3 shows another sketch of an idealized neuron. This neuron has been impaled with one electrode to measure the resting potential and a second electrode called the stimulating electrode. The stimulating electrode is connected through a switch to a battery. If the battery is oriented such that the positive pole is connected to the switch, closing the switch will make the inside of the
Figure 2. See Neuroscience Online for animation.
Figure 3. See Neuroscience Online for animation.

Medical Neuroscience 2013 : Block 1
Page 8Introduction to Neuroscience
cell somewhat more positive depending upon the size of the battery. (Such a decrease in the polarized state of a membrane is called a depolarization.) Figure 3 is an animation (see Neuroscience Online) in which the switch is repeatedly opened and closed and each time it is closed a larger battery is applied to the circuit. Initially, the switch closure produces only small depolarizations. However, the potentials become larger and eventually the depolarization is sufficiently large to trigger an action potential, also known as a spike or an impulse. The action potential is associated with a very rapid depolarization to achieve a peak value of about +40 mV in just 0.5 milliseconds (msec). The peak is followed by an equally rapid repolarization phase.
The voltage at which the depolarization becomes sufficient to trigger an action potential is called the threshold. If a larger battery is used to generate a suprathreshold depolarization, a single action potential is still generated and the amplitude of that action potential is the same as the action potential trigged by a just-threshold stimulus. The simple recording in Figure 3 illustrates two very important features of action potentials. First, they are elicited in an all-or-nothing fashion. Either an action potential is elicited with stimuli at or above threshold, or an action potential is not elicited. Second, action potentials are very brief events of only about several milliseconds in duration. Initiating an action potential is somewhat analogous to applying match to a fuse. A certain temperature is needed to ignite the fuse (i.e., the fuse has a threshold). A match that generates a greater amount of heat than the threshold temperature will not cause the fuse to burn any brighter or faster. Just as action potentials are elicited in an all-or-nothing fashion, they are also propagated in an all-or-nothing fashion. Once an action potential is initiated in one region of a neuron such as the cell body, that action potential will propagate along the axon (like a burning fuse) and ultimately invade the synapse where it can initiate the process of synaptic transmission.
In the example in Figure 3, only a single action potential was generated because the duration of each of the two suprathreshold stimuli was so brief that sufficient time was only available to initiate a single action potential (i.e., the stimulus ended before the action potential completed its depolarization-repolarization cycle). But, as shown in the animations of Figure 4 (see Neuroscience Online), longer-duration stimuli can lead to the initiation of multiple action potentials, the frequency of which is dependent on the intensity of the stimulus. Therefore, it is evident that the nervous system encodes information not in terms of the changes in the amplitude of action potentials, but rather in their frequency. This is a very universal property. The greater the intensity of a mechanical stimulus to a touch receptor, the greater the number of action potentials; the greater the amount of stretch to a muscle stretch receptor, the greater the number of action potentials; the greater the intensity of a light, the greater the number of action potentials that is transmitted to the central nervous system. Similarly, in the motor system, the greater the number of action potentials in a motor neuron, the greater will be the contraction of the muscle that receives a synaptic connection from that motor neuron. Engineers call this type of information coding pulse frequency modulation.

Medical Neuroscience 2013 : Block 1
Page 9Introduction to Neuroscience
Figure 4. See Neuroscience Online for animation.

Medical Neuroscience 2013 : Block 1
Page 10Introduction to Neuroscience
syNaptic poteNtials aNd syNaptic iNtegratioNFigure 5 illustrates three neurons. The one colored green will be referred to as an excitatory neuron for reasons that will become clear shortly. It makes a connection to the postsynaptic neuron colored blue. The traces below (see Neuroscience Online for animation) illustrate the consequences of initiating an action potential in the green neuron. That action potential in the presynaptic neuron leads to a decrease in the membrane potential of the postsynaptic cell. The membrane potential changes from its resting value of about -60 millivolts to a more depolarized state. This potential is called an excitatory postsynaptic potential (EPSP). It is “excitatory” because it moves the membrane potential toward the threshold and it is “postsynaptic” because it is a potential recorded on the postsynaptic side of the synapse. Generally (and this is an important point), a single action potential in a presynaptic cell does not produce an EPSP large enough to reach threshold and trigger an action potential. But, if multiple action potentials are fired in the presynaptic cell, the corresponding multiple excitatory potentials can summate through a process called temporal summation to reach threshold and triggering an action potential. EPSPs can be viewed as a “go signal” to the postsynaptic neuron to transmit information through a network pathway.
The red-colored neuron in Figure 5 is referred to as an inhibitory neuron. Like the green neuron, it also makes a synaptic contact with the blue postsynaptic neuron. It also releases a chemical transmitter messenger, but the consequences of the transmitter from the blue cell binding to receptors on the postsynaptic cell is opposite to the consequences of the transmitter released by the green neuron. The consequence of action potential in the red presynaptic neuron is to produce an increase in the membrane potential of the blue postsynaptic neuron. The membrane potential is more negative than it was before (a hyperpolarization) and therefore the membrane potential is farther away from threshold. This type of potential is called an inhibitory postsynaptic potential (IPSP) because it tends to prevent the postsynaptic neuron from firing an action potential. This is a “stop signal” for the postsynaptic cell. So the green neuron says “go” and the red neuron says “stop”. Now what is the postsynaptic neuron to do?
Neurons are like adding machines. They are constantly adding up the excitatory and the in-hibitory synaptic input in time (temporal summation) and over the area of the dendrites receiving synaptic contacts (spatial summation), and if that summation is at or above threshold they fire an action potential. If the sum is below threshold, no action potential is initiated. This is a process called synaptic integration and is illustrated in Figure 5. Initially, two action potentials in the green neuron produced summating EPSPs that fired an action potential in the blue neuron. But, if an IPSP from the inhibitory neuron occurs just before two action potentials in the excitatory neu-ron, the summation of the one IPSP and the two EPSPs is below threshold and no action potential is elicited in the postsynaptic cell. The inhibitory neuron (and inhibition in general) is a way of gating or regulating the ability of an excitatory signal to fire a postsynaptic cell.
Figure 5. See Neuroscience Online for animation.

Medical Neuroscience 2013 : Block 1
Page 11Introduction to Neuroscience
neuronal neTworks
MicroNetwork MotiFsAs indicated earlier in the chapter, a neuron can receive contacts from up to 10,000 presynaptic neurons, and, in turn, any one neuron can contact up to 10,000 postsynaptic neurons. The combinatorial possibility could give rise to enormously complex neuronal circuits or network topologies, which might be very difficult to understand. But despite the potential vast complexity, much can be learned about the functioning of neuronal circuits by examining the properties of a subset of simple circuit configurations. Figure 6 illustrates some of those microcircuit or micronetwork motifs. Although simple, they can do much of what needs to be done by a nervous system.Feedforward excitation. Allows one neuron to relay information to its neighbor. Long chains of these can be used to propagate information through the nervous system.
Feedforward inhibition. A presynaptic cell excites an inhibitory interneuron (an interneuron is a neuron interposed between two neurons) and that inhibitory interneuron then inhibits the next follower cell. This is a way of shutting down or limiting excitation in a downstream neuron in a neural circuit.
Convergence/Divergence. One postsynaptic cell receives convergent input from a number of different presynaptic cells and any individual neuron can make divergent connections to many different postsynaptic cells. Divergence allows one neuron to communicate with many other neurons in a network. Convergence allows a neuron to receive input from many neurons in a network.
Lateral inhibition. A presynaptic cell excites inhibitory interneurons and they inhibit neighboring cells in the network. As described in detail later in the Chapter, this type of circuit can be used in sensory systems to provide edge enhancement.
Figure 6.

Medical Neuroscience 2013 : Block 1
Page 12Introduction to Neuroscience
Feedback/recurrent inhibition. In Panel E1, a presynaptic cell connects to a postsynaptic cell, and the postsynaptic cell in turn connects to an interneuron, which then inhibits the presynaptic cell. This circuit can limit excitation in a pathway. Some initial excitation would be shut off after the red interneuron becomes active. In Panel E2, each neuron in the closed chain inhibits the neuron to which it is connected. This circuit would appear to do nothing, but, as will be seen later in the Chapter, it can lead to the generation of complex patterns of spike activity.
Feedback/recurrent excitation. In Panel F1, a presynaptic neuron excites a postsynaptic neuron and that postsynaptic neuron excites the presynaptic neuron. This type of circuit can serve a switch-like function because once the presynaptic cell is activated that activation could be perpetuated. Activation of the presynaptic neuron could switch this network on and it could stay on. Panel F2 shows variants of feedback excitation in which a presynaptic neuron excites a postsynaptic neuron that can feedback to excite itself (a, an autapse) or other neurons which ultimately feedback (b) to itself.
These simple motifs are ubiquitous components of many neural circuits. Let’s examine some examples of what these networks can do.
FeedForward excitatioN aNd FeedForward iNhiBitioNOne of the best understood microcircuits is the circuit that mediates simple reflex behaviors. Figure 7 illustrates the circuit for the so-called knee jerk or stretch reflex. A neurologist strikes the knee with a rubber tapper, which elicits an extension of the leg. This test is used as a simple way to examine the integrity of some of the sensory and motor pathways in the spinal cord. The tap of hammer stretches the muscle and leads to the initiation of action potentials in sensory neurons within the muscle that are sensitive to stretch. (The action potentials are represented by the small bright “lights” in the animation (see Neuroscience Online).) The action potentials are initiated in an all-or-nothing fashion and propagate into the spinal cord where the axon splits (bifurcates) into two branches.
Let’s first discuss the branch to the left that forms a synaptic connection (green triangle) with an Extensor (E) motor neuron (colored blue). The action potential in the sensory neuron invades the synaptic terminal of the sensory neuron causing the release of transmitter and subsequent excitation of the motor neuron. The stretch to the muscle leads to an action potential in the motor neuron (MN), which then propagates out the peripheral nerve to invade the synapse at the muscle, causing the release of transmitter and an action potential in the muscle. The action potential in the muscle cell leads to a contraction of the muscle and an extension of the limb. So, here we have a simple feedforward excitation circuit that mediates a behavior.
Now let’s examine the right branch of the axon of the sensory neuron of Figure 7. The action potential in the sensory neuron invades the synaptic terminal of the sensory neuron causing the release
Figure 7. See Neuroscience Online for animation.

Medical Neuroscience 2013 : Block 1
Page 13Introduction to Neuroscience
of transmitter, and subsequent excitation of the postsynaptic interneuron colored black. This neuron is called an interneuron because it is interposed between one neuron (here the SN) and another neuron (here the MN). The excitation of the interneuron leads to the initiation of an action and the subsequent release of transmitter from the presynaptic terminal of the interneuron (black triangle), but for this branch of the circuit, the transmitter leads to an IPSP in the postsynaptic flexor (F) motor neuron (colored red). The functional consequences of this feedforward inhibition it is to decrease the probability of the flexor motor neuron becoming active and producing an inappropriate flexion of the leg.
coNvergeNce aNd divergeNceThe simplified circuit mediating the stretch reflex is summarized in Figure 8. However, the proper function of the circuit of the stretch reflex also relies on convergence and divergence. A single sensory has multiple branches that diverge and make synaptic connections with many individual motor neurons. Therefore, when the muscle contracts as a result of the neurologist’s tapper, it does so because multiple muscle fibers are activated simultaneously by multiple motor neurons. Also, when the muscle is stretched, not one, but multiple sensory neuron are activated and these sensory neurons all project into the spinal cord where they converge on to individual extensor motor neurons. So, the stretch reflex is due to the combined effects of the activation of multiple sensory neurons and extensor motor neurons.
lateral iNhiBitioNEdge enhancement. Lateral inhibition is very important for processing sensory information. One example is a phenomenon in the visual system called edge enhancement. Figure 9 illustrates two bands, a dark gray band on the left, and a light gray band on the right. Although the dark band and the light band are of uniform luminance throughout each field, a close examination reveals that the light gray band appears somewhat lighter at the border of the dark gray band than it is in the other regions of the field. In contrast, the dark gray band appears somewhat darker at the border than at other regions of the dark field. This is a phenomenon of edge enhancement, which helps the visual system to extract important information from visual scenes. Edge enhancement is mediated, at least in part by lateral inhibition in the retina.
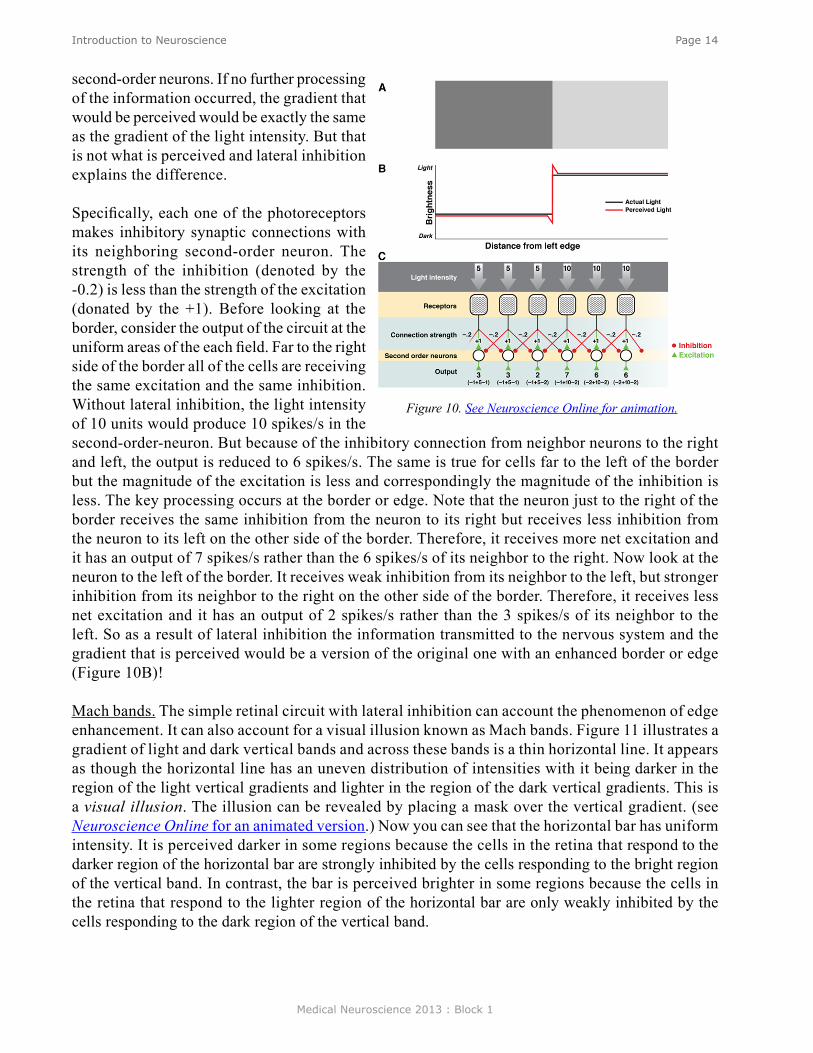
Figure 10 illustrates a simplified circuit with lateral inhibition. Light falls on the retina (Part A) and the intensity can be described by the step-like gradient (Part B). As a simplification, assume that the dark gray region has an intensity of five units and the light gray region has an intensity of ten units. The gradient of light activates the photoreceptors and the photoreceptors make synaptic connections to second-order neurons. Assume that the light intensity of 5 units leads to 5 spikes/s and the light intensity of 10 units leads to 10 spikes/s (Part C) in the photo receptors, and that the synaptic strength is sufficient (here indicated as +1) so that the light intensity of 5 units would lead to 5 spikes/s and the light intensity of 10 units would lead to 10 spikes/s respectively in the
Figure 8
Figure 9
Medical Neuroscience 2013 : Block 1
Page 14Introduction to Neuroscience
second-order neurons. If no further processing of the information occurred, the gradient that would be perceived would be exactly the same as the gradient of the light intensity. But that is not what is perceived and lateral inhibition explains the difference.
Specifically, each one of the photoreceptors makes inhibitory synaptic connections with its neighboring second-order neuron. The strength of the inhibition (denoted by the -0.2) is less than the strength of the excitation (donated by the +1). Before looking at the border, consider the output of the circuit at the uniform areas of the each field. Far to the right side of the border all of the cells are receiving the same excitation and the same inhibition. Without lateral inhibition, the light intensity of 10 units would produce 10 spikes/s in the second-order-neuron. But because of the inhibitory connection from neighbor neurons to the right and left, the output is reduced to 6 spikes/s. The same is true for cells far to the left of the border but the magnitude of the excitation is less and correspondingly the magnitude of the inhibition is less. The key processing occurs at the border or edge. Note that the neuron just to the right of the border receives the same inhibition from the neuron to its right but receives less inhibition from the neuron to its left on the other side of the border. Therefore, it receives more net excitation and it has an output of 7 spikes/s rather than the 6 spikes/s of its neighbor to the right. Now look at the neuron to the left of the border. It receives weak inhibition from its neighbor to the left, but stronger inhibition from its neighbor to the right on the other side of the border. Therefore, it receives less net excitation and it has an output of 2 spikes/s rather than the 3 spikes/s of its neighbor to the left. So as a result of lateral inhibition the information transmitted to the nervous system and the gradient that is perceived would be a version of the original one with an enhanced border or edge (Figure 10B)!
Mach bands. The simple retinal circuit with lateral inhibition can account the phenomenon of edge enhancement. It can also account for a visual illusion known as Mach bands. Figure 11 illustrates a gradient of light and dark vertical bands and across these bands is a thin horizontal line. It appears as though the horizontal line has an uneven distribution of intensities with it being darker in the region of the light vertical gradients and lighter in the region of the dark vertical gradients. This is a visual illusion. The illusion can be revealed by placing a mask over the vertical gradient. (see Neuroscience Online for an animated version.) Now you can see that the horizontal bar has uniform intensity. It is perceived darker in some regions because the cells in the retina that respond to the darker region of the horizontal bar are strongly inhibited by the cells responding to the bright region of the vertical band. In contrast, the bar is perceived brighter in some regions because the cells in the retina that respond to the lighter region of the horizontal bar are only weakly inhibited by the cells responding to the dark region of the vertical band.
Figure 10. See Neuroscience Online for animation.

Medical Neuroscience 2013 : Block 1
Page 15Introduction to Neuroscience
FeedBack/recurreNt iNhiBitioNFeedback inhibition in microcircuits. Feedback inhibition plays a general role in damping excitation through a neural circuit. A classic example is the Renshaw cell in the spinal cord. The axon of a spinal motor neuron branches. One branch innervates muscle as described earlier (e.g., Figure 7) and the other branch makes an excitatory synaptic connection with an interneuron called the Renshaw cell. The interneuron in turn inhibits the motor neuron, thereby closing the loop. Another example of feedback inhibition is found in the hippocampus. CA3-type pyramidal cells make excitatory connections to basket cells and the basket cells feedback to inhibit the CA3 cells. The term recurrent inhibition is applied to simple feedback inhibition circuits such as the Renshaw circuit in the spinal cord and the basket cell circuit in the hippocampus.
Feedback inhibition in nanocircuits. Feedback inhibition is not only prevalent in many neuronal circuits; it is also prevalent in biochemical circuits. Here it can serve as a substrate for generating oscillations. These can cover multiple time scales from seconds to days depending on the molecular components of the circuit.
1. Endogenous bursting behavior in neurons. The idealized neuron described earlier in the Chapter was silent in the absence of stimulation (e.g., Figure 3). However, some neurons fire action potentials in the absence of stimulation, and, in some cases, the firing patterns can exhibit a bursting pattern in which successive high frequency spike activity is followed by quiescent periods. Such neuronal properties could be important for generating rhythmic
Figure 11. See Neuroscience Online for animation.

Medical Neuroscience 2013 : Block 1
Page 16Introduction to Neuroscience
behaviors such as respiration. Figure 12 is an example of a recording from an invertebrate neuron that has an endogenous bursting rhythm. This particular neuron is called the parabolic burster because the nature of the inter-spike intervals being long in the beginning and end of the burst cycle, but very brief in the middle of the cycle. The cell fires a burst of action potentials and then becomes silent, but soon another burst occurs and this process continues indefinitely about every ten to fifteen seconds. The bursting occurs even if the neuron is surgically removed from the ganglion and placed it in culture so there are no synaptic connections from other neurons. So a neuronal network is not necessary for this rhythm - it is endogenous. But, it does involve a nanocircuit within the cell. Figure 13A illustrates a very simplified version of that network that emphasizes the key principle of operation. Critical for this network function is a channel in the membrane (labeled gSI), which is permeable to Ca2+. Because the concentration of Ca2+ is relatively high in the extracellular medium and low inside the cell, Ca2+ will move down its concentration gradient and in so doing will depolarize the cell. Eventually, the depolarization reaches threshold and the cell begins to fire. The firing leads to additional influx of Ca2+ (green arrow) and accumulation of Ca2+ within the cell. The key step is that the accumulation of Ca2+ inhibits (red arrow) the further influx of calcium and terminates the burst. The burst remains terminated as long as the levels of intracellular Ca2+ remain elevated. But the levels of Ca2+ do not remain elevated for long. They are reduced by intracellular buffers and removed from the cell by pumps (INaCa and ICaP). As intracellular levels of Ca2+ are reduced, the inhibition of the channel is removed (disinhibition) and the neuron begins to depolarize again and another burst is initiated. Essentially, what is seen here at the nanonetwork level is a recapitulation of a feedback inhibitory network (Figure 13B). An initial excitatory process leads to activation of an inhibitory process, which feeds back to shut off the excitatory process. In such a network, oscillations will result if the excitatory drive is continuous, but the inhibitory process wanes in its effectiveness.
2. Circadian rhythms. A second example of feedback inhibition is the nanocircuit for the gene regulation that underlies circadian rhythms. The vertebrate circadian rhythm is due to the operation of a group of neurons in a region of the brain called the suprachiasmatic nucleus, which is located just above the optic nerve. Those neurons have profound effects on both hormonal release such as melatonin, cortisol, as well as on autonomic functions such as body temperature (Figure 14). Despite the profound effects of this oscillator, its operation
Figure 12
Figure 13

Medical Neuroscience 2013 : Block 1
Page 17Introduction to Neuroscience
reduces to a very simple circuit, and indeed not a neural circuit, but rather another nanocircuit. The basic mechanism seems to be conserved across all animal species including man. Figure 15 is a simplified schematic diagram of the basic components. Several genes are involved but the core mechanism involves a gene called per, where per is for period. This gene was first identified from the fruit fly Drosophila but is also present in vertebrates. The per gene leads to the production of per messenger RNA. The per mRNA leaves the nucleus and enters the cytoplasm where it leads to the synthesis of PER protein. PER diffuses or is transported back into the nucleus where it represses the further transcription of the per gene. Conceptually, this system is very similar to that of the mechanism for the bursting neuron discussed above. The gene is activated, it produces the message and the protein, and the protein feedbacks to inhibit the gene expression. But how does the cycle repeat itself? The key mechanism is degradation of the PER. PER protein is degraded and it is degraded over a 24 hour period. So as the PER protein is degraded the inhibition or repression is removed (disinhibition) allowing this gene to start making messenger RNA and protein all over again. So once this cycle begins, it is repeated over and over again at a 24 hour period. This is the core mechanism underlying circadian rhythms and the powerful affects that they have on a number of different physiological systems. Basically, our circadian rhythms all start with a molecular feedback inhibition network.
Feedback inhibition in ring circuits. Recurrent inhibition can, at least in principle, explain the generation of complex motor patters, an example of which is quadrupedal locomotion. Quadrupedal location is interesting because quadrupeds are capable of not only moving their four legs, but generating different types of cycles of activity called gaits. Figure 16 illustrates four gaits (see Neuroscience Online for an animation). The first panel of Figure 16 is a walk. The sequence begins with extension of the left front limb. This is followed by extensions of the right hind limb, the right front limb and the left hind limb. In the trot (second panel), the left front and right hind limbs are in phase with each other and 180 degrees out of phase with the right front and left hind limbs. In the bound (third panel) the left front and right front limbs are in phase, but 180 degrees out of phase with the left hind and rear hind limbs. The gallop (fourth panel) is a variant of the bound in which there is a slight phase difference between the right and left front limbs and rear limbs.
Figure 14
Figure 15

Medical Neuroscience 2013 : Block 1
Page 18Introduction to Neuroscience
How does the nervous system generate these gaits? And, are separate neuronal circuits necessary for each one? Unfortunately, neuroscientists do not know the answers to these questions, but it is instructive to examine some possibilities. This is an approach in the field of neuroscience called Computational and Theoretical Neuroscience. One way to generate a gait is illustrated in Figure 17. Take four individual neurons each with endogenous bursting activity like the one illustrated previously in Figure 12, and assign activity in each one of these neurons to the control of a specific limb. The neurons could be “started” so that they have the appropriate phase relationships to generate a gait such as the bound illustrated in Figure 16. The difficulty would be in starting the neurons at exactly the precise time. Another problem would be slight “drifts” in the oscillatory periods of the four independent neurons that over time would cause the pattern to become uncoordinated. This dog is not going to win any races and it is probably not going to be able to walk.
So clearly the neurons need to be coupled. One way of doing this is to use a recurrent inhibition circuit consisting of four coupled neurons to form a so called “ring” circuit where each neuron in the circuit has endogenous bursting activity and each neuron is coupled to the next with an inhibitory synaptic connection (Figure 18A).
To obtain the correct phase relationships for gaits, rather than assigning Neuron 3 to be the right front, it is assigned to control the right hind limb, and Neuron 4 is assigned to control the right front limb (a simple twist of the circuit) (Figure 18B). When implemented with a computer simulation, this single circuit is capable of generating quadrupedal gates. Moreover, the same circuit, with just small changes in the properties of the individual neurons, can generate each of the four gaits illustrated in Figure 16 (see Neuroscience Online for an animation). This result indicates an important point about neural networks. In order to understand them it is necessary to understand not just the topology of the network, but also the nature of the connections between the neurons (whether they are excitatory or inhibitory), as well as the properties of the individual nodes (i.e., the neurons). Also, this simulation
Figure 16. See Neuroscience Online for animation.WALK TROT BOUND GALLOP
Figure 17
Figure 18

Medical Neuroscience 2013 : Block 1
Page 19Introduction to Neuroscience
illustrates a phenomenon called dynamic reconfiguration. It is not necessary to have four different networks to generate these four different gaits - it can all be done with a single circuit.
summary
Considerable progress has been made in understanding how different simple neural networks are involved in information processing and mediating behavior. Feedforward excitation and feedforward inhibition mediate reflex behaviors. Lateral inhibition is important for edge enhancement. Recurrent excitation is an important mechanism for memory. Recurrent inhibition can be important for generating locomotor behavior. Convergence and divergence are embedded in these microcircuits. The same kinds of network motifs are recapitulated in biochemical and gene networks.
The next level of understanding is at the level of the neuronal networks that mediate more complex, so called higher-order functions of the brain. Their understanding is becoming possible through the use of electrophysiological and optical recording techniques, and modern imaging techniques such as functional magnetic resonance imaging (fMRI) and diffusion tensor imaging (DTI). fMRI allows investigators to identify areas of the brain that are engaged in cognitive tasks, whereas DTI allows visualization of pathways linking one brain region to another. Object recognition is an example where progress is being made in understanding macrocircuits. Processing of visual information starts in the retina and then engages multiple cortical regions such as the occipital cortex and the temporal cortex. Within this macrocircuit are modules that extract higher-order information. Each module presumably involves hundreds, if not thousands of individual microcircuits. The challenge for the future is to determine how these modules work and how they interact with other modules. Although feedforward connections are present, feedback connections and lateral connections are widespread. The challenge is enormous but perhaps achievement of the goal will be facilitated by taking advantage of what has been learned about the principles of nanocircuits and microcircuits. To understand the macrocircuits it will be necessary to know more than the topology of the network interconnections. It will be necessary to know how each module functions and about the dynamics of the inter module connections.

Medical Neuroscience 2013 : Block 1
Page 20Synaptic Formation, Development & Regeneration
Synaptic Formation/Development and Regeneration
assignmenT
I. IntroductionII. Neuronal Migration
III. Axon PathfindingIV. Sources of Axon Guidance InformationV. Presynaptic and Postsynaptic Differentiation
VI. Synapse EliminationVII. Properties of NGF as a Model for Other Growth Factors
VIII. Injury Repair Recapitulates Developmental ProgramsIX. Summary
major objeCTives
1. Know the general principles underlying axonal guidance and connectional specificity between neurons and their other targets.
2. Know the concepts of presynaptic and postsynaptic differentiation, as well as synapse elimination.
3. Know the cellular events associated with neuronal degeneration and regeneration. 4. Know the properties of NGF as a model for trophic factors 5. Know the potential therapeutic application of tropic factors and tissue/cell transplantation
to degenerative disease.
required reading
● Bean, A.J., Synapse Formation, Synapse Formation/Development, Lecture Supplement Volume I.
addiTional reading
● Bean, A.J. Synapse Formation, Survival and Elimination, Neuroscience Online. http://nba.uth.tmc.edu/neuroscience/s1/chapter09.html
Lecturer: Andrew Bean, Ph.D.January 7, 2013 | 9:00 AM

