
RET/GFRα Signals Are Dispensable for Thymic T Cell Development In Vivo
Upload
independentCategory
view
0download
0
www.elsevier.com/locate/ydbio
Developmental Biology 272 (2004) 118–133
Neural cells in the esophagus respond to glial cell line-derived
neurotrophic factor and neurturin, and are RET-dependent
Hui Yan,a Annette J. Bergner,a Hideki Enomoto,b,c Jeffrey Milbrandt,c
Donald F. Newgreen,d and Heather M. Younga,*
aDepartment of Anatomy and Cell Biology, University of Melbourne, Parkville, 3010 Victoria, AustraliabLaboratory for Neuronal Differentiation and Regeneration, RIKEN Center for Developmental Biology, Chuo, Kobe, Hyogo 650-0047, Japan
cDepartments of Pathology and Internal Medicine, Washington University School of Medicine, St. Louis, MO 63110, USAdMurdoch Childrens Research Institute, Royal Childrens Hospital, Parkville, 3052 Victoria, Australia
Received for publication 12 February 2004, revised 12 April 2004, accepted 12 April 2004
Available online 24 May 2004
Abstract
Glial cell line-derived neurotrophic factor (GDNF) is expressed in the gastrointestinal tract of the developing mouse and appears to
play an important role in the migration of enteric neuron precursors into and along the small and large intestines. Two other GDNF family
members, neurturin and artemin, are also expressed in the developing gut although artemin is only expressed in the esophagus. We
examined the effects of GDNF, neurturin, and artemin on neural crest cell migration and neurite outgrowth in explants of mouse
esophagus, midgut, and hindgut. Both GDNF and neurturin induced neural crest cell migration and neurite outgrowth in all regions
examined. In the esophagus, the effect of GDNF on migration and neurite outgrowth declined with age between E11.5 and E14.5, but
neurturin still had a strong neurite outgrowth effect at E14.5. Artemin did not promote neural migration or neurite outgrowth in any region
investigated. The effects of GDNF family ligands are mediated by the Ret tyrosine kinase. We examined the density of neurons in the
esophagus of Ret�/� mice, which lack neurons in the small and large intestines. The density of esophageal neurons in Ret�/� mice was
only about 4% of the density of esophageal neurons in Ret+/� and Ret+/+ mice. These results show that GDNF and neurturin promote
migration and neurite outgrowth of crest-derived cells in the esophagus as well as the intestine. Moreover, like intestinal neurons, the
development of esophageal neurons is largely Ret-dependent.
D 2004 Elsevier Inc. All rights reserved.
Keywords: GDNF; Neurturin; Cell migration; Neurite outgrowth; Ret�/�; Esophagus; Gastrointestinal tract
Introduction ligand binding molecule, GFRa1, and a receptor tyrosine
The enteric nervous system is an extensive system of
neurons and glial cells within the wall of the gastrointestinal
tract. Most enteric neurons arise from neural crest cells that
emigrate from the vagal level of the neural axis (post-otic
hindbrain adjacent to somites 1–7) (Burns and Le Douarin,
1998; Le Douarin and Teillet, 1973; Yntema and Hammond,
1954).
The glial cell line-derived neurotrophic factor (GDNF)
signaling pathway is essential for the development of the
enteric nervous system in most regions of the gastrointes-
tinal tract. GDNF acts at a receptor complex consisting of a
0012-1606/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.ydbio.2004.04.025
* Corresponding author. Fax: +61-613-9347-5219.
E-mail address: [email protected] (H.M. Young).
kinase, Ret (Airaksinen et al., 1999). Mice lacking GDNF,
GFRa1, or Ret lack enteric neurons in the small and large
intestines (Cacalano et al., 1998; Enomoto et al., 1998;
Moore et al., 1996; Pichel et al., 1996; Sanchez et al., 1996;
Schuchardt et al., 1994; Tomac et al., 2000). GDNF appears
to play multiple roles in enteric neuron development,
including survival, proliferation, and differentiation (Chala-
zonitis et al., 1998; Focke et al., 2001, 2003; Gianino et al.,
2003; Hearn et al., 1998; Shen et al., 2002; Taraviras et al.,
1999; Worley et al., 2000). In addition, GDNF is chemo-
attractive to enteric neural crest-derived cells, and appears to
play an important role in promoting the migration of vagal
neural crest cells into and along the gastrointestinal tract
(Barlow et al., 2003; Iwashita et al., 2003; Kruger et al.,
2003; Natarajan et al., 2002; Young et al., 2001). Despite
the absence of enteric neurons in the small and large

Fig. 1. Diagram of the gastrointestinal tract from an E11.5 mouse showing
the location of neural crest-derived cells (shown in grey; see Young and
H. Yan et al. / Developmental Biology 272 (2004) 118–133 119
intestines of mice with null mutations in Gdnf, Gfra1, orRet, enteric neurons are reported to be present in the
esophagus of Ret�/� mice, and, therefore, it has been
proposed that the development of neurons in the esophagus
is Ret-independent (Durbec et al., 1996). Thus, enteric
neuron precursors in different parts of the gastrointestinal
tract may differ in their requirements for GDNF family
members.
In addition to GDNF, two other members of the GDNF
family of ligands are expressed in the developing gastroin-
testinal tract, neurturin and artemin. The effects of neurturin
are mediated by a receptor complex consisting of its
preferred co-receptor, GFRa2, and Ret, and the receptor
complex for artemin consists of GFRa3 and Ret, although
neurturin and artemin may also bind weakly to GFRa1
(Airaksinen and Saarma, 2002). Neurturin is expressed
throughout the developing gastrointestinal tract (Golden et
al., 1999; Widenfalk et al., 1997; Xian et al., 1999), whereas
artemin is only expressed in the esophagus (Enomoto et al.,
2001). Neurturin promotes the proliferation and differenti-
ation of enteric neural crest-derived cells in vitro (Heuck-
eroth et al., 1998; Taraviras et al., 1999). Nonetheless, mice
with null mutations in neurturin or GFRa2 have neurons
present in normal numbers throughout the gastrointestinal
tract, but show a decreased density of excitatory nerve fibres
in the circular muscle (Gianino et al., 2003; Heuckeroth et
al., 1999; Rossi et al., 1999, 2003). Studies of mice with a
null mutation in artemin or GFRa3, the binding molecule
for artemin, have not reported any defects in the enteric
nervous system (Honma et al., 2002; Nishino et al., 1999).