Medical Neuroscience 2013 : Block 1
Page 21Synaptic Formation, Development & Regeneration
inTroduCTion
Perhaps the most remarkable feature of the nervous system is the accuracy of its synaptic connec-tions. The networks of circuits formed by neuronal interactions are responsible for the generation of behavior. Synapse formation is finely regulated. It involves processes at the cellular and subcellular levels that result in axons finding their appropriate targets from an array of choices, synapses being formed on the correct cellular compartment, and formation of pre- and postsynaptic specializations that allow for efficient information transfer.
Figure 1. B. In some areas of the brain neurons migrate by wrapping themselves around preexisting glial cells. Some neurons migrate to these final positions before they extend processes (e.g., motor neurons). Other neurons extend axons as they migrate (e.g., cerebellar granule neurons). (See Neuroscience Online for animation.)
Figure 1. A. Various modes of neuronal migration in the developing cerebral cortex. Early in development radial migration is prevalent and is mediated by somal translocation (a). As development proceeds, glia-guided movement predominates (b). Cortical interneurons follow tangential pathways (c) and also use ventricle-directed migration) (d). A subset of neurons switch from radial to tangential movement to allow movement across cortical modules (e).(From B. Nadarajah, P. Alifragis, R.O.L. Wong and J.G. Parnavelas, Neuronal Migration in the Developing Cerebral Cortex: Observations Based on Real-time Imaging. Cereb. Cortex (2003) 13 (6): 607-611.)
Medical Neuroscience 2013 : Block 1
Page 22Synaptic Formation, Development & Regeneration
axon PaThfinding
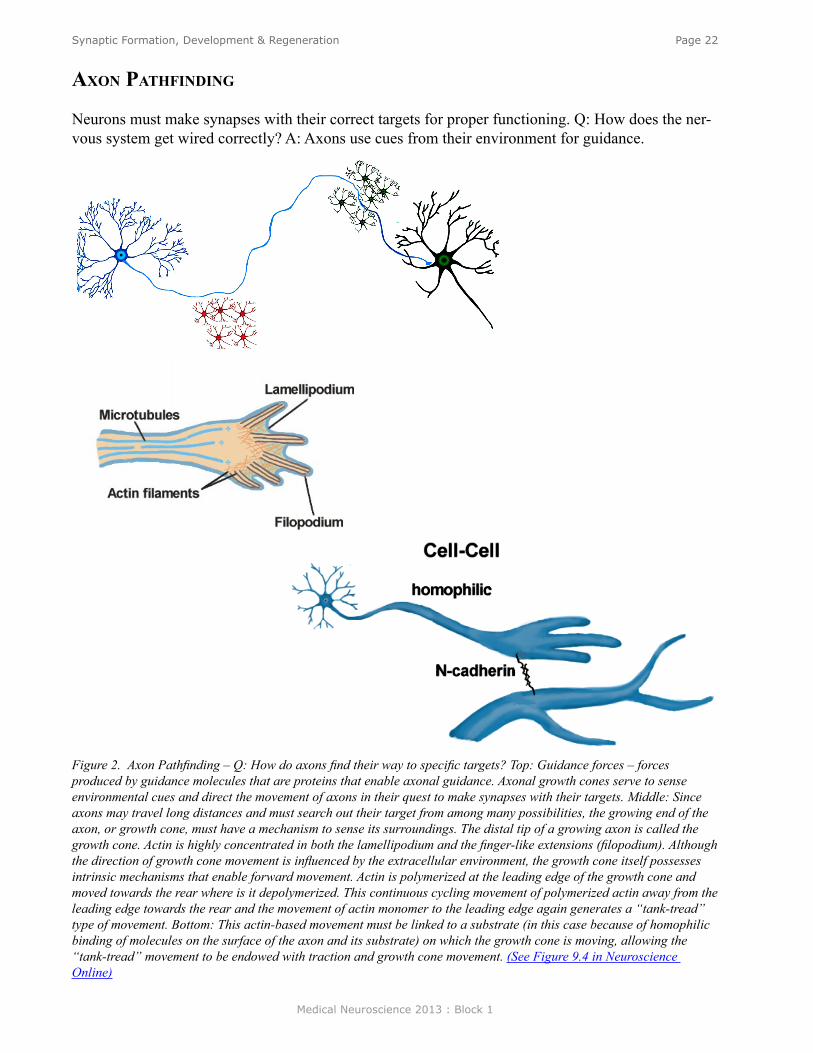
Neurons must make synapses with their correct targets for proper functioning. Q: How does the ner-vous system get wired correctly? A: Axons use cues from their environment for guidance.
Figure 2. Axon Pathfinding – Q: How do axons find their way to specific targets? Top: Guidance forces – forces produced by guidance molecules that are proteins that enable axonal guidance. Axonal growth cones serve to sense environmental cues and direct the movement of axons in their quest to make synapses with their targets. Middle: Since axons may travel long distances and must search out their target from among many possibilities, the growing end of the axon, or growth cone, must have a mechanism to sense its surroundings. The distal tip of a growing axon is called the growth cone. Actin is highly concentrated in both the lamellipodium and the finger-like extensions (filopodium). Although the direction of growth cone movement is influenced by the extracellular environment, the growth cone itself possesses intrinsic mechanisms that enable forward movement. Actin is polymerized at the leading edge of the growth cone and moved towards the rear where is it depolymerized. This continuous cycling movement of polymerized actin away from the leading edge towards the rear and the movement of actin monomer to the leading edge again generates a “tank-tread” type of movement. Bottom: This actin-based movement must be linked to a substrate (in this case because of homophilic binding of molecules on the surface of the axon and its substrate) on which the growth cone is moving, allowing the “tank-tread” movement to be endowed with traction and growth cone movement. (See Figure 9.4 in Neuroscience Online)
Medical Neuroscience 2013 : Block 1
Page 23Synaptic Formation, Development & Regeneration
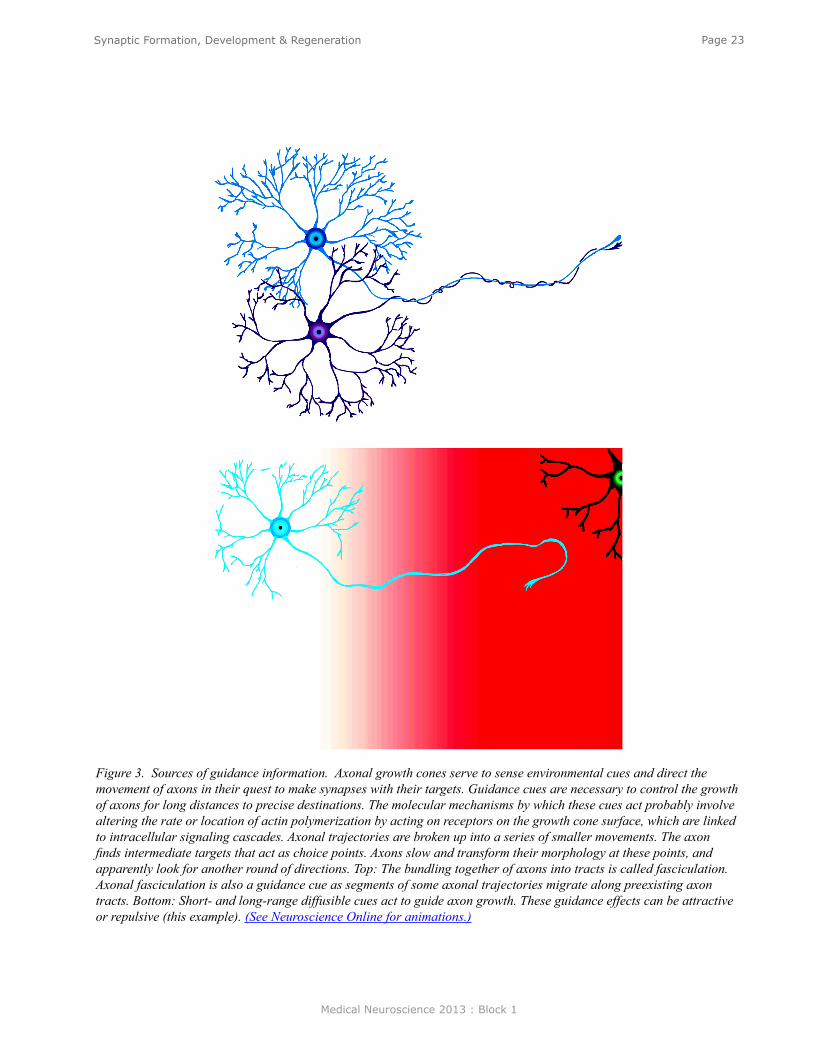
Figure 3. Sources of guidance information. Axonal growth cones serve to sense environmental cues and direct the movement of axons in their quest to make synapses with their targets. Guidance cues are necessary to control the growth of axons for long distances to precise destinations. The molecular mechanisms by which these cues act probably involve altering the rate or location of actin polymerization by acting on receptors on the growth cone surface, which are linked to intracellular signaling cascades. Axonal trajectories are broken up into a series of smaller movements. The axon finds intermediate targets that act as choice points. Axons slow and transform their morphology at these points, and apparently look for another round of directions. Top: The bundling together of axons into tracts is called fasciculation. Axonal fasciculation is also a guidance cue as segments of some axonal trajectories migrate along preexisting axon tracts. Bottom: Short- and long-range diffusible cues act to guide axon growth. These guidance effects can be attractive or repulsive (this example). (See Neuroscience Online for animations.)
Medical Neuroscience 2013 : Block 1
Page 24Synaptic Formation, Development & Regeneration
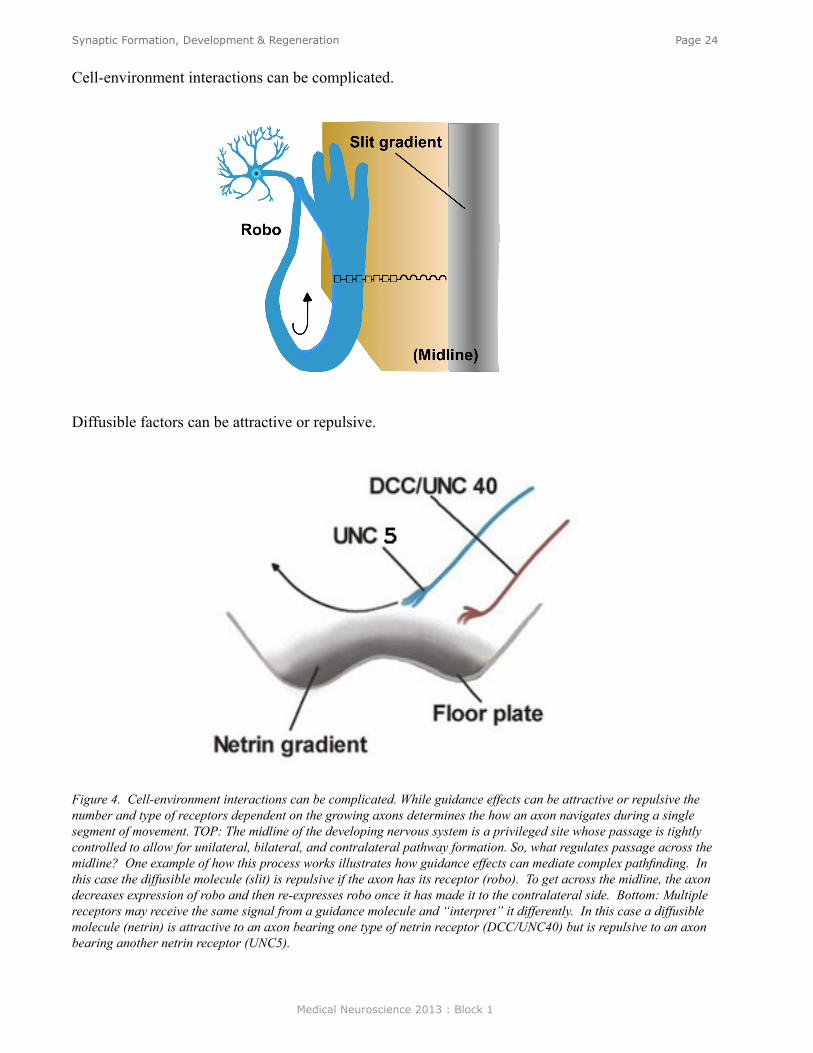
Cell-environment interactions can be complicated.
Diffusible factors can be attractive or repulsive.
Figure 4. Cell-environment interactions can be complicated. While guidance effects can be attractive or repulsive the number and type of receptors dependent on the growing axons determines the how an axon navigates during a single segment of movement. TOP: The midline of the developing nervous system is a privileged site whose passage is tightly controlled to allow for unilateral, bilateral, and contralateral pathway formation. So, what regulates passage across the midline? One example of how this process works illustrates how guidance effects can mediate complex pathfinding. In this case the diffusible molecule (slit) is repulsive if the axon has its receptor (robo). To get across the midline, the axon decreases expression of robo and then re-expresses robo once it has made it to the contralateral side. Bottom: Multiple receptors may receive the same signal from a guidance molecule and “interpret” it differently. In this case a diffusible molecule (netrin) is attractive to an axon bearing one type of netrin receptor (DCC/UNC40) but is repulsive to an axon bearing another netrin receptor (UNC5).
Medical Neuroscience 2013 : Block 1
Page 25Synaptic Formation, Development & Regeneration
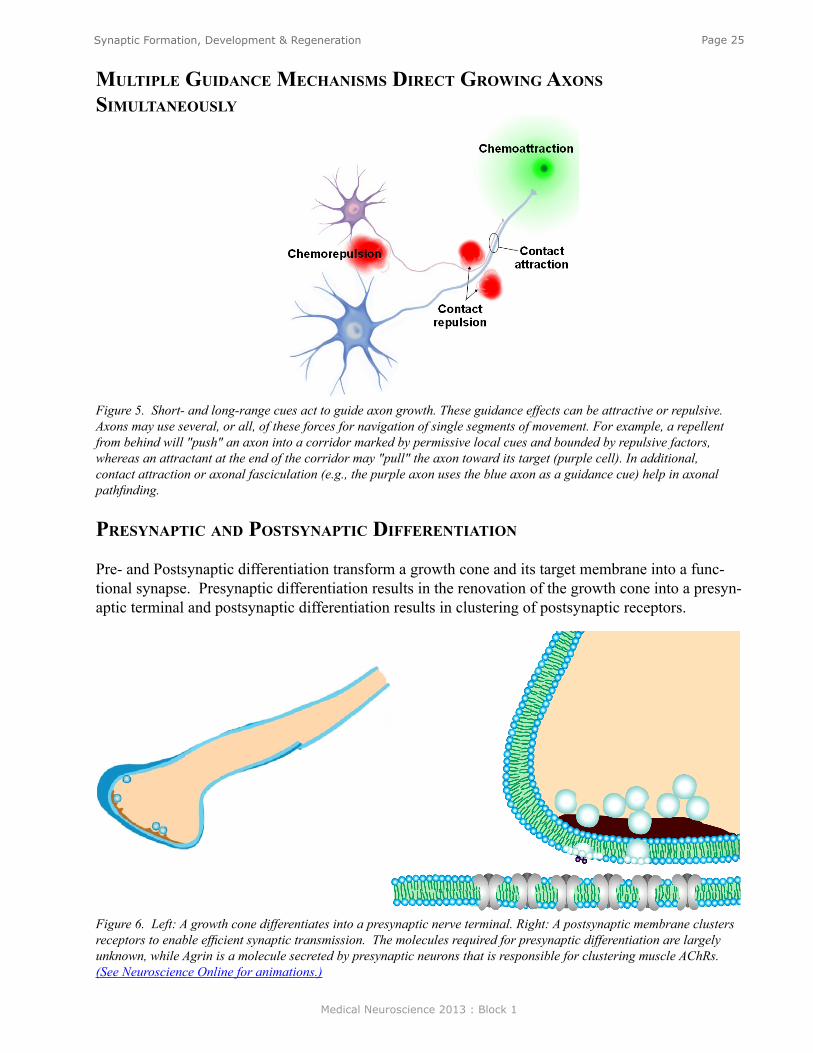
mulTiPle guidanCe meChanisms direCT growing axons simulTaneously
Figure 5. Short- and long-range cues act to guide axon growth. These guidance effects can be attractive or repulsive. Axons may use several, or all, of these forces for navigation of single segments of movement. For example, a repellent from behind will "push" an axon into a corridor marked by permissive local cues and bounded by repulsive factors, whereas an attractant at the end of the corridor may "pull" the axon toward its target (purple cell). In additional, contact attraction or axonal fasciculation (e.g., the purple axon uses the blue axon as a guidance cue) help in axonal pathfinding.
PresynaPTiC and PosTsynaPTiC differenTiaTion
Pre- and Postsynaptic differentiation transform a growth cone and its target membrane into a func-tional synapse. Presynaptic differentiation results in the renovation of the growth cone into a presyn-aptic terminal and postsynaptic differentiation results in clustering of postsynaptic receptors.
Figure 6. Left: A growth cone differentiates into a presynaptic nerve terminal. Right: A postsynaptic membrane clusters receptors to enable efficient synaptic transmission. The molecules required for presynaptic differentiation are largely unknown, while Agrin is a molecule secreted by presynaptic neurons that is responsible for clustering muscle AChRs. (See Neuroscience Online for animations.)
Medical Neuroscience 2013 : Block 1
Page 26Synaptic Formation, Development & Regeneration
synaPse eliminaTion
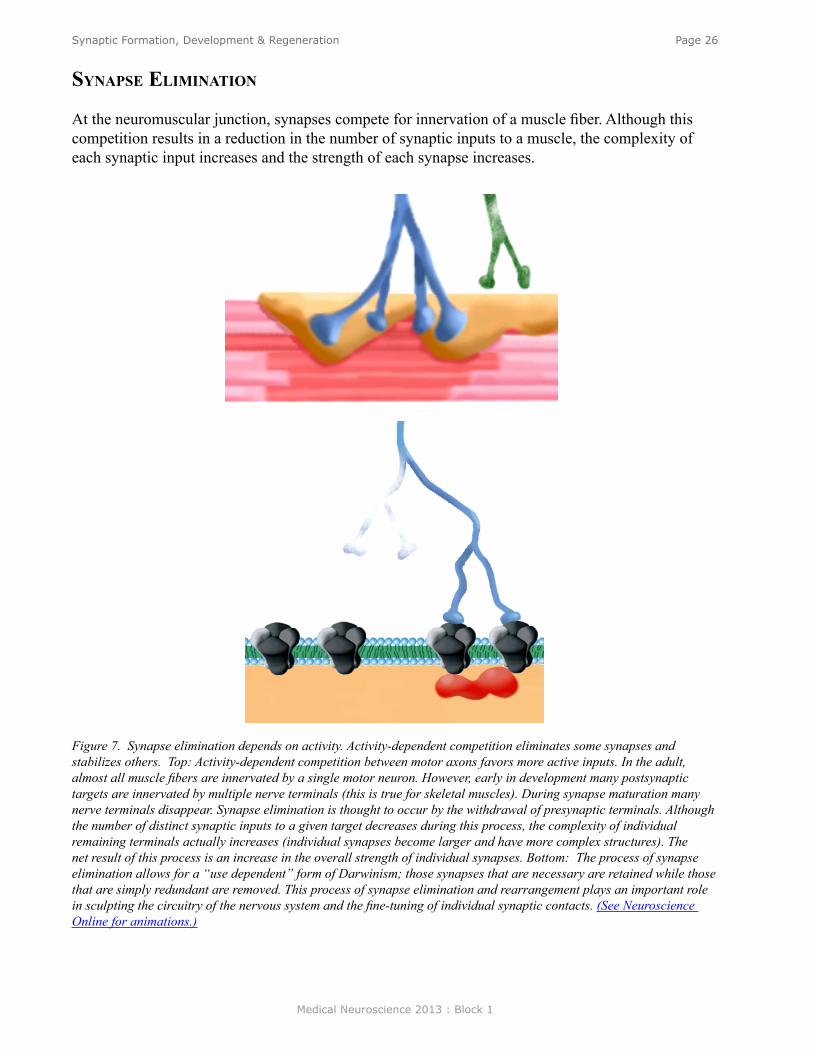
At the neuromuscular junction, synapses compete for innervation of a muscle fiber. Although this competition results in a reduction in the number of synaptic inputs to a muscle, the complexity of each synaptic input increases and the strength of each synapse increases.
Figure 7. Synapse elimination depends on activity. Activity-dependent competition eliminates some synapses and stabilizes others. Top: Activity-dependent competition between motor axons favors more active inputs. In the adult, almost all muscle fibers are innervated by a single motor neuron. However, early in development many postsynaptic targets are innervated by multiple nerve terminals (this is true for skeletal muscles). During synapse maturation many nerve terminals disappear. Synapse elimination is thought to occur by the withdrawal of presynaptic terminals. Although the number of distinct synaptic inputs to a given target decreases during this process, the complexity of individual remaining terminals actually increases (individual synapses become larger and have more complex structures). The net result of this process is an increase in the overall strength of individual synapses. Bottom: The process of synapse elimination allows for a “use dependent” form of Darwinism; those synapses that are necessary are retained while those that are simply redundant are removed. This process of synapse elimination and rearrangement plays an important role in sculpting the circuitry of the nervous system and the fine-tuning of individual synaptic contacts. (See Neuroscience Online for animations.)
Medical Neuroscience 2013 : Block 1
Page 27Synaptic Formation, Development & Regeneration
growTh faCTors
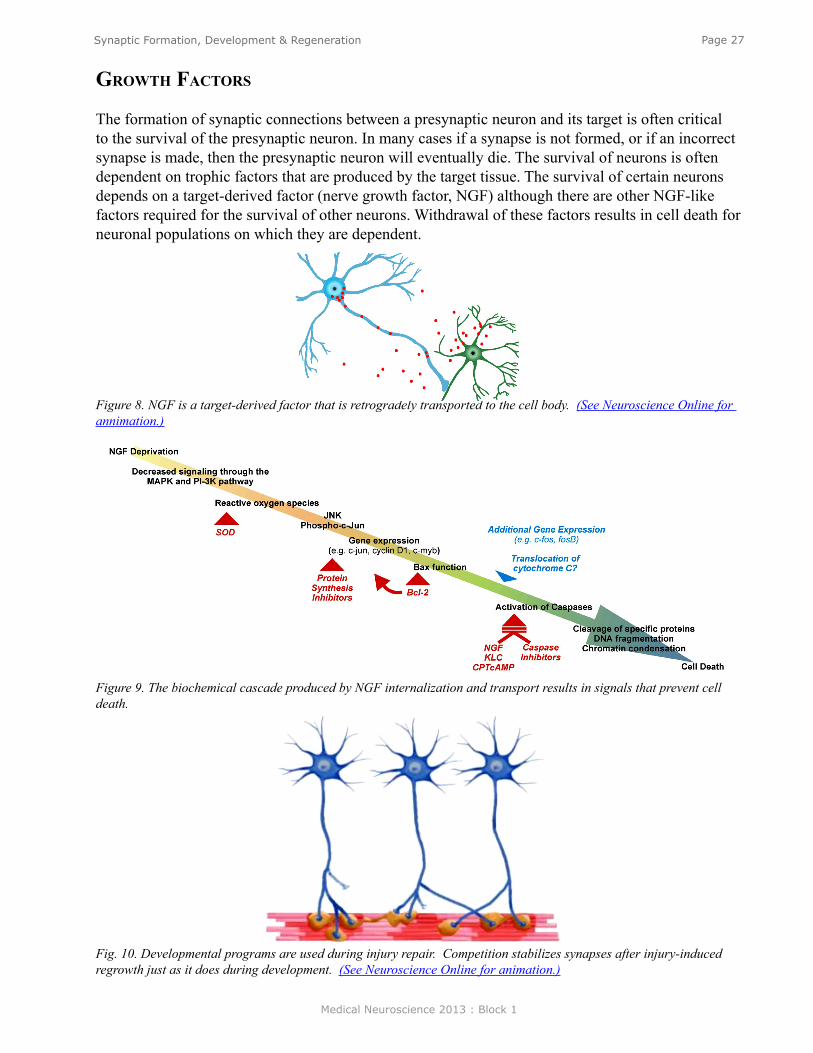
The formation of synaptic connections between a presynaptic neuron and its target is often critical to the survival of the presynaptic neuron. In many cases if a synapse is not formed, or if an incorrect synapse is made, then the presynaptic neuron will eventually die. The survival of neurons is often dependent on trophic factors that are produced by the target tissue. The survival of certain neurons depends on a target-derived factor (nerve growth factor, NGF) although there are other NGF-like factors required for the survival of other neurons. Withdrawal of these factors results in cell death for neuronal populations on which they are dependent.
Figure 8. NGF is a target-derived factor that is retrogradely transported to the cell body. (See Neuroscience Online for annimation.)
Figure 9. The biochemical cascade produced by NGF internalization and transport results in signals that prevent cell death.
Fig. 10. Developmental programs are used during injury repair. Competition stabilizes synapses after injury-induced regrowth just as it does during development. (See Neuroscience Online for animation.)
Medical Neuroscience 2013 : Block 1
Page 28Synaptic Formation, Development & Regeneration
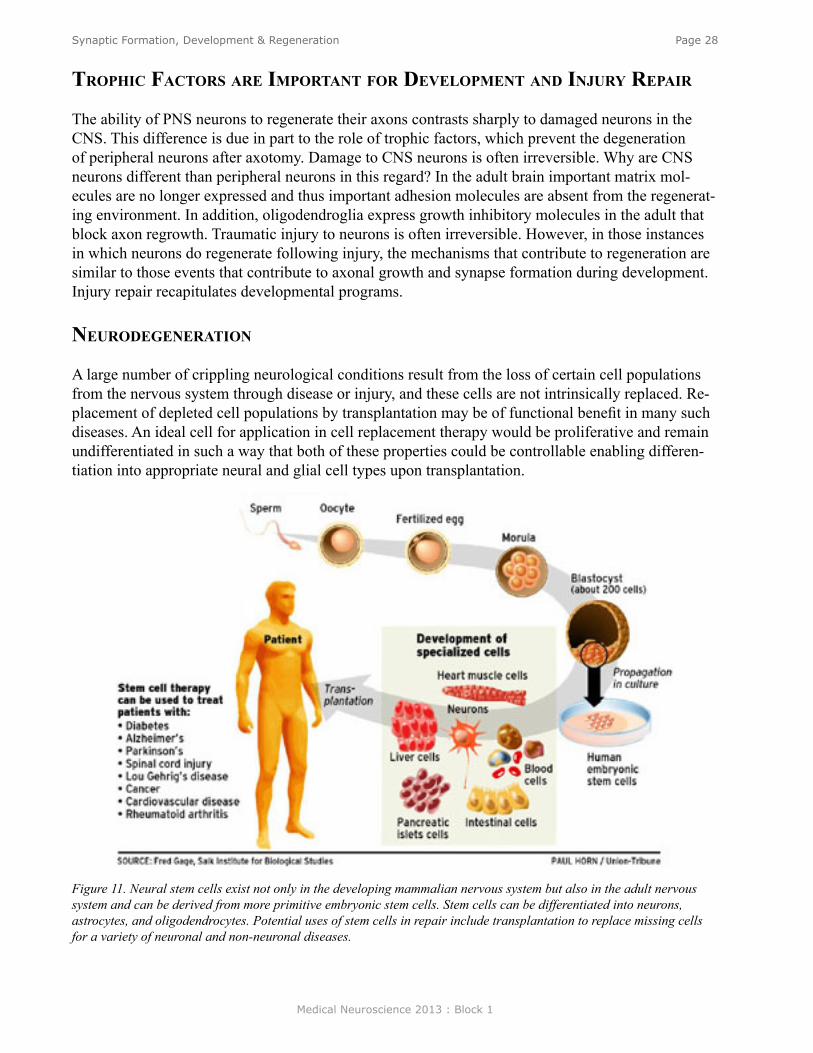
TroPhiC faCTors are imPorTanT for develoPmenT and injury rePair
The ability of PNS neurons to regenerate their axons contrasts sharply to damaged neurons in the CNS. This difference is due in part to the role of trophic factors, which prevent the degeneration of peripheral neurons after axotomy. Damage to CNS neurons is often irreversible. Why are CNS neurons different than peripheral neurons in this regard? In the adult brain important matrix mol-ecules are no longer expressed and thus important adhesion molecules are absent from the regenerat-ing environment. In addition, oligodendroglia express growth inhibitory molecules in the adult that block axon regrowth. Traumatic injury to neurons is often irreversible. However, in those instances in which neurons do regenerate following injury, the mechanisms that contribute to regeneration are similar to those events that contribute to axonal growth and synapse formation during development. Injury repair recapitulates developmental programs.
neurodegeneraTion
A large number of crippling neurological conditions result from the loss of certain cell populations from the nervous system through disease or injury, and these cells are not intrinsically replaced. Re-placement of depleted cell populations by transplantation may be of functional benefit in many such diseases. An ideal cell for application in cell replacement therapy would be proliferative and remain undifferentiated in such a way that both of these properties could be controllable enabling differen-tiation into appropriate neural and glial cell types upon transplantation.
Figure 11. Neural stem cells exist not only in the developing mammalian nervous system but also in the adult nervous system and can be derived from more primitive embryonic stem cells. Stem cells can be differentiated into neurons, astrocytes, and oligodendrocytes. Potential uses of stem cells in repair include transplantation to replace missing cells for a variety of neuronal and non-neuronal diseases.

Medical Neuroscience 2013 : Block 1
Page 29Synaptic Formation, Development & Regeneration
Test Your Knowledge(See Neuroscience Online for answers to questions)
1. Which of the following is a guidance cue for axonal outgrowth and pathfinding?A. ActinB. Stem cellsC. FasciculationD. FilopodiumE. Lamellopodium
2. Synapses...A. ... are stabilized by growth factor withdrawl.B. ... are stabilized by chemoattraction.C. ... are eliminated by oligodendrocytesD. ... are eliminated if they are not acive.E. ... are eliminated by agrin.

Medical Neuroscience 2013 : Block 1
Page 30Organization of Cell Types
Organization of Cell Types
assignmenT
I. Neurons A. Structure of a model neuron
1. Cell body2. Dendrite3. Axon4. Initial segment and axon hillock5. Nerve ending
B. Variations in neuronal structure C. Naming neuronal types D. Glossary of terms
II. Glial cells and functionA. Macroglia
1. Proloplasmic and fibrous astrocytes2. Oligodendroglia3. Schwann cells 4. Ependyma
B. Microglia
major objeCTives
1. Review the basic structure of neurons and glia.2. Understand the relationships between structure and function of CNS cells.
required reading
● Waymire, J.C. Organization of Cell Types, Lecture Supplement Volume I.
suggesTed reading (noT required)
● Waymire, J.C., Organization of Cell Types, Neuroscience Online http://nba.uth.tmc.edu/neuroscience/s1/chapter08.html
Lecturer: Jack C. Waymire, Ph.D.January 9, 2013 | 8:00 AM
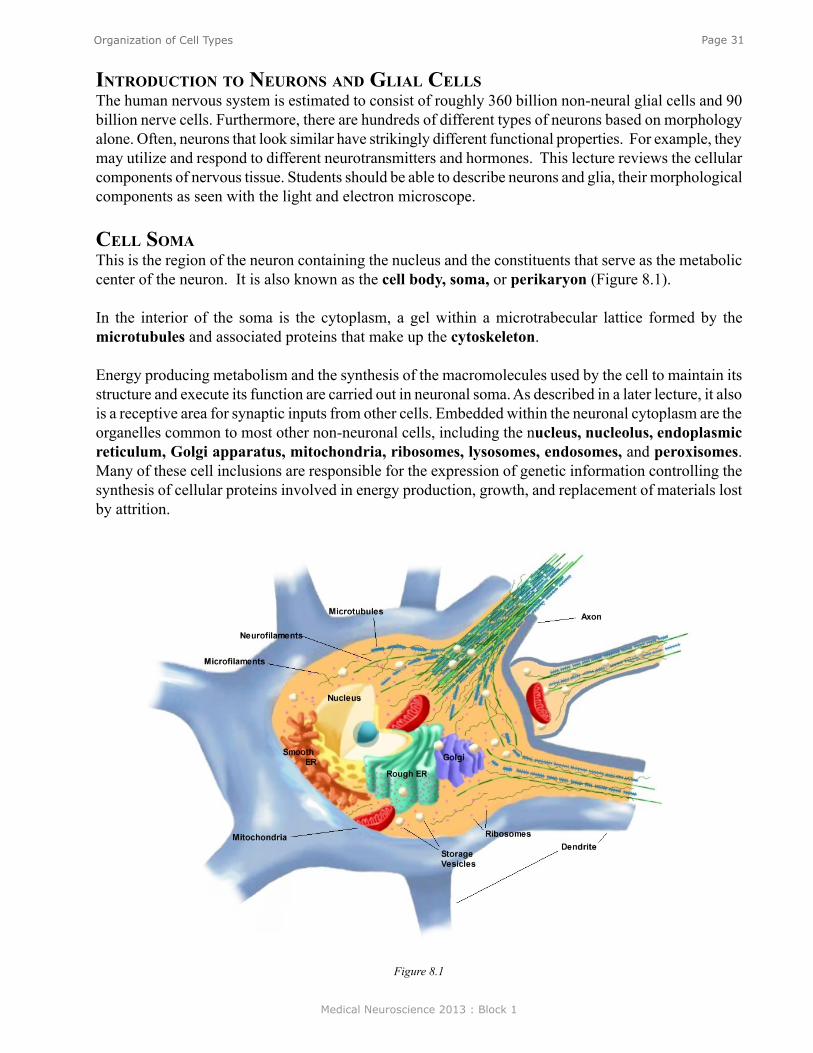
Medical Neuroscience 2013 : Block 1
Page 31Organization of Cell Types
inTroduCTion To neurons and glial CellsThe human nervous system is estimated to consist of roughly 360 billion non-neural glial cells and 90 billion nerve cells. Furthermore, there are hundreds of different types of neurons based on morphology alone. Often, neurons that look similar have strikingly different functional properties. For example, they may utilize and respond to different neurotransmitters and hormones. This lecture reviews the cellular components of nervous tissue. Students should be able to describe neurons and glia, their morphological components as seen with the light and electron microscope.
Cell somaThis is the region of the neuron containing the nucleus and the constituents that serve as the metabolic center of the neuron. It is also known as the cell body, soma, or perikaryon (Figure 8.1).
In the interior of the soma is the cytoplasm, a gel within a microtrabecular lattice formed by the microtubules and associated proteins that make up the cytoskeleton.
Energy producing metabolism and the synthesis of the macromolecules used by the cell to maintain its structure and execute its function are carried out in neuronal soma. As described in a later lecture, it also is a receptive area for synaptic inputs from other cells. Embedded within the neuronal cytoplasm are the organelles common to most other non-neuronal cells, including the nucleus, nucleolus, endoplasmic reticulum, Golgi apparatus, mitochondria, ribosomes, lysosomes, endosomes, and peroxisomes. Many of these cell inclusions are responsible for the expression of genetic information controlling the synthesis of cellular proteins involved in energy production, growth, and replacement of materials lost by attrition.
Figure 8.1

Medical Neuroscience 2013 : Block 1
Page 32Organization of Cell Types
dendriTesThe membrane of the neuron functions as a receptive surface over its entire extent; however, specific inputs (termed afferents) from other cells are received primarily on the surface of the cell body and on the surface of the specialized processes known as dendrites. The dendritic processes may branch extensively and are covered with projections known as dendritic spines. These spines provide a tremendous increase in the surface area available for synaptic contacts. The dendritic processes and spines of neurons are essentially expansions of cytoplasm containing most of the organelles found in the cell body. Dendrites contain numerous orderly arrays of microtubules and fewer neurofilaments (see next page). The microtubule associated proteins (MAPs) found in dendrites are distinct in that they have a molecular weight that is higher than those found in the axon. An example is MAP2. Microtubules in dendrites have their positive ends pointed toward the cell soma. The Mitochondria in dendrites are often arranged longitudinally. Rough endoplasmic reticulum and ribosomes are present in larger but not smaller dendrites. The shape and extent of the "dendritic tree" of an individual neuron is indicative of the quantity of information received and processed by that neuron. The dendritic spines usually contain microfilaments. As you learned from Dr. Byrne’s lectures, these structures are the cytoskeletal element responsible for changes in spine shape observed in some examples of synaptic plasticity.
Information is received by the dendrite through receptors on dendrite surface that react to transmitters released from the axon terminals of other neurons. Dendrites may consist of a single twig-like extension from the soma or a multi-branched network capable of receiving inputs from thousands of other cells. For instance, an average spinal motor neuron with a moderate-sized dendritic tree, receives 10,000 contacts, with 2,000 of these on the soma and 8,000 on the dendrites.
iniTial segmenT and axon hilloCk The cone-shaped region of the cell body where the axon originates is termed the axon hillock (Figure 8.2). This area is free of ribosomes and most other cell organelles, with the exception of cytoskeletal elements and organelles that are being transported down the axon. The neurofilaments in the axon hillock become clustered together as fascicles. The region between the axon hillock and the beginning of
Figure 8.2 Diagrammatic representation of the initial segment of a neuron
emphasizing the areas at which the action potential is initiated. (See Neuroscience
Online for enlarged view)

Medical Neuroscience 2013 : Block 1
Page 33Organization of Cell Types
the myelin sheath is known as the initial segment. In many cases, this region is the anatomical location for the initiation of the action potential. The area under the axolemma in this region has material that stains darkly when viewed by EM. This region is shown in Figure 8.3. At the distal-most end of the axon and its collaterales are small branches whose tips are button-shaped cytoplasmic enlargements called terminal boutons or nerve endings.
axon The other type of process in the idealized neuron is the axon. Each neuron has only one axon and it is usually straighter and smoother than the dendritic profiles. Axons also contain bundles of microtubules and neurofilaments and scattered mitochondria. The most MAPs in an axon have a lower molecular weight than those in the dendrite. A predominant MAP in axons is tau. Microfilaments within the axon are usually associated with an area adjacent to the plasmalemma and often are the most dense at the nodes of Ranvier. Beyond the initial segments, the axoplasm lacks rough endoplasmic reticulum and free ribosomes. The branches of axons are known as axon collaterales. The axon itself is often surrounded by a membranous material, called the myelin sheath, formed by glia cells. The myelin sheath acts to insulate the plasmalemma of the axon in a way that necessitates the more rapid spread of the depolarization of the plasmalemma and increases the speed of conduction of the nerve impulse.
nerve endings The part of the plasma membrane of the nerve ending that is specialized to form functional a contact with other cells is the synapse. When neurons interact with muscle fibers, the region of functional contact is called the neuromuscular junction or motor endplate. According to the classical definition of synapse, when a nerve ending synapses on a dendrite or soma of a second neuron it is termed either an axodendritic or an axosomatic synapse, respectively. However, almost all possible combinations of pre- and postsynaptic elements have been found in the central nervous system. These different types of synapse are designated by combining the name of the structure of the presynaptic element with that of the postsynaptic structure. For example, when the transfer of information occurs from an axon to axon or from one terminal to another, the synapse involved is called an axoaxonic synapse.
Cellular elemenTs aT The TyPiCal nerve Terminal Regions of functional contacts between neurons (synapses) have distinct morphological characteristics. Although a great deal of variation exists in the size and shape of boutons of individual neurons, synapses can be identified by the presence of the following:
Figure 8.3. Excitatory nerve ending and post synaptic cell

Medical Neuroscience 2013 : Block 1
Page 34Organization of Cell Types
1. A presynaptic complement of membrane-bound synaptic vesicles exists. These are spherical vesicles in excitatory nerve endings, shown in Figure 8.3. In inhibitory neurons the synaptic vesicle are often flattened.
2. The nerve ending often has aggregations of dense material in the cytoplasm immediately adjacent to the membrane on the pre- and postsynaptic side of the junction (these are known as presynaptic density or postsynaptic density, respectively.) This dense material on the presynaptic side is thought to be the site of vesicle attachment. The dense material on the postsynaptic side is a site where receptor proteins and channels are prevalent.
3. Many mitochondria are present, especially in the nerve terminal; and 4. There is a distinct synaptic cleft or intercellular space of approximately 20-40 nm.5. An endoplasmic reticulum is present that regulates Ca2+ level. 6. Endosomal membrane that functions in the recycling of synaptic vesicles.
sTruCTural variaTionsNumerous variations of the “model” neuron described above exist. An important modification, which occurs especially in receptor neurons, involves the designation of a neuronal process as a dendrite or as an axon. Classically, the axon has been identified as the myelinated or unmyelinated process that transmits signals away from the cell body. The classical view of the dendrite is that of an unmyelinated tube of cytoplasm which carries information toward the cell body. However, this distinction does not hold for ALL neurons. Some cells have a myelinated process that transmits signals toward the cell body. Morphologically the “dendrite” and the “axon” may, therefore, be indistinguishable. Neither the position of the cell body nor the presence or absence of myelin is always a useful criterion for understanding the orientation of the neuron. The region of impulse initiation is more reliable guide to understanding the functional focal point of the cell. This region is analogous to the initial segment of the model neuron, discussed above. Routinely the fiber or process, which contains the initial segment or trigger zone, is referred to as an axon. Note, as shown in Figure 8.4, the trigger zone does not have to be immediately adjacent to the cell body.
Figure 8.4 Comparison of variations in the structure of neurons (See Neuroscience Online for animation)

Medical Neuroscience 2013 : Block 1
Page 35Organization of Cell Types
naming neuronsA number of conventions have evolved to classify and name neurons as described below:
Complexity of dendritic tree: One of the oldest conventiopns devised by Golgi in the late 1800’s is based on the complexity of the dendritic tree of the neuron. Through this approach cells are classified as unipolar, bipolar and multipolar, as shown in Figure 8.4.
Unipolar cells have only one cell process, and are primarily found during development in vertebrates and as well as in many invertebrate organisms.
Bipolar cells are present in the retina and the olfactory bulb.
Multipolar cells make up the remainder of neuronal types and are, consequently, the most numerous type. These have been further sub-categorized into Golgi type I cells that are large multipolar neurons, usually interneurons, and Golgi type II cells that are small multipolar neurons.
Shape: Cells are also named for their shape (e.g., pyramidal cells, granular cells, stellate cells).
Historical reference: Several cells are named for the person who first described them (e.g., Purkinje cells, Golgi cells).
Neurotransmitter: More recently, neurons have been named for the neurotransmitter they contain (e.g., CNS dopamine, norepinephrine and serotonin cell groups as covered in Chapter 13 of Neuroscience Online). This description is possible because of the development of histochemical and immunocytochemical methods to specifically identify the neurotransmitter released by specific neurons.
glial Cells and funCTionThe most numerous cellular constituents of the central nervous system are the non-neuronal, neuroglial ("nerve glue") cells that occupy the space between neurons. It is estimated that there are roughly 360 billion glial cells, which comprise 80-90% of the cells in the CNS. This section covers the general classifications of the neuroglial cells and describes some of the general properties that distinguish neuroglia from neurons.
Neuroglia differ from neurons in several general ways including neuroglia:
1. do not form synapses,2. have essentially only one type of process,3. retain the ability to divide, and4. are less electrically excitable than neurons.
Classification: Neuroglia are broadly classified based on cell size into 1) macroglia and 2) microglia. In addition neuropathologists use alkaline (basic) dyes to show nuclear morphology and from this morphology delineate microglia from macroglia. Further, nuclear size, shape, staining intensity, and distribution of chromatin, are used to evaluate the relative
Figure 8.5. Types of neuroglia. (See Neuroscience Online for more detailed information)

Medical Neuroscience 2013 : Block 1
Page 36Organization of Cell Types
number of cell types in pathological material. In research metal stains are used show the shape of the cell and cytoplasmic architecture as well the relative abundance of cytoskeleton.
maCroglia There are three types of macroglia: Oligodendroglia, Ependyma, and Astrocytes. This section discusses the two types of astrocytes: Protoplasmic and Fibrous.
ProToPlasmiC and fibrous asTroCyTesProtoplasmic astrocytes are found primarily in gray matter. With silver or glial specific stains, their cell bodies and processes are very irregular. The processes range from large to very fine, sometimes forming sheets that run between axons and dendrites, and surround synapses. These fine sheet-like processes give the protoplasmic astrocyte a "fuzzy" or murky appearance. Using metal stains bundles of fine fibrils can be seen within the cytoplasm. The nucleus of a protoplasmic astrocyte is ellipsoid or bean-shaped with characteristic flecks of chromatin. Intercellular junctions have been noted between processes of protoplasmic astrocytes that are believed to mediate ion exchange between cells.
Fibrous astrocytes are found primarily in white matter. With glial specific metal stains these cells have a smoother cell body contour than protoplasmic astrocytes and have processes that emerge radially from the cell body. The processes are narrower and branch often to form end feet on blood vessels, ependyma, and pia. The processes of fibrous astrocytes do not form sheets and do not tend to conform to the shape of the surrounding neurons or vascular elements. The major distinguishing feature of fibrous astrocytes, as the name suggests, is an abundance of glial fibrils arranged in parallel arrays in the cytoplasm and extending into the astrocytic processes as seen with heavy metal stains.
In Nissl stains, the fibrous astrocytes also have a nucleus which is ellipsoid or bean-shaped with characteristic flecks of chromatin essentially the same as that of the protoplasmic type. Intercellular adherences have also been observed between fibrous astrocytes.
Both types of astrocytes function to support the neurons by 1) providing a physical barrier between cells, 2) maintaining the ionic and pH equilibrium of the extracellular space around neurons, and 3) continually modifying the chemical environment of the neighboring cells. 4) astrocytes form a complete lining around the external surface of the CNS. This is termed the glial limitans. They also 5) form a barrier around blood vessels. These are termed perivascular feet. As discussed by Dr. Bean, during development, 6) astrocytes form scaffolding along which nerve cells migrate to achieve their mature location and structure. Importantly, during injury astrocytes 7) proliferate and phagocytize dead cells. This can lead to the formation of glial scar.
In addition to these general functions, astrocytes also act in many specialized ways to facilitate neuronal function. For example, as will be discussed in subsequent lectures, astrocytes 8) metabolize neurotransmitters by removing them from the synaptic cleft. For example, the amino acid glutamate is taken up by astrocytes and inactivated by conversion to glutamine. Glutamine is then transported to the neuron to be re-synthesized into glutamate (see Chapter 13). More recent evidence indicates that the astrocytes can dramatically change size as part of their physiological regulation of the neuronal environment. Many of these functions will be discussed in later sections.

Medical Neuroscience 2013 : Block 1
Page 37Organization of Cell Types
oligodendrogliaOligodendrocyte, although located in both gray and white matter, are the predominant cell type in white matter where they are often located as rows of cells between groups of neuronal processes. These are termed interfascicular oligodendroglia and are involved in the formation and maintenance of the myelin of nearby axonal processes. In gray matter, oligodendroglia are often located near neurons and, therefore, are known as perineuronal satellite cells. Cell bodies of oligodendroglia are located near capillaries, but they lack the definite perivascular end feet characteristic of astrocytes.
Anatomically, oligodendrocytes have fewer and more delicate processes than astrocytes. They can be distinguished from macroglia by a smaller eccentrically located nucleus that contains clumps of chromatin. Silver staining of the cytoplasm of oligodendrocytes tends to be much darker than that of astrocytes, and does not contain glial fibrils.
The role of oligodendroglia in the central nervous system, particularly of the interfascicular oligodendrocytes, is the formation and maintenance of myelin. Myelin is composed of concentric layers of membranes compacted against one another with an internal (i.e. against the nerve fiber) and an external collar of cytoplasm. A single oligodendrocyte contributes to the myelination of several adjacent nerve processes. Moreover, more than one oligodendrocyte contributes to the myelination of a single internode of an axon. The lamellae of myelin membranes result from the spiral wrapping of the axon by cytoplasmic processes of the oligodendroglia. Interestingly, the oligodendrocyte forming a particular myelin internode (i.e. the myelin between two nodes) is seldom seen directly adjacent to the myelin-wrapped process. This is because thin cytoplasmic bridges connect the region of the oligodendrocyte cell body to the external wrap of myelin. It is important to note that the regions of the axon exposed at the nodes of Ranvier are not bare. They are sites of branching of the axon, the sites of synaptic contacts, or are covered with various glial processes.
In the peripheral nervous system (PNS), Schwann cells, rather than oligodendroglia form the myelin. In contrast to oligodendroglia, Schwann cells migrate around the axon, laying a membrane covering around the axon by squeezing out their own cytoplasm. Also, a single Schwann cell form the myelin for a single intermodal region. In addition unmyelinated axons in the PNS are also enclosed by membranes formed by Schwann cells.
ePendymaEpendymal cells are derived from the early germinal epithelium lining the lumen of the neural tube and thus are also ectodermal derivatives (along with neurons, astrocytes, and oligodendrocytes). Ependymal cells from the lining of the ventricles of the brain and the central canal of the spinal cord. They are arranged in a single-layered columnar epithelium, and have many of the histological characteristics of simple epithelium, which vary from squamous to cuboidal depending upon their location. The ependyma that form the ventricular lining do not connect to a basal lamina, but rest directly upon underlying nervous tissue. As shown in Figure 8.6, the surface facing the ventricle contains many microvilli and cilia. These cilia move cerebrospinal fluid (CSF) in the
Figure 8.6 Diagrammatic representation of the arrangement of ependymal cells to form the ciliated lining of the ventricles. (See Neuroscience Online to
view ependymal layer)

Medical Neuroscience 2013 : Block 1
Page 38Organization of Cell Types
ventricles. The lateral borders of the ependymal cells are relatively straight and form junctions with adjacent cells.
Ependymal cells are modified in various regions of the ventricles into layers of cuboidal epithelium, which do lie on a basement membrane (formed by an outgrowth of the pia) over a rich bed of vasculature and connective tissue. This is the choroid plexus studied in the Laboratory that is responsible for the secretion, uptake and transport of substances to and from the CSF.
miCrogliaMicroglia, in contrast to the other types of glial cells, originate from embryonic mesoderm. Although they are present throughout the central nervous system, they are inconspicuous in mature normal tissue and are difficult to identify with the light or electron microscope. Careful studies have shown that they are more abundant in gray matter, and may compromise up to 5-10% of the neuroglia in the cerebral cortex.
The general appearance of microglia is similar to oligodendrocytes, although they are smaller and have undulating processes with spine-like projections. Microglial nuclei are elongated or triangular and stain deeply with alkaline dyes.
Following damage to nervous tissue, microglia proliferate and migrate to the site of injury where they clear cellular debris by phagocytosis. The reacting microglia have a swollen form with shortened processes and are difficult to discriminate from phagocytes from the periphery or migrating perivascular cells. It is estimated that at least one third of the phagocytes appearing in the area of a CNS lesion are of CNS origin.
Test Your Knowledge
(See Neuroscience Online for answer to the question.)
8. Which of the following cell types proliferate in the CNS in response to injury? (Note: There is more than one correct answer.)
A. Neurons B. MicrogliaC. Fibrous astrocytesD. Protoplasmic astrocytesE. Macrophages
8. Which of the following cell types is responsible for the maintenance of pH of the extracellular space of the CNS? (Note: There is more than one correct answer.)
A. MicrogliaB. Fibrous AstrocytesC. Protoplasmic AstrocytesD. Ependymal cellsE. Macrophages

Medical Neuroscience 2013 : Block 1
Page 39Resting Potentials and Action Potentials
Resting Potentials and Action Potentials
assignmenT
I. Basic features of the action potential in neuronsII. Intracellular recording of the resting potential and action potential
III. Ionic mechanisms of the resting potentialA. The equilibrium potential and the Nernst equationB. Bernstein’s hypothesis for the ionic basis of the resting potentialC. Goldman-Hodgkin-Katz model of the resting potential
IV. Role of sodium ions in the generation of the action potential: The Hodgkin and Katz experiment
major objeCTives
1. Understand the ways in which electrical potentials can be measured from nerve cells.2. Understand the concept of an equilibrium potential.3. Know the Nernst equation and Goldman-Hodgkin-Katz equation.4. Understand why the resting potential is not equal to the potassium equilibrium potential.5. Know the different phases of the nerve action potential.
required reading
● Byrne, J.H. Resting Potentials and Action Potentials/Ionic Mechanisms of Action Potentials; Neuroscience Online: http://nba.uth.tmc.edu/neuroscience/s1/chapter01.html; or
● Byrne, J.H. Resting Potentials and Action Potentials, Lecture Supplement Volume I.
suggesTed reading
● Byrne, J. H., Roberts, J. L., From Molecules to Networks: An Introduction to Cellular and Molecular Neuroscience, 2nd Edition, Elsevier, Amsterdam, 2009, Chapter 5.
Lecturer: John H. Byrne, Ph.D.January 9, 2013 | 9:00 AM

Medical Neuroscience 2013 : Block 1
Page 40Resting Potentials and Action Potentials
Despite the enormous complexity of the brain, it is possible to obtain an understanding of its function by paying attention to two major details:
• First, the ways in which individual neurons, the components of the nervous system, are wired together to generate behavior.
• Second, the biophysical, biochemical, and electrophysiological properties of the individual neurons.
A good place to begin is with the components of the nervous system and how the electrical properties of the neurons endow nerve cells with the ability to process and transmit information.
Important insights into the nature of electrical signals used by nerve cells were obtained more than 50 years ago. Electrodes were placed on the surface of an optic nerve of an invertebrate eye. (By placing electrodes on the surface of a nerve, it is possible to obtain an indication of the changes in membrane potential that are occurring between the outside and inside of the nerve cell.) Then 1-sec duration flashes of light of varied intensities were presented to the eye; first dim light, then brighter lights. Very dim lights produced no changes in the activity, but brighter lights produced small repetitive spike-like events. These spike-like events are called action potentials, nerve impulses, or sometimes simply spikes. Action potentials are the basic events the nerve cells use to transmit information from one place to another.
feaTures of aCTion PoTenTials
The recordings in the figure above illustrate three very important features of nerve action potentials. First, the nerve action potential has a short duration (about 1 msec). Second, nerve action potentials are elicited in an all-or-nothing fashion. Third, nerve cells code the intensity of information by the frequency of action potentials. When the intensity of the stimulus is increased, the size of the action potential does not become larger. Rather, the frequency or the number of action potentials increases. In general, the greater the intensity of a stimulus, (whether it be a light stimulus to a photoreceptor, a mechanical stimulus to the skin, or a stretch to a muscle receptor) the greater the number of action potentials elicited. Similarly, for the motor system, the greater the number of action potentials in a motor neuron, the greater the intensity of the contraction of a muscle that is innervated by that motor neuron.
Figure 1.1 See Neuroscience Online for animation.

Medical Neuroscience 2013 : Block 1
Page 41Resting Potentials and Action Potentials
Action potentials are of great importance to the functioning of the brain since they propagate information in the nervous system to the central nervous system and propagate commands initiated in the central nervous system to the periphery. Consequently, it is necessary to understand thoroughly their properties. To answer the questions of how action potentials are initiated and propagated, we need to record the potential between the inside and outside of nerve cells using intracellular recording techniques.
inTraCellular reCordings from neurons
The potential difference across a nerve cell membrane can be measured with a microelectrode whose tip is so small (about a micron) that it can penetrate the cell without producing any damage. When the electrode is in the bath (the extracellular medium) there is no potential recorded because the bath is isopotential. If the microelectrode is carefully inserted into the cell, there is a sharp change in potential. The reading of the voltmeter instantaneously changes from 0 mV, to reading a potential difference of -60 mV inside the cell with respect to the outside. The potential that is recorded when a living cell is impaled with a microelectrode is called the resting potential, and varies from cell to cell. Here it is shown to be -60 mV, but can range between -80 mV and -40 mV, depending on the particular type of nerve cell. In the absence of any stimulation, the resting potential is generally constant.
Figure 1.2. See Neuroscience Online for animation.
It is also possible to record and study the action potential. Figure 1.3 illustrates an example in which a neuron has already been impaled with one microelectrode (the recording electrode), which is connected to a voltmeter. The electrode records a resting potential of -60 mV. The cell has also been impaled with a second electrode called the stimulating electrode. This electrode is connected to a battery and a device that can monitor the amount of current (I) that flows through the electrode. Changes in membrane potential are produced by closing the switch and by systematically changing both the size and polarity of the battery. If the negative pole of the battery is connected to the inside of the cell as in Figure 1.3A, an instantaneous change in the amount of current will flow through the stimulating electrode, and the membrane potential becomes transiently more negative. This result should not be surprising. The negative pole of the battery makes the inside of the cell more negative than it was before. A change in potential that increases the polarized state of a membrane is called a hyperpolarization. The cell is more polarized than it was normally. Use yet a larger battery and the potential becomes even larger. The resultant hyperpolarizations are graded functions of the magnitude of the stimuli used to produce them.
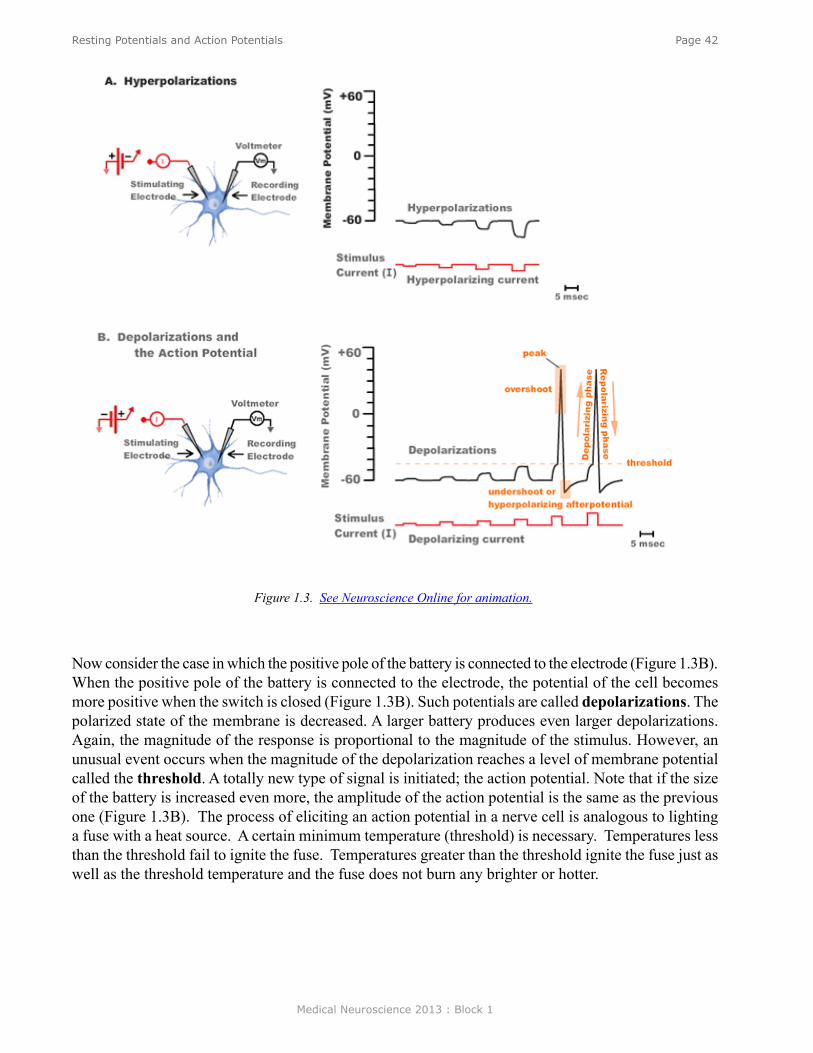
Medical Neuroscience 2013 : Block 1
Page 42Resting Potentials and Action Potentials
Now consider the case in which the positive pole of the battery is connected to the electrode (Figure 1.3B). When the positive pole of the battery is connected to the electrode, the potential of the cell becomes more positive when the switch is closed (Figure 1.3B). Such potentials are called depolarizations. The polarized state of the membrane is decreased. A larger battery produces even larger depolarizations. Again, the magnitude of the response is proportional to the magnitude of the stimulus. However, an unusual event occurs when the magnitude of the depolarization reaches a level of membrane potential called the threshold. A totally new type of signal is initiated; the action potential. Note that if the size of the battery is increased even more, the amplitude of the action potential is the same as the previous one (Figure 1.3B). The process of eliciting an action potential in a nerve cell is analogous to lighting a fuse with a heat source. A certain minimum temperature (threshold) is necessary. Temperatures less than the threshold fail to ignite the fuse. Temperatures greater than the threshold ignite the fuse just as well as the threshold temperature and the fuse does not burn any brighter or hotter.
Figure 1.3. See Neuroscience Online for animation.
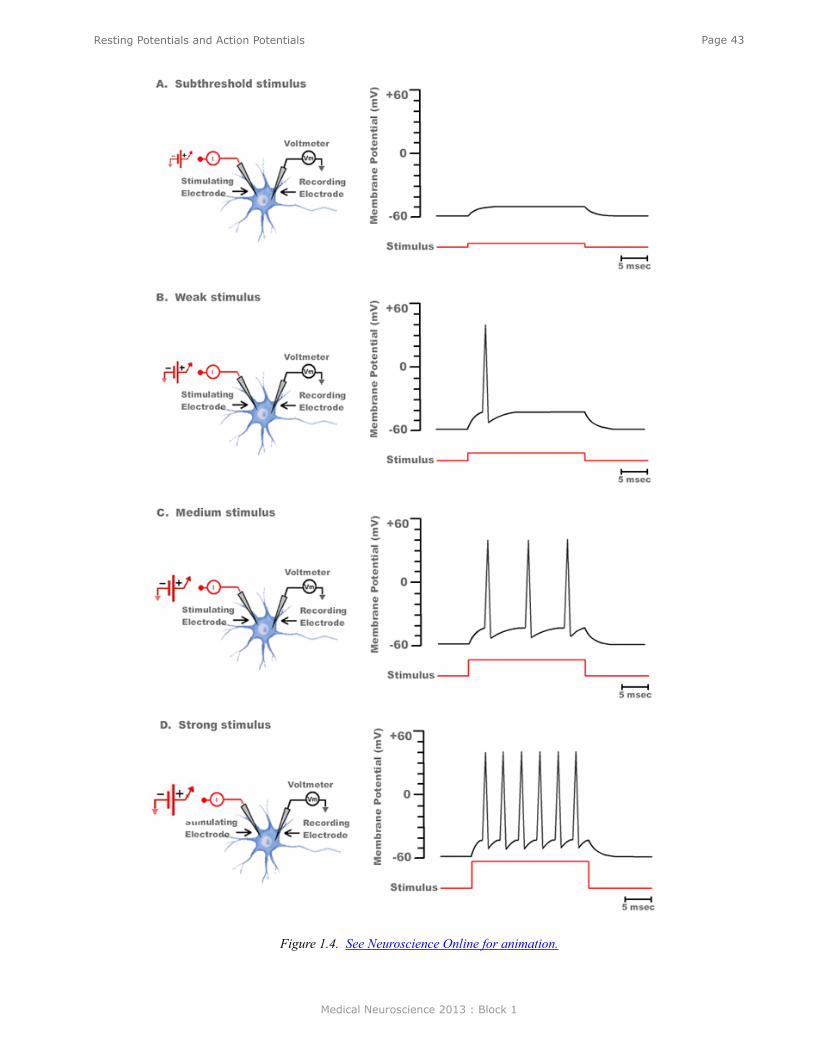
Medical Neuroscience 2013 : Block 1
Page 43Resting Potentials and Action Potentials
Figure 1.4. See Neuroscience Online for animation.

Medical Neuroscience 2013 : Block 1
Page 44Resting Potentials and Action Potentials
If the suprathreshold current stimulus is long enough, however, a train of action potentials will be elicited. In general, the action potentials will continue to fire as long as the stimulus continues, with the frequency of firing being proportional to the magnitude of the stimulus (Figure 1.4).
Action potentials are not only initiated in an all-or-nothing fashion, but they are also propagated in an all-or-nothing fashion. An action potential initiated in the cell body of a motor neuron in the spinal cord will propagate in an undecremented fashion all the way to the synaptic terminals of that motor neuron. Again, the situation is analogous to a burning fuse. Once the fuse is ignited, the flame will spread to its end.
ComPonenTs of The aCTion PoTenTials
The action potential consists of several components (Figure 1.3B). The threshold is the value of the membrane potential which, if reached, leads to the all-or-nothing initiation of an action potential. The initial or rising phase of the action potential is called the depolarizing phase or the upstroke. The region of the action potential between the 0 mV level and the peak amplitude is the overshoot. The return of the membrane potential to the resting potential is called the repolarization phase. There is also a phase of the action potential during which time the membrane potential can be more negative than the resting potential. This phase of the action potential is called the undershoot or the hyperpolarizing afterpotential. In Figure 1.4 the undershoots of the action potentials do not become more negative than the resting potential because they are “riding” on the constant depolarizing stimulus.
ioniC meChanisms of resTing PoTenTials
Before examining the ionic mechanisms of action potentials, it is first necessary to understand the ionic mechanisms of the resting potential. The two phenomena are intimately related. The story of the resting potential goes back to the early 1900’s when Julius Bernstein suggested that the resting potential (Vm) was equal to the potassium equilibrium potential (EK). Where
the key to understanding the resting potential is the fact that ions are distributed unequally on the inside and outside of cells, and that cell membranes are selectively permeable to different ions. K+ is particularly important for the resting potential. The membrane is highly permeable to K+. In addition, the inside of the cell has a high concentration of K+ ([K+]i) and the outside of the cell has a low concentration of K+ ([K+]o). Thus, K+ will naturally move by diffusion from its region of high concentration to its region of low concentration. Consequently, the positive K+ ions leaving the inner surface of the membrane leave behind some negatively charged ions. That negative charge attracts the positive charge of the K+ ion that is leaving and tends to "pull it back". Thus, there will be an electrical force directed inward that will tend to counterbalance the diffusional force directed outward. Eventually, an equilibrium will be established; the concentration force moving K+ out will balance the electrical force holding it in. The potential at which that balance is achieved is called the Nernst Equilibrium Potential.
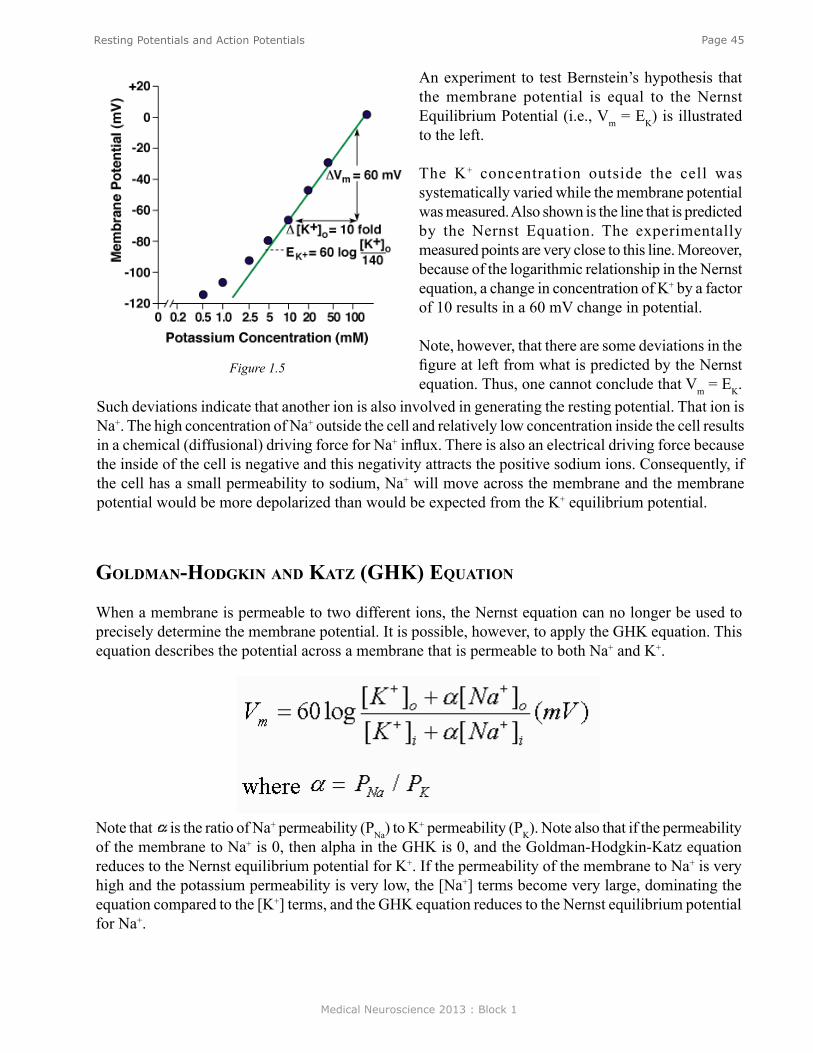
Medical Neuroscience 2013 : Block 1
Page 45Resting Potentials and Action Potentials
An experiment to test Bernstein’s hypothesis that the membrane potential is equal to the Nernst Equilibrium Potential (i.e., Vm = EK) is illustrated to the left.
The K+ concentration outside the cell was systematically varied while the membrane potential was measured. Also shown is the line that is predicted by the Nernst Equation. The experimentally measured points are very close to this line. Moreover, because of the logarithmic relationship in the Nernst equation, a change in concentration of K+ by a factor of 10 results in a 60 mV change in potential.
Note, however, that there are some deviations in the figure at left from what is predicted by the Nernst equation. Thus, one cannot conclude that Vm = EK.
Figure 1.5
Such deviations indicate that another ion is also involved in generating the resting potential. That ion is Na+. The high concentration of Na+ outside the cell and relatively low concentration inside the cell results in a chemical (diffusional) driving force for Na+ influx. There is also an electrical driving force because the inside of the cell is negative and this negativity attracts the positive sodium ions. Consequently, if the cell has a small permeability to sodium, Na+ will move across the membrane and the membrane potential would be more depolarized than would be expected from the K+ equilibrium potential.
goldman-hodgkin and kaTz (ghk) equaTion
When a membrane is permeable to two different ions, the Nernst equation can no longer be used to precisely determine the membrane potential. It is possible, however, to apply the GHK equation. This equation describes the potential across a membrane that is permeable to both Na+ and K+.
Note that is the ratio of Na+ permeability (PNa) to K+ permeability (PK). Note also that if the permeability of the membrane to Na+ is 0, then alpha in the GHK is 0, and the Goldman-Hodgkin-Katz equation reduces to the Nernst equilibrium potential for K+. If the permeability of the membrane to Na+ is very high and the potassium permeability is very low, the [Na+] terms become very large, dominating the equation compared to the [K+] terms, and the GHK equation reduces to the Nernst equilibrium potential for Na+.

Medical Neuroscience 2013 : Block 1
Page 46Resting Potentials and Action Potentials
If the GHK equation is applied to the same data in Figure 1.5, there is a much better fit. The value of alpha needed to obtain this good fit was 0.01. This means that the potassium K+ permeability is 100 times the Na+ permeability. In summary, the resting potential is due not only to the fact that there is a high permeability to K+. There is also a slight permeability to Na+, which tends to make the membrane potential slightly more positive than it would have been if the membrane were permeable to K+ alone.
Figure 1.6
membrane PoTenTial laboraTory
See Neuroscience Online for the interactive Membrane Potential Laboratory. It allows one to experiment with the effects of altering external or internal potassium ion concentration and membrane permeability to sodium and potassium ions. Predictions are made using the Nernst and the Goldman, Hodgkin, Katz equations.
Test Your Knowledge (See Neuroscience Online for answers to the questions.)
1. If a nerve membrane suddenly became equally permeable to both Na+ and K+, the membrane potential would:
A. Not change B. Approach the new K+ equilibrium potential C. Approach the new Na+ equilibrium potential D. Approach a value of about 0 mV E. Approach a constant value of about +55 mV
2. If the concentration of K+ in the cytoplasm of an invertebrate axon is changed to a new value of 200 mM (Note: for this axon normal [K]o = 20 mM and normal [K]i = 400 mM):
F. The membrane potential would become more negative G. The K+ equilibrium potential would change by 60 mV H. The K+ equilibrium potential would be about -60 mV I. The K+ equilibrium potential would be about -18 mV J. An action potential would be initiated

Medical Neuroscience 2013 : Block 1
Page 47Ionic Mechanisms of Action Potentials
Ionic Mechanisms of Action Potentials
assignmenT
I. Sequence of permeability changes underlying the nerve action potential: The Hodgkin and Huxley model A. Voltage clamp analysisB. Voltage-dependent Na+ permeabilityC. Na+ - inactivationD. Voltage-dependent K+ permeabilityE. Quantitative reconstruction of the action potential
II. Pharmacology of voltage-dependent membrane channelsA. Tetrodotoxin (TTX)B. Tetraethylammonium (TEA)
III. Types of membrane channelsIV. ChannelopathiesV. Refractory periods
A. AbsoluteB. Relative
major objeCTives
1. Have a detailed understanding of the ionic mechanisms underlying the initiation and repolarization of the nerve action potential.
2. Know the specific role played by individual membrane channels in the action potential.3. Know the ionic basis of the absolute and relative refractory periods.
required reading
● Byrne, J.H. Ionic Mechanisms of Action Potentials. Neuroscience Online: http://nba.uth.tmc.edu/neuroscience/s1/chapter02.html; or
● Byrne, J.H. Ionic Mechanisms of Action Potentials, Lecture Supplement Volume I.
suggesTed reading
● Byrne, J. H., Roberts, J. L., From Molecules to Networks: An Introduction to Cellular and Molecular Neuroscience, 2nd Edition, Elsevier, Amsterdam, 2009, Chapters 5, 6, 7.
Lecturer: John H. Byrne, Ph.D.January 11, 2013 | 8:00 AM
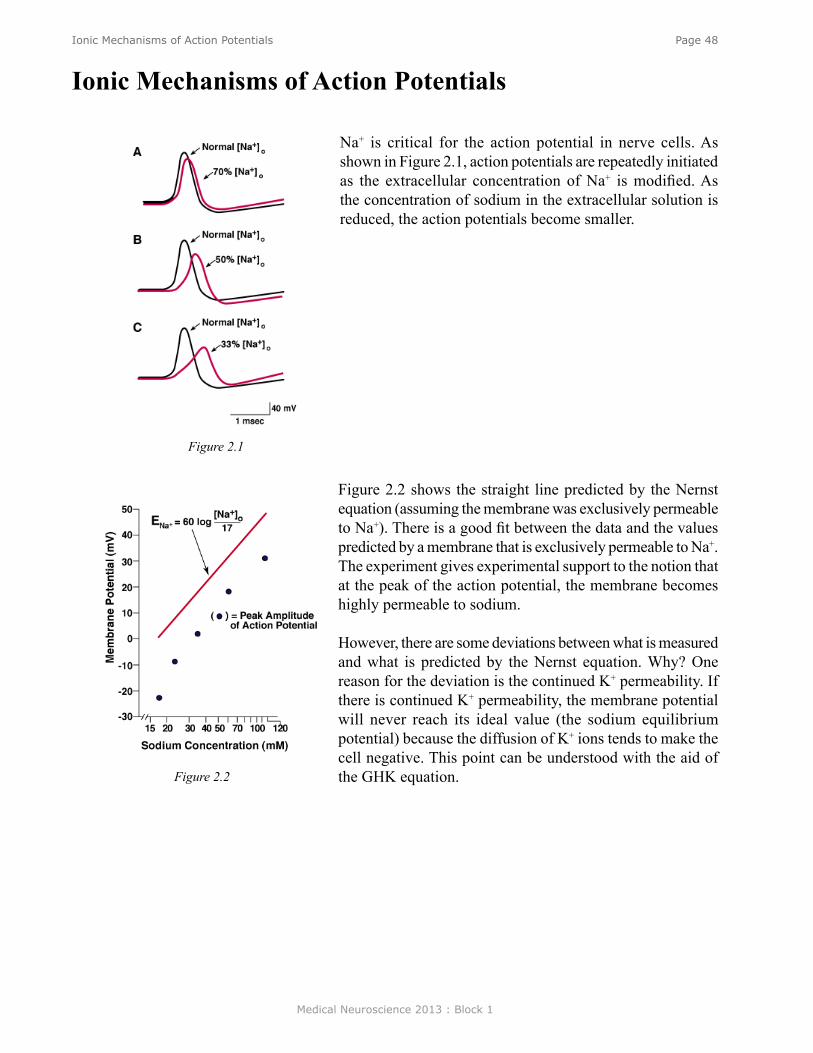
Medical Neuroscience 2013 : Block 1
Page 48Ionic Mechanisms of Action Potentials
Na+ is critical for the action potential in nerve cells. As shown in Figure 2.1, action potentials are repeatedly initiated as the extracellular concentration of Na+ is modified. As the concentration of sodium in the extracellular solution is reduced, the action potentials become smaller.
Ionic Mechanisms of Action Potentials
Figure 2.2 shows the straight line predicted by the Nernst equation (assuming the membrane was exclusively permeable to Na+). There is a good fit between the data and the values predicted by a membrane that is exclusively permeable to Na+. The experiment gives experimental support to the notion that at the peak of the action potential, the membrane becomes highly permeable to sodium.
However, there are some deviations between what is measured and what is predicted by the Nernst equation. Why? One reason for the deviation is the continued K+ permeability. If there is continued K+ permeability, the membrane potential will never reach its ideal value (the sodium equilibrium potential) because the diffusion of K+ ions tends to make the cell negative. This point can be understood with the aid of the GHK equation.
Figure 2.1
Figure 2.2
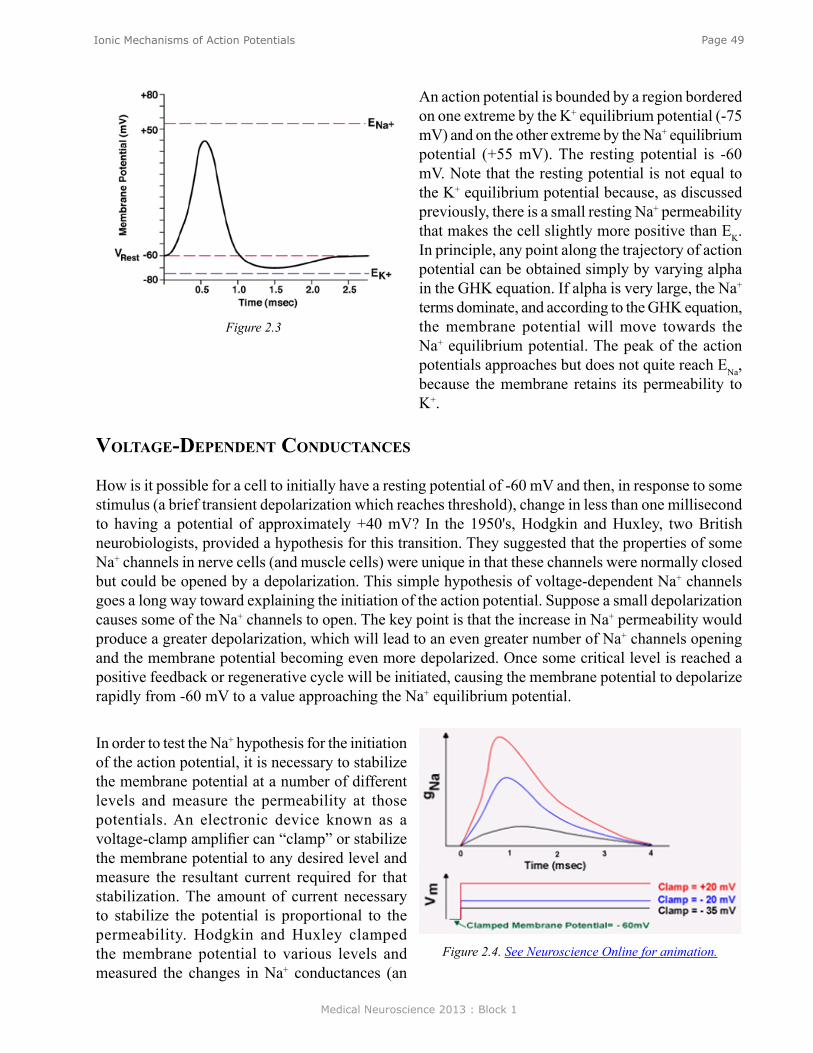
Medical Neuroscience 2013 : Block 1
Page 49Ionic Mechanisms of Action Potentials
An action potential is bounded by a region bordered on one extreme by the K+ equilibrium potential (-75 mV) and on the other extreme by the Na+ equilibrium potential (+55 mV). The resting potential is -60 mV. Note that the resting potential is not equal to the K+ equilibrium potential because, as discussed previously, there is a small resting Na+ permeability that makes the cell slightly more positive than EK. In principle, any point along the trajectory of action potential can be obtained simply by varying alpha in the GHK equation. If alpha is very large, the Na+ terms dominate, and according to the GHK equation, the membrane potential will move towards the Na+ equilibrium potential. The peak of the action potentials approaches but does not quite reach ENa, because the membrane retains its permeability to K+.
Figure 2.3
volTage-dePendenT ConduCTanCes
How is it possible for a cell to initially have a resting potential of -60 mV and then, in response to some stimulus (a brief transient depolarization which reaches threshold), change in less than one millisecond to having a potential of approximately +40 mV? In the 1950's, Hodgkin and Huxley, two British neurobiologists, provided a hypothesis for this transition. They suggested that the properties of some Na+ channels in nerve cells (and muscle cells) were unique in that these channels were normally closed but could be opened by a depolarization. This simple hypothesis of voltage-dependent Na+ channels goes a long way toward explaining the initiation of the action potential. Suppose a small depolarization causes some of the Na+ channels to open. The key point is that the increase in Na+ permeability would produce a greater depolarization, which will lead to an even greater number of Na+ channels opening and the membrane potential becoming even more depolarized. Once some critical level is reached a positive feedback or regenerative cycle will be initiated, causing the membrane potential to depolarize rapidly from -60 mV to a value approaching the Na+ equilibrium potential.
In order to test the Na+ hypothesis for the initiation of the action potential, it is necessary to stabilize the membrane potential at a number of different levels and measure the permeability at those potentials. An electronic device known as a voltage-clamp amplifier can “clamp” or stabilize the membrane potential to any desired level and measure the resultant current required for that stabilization. The amount of current necessary to stabilize the potential is proportional to the permeability. Hodgkin and Huxley clamped the membrane potential to various levels and measured the changes in Na+ conductances (an
Figure 2.4. See Neuroscience Online for animation.
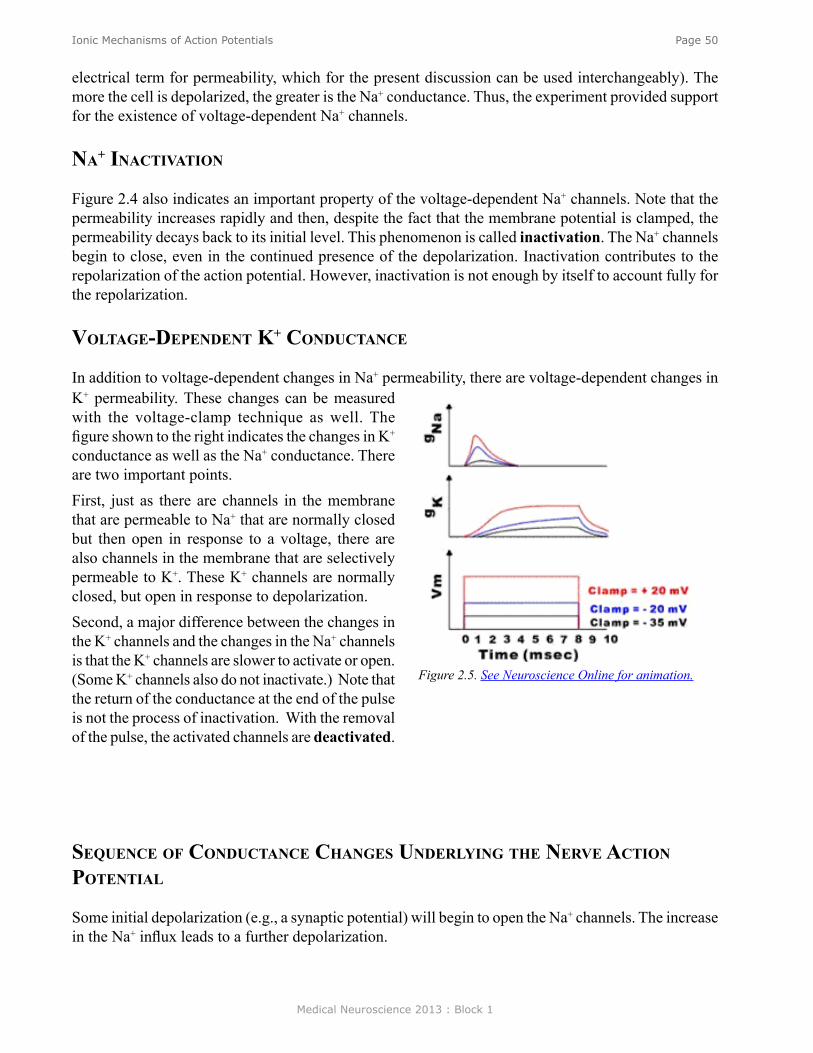
Medical Neuroscience 2013 : Block 1
Page 50Ionic Mechanisms of Action Potentials
electrical term for permeability, which for the present discussion can be used interchangeably). The more the cell is depolarized, the greater is the Na+ conductance. Thus, the experiment provided support for the existence of voltage-dependent Na+ channels.
na+ inaCTivaTion
Figure 2.4 also indicates an important property of the voltage-dependent Na+ channels. Note that the permeability increases rapidly and then, despite the fact that the membrane potential is clamped, the permeability decays back to its initial level. This phenomenon is called inactivation. The Na+ channels begin to close, even in the continued presence of the depolarization. Inactivation contributes to the repolarization of the action potential. However, inactivation is not enough by itself to account fully for the repolarization.
volTage-dePendenT k+ ConduCTanCe
In addition to voltage-dependent changes in Na+ permeability, there are voltage-dependent changes in K+ permeability. These changes can be measured with the voltage-clamp technique as well. The figure shown to the right indicates the changes in K+ conductance as well as the Na+ conductance. There are two important points. First, just as there are channels in the membrane that are permeable to Na+ that are normally closed but then open in response to a voltage, there are also channels in the membrane that are selectively permeable to K+. These K+ channels are normally closed, but open in response to depolarization. Second, a major difference between the changes in the K+ channels and the changes in the Na+ channels is that the K+ channels are slower to activate or open. (Some K+ channels also do not inactivate.) Note that the return of the conductance at the end of the pulse is not the process of inactivation. With the removal of the pulse, the activated channels are deactivated.
Figure 2.5. See Neuroscience Online for animation.
sequenCe of ConduCTanCe Changes underlying The nerve aCTion PoTenTial
Some initial depolarization (e.g., a synaptic potential) will begin to open the Na+ channels. The increase in the Na+ influx leads to a further depolarization.
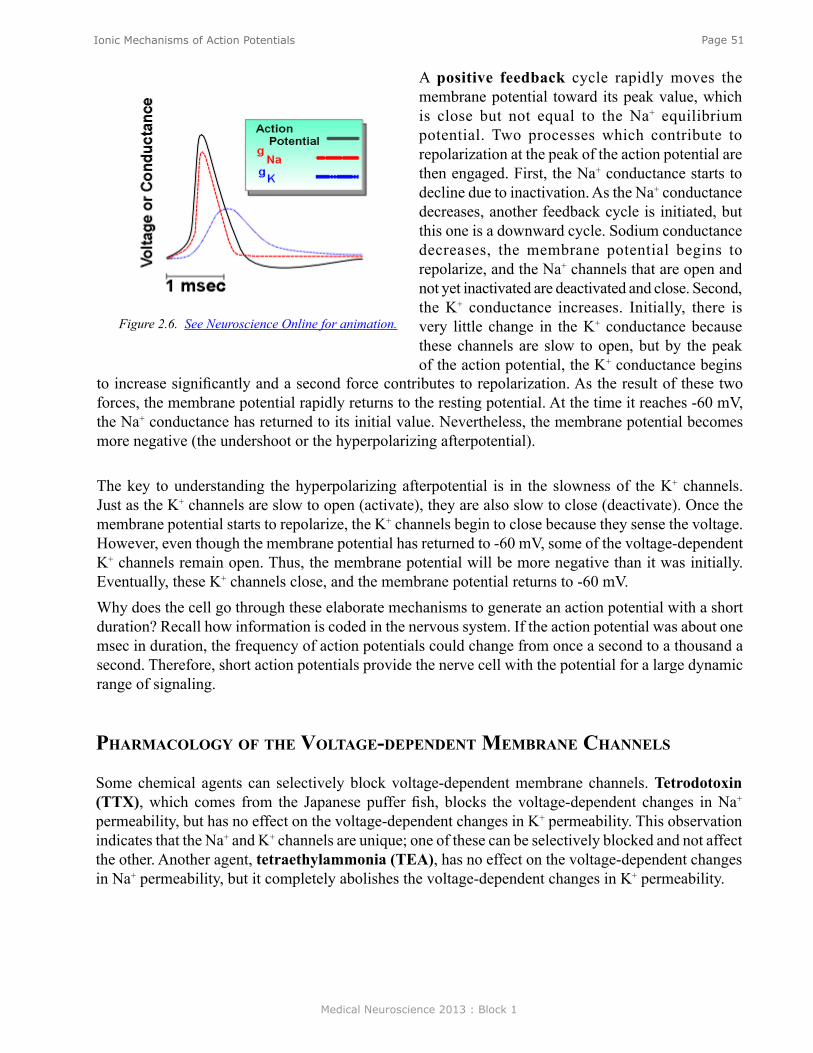
Medical Neuroscience 2013 : Block 1
Page 51Ionic Mechanisms of Action Potentials
A positive feedback cycle rapidly moves the membrane potential toward its peak value, which is close but not equal to the Na+ equilibrium potential. Two processes which contribute to repolarization at the peak of the action potential are then engaged. First, the Na+ conductance starts to decline due to inactivation. As the Na+ conductance decreases, another feedback cycle is initiated, but this one is a downward cycle. Sodium conductance decreases, the membrane potential begins to repolarize, and the Na+ channels that are open and not yet inactivated are deactivated and close. Second, the K+ conductance increases. Initially, there is very little change in the K+ conductance because these channels are slow to open, but by the peak of the action potential, the K+ conductance begins
Figure 2.6. See Neuroscience Online for animation.
to increase significantly and a second force contributes to repolarization. As the result of these two forces, the membrane potential rapidly returns to the resting potential. At the time it reaches -60 mV, the Na+ conductance has returned to its initial value. Nevertheless, the membrane potential becomes more negative (the undershoot or the hyperpolarizing afterpotential).
The key to understanding the hyperpolarizing afterpotential is in the slowness of the K+ channels. Just as the K+ channels are slow to open (activate), they are also slow to close (deactivate). Once the membrane potential starts to repolarize, the K+ channels begin to close because they sense the voltage. However, even though the membrane potential has returned to -60 mV, some of the voltage-dependent K+ channels remain open. Thus, the membrane potential will be more negative than it was initially. Eventually, these K+ channels close, and the membrane potential returns to -60 mV. Why does the cell go through these elaborate mechanisms to generate an action potential with a short duration? Recall how information is coded in the nervous system. If the action potential was about one msec in duration, the frequency of action potentials could change from once a second to a thousand a second. Therefore, short action potentials provide the nerve cell with the potential for a large dynamic range of signaling.
PharmaCology of The volTage-dePendenT membrane Channels
Some chemical agents can selectively block voltage-dependent membrane channels. Tetrodotoxin (TTX), which comes from the Japanese puffer fish, blocks the voltage-dependent changes in Na+ permeability, but has no effect on the voltage-dependent changes in K+ permeability. This observation indicates that the Na+ and K+ channels are unique; one of these can be selectively blocked and not affect the other. Another agent, tetraethylammonia (TEA), has no effect on the voltage-dependent changes in Na+ permeability, but it completely abolishes the voltage-dependent changes in K+ permeability.

Medical Neuroscience 2013 : Block 1
Page 52Ionic Mechanisms of Action Potentials
PumPs and leaks
It is easy to receive the impression that there is a "gush" of Na+ that comes into the cell with each action potential. Although, there is some influx of Na+, it is minute compared to the intracellular concentration of Na+. The influx is insufficient to make any noticeable change in the intracellular concentration of Na+. Therefore, the Na+ equilibrium potential does not change during or after an action potential. For any individual action potential, the amount of Na+ that comes into the cell and the amount of K+ that leaves
Figure 2.7 Figure 2.8
Use these two agents (TTX and TEA) to test your understanding of the ionic mechanisms of the action potential. (Figure 2.9) What effect would treating an axon with TTX have on an action potential? An action potential would not occur because an action potential in an axon cannot be initiated without voltage-dependent Na+ channels. How would TEA affect the action potential? It would be longer and would not have an undershoot.
In the presence of TEA the initial phase of the action potential is identical, but note that it is much longer and does not have an after-hyperpolarization. There is a repolarization phase, but now the repolarization is due to the process of Na+ inactivation alone. Note that in the presence of TEA, there is no change in the resting potential. The channels in the membrane that endow the cell with the resting potential are different from the ones that are opened by voltage. They are not blocked by TEA. TEA only affects the voltage-dependent changes in K+ permeability.
Figure 2.9

Medical Neuroscience 2013 : Block 1
Page 53Ionic Mechanisms of Action Potentials
Test Your Knowledge (See Neuroscience Online for answer to the question.)
3. Drug X, when applied to a nerve axon, results in both a gradual decrease in the amplitude of individual action potentials and a depolarization of the resting potential, both of which develop over a period of several hours. The drug is most likely:
A. Blocking the voltage-dependent Na+ permeabilityB. Blocking the voltage-dependent K+ permeability C. Blocking the (Na+ -K+) pump D. Blocking the process of Na+ inactivation E. Increasing the rate at which voltage-dependent changes in K+
permeability occur
are insignificant and have no effect on the bulk concentrations. However, without some compensatory mechanism, over the long-term (many spikes), Na+ influx and K+ efflux would begin to alter the concentrations and the resultant Na+ and K+ equilibrium potentials. The Na+-K+ pumps in nerve cells provide for the long-term maintenance of these concentration gradients. They keep the intracellular concentrations of K+ high and the Na+ low, and thereby maintain the Na+ equilibrium potential and the K+ equilibrium potential. The pumps are necessary for the long-term maintenance of the "batteries" so that resting potentials and action potentials can be supported.
TyPes of membrane Channels
So far, two basic classes of channels, voltage-dependent or voltage-gated channels and voltage-independent channels, have been considered. Voltage-dependent channels can be further divided based on their permeation properties into voltage-dependent Na+ channels and voltage-dependent K+
channels. There are also voltage-dependent Ca2+ channels (see chapter on Synaptic Transmission). Indeed, there are multiple types of Ca2+ channels and voltage-dependent K+ channels. Nevertheless, all these channels are conceptually similar. They are membrane channels that are normally closed and as a result of changes in potential, the channel (pore) is opened. The amino acid sequence of these channels is known in considerable detail and specific amino acid sequences have been related to specific aspects of channel function (e.g., ion selectivity, voltage gating, inactivation). A third major channel class, the transmitter-gated or ligand-gated channels, will be described later.
ChanneloPaTies
Ion channel mutations have been identified as a possible cause of a wide variety of inherited disor-ders. Several disorders involving muscle membrane excitability have been asscociated with muta-tions in calcium, sodium and chloride channels as well as acetylcholine receptors and have been labeled ‘channelopathies’. It is possible that movement disorders, epilepsy and headache, as well as other rare inherited diseases, might be linked to ion channels. The manifestations and mechanisms of channelopathies affecting neurons are reviewed in Kullman, 2002. The existence of channelopa-thies may provide insights into the variety of cellular mechanisms associated with the misfunction-ing of neuronal circuits.