The effects of neurturin and artemin on enteric neural crest
cell-derived migration have not previously been examined.
In this study, we examined the effect of GDNF, neurturin,
and artemin on neural migration and neurite outgrowth from
neural precursors in different regions of the E10.5–E14.5
gastrointestinal tract (esophagus, midgut, and hindgut). Our
results show that both GDNF and neurturin induce neural
migration and neurite outgrowth in the esophagus as well as
the intestine, but the effect of GDNF declines with age.
Although artemin is expressed in the esophagus, artemin did
not induce migration or neurite outgrowth in the esophagus
or any gastrointestinal region examined. In addition, al-
though mice lacking Ret have been reported to possess
enteric neurons in the esophagus, the density of enteric
neurons in the esophagus of Ret�/� mice has not been
previously been quantified. We show that the density of
neurons in the esophagus of Ret�/� mice is dramatically
(about 25 times) lower than that in Ret+/� or Ret+/+ mice,
suggesting that the development of enteric neurons in all
regions of the gastrointestinal tract is largely Ret-dependent.
Newgreen, 2001). The regions labelled ‘‘a’’ (esophagus), ‘‘b’’ (midgut), and
‘‘c’’ (hindgut) were removed, suspended between a ‘‘V’’ cut into a piece of
filter paper and grown in tissue culture medium for 4 days as described
previously (Hearn et al., 1999). At the time of explantation, neural crest-
derived cells (grey) were present throughout the explants of esophagus and
midgut, whereas in the hindgut, they were only present at the rostral end of
the explants (lower panels).
Materials and methods
Embryonic BALB/c mice were used for all organ culture
experiments. The day at which a vaginal plug was found
was designated E0.5. Pregnant mothers were killed by
cervical dislocation, and the embryos removed using aseptic
conditions.
Suspension (catenary) culture
Segments of esophagus, midgut, and hindgut from E11.5
mice, esophagus from E10.5, and midgut from E12.5 mice
were dissected. The entire esophagus, which was 1.2–1.5
mm long, was dissected and explanted. Explants of midgut
were from the caudal region, rostral to the caecum, and
include regions that will develop into the ileum and jeju-
num. A ‘‘V’’ was cut into a 3 � 3-mm piece of Millipore
filter paper using a scalpel, and an explant was suspended
across the ‘‘V’’ as described previously (Hearn et al., 1999;
Fig. 1). The orientation of the hindgut explant was indicated
by removing the corner of the filter paper adjacent to the
caudal end. The whole preparation, including the filter paper
and the attached explant, was placed in a Terasaki well in 20
Al of tissue culture medium and cultured in a 5% CO2
incubator at 37jC for 4 days. The tissue culture medium
contained DMEM (Trace Scientific Ltd., Noble Park, Vic-
toria, Australia) with 10% fetal bovine serum, 2 mM L-
glutamine, 0.075% sodium bicarbonate, and penicillin/strep-
tomycin sulfate solution. GDNF, artemin, or neurturin (all at
100 ng/ml, PeproTech Inc., Rocky Hill, NJ, USA) was
added to the culture medium of some explants. The culture
medium was changed after 2 days.

Table 1
Primary and secondary antibodies used
Primary antibodies
Host Dilution Source
PGP 9.5 Rabbit 1:2000 The Binding Site,
Birmingham, UK
Neurofilament-M
(145 kDa)
Rabbit 1:1000 Chemicon, Temecula,
CA, USA
Hu Human 1:2000 Fairman et al., 1995
NOS Sheep 1:2000 Norris et al., 1995
Phox2b Rabbit 1:700 Pattyn et al., 1997
GFRa1, GFRa3 Goat 1:25 R&D systems, Inc.,
Minneapolis, MN, USA
Secondary antibodies
Species in which
primary antibodies
were raised
Secondary antibodies
Rabbit Goat anti-rabbit Alexa 488 (1:250, Molecular
Probes, Eugene, OR, USA)
Human Donkey anti-human Texas Red (1:100, Jackson
ImmunoResearch, West Grove, PA, USA)
Sheep or goat Donkey anti-sheep FITC (1:100, Jackson
ImmunoResearch), or Biotinylated donkey
anti-sheep (1:100, Jackson ImmunoResearch)
followed by streptavidin-Cy5
(1:100, Amersham)
tal Biology 272 (2004) 118–133
Slice explants grown on collagen gel
The esophagus and midgut from E11.5, E12.5, and E14.5
mice were dissected, cut into transverse sections about 0.5–1
mm thick, and the slices placed on collagen gels in a 5% CO2
incubator at 37jC for 4 days. The collagen gels were made
by restoring 4 mg/ml acidic collagen solution (Upstate,
Parkville, Victoria, Australia) to normal osmolality with
5� DMEM and normal pH with 200 mM NaOH, on ice.
This solution was diluted to 1 mg/ml collagen with tissue
culture medium. GDNF, artemin, or neurturin was added to
some collagen solutions before gelling to give a final
concentration of 100 ng/ml.
Slice explants grown on filter paper
Transverse slices of esophagus and midgut from E11.5
mice were placed on 2.5 � 2.5 mm squares of filter papers.
Each preparation was placed in a Terasaki well in 20 Al oftissue culture medium with or without 100 ng/ml GDNF,
artemin, or neurturin and cultured in a 5% CO2 incubator at
37jC for 4 days. The tissue culturemediumwas changed after
2 days.
Immunohistochemistry
Explants were fixed in 4% paraformaldehyde in 0.1 M
phosphate buffer for 4–24 hours. Explants grown on filter
paper supports were left attached to the filter paper. After
fixation, the explants were processed for immunohistochem-
istry using the antisera shown in Table 1. Whole-mount
preparations of the gastrointestinal tract from the esophagus
to the rectum from E12.5 and E14.5 mice were fixed for 2–
4 hours in 4% paraformaldehyde in 0.1 M phosphate buffer
or in Zamboni’s fixative for 2–4 hours, and then processed
for immunohistochemistry using GFRa1 and GFRa3 anti-
sera (see Table 1).