Medical Neuroscience 2013 : Block 1
Page 54Ionic Mechanisms of Action Potentials
a change in the Na+ conductance, which is identical to the first pulse. However, if the second pulse comes soon after the first pulse, then the change in Na+ conductance produced by the second pulse is less than that produced by the first. Indeed, if the second pulse occurs immediately after the first pulse, the second pulse produces no change in the Na+ conductance. Therefore, when the Na+ channels open and spontaneously inactivate, it takes time (several msec) for them to recover from that inactivation. This process of recovery from inactivation underlies the absolute refractory period. During an action potential the Na+ channels open and then they become inactivated. Therefore, if a second stimulus is delivered soon after the one that initiated the first spike, there will be few Na+ channels available to be opened by the second stimulus because they have been inactivated by the first action potential.
Figure 2.10
aCTion PoTenTial laboraTory
See Neuroscience Online version for the interactive Action Potential Laboratory. The laboratory allows one to examine the ways in which the action potential is affected by changes in the Na+ conductance, K+ conductance and equilibrium potentials for Na+ and K+.
absoluTe and relaTive refraCTory Periods
The absolute refractory period is a period of time after the initiation of one action potential when it is impossible to initiate a second action potential no matter how much the cell is depolarized. The relative refractory period is a period after one action potential is initiated when it is possible to initiate a second action potential, but only with a greater depolarization than was necessary to initiate the first. The relative refractory period can be understood at least in part by the hyperpolarizing afterpotential. Assume that an initial stimulus depolarized a cell from -60 mV to -45 mV in order to reach threshold and then consider delivering the same 15-mV stimulus sometime during the after-hyperpolarization. The stimulus would again depolarize the cell but the depolarization would be below threshold and insufficient to trigger an action potential. If the stimulus was made larger, however, such that it again was capable of depolarizing the cell to threshold (-45 mV), an action potential could be initiated.
The absolute refractory period can be explained by the dynamics of the process of Na+-inactivation, the features of which are illustrated in Figure 2.10. Here, two voltage clamp pulses are delivered. The first pulse produces a voltage-dependent increase in the Na+ permeability which then undergoes the process of inactivation. If the two pulses are separated sufficiently in time, the second pulse produces

Medical Neuroscience 2013 : Block 1
Page 55Propagation of Action Potentials
Propagation of Action Potentials
assignmenT
I. Passive electrical properties of the membrane A. Time constant B. Space (length) constant
II. Effect of passive membrane properties on the propagation velocity of the action potentialIII. Saltatory conduction in myelinated axons
major objeCTives
1. Know the determinants of the passive properties of the membrane.2. Know about the ways in which the propagation velocity is affected by the passive
properties of membranes.3. Be able to compare and contrast the properties of myelinated and unmyelinated axons.
required reading
● Byrne, J.H. Propagation of Action Potentials. Neuroscience Online: http://nba.uth.tmc.edu/neuroscience/s1/chapter03.html or
● Byrne, J.H. Propagation of Action Potentials, Lecture Supplement Volume I
suggesTed reading ● Byrne, J. H., Roberts, J. L., From Molecules to Networks: An Introduction to Cellular and
Molecular Neuroscience, 2nd Edition, Elsevier, Amsterdam, 2009, Chapters 4 and 5.
Lecturer: John H. Byrne, Ph.D.January 11, 2013 | 9:00 AM

Medical Neuroscience 2013 : Block 1
Page 56Propagation of Action Potentials
Propagation of Action Potentials
Changes in The sPaTial disTribuTion of Charge
Once an action potential is initiated at one point in the nerve cell, how does it propagate to the synaptic terminal region in an all-or-nothing fashion?
Figure 3.1 shows a schematic diagram of an axon and the charge distributions that would be expected to occur along the membrane of that axon. Positive charges exist on the outside of the axon and negative charges on the inside. Now consider the consequences of delivering some stimulus to a point in the middle of the axon. If the depolarization is sufficiently large, voltage-dependent sodium channels will be opened, and an action potential will be initiated. Consider for the moment “freezing” the action potential at its peak value. Its peak value now will be about +40 mV inside with respect to the outside. Unlike charges attract, so the positive charge will move to the adjacent region of the
membrane. As the charge moves to the adjacent region of the membrane, the adjacent region of the membrane will depolarize. If it depolarizes sufficiently, as it will, voltage-dependent sodium channels in the adjacent region of the membrane will be opened and a “new” action potential will be initiated. This charge distribution will then spread to the next region and initiate other “new” action potentials. One way of viewing this process is with a thermal analogue. You can think of an axon as a piece of wire coated with gunpowder (the gunpowder is analogous to the sodium channels). If a sufficient stimulus (heat) is delivered to the wire, the gunpowder will ignite, generate heat, and the heat will spread along the wire to adjacent regions and cause the gunpowder in the adjacent regions to ignite.
deTerminanTs of ProPagaTion veloCiTy
A great variability is found in the velocity of the propagation of action potentials. In fact, the propagation velocity of the action potentials in nerves can vary from 100 meters per second (580 miles per hour) to less than a tenth of a meter per second (0.6 miles per hour). Why do some axons propagate information very rapidly and others slowly? In order to understand how this process works, it is necessary to consider two so-called passive properties of membranes, the time constant and the space or length constant. Why are these called passive properties? They have nothing to do with any of the voltage-dependent conductances discussed earlier. They have nothing to do with any pumps or exchangers. They are intrinsic properties of all biological membranes.
tiMe coNstaNt.First, consider a thermal analogue. Place a block of metal at 10oC on a hotplate at 100oC. How would the temperature change? It will increase from its initial value of 10oC to a final value of 100oC. But the temperature will not change instantly. In fact, it would change as an exponential function of time. An analogous situation occurs in nerve cells, when they receive an instantaneous stimulus. The figure at right represents an idealized nerve cell. The recording electrode initially measures a potential of -60 mV (the resting potential). At some point in time (time 0), the switch is closed. The switch closure
Figure 3.1. See Neuroscience Online for animation.
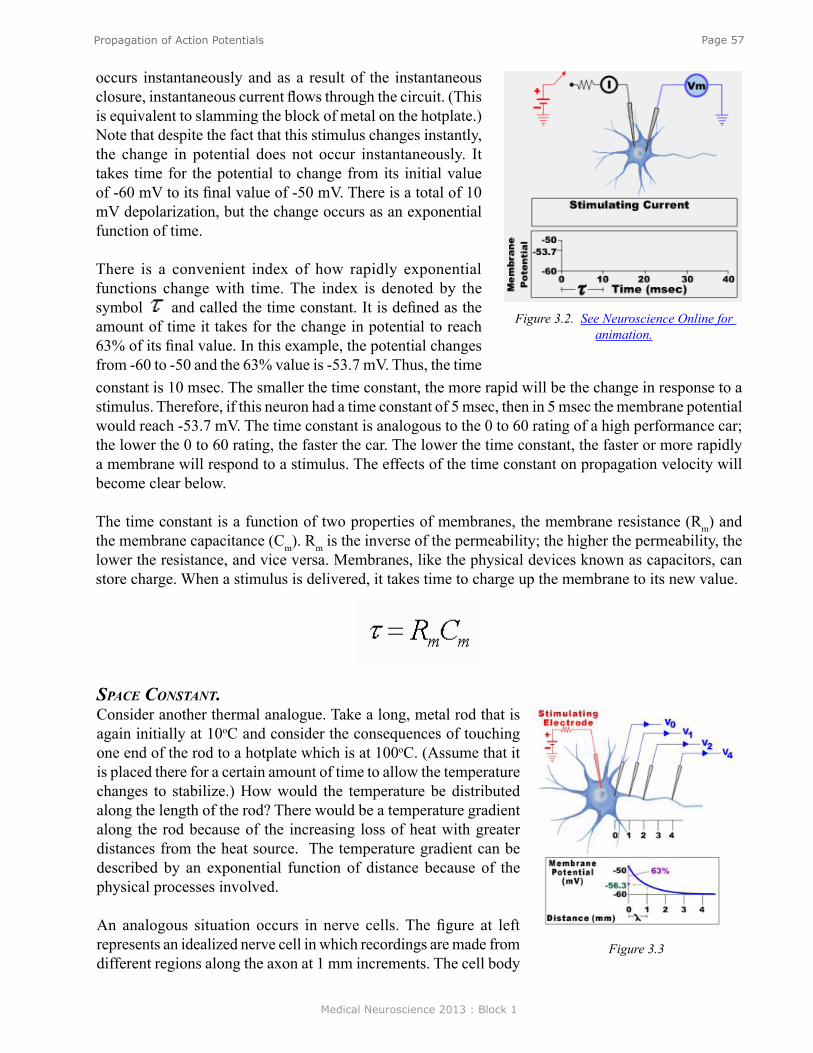
Medical Neuroscience 2013 : Block 1
Page 57Propagation of Action Potentials
occurs instantaneously and as a result of the instantaneous closure, instantaneous current flows through the circuit. (This is equivalent to slamming the block of metal on the hotplate.) Note that despite the fact that this stimulus changes instantly, the change in potential does not occur instantaneously. It takes time for the potential to change from its initial value of -60 mV to its final value of -50 mV. There is a total of 10 mV depolarization, but the change occurs as an exponential function of time.
There is a convenient index of how rapidly exponential functions change with time. The index is denoted by the symbol and called the time constant. It is defined as the amount of time it takes for the change in potential to reach 63% of its final value. In this example, the potential changes from -60 to -50 and the 63% value is -53.7 mV. Thus, the time
Figure 3.2. See Neuroscience Online for animation.
constant is 10 msec. The smaller the time constant, the more rapid will be the change in response to a stimulus. Therefore, if this neuron had a time constant of 5 msec, then in 5 msec the membrane potential would reach -53.7 mV. The time constant is analogous to the 0 to 60 rating of a high performance car; the lower the 0 to 60 rating, the faster the car. The lower the time constant, the faster or more rapidly a membrane will respond to a stimulus. The effects of the time constant on propagation velocity will become clear below.
The time constant is a function of two properties of membranes, the membrane resistance (Rm) and the membrane capacitance (Cm). Rm is the inverse of the permeability; the higher the permeability, the lower the resistance, and vice versa. Membranes, like the physical devices known as capacitors, can store charge. When a stimulus is delivered, it takes time to charge up the membrane to its new value.
space coNstaNt. Consider another thermal analogue. Take a long, metal rod that is again initially at 10oC and consider the consequences of touching one end of the rod to a hotplate which is at 100oC. (Assume that it is placed there for a certain amount of time to allow the temperature changes to stabilize.) How would the temperature be distributed along the length of the rod? There would be a temperature gradient along the rod because of the increasing loss of heat with greater distances from the heat source. The temperature gradient can be described by an exponential function of distance because of the physical processes involved.
An analogous situation occurs in nerve cells. The figure at left represents an idealized nerve cell in which recordings are made from different regions along the axon at 1 mm increments. The cell body
Figure 3.3

Medical Neuroscience 2013 : Block 1
Page 58Propagation of Action Potentials
is impaled with a stimulating electrode connected to a battery, the value of which changes the potential of the cell body to -50 mV (the equivalent of putting a 10oC rod on a 100oC hot plate). This axon, even though it initially had a spatially uniform resting potential of -60 mV, now has a potential of -50 mV in the soma because that is the region in which the stimulus is applied. However, the potential is not -50 mV all along the axon; it varies as a function of distance from the soma. One mm away the potential is -56 mV; at 2 mm away it is even closer to -60 mV; and far enough along the axon, the potential of the axon is -60 mV, the resting potential. Just as there is an index for how a change in potential changes with the time (the time constant), there is also an index denoted by the symbol λ (called the space constant or the length constant) which is an indication of how far a potential will spread along an axon in response to a subthreshold stimulus at another point. In Figure 3.3, the space constant or length constant is 1 mm. In 1 mm the potential will change by 63% of its final value. If λ was greater than 1 mm, the potential would spread a greater distance. If λ was 1/2 mm, the potential would spread less along the axon. Thus, whereas the time constant is an index of how rapidly a membrane would respond to a stimulus in time, the space constant is an index of how well a subthreshold potential will spread along an axon as a function of distance. The space constant is a passive property of membranes. Although it influences the rate of propagation of the action potentials, it is an independent process. It is like the surface of a race track and the action potential is like the race car. If the surface is muddy, the car will go slow, if it is firm and paved, the same car will be able to go much faster.
The length constant can be described in terms of the physical parameters of the axon, where d is the diameter of the axon, Rm is, as before, the membrane resistance, the inverse of the permeability, and Ri is the internal resistance (resistance of the axoplasm). Ri is an indicator of the ability of charges to move along the inner surface of the axon. A small subthreshold change in the charge distribution at one point along an axon will spread along the axon, but as it does
some will diffuse back out of the membrane and some will continue to move along the axon. If the resistance of the membrane (Rm) is high, less will leak out and relatively more will move along the axon. Increasing Rm is like putting insulation on a metal rod and heating the rod at one end. With more insulation (more resistance to heat loss to the outside of the rod), more heat will travel along the inside of the rod.
propagatioN velocity. How are the time constant and the space constant related to propagation velocity of action potentials? The smaller the time constant, the more rapidly a depolarization will affect the adjacent region. If a depolarization more rapidly affects an adjacent region, it will bring the adjacent region to threshold sooner. Therefore, the smaller the time constant, the more rapid will be the propagation velocity. If the space constant is large, a potential change at one point would spread a greater distance along the axon and bring distance regions to threshold sooner. Therefore, the greater the space constant, the more rapidly distant regions will be brought to threshold and the more rapid will be the propagation velocity. Thus, the propagation velocity is directly proportional to the space constant and inversely proportional to the time constant. There are separate equations that describe both the time constant and the space constant. The insight above allows us to make a new equation that combines the two.

Medical Neuroscience 2013 : Block 1
Page 59Propagation of Action Potentials
The equation provides insights into how it is possible for different axons to have different propagation velocities. One way of endowing an axon with a high propagation velocity is to increase the diameter. However, there is one serious problem in changing the propagation velocity by simply changing the diameter. To double the velocity, it is necessary to quadruple the diameter. Clearly there must be a better way of increasing propagation velocity than by simply increasing the diameter.
Figure 3.4. See Neuroscience Online for animation.
Another way to increase the propagation velocity is to decrease the membrane capacitance. This can be achieved by coating axons with a thick insulating sheath known as myelin. One potential problem with this approach is that the process of covering the axon would cover voltage-dependent Na+ channels. If Na+ channels are occluded, it would be impossible to generate an action potential. Instead of coating the entire axon with the myelin, only sections are coated and some regions called nodes are left bare.
ProPagaTion in myelinaTed fibers
Propagation of action potentials in myelinated fibers is illustrated in Figure 3.4. Start with an action potential at a node on the left. In the absence of myelin, the action potential would propagate actively through the simple mechanisms discussed above. However, now the myelin occludes all the voltage-dependent sodium channels so the action potential can not propagate actively. (In fact, myelinated axons do not even have sodium channels in the internodal region.) Rather, the potential change produced by the action potential at one node spreads in the internodal region along the axon passively just as the temperature would spread along a long metal rod. The potential spreads, but gets smaller (decrements), just as a temperature change induced at one end of a rod would get smaller as it spreads along a rod.
Now consider the point at which the passively spreading potential reaches the next node. A “new” action potential will be initiated. The stimulus for this action potential is the depolarization that emerges from the end of the myelin. Think of the gunpowder analogue again, but this time coat the rod with some insulation and put gunpowder only at the bare regions. Because of the insulation, a temperature change produced by the ignition of the gunpowder will spread effectively along the metal rod. The temperature will be sufficient to ignite the gunpowder at the next region and the process will repeat itself.

Medical Neuroscience 2013 : Block 1
Page 60Synaptic Transmission at Skeletal Neuromuscular Junction
Synaptic Transmission at Skeletal Neuromuscular Junction and
Mechanisms of Neurotransmitter Release
assignmenT
I. Motor end-plateII. End-plate potential (EPP)
A. Action of curareB. Electronic propagationC. Changes in permeability to Na+ and K+
III. Acetylcholine (ACh) hypothesis of transmitter releaseA. Acetylcholinesterase (AChE)B. Iontophoresis of ACh
IV. Presynaptic events underlying the release of AChA. Effects of action potential amplitude and durationB. Ca2+ hypothesis
V. Quantal nature of acetylcholine releaseA. Miniature end-plate potentials (MEPPs)B. Unit end-plate potentialsC. Role of Ca2+
major objeCTives
1. Have a complete mechanistic understanding of the sequence of events underlying synaptic transmission at the skeletal neuromuscular junction.
2. Understand the Ca2+ hypothesis for chemical synaptic transmission.3. Understand the differences between EPPs and MEPPs.4. Understand the ways in which ACh is released and is degraded.
required reading ● Byrne, J.H. Synaptic Transmission at Skeletal Neuromuscular Junction/Mechanisms of
Neurotransmitter Release; Neuroscience Online: http://nba.uth.tmc.edu/neuroscience/s1/chapter04.html or
● Byrne, J.H. Synaptic Transmission at Skeletal Neuromuscular Junction/Mechanisms of Neurotransmitter Release, Lecture Supplement Volume I.
suggesTed reading ● Byrne, J. H., Roberts, J. L., From Molecules to Networks: An Introduction to Cellular and
Molecular Neuroscience, 2nd Edition, Elsevier, Amsterdam, 2009, Chapters 8, 9, 11, 16.
Lecturer: John H. Byrne, Ph.D.January 14, 2013 | 8:00 AM

Medical Neuroscience 2013 : Block 1
Page 61Synaptic Transmission at Skeletal Neuromuscular Junction
The synapse is a specialized structure that allows one neuron to communicate with another neuron or a muscle cell. There are billions of nerve cells in the brain and each nerve cell can make and receive up to 10,000 synaptic connections with other nerve cells. Also, the strength of the synapse is modifiable. Changes in the strength of synapses endow the nervous system with the ability to store information.
anaTomy of The neuromusCular junCTion
The synapse for which most is known is the one formed between a spinal motor neuron and a skeletal muscle cell. Historically, it has been studied extensively because it is relatively easy to analyze. However, the basic properties of synaptic transmission at the skeletal neuromuscular junction are very similar to the process of synaptic transmission in the central nervous system. Consequently, an understanding of this synapse leads to an understanding of the others. Therefore, we will first discuss the process of synaptic transmission at the skeletal neuromuscular junction.
Figure 4.1. See Neuroscience Online for an enlarged view.
The features of the synaptic junction at the neuromuscular junction are shown in the figure at left. Skeletal muscle fibers are innervated by motor neurons whose cell bodies are located in the ventral horn of the spinal cord. The terminal region of the axon gives rise to very fine processes that run along skeletal muscle cells. Along these processes are specialized structures known as synapses. The particular synapse made between a spinal motor neuron and skeletal muscle cell is called the motor endplate because of its specific structure.
The synapse at the neuromuscular junction has three characteristic features of chemical synapses in the nervous system. First, there is a distinct separation between the presynaptic and the postsynaptic membrane. The space between the two is known as the synaptic cleft. The space tells us there must be some intermediary signaling mechanism between the presynaptic neuron and the postsynaptic neuron in order to have information flow across the synaptic cleft. Second, there is a characteristic high density of small spherical vesicles.
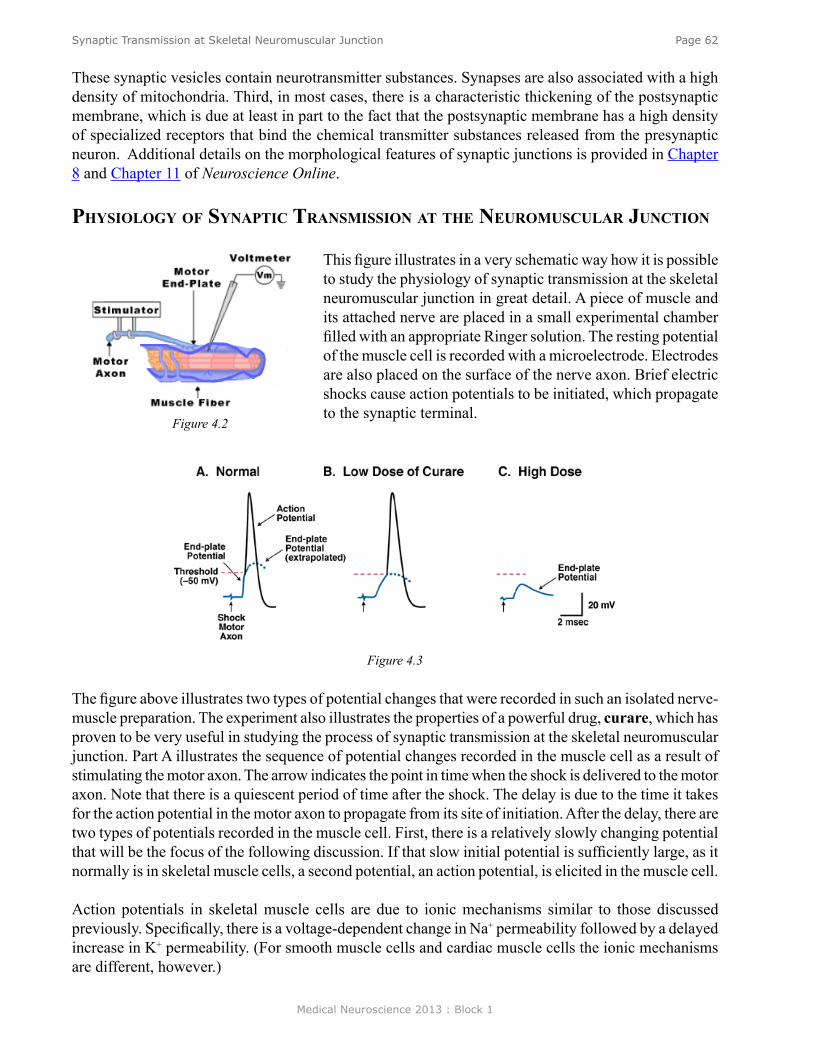
Medical Neuroscience 2013 : Block 1
Page 62Synaptic Transmission at Skeletal Neuromuscular Junction
These synaptic vesicles contain neurotransmitter substances. Synapses are also associated with a high density of mitochondria. Third, in most cases, there is a characteristic thickening of the postsynaptic membrane, which is due at least in part to the fact that the postsynaptic membrane has a high density of specialized receptors that bind the chemical transmitter substances released from the presynaptic neuron. Additional details on the morphological features of synaptic junctions is provided in Chapter 8 and Chapter 11 of Neuroscience Online.
Physiology of synaPTiC Transmission aT The neuromusCular junCTion
This figure illustrates in a very schematic way how it is possible to study the physiology of synaptic transmission at the skeletal neuromuscular junction in great detail. A piece of muscle and its attached nerve are placed in a small experimental chamber filled with an appropriate Ringer solution. The resting potential of the muscle cell is recorded with a microelectrode. Electrodes are also placed on the surface of the nerve axon. Brief electric shocks cause action potentials to be initiated, which propagate to the synaptic terminal.
The figure above illustrates two types of potential changes that were recorded in such an isolated nerve-muscle preparation. The experiment also illustrates the properties of a powerful drug, curare, which has proven to be very useful in studying the process of synaptic transmission at the skeletal neuromuscular junction. Part A illustrates the sequence of potential changes recorded in the muscle cell as a result of stimulating the motor axon. The arrow indicates the point in time when the shock is delivered to the motor axon. Note that there is a quiescent period of time after the shock. The delay is due to the time it takes for the action potential in the motor axon to propagate from its site of initiation. After the delay, there are two types of potentials recorded in the muscle cell. First, there is a relatively slowly changing potential that will be the focus of the following discussion. If that slow initial potential is sufficiently large, as it normally is in skeletal muscle cells, a second potential, an action potential, is elicited in the muscle cell. Action potentials in skeletal muscle cells are due to ionic mechanisms similar to those discussed previously. Specifically, there is a voltage-dependent change in Na+ permeability followed by a delayed increase in K+ permeability. (For smooth muscle cells and cardiac muscle cells the ionic mechanisms are different, however.)
Figure 4.2
Figure 4.3

Medical Neuroscience 2013 : Block 1
Page 63Synaptic Transmission at Skeletal Neuromuscular Junction
The underlying event that triggers the action potential can be revealed by taking advantage of curare, an arrow poison used by some South American Indians. A low dose of curare (Part B) reduces the underlying event, but it is still not sufficiently reduced to fall below threshold. If a somewhat higher dose of curare is delivered (Part C), the slow underlying event becomes subthreshold. The underlying signal is known as the endplate potential (EPP) because it is a potential change recorded at the motor endplate. Generally, it is known as an excitatory postsynaptic potential (EPSP).
Curare blocks the endplate potential because it is a competitive inhibitor of acetylcholine (ACh), the transmitter released at the presynaptic terminal. Curare does not block the voltage-dependent Na+ conductance or the voltage-dependent K+ conductance that underlies the muscle action potential. Curare affects the stimulus (the EPSP) which normally leads to the initiation of the muscle action potential. An animal that is poisoned with curare will asphyxiate because the process of neuromuscular transmission at respiratory muscles is blocked.
Normally, the magnitude of the endplate potential is quite large. It more than reaches threshold; it goes beyond threshold. In fact, the amplitude of the endplate potential is about 50 mV, but only about 30 mV is needed to reach threshold. The extra 20 mV is called the safety factor. Note that even if the endplate potential were to become somewhat smaller, the EPP would reach threshold and there would still be a one-to-one relationship between an axon potential and the motor axon and an action potential in the muscle cell.
ProPagaTion of The ePP
What are the properties of the EPP and how does it compare with the properties of the action potential? Is the endplate potential due to a voltage-dependent change in Na+ and K+ permeabilities like the action potential? Is the endplate potential propagated in an all-or-nothing fashion like the action potential?
The figure on the right illustrates an experiment that examines the propagation of the endplate potential. The muscle fiber is impaled repeatedly with electrodes at 1 mm intervals. (Note that the endplate potential is small because this experiment is done in the presence of a low concentration of curare so the endplate potential can be recorded without the complications of triggering an action potential.) The endplate potential is not propagated in an all-or-nothing fashion. It does spread along the muscle, but it does so with decrement. Thus, the spread of the endplate potential from its site of initiation to other sites along the muscle cell occurs passively and with decrement, just as a subthreshold potential change in one portion of the axon spreads along the axon, or just as a change in temperature at one point on a metal rod spreads along the rod.
Figure 4.4

Medical Neuroscience 2013 : Block 1
Page 64Synaptic Transmission at Skeletal Neuromuscular Junction
overview of The sequenCe of evenTs underlying The ePP
What are the other steps in the process of chemical synaptic transmission? Figure 4.5 provides an overview. Two factors control the duration of the EPSP at the neuromuscular junction. First, ACh is removed by diffusion. Second, a substance in the synaptic cleft, called acetylcholinesterase (AChE), hydrolyzes or breaks down ACh.
Figure 4.5. See Neuroscience Online for animation.
role of aChe
An important family of substances, one of which is neostigmine, inhibits the action of AChE. Neostigmine blocks the action of AChE, and thereby makes the endplate potential larger and longer in duration. This figure illustrates two endplate potentials. One was recorded in saline and curare and a second recorded after neostigmine was added to the solution. (Curare is added so that the properties of the EPP can be studied without triggering an action potential in the muscle cell.) After applying neostigmine the endplate potential is much larger and longer in duration.
myasThenia gravis
Myasthenia gravis is associated with severe muscular weakness because of a decrease in the number of acetylcholine receptors in the muscle cell. If the endplate potential is smaller, the endplate potential will fail to reach threshold. If it fails to reach threshold, there will be no action potential in the muscle cell and no contraction of the muscle, which causes muscular weak-ness. Neostigmine and other inhibitors of AChE are used to treat patients with myasthenia gravis. These agents make the amount of acetylcholine that is released more effectively reach the remain-ing acetylcholine receptors.
Figure 4.6

Medical Neuroscience 2013 : Block 1
Page 65Synaptic Transmission at Skeletal Neuromuscular Junction
ionToPhoresis of aCh
Iontophoresis is an interesting technique that can be used to further test the hypothesis that ACh is the neurotransmitter substance at the neuromuscular junction. If ACh is the transmitter that is released by this synapse, one would predict that it should be possible to substitute artificial application of the transmitter for the normal release of the transmitter. Because ACh is a positively charged molecule, it can be forced out of a microelectrode to simulate the release of ACh from a presynaptic terminal.
Figure 4.7. See Neuroscience Online for animation.
Figure 4.8
Indeed, minute amounts of ACh can be applied to the vicinity of the neuromuscular junction. This figure compares an EPP produced by stimulation of the motor axon and the response to ejections of ACh. The potential change looks nearly identical to the endplate potential produced by the normal release of ACh. This experiment provides experimental support for the concept that ACh is the natural transmitter at this synapse.
The response to the ejection of ACh has some other interesting properties that are all consistent with the cholinergic nature of the synapse at the skeletal neuromuscular junction. Neostigmine makes the response to the iontophoresis of ACh longer and larger. Curare reduces the response because it competes with the normal binding of ACh. If ACh is ejected into the muscle cell, nothing happens because the receptors for acetylcholine are not in the inside; they are on the outside of the muscle cell. Application of acetylcholine to regions of the muscle away from the end-plate produces no response because the receptors for the ACh are concentrated at the synaptic region.
To test your understanding so far, consider how an agent such as TTX would affect the generation of both an EPP and the response of a muscle fiber to the iontophoretic application of ACh? TTX has no effect on the response to ACh, but it does block the EPP. The reason the response to ACh is unaffected is clear, but many expect that if there is no effect here, there should be no effect on the EPP either. Tetrodotoxin does not affect the binding of acetylcholine to the receptors and therefore will not affect the response to direct application of ACh. However, tetrodotoxin will affect the ability of an action potential to be elicited in the motor axon. If an action potential cannot be elicited in the motor axon, it cannot cause the release of transmitter. Thus, tetrodotoxin would totally abolish the EPP. The block would not be due to a block of ACh receptors, but rather to a block of some step prior to the release of the transmitter.

Medical Neuroscience 2013 : Block 1
Page 66Synaptic Transmission at Skeletal Neuromuscular Junction
ioniC meChanisms of The ePP
Bernard Katz and his colleagues were pioneers in investigating mechanisms of synaptic transmission at the neuromuscular junction. They suggested that the channel opened by ACh was one that had equal permeability to both Na+ and K+. Because it was equally permeable to Na+ and K+, Katz suggested that, as a result of the opening of these channels, the membrane potential would move toward 0 mV. (A value of alpha in the GHK equation equal to one, which when substituted into the equation, yields a potential of about 0 mV.)
Figure 4.9. See Neuroscience Online for animation.
Test Your Knowledge(see Neuroscience Online for answer to question)
4. An endplate potential in a skeletal muscle cell could in principle be produced by a decreased permeability to which of the following ions(s)? (Assume that there is a finite initial permeability to each of the ions listed below and that physiological concentration gradients are present.):
A. Na+ B. Na+ and Ca2+ C. Ca2+ D. K+
This potential is also called the reversal potential. As a result of ACh binding to receptors, specific channels become equally permeable to Na+ and K+. (Figure 4.9) This permeability change tends to move the membrane potential from wherever it is initially towards a new potential of 0 mV.
Why does the normal endplate potential never actually reach 0 mV? One reason is that the sequence of permeability changes that underlie the action potential "swamp out" the changes produced by the EPP. But even if an action potential was not triggered, the EPP still would not reach 0 mV. This is because the ACh channels are only a small fraction of the total number of channels in muscle fibers. The K+ channels that endow the muscle cells with its resting potential are present as well. Their function is to try to maintain the cell at the resting potential.
The channel opened by ACh is a member of a general class of channels called ligand-gated channels or ionotropic receptors. As illustrated in Figure 4.9, the transmitter binding site is part of the channel itself. As a result of transmitter binding to the receptor (generally two molecules are necessary), there is a conformational change in the protein allowing a pore region to open and ions to flow down their electrochemical gradients. Additional details of the channel are presented in Chapter 11 of Neuroscience Online.
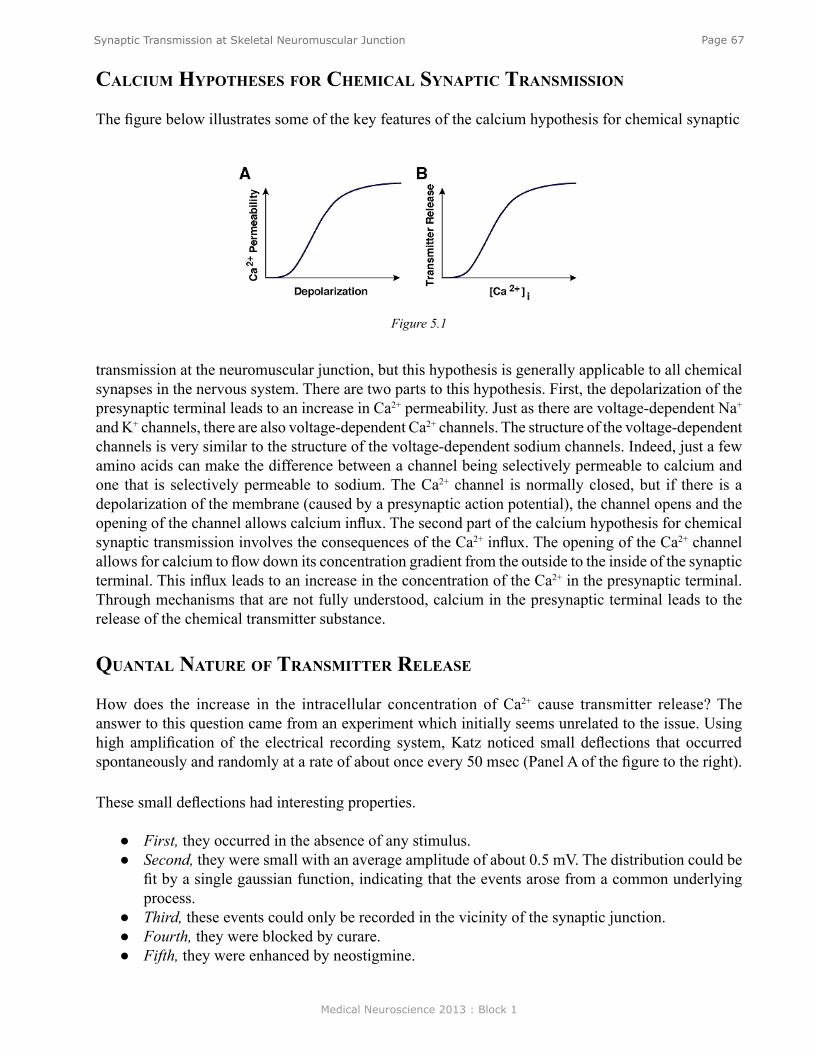
Medical Neuroscience 2013 : Block 1
Page 67Synaptic Transmission at Skeletal Neuromuscular Junction
Figure 5.1
CalCium hyPoTheses for ChemiCal synaPTiC Transmission
The figure below illustrates some of the key features of the calcium hypothesis for chemical synaptic
transmission at the neuromuscular junction, but this hypothesis is generally applicable to all chemical synapses in the nervous system. There are two parts to this hypothesis. First, the depolarization of the presynaptic terminal leads to an increase in Ca2+ permeability. Just as there are voltage-dependent Na+ and K+ channels, there are also voltage-dependent Ca2+ channels. The structure of the voltage-dependent channels is very similar to the structure of the voltage-dependent sodium channels. Indeed, just a few amino acids can make the difference between a channel being selectively permeable to calcium and one that is selectively permeable to sodium. The Ca2+ channel is normally closed, but if there is a depolarization of the membrane (caused by a presynaptic action potential), the channel opens and the opening of the channel allows calcium influx. The second part of the calcium hypothesis for chemical synaptic transmission involves the consequences of the Ca2+ influx. The opening of the Ca2+ channel allows for calcium to flow down its concentration gradient from the outside to the inside of the synaptic terminal. This influx leads to an increase in the concentration of the Ca2+ in the presynaptic terminal. Through mechanisms that are not fully understood, calcium in the presynaptic terminal leads to the release of the chemical transmitter substance.
quanTal naTure of TransmiTTer release
How does the increase in the intracellular concentration of Ca2+ cause transmitter release? The answer to this question came from an experiment which initially seems unrelated to the issue. Using high amplification of the electrical recording system, Katz noticed small deflections that occurred spontaneously and randomly at a rate of about once every 50 msec (Panel A of the figure to the right).
These small deflections had interesting properties.
● First, they occurred in the absence of any stimulus. ● Second, they were small with an average amplitude of about 0.5 mV. The distribution could be
fit by a single gaussian function, indicating that the events arose from a common underlying process.
● Third, these events could only be recorded in the vicinity of the synaptic junction. ● Fourth, they were blocked by curare. ● Fifth, they were enhanced by neostigmine.

Medical Neuroscience 2013 : Block 1
Page 68Synaptic Transmission at Skeletal Neuromuscular Junction
Based on these considerations, Katz called these events miniature endplate potentials or MEPPs. They appeared very similar to endplate potentials, but they were only about 0.5 mV in amplitude compared to the 50 mV amplitude of the normal EPP. Katz suggested that MEPPs were due to the spontaneous and random release of ACh. This idea intuitively makes good sense. If there is an abundance of ACh in the presynaptic terminal, perhaps some will leak out and diffuse across the cleft, bind to ACh receptors, and produce a small potential change. ACh is likely to be spontaneously released occasionally because there is a basal level of calcium in the presynaptic terminal. Each vesicle actually contains enough transmitter to open about 1,000 individual ACh-sensitive channels. (This information is necessary to answer "Test Your Knowledge" Question #5.)
(The designation MEPP has a very specific meaning. It refers to those small endplate potentials that occur randomly in the absence of any stimulation. For example, small endplate potentials (EPPs) can be recorded in the presence of curare or low levels of extracellular Ca2+ , but they are not MEPPs.)
Katz suggested, as a result of the experiment illustrated in Figure 5.2, that the normal EPP is due to the summation effects of many vesicles being released at the same time. One vesicle produces a potential of about 0.5 mV. The release of 100 of those vesicles at the same time could produce a potential which is 100 times as great (50 mV). The illustration below (Figure 5.3) shows one of these vesicles in the process of fusing with the membrane and releasing its contents into the synaptic cleft through a process called exocytosis. For illustrative purposes, each synaptic vesicle is shown to contain three molecules of transmitter. In reality, each vesicle contains about 10,000 molecules of transmitter. Vesicles ready to be released are found in a region near the presynaptic terminal membrane called the readily releasable pool. Newly synthesized vesicles are found in the storage or reserve pool. The process by which a vesicle migrates from the reserve pool to the readily releasable pool is called mobilization. After fusing with the membrane and releasing its contents, the membrane is recycled to form new synaptic vesicles. This process is called recycling. Additional details of this process are found in Chapter 10 of Neuroscience Online. Figure 5.4, below, illustrates a summary of the steps involved in the process of synaptic transmission at the neuromuscular junction.
Figure 5.2
Figure 5.3. See Neuroscience Online for animation.
Figure 5.4. See Neuroscience Online for animation.

Medical Neuroscience 2013 : Block 1
Page 69Synaptic Transmission at Skeletal Neuromuscular Junction
Test Your Knowledge(see Neuroscience Online for answers to questions)
5. In a normal, healthy individual the number of ACh-sensitive channels in a skeletal muscle cell opened as a result of a single action potential in a motor axon would be approximately.
A. 1B. 100C. 10,000D. 100,000E. 10,000,000
6. An isolated nerve muscle preparation treated with a moderate dose of curare would be associated with a decrease in the (Note: There is more than one correct answer.)
A. Frequency of MEPPsB. Amplitude of MEPPsC. Release of transmitter from the terminal of the motor axon
triggered by a presynaptic action potentialD. Amplitude of the EPP
7. An isolated nerve muscle preparation exposed to a low concentration of extracellular Ca2+ would be associated with a decrease in the (Note: There is more than one correct answer.)
A. Amplitude of the EPPB. Release of transmitter from the terminal of the motor axon
triggered by a presynaptic action potentialC. Influx of Ca2+ into the presynaptic terminalD. Amplitude of MEPPs

Medical Neuroscience 2013 : Block 1
Page 70Synaptic Transmission in the CNS / Synaptic Plasticity
Synaptic Transmission in the Central Nervous System/Synaptic Plasticity
assignmenTI. Postsynaptic potentials
A. Excitatory postsynaptic potentials (EPSPs)1. Ionic mechanisms2. Temporal summation3. Spatial summation
B. Inhibitory postsynaptic potentials (IPSPs)1. General features2. Ionic mechanisms3. Role of glycine in mediating IPSPs in the spinal cord
C. IntegrationD. Decreased conductance and slow synaptic potentials
1. Features2. Ionic mechanisms3. Role of second messengers
II. Modulation of synaptic efficacy (synaptic plasticity)A. Intrinsic regulation
1. Depression2. Facilitation3. Post-tetanic potentiation
B. Extrinsic regulation1. Morphological basis2. Presynaptic inhibition3. Presynaptic facilitation
major objeCTives1. Be able to compare and contrast features of synaptic transmission at the neuromuscular
junction with those at synapses in the CNS and spinal cord.2. Understand the differences between EPSPs and IPSPs.3. Know the principles of synaptic integration.4. Be able to compare and contrast the different types of synaptic plasticity and their
underlying mechanisms.
Lecturer: John H. Byrne, Ph.D.January 14 2013 | 9:00 AM and January 16, 2013 | 8:00 AM

Medical Neuroscience 2013 : Block 1
Page 71Synaptic Transmission in the CNS / Synaptic Plasticity
required reading ● Byrne, J.H. Synaptic Transmission in the Central Nervous System/Synaptic Plasticity.
Neuroscience Online: http://nba.uth.tmc.edu/neuroscience/s1/chapter06.html and http://nba.uth.tmc.edu/neuroscience/s1/chapter07.html; or
● Byrne, J.H. Synaptic Transmission in the Central Nervous System/Synaptic Plasticity, Lecture Supplement Volume I.
suggesTed reading ● Byrne, J. H., Roberts, J. L., From Molecules to Networks: An Introduction to Cellular and
Molecular Neuroscience, 2nd Edition, Elsevier, Amsterdam, 2009, Chapters 8, 9, 11, 12, 16, 17, and 19.
synaPTiC Transmission in a simPle reflex CirCuiT
One of the simplest behaviors mediated by the central nervous system is knee-jerk or stretch reflex. In response to a neurologist’s hammer to the patella tendon, there is a reflex extension of the leg. This figure illustrates the neurocircuitry that controls that reflex response. The stretch to the patella tendon stretches the extensor muscle. More specifically, it stretches a group of specific receptors known as muscle spindle receptors or simply stretch receptors. The stretch elicits action potentials in the stretch
Figure 6.1See Neuroscience Online for animation.
receptors which then propagate over type 1A afferent fibers, the somata of which are located in the dorsal root ganglion. Processes of these sensory neurons then enter the spinal cord and make synaptic connections with two types of cells. First, a synaptic connection is formed with the extensor motor neuron located in the ventral horn of the spinal cord. As the result of synaptic activation of this motor neuron, action potentials are elicited in the motor neuron and propagate out the ventral roots, ultimately invading the terminal regions of the motor axon (i.e., the neuromuscular junction, causing release of acetylcholine, depolarization of the muscle cell, formation of an action potential in the muscle cell, and a subsequent contraction of the muscle.
The sensory neurons also make synaptic connections with another type of neuron in the spinal cord called an interneuron. Interneurons are so named because they are interposed between one type of neuron and another. The particular interneuron shown is an inhibitory interneuron. As a result of its activation through the process of synaptic transmission, action potentials are elicited in the interneuron. An action potential in the inhibitory neuron leads to the release of a chemical transmitter substance that inhibits the flexor motor neuron, thereby preventing an improper movement from occurring. This particular reflex is known as the monosynaptic stretch reflex because this reflex is mediated by a single excitatory synaptic relay in the central nervous system.

Medical Neuroscience 2013 : Block 1
Page 72Synaptic Transmission in the CNS / Synaptic Plasticity
ioniC meChanisms of iPsPs
synaPTiC PoTenTials
Figure 6.2 illustrates how it is possible to experimentally examine some of the components of synaptic transmission in the reflex pathway that mediates the stretch reflex. Normally, the sensory neuron is activated by a stretch to the stretch receptor, but this process can be bypassed by injecting a depolarizing current into the sensory neuron. That stimulus initiates an action potential in the sensory neuron which
Figure 6.2
leads to a change in the potential of the motor neuron. This potential is known as an excitatory postsynaptic potential (EPSP); excitatory because it tends to depolarize the cell, thereby tending to increase the probability of firing an action potential in the motor neuron and postsynaptic because it is a potential recorded on the postsynaptic side of the synapse.
The ionic mechanisms for the EPSP in the spinal motor neuron are essentially identical to the ionic mechanisms for the EPSP at the neuromuscular junction. Specifically, the transmitter substance diffuses across the synaptic cleft and binds to specific ionotropic receptors on the postsynaptic membrane, leading to a simultaneous increase in the sodium and potassium permeability (See Figure 4.10). The mechanisms for release are also identical to those at the neuromuscular junction. An action potential in the presynaptic terminal leads to the opening of voltage dependent Ca2+ channels, and the Ca2+ influx causes transmitter substance to be released. differenCes beTween The ePsP aT The skeleTal neuromusCular junCTion and ePsPs in The Cns
There are two fundamental differences between the process of synaptic transmission at the sensorimotor synapse in the spinal cord and the process of synaptic transmission at the neuromuscular junction. First, transmitter substance released by the sensory neuron is not ACh but rather the amino acid glutamate. Indeed, there are many different transmitters in the central nervous system - up to 50 or more and the list grows every year. Fortunately, these 50 or more transmitter substances can be conveniently grouped into four basic categories: acetylcholine, monoamines, peptides, and the amino acids. Second, in contrast to the 50-mV amplitude of the synaptic potential at the neuromuscular junction, the amplitude of the synaptic potential in a spinal motor neuron, as a result of an action potential in a 1A afferent fiber, is only about 1 mV.