Neuron counts within and outside suspended explants of gut
The total number of PGP9.5+ neurons present on the
filter paper supports outside the gut proper was counted,
and the density of PGP9.5+ neurons on the top surface of
the gut explants was determined. To count the total
number of PGP9.5+ cells on the filter paper, images were
taken on a fluorescence microscope using a 5� lens. A
montage was made of the images using CorelDRAW10
and printed, and PGP9.5+ neurons were counted from the
print. In regions where the cells were clumped or over-
lapped, a 40� lens was used to count the neurons directly
under the microscope. The total numbers of PGP9.5+
neurons on the paper supports at both rostral and caudal
ends were counted and added. Within the gut, the density
of PGP9.5+ neurons on the top surface only of the
suspended explants was determined as the neurons along
the bottom and sides of the gut explants could not be
H. Yan et al. / Developmen120
visualized clearly because of the tubular shape of the gut.
The preparations were viewed on Bio-Rad MRC1000 or
1024 confocal microscopes (Bio-Rad, Richmond, CA) and
images of PGP9.5+ neurons in the suspended gut were
taken by using a 20� objective lens. Images were printed
and the number of neurons counted. The area of gut
explant within which the neurons were counted was
measured using Scion image software and then the density
of neurons was calculated.
Enteric neuron density within the esophagus of Ret�/�,
Ret+/�, and Ret+/+ mice
Mice in which cDNA encoding tau-EGFP-myc (TGM)
had been inserted into the first coding exon of the receptor
tyrosine kinase gene, Ret, were used (Enomoto et al., 2001).
Mice heterozygous for the Ret-TGM mutation (RetTGM/+
mice) were mated. The genotype of adult and embryonic
RetTGM mice was determined by PCR using the primers and
conditions reported by Enomoto et al. (2001). The identity
of E18.5 Ret�/� mice was confirmed by processing samples
of duodenum from each embryo for NADPH diaphorase
histochemistry (Ward et al., 1999). Samples of duodenum
were fixed in 4% paraformaldehyde in 0.1M phosphate
buffer for 1 h, washed, incubated in 10 mg h-NADPH,2.5 mg nitroblue tetrazolium, and 20 Al Triton X in 10 ml
0.1 M Tris/HCl (pH 8.0) for 15–20 minutes at 37jC and
then examined to determine if there were any NADPH
diaphorase-stained neurons. E18.5 esophagus was processed
for immunohistochemistry using antibodies to Hu (Table 1).
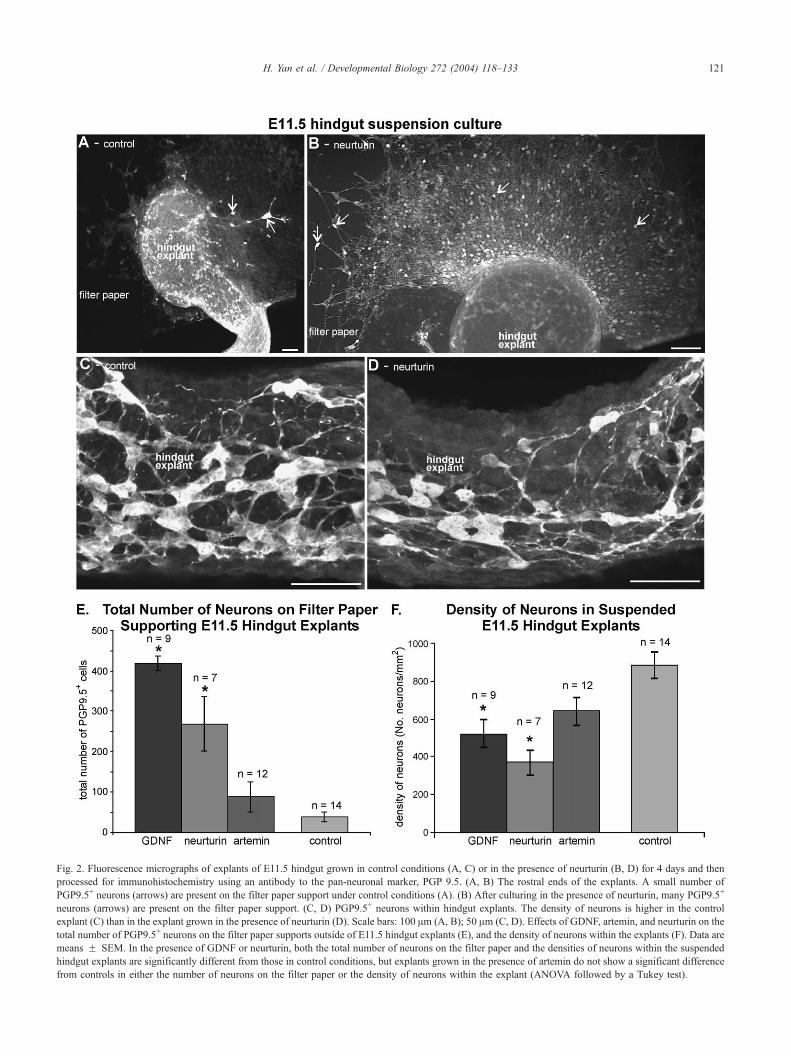
Fig. 2. Fluorescence micrographs of explants of E11.5 hindgut grown in control conditions (A, C) or in the presence of neurturin (B, D) for 4 days and then
processed for immunohistochemistry using an antibody to the pan-neuronal marker, PGP 9.5. (A, B) The rostral ends of the explants. A small number of
PGP9.5+ neurons (arrows) are present on the filter paper support under control conditions (A). (B) After culturing in the presence of neurturin, many PGP9.5+
neurons (arrows) are present on the filter paper support. (C, D) PGP9.5+ neurons within hindgut explants. The density of neurons is higher in the control
explant (C) than in the explant grown in the presence of neurturin (D). Scale bars: 100 Am (A, B); 50 Am (C, D). Effects of GDNF, artemin, and neurturin on the
total number of PGP9.5+ neurons on the filter paper supports outside of E11.5 hindgut explants (E), and the density of neurons within the explants (F). Data are
means F SEM. In the presence of GDNF or neurturin, both the total number of neurons on the filter paper and the densities of neurons within the suspended
hindgut explants are significantly different from those in control conditions, but explants grown in the presence of artemin do not show a significant difference
from controls in either the number of neurons on the filter paper or the density of neurons within the explant (ANOVA followed by a Tukey test).
H. Yan et al. / Developmental Biology 272 (2004) 118–133 121
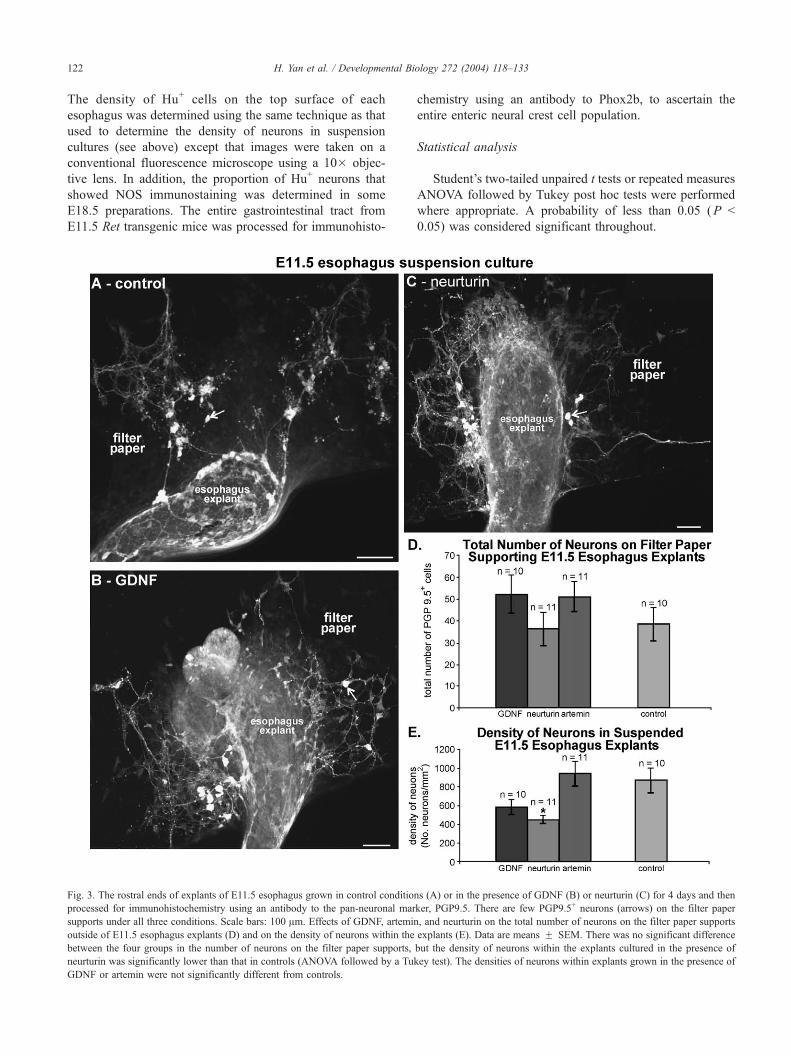
H. Yan et al. / Developmental Biology 272 (2004) 118–133122
The density of Hu+ cells on the top surface of each
esophagus was determined using the same technique as that
used to determine the density of neurons in suspension
cultures (see above) except that images were taken on a
conventional fluorescence microscope using a 10� objec-
tive lens. In addition, the proportion of Hu+ neurons that
showed NOS immunostaining was determined in some
E18.5 preparations. The entire gastrointestinal tract from
E11.5 Ret transgenic mice was processed for immunohisto-
Fig. 3. The rostral ends of explants of E11.5 esophagus grown in control conditio
processed for immunohistochemistry using an antibody to the pan-neuronal mar
supports under all three conditions. Scale bars: 100 Am. Effects of GDNF, artemin
outside of E11.5 esophagus explants (D) and on the density of neurons within the
between the four groups in the number of neurons on the filter paper supports,
neurturin was significantly lower than that in controls (ANOVA followed by a Tu
GDNF or artemin were not significantly different from controls.
chemistry using an antibody to Phox2b, to ascertain the
entire enteric neural crest cell population.
Statistical analysis
Student’s two-tailed unpaired t tests or repeated measures
ANOVA followed by Tukey post hoc tests were performed
where appropriate. A probability of less than 0.05 (P <
0.05) was considered significant throughout.