Medical Neuroscience 2013 : Block 1
Page 73Synaptic Transmission in the CNS / Synaptic Plasticity
TemPoral and sPaTial summaTion
If the amplitude of the postsynaptic potential is only 1 mV, how can an action potential in the motor neuron be triggered and the reflex function? Note that a 1-mV EPSP is unlikely to be sufficient to drive the membrane potential of the motor neuron to threshold to fire a spike. If there is no spike, there will be no contraction of the muscle. The answer is that the stretch of the muscle fires multiple action potentials in many different stretch receptors. In fact, the greater the stretch, the greater is the probability of activating more stretch receptors. This process is referred to as recruitment. Therefore, multiple 1A afferents will converge onto the spinal motor neuron and participate in its activation. This is not the whole answer, however. Recall that the greater the intensity of the stimulus, the greater is the number of action potentials elicited in a sensory receptor. The greater the stretch, the greater the number of action potentials elicited in a single sensory neuron and the greater number of EPSPs produced in the motor neuron from that train of action potentials in the sensory cell. The processes by which the multiple EPSPs from presynaptic neurons summate over space and time are called temporal and spatial summation.
Figure 6.3
Temporal summationA single action potential in sensory neuron 1 produces a 1-mV EPSP in the motor neuron. Now consider the consequences of firing two action potentials in quick succession (See figure above). Two EPPs are elicited, the second of which summates on the falling edge of the first. As a result of two action potentials, a summated potential about 2 mV in amplitude occurs. If there were three presynaptic action potentials, and they occurred rapidly enough, the total potential would be about 3 mV, and so forth. Temporal summation is strictly a passive property of nerve cells. Special ionic conductive mechanisms are not needed to explain it. The potentials summate because of the passive properties of the nerve cell membrane, specifically the ability of membranes to store charge. The membrane temporarily stores the charge of the first PSP and then the charge from the second PSP is added to it to produce a potential twice as large at first. This process of temporal summation is very much dependent upon the duration of the synaptic potential. The temporal summation occurs when the presynaptic action potentials occur in quick succession. The time frame is dependent upon the passive properties of the membrane, specifically the time constant.
Spatial summationNow consider a motor neuron that receives two inputs. An action potential produced in sensory neuron 1 produces a 1-mV EPSP and a single action potential in sensory neuron 2 also produces a 1-mV EPSP. If action potentials are produced simultaneously in sensory neuron 1 and in sensory neuron 2, the EPSPs summate to produce a summated EPSP which is twice that of the individual EPSPs. Spatial summation

Medical Neuroscience 2013 : Block 1
Page 74Synaptic Transmission in the CNS / Synaptic Plasticity
in nerve cells occurs because of the space constant, the ability of a charge produced in one region of the cell to spread to other regions of the cell.
iPsPs
Whether a neuron fires in response to a synaptic input is dependent upon how many action potentials are being fired in any one afferent input, as well as how many individual afferent pathways are activated.
The decision to fire also depends on the presence of inhibitory synaptic inputs. Artificially depolarizing the interneuron to initiate an action potential produces a transient hyperpolarization of the membrane potential of the motor neuron (See Figure 6.2). The time course of this hyperpolarization looks very similar to that of an EPSP, but it is reversed in sign. The synaptic potential in the motor neuron is called an inhibitory postsynaptic potential (IPSP) because it tends to move the membrane potential away from the threshold, thereby decreasing the probability of this neuron initiating an action potential.
ioniC meChanisms for iPsPs
The membrane potential of the flexor motor neuron is about -65 mV, so one might predict that the IPSP would be due to an increase in the permeability or the conductance of an ion whose equilibrium potential is more negative than -65 mV. One possibility is potassium. Potassium does mediate some inhibitory synaptic potentials in the central nervous system, but not at the particular synapse between a spinal interneuron and spinal motor neuron. At this particular synapse, the IPSP is due to a selective increase in chloride permeability. Note that the equilibrium potential for chloride is about -70 mV. The transmitter released by the spinal interneuron binds to a special class of ionotropic receptors which are normally closed, but open and become selectively permeable to chloride ions as a result of the binding of the transmitter. As a result of the increase in Cl- permeability, the membrane potential moves from its resting value of -65 mV towards the Cl- equilibrium potential. (Note that in principle, decreasing the resting conductance of Na+ could also produce an IPSP.)
TransmiTTer subsTanCe of The sPinal inhibiTory neuron
What about the transmitter substance that is released by the inhibitory interneuron in the spinal cord? The transmitter substance is glycine, an amino acid which is used frequently in the central nervous system as a transmitter that produces inhibitory actions. It is not the most common, however. The most common transmitter with inhibitory actions is gamma amino butyric acid (GABA).
meTaboTroPiC synaPTiC resPonses
In addition to the responses mediated by ionotropic receptors, there is an entirely separate class of synaptic potentials that have durations with orders of magnitude greater than the durations of the classical EPSPs. These are so-called slow synaptic potentials and they are mediated by metabotropic receptors. Slow synaptic potentials are not observed at every postsynaptic neuron but they are certainly observed at many. The figure below illustrates a postsynaptic neuron which receives two inputs. An action potential in neuron 1 produces an excitatory postsynaptic potential or EPSP in the postsynaptic cell whose duration is about 20 msec. Neuron 2 can also produce a postsynaptic potential but its duration is more than three orders of magnitude longer than that of the conventional type of synaptic

Medical Neuroscience 2013 : Block 1
Page 75Synaptic Transmission in the CNS / Synaptic Plasticity
potential. The mechanism of these slow synaptic responses involves changes in metabolism of the cell.
One mechanism for a slow synaptic potential is shown in the illustration above (Figure 6.5) and in Figure 12.11. In contrast to the ionotropic receptor for which the receptors are actually part of the channel complex, the channels that produce the slow synaptic potentials are not directly coupled to the transmitter receptors. Rather, the receptors are separate from the channel. These receptors are known as metabotropic because they involve changes in the metabolism of the cell and, in general, changes in activation of specific second messenger systems. The figure above illustrates an example of one type of response that involves the cyclic AMP cascade. Slow PSPs are in some cases mediated by cyclic AMP but they are also mediated by other protein kinases. For the response in Figure 6.5, the transmitter activates G proteins that lead to the increased synthesis of cyclic AMP. Cyclic AMP then leads to the activation of cyclic AMP-dependent kinase (PKA), which phosphorylates a channel protein or a component of the channel and then produces a conformational change in the channel and a change in its ionic permeability. In contrast to a direct conformational change produced by the binding of a transmitter to the receptor channel complex (seen in responses mediated by ionotropic receptors), the conformational change is produced by phosphorylation. The particular channel is one that is selectively permeable to K+ and is normally open. As a result of the channel phosphorylation by PKA, the channel closes and becomes less permeable to K+. Since the normal resting potential is due to a balance of Na+ and K+, decreasing the K+ conductance favors the effects of the Na+ conductance and a depolarization is produced.
Figure 6.4
Figure 6.5

Medical Neuroscience 2013 : Block 1
Page 76Synaptic Transmission in the CNS / Synaptic Plasticity
It is interesting to point out that the activation of metabotropic receptors can produce effects which are much longer than several hundred seconds. For example, protein kinase A can diffuse in the nucleus where it can phosphorylate proteins (i.e., transcription factors) that regulate gene expression.
neuroToxins
The discovery of certain toxins has greatly facilitated the analysis of voltage and chemically gated channels as well as the process of synaptic transmission. The following table illustrates some that have been particularly useful.
synaPTiC PlasTiCiTy
Historically, it was generally thought that the role of the synapse was to simply transfer information between one neuron and another neuron or between a neuron and a muscle cell. In addition, it was thought that these connections, once established during development, were relatively fixed in their strength, much like a solder joint between two electronic components. One exciting development in neurobiology over the past forty years is the realization that most synapses are extremely plastic; they are able to change their strength as a result of either their own activity or through activity in another pathway. Many think that this synaptic plasticity is central to understanding the mechanisms of learning and memory.
Some Important Neurotoxinstetrodotoxin (TTX): Fish toxin that blocks the pore of voltage-dependent Na+ channels.
µ-conotoxin (µ-CTX): Fish-hunting cone snail toxin with properties similar to TTX.
saxitoxin (STX): Toxin from marine dinoflagellates with properties similar to TTX. STX is also known as paralytic shellfish poison.
ω-conotoxin (ω-CTX): Fish-hunting cone snail toxin that blocks certain types of voltage-dependent Ca2+ channels.
funnel web spider toxin(ω-Aga): Toxin from funnel web spider which blocks certain types of voltage-dependent Ca2+ channels.
apamin: Bee venom toxin that blocks certain types of Ca2+-activated K+ channels.
charybdotoxin (ChTX): Scorpion venom toxin that blocks pore of some Ca2+-activated K+ channels and voltage-dependent K+ channels.
curare (d-tubocuraine): Plant toxin that is a competitive inhibitor of nicotinic ACh receptors.
α-bungarotoxin: Snake toxin that is competitive and highly irreversible inhibitor of nicotinic ACh receptors.
picrotoxin: GABAA receptor blocker isolated from the seed of Anamirta cocculus.
strychnine: Glycine receptor blocker isolated from the seed of the East Indian tree Strychnos nux-vomica.
tetanus toxin: Clostridial neurotoxin with zinc-dependent protease activity; Cleaves synaptic vesicle proteins in the CNS and thereby blocks release of neurotransmitters.
botulinum toxin: Clostridial neurotoxin with zinc-dependent protease activity; Cleaves synaptic vesicle proteins at the neuromuscular junction and thereby blocks release of ACh.

Medical Neuroscience 2013 : Block 1
Page 77Synaptic Transmission in the CNS / Synaptic Plasticity
There are two general forms of synaptic plasticity, intrinsic and extrinsic. Intrinsic mechanisms, also known as homosynaptic mechanisms, refer to changes in the strength of a synapse that are brought about by its own activity. (Homo from the Greek meaning the same.) Extrinsic plasticity, or heterosynaptic plasticity, is a change in the strength of a synapse brought about by activity in another pathway.
Homosynaptic PlasticityThere are two types of intrinsic or homosynaptic plasticity, synaptic depression and synaptic facilitation. Synaptic depression and facilitation are not always found at the same synapse. Some synapses exhibit one but not the other, whereas some synapses exhibit both. Figure 7.1B illustrates homosynaptic plasticity
Figure 7.1
at the synapse between a 1A afferent fiber and a spinal motor neuron. An action potential in the sensory neuron produces an EPSP in the motor neuron. A second action potential in the sensory neuron, 200 msec after the first, produces an EPSP that is smaller than that produced by the first action potential. This phenomenon is called synaptic depression. The efficacy of synaptic transmission is not constant; it varies depending upon the frequency of stimulation. The mechanisms of synaptic depression vary but one common mechanism is depletion of the available transmitter. The second of two action potentials will release less transmitter because less transmitter is available to be released. (See Figure 7.2A)
The figure at bottom illustrates the second form of homosynaptic plasticity, synaptic facilitation. This particular example is known as paired-pulse or twin-pulse facilitation. Two action potentials in the presynaptic cell produce two EPSPs in the postsynaptic cell. The first action potential produces a 1 mV EPSP, but the second action potential, which occurs about 20 msec after the first, produces an EPSP that is larger than the EPSP produced by the first. In this example, it is twice as large as the first one. This doubling of the EPSP represents the synaptic facilitation. The net EPSP is 3 mV. Through the process of temporal summation the second EPSP (2mV) adds to the amplitude of the first EPSP (1mV).
One mechanism contributing to twin pulse facilitation is residual calcium. An action potential leads to the opening of Ca2+ channels and the influx of Ca2+, which leads to the release of transmitter. Now consider the fate of the calcium after the first action potential (Figure 7.2B). Ca2+ levels will decline back to their initial level, but this recovery will not occur instantaneously. Thus, if a second action potential is initiated at a time during which the calcium has not yet recovered to its basal level, the calcium influx associated with the second spike will add to the “residual calcium” that is left over from the first.
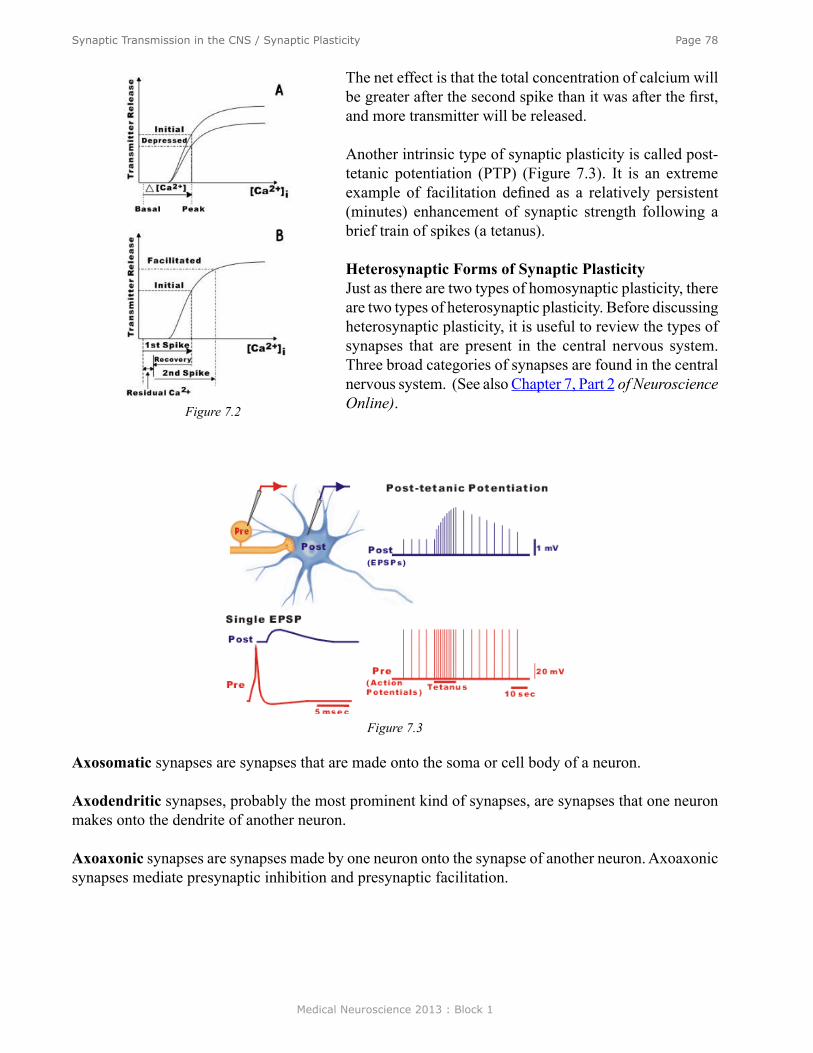
Medical Neuroscience 2013 : Block 1
Page 78Synaptic Transmission in the CNS / Synaptic Plasticity
The net effect is that the total concentration of calcium will be greater after the second spike than it was after the first, and more transmitter will be released.
Another intrinsic type of synaptic plasticity is called post-tetanic potentiation (PTP) (Figure 7.3). It is an extreme example of facilitation defined as a relatively persistent (minutes) enhancement of synaptic strength following a brief train of spikes (a tetanus).
Heterosynaptic Forms of Synaptic PlasticityJust as there are two types of homosynaptic plasticity, there are two types of heterosynaptic plasticity. Before discussing heterosynaptic plasticity, it is useful to review the types of synapses that are present in the central nervous system. Three broad categories of synapses are found in the central nervous system. (See also Chapter 7, Part 2 of Neuroscience Online). Figure 7.2
Figure 7.3
Axosomatic synapses are synapses that are made onto the soma or cell body of a neuron.
Axodendritic synapses, probably the most prominent kind of synapses, are synapses that one neuron makes onto the dendrite of another neuron.
Axoaxonic synapses are synapses made by one neuron onto the synapse of another neuron. Axoaxonic synapses mediate presynaptic inhibition and presynaptic facilitation.

Medical Neuroscience 2013 : Block 1
Page 79Synaptic Transmission in the CNS / Synaptic Plasticity
Figure 7.5 illustrates the two major types of heterosynaptic plasticity; presynaptic inhibition and presynaptic facilitation. Presynaptic inhibition is not an esoteric phenomenon. It is very prominent in the spinal cord and regulates the propagation of information to higher brain centers. An action potential in the presynaptic cell produces an EPSP in the postsynaptic cell. The modulatory cell (M1) makes an axoaxonic synapse with the presynaptic cell. After firing cell M1, the EPSP in the postsynaptic cell is smaller. This phenomenon is called presynaptic inhibition, because cell M1 regulates the ability of the presynaptic cell to release transmitter. The modulatory transmitter engages metabotropic-type receptors that activate a second messenger system that phosphorylates Ca2+ channels in such a way that the Ca2+ channels open less readily. Fewer Ca2+ channels are opened with a subsequent action potential in Pre and therefore the Ca2+ influx (ICa) will be reduced. Less Ca2+ influx leads to less transmitter release and a smaller EPSP.
The phenomenon complementary to presynaptic inhibition is presynaptic facilitation. M1 is capable of increasing the strength of the synaptic pathway. As a result of the activation of a second messenger cascade by M1, an action potential in the presynaptic terminal leads to a greater amount of Ca2+ influx, and therefore more transmitter is released.
Figure 7.4
Figure 7.5
Long-Term Potentiation (LTP)A very enduring form of synaptic plasticity is called long-term potentiation (LTP). It can have both homosynaptic and heterosynaptic components. An electric shock (trest stimulus) to afferent fibers produces an EPSP (Figure 7.6). If the pathway is repeatedly stimulated (e.g., every minute), the amplitude of EPSP is constant. A tetanus produces post-tetanic potentiation (PTP) that dies away after several minutes. What is left is a very enduring enhancement of the EPSP. There is excitement about LTP because it is the kind of mechanism necessary to store memory (Figure 7.7).
The NMDA-type receptor is critical for some forms of LTP, in particular LTP at the CA3-CA1 synapse in the hippocampus. The postsynaptic spines of CA1 neurons have two types of glutamate receptors; NMDA-type glutamate receptors and the AMPA-type glutamate receptors (Figures 7.8 and 7.9). Both receptors are permeable to Na+ and K+, but the NMDA-type has two additional features.
● First, in addition to being permeable to Na+, it also has a significant permeability to Ca2+. ● Second, this channel is normally blocked by Mg2+.
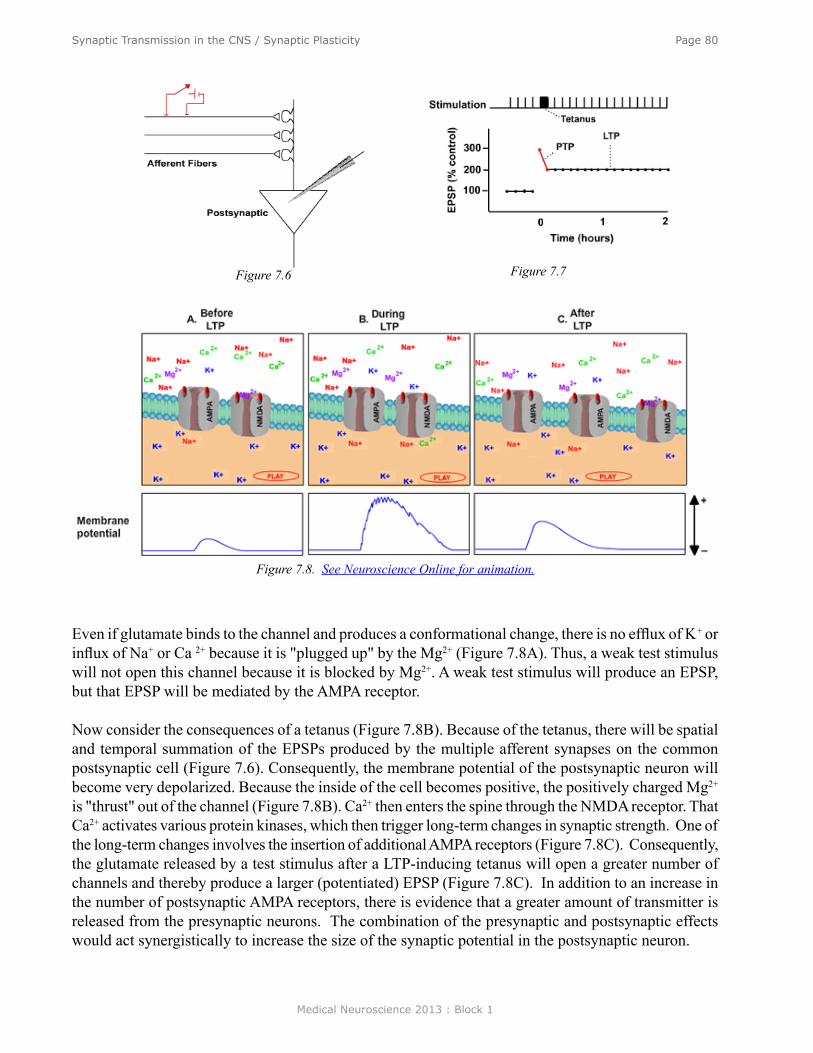
Medical Neuroscience 2013 : Block 1
Page 80Synaptic Transmission in the CNS / Synaptic Plasticity
Figure 7.6 Figure 7.7
Even if glutamate binds to the channel and produces a conformational change, there is no efflux of K+ or influx of Na+ or Ca 2+ because it is "plugged up" by the Mg2+ (Figure 7.8A). Thus, a weak test stimulus will not open this channel because it is blocked by Mg2+. A weak test stimulus will produce an EPSP, but that EPSP will be mediated by the AMPA receptor.
Now consider the consequences of a tetanus (Figure 7.8B). Because of the tetanus, there will be spatial and temporal summation of the EPSPs produced by the multiple afferent synapses on the common postsynaptic cell (Figure 7.6). Consequently, the membrane potential of the postsynaptic neuron will become very depolarized. Because the inside of the cell becomes positive, the positively charged Mg2+ is "thrust" out of the channel (Figure 7.8B). Ca2+ then enters the spine through the NMDA receptor. That Ca2+ activates various protein kinases, which then trigger long-term changes in synaptic strength. One of the long-term changes involves the insertion of additional AMPA receptors (Figure 7.8C). Consequently, the glutamate released by a test stimulus after a LTP-inducing tetanus will open a greater number of channels and thereby produce a larger (potentiated) EPSP (Figure 7.8C). In addition to an increase in the number of postsynaptic AMPA receptors, there is evidence that a greater amount of transmitter is released from the presynaptic neurons. The combination of the presynaptic and postsynaptic effects would act synergistically to increase the size of the synaptic potential in the postsynaptic neuron.
Figure 7.8. See Neuroscience Online for animation.

Medical Neuroscience 2013 : Block 1
Page 81Synaptic Transmission in the CNS / Synaptic Plasticity
summary
A given postsynaptic neuron receives synaptic input from a number of different sources (Figure 7.9). There are the traditional types of axosomatic and axodendritic synapses. These can be either excitatory or inhibitory. In addition, the synaptic responses can be mediated by both ionotropic and metabotropic receptors. The presynaptic cells can be modulated through presynaptic inhibition and presynaptic facilitation.
Consider that any one postsynaptic cell makes and receives 10,000 connections with other cells and that this module can be recapitulated in each of the billions of cells in the nervous system. It is this enormous pattern of synaptic connections and the plasticity that occurs at each one of these synapses which makes the nervous system so extraordinary.
It is very difficult to overestimate the importance of synaptic transmission. It is critical to the basic functioning of the nervous system and appears to be critical in learning and memory. Also, changes in synaptic transmission seem to be central to understanding a number of neurological disorders such as myasthenia gravis and Parkinson’s disease. Synaptic transmission is central to understanding mental diseases such as schizophrenia, anxiety, and depression. A major theme of neuroscience is to identify the specific transmitter systems involved in these brain diseases and design appropriate interventions. Finally, most of the psychoactive drugs function by affecting some aspects of synaptic transmission.
Figure 7.9

Medical Neuroscience 2013 : Block 1
Page 82Transport and Molecular Mechanisms of Secretion
Transport and Molecular Mechanisms of Secretion
assignmenT
I. Synthesis of vesicles and protein II. Transport of vesicles
III. Secretory mechanismIV. Vesicle recyclingV. Retrograde axoplasmic transport
major objeCTives
1. Understand the elements and basic properties of neurotransmitter.2. Have a basic knowledge of the principles underlying exocytosis.3. Know how axoplasmic transport functions and why it is important.
required reading
● Waymire, J.C. Notes on Synaptic Vesicles, Synthesis, Transport, Exocytosis And Retrieval, Lecture Supplement Volume I.
suggesTed reading (noT required)
● Waymire, J.C. Transport and the Molecular Mechanism of Secretion, Neuroscience Online, http://nba.uth.tmc.edu/neuroscience/s1/chapter10.html
Lecturer: Jack C. Waymire, Ph.D.January 16, 2013 | 9:00 AM
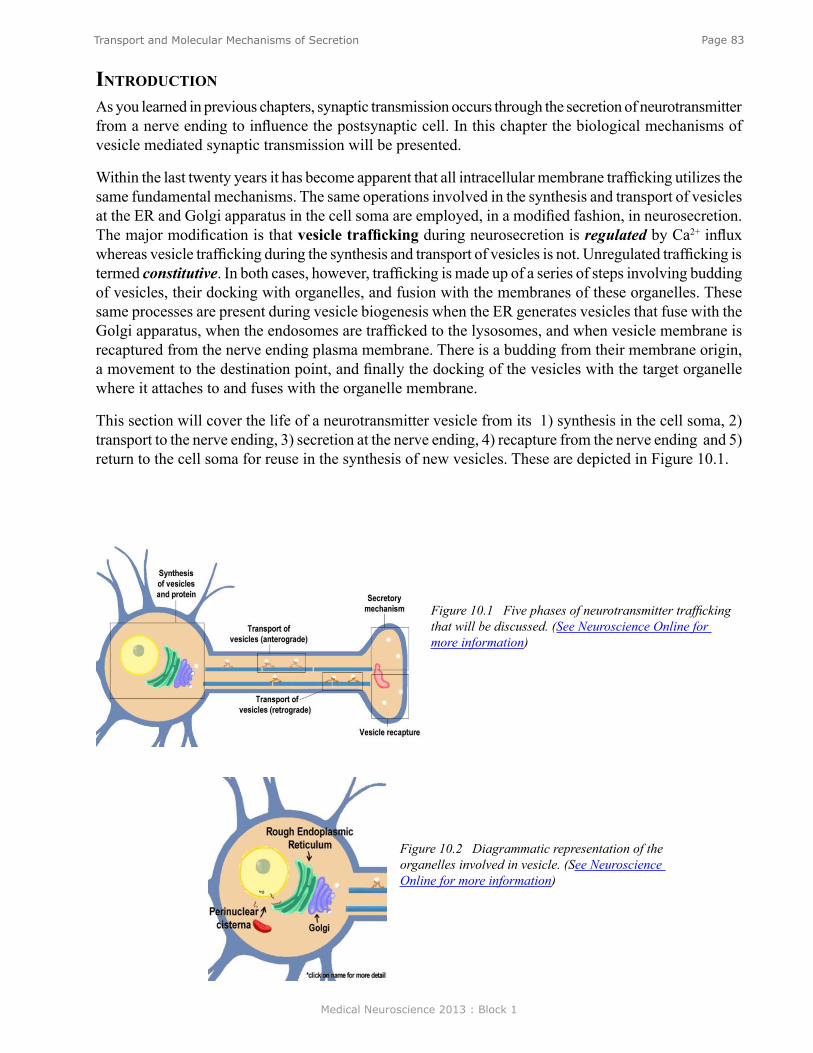
Medical Neuroscience 2013 : Block 1
Page 83Transport and Molecular Mechanisms of Secretion
inTroduCTion
As you learned in previous chapters, synaptic transmission occurs through the secretion of neurotransmitter from a nerve ending to influence the postsynaptic cell. In this chapter the biological mechanisms of vesicle mediated synaptic transmission will be presented.
Within the last twenty years it has become apparent that all intracellular membrane trafficking utilizes the same fundamental mechanisms. The same operations involved in the synthesis and transport of vesicles at the ER and Golgi apparatus in the cell soma are employed, in a modified fashion, in neurosecretion. The major modification is that vesicle trafficking during neurosecretion is regulated by Ca2+ influx whereas vesicle trafficking during the synthesis and transport of vesicles is not. Unregulated trafficking is termed constitutive. In both cases, however, trafficking is made up of a series of steps involving budding of vesicles, their docking with organelles, and fusion with the membranes of these organelles. These same processes are present during vesicle biogenesis when the ER generates vesicles that fuse with the Golgi apparatus, when the endosomes are trafficked to the lysosomes, and when vesicle membrane is recaptured from the nerve ending plasma membrane. There is a budding from their membrane origin, a movement to the destination point, and finally the docking of the vesicles with the target organelle where it attaches to and fuses with the organelle membrane.
This section will cover the life of a neurotransmitter vesicle from its 1) synthesis in the cell soma, 2) transport to the nerve ending, 3) secretion at the nerve ending, 4) recapture from the nerve ending and 5) return to the cell soma for reuse in the synthesis of new vesicles. These are depicted in Figure 10.1.
Figure 10.1 Five phases of neurotransmitter trafficking that will be discussed. (See Neuroscience Online for more information)
Figure 10.2 Diagrammatic representation of the organelles involved in vesicle. (See Neuroscience Online for more information)

Medical Neuroscience 2013 : Block 1
Page 84Transport and Molecular Mechanisms of Secretion
synThesis of vesiCles and ProTeins
The cycle of neurotransmitter vesicles begins in the ER where the vesicles proteins are synthesized. Vesicle biosynthesis continues as the proteins migrate through the smooth ER and the Golgi apparatus to eventually emerge to be transported to the nerve ending by axoplasmic transport.
PerinuClear CisTernae and ribosomal ProTein synThesis
The outer layer of the nuclear envelope is contiguous with the endoplasmic reticulum, which is in turn contiguous with the Golgi apparatus. As shown in Figure 10.3, after the mRNA (destined to make vesicle protein) is transcribed from the DNA in the nucleus, it migrates through pores (perinuclear cisternae label (A) in Figure 10.3). As the mRNA reaches the cytoplasm it encounters free ribosomes, labeled (B) where cytoplasmic protein synthesis occurs. Proteins that are destined to be associated with membrane structures, such as neurotransmitter vesicles or the plasmalemma are synthesized on ribosomes within the endoplasmic reticulum. Recall that the presence of ribosomes defines this part of the endoplasmic reticulum as rough endoplasmic reticulum.
Figure 10.3 Diagrammatic representation of the sites of cellular protein synthesis.
In (C) of Figure 10.3, the polypeptide (protein) being synthesized has no signal peptide. Consequently, this protein will become a non-membrane bound (soluble) protein that resides within lumen of vesicles. While in the lumen, the protein undergoes further processing as it proceeds through the smooth ER, Golgi apparatus, and the secretory vesicle as mature secretory proteins are formed.
In (D) of Figure 10.3, a signal peptide is a part of the polypeptide being synthesized (this is signified in the figure by the fact that the peptide is anchored in the ribosomal membrane). Consequently, this protein will be an integral membrane protein. As the ribosome moves along the mRNA template, amino acids are added. The signal peptide inserts in the membrane of the endoplasmic reticulum and maintains the protein's association with membrane. This will ensure that the protein will associate with a vesicular structure, such as a neurotransmitter storage vesicle. Figure 10.3 shows the protein synthesis from ribosomes in the rough ER to synthesize membrane bound protein.

Medical Neuroscience 2013 : Block 1
Page 85Transport and Molecular Mechanisms of Secretion
Figure 10.4 Diagram of the budding of vesicle from the SER and fusion with the Golgi Apparatus.
rough endoPlasmiC reTiCulum
Figure 10.4 illustrates the movement of the secretory vesicles through the rough and the smooth ER. The smooth ER extends from the RER and serves as a site for lipid biosynthesis for the production of endosomes, lysosomes and plasma membrane as well as for the neurotransmitter vesicles. New membrane protein that begins its synthesis in the RER continues in the SER where pieces of the SER bud off to form transport vesicles that shuttle to the Golgi apparatus with their contents.
golgi
As shown in Figure 10.5, in the Golgi apparatus the vesicles fuse to form the outermost of the Golgi apparatus cisternae stacks. Each cisterna migrates in a stepwise fashion through the Golgi apparatus toward the concave surface. During this migration, proteins become more concentrated and undergo various types of biochemical modification to produce mature functional proteins. These modifications include phosphorylation, glycosylation, proteolysis and addition of fatty acids, as well as others. The migration proceeds from the cis face close to the SER to the concave trans face. Upon reaching the concave face the cisternae round up into small vacuoles, then coalesce to form a larger condensing vacuole. The condensing vacuoles then give rise to a number of dense spherical transport vesicles. These vesicles bud off and are transported to the various destinations within the neuron where they become cell membrane, lysosomes, endosomes or neurotransmitter vesicles.
Figure 10.5 Diagram of the budding of vesicle from the Golgi Apparatus.

Medical Neuroscience 2013 : Block 1
Page 86Transport and Molecular Mechanisms of Secretion
All cellular constituents that are transported to the nerve terminal are done so through either fast transport, as described above, or a slower transport that proceeds at a speed approximately 100 x slower than fast transport. This too is mediated by a kinesin. This transport is probably slower because it has many stops and starts giving it a very jerky appearance when viewed in video enhanced microscopy. Many cytoskeletal elements are transported by this slower transport. There are two classes of slow anterograde transport: Component A which carries microtubules & neurofilaments at 0.1-1 mm per day, and component B which carries over 200 diverse proteins as well as actin at a rate of up to 6 mm a day.
seCreTory meChanism
As discussed in Chapter 5 of Neuroscience Online, neurotransmitter is secreted at the nerve ending through the Ca2+-dependent fusion of neurotransmitter storage vesicle with the plasma membrane with the neurotransmitter being secreted (released) into the synaptic cleft to influence the postsynaptic cell. This process is termed exocytosis. An important concept to emerge is that the same mechanisms that occur in vesicle fusion with membranes in the Golgi during vesicle biosynthesis are also employed at the nerve ending for neurotransmitter release. This is true for vesicle synthesis, even very simple cells like yeast. This conservation of mechanisms has permitted the use of simple systems to help understand the molecular mechanisms of neurotransmitter release.
The first event that must occur (with the exception of neuropeptide neurotransmitters) is the filling of vesicles with neurotransmitter through specific neurotransmitter uptake (NT Uptake). This uptake will be covered in subsequent chapters that discuss each of the specific neurotransmitters.
Figure 10.6 Anterograde axoplasmic transport mediated by kinesin association with microtubules.
anTerograde TransPorT of vesiCles
Vesicles formed in the cell soma are moved to the site at which they will be used in synaptic transmission. This step in the trafficking of the vesicle is mediated by a process termed fast anterograde axoplasmic transport. As depicted in Figure 10.6, transport is mediated through the interaction of the vesicle with the microtubule. Transport is an energy dependent process in which a motor protein, kinesin, associates with vesicles and moves down the microtubule in a series of attachment-detachment steps. Evidence for the existence of axoplasmic transport comes from a variety of observations, including the movement of radioactive proteins synthesized in the cell soma down the axon to the nerve endings. The speed of transport is 50 – 400 mm/ day. Ca2+ is also required for transport. The vinca alkaloid drugs colchicine and vinblastine prevent axoplasmic transport through their disruption of microtubules.

Medical Neuroscience 2013 : Block 1
Page 87Transport and Molecular Mechanisms of Secretion
The vesicles remain in reserve until needed for secretion. When needed for secretion, a translocation occurs, which is also referred to as mobilization. The vesicles move to a region of plasma membrane called the active zone. The active zone is the release site and is characterized by the appearance of dense material adjacent to the plasma membrane. The influx of Ca2+ is believed to increase translocation by increasing the Ca2+ dependent phosphorylation of a vesicle binding protein termed synapsin. Evidence supports the Ca2+ dependent phosphorylation of synapsin which frees the vesicles from binding to actin microfilaments. The vesicles then move to the active zone of the plasma membrane where they are in position to undergo release of their neurotransmitter.
The association of the vesicle with the plasma membrane is termed docking and serves to prime the vesicle for secretion. The docking occurs through the binding of proteins in the vesicle membrane to proteins in the plasma membrane. Several of these proteins have been discovered because they are targets for clostridia bacterial toxins. These toxins act by blocking synaptic transmission. Several of these toxins and the proteins they interact with are shown in Table I. The toxins are so toxic that a single molecule can poison a whole nerve terminal. VAMP is a synaptic vesicle protein and syntaxin and SNAP-25 are two of the synaptic plasma membrane proteins.
Table I
Toxin Synaptic protein Location
Botulinum toxins A & E SNAP-25 Synaptic plasma
membrane
Botulinum toxin C1 Syntaxins Synaptic plasma membrane
Botulinum toxin B, D, F & G & tetanus toxin VAMPs Synaptic vesicle
Figure 10.7 Diagrammatic representation of the interaction of vesicle and membrane proteins during synaptic vesicle docking, fusion and fission. (See Neuroscience Online for animation)
A third plasma membrane protein, n-sec-1, has a loose association with the plasma membrane and thereby prevent the interaction of the neurotransmitter vesicle proteins until n-sec-1 is displaced (the mechanism of n-sec-1 displacement is currently not understood). This and subsequent steps in the secretory process are shown in Figure 10.7. The vesicle and plasma membrane proteins are hypothesized to complex with one another upon the displacement of n-sec-1 to form a "trimeric complex" (SNAP-25, Syntaxin and VAMP). This three-member complex has been isolated, intact, from the nerve endings of animals. This association of the three proteins initiates fusion. Vesicles at this stage are primed for release.

Medical Neuroscience 2013 : Block 1
Page 88Transport and Molecular Mechanisms of Secretion
The final stage of release, also shown in Figure 10.7, is the fission of the membrane at the point of contact between the vesicle and the plasma membrane. Exocytosis of neurotransmitter into the synaptic cleft occurs when fission takes place. This step is initiated by Ca2+, but the mechanism of the Ca2+ trigger is unknown. One hypothesis is that the vesicle associated protein synaptotagmin binds Ca2+ to initiate fission. Support for synaptotagmin, as the Ca2+ sensor, is that it possesses two binding sites for Ca2+. Additional evidence comes from studies of mice in which synaptotagmin has been knocked out. In these mice fast Ca2+-triggered synaptic vesicle exocytosis is severely limited.
Many aspects of the fusion-fission mechanism remain to be understood, including: what causes the dissociation of n-sec-1 from the complex, how Ca2+ functions in the release process and how all the proteins that are involved in release become reconstituted into vesicle membrane following release as the vesicle membrane is recycled.
vesiCle reCaPTure
Although the original source of vesicles for neurotransmitter secretion comes from biosynthesis in the Golgi apparatus at the cell body and subsequent transport to the nerve ending, evidence indicates that local re-synthesis of synaptic vesicle at the nerve ending is responsible for the continual supply of vesicles for neurotransmission. Two mechanisms have been observed that are responsible for this resupply. Both utilize the recapture of vesicle membrane from the nerve ending. In one, vesicles bud off the plasma membrane through the formation of pits that migrate directly to become a functional neurotransmitter vesicle as soon as it can be refilled with neurotransmitter through the neurotransmitter uptake process. This mechanism is referred to as "kiss and run". The second mechanism involves the formation of clathrin coated pits to recapture vesicle membrane, and the vesicles then cycle through the endosomal compartment in the nerve ending to become functional synaptic vesicles. The vesicles bud off the endosome to form the neurotransmitter vesicle and are then filled with neurotransmitter. It is believed that both mechanisms can exist in the same nerve ending or only one of the two can be present. Both are important in that they recover vesicle protein to permit a plentiful supply for synaptic transmission. This mechanism also explains the lack of enlargement of the nerve ending that would occur if vesicle membrane were not recaptured. No matter which mechanism is involved, the supply and resupply of vesicles can only keep pace with a high rate of synaptic transmission for a few minutes.
reTrograde axoPlasmiC TransPorT
Eventually the proteins utilized for synaptic transmission in the nerve ending are targeted and returned to the cell body of the neuron to be recycled to make new protein and vesicles. The proteins are returned to the soma through a retrograde axoplasmic transport that is analogous to anterograde transport but uses a different motor protein, dynein. Transport is mediated by the interaction of dynein with microtubules, depicted in Figure 10.8, and proceeds at a rate (100-200 mm per day) similar to that of fast anterograde transport. In addition to returning proteins to the soma, retrograde transport serves as a means of communication between the nerve endings and the cell soma. This is a mechanism to transport signaling molecules to regulate the development and maintenance of axonal contacts with postsynaptic cells. In addition, several viruses (e.g. rabies and herpes) are transported to the CNS through this mechanism.