ns (A) or in the presence of GDNF (B) or neurturin (C) for 4 days and then
ker, PGP9.5. There are few PGP9.5+ neurons (arrows) on the filter paper
, and neurturin on the total number of neurons on the filter paper supports
explants (E). Data are means F SEM. There was no significant difference
but the density of neurons within the explants cultured in the presence of
key test). The densities of neurons within explants grown in the presence of
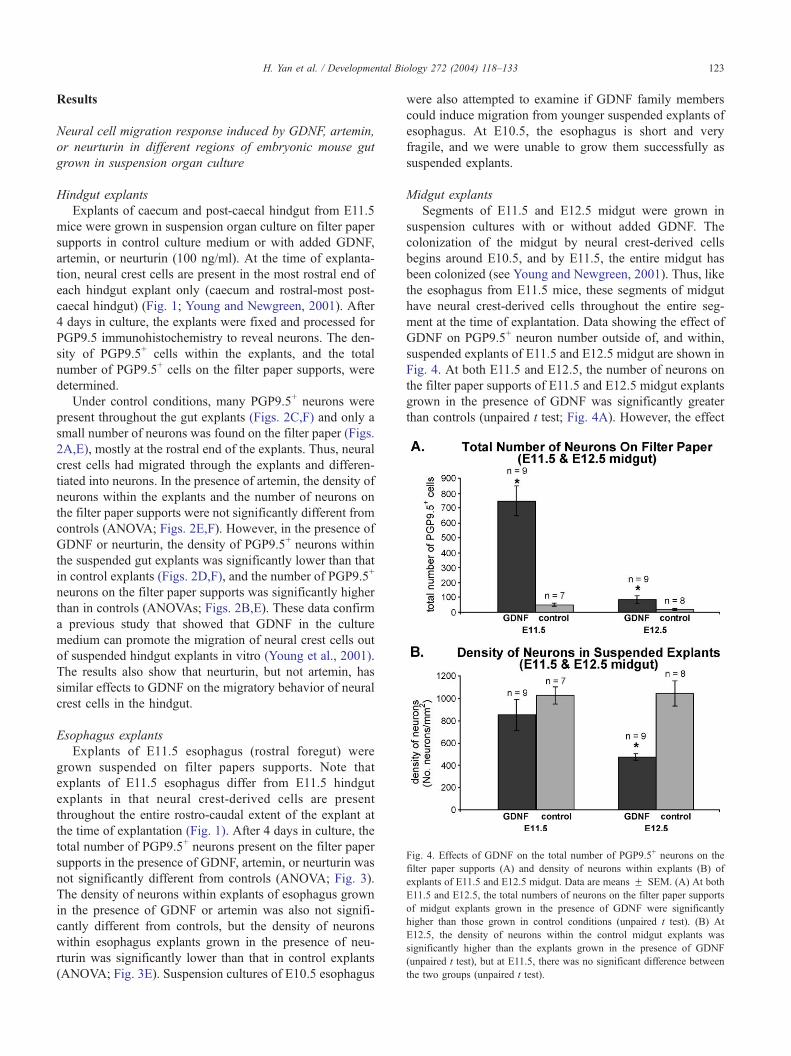
Fig. 4. Effects of GDNF on the total number of PGP9.5+ neurons on the
filter paper supports (A) and density of neurons within explants (B) of
explants of E11.5 and E12.5 midgut. Data are means F SEM. (A) At both
E11.5 and E12.5, the total numbers of neurons on the filter paper supports
of midgut explants grown in the presence of GDNF were significantly
higher than those grown in control conditions (unpaired t test). (B) At
E12.5, the density of neurons within the control midgut explants was
significantly higher than the explants grown in the presence of GDNF
(unpaired t test), but at E11.5, there was no significant difference between
the two groups (unpaired t test).
H. Yan et al. / Developmental Biology 272 (2004) 118–133 123
Results
Neural cell migration response induced by GDNF, artemin,
or neurturin in different regions of embryonic mouse gut
grown in suspension organ culture
Hindgut explants
Explants of caecum and post-caecal hindgut from E11.5
mice were grown in suspension organ culture on filter paper
supports in control culture medium or with added GDNF,
artemin, or neurturin (100 ng/ml). At the time of explanta-
tion, neural crest cells are present in the most rostral end of
each hindgut explant only (caecum and rostral-most post-
caecal hindgut) (Fig. 1; Young and Newgreen, 2001). After
4 days in culture, the explants were fixed and processed for
PGP9.5 immunohistochemistry to reveal neurons. The den-
sity of PGP9.5+ cells within the explants, and the total
number of PGP9.5+ cells on the filter paper supports, were
determined.
Under control conditions, many PGP9.5+ neurons were
present throughout the gut explants (Figs. 2C,F) and only a
small number of neurons was found on the filter paper (Figs.
2A,E), mostly at the rostral end of the explants. Thus, neural
crest cells had migrated through the explants and differen-
tiated into neurons. In the presence of artemin, the density of
neurons within the explants and the number of neurons on
the filter paper supports were not significantly different from
controls (ANOVA; Figs. 2E,F). However, in the presence of
GDNF or neurturin, the density of PGP9.5+ neurons within
the suspended gut explants was significantly lower than that
in control explants (Figs. 2D,F), and the number of PGP9.5+
neurons on the filter paper supports was significantly higher
than in controls (ANOVAs; Figs. 2B,E). These data confirm
a previous study that showed that GDNF in the culture
medium can promote the migration of neural crest cells out
of suspended hindgut explants in vitro (Young et al., 2001).
The results also show that neurturin, but not artemin, has
similar effects to GDNF on the migratory behavior of neural
crest cells in the hindgut.
Esophagus explants
Explants of E11.5 esophagus (rostral foregut) were
grown suspended on filter papers supports. Note that
explants of E11.5 esophagus differ from E11.5 hindgut
explants in that neural crest-derived cells are present
throughout the entire rostro-caudal extent of the explant at
the time of explantation (Fig. 1). After 4 days in culture, the
total number of PGP9.5+ neurons present on the filter paper
supports in the presence of GDNF, artemin, or neurturin was
not significantly different from controls (ANOVA; Fig. 3).
The density of neurons within explants of esophagus grown
in the presence of GDNF or artemin was also not signifi-
cantly different from controls, but the density of neurons
within esophagus explants grown in the presence of neu-
rturin was significantly lower than that in control explants
(ANOVA; Fig. 3E). Suspension cultures of E10.5 esophagus
were also attempted to examine if GDNF family members
could induce migration from younger suspended explants of
esophagus. At E10.5, the esophagus is short and very
fragile, and we were unable to grow them successfully as
suspended explants.
Midgut explants
Segments of E11.5 and E12.5 midgut were grown in
suspension cultures with or without added GDNF. The
colonization of the midgut by neural crest-derived cells
begins around E10.5, and by E11.5, the entire midgut has
been colonized (see Young and Newgreen, 2001). Thus, like
the esophagus from E11.5 mice, these segments of midgut
have neural crest-derived cells throughout the entire seg-
ment at the time of explantation. Data showing the effect of
GDNF on PGP9.5+ neuron number outside of, and within,
suspended explants of E11.5 and E12.5 midgut are shown in
Fig. 4. At both E11.5 and E12.5, the number of neurons on
the filter paper supports of E11.5 and E12.5 midgut explants
grown in the presence of GDNF was significantly greater
than controls (unpaired t test; Fig. 4A). However, the effect
H. Yan et al. / Developmental Biology 272 (2004) 118–133124
of GDNF on the number of neurons on the filter paper
declined dramatically with age. In the presence of GDNF,
the density of neurons within the gut explants was signif-
icantly lower than in the controls at E12.5 (unpaired t test),
but not at E11.5 (unpaired t test; Fig. 4B). This difference
may reflect differing effects of GDNF on proliferation and/
or differentiation at different ages. These data demonstrate
that even when neural crest cells are present throughout
midgut explants at the time of explantation, GDNF is
capable of inducing an increase in the number of neurons
on the filter paper supports, but that the effect of GDNF on
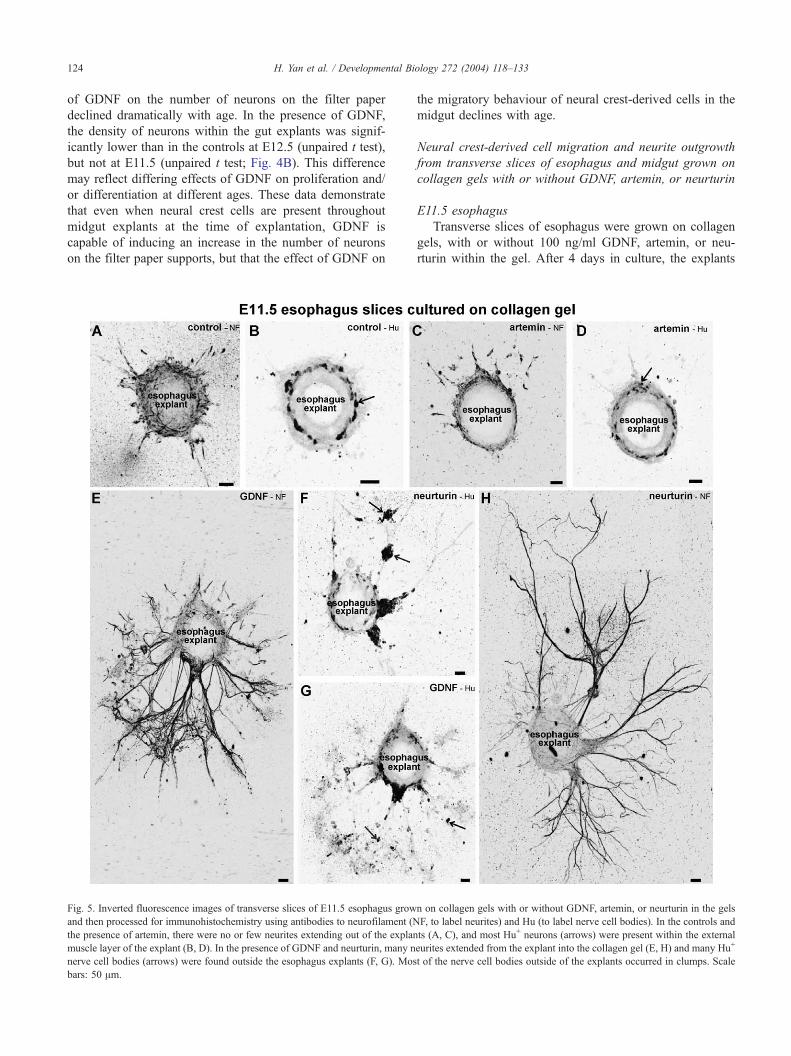
Fig. 5. Inverted fluorescence images of transverse slices of E11.5 esophagus grow
and then processed for immunohistochemistry using antibodies to neurofilament (N
the presence of artemin, there were no or few neurites extending out of the explan
muscle layer of the explant (B, D). In the presence of GDNF and neurturin, many n
nerve cell bodies (arrows) were found outside the esophagus explants (F, G). Mos
bars: 50 Am.
the migratory behaviour of neural crest-derived cells in the
midgut declines with age.
Neural crest-derived cell migration and neurite outgrowth
from transverse slices of esophagus and midgut grown on
collagen gels with or without GDNF, artemin, or neurturin
E11.5 esophagus
Transverse slices of esophagus were grown on collagen
gels, with or without 100 ng/ml GDNF, artemin, or neu-
rturin within the gel. After 4 days in culture, the explants
n on collagen gels with or without GDNF, artemin, or neurturin in the gels
F, to label neurites) and Hu (to label nerve cell bodies). In the controls and
ts (A, C), and most Hu+ neurons (arrows) were present within the external
eurites extended from the explant into the collagen gel (E, H) and many Hu+
t of the nerve cell bodies outside of the explants occurred in clumps. Scale
H. Yan et al. / Developmental Biology 272 (2004) 118–133 125
were immunostained for Hu to reveal neural cell bodies and
for neurofilament to reveal neurites. In control conditions, or
with artemin added to the gels, many Hu+ cells were present
within the explant, and they formed a discrete layer in the
outer part of the mesenchyme (Figs. 5B,D). A small number
of Hu+ cells was found close to, but outside, the explants in
a few specimens (2/12 controls and 1/12 with artemin
present). In control or artemin-treated explants, there were
either no neurites, or only sparse neurites, that extended
away from the explants (Figs. 5A,C). In the presence of
GDNF or neurturin, many Hu+ cells were observed on the
gel outside of the explants (Figs. 5F,G). The Hu+ cells
outside the explants were mainly found in clumps (Figs.