Medical Neuroscience 2013 : Block 1
Page 89Transport and Molecular Mechanisms of Secretion
Figure 10.8 Retrograde axoplasmic transport mediated by dynein association with microtubules
Figure 10.9 shows a summary of retrograde and anterograde axoplasmic transport. It shows the motor proteins, kinesin and dynein, mediating the movement of vesicles and mitochondria anterogradely and vesicles retrogradely along microtubules. The animation shows the motor proteins as a part of the organelle that is transported. The other possible relationship between the motor protein and microtubules is that the motor proteins are a part of the microtubule and pass the vesicles along the microtubule. (See Neuroscience Online for animation)
Test Your Knowledge(see Neuroscience Online for answer to question)
11. Which of the following processes dictate the amount of neurotransmitter released from a nerve ending on a short-term, minute-to-minute, basis? (Note: There is more than one correct answer.)
A. Neurotransmitter synthesisB. Vesicle synthesis in the cell somaC. Vesicle recycling in the nerve endingD. Axoplasmic transportE. Calcium availability

Medical Neuroscience 2013 : Block 1
Page 90Acetylcholine Neurotransmission
Acetylcholine Neurotransmission
assignmenT
I. Introduction and ACh structure II. Anatomical distribution
A. ANSB. PNSC. CNS
III. Cell biology and pharmacology of acetylcholineA. Synthesis of acetylcholineB. Storage of acetylcholineC. Release of acetylcholineD. Acetylcholine receptors
1. Nicotinic - ion channels2. Muscarinic - G protein linked
E. Acetylcholine inactivation by acetylcholine esteraseIV. Termination of ACh actionV. Physiology and behavior
VI. Clinical importanceA. Alzheimer’s diseaseB. Myasthenia gravis
VII. Pharmacology
major objeCTives
1. Know the biosynthesis, degradation, release, and function of acetylcholine. 2. Understand the basic differences between nicotinic and muscarinic cholinergic receptors.3. Know how cholinergic drugs are used.
required reading
● Waymire, J.C. Notes on Acetylcholine Neurotransmission, Lecture Supplement Volume I.
suggesTed reading (noT required)
● Waymire, J.C. Acetylcholine Neurotransmission, Neuroscience Online: http://nba.uth.tmc.edu/neuroscience/s1/chapter11.html
Lecturer: Jack C. Waymire, Ph.D.January 18, 2013 | 8:00 AM

Medical Neuroscience 2013 : Block 1
Page 91Acetylcholine Neurotransmission
inTroduCTion
Acetylcholine, the first neurotransmitter discovered, was originally described as “vagus stuff” by Otto Loewi because of its ability to mimic the electrical stimulation of the vagus nerve. It is now known to be a neurotransmitter at all autonomic ganglia, at many autonomically innervated organs, at the neuromuscular junction, and at many synapses in the CNS.
In this chapter we will discuss the acetylcholine anatomy, cell biology, physiological effects, role in behavior, and clinical applications.
aCeTylCholine in The auTonomiC nervous sysTem
In the autonomic nervous system, acetylcholine (ACh) is the neurotransmitter in the preganglionic sympathetic and parasympathetic neurons. These are shown in Figure 11.1 as the ACh in the ganglion. ACh is also the neurotransmitter at the adrenal medulla and serves as the neurotransmitter at all the parasympathetic innervated organs. ACh is also the neurotransmitter, at the sweat glands, and at the piloerector muscle of the sympathetic ANS.
Figure 11.1 Peripheral and autonomic sites where ACh is neurotransmitter.
aCh in The PeriPheral nervous sysTem
In the peripheral nervous system, ACh is the neurotransmitter at the neuromuscular junction between the motor nerve and skeletal muscle.
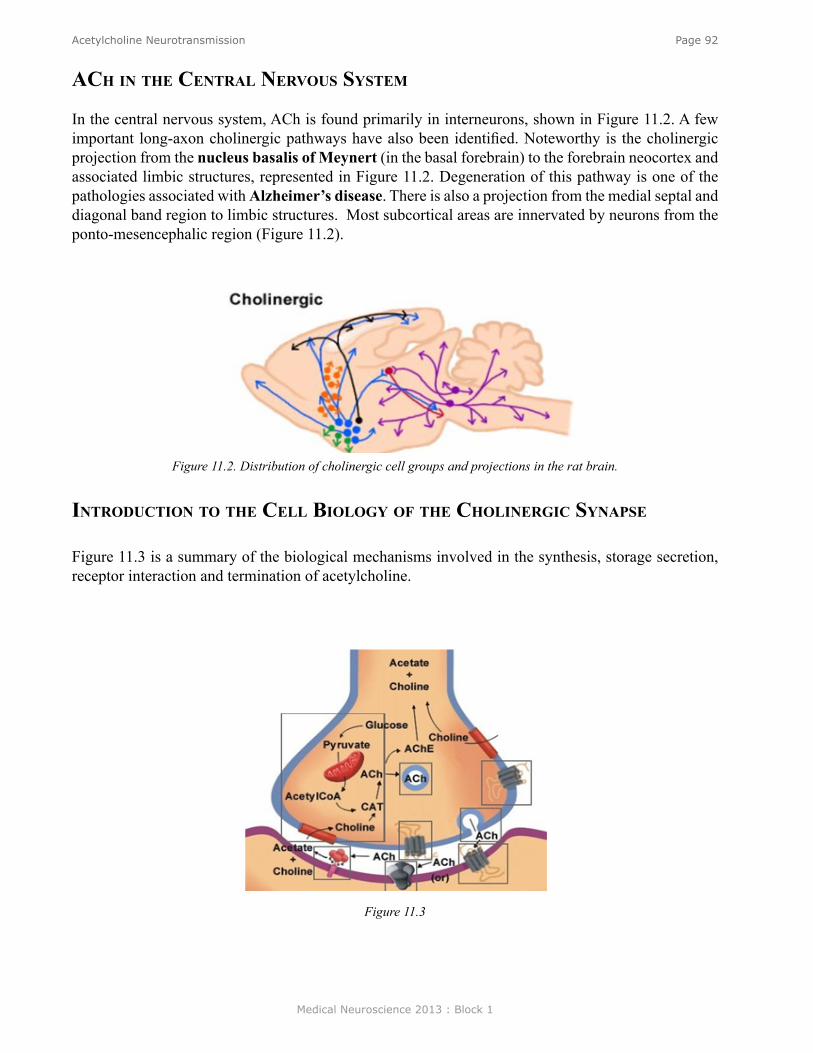
Medical Neuroscience 2013 : Block 1
Page 92Acetylcholine Neurotransmission
aCh in The CenTral nervous sysTem
In the central nervous system, ACh is found primarily in interneurons, shown in Figure 11.2. A few important long-axon cholinergic pathways have also been identified. Noteworthy is the cholinergic projection from the nucleus basalis of Meynert (in the basal forebrain) to the forebrain neocortex and associated limbic structures, represented in Figure 11.2. Degeneration of this pathway is one of the pathologies associated with Alzheimer’s disease. There is also a projection from the medial septal and diagonal band region to limbic structures. Most subcortical areas are innervated by neurons from the ponto-mesencephalic region (Figure 11.2).
Figure 11.2. Distribution of cholinergic cell groups and projections in the rat brain.
inTroduCTion To The Cell biology of The CholinergiC synaPse
Figure 11.3 is a summary of the biological mechanisms involved in the synthesis, storage secretion, receptor interaction and termination of acetylcholine.
Figure 11.3

Medical Neuroscience 2013 : Block 1
Page 93Acetylcholine Neurotransmission
synThesis of aCh
Choline acetyltransferase (CAT): As shown in Figure 11.4, ACh is synthesized by a single step reaction catalyzed by the biosynthetic enzyme choline acetyltransferase. As is the case for all nerve terminal proteins, CAT is produced in the cholinergic cell body and transported down the axon to the nerve endings. Both CAT and ACh may be found throughout the neuron, but their highest concentration is in axon terminals. The presence of CAT is the “marker” that a neuron is cholinergic, only cholinergic neurons contain CAT.
Figure 11.4. Diagram showing the role of acetyl-CoA from glucose metabolism and choline from the high affinity uptake in ACh biosynthesis.
The rate-limiting steps in ACh synthesis are the availability of choline and acetyl-CoA. During increased neuronal activity the availability of acetyl-CoA from the mitochondria is regulated as is the uptake of choline into the nerve ending from the synaptic cleft. Ca 2+ appears to be involved in both of these regulatory mechanisms. As will be described later, the inactivation of ACh is converted by metabolism to choline and acetic acid. Consequently much of the choline used for ACh synthesis comes from the recycling of choline from metabolized ACh. Another source is the breakdown of the phospholipid, phosphatidylcholine. One of the strategies to increase ACh neurotransmission is the administration of choline in the diet. However, this has not been effective, probably because the administration of choline does not increase the availability of choline in the CNS.
sTorage of aCh
The majority of the ACh in nerve endings is contained in clear (as viewed in the electron microscope) 100 um vesicles. A small amount is also free in the cytosol. Vesicle-bound ACh is not accessible to degradation by acetylcholinesterase (see below). The uptake of ACh into storage vesicle occurs through an energy-dependent pump that acidifies the vesicle. The acidified vesicle then uses a vesicular ACh transporter (VAChT) to exchange protons for ACh molecules. No useful pharmacological agents are available to modify cholinergic function through interaction with the storage of ACh. Interestingly, the gene for VAChT is contained on the first intron of the choline acetyltransferase gene. This proximity implies the two important cholinergic proteins are probably regulated coordinately.
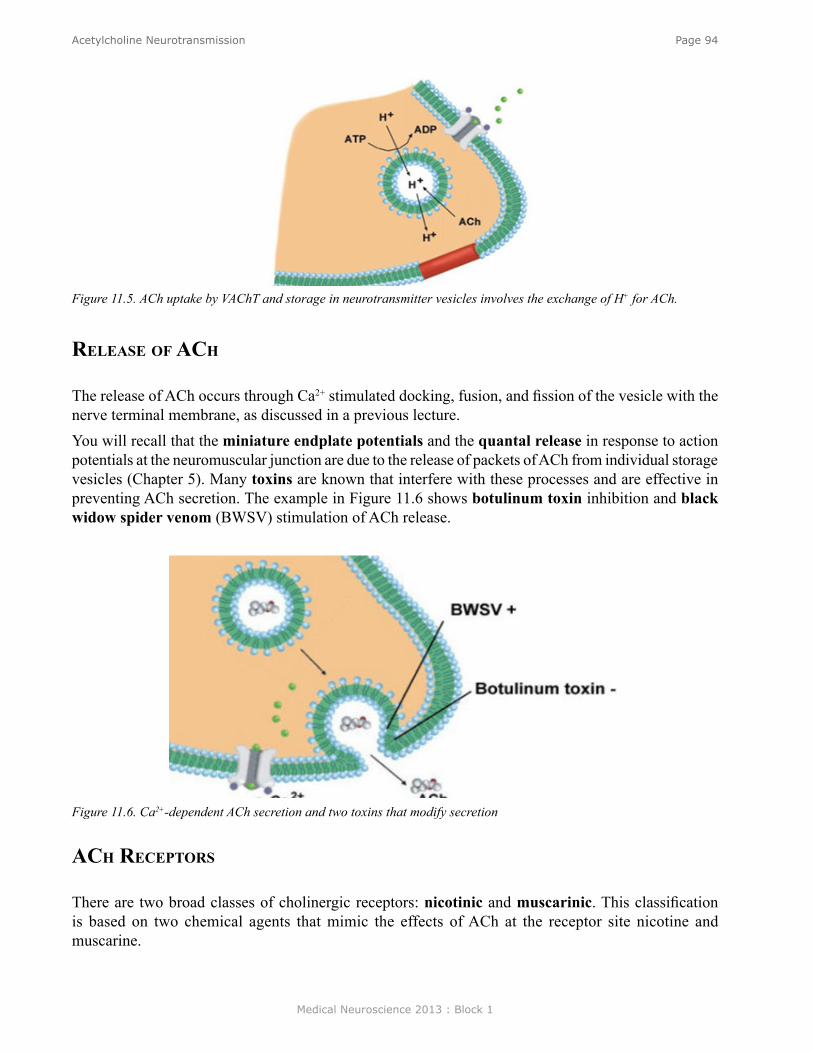
Medical Neuroscience 2013 : Block 1
Page 94Acetylcholine Neurotransmission
Figure 11.5. ACh uptake by VAChT and storage in neurotransmitter vesicles involves the exchange of H+ for ACh.
release of aCh
The release of ACh occurs through Ca2+ stimulated docking, fusion, and fission of the vesicle with the nerve terminal membrane, as discussed in a previous lecture. You will recall that the miniature endplate potentials and the quantal release in response to action potentials at the neuromuscular junction are due to the release of packets of ACh from individual storage vesicles (Chapter 5). Many toxins are known that interfere with these processes and are effective in preventing ACh secretion. The example in Figure 11.6 shows botulinum toxin inhibition and black widow spider venom (BWSV) stimulation of ACh release.
Figure 11.6. Ca2+-dependent ACh secretion and two toxins that modify secretion
aCh reCePTors
There are two broad classes of cholinergic receptors: nicotinic and muscarinic. This classification is based on two chemical agents that mimic the effects of ACh at the receptor site nicotine and muscarine.

Medical Neuroscience 2013 : Block 1
Page 95Acetylcholine Neurotransmission
Table I summarizes some of the properties of nicotinic and muscarinic receptors.
Table I Nicotinic and Mauscarinic Receptors and their actions
Nicotinic Muscarinic Bind nicotine Bind muscarine Blocked by curare (tubocurarine) Blocked by atropine Linked to ionic channels Linked to 2nd messenger systems through G proteins (see below) Response is brief and fast Response is slow and prolonged Located at neuromuscular junctions, autonomic ganglia, and to a small extent in the CNS
Found on myocardial muscle, certain smooth muscle, and in discrete CNS regions
Mediate excitation in target cells Mediate inhibition and excitation in target cells Postsynaptic Both pre- and postsynaptic
The niCoTiniC reCePTor is an ion Channel
As mentioned in Table 1, nicotinic receptors are located at the NMJ, autonomic ganglia and sparsely in the CNS.
Figure 11.7. Schematic of the five subunit nicotinic ACh receptor in the postsynaptic membrane at the NMJ. ACh binds to the two α subunits. The bottom half shows the molecular structure of each α subunit of the nicotinic receptor based on cDNA derived amino acid sequence. The β, γ and δ subunits have an analogous structure to the α subunit.
The NMJ nicotinic ACh receptor consists of five polypeptide subunits: two α subunits and one each of ß, δ , and γ (see Figure 11.7). A funnel-shaped internal ion channel is surrounded by the five subunits. The binding surface of the receptor appears to be primarily on the α subunits, near the outer surface of the molecule. The subunits contain recognition sites for agonists, reversible antagonists, and α -toxins (cobra α-toxin and α-bungarotoxin). Whereas the NMJ nicotinic receptor is composed of four different species of subunit (2 α, β, γ, δ), the neuronal nicotinic receptor also is composed of only two subunit types (2 α and 3 β).

Medical Neuroscience 2013 : Block 1
Page 96Acetylcholine Neurotransmission
The musCariniC reCePTor is CouPled To g-ProTeins
Muscarinic receptors, classified as G protein coupled receptors (GPCR), are located at parasympathetic autonomically innervated visceral organs, on the sweat glands and piloerector muscles and both post-synaptically and pre-synaptically in the CNS (Table 1). The muscarinic receptor is composed of a single polypeptide. Seven regions of the polypeptide are made up of 20-25 amino acids arranged in an α helix. Because each of these regions of the protein is markedly hydrophobic, they span the cell membrane seven times as depicted in Figure 11.8. The third internal loop and the carboxyl-terminal tail of the polypeptide receptor are believed to be the site of the interaction of the muscarinic receptor with G proteins (see right). The site of agonist binding is a circular pocket formed by the upper portions of the seven membrane-spanning regions.
Figure 11.8. ACh released into the extracellular space interacts with muscarinic receptors on both the innervated cell and the ACh nerve ending.
ACh has excitatory actions at the neuromuscular junction, at autonomic ganglion, at certain glandular tissues and in the CNS. It has inhibitory actions at certain smooth muscles and at cardiac muscle. The biochemical responses to stimulation of muscarinic receptor involve the receptor occupancy causing an altered conformation of an associated GTP-binding protein (G protein). G protein is made up of the three subunits α, β and γ. In response to the altered conformation of the muscarinic receptor, the α subunit of the G protein releases bound guanosine diphosphate (GDP) and simultaneously binds guanosine triphosphate (GTP). The binding of GTP "activates" the G protein, allowing dissociation of the α subunit from the trimeric complex and for it to interact with effector systems to mediate specific responses. An inherent GTPase catalytic activity of the G protein hydrolyzes the GTP back to GDP. This hydrolysis terminates the action of the G protein. The rate of hydrolysis of the GTP thus dictates the length of time the G protein remains activated. The responses mediated by muscarinic receptors through G proteins include:
Inhibition of Adenylyl Cyclase: The muscarinic receptor, through interaction with an inhibitory GTP-binding protein, acts to inhibit adenylyl cyclase. Reduced cAMP production leads to reduced activation of cAMP-dependent protein kinase, reduced heart rate, and contraction strength.

Medical Neuroscience 2013 : Block 1
Page 97Acetylcholine Neurotransmission
Stimulation of Phospholipase C: The muscarinic receptor activates phosphoinositide-specific phospholipase C (PLCβ) through interaction with a GTP-binding protein. The hydrolysis of phosphatidylinositol bis-phosphate yields two second messengers; inositol tris-phosphate (IP3) and diacylglycerol (DAG). The DAG activates protein kinase C (not shown). Cellular responses are influenced by PKC’s phosphorylation of target proteins. The IP3 diffuses to the smooth endoplasmic reticulum (ER) where it interacts with IP3 receptors to increase Ca2+ release from this intracellular storage site.
TerminaTion of aCh aCTion
ACh binds only briefly to the pre- or postsynaptic receptors. Following dissociation from the receptor, the ACh is rapidly hydrolyzed by the enzyme acetylcholinesterase (AChE) as shown in Figure 11.9. This enzyme has a very high catalysis rate, one of the highest known in biology. AChE is synthesized in the neuronal cell body and distributed throughout the neuron by axoplasmic transport. AChE exists as alternatively spliced isoforms that vary in their subunit composition. The variation at the NMJ is a heteromeric protein composed of four subunits coupled to a collagen tail that anchors the multi-subunit enzyme to the cell membrane of the postsynaptic cell (Figure 11.9). This four-subunit form is held together by sulfhydryl bonds and the tail anchors the enzyme in the extracellular matrix at the NMJ. Other isoforms are homomeric and freely soluble in the cytoplasm of the presynaptic cell. AChE, unlike ChAT, is found in non-cholinergic neurons as well. In addition, other cholinesterases exist throughout the body, which are also able to metabolize acetylcholine. These are termed pseudocholinesterases.
Figure 11.9 Hydrolysis of ACh to acetate and choline at the NMJ and cholinergic synapses.
Drugs that inhibit ACh breakdown are effective in altering cholinergic neurotransmission. In fact, the irreversible inhibition of AChE by isopropylfluoroesters is so toxic that they can be incompatible with life—inhibiting the muscles for respiration. This inhibition is produced because ACh molecules accumulate in the synaptic space, keep the receptors occupied, and cause paralysis. Two notable examples are insecticides and the gases used in biological warfare. The mechanism of action of these irreversible inhibitors of AChE is that they carbamylate the AChE, rendering it inactive. The carbamylation inactivates both the acetyl and choline binding domains. A recently developed antidote to these inhibitors cleaves the nerve gas so that it will dissociate from the AChE.

Medical Neuroscience 2013 : Block 1
Page 98Acetylcholine Neurotransmission
In contrast to the irreversible inhibitors, the reversible AChE inhibitors are effective in transiently increasing the ACh level and are effective in diseases and conditions where an increased ACh level is desired. The clinically important compound, eserine (physostigmine), reversibly inhibits AChE.
Physiology
Nicotinic receptor activation causes the opening of the channel formed by the receptor. This increases the Na+ movement into the target cell, leading to depolarization and generation of the action potential. This rapidly developing change, termed a fast EPSP, is illustrated in Figures 4.3 and 6.2. The action of ACh on nicotinic receptors is always excitatory.Muscarinic receptor activation of postsynaptic cells can be either excitatory or inhibitory and is always slow in onset and long in duration (Table 1). As described earlier, G protein activation underlies all actions of the muscarinic receptors, thus accounting for their slow onset.
behavior
The rapid nature of the synaptic transmission mediated by the nicotinic receptor is consistent with its role at the NMJ and in the ganglion of the ANS. Little is known about the role of the nicotinic recep-tor role in CNS behavior. Clearly, nicotine stimulation is related in some manner to reinforcement, as indicated by the prevalence of nicotine addiction among humans. Muscarinic receptors, in contrast, are important mediators of behavior in the CNS. One example is their role in modulating motor control circuits in the basal ganglia. A second example is their participation in learning and memory. The latter is inferred from two types of observations: 1) muscarinic antagonists are amnesic agents, and 2) deterioration of the cholinergic innervation of the neocortex is associated with memory loss in Alzheimer’s disease.
CliniCal
Alzheimer's disease: A disease in which a marked deterioration occurs in the CNS, the hallmark of which is a progressive dementia. One of the characteristics of this disease is a marked decrease in ACh concentrations in the cerebral cortex and caudate nucleus. Myasthenia gravis: A disease of the neuromuscular junction in which the receptors for ACh are destroyed through the actions of the patient's own antibodies.Cholinergic Pharmacology: Numerous drugs are used clinically to interact with the cholinergic systems. Table II summarizes the major uses for cholinergic drugs.
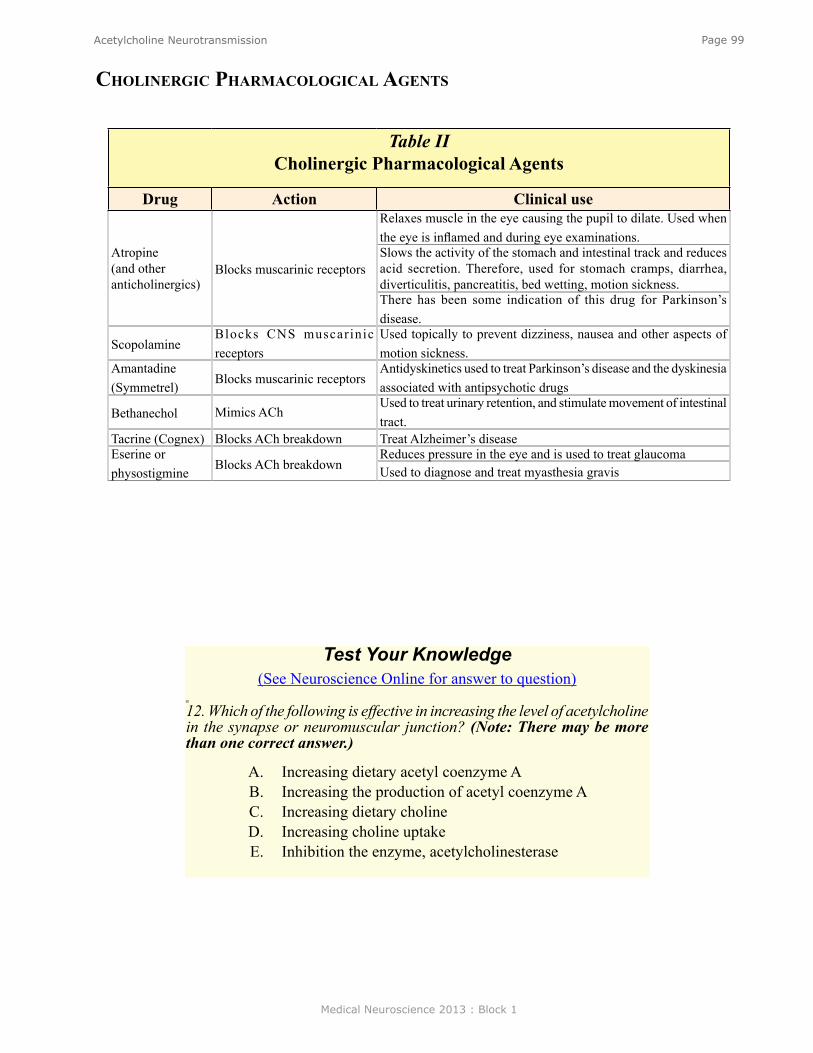
Medical Neuroscience 2013 : Block 1
Page 99Acetylcholine Neurotransmission
Table II Cholinergic Pharmacological Agents
Drug Action Clinical use
Atropine (and other anticholinergics)
Blocks muscarinic receptors
Relaxes muscle in the eye causing the pupil to dilate. Used when the eye is inflamed and during eye examinations. Slows the activity of the stomach and intestinal track and reduces acid secretion. Therefore, used for stomach cramps, diarrhea, diverticulitis, pancreatitis, bed wetting, motion sickness. There has been some indication of this drug for Parkinson’s disease.
Scopolamine Blocks CNS muscarinic receptors
Used topically to prevent dizziness, nausea and other aspects of motion sickness.
Amantadine (Symmetrel) Blocks muscarinic receptors
Antidyskinetics used to treat Parkinson’s disease and the dyskinesia associated with antipsychotic drugs
Bethanechol Mimics ACh Used to treat urinary retention, and stimulate movement of intestinal tract.
Tacrine (Cognex) Blocks ACh breakdown Treat Alzheimer’s disease Eserine or physostigmine Blocks ACh breakdown
Reduces pressure in the eye and is used to treat glaucoma Used to diagnose and treat myasthesia gravis
CholinergiC PharmaCologiCal agenTs
Test Your Knowledge(See Neuroscience Online for answer to question)
12. Which of the following is effective in increasing the level of acetylcholine in the synapse or neuromuscular junction? (Note: There may be more than one correct answer.)
A. Increasing dietary acetyl coenzyme A B. Increasing the production of acetyl coenzyme AC. Increasing dietary cholineD. Increasing choline uptakeE. Inhibition the enzyme, acetylcholinesterase

Medical Neuroscience 2013 : Block 1
Page 100Biogenic Amine Neurotransmitters
Biogenic Amine Neurotransmitters
assignmenT
I. Introduction and classificationA. CatecholaminesB. IndolaminesC. Histamine
II. Anatomical distribution A. Autonomic Nervous System (ANS)B. Central Nervous System (CNS)
III. Cell biology A. Biosynthesis of monamines (MA)B. Storage of monaminesC. Release of monamines D. Monoamine receptors E. Inactivation of monoamines
IV. Physiological and behavioral actions V. Clinical importance and pharmacology
major objeCTives
1. Know the biosynthesis, release, inactivation, and receptor properties of the monoamine neurotransmitters.
2. Have a basic understanding of the anatomical distribution of monamine neurons and the function of the monoamine neurotransmitters in normal behavior and disease.
required reading
● Waymire, J.C. Notes on Biogenic Amine Neurotransmitters, Lecture Supplement Volume I.
suggesTed reading (noT required)
● Waymire, J. C. Biogenic Amine Neurotransmitters, Neuroscience Online: http://nba.uth.tmc.edu/neuroscience/s1/chapter12.html
Lecturer: Jack C. WaymireJanuary 23, 2013 | 8:00 AM

Medical Neuroscience 2013 : Block 1
Page 101Biogenic Amine Neurotransmitters
inTroduCTion
Monoamines (also known as "biogenic amines") include three classes of neurotransmitters: ● Catecholamines
○ Dopamine (DA), norepinephrine (NE, also called noradrenaline) and epinephrine (E, also called adrenaline) make up a class of neurotransmitters named on the basis of the hydroxylated phenol ring, termed a catechol nucleus.
● Indolamines ○ Serotonin (5-hydroxytryptamine; 5-HT); is the principal member of this group of compounds.
The name serotonin is derived from the fact that this substance was first isolated from the serum based on its ability to cause an increase in blood pressure. Melatonin, a second indolamine, is restricted to the pineal and is released into the blood stream in a manner that is regulated by the diurnal cycle. Melatonin will not be covered further in this chapter.
● Histamine ○ Histamine has been recognized as a neurotransmitter in the CNS only within the past fifteen
years.
anaTomy of CaTeCholamines
Catecholamines are neurotransmitters in a sympathetic limb of the autonomic nervous system and in the CNS.
autoNoMic Nervous systeMsNorepinephrine is the neurotransmitter in postganglionic sympathetic neurons where it acts on smooth muscle to cause either contraction or relaxation, depending on the types of receptors present (see below). Epinephrine and norepinephrine are neurohumoral agents released into the circulation by the adrenal medulla. The ratio of E to NE released is 4 to 1.
ceNtral Nervous systeMGenerally, the cell bodies of catecholamine neurons are found in clusters in the brain stem or midbrain and project to other regions of the brain and spinal cord. NE, for example, projects to almost every area of the brain. In contrast, DA has a more restricted projection.
dopaMiNe - aNatoMy The major site of DA cell bodies is the midbrain. These clusters of cells give rise to four DA systems: 1) Mesostriatal system; 2) Mesolimbocortical system; 3) Periventricular system; 4) Tuberohypophyseal system.
NorepiNephriNe - aNatoMyThe major site of NE cell bodies is the medulla and pons. The NE cells consist of three main groups: 1) locus coeruleus complex; 2) lateral tegmental system; 3) dorsal medullary system
NE neurons project diffusely to broad regions of the brain where their nerve terminals lack conventional synaptic junctions. Release of transmitter from these cells is described as volume transmission, because NE, once released, is thought to diffuse and influence a number of adjacent cells.

Medical Neuroscience 2013 : Block 1
Page 102Biogenic Amine Neurotransmitters
serotoNiN - aNatoMySerotonin cells are located in two clusters: 1) a caudal system in the medulla (B1-B4); and 2) a rostral system in the midbrain (B5-B9). Both project widely throughout the CNS.
histaMiNe - aNatoMyHistamine cells (HA) are located exclusively in the basal posterior hypothalamus. These cells project extensively throughout the neural axis in a manner analogous to the NE and 5-HT systems. Although HA has not been investigated extensively, based on its diffuse innervation of the CNS and lack of classic synaptic contacts, it is likely that histamine has a broad behavioral and physiological function. Histamine is also the major active substance released from mast cells. The presence of mast cells in the blood in the CNS has hindered the analysis of the role of histamine as a neurotransmitter.
Cell biology
The monoamines will be considered as a group in discussing the cell biology of their 1) synthesis, 2) storage and 3) release. Monoamine receptors and termination of action of each monoamine will be considered separately.
BiosyNthesis oF MoNoaMiNes All monoamine (MA) neurotransmitters are synthesized from amino acids through a series of enzyme catalyzed reactions in which hydroxylation, decarboxylation and/or methylation convert the precursor amino acid into the active monoamine neurotransmitter.
BiosyNthesis oF dopaMiNe (da), NorepiNephriNe (Ne), aNd 5-hydroxytryptaMiNe (5-ht) Biosynthesis of all monoamines occurs primarily in the nerve terminal. As shown in Figure 12.1, the first step in the synthesis of catecholamines (DA and NE, as well as E, not shown) is the hydroxylation of the tyrosine to form DOPA. An analogous reaction, the hydroxylation of tryptophan to 5 hydroxytryptophane (5-HTP) is the first step in the biosynthesis of 5-HT. Both tyrosine hydroxylase and tryptophan hydroxylase are the rate-limiting steps in the biosynthetic pathway of the respective monoamines. Both enzymes are mixed function mono-oxygenases requiring molecular oxygen, iron and the cofactor, tetrahydrobiopterin (BH4) for activity. BH4 is converted to BH2 during the hydroxylation and must be regenerated to BH4 in order for monoamine biosynthesis to continue. As shown in Figure 12.1, the enzyme pteridine reductase regenerates the active cofactor. Pteridine reductase is therefore also an essential enzyme in the synthesis of catecholamines. The next step in the biosynthesis of monoamines is the decarboxylation by aromatic amino acid decarboxylase (AADC) to form the corresponding monoamine (Dopamine and 5 hydroxytryptamine 5-HT, respectively). NE is then formed from dopamine through an additional reaction, the hydroxylation of the 2nd carbon of the DA side chain. This last hydroxylation step occurs within the monoamine storage vesicle (see Figure 12.3a) and is catalyzed by dopamine β hydroxylase.
Two additional cofactors are required for the synthesis of monoamines; vitamin B6 is necessary as a cofactor for AADC catalyzed decarboxylation. Vitamin C is required as a cofactor for DA conversion to NE in the storage vesicle (see Figure 12.3a).
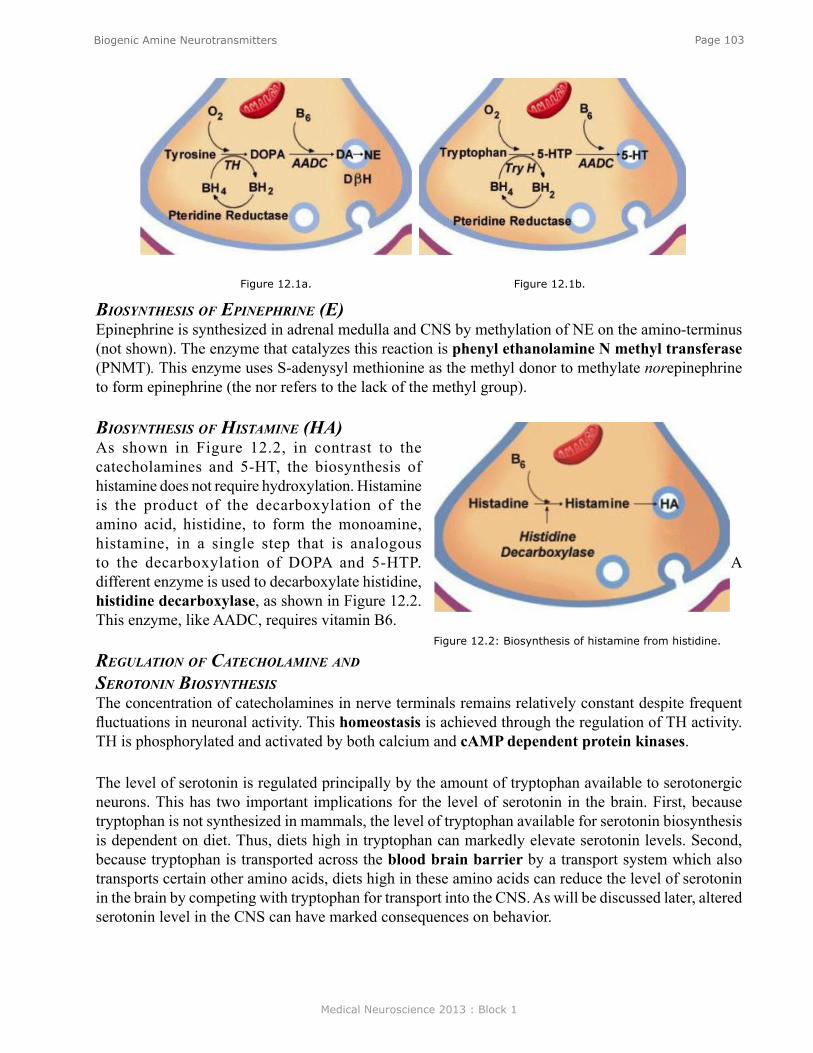
Medical Neuroscience 2013 : Block 1
Page 103Biogenic Amine Neurotransmitters
BiosyNthesis oF epiNephriNe (e) Epinephrine is synthesized in adrenal medulla and CNS by methylation of NE on the amino-terminus (not shown). The enzyme that catalyzes this reaction is phenyl ethanolamine N methyl transferase (PNMT). This enzyme uses S-adenysyl methionine as the methyl donor to methylate norepinephrine to form epinephrine (the nor refers to the lack of the methyl group).
BiosyNthesis oF histaMiNe (ha) As shown in Figure 12.2, in contrast to the catecholamines and 5-HT, the biosynthesis of histamine does not require hydroxylation. Histamine is the product of the decarboxylation of the amino acid, histidine, to form the monoamine, histamine, in a single step that is analogous to the decarboxylation of DOPA and 5-HTP. A different enzyme is used to decarboxylate histidine, histidine decarboxylase, as shown in Figure 12.2. This enzyme, like AADC, requires vitamin B6.
regulatioN oF catecholaMiNe aNd serotoNiN BiosyNthesis The concentration of catecholamines in nerve terminals remains relatively constant despite frequent fluctuations in neuronal activity. This homeostasis is achieved through the regulation of TH activity. TH is phosphorylated and activated by both calcium and cAMP dependent protein kinases.
The level of serotonin is regulated principally by the amount of tryptophan available to serotonergic neurons. This has two important implications for the level of serotonin in the brain. First, because tryptophan is not synthesized in mammals, the level of tryptophan available for serotonin biosynthesis is dependent on diet. Thus, diets high in tryptophan can markedly elevate serotonin levels. Second, because tryptophan is transported across the blood brain barrier by a transport system which also transports certain other amino acids, diets high in these amino acids can reduce the level of serotonin in the brain by competing with tryptophan for transport into the CNS. As will be discussed later, altered serotonin level in the CNS can have marked consequences on behavior.
Figure 12.1a. Figure 12.1b.
Figure 12.2: Biosynthesis of histamine from histidine.

Medical Neuroscience 2013 : Block 1
Page 104Biogenic Amine Neurotransmitters
sTorage of monoamines
Monoamine neurotransmitters are stored in vesicles that appear dark at the EM level and are thus referred to as dense core vesicles . MA neurotransmitters are stored at a high concentration and are complexed with ATP. As shown in Figure 12.3, MA neurotransmitters are taken into the vesicles by an exchange of H+ for the MA. In NE cells DA is taken up and converted to NE by DβH.
release of monoamines
Neuronal activation elicits the release of MA neurotransmitters by a calcium-dependent exocytosis, as described previously. Because there is no classic postsynaptic specialization associated with the majority of MA nerve endings, the released MA neurotransmitters diffuse to postsynaptic cells in the vicinity where they stimulate MA receptors (volume neurotransmission).
MA neurotransmitters also act on the presynaptic cell, as shown in Figure 12.4 to influence their cell biology in a feed back manner. The interaction with the presynaptic receptors (termed autoreceptors) can both stimulate MA biosynthesis and inhibit the further release of neurotransmitter. Both the pre- and postsynaptic MA receptors are G protein linked, seven trans-membrane receptors. Their structure is similar to the muscarinic receptors discussed in the lecture on cholinergic neurotransmission.
ProPerTies of monoamine reCePTors
The vast majority of the MA receptors are seven transmembrane, G-protein coupled receptors (GPCR) that mediate MA action through one of a few mechanisms. These are the same mechanisms employed by other GPCR, such as the muscarinic receptors (Chapter 11, Part 5 of Neuroscience Online) and GPC-glutamate receptors (Chapter 13, Part 3 of Neuroscience Online). These mechanisms are:
1. Stimulation or inhibition of adenylyl cyclase,2. Stimulation of PLCβ or PLA, and 3. Direct action on ion channel.
As will be described below, one type of MA receptor, 5-HT3, is unusual in that it is NOT LINKED TO G PROTEIN RECEPTORS. Instead, this receptor is a ligand gated ion channel, similar in structure and function to ionotropic nicotinic cholinergic receptors and glutamate receptors.
Figure 12.3a Figure 12.3b
An antiporter that exchanges protons for monoamines (MA) mediates storage of monoamines in dense core vesicles. Left: In NE cells, DA is taken up then converted to NE within the vesicle by the
enzyme DBH. Right: All other monoamine cells merely store the MA neurotransmitters.
Figure 12.4 MA release and interaction with both presynaptic and
postsynaptic receptors.

Medical Neuroscience 2013 : Block 1
Page 105Biogenic Amine Neurotransmitters
ne and e PeriPheral reCePTors
The receptors for NE and E were originally classified based on the observation that some physiological actions were mimicked by the catecholamine analog, isoproterenol, whereas others were not. This observation led to the convention that actions that could be mimicked by isoproterenol were classified as mediated by β-receptors. Those actions that were not mimicked by isoproterenol were classified as mediated by α-receptors. This classification has since been extended to include subclasses of α and ß receptors based on the capacity of drugs to selectively activate (or block) specific physiological responses to NE and E. The molecular cloning of mRNAs for distinct subclasses of NE and E receptors also aided in the classification of receptors. Noteworthy is the fact that most α receptor responses are excitatory, while most β responses are inhibitory (although some exceptions exist, e.g. cardiac muscle). Also, the α receptor is invariably linked to IP3 production, whereas the β receptor is associated with increased levels of cAMP.
Cns ne and da reCePTors
Ne receptorsThe distribution of NE receptors in the CNS is complex and not yet well resolved. Generally, both α and β receptors are believed to be modulators of the actions of other neurotransmitters. α1 receptors are often excitatory, acting via IP3. In contrast, α2 receptors are inhibitory acting via decreased levels of cAMP. β receptors are inhibitory and act through increased levels of cAMP. The anatomical location of the specific receptor subtypes is not yet clearly delineated.
da receptors In the CNS, dopamine receptors, designated by the letter D, are grouped into two large families based on cDNA-derived structural similarities, synaptic action and signaling mechanism. The D1 family (D1 and D5) increases cAMP level, and has a positive influence on the excitability of its target cell. The D2 family (D2, D3, and D4) decreases cAMP level and decreases the excitability of the target cell. D1 are usually post synaptic while D2 are usually presynaptic.
5-ht receptorsAll but one of the 5-HT receptors belongs to the G protein coupled receptor superfamily. As mentioned above, the one exception is the 5-HT3 receptor, which is a ligand gated ion channel. All other 5-HT mediated actions occur through the same types of second messenger mechanisms as cholinergic and catecholamine G protein linked receptors.
Two classes of 5-HT receptors, 5-HT1B and 5-HT1D, appear to predominantly act as presynaptic autoreceptors to modulate the synthesis and release of 5-HT from the terminals of serotonergic neurons.
histaMiNe receptorsThree subtypes of histamine receptors have been identified. All three are G protein linked and all three are present in the CNS as well as the periphery.

Medical Neuroscience 2013 : Block 1
Page 106Biogenic Amine Neurotransmitters
inaCTivaTion of ma neuroTransmiTTers by reuPTake and meTabolism
The major mechanism for the inactivation of secreted MA is the reuptake into the nerve terminal from which the MA was released. Under conditions of very high neuronal activity, the MA will also be taken up by neighboring glial cells and will overflow into the capillaries perfusing the CNS. Under all three situations, a portion of the MA will be metabolized by enzymes that inactivate the MA, converting them to inactive products. As described below, measurement of these metabolites is used clinically and in research to monitor the activity of the MA systems.
reuPTake of ma neuroTransmiTTers
High affinity transport (reuptake) into axon terminals is the main process of inactivation of released monoamines. Reuptake requires sodium ions and a source of energy (e.g., ATP) and is mediated by a protein carrier located on the plasma membrane of the monamine neurons. Tricyclic antidepressants and cocaine inhibit the transporters for DA, NE and 5-HT. Within the past ten years the structure of several MA transporters has been determined and shown to consist of a twelve transmembrane protein with both the N and C terminal ends residing within the cytoplasm (Figure 12.5). The powerful addictive drugs cocaine and amphetamine increase the level of MA neurotransmitters in the extracellular space. Cocaine acts by blocking the transport of MA (Figure 12.5) neurotransmitters into the terminal and as a consequence increases MA in the extracellular space. In contrast, amphetamine reverses the transport direction transporting MA neurotransmitters out of the nerve terminal.
A portion of released catecholamines diffuses to the extracellular space where monoamine oxidase (MAO) and/or catechol-0-methyl-transferase (COMT) catabolize it. This route of inactivation is more prominent following extremely high levels of catecholaminergic neuronal activity.
meTabolism of ma neuroTransmiTTers
catecholaMiNes aNd 5-htThe enzymatic metabolism of MA neurotransmitters is carried out by MAO, COMT and histamine methyl transferase. These enzymes are widely distributed in tissues.
MoNoaMiNe oxidase (Mao)This metabolic enzyme is located on the outer membrane of the mitochondrion and metabolizes DA, NE and 5-HT by oxidative deamination of to the corresponding aldehyde. These metabolites are excreted (see Table I), or further metabolized by methylation through the action of catechol- O-methyltransferase and then excreted (see below).
catechol-o -Methyl-traNsFerase (coMt)This extraneuronal enzyme inactivates catecholamines by methylation of the hydroxyls on the catechol ring. COMT methylates either catecholamines that have already been metabolized by MAO or those that have not. The metabolites of catecholamines are shown in Table I.
Figure 12.5 Reuptake of MA neurotransmitters by a transporter with a twelve transmembrane structure.
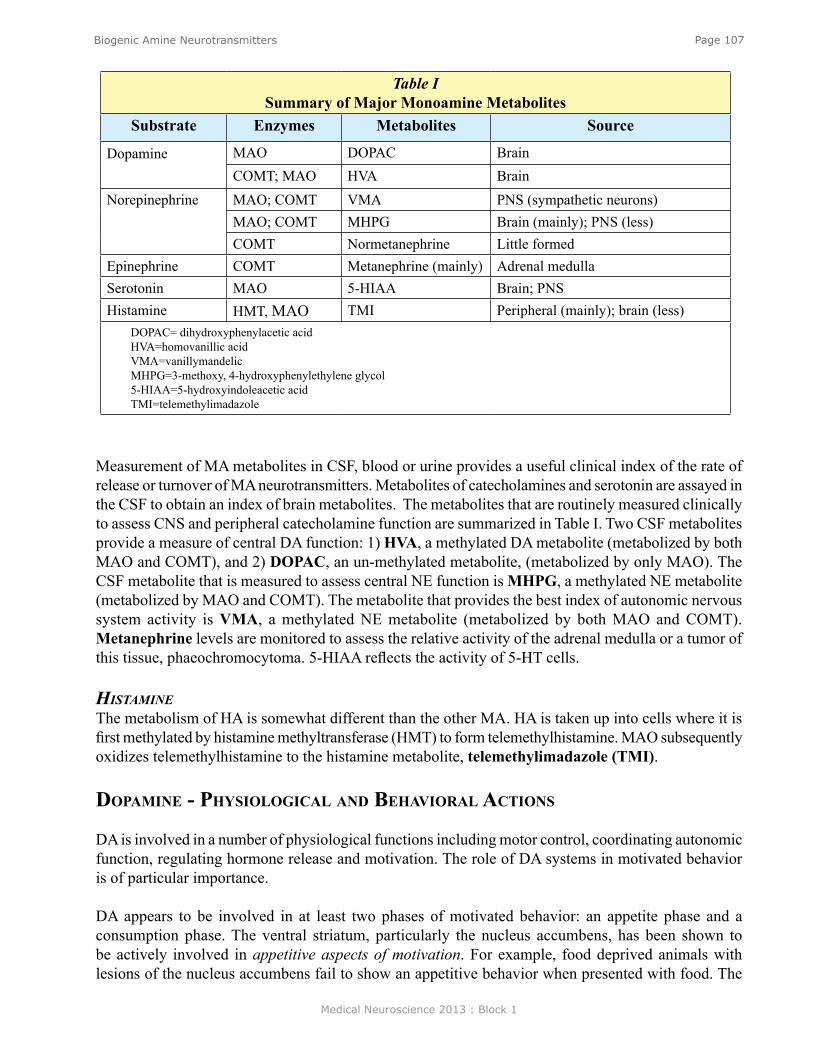
Medical Neuroscience 2013 : Block 1
Page 107Biogenic Amine Neurotransmitters
Measurement of MA metabolites in CSF, blood or urine provides a useful clinical index of the rate of release or turnover of MA neurotransmitters. Metabolites of catecholamines and serotonin are assayed in the CSF to obtain an index of brain metabolites. The metabolites that are routinely measured clinically to assess CNS and peripheral catecholamine function are summarized in Table I. Two CSF metabolites provide a measure of central DA function: 1) HVA, a methylated DA metabolite (metabolized by both MAO and COMT), and 2) DOPAC, an un-methylated metabolite, (metabolized by only MAO). The CSF metabolite that is measured to assess central NE function is MHPG, a methylated NE metabolite (metabolized by MAO and COMT). The metabolite that provides the best index of autonomic nervous system activity is VMA, a methylated NE metabolite (metabolized by both MAO and COMT). Metanephrine levels are monitored to assess the relative activity of the adrenal medulla or a tumor of this tissue, phaeochromocytoma. 5-HIAA reflects the activity of 5-HT cells.
histaMiNeThe metabolism of HA is somewhat different than the other MA. HA is taken up into cells where it is first methylated by histamine methyltransferase (HMT) to form telemethylhistamine. MAO subsequently oxidizes telemethylhistamine to the histamine metabolite, telemethylimadazole (TMI).
doPamine - PhysiologiCal and behavioral aCTions
DA is involved in a number of physiological functions including motor control, coordinating autonomic function, regulating hormone release and motivation. The role of DA systems in motivated behavior is of particular importance.
DA appears to be involved in at least two phases of motivated behavior: an appetite phase and a consumption phase. The ventral striatum, particularly the nucleus accumbens, has been shown to be actively involved in appetitive aspects of motivation. For example, food deprived animals with lesions of the nucleus accumbens fail to show an appetitive behavior when presented with food. The
Table I Summary of Major Monoamine Metabolites
Substrate Enzymes Metabolites Source
Dopamine MAO DOPAC BrainCOMT; MAO HVA Brain
Norepinephrine MAO; COMT VMA PNS (sympathetic neurons)MAO; COMT MHPG Brain (mainly); PNS (less)COMT Normetanephrine Little formed
Epinephrine COMT Metanephrine (mainly) Adrenal medullaSerotonin MAO 5-HIAA Brain; PNSHistamine HMT, MAO TMI Peripheral (mainly); brain (less)
DOPAC= dihydroxyphenylacetic acidHVA=homovanillic acidVMA=vanillymandelicMHPG=3-methoxy, 4-hydroxyphenylethylene glycol5-HIAA=5-hydroxyindoleacetic acidTMI=telemethylimadazole

Medical Neuroscience 2013 : Block 1
Page 108Biogenic Amine Neurotransmitters
consumption of the food, in contrast, is unimpaired. On the other hand, the dorsal striatum, particularly the caudate-putamen, appears to be involved in consummatory aspects of motivated behavior. Animals with lesions in this region will approach food but will not consume it. These and other observations have led to the proposition that DA mediates a performance activating effect of motivated behavior, as well as conveying internal reward signals. These characteristics of DA neurotransmission make it an extremely important neurotransmitter in motivational behavior as well as in the neuropharmacology of reward. Importantly, DA neurotransmission is hypothesized to be at the heart of the mechanisms of ALL addictive-drugs and addictive-behaviors.
norePinePhrine - PhysiologiCal and behavioral aCTions
NE is involved in a broad range of physiological functions and behaviors. One of the most important is its role in attention and arousal.
NE neurons appear to be involved in the regulation of an organism’s vigilance. The broad projection of the NE neurons in the locus coeruleus (LC) makes it especially well suited to act as a mechanism to alert cortical and thalamic areas to incoming sensory stimuli. The LC is electrophysiologically quiet during low vigilance states such as sleep or in the lack of sensory input. When exposed to a strong stimulus, the LC markedly increases its firing rate. The broad influence of the activated LC is to filter weak stimuli and enhance moderate stimuli. This filtering and enhancement by NE is believed to aid in CNS processing of sensory information. In support of this conclusion, the application of NE to cortical neurons reduces responsiveness to weak stimuli, and enhances responses to strong stimuli. α2 receptors appear to be important for these responses.
seroTonin - PhysiologiCal and behavioral aCTions
Serotonin is a key neurotransmitter in a number of physiological regulatory mechanisms and behaviors, including appetite, sleep, and aggression.
Serotonin is important in the regulation of appetite, and appears to act in a pathway that monitors the carbohydrate intake, acting as a negative regulator of the motivation to ingest carbohydrate. This response appears to be mediated by 5-HT in the hypothalamus and has led to the use of serotonin uptake blockers, such as fenfluramine, as obesity pills (Table VII).
Many clinical observations and animal behavioral studies support the conclusion that serotonin is an important factor in aggressive behavior and the expression of dominance versus submissive behavior. For example the use of pharmacological agents to decrease levels of 5-HT at synapses in animal studies consistently demonstrates that low 5-HT is associated with both increased aggressiveness and decreased dominance. Similarly, the measurement of 5-HT metabolites in CSF and blood of patients or experimental animals shows that low 5-HIAA predicts aggressiveness as well as risk taking and a lower social rank. This correlation between decreased 5-HT activity and increased aggression was recently supported by the observation that 5-HT1B receptor knock-out mice have a marked increase in aggressive behavior.

Medical Neuroscience 2013 : Block 1
Page 109Biogenic Amine Neurotransmitters
doPamine - CliniCal imPorTanCe and PharmaCology
DA is implicated in psychiatric illnesses (especially schizophrenia) and disorders of movement control.
schizophreNiaThe DA theory of schizophrenia is based on the observation that DA antagonists are effective antipsychotic drugs. Their capacity to inhibit DA receptors correlates well with their antipsychotic efficacy. Currently, clinical studies are attempting to develop DA antagonists with specific DA receptor subtype efficacy that will most effectively decrease the antipsychotic symptoms without influencing other DA actions, such as movement control.
disorders oF MoveMeNt coNtrolIt is now well accepted that decreased DA in the substantia nigra and striatum is the critical lesion in Parkinson’s disease. Autopsy shows that nearly all DA is lost during the course of this disease, apparently due to the degeneration of DA neurons. l-DOPA is used to treat this disease’s symptoms because it can be converted to DA by AADC in the cells in the vicinity of the degenerated nerve endings to replace the missing endogenous DA. Some preparations of l-DOPA include a peripheral AADC inhibitor so that more of the l-DOPA will be available to the CNS. Other drugs that are effective in treating Parkinson’s symptoms are DA agonists, as well as drugs that prevent DA metabolism, MAO and COMT inhibitors.
norePinePhrine - CliniCal imPorTanCe and PharmaCology
aFFective disordersNE is believed to be involved in the etiology of some unipolar and bipolar affective disorders. This conclusion is based to a large degree on the observation that drugs that are effective in treating depression are also good at either 1) preventing the metabolism of NE (MAO inhibitors), or 2) preventing the removal of NE from the extracellular space by uptake into nerve endings. There is also evidence that the levels of the CNS NE metabolites are lower in depressed patients and higher during the manic phase of bipolar disorder in manic patients.
seroTonin - CliniCal imPorTanCe and PharmaCology
aFFective disordersLow levels of 5-HT and metabolites are associated with depression and especially a type of depression that is more likely to lead to suicide. Several studies have shown reduced 5-HT in brains of suicide victims as well as a low 5HIAA in CSF of depressed patients who have high incidence of suicide attempts. Recent studies indicate that this type of 5-HT influence may start early in life; low levels of 5HIAA have been found in children and adolescents with disruptive behavioral disorders. Some of the drugs that are effective antidepressant treatments are nonspecific with respect to their relative influence on NE versus 5-HT disposition, thus it has been difficult to know for certain which monoamine is responsible for the treatment effects. More recently selective serotonin reuptake inhibitors (termed SSRIs) have been introduced for the treatment of depression that are among the more effective drugs available for this purpose.

Medical Neuroscience 2013 : Block 1
Page 110Biogenic Amine Neurotransmitters
oTher CliniCal uses of 5-hT drugs
oBsessive coMpulsive disorder5-HT dysfunction has been associated with obsessive compulsive disorder. Accordingly, selective 5-HT uptake blockers are used as a therapy for this condition.
aggressioNAlthough controversial, 5-HT reuptake blockers are used for the treatment of aggression.
eatiNg disordersAlso controversial is the use of the drug fenfluramine to treat eating disorders because of the toxic effects that occur in some individuals. Fenfluramine blocks 5-HT reuptake into nerve terminals.
schizophreNiaA number of recently introduced antipsychotic drugs are producing favorable results in treating the symptoms of schizophrenia. These drugs are interesting pharmacologically in that they block both DA and 5-HT receptors as well as ACh and HA (Table VII, Olanzapine).
MigraiNe headaches5-HT1 agonists are used for the treatment of migraine headache.
iNsoMNiaThe role of 5-HT in sleep regulation has lead to the hypothesis that reduced levels 5-HT may induce insomnia. Some clinicians are treating patients with 5-HT uptake blockers for this ailment.
Test Your Knowledge(See Neuroscience Online for answer to question)
13. Which of the following can be administered orally to increase dopamine levels in the CNS? (Note: There is more than one correct answer.)
A. Dopamine B. TyrosineC. Acetyl coenzyme AD. l-DOPAE. l-DOPA plus an aromatic L-amino acid decarboxylase inhibitor

Medical Neuroscience 2013 : Block 1
Page 111Amino Acid Neurotransmitters
Amino Acid Neurotransmitters
assignmenTI. Introduction and review
II. Structure of amino acid transmittersIII. Biosynthesis
A. Glutamate and aspartateB. GlycineC. GABA
IV. Release - Ca2+ - dependentV. Receptors
A. Glutamate1. N-methyl-D-aspartate (NMDA)2. Kainate/AMPA/Quisqualate (a)
B. GABA (a) C. GlycineD. Quisqualate (b) and GABA (b)
VI. Termination of actionVII. Diseases associated with glutamate
VIII. Diseases associated with GABA
major objeCTives1. Know how amino acid transmitters are synthesized.2. Know the difference between the glutamate receptor subtypes.3. Know the differences between the physiologic effects of excitatory amino acids and
inhibitory amino acids.4. Know the differences between ionotropic and G-protein coupled receptors.5. Know how the action of amino acid transmitters is terminated.
required reading ● Waxham, M.N. Notes on Amino Acid Neurotransmitters, Lecture Supplement Volume I; or ● Waxham, M.N. Amino Acid Neurotransmitters, Neuroscience Online:
http://nba.uth.tmc.edu/neuroscience/s1/chapter13.htmlsuggesTed reading
● Waxham, M.N. (2009) Neurotransmitter Receptors. In J.H. Byrne and J.L. Roberts (Eds.), From Molecules to Networks: An Introduction to Cellular and Molecular Neuroscience, Second Edition (pp. 321-358). Academic Press, Elsevier, Inc.
● See also Byrne, J.H. Synaptic Transmission in the Central Nervous System. Neuroscience Online: http://nba.uth.tmc.edu/neuroscience/s1/chapter06.html
Lecturer: M. Neal Waxham, Ph.D.January 23, 2013 | 9:00 AM
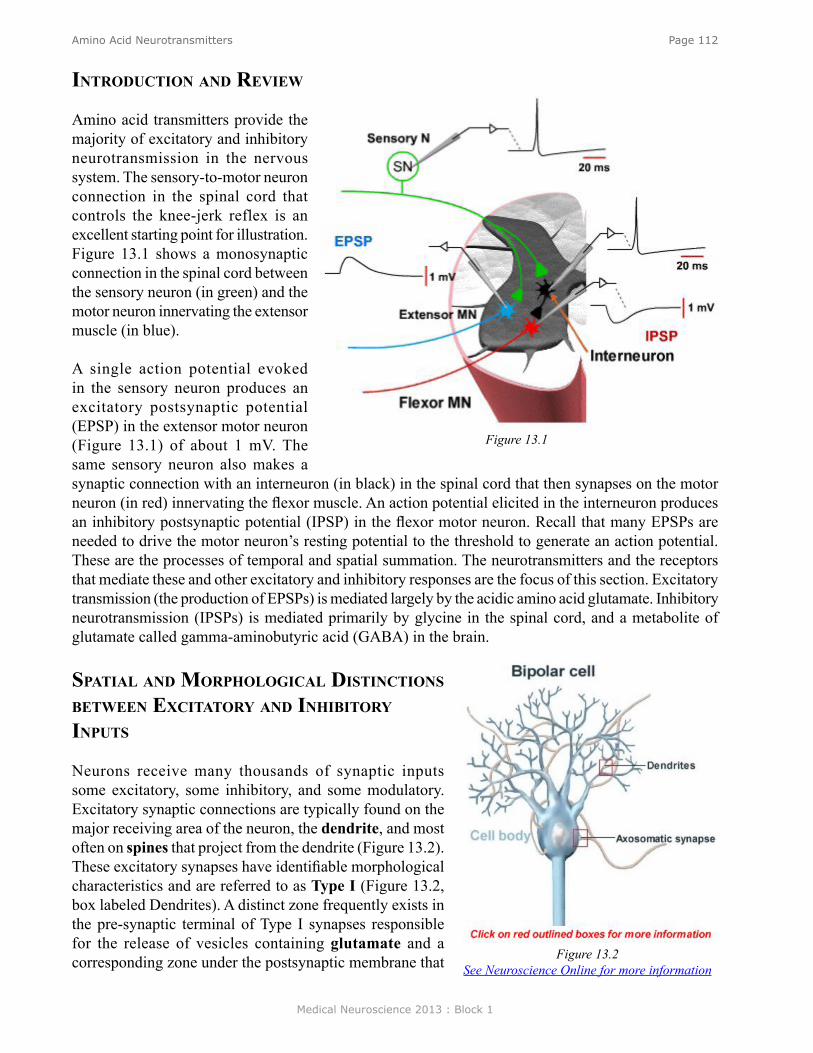
Medical Neuroscience 2013 : Block 1
Page 112Amino Acid Neurotransmitters
inTroduCTion and review
Amino acid transmitters provide the majority of excitatory and inhibitory neurotransmission in the nervous system. The sensory-to-motor neuron connection in the spinal cord that controls the knee-jerk reflex is an excellent starting point for illustration. Figure 13.1 shows a monosynaptic connection in the spinal cord between the sensory neuron (in green) and the motor neuron innervating the extensor muscle (in blue).
A single action potential evoked in the sensory neuron produces an excitatory postsynaptic potential (EPSP) in the extensor motor neuron (Figure 13.1) of about 1 mV. The same sensory neuron also makes a synaptic connection with an interneuron (in black) in the spinal cord that then synapses on the motor neuron (in red) innervating the flexor muscle. An action potential elicited in the interneuron produces an inhibitory postsynaptic potential (IPSP) in the flexor motor neuron. Recall that many EPSPs are needed to drive the motor neuron’s resting potential to the threshold to generate an action potential. These are the processes of temporal and spatial summation. The neurotransmitters and the receptors that mediate these and other excitatory and inhibitory responses are the focus of this section. Excitatory transmission (the production of EPSPs) is mediated largely by the acidic amino acid glutamate. Inhibitory neurotransmission (IPSPs) is mediated primarily by glycine in the spinal cord, and a metabolite of glutamate called gamma-aminobutyric acid (GABA) in the brain.
sPaTial and morPhologiCal disTinCTions beTween exCiTaTory and inhibiTory inPuTs
Neurons receive many thousands of synaptic inputs some excitatory, some inhibitory, and some modulatory. Excitatory synaptic connections are typically found on the major receiving area of the neuron, the dendrite, and most often on spines that project from the dendrite (Figure 13.2). These excitatory synapses have identifiable morphological characteristics and are referred to as Type I (Figure 13.2, box labeled Dendrites). A distinct zone frequently exists in the pre-synaptic terminal of Type I synapses responsible for the release of vesicles containing glutamate and a corresponding zone under the postsynaptic membrane that
Figure 13.1
Figure 13.2See Neuroscience Online for more information

Medical Neuroscience 2013 : Block 1
Page 113Amino Acid Neurotransmitters
Figure 13.3Figure 13.3 i l lustrates the structure of four key amino acid neurotransmitters. Note that the excitatory amino acids carry two negative charges from the two carboxylate groups (COO-, red balls) as opposed to one for the inhibitory amino acids. Recognize that N-methyl-D-Aspartate is a synthetic compound not found in the brain and is technically not a neurotransmitter. It is a highly useful agonist that can mimic the actions of glutamate on a particular subset of glutamate receptors.
serves to anchor the receptors for glutamate (click on the box for details). In addition, vesicles that contain glutamate are small (~50 nm in diameter) and tend to have a spherical appearance.
Inhibitory synapses (like those utilizing glycine and GABA) tend to be localized near the neuronal soma and are referred to as Type II (Figure 13.2, labeled Axosomatic synapse). Morphologically, the synapses again have specializations for release of vesicles and for anchoring receptors. However, the zones of contact tend to be smaller than for excitatory synapses (See Neuroscience Online for more details). For unknown reasons, the vesicles containing glycine or GABA are often elliptical in shape.
Functionally, the location of these synaptic contacts has profound influences on the postsynaptic neuron. In general, the further from the cell body, the more the EPSP is attenuated by the passive properties of the membrane (these potentials are not propagating action potentials; they are synaptic potentials). Therefore, for neurons lacking regenerative processes in their dendrites, EPSPs that are far from the point of action potential generation (the cell soma and axon hillock) attenuate to a greater degree than IPSPs which are generated closer to the neuron’s soma. Due to this spatial arrangement and the relatively small size of each EPSP (1 mV), many distant EPSPs must summate to cause the initiation of an action potential. In contrast, fewer local IPSPs on the cell soma are required to inhibit production of action potentials. On a typical cortical neuron, one might find 10,000 axodendritic excitatory synapses and only 10-50 axosomatic inhibitory synapses.
sTruCTure of amino aCid TransmiTTers
Initially, amino acids were not considered viable candidates for neurotransmitters since they are ubiquitous cellular constituents and are required for protein synthesis. Also, unlike the specific enzymes in neurons that synthesize ACh and catecholamines, enzymes that synthesize glutamate, aspartate and glycine are not unique to neurons. Whereas antibodies to choline acetyltransferase can be used to identify neurons as cholinergic, no such markers are available for neurons that use the amino acids as transmitters. Nevertheless, it is now known that amino acids constitute the major group of substances used for generating excitatory and inhibitory synaptic potentials in the CNS. Amino acids used for synaptic transmission are compartmentalized. For example, glutamate to be used as a neurotransmitter is compartmentalized from metabolic glutamate used for protein synthesis by packaging the transmitter into synaptic vesicles for subsequent Ca2+-dependent release.
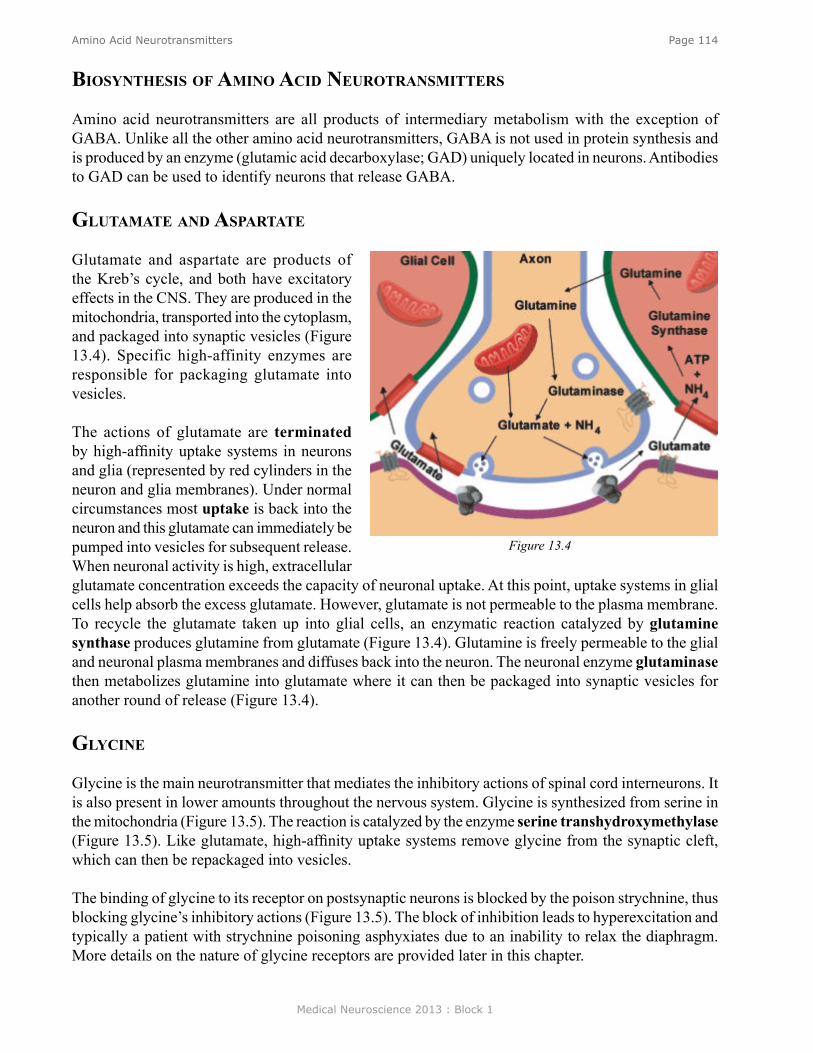
Medical Neuroscience 2013 : Block 1
Page 114Amino Acid Neurotransmitters
biosynThesis of amino aCid neuroTransmiTTers
Amino acid neurotransmitters are all products of intermediary metabolism with the exception of GABA. Unlike all the other amino acid neurotransmitters, GABA is not used in protein synthesis and is produced by an enzyme (glutamic acid decarboxylase; GAD) uniquely located in neurons. Antibodies to GAD can be used to identify neurons that release GABA.
gluTamaTe and asParTaTe
Glutamate and aspartate are products of the Kreb’s cycle, and both have excitatory effects in the CNS. They are produced in the mitochondria, transported into the cytoplasm, and packaged into synaptic vesicles (Figure 13.4). Specific high-affinity enzymes are responsible for packaging glutamate into vesicles.
The actions of glutamate are terminated by high-affinity uptake systems in neurons and glia (represented by red cylinders in the neuron and glia membranes). Under normal circumstances most uptake is back into the neuron and this glutamate can immediately be pumped into vesicles for subsequent release. When neuronal activity is high, extracellular glutamate concentration exceeds the capacity of neuronal uptake. At this point, uptake systems in glial cells help absorb the excess glutamate. However, glutamate is not permeable to the plasma membrane. To recycle the glutamate taken up into glial cells, an enzymatic reaction catalyzed by glutamine synthase produces glutamine from glutamate (Figure 13.4). Glutamine is freely permeable to the glial and neuronal plasma membranes and diffuses back into the neuron. The neuronal enzyme glutaminase then metabolizes glutamine into glutamate where it can then be packaged into synaptic vesicles for another round of release (Figure 13.4).
glyCine
Glycine is the main neurotransmitter that mediates the inhibitory actions of spinal cord interneurons. It is also present in lower amounts throughout the nervous system. Glycine is synthesized from serine in the mitochondria (Figure 13.5). The reaction is catalyzed by the enzyme serine transhydroxymethylase (Figure 13.5). Like glutamate, high-affinity uptake systems remove glycine from the synaptic cleft, which can then be repackaged into vesicles.
The binding of glycine to its receptor on postsynaptic neurons is blocked by the poison strychnine, thus blocking glycine’s inhibitory actions (Figure 13.5). The block of inhibition leads to hyperexcitation and typically a patient with strychnine poisoning asphyxiates due to an inability to relax the diaphragm. More details on the nature of glycine receptors are provided later in this chapter.
Figure 13.4
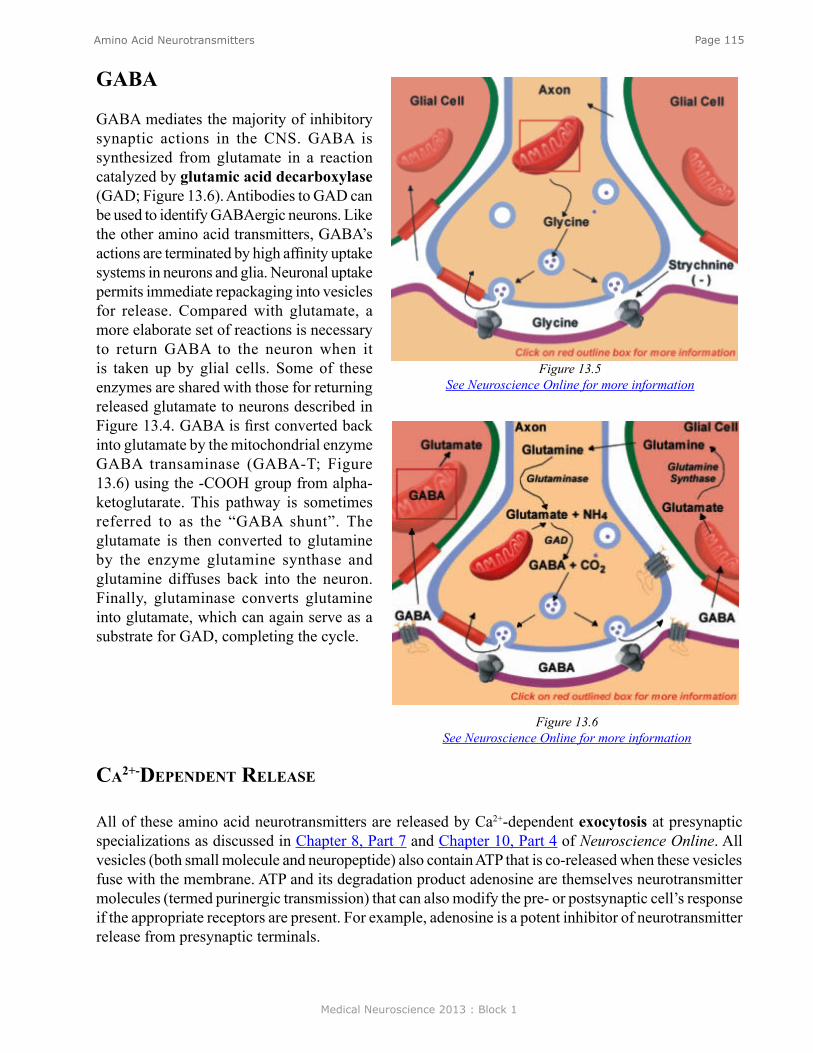
Medical Neuroscience 2013 : Block 1
Page 115Amino Acid Neurotransmitters
gaba
GABA mediates the majority of inhibitory synaptic actions in the CNS. GABA is synthesized from glutamate in a reaction catalyzed by glutamic acid decarboxylase (GAD; Figure 13.6). Antibodies to GAD can be used to identify GABAergic neurons. Like the other amino acid transmitters, GABA’s actions are terminated by high affinity uptake systems in neurons and glia. Neuronal uptake permits immediate repackaging into vesicles for release. Compared with glutamate, a more elaborate set of reactions is necessary to return GABA to the neuron when it is taken up by glial cells. Some of these enzymes are shared with those for returning released glutamate to neurons described in Figure 13.4. GABA is first converted back into glutamate by the mitochondrial enzyme GABA transaminase (GABA-T; Figure 13.6) using the -COOH group from alpha-ketoglutarate. This pathway is sometimes referred to as the “GABA shunt”. The glutamate is then converted to glutamine by the enzyme glutamine synthase and glutamine diffuses back into the neuron. Finally, glutaminase converts glutamine into glutamate, which can again serve as a substrate for GAD, completing the cycle.
Ca2+-dePendenT release
All of these amino acid neurotransmitters are released by Ca2+-dependent exocytosis at presynaptic specializations as discussed in Chapter 8, Part 7 and Chapter 10, Part 4 of Neuroscience Online. All vesicles (both small molecule and neuropeptide) also contain ATP that is co-released when these vesicles fuse with the membrane. ATP and its degradation product adenosine are themselves neurotransmitter molecules (termed purinergic transmission) that can also modify the pre- or postsynaptic cell’s response if the appropriate receptors are present. For example, adenosine is a potent inhibitor of neurotransmitter release from presynaptic terminals.
Figure 13.6See Neuroscience Online for more information
Figure 13.5See Neuroscience Online for more information

Medical Neuroscience 2013 : Block 1
Page 116Amino Acid Neurotransmitters
reCePTors for amino aCid neuroTransmiTTers
Receptors for each of the amino acid neurotransmitters can either directly open an ion channel (ionotropic) or couple to a G-protein (G-protein coupled receptor; GPCR) except for glycine. There is no known GPCR for glycine and all of glycine’s effects are mediated through an ion channel permeable to Cl-. In contrast, glutamate and GABA can produce fast responses by directly opening ion channels and can produce slow responses by activating receptors coupled to G-proteins. Examples of ionotropic receptors and GPCRs are compared in Figures 13.7 and 13.8, respectively.
gluTamaTe reCePTors
There are at least three distinct types of glutamate receptors. Two are ionotropic since glutamate binding directly opens an ion channel and the other is a GPCR, producing alterations in intracellular messengers (Figures 13.7 and 13.8). These three distinct types of glutamate receptors have been characterized by using agonists that specifically activate each type. The agonists and the subset of glutamate receptors they activate are termed
I. NMDA (N-methyl-D-aspartate, a synthetic compound that acts as an agonist for this particular glutamate receptor subtype),
II. non-NMDA (also known as kainate/AMPA receptors since these agonists activate this receptor subtype), and
III. G-protein coupled glutamate receptor.
Ionotropic glutamate receptors open channels that cause the cell to depolarize and are therefore excitatory (driving the membrane potential towards firing an action potential). The reversal potential (near 0 mV) of the EPSP indicates that glutamate opens receptors selectively permeable to cations (Na+, K+, and Ca2+).
Opening of non-NMDA receptors causes the majority of the excitatory postsynaptic potentials (EPSPs) in the nervous system. This receptor is mainly permeable to Na+ and K+ (Figure 13. 9). The structure of non-NMDA receptors loosely resembles the nicotinic ACh receptor, although glutamate receptors have
Figure 13.7 Figure 13.8
See Neuroscience Online for more information
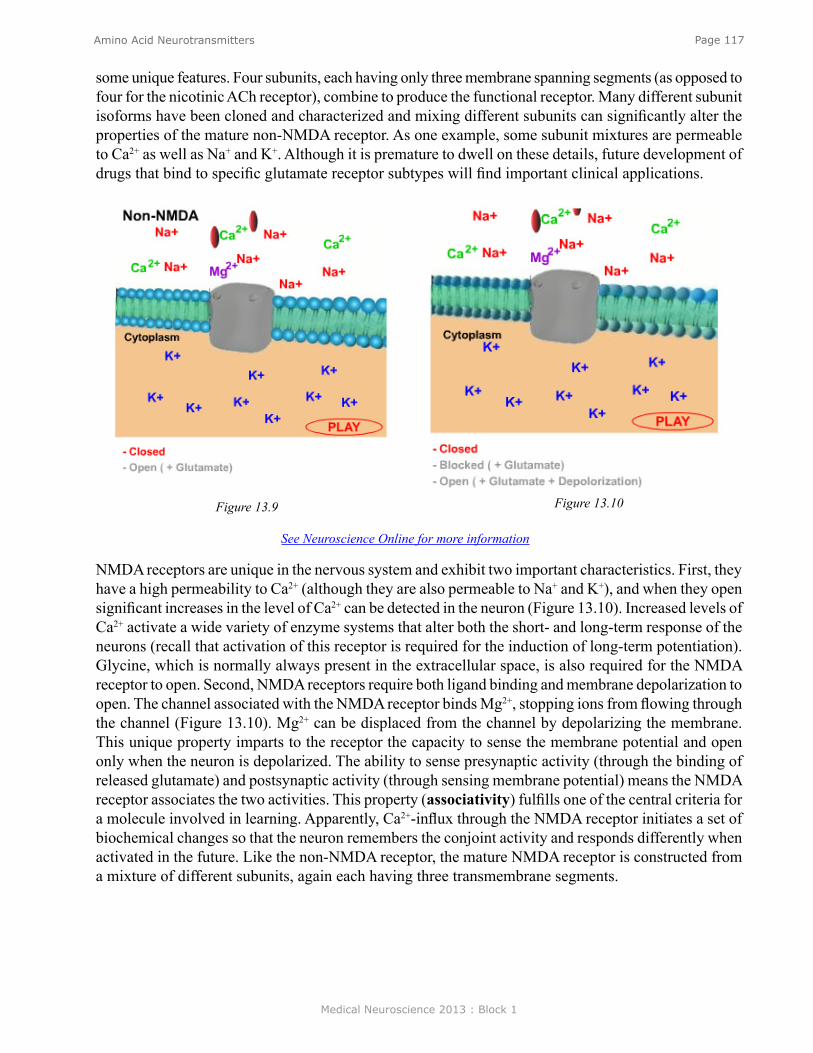
Medical Neuroscience 2013 : Block 1
Page 117Amino Acid Neurotransmitters
some unique features. Four subunits, each having only three membrane spanning segments (as opposed to four for the nicotinic ACh receptor), combine to produce the functional receptor. Many different subunit isoforms have been cloned and characterized and mixing different subunits can significantly alter the properties of the mature non-NMDA receptor. As one example, some subunit mixtures are permeable to Ca2+ as well as Na+ and K+. Although it is premature to dwell on these details, future development of drugs that bind to specific glutamate receptor subtypes will find important clinical applications.
NMDA receptors are unique in the nervous system and exhibit two important characteristics. First, they have a high permeability to Ca2+ (although they are also permeable to Na+ and K+), and when they open significant increases in the level of Ca2+ can be detected in the neuron (Figure 13.10). Increased levels of Ca2+ activate a wide variety of enzyme systems that alter both the short- and long-term response of the neurons (recall that activation of this receptor is required for the induction of long-term potentiation). Glycine, which is normally always present in the extracellular space, is also required for the NMDA receptor to open. Second, NMDA receptors require both ligand binding and membrane depolarization to open. The channel associated with the NMDA receptor binds Mg2+, stopping ions from flowing through the channel (Figure 13.10). Mg2+ can be displaced from the channel by depolarizing the membrane. This unique property imparts to the receptor the capacity to sense the membrane potential and open only when the neuron is depolarized. The ability to sense presynaptic activity (through the binding of released glutamate) and postsynaptic activity (through sensing membrane potential) means the NMDA receptor associates the two activities. This property (associativity) fulfills one of the central criteria for a molecule involved in learning. Apparently, Ca2+-influx through the NMDA receptor initiates a set of biochemical changes so that the neuron remembers the conjoint activity and responds differently when activated in the future. Like the non-NMDA receptor, the mature NMDA receptor is constructed from a mixture of different subunits, again each having three transmembrane segments.
Figure 13.9 Figure 13.10
See Neuroscience Online for more information

Medical Neuroscience 2013 : Block 1
Page 118Amino Acid Neurotransmitters
reCePTors-gabaa and glyCine
GABA and glycine ionotropic receptors are selectively permeable to Cl- (reversal potential near -70 mV). When they open, they cause the neuron to hyperpolarize and therefore drive the membrane potential away from the threshold for firing an action potential. GABA, like glutamate, also binds to and activates a GPCR. In contrast, glycine only binds to ionotropic receptors.
gaba reCePTors
The ionotropic and G-protein coupled metabotropic GABA receptors are referred to as GABAA and GABAB, respectively. Some of the main features of GABAA and GABAB receptors are as follows:
CharaCTerisTiCs of gabaa reCePTor
The GABAA receptor is composed of five subunits that each contain four membrane spanning domains. GABAA subunits are highly related to those of the nicotinic ACh receptor. Important differences exist to produce a channel that permits the permeation of the negatively charged Cl- ion. Specifically, there are positively charged amino acids placed at strategic positions within the channel portion of the receptors that permit Cl- passage. The different subunits of the GABAA receptor are responsible for the binding of different drugs.
IV. GABA binds predominantly to the alpha subunit (Figure 13.11).V. Benzodiazepines (like Valium and Librium) bind to the gamma subunit.
VI. Barbiturates (Phenobarbital and secobarbital) bind to both the alpha and beta subunits.VII. Picrotoxin blocks ion flow through the receptor (Figure 13.11).
The pharmacology of GABAA receptors is complex and clinically important. When GABA is released into the synapse, it binds to a population of the available receptors, but typically not all of them. If benzodiaze-pines are present, the effectiveness of GABA binding to its receptor is increased signifi-cantly. Therefore, effective doses of benzo-diazepines enhance the ability of GABA to hyperpolarize the neuron by increasing the number of GABA receptors that open at a fixed concentration of GABA. Inhibition is produced by increasing the amount of
Table I: GABA ReceptorsGABAA GABAB
Largely Postsynaptic Largely Presynaptic
Opens a Cl- Channel Alters Second Messengers
Rapid Response (15 msec) Slow Response (300-500 msec and longer)
Multisubunit, Binds Modulators Single Subunit
Figure 13.11

Medical Neuroscience 2013 : Block 1
Page 119Amino Acid Neurotransmitters
Cl- that flows into the neuron. Recognize that benzodiazepines themselves do not open the receptor but simply enhance GABA binding. Barbiturates also produce their sedative effects by increasing the effectiveness of GABA binding to its receptor. The naturally occurring toxin called picrotoxin is a potent inhibitor of the GABAA receptor and works by preventing Cl- flow through the receptor (Figure 13.11). glyCine reCePTor
The glycine receptor, like the GABAA receptor also permits the influx of Cl- into neurons and displays a reversal potential near -70 mV. This Cl--permeable glycine receptor can be blocked by the rat poison strychnine. The mature glycine receptor is constructed from mixtures of at least two subunits each of which has four membrane spanning domains.
g-ProTein CouPled gluTamaTe and gabab reCePTors
Glutamate GPCRs are members of a large family of receptors that couple with G proteins to produce their effects. These receptors like those for serotonin, norepinephrine, epinephrine, muscarinic ACh, and dopamine, produce the large majority of their effects through alterations in the activity of metabolic enzymes and not by directly opening ion channels in the membranes. All of these receptors are single polypeptides that span the membrane seven times (See Fig. 12.10 and Fig. 13.8).
The glutamate GPCR’s best known effects are the activation of phospholipase C which generates inositol-trisphosphate (IP3) and diacylglycerol (DAG) from the precursor lipid phosphatidylinositol bisphosphate (See Figure 13.8). Inositol-trisphosphate binds to receptors on intracellular organelles causing the release of Ca2+. Among several other things, increased Ca2+ along with diacylglycerol lead to the activation of protein kinase C which produces a variety of alterations in the enzymatic machinery of the cell including the regulation of ion channels that affect the electrical properties of the neuron.
The GABAB receptor, like the glutamate GPCR, produces its effects not by directly opening ion channels, but by coupling to G-proteins and enzymes that influence metabolites within the neuron. Reported effects include alterations (either increases or decreases) in cAMP levels, increases in K+-conductance, and decreases in Ca2+-conductance. Some of the ion channel effects detected are due to the components of the activated G-protein binding directly to ion channels, influencing their properties (See Figure 6.5, Synaptic Transmission of the Central Nervous System, Neuroscience Online). TerminaTion of aCTion
Two basic mechanisms, diffusion and high affinity uptake, terminate the response to amino acid transmitters. The high affinity uptake mechanism is the most predominant. The proteins involved in transmitter uptake are related and each contains 12 membrane-spanning domains. Transporters use energy derived either from the hydrolysis of ATP or electrochemical ion gradients established across the membrane to pump the transmitters into neurons and glia. The energy-dependent nature of these receptors means that in times of metabolic stress, such as during an ischemic episode, the pumps fail and toxic levels of these transmitters build up.

Medical Neuroscience 2013 : Block 1
Page 120Amino Acid Neurotransmitters
CliniCal manifesTaTions of alTered gluTamaTe levels
The neurotransmitter glutamate is highly toxic to neurons when present for extended periods. One of the best understood clinical conditions involving glutamate is neuronal injury following stroke or trauma. Both events produce massive release of glutamate in the brain that over-stimulates glutamate receptors. The absence of energy prevents the pumps from removing glutamate from the synapse. As a consequence, the uncontrolled opening of glutamate receptors causes a large influx of Na+ followed by water that produces swelling and a large and sustained influx of Ca2+ that leads to hyperactivation of many calcium-dependent enzymes. The Ca2+ influx through the NMDA receptor appears to be one of the keys to producing neuronal damage since specifically blocking activation of this receptor attenuates some of the neuronal injury following stroke. The key to minimizing damage following stroke is well-controlled reestablishment of blood flow so that ATP production is supported and homeostasis is reestablished. Clot breaking agents such as tissue plasminogen activator (tPA) are now used commonly to reestablish blood flow.
Because glutamate is the major excitatory transmitter in the human brain, derangements in glutamate metabolism or receptor activation have been implicated in a wide variety of pathologic conditions. These include diseases such as Alzheimer’s and Huntington’s chorea.
diseases assoCiaTed wiTh gaba
One explanation for the establishment of focal epilepsy is decreased local GABA-mediated inhibition. Many facets of epilepsy can be elicited experimentally by blocking GABA receptors with the toxin picrotoxin previously described. The decrease in GABA inhibition permits cells to fire synchronously, thus producing massive local excitation and initiation of a seizure. Clinically, seizures can often be terminated by inducing a barbiturate coma. High dose barbiturates presumably potentiate GABA’s inhibitory effects, preventing local hyperexcitation by hyperpolarizing the cell membranes.
Mood disorders (generalized anxiety disorder) can also be controlled by drugs which potentiate GABA’s inhibitory activity. Some of the most widely prescribed drugs-benzodiazepines (Librium and Valium)-produce their pharmacological effects by increasing GABA’s ability to hyperpolarize neuronal membranes, thereby quieting the system. This finding suggests that some initial imbalance in the GABAergic system may underlie aspects of this disorder.