5F,G). Most esophagus explants grown on gels containing
GDNF or neurturin also possessed many bundles of neuro-
filament+ neurites that extended away from the explants (8/
12 with GDNF present in the gel, 10/12 with neurturin;
Figs. 5E,H). The clumps of Hu+ cells were usually found
associated with bundles of neurites. Hence, in contrast to
suspended explants of E11.5 esophagus, GDNF and neu-
rturin induced a migratory and neurite outgrowth response
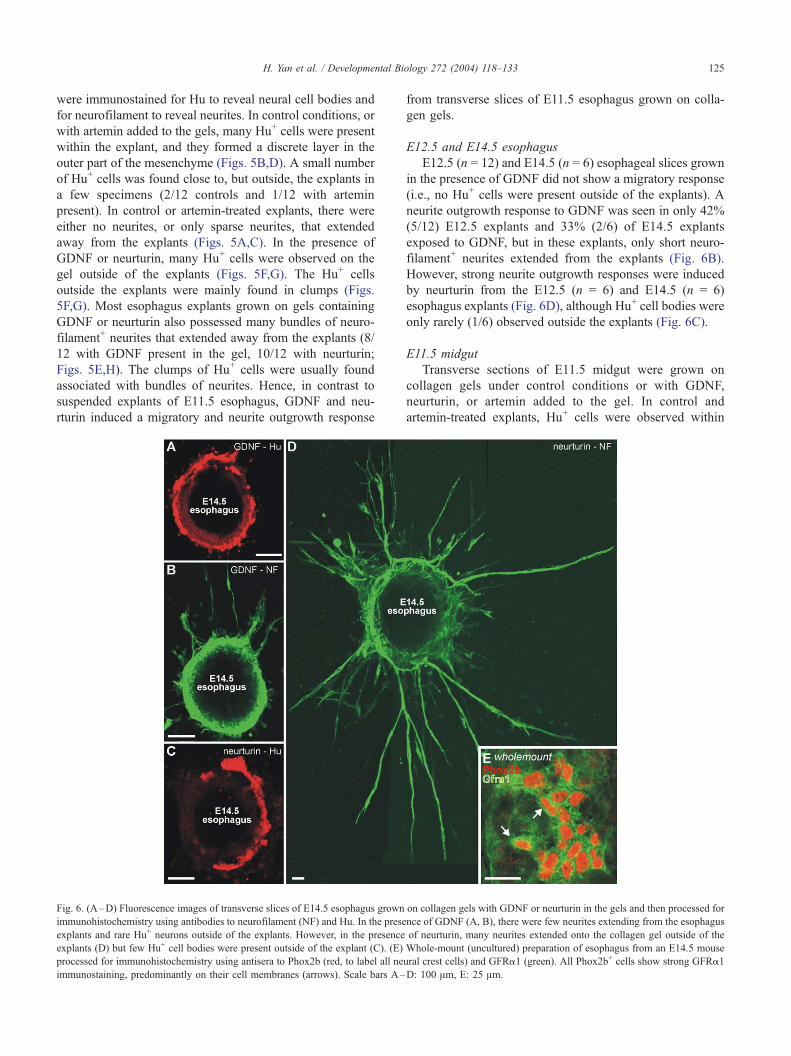
Fig. 6. (A–D) Fluorescence images of transverse slices of E14.5 esophagus grown
immunohistochemistry using antibodies to neurofilament (NF) and Hu. In the prese
explants and rare Hu+ neurons outside of the explants. However, in the presence
explants (D) but few Hu+ cell bodies were present outside of the explant (C). (E)
processed for immunohistochemistry using antisera to Phox2b (red, to label all ne
immunostaining, predominantly on their cell membranes (arrows). Scale bars A–
from transverse slices of E11.5 esophagus grown on colla-
gen gels.
E12.5 and E14.5 esophagus
E12.5 (n = 12) and E14.5 (n = 6) esophageal slices grown
in the presence of GDNF did not show a migratory response
(i.e., no Hu+ cells were present outside of the explants). A
neurite outgrowth response to GDNF was seen in only 42%
(5/12) E12.5 explants and 33% (2/6) of E14.5 explants
exposed to GDNF, but in these explants, only short neuro-
filament+ neurites extended from the explants (Fig. 6B).
However, strong neurite outgrowth responses were induced
by neurturin from the E12.5 (n = 6) and E14.5 (n = 6)
esophagus explants (Fig. 6D), although Hu+ cell bodies were
only rarely (1/6) observed outside the explants (Fig. 6C).
E11.5 midgut
Transverse sections of E11.5 midgut were grown on
collagen gels under control conditions or with GDNF,
neurturin, or artemin added to the gel. In control and
artemin-treated explants, Hu+ cells were observed within
on collagen gels with GDNF or neurturin in the gels and then processed for
nce of GDNF (A, B), there were few neurites extending from the esophagus
of neurturin, many neurites extended onto the collagen gel outside of the
Whole-mount (uncultured) preparation of esophagus from an E14.5 mouse
ural crest cells) and GFRa1 (green). All Phox2b+ cells show strong GFRa1
D: 100 Am, E: 25 Am.
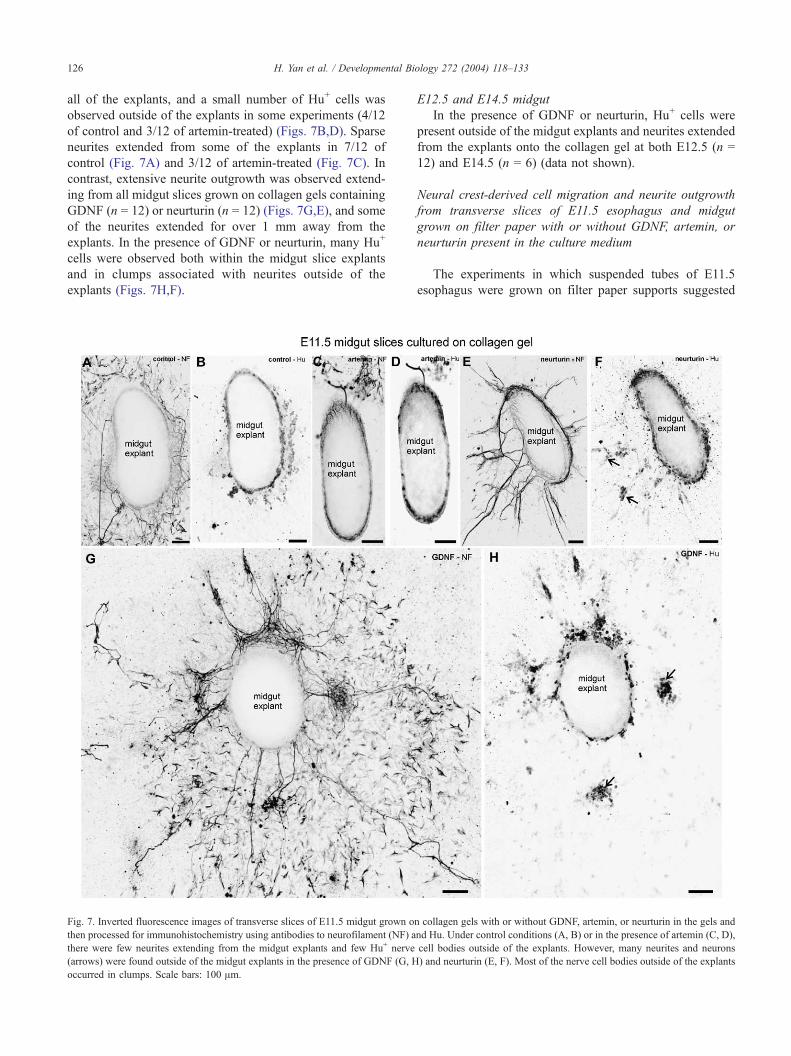
H. Yan et al. / Developmental Bi126
all of the explants, and a small number of Hu+ cells was
observed outside of the explants in some experiments (4/12
of control and 3/12 of artemin-treated) (Figs. 7B,D). Sparse
neurites extended from some of the explants in 7/12 of
control (Fig. 7A) and 3/12 of artemin-treated (Fig. 7C). In
contrast, extensive neurite outgrowth was observed extend-
ing from all midgut slices grown on collagen gels containing
GDNF (n = 12) or neurturin (n = 12) (Figs. 7G,E), and some
of the neurites extended for over 1 mm away from the
explants. In the presence of GDNF or neurturin, many Hu+
cells were observed both within the midgut slice explants
and in clumps associated with neurites outside of the
explants (Figs. 7H,F).