Medical Neuroscience 2013 : Block 1
Page 121Neuropeptides and Nitric Oxide
Neuropeptides and Nitric Oxide
assignmenT
I. Peptides as Neurotransmitters and Neuromodulators A. Classification of peptides by familiesB. Cell biology
1. Biosynthesis and regulation2. Release3. Termination of action
C. Receptors - G protein linkedII. Nitric Oxide (NO) - an unconventional extracellular messenger
A. Characteristics of NOB. Synthesis by nitric oxide synthase (NOS) and releaseC. Receptors for NOD. Biological Effects of NO
1. Vasodilator2. Neuromodulator3. Toxicity
major objeCTives
1. Know the characteristics of neuropeptide receptors and how they compare with those of non-peptide neurotransmitters.
2. Know how a neuropeptide is synthesized, released and degraded.3. Know the unique characteristics of NO as a “transmitter”.4. Know how NO is synthesized and what enzymes it influences when released.5. Know the biological effects of NO.
required reading ● Waxham, M.N., Notes on Neuropeptides and Nitric Oxide, Lecture Supplement Volume I; or ● Waxham, M.N., Neuropeptides and Nitric Oxide. Neuroscience Online: http://nba.uth.tmc.
edu/neuroscience/s1/chapter14.html
suggesTed reading ● Deutch, A.Y., Giuffrida, A., and Roberts, J.L. (2009) Nonclassic Signaling in the Brain. In
J.H. Byrne and J.L. Roberts (Eds.), From Molecules to Networks: An Introduction to Cellular and Molecular Neuroscience, Second Edition (pp. 301-319). Academic Press, Elsevier, Inc.
Lecturer: M. Neal Waxham, Ph.D.January 25, 2013 | 8:00 AM

Medical Neuroscience 2013 : Block 1
Page 122Neuropeptides and Nitric Oxide
In addition, many neuropeptides do not produce an obvious electrophysiological change in the postsynaptic neuron. For example, an action potential in neuron 3 (Figure 14.1; pink) produces no response in the postsynaptic neuron even though one can confirm through other means that neuropeptides were released. However, changes have occurred in the postsynaptic neuron as revealed in the example shown in the panels labeled "C". If one first measures the glutamate response (C1) by stimulating neuron 1, a typical response is seen. Stimulation of neuron 3 produces no response (C2) as before. However, if neuron 1 is stimulated again after the stimulation of neuron 3, the EPSP measured in the postsynaptic neuron is
inTroduCTion
Classical neurotransmitters (e.g., glutamate and ACh) work by the directional transfer of information between presynaptic release sites and postsynaptic clusters of receptors. The process is generally well organized spatially with some, but very limited, "spill-over" of transmitter between neighboring synapses. The situation is quite different for transmission mediated by neuropeptides and nitric oxide (NO). These molecules ignore directionality of information flow and instead influence a volume of area from their point of release. This feature is loosely analogous to the release of peptide hormones into the blood stream, which can be carried to distant sites to produce their effects. For example, NO can be produced in a postsynaptic neuron and diffuse back to the presynaptic neuron or to other neighboring neurons affecting the subsequent release of transmitter. This property is termed retrograde transmission, since the signal travels in the opposite direction (post- to pre-) as compared to the classical directionality of neurotransmission.
neuroPePTides
Many neuropeptides were originally described as hormones (e.g., somatostatin and cholecystokinin), although once characterized they were subsequently also found in neurons within the CNS. Neuropeptides typically produce neuronal responses with slow onset and long duration (Figure 14.1), and every characterized neuropeptide receptor falls into the GPCR (G-protein coupled receptor) class. For comparison, Figure 14.1 shows typical responses produced by the action of a classical neurotransmitter (glutamate) acting at an ionotropic receptor and a neuropeptide. An action potential elicited in neuron 1 (in green), a glutamate releasing neuron, produces the fast (5 msec) EPSP typical of glutamate mediated synaptic transmission. In contrast, an action potential elicited in neuron 2 (purple) that releases neuropeptide produces a slow onset (5 sec to peak) and long-duration (10 sec) EPSP. These slow EPSPs are typical of neuropeptide actions since all neuropeptide receptors are G-protein coupled.
Figure 14.1

Medical Neuroscience 2013 : Block 1
Page 123Neuropeptides and Nitric Oxide
Table INeuropeptide Families
Tachykinins: substance P, bombesin, substance Insulins: insulin, insulin-like growth factors Somatostatins: somatostatin, pancreatic polypeptide Gastrins: gastrin, cholecystokinin Opioids: opiocortins, enkephalins, dynorphin
For example, opiates are grouped as a family due to the identical amino acid sequence Tyr-Gly-Gly-Phe--(Table II) found at the N-terminus of each.
Table II Opiate Family-Sequence Identity in Italics
Name Amino Acid Sequence Leu-enkephalin Tyr-Gly-Gly-Phe-Leu-OHMet-enkephalin Tyr-Gly-Gly-Phe-Met-OH
Beta-endorphin
Tyr-Gly-Gly-Phe-Met-Thr-Ser-Glu-Lys-Ser-Gln-Thr-Pro-Leu-Val-Thr-Leu-
Phe-Lys-Asn-Ala-Ile-Val-Lys-Asn-Ala-His-Lys-Gly-Gln-His-OH
Dynorphin Tyr-Gly-Gly-Phe-Leu-Arg-Arg-Ile-Arg- Pro-Lys-Leu-Lys-Trp-Asp-Asn-Gln-OH
enhanced (C3) (the original EPSP is the solid line and the EPSP following neuron 3 stimulation is the dashed line). Thus, the neuropeptide released from neuron 3 altered the postsynaptic neuron's response to another neurotransmitter. These neuropeptides are said to be neuromodulators since their effects are to potentiate or depress the effects of a second transmitter. Other examples of such heterosynaptic plasticity are described in Chapter 7, Part 2 of Neuroscience Online.
ClassifiCaTion of PePTides by families
Neuropeptides can be grouped into families based on similarities in their amino acid sequences (Tables I and II).
biosynThesis and regulaTion
Neuropeptides are derived from larger precursors by proteolytic processing. They are all initially synthesized within the soma (Figure 14.2).
Precursors are initially formed by translation on polyribosomes attached to the endoplasmic reticulum near the cell body. From there, they progress through the Golgi apparatus where further modifications take place, including glycosylation. They are then packaged into secretory granules that are transported to terminals by fast axonal transport. Fast is a relative term, however, and due to the relatively long distances that some neuropeptides must travel. Replenishing the pool of releasable neuropeptide might require many hours. During the transport to the nerve terminal, proteases that are packaged within the vesicle begin to cleave the precursor neuropeptide into its final mature form. This processing is essential for the activation of the neuropeptides since the precursors are biologically inactive. At least three types of processing occur within the vesicles (Figure 14.2 , B, C). First, an endopeptidase cleaves the
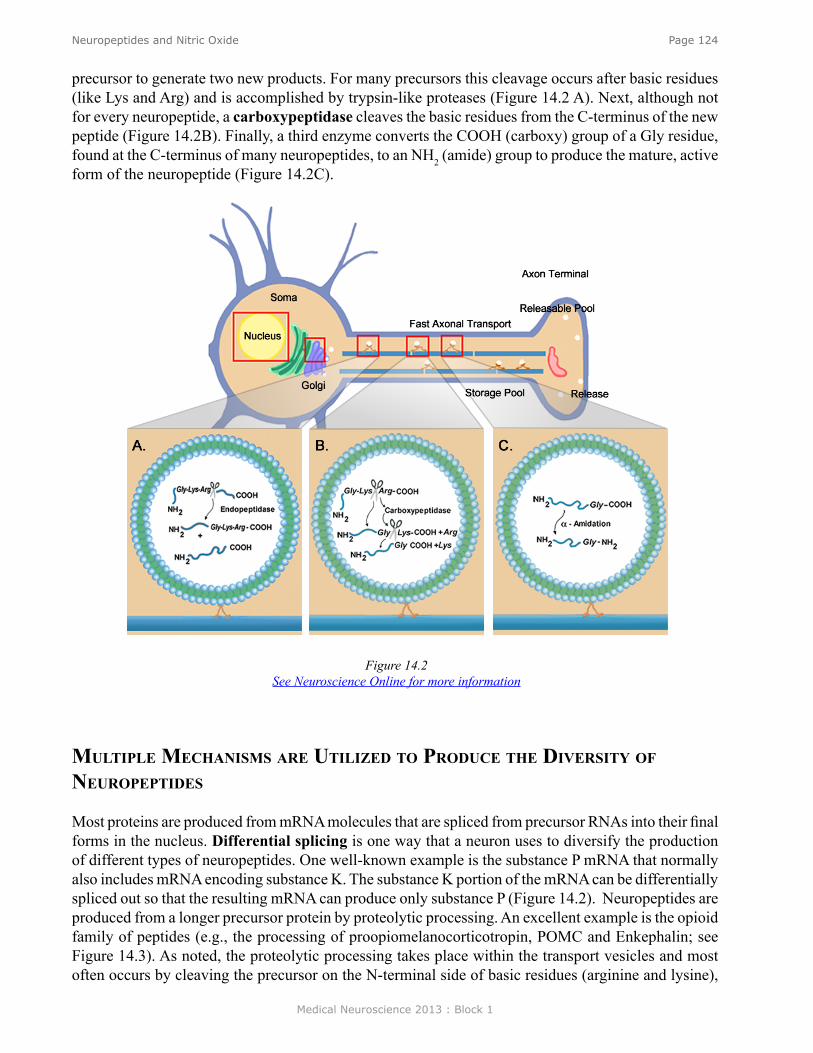
Medical Neuroscience 2013 : Block 1
Page 124Neuropeptides and Nitric Oxide
mulTiPle meChanisms are uTilized To ProduCe The diversiTy of neuroPePTides
Most proteins are produced from mRNA molecules that are spliced from precursor RNAs into their final forms in the nucleus. Differential splicing is one way that a neuron uses to diversify the production of different types of neuropeptides. One well-known example is the substance P mRNA that normally also includes mRNA encoding substance K. The substance K portion of the mRNA can be differentially spliced out so that the resulting mRNA can produce only substance P (Figure 14.2). Neuropeptides are produced from a longer precursor protein by proteolytic processing. An excellent example is the opioid family of peptides (e.g., the processing of proopiomelanocorticotropin, POMC and Enkephalin; see Figure 14.3). As noted, the proteolytic processing takes place within the transport vesicles and most often occurs by cleaving the precursor on the N-terminal side of basic residues (arginine and lysine),
Figure 14.2See Neuroscience Online for more information
precursor to generate two new products. For many precursors this cleavage occurs after basic residues (like Lys and Arg) and is accomplished by trypsin-like proteases (Figure 14.2 A). Next, although not for every neuropeptide, a carboxypeptidase cleaves the basic residues from the C-terminus of the new peptide (Figure 14.2B). Finally, a third enzyme converts the COOH (carboxy) group of a Gly residue, found at the C-terminus of many neuropeptides, to an NH2 (amide) group to produce the mature, active form of the neuropeptide (Figure 14.2C).
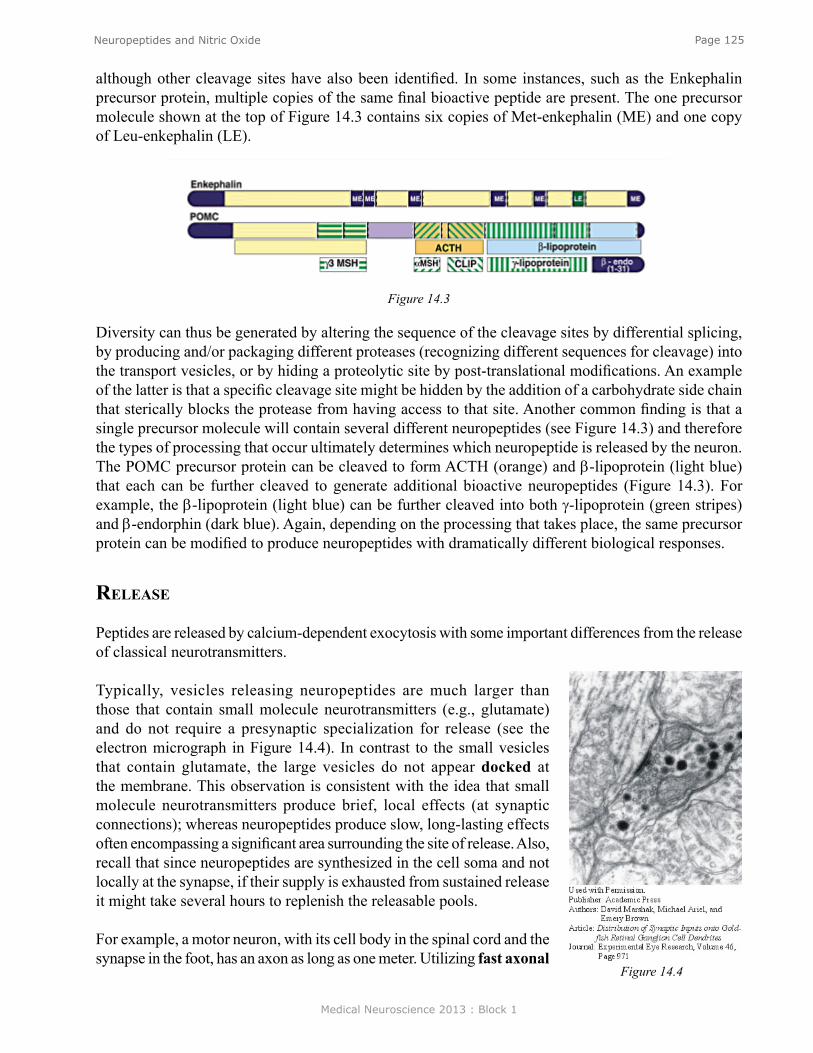
Medical Neuroscience 2013 : Block 1
Page 125Neuropeptides and Nitric Oxide
Figure 14.3
Diversity can thus be generated by altering the sequence of the cleavage sites by differential splicing, by producing and/or packaging different proteases (recognizing different sequences for cleavage) into the transport vesicles, or by hiding a proteolytic site by post-translational modifications. An example of the latter is that a specific cleavage site might be hidden by the addition of a carbohydrate side chain that sterically blocks the protease from having access to that site. Another common finding is that a single precursor molecule will contain several different neuropeptides (see Figure 14.3) and therefore the types of processing that occur ultimately determines which neuropeptide is released by the neuron. The POMC precursor protein can be cleaved to form ACTH (orange) and β-lipoprotein (light blue) that each can be further cleaved to generate additional bioactive neuropeptides (Figure 14.3). For example, the β-lipoprotein (light blue) can be further cleaved into both γ-lipoprotein (green stripes) and β-endorphin (dark blue). Again, depending on the processing that takes place, the same precursor protein can be modified to produce neuropeptides with dramatically different biological responses.
release
Peptides are released by calcium-dependent exocytosis with some important differences from the release of classical neurotransmitters.
Typically, vesicles releasing neuropeptides are much larger than those that contain small molecule neurotransmitters (e.g., glutamate) and do not require a presynaptic specialization for release (see the electron micrograph in Figure 14.4). In contrast to the small vesicles that contain glutamate, the large vesicles do not appear docked at the membrane. This observation is consistent with the idea that small molecule neurotransmitters produce brief, local effects (at synaptic connections); whereas neuropeptides produce slow, long-lasting effects often encompassing a significant area surrounding the site of release. Also, recall that since neuropeptides are synthesized in the cell soma and not locally at the synapse, if their supply is exhausted from sustained release it might take several hours to replenish the releasable pools.
For example, a motor neuron, with its cell body in the spinal cord and the synapse in the foot, has an axon as long as one meter. Utilizing fast axonal
although other cleavage sites have also been identified. In some instances, such as the Enkephalin precursor protein, multiple copies of the same final bioactive peptide are present. The one precursor molecule shown at the top of Figure 14.3 contains six copies of Met-enkephalin (ME) and one copy of Leu-enkephalin (LE).
Figure 14.4

Medical Neuroscience 2013 : Block 1
Page 126Neuropeptides and Nitric Oxide
transport it would potentially take more than a day for a newly synthesized neuropeptide to arrive at this synapse from the soma. It should also be evident that endogenous pain-killing neuropeptides, like beta-endorphin, could be “used-up” in times of persistent stimulation leading to situations where pain can no longer be controlled by endogenous mechanisms.
A typical mature neuron will often release one small molecule neurotransmitter and one or more neuropeptides (as in the example shown in Figure 14.4). If more than one neuropeptide is released they most often come from the same single precursor molecule. An example is the co-release of both ACh and calcitonin gene-related peptide from spinal motor neurons. CGRP activates adenylate cyclase, raising cAMP levels, and potentiates the force of contraction produced by ACh activation of the nicotinic ACh receptor. In this case, the neuropeptide is modulatory as described in Figure 14.1. However, in this instance, the effect potentiates muscle contraction instead of increasing the magnitude of the EPSP. In both examples, the potentiated response is due to increased sensitivity of the system to a constant amount of released neurotransmitter.
TerminaTion of aCTion
Neuropeptides are slowly removed from the extracellular space; a feature which also contributes to their relatively long lasting effects. Inactivation occurs by both diffusion and breakdown by extracellular proteases. No evidence has been found for peptide re-uptake as a means of terminating their action.
reCePTors are all g-ProTein linked
All known neuropeptide receptors produce their effects by altering the levels of intracellular second messengers. These receptors are seven typical transmembrane spanning proteins that are linked through G proteins (GPCRs) to alter the activation of other cellular enzymes. This property is consistent with neuropeptides inducing a slower response and is well suited for a modulatory role. One important distinction between small and neuropeptide molecule transmitters is that neuropeptide receptors have a high affinity for binding (nanomolar) as opposed to micro- or millimolar affinities measured for small molecule neurotransmitters (like glutamate). As neuropeptides are not released directionally into the confined volume of a synapse, their concentrations do not achieve very high levels and the receptors then must have high affinities to react to these small concentrations. This high affinity slows the dissociation of the neuropeptide from its receptor and also contributes to the persistent effects of these molecules.
niTriC oxide (no)
Nitric oxide has recently gained widespread attention as the founding member of a new class of gaseous messenger molecules. NO is the active molecule that sublingual nitroglycerin produces to increase vasodilation in the relief of angina. Other important biological effects of NO are now recognized. For example, in the nervous system, NO is important in the regulation of cerebral blood flow, in the modulation of neurotransmission, and in toxicity associated with various pathologic states.
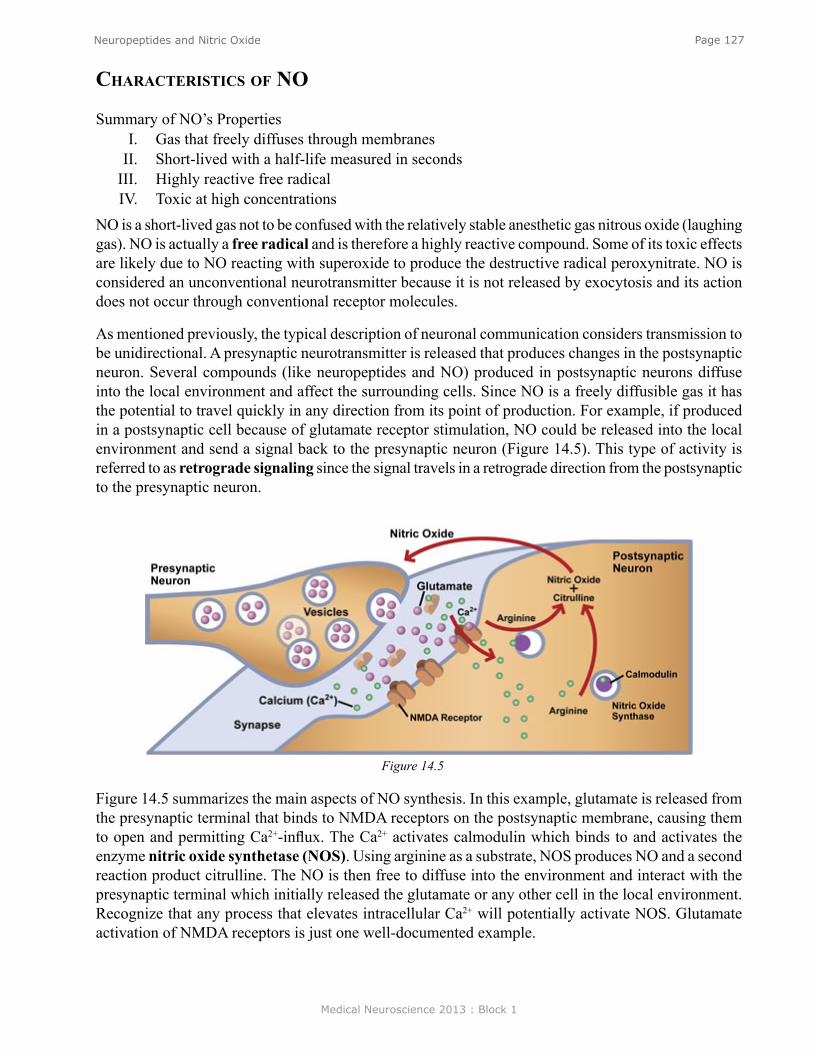
Medical Neuroscience 2013 : Block 1
Page 127Neuropeptides and Nitric Oxide
Figure 14.5
CharaCTerisTiCs of no
Summary of NO’s PropertiesI. Gas that freely diffuses through membranes
II. Short-lived with a half-life measured in secondsIII. Highly reactive free radicalIV. Toxic at high concentrations
NO is a short-lived gas not to be confused with the relatively stable anesthetic gas nitrous oxide (laughing gas). NO is actually a free radical and is therefore a highly reactive compound. Some of its toxic effects are likely due to NO reacting with superoxide to produce the destructive radical peroxynitrate. NO is considered an unconventional neurotransmitter because it is not released by exocytosis and its action does not occur through conventional receptor molecules.
As mentioned previously, the typical description of neuronal communication considers transmission to be unidirectional. A presynaptic neurotransmitter is released that produces changes in the postsynaptic neuron. Several compounds (like neuropeptides and NO) produced in postsynaptic neurons diffuse into the local environment and affect the surrounding cells. Since NO is a freely diffusible gas it has the potential to travel quickly in any direction from its point of production. For example, if produced in a postsynaptic cell because of glutamate receptor stimulation, NO could be released into the local environment and send a signal back to the presynaptic neuron (Figure 14.5). This type of activity is referred to as retrograde signaling since the signal travels in a retrograde direction from the postsynaptic to the presynaptic neuron.
Figure 14.5 summarizes the main aspects of NO synthesis. In this example, glutamate is released from the presynaptic terminal that binds to NMDA receptors on the postsynaptic membrane, causing them to open and permitting Ca2+-influx. The Ca2+ activates calmodulin which binds to and activates the enzyme nitric oxide synthetase (NOS). Using arginine as a substrate, NOS produces NO and a second reaction product citrulline. The NO is then free to diffuse into the environment and interact with the presynaptic terminal which initially released the glutamate or any other cell in the local environment. Recognize that any process that elevates intracellular Ca2+ will potentially activate NOS. Glutamate activation of NMDA receptors is just one well-documented example.
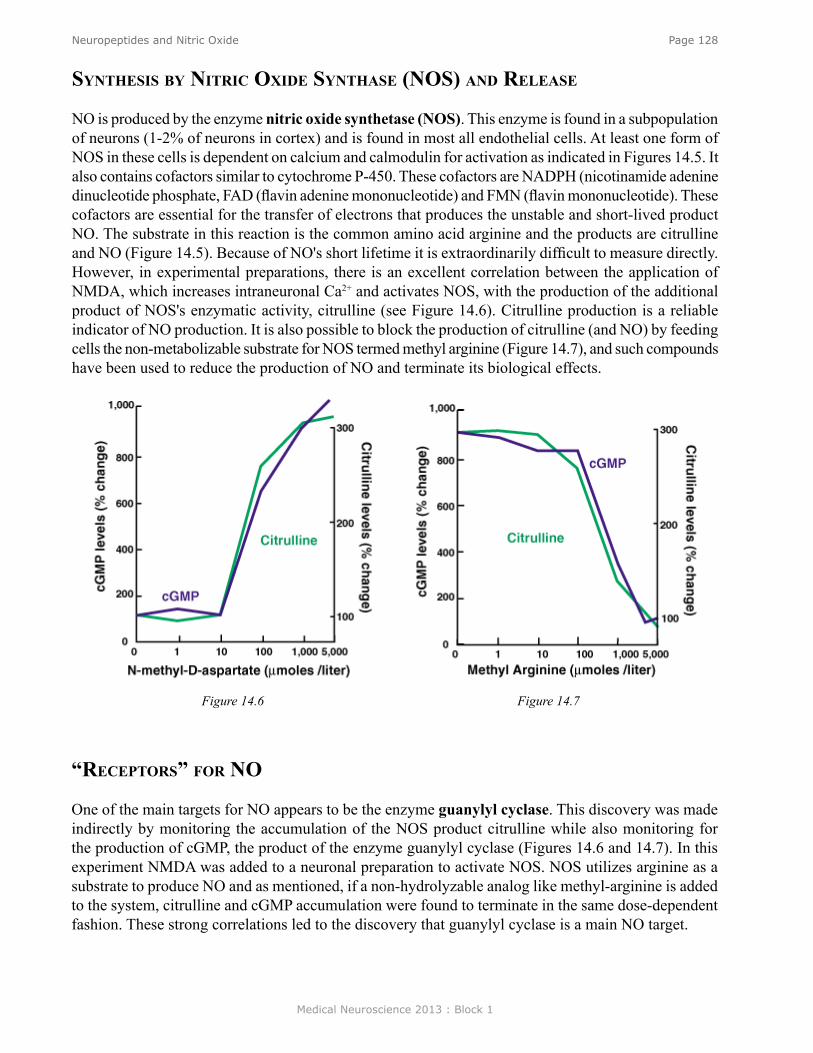
Medical Neuroscience 2013 : Block 1
Page 128Neuropeptides and Nitric Oxide
“reCePTors” for no
One of the main targets for NO appears to be the enzyme guanylyl cyclase. This discovery was made indirectly by monitoring the accumulation of the NOS product citrulline while also monitoring for the production of cGMP, the product of the enzyme guanylyl cyclase (Figures 14.6 and 14.7). In this experiment NMDA was added to a neuronal preparation to activate NOS. NOS utilizes arginine as a substrate to produce NO and as mentioned, if a non-hydrolyzable analog like methyl-arginine is added to the system, citrulline and cGMP accumulation were found to terminate in the same dose-dependent fashion. These strong correlations led to the discovery that guanylyl cyclase is a main NO target.
Figure 14.6 Figure 14.7
synThesis by niTriC oxide synThase (nos) and release
NO is produced by the enzyme nitric oxide synthetase (NOS). This enzyme is found in a subpopulation of neurons (1-2% of neurons in cortex) and is found in most all endothelial cells. At least one form of NOS in these cells is dependent on calcium and calmodulin for activation as indicated in Figures 14.5. It also contains cofactors similar to cytochrome P-450. These cofactors are NADPH (nicotinamide adenine dinucleotide phosphate, FAD (flavin adenine mononucleotide) and FMN (flavin mononucleotide). These cofactors are essential for the transfer of electrons that produces the unstable and short-lived product NO. The substrate in this reaction is the common amino acid arginine and the products are citrulline and NO (Figure 14.5). Because of NO's short lifetime it is extraordinarily difficult to measure directly. However, in experimental preparations, there is an excellent correlation between the application of NMDA, which increases intraneuronal Ca2+ and activates NOS, with the production of the additional product of NOS's enzymatic activity, citrulline (see Figure 14.6). Citrulline production is a reliable indicator of NO production. It is also possible to block the production of citrulline (and NO) by feeding cells the non-metabolizable substrate for NOS termed methyl arginine (Figure 14.7), and such compounds have been used to reduce the production of NO and terminate its biological effects.
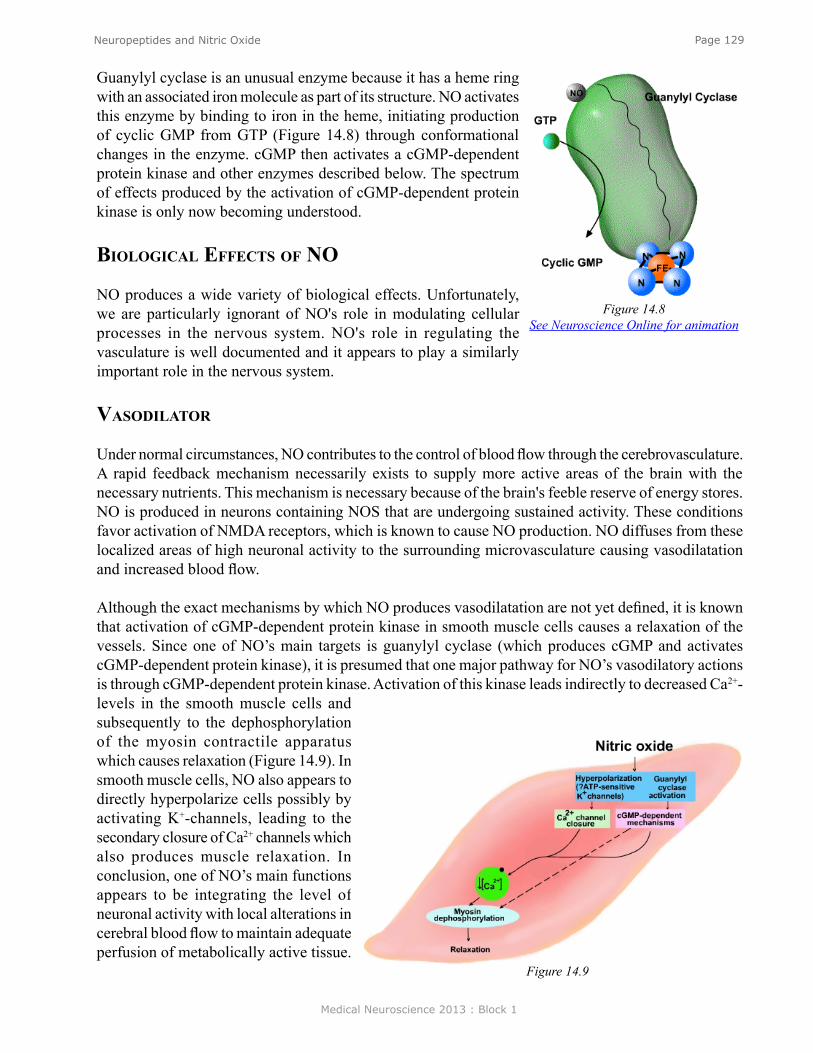
Medical Neuroscience 2013 : Block 1
Page 129Neuropeptides and Nitric Oxide
Guanylyl cyclase is an unusual enzyme because it has a heme ring with an associated iron molecule as part of its structure. NO activates this enzyme by binding to iron in the heme, initiating production of cyclic GMP from GTP (Figure 14.8) through conformational changes in the enzyme. cGMP then activates a cGMP-dependent protein kinase and other enzymes described below. The spectrum of effects produced by the activation of cGMP-dependent protein kinase is only now becoming understood.
biologiCal effeCTs of no
NO produces a wide variety of biological effects. Unfortunately, we are particularly ignorant of NO's role in modulating cellular processes in the nervous system. NO's role in regulating the vasculature is well documented and it appears to play a similarly important role in the nervous system.
vasodilaTor
Under normal circumstances, NO contributes to the control of blood flow through the cerebrovasculature. A rapid feedback mechanism necessarily exists to supply more active areas of the brain with the necessary nutrients. This mechanism is necessary because of the brain's feeble reserve of energy stores. NO is produced in neurons containing NOS that are undergoing sustained activity. These conditions favor activation of NMDA receptors, which is known to cause NO production. NO diffuses from these localized areas of high neuronal activity to the surrounding microvasculature causing vasodilatation and increased blood flow.
Although the exact mechanisms by which NO produces vasodilatation are not yet defined, it is known that activation of cGMP-dependent protein kinase in smooth muscle cells causes a relaxation of the vessels. Since one of NO’s main targets is guanylyl cyclase (which produces cGMP and activates cGMP-dependent protein kinase), it is presumed that one major pathway for NO’s vasodilatory actions is through cGMP-dependent protein kinase. Activation of this kinase leads indirectly to decreased Ca2+-levels in the smooth muscle cells and subsequently to the dephosphorylation of the myosin contractile apparatus which causes relaxation (Figure 14.9). In smooth muscle cells, NO also appears to directly hyperpolarize cells possibly by activating K+-channels, leading to the secondary closure of Ca2+ channels which also produces muscle relaxation. In conclusion, one of NO’s main functions appears to be integrating the level of neuronal activity with local alterations in cerebral blood flow to maintain adequate perfusion of metabolically active tissue.
Figure 14.8See Neuroscience Online for animation
Figure 14.9

Medical Neuroscience 2013 : Block 1
Page 130Neuropeptides and Nitric Oxide
neuromodulaTor
NO is also thought to act as a locally diffusible messenger. It is produced by any action that elevates Ca2+ in cells containing NOS, such as glutamate stimulation of NMDA receptors. Through subsequent activation of guanylyl cyclase and production of cGMP, NO production influences a variety of secondary processes. These include direct modulation of ion channels, stimulation of cGMP-dependent protein kinase, and both up-regulation or down-regulation of cAMP-phosphodiesterase. Downstream effects are then numerous and include up and down regulation of Ca2+ channels, increased excitability (increases neuronal firing rate), increased or decreased neurotransmitter release, and changes in neuron morphology.
ToxiCiTy
NO in excess is toxic to cells. However, a paradox exists for NO toxicity. Apparently, cells that produce high levels of NO are resistant to its toxic effects. For example, NO toxicity is used by macrophages and neutrophils as a mechanism to kill tumor cells and bacteria. However, neither cell type producing NO is susceptible to its damaging effects. This finding is also true for neurons in the central nervous system. Excess glutamate induces neurotoxicity in the brain and is thought to be the primary cause of neuronal death in diseases such as Huntington's or Alzheimer's or after acute stroke or trauma. Excess production of NO is thought to play some role in this neuronal loss due to its toxicity when produced in excess. Interestingly, cells that stain positive for the enzyme NOS are spared in degenerating areas of the brain affected by these diseases. The resistance of these cells appears to be similar to the resistance of the immune cells described above.

Medical Neuroscience 2013 : Block 1
Page 131Principles of Structural Neuroimaging
Principles of Structural Neuroimaging
assignmenT
I. Introduction to NeuroimagingII. CT — X-Ray Computed Tomography
III. MRI — Magnetic Resonance ImagingIV. PET and SPECT — Positron Emission Tomography and Single Photon Emission
Computed TomographyV. Functional Neuroimaging
A. Functional MRI (fMRI) B. Magnetoencephalography (MEG)
VI. Examples of Images from Each Technique
major objeCTives
1. Identify the major neuroimaging modalities.2. Understand the basic principles underlying each modality.3. Differentiate modalities that image the structure versus function of the brain.
referenCes
● Suetens, P. (2002). Fundamentals of Medical Imaging. Cambridge, UK: Cambridge University Press.
Lecturer: Michael Beauchamp, Ph.D.January 25, 2013 | 9:00 AM
Material prepared by Rebecca Billingsley-Marshall, Ph.D.

Medical Neuroscience 2013 : Block 1
Page 132Principles of Structural Neuroimaging
inTroduCTion To neuroimaging
Advances in imaging technologies have improved clinical diagnosis and surgical planning by mak-ing it possible to produce detailed images of brain structures. Newer digital image acquisition and processing techniques have also allowed for the imaging of brain function, in addition to its detailed structure. This outline provides an overview of the most common structural and functional brain imaging techniques, which include computed tomography (CT), magnetic resonance imaging (MRI), positron emission tomography (PET), single photon emission tomography (SPECT), and magnetoen-cephalography (MEG). Each technique provides different kinds of information about brain structure and function. Basic background information and a summary of the primary clinical uses of each technique are discussed.
x-ray ComPuTed TomograPhy (CT)
Background. Computed tomography, or CT, is an imaging modality that produces cross-sectional images of the brain based on the x-ray attenuation properties of tissues being imaged. X-rays are electromagnetic waves consisting of photons which, when accelerated, release energy. These waves interact with biological tissue in different ways; CT is a recording of the attenuation of these waves as they encounter biological tissue.
Imaging Procedure. During a CT scan, a set of lines is scanned over a given field of view using a thin x-ray beam. This process is repeated for a large number of different angles so that attenuation at each point in a given field can be reconstructed. The attenuation of each beam is determined by a set of detectors that record the angle of the beam as it exits the body. The information collected by these detectors is converted into digital data and processed with a computer for image reconstruction.
The degree of attenuation depends on the tissue density: very dense tissue, like bone, has a high de-gree of attenuation, whereas grey matter and fluid have lower attenuation coefficients. The attenua-tion of each beam at a given point is represented by a gray level transformation of the numeric value of the attenuation. A two-dimensional image is reconstructed from multiple projections of the slice taken at different views. 3-D objects are reconstructed from multiple 2-D slices; each slice consists of a matrix of attenuation coefficients corresponding to the different tissues in the slice. Contrasts in a given image are therefore due to differences in attenuation values between tissues.
Clinical Use. CT is commonly used to obtain anatomical images of the head and neck. Contrast agents are often used in CT for better visualization of increased or decreased vascularization of tis-sues. Radiation doses are relatively high in CT, about 10 to 100 times higher than a radiographic im-age of the same region. CT is likely to remain the preferred imaging modality for the visualization of the skeleton and calcifications. Other investigations are likely to be replaced by MRI. CT will remain, however, the alternative for patients who are not able to enter the MR environment because of metallic implants and electrical stimulators.
magneTiC resonanCe imaging (mri)
Background. MRI is used to obtain images of the brain without using ionizing radiation. It relies on the atomic properties of hydrogen, a common element in biological tissues. Each proton of a hydro-gen atom has a natural property of spinning around its axis. The orientation of the spin of any set of

Medical Neuroscience 2013 : Block 1
Page 133Principles of Structural Neuroimaging
protons in the body is random, making the net magnetization equal to zero.
However, in the presence of a static magnetic field B, such as the field of an MRI scanner, the axes of the spins change orientation and align with that field. The number of protons that align with B in a parallel manner is larger than the number of protons that align in an antiparallel manner, which results in a net magnetization vector parallel to the B field.
The magnetization vector of each proton in static magnetic field B, which is its magnetic strength in a particular direction, has two components: a longitudinal and a transverse component. The longitu-dinal component is parallel to static field B, whereas the transverse component is at a right angle to the longitudinal component. The net magnetization of a set of protons is the sum of their longitudi-nal and transverse components.
The MRI signal. In order to generate an electromagnetic signal that can be detected at the surface of the head in the MRI environment, an additional magnetic field is applied. This field, which pulsates at a high frequency, is called a Radio Frequency (RF) pulse and is delivered along the transverse plane, at a right angle to the static field B. This pulse adds energy to the spinning and precessing protons, and changes their net magnetization. When the RF stops, the protons begin to return to their previous state, resulting in an emission of energy and a recovery of the longitudinal magnetization. This emission of energy is what is observed as the MRI signal; the same coil used to apply the RF pulse is used to record the transverse field.
MRI contrast. The strength of the MRI signal depends primarily on three parameters: the proton den-sity in a tissue (the higher the density of protons, the larger the signal), T1, and T2. T1 is the spin-lattice relaxation time, which is the process by which the spins return to their preferred lower energy state, and the longitudinal component of the net magnetization grows toward its equilibrium value. T2 is the spin-spin relaxation time, or the exponential decay of the transverse magnetization.
The contrast between brain tissues depends upon how these 3 parameters differ in the tissues. For most "soft" tissues in the body, the proton density does not vary dramatically and therefore does not contribute much to signal differences observed in an image. T1 and T2, on the other hand, often differ dramatically between tissues and are primarily responsible for the contrasts observed between soft tissues.
Clinical Use. In clinical practice the MRI signal is manipulated to allow the user to acquire images in which T1, T2, or proton density dominantly influences the gray values displayed on an image. The dependence of the image on these characteristics is influenced by the length and timing of the RF pulses. Obtaining multiple sequences of the same region allows the user to better characterize normal and diseased tissue. Contrast agents are also used in MRI for better visualization. Two com-mon contrast agents are gadolinium compounds and iron chloride.
In addition to the acquisition of structural images, several MRI techniques are used to assess dy-namic characteristics of the brain, including indices of blood flow (MR Angiography), biochemical changes (MR Spectroscopy), and neuronal function (Functional MRI). These are briefly described below.
MRA–MR Angiography (Blood Flow Imaging): MRA is based on the sensitivity of MRI signal to

Medical Neuroscience 2013 : Block 1
Page 134Principles of Structural Neuroimaging
motion of spins. It uses signal changes resulting from flowing blood to quantify flow direction, velocity, and volume, and to visualize differences in vascular structures. MRA better displays the vascular structures without superimposition of the brain (see example in Section VI).
MRS – Magnetic Resonance Spectroscopy: MRS is based on the different resonant frequencies of chemical compounds. MRS is used for the serial monitoring of biochemical and metabolic changes in conditions such as brain tumors, multiple sclerosis, and temporal lobe epilepsy. Commonly used proton spectra in the brain include N-acetyl aspartate (NAA), Creatine (Cr), and lactate (not normal-ly found in the brain; marker of non-oxidative metabolism).
PosiTron emission TomograPhy (PeT) and single PhoTon emission ComPuTed TomograPhy (sPeCT)
Background. PET and SPECT measure emissions from radioactively labeled tracer molecules that have been injected into the bloodstream and carried to the brain. A tracer molecule carries an unsta-ble isotope, called a radionuclide. The molecule becomes part of the metabolic process in the body and produces gamma rays, which allow for the measurement of the tracer molecule in the brain as a function of position (space) and time.
PET. PET studies use a cyclotron to produce radioactive tracers. Positron emitting isotopes that are created include Oxygen-15, Nitrogen-13, Carbon-11, and Fluorine-18, which can combine with other elements to create organic molecules that may substitute for natural substances, such as water, glu-cose, or different neurochemicals (often derived from drugs that target neurotransmitter pathways). Each emitted particle, called a positron, interacts with electrons and produces photons which can be detected at the surface of the head. In the PET scanner, two of the detectors form a ring around the patient’s head and detect photon pairs (coincidence detection resulting from positron annihilation). The reconstructed image depicts the location and concentration of the radioisotope within a plane of the brain. Using different compounds, PET can show blood flow, oxygen and glucose metabolism, and drug concentrations in brain tissues.
SPECT. Similar to PET, SPECT also uses radioactive tracers, but it involves the detection of indi-vidual photons emitted from the radionuclide. The SPECT scanner uses two or three scintillation cameras that rotate around the patient’s head to record data at different angles. Typical radionuclides used in SPECT include Technetium-99m, Iodine-123, and Thallium-201. SPECT tracers are gener-ally more limited than PET tracers in the kinds of brain activity they can monitor, but they are longer lasting. For this reason, SPECT does not require an onsite cyclotron. PET has greater spatial reso-lution than SPECT, particularly for subcortical structures, but SPECT is much less expensive than PET.
Clinical Use: PET and SPECT are used to detect local metabolism in brain structures. A successful tracer for assessing glucose metabolism is 18FDG (fluoro-deoxy-glucose). FDG accumulates propor-tionally to glucose consumption, which can be considerably higher in some tumors than in surround-ing tissue. Oxygen-15 is commonly used to trace oxygen consumption in PET studies of cognitive function. In drug abuse research, PET and SPECT are is used to identify the sites where drugs and neurotransmitters act, to show how quickly drugs reach neural receptors, and to monitor how long drugs occupy these receptors.

Medical Neuroscience 2013 : Block 1
Page 135Principles of Structural Neuroimaging
CT Scan
T1-weighted MRI scan and T2-weighted MRI scan
funCTional neuroimaging
Functional MRI: Brain tissue relaxation is dependent on the oxygenation level of the blood. This dependence allows for the ability to observe brain function with MRI. Via hemoglobin, the vascular system delivers greater amounts of oxygen to brain cells as their metabolic needs increase. Oxyhe-moglobin is carried by the arteries and, at the capillary level, the hemoglobin molecule delivers part of its oxygen to the neurons and becomes deoxyhemoglobin. Oxyhemoglobin is diamagnetic, where-as doxyhemoglobin is a paramagnetic substance that produces magnetic field inhomogeneities that decrease the transverse relation time of the blood and surrounding tissue. This is called the BOLD (blood oxygenation-level dependent) phenomenon, on which functional MRI (fMRI) is based. Func-tional MRI is used widely for mapping cortical and subcortical regions that contribute to sensory, motor, language, and other cognitive functions. This information can be used for pre-surgical map-ping purposes.
examPles of images from eaCh modaliTy

Medical Neuroscience 2013 : Block 1
Page 136Principles of Structural Neuroimaging
Proton Density MRI Scan
MR Angiography
MR Spectroscopy (with example of an N-acetyl aspartate (NAA) peak (right))
Medical Neuroscience 2013 : Block 1
Page 137Principles of Structural Neuroimaging
Functional MRI (activation while watching a movie)
PET (using FDG)