Fig. 7. Inverted fluorescence images of transverse slices of E11.5 midgut grown o
then processed for immunohistochemistry using antibodies to neurofilament (NF) a
there were few neurites extending from the midgut explants and few Hu+ nerve
(arrows) were found outside of the midgut explants in the presence of GDNF (G, H
occurred in clumps. Scale bars: 100 Am.
E12.5 and E14.5 midgut
In the presence of GDNF or neurturin, Hu+ cells were
present outside of the midgut explants and neurites extended
from the explants onto the collagen gel at both E12.5 (n =
12) and E14.5 (n = 6) (data not shown).
Neural crest-derived cell migration and neurite outgrowth
from transverse slices of E11.5 esophagus and midgut
grown on filter paper with or without GDNF, artemin, or
neurturin present in the culture medium
The experiments in which suspended tubes of E11.5
esophagus were grown on filter paper supports suggested
ology 272 (2004) 118–133
n collagen gels with or without GDNF, artemin, or neurturin in the gels and
nd Hu. Under control conditions (A, B) or in the presence of artemin (C, D),
cell bodies outside of the explants. However, many neurites and neurons
) and neurturin (E, F). Most of the nerve cell bodies outside of the explants
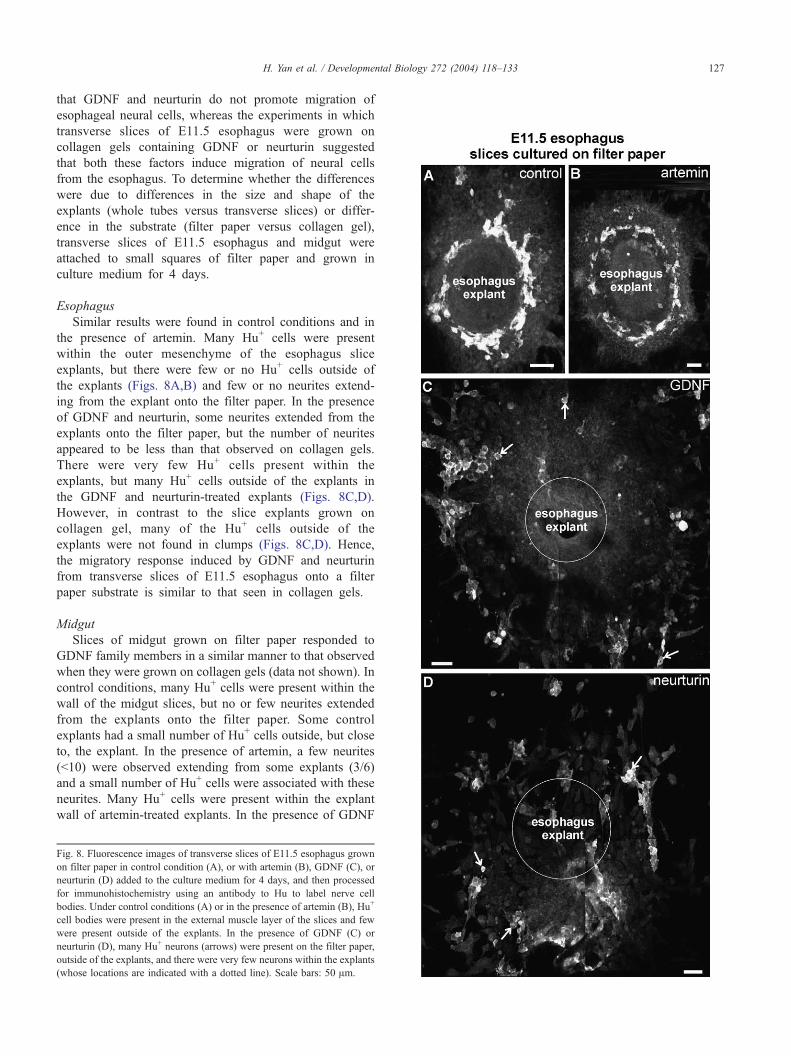
H. Yan et al. / Developmental Biology 272 (2004) 118–133 127
that GDNF and neurturin do not promote migration of
esophageal neural cells, whereas the experiments in which
transverse slices of E11.5 esophagus were grown on
collagen gels containing GDNF or neurturin suggested
that both these factors induce migration of neural cells
from the esophagus. To determine whether the differences
were due to differences in the size and shape of the
explants (whole tubes versus transverse slices) or differ-
ence in the substrate (filter paper versus collagen gel),
transverse slices of E11.5 esophagus and midgut were
attached to small squares of filter paper and grown in
culture medium for 4 days.
Esophagus
Similar results were found in control conditions and in
the presence of artemin. Many Hu+ cells were present
within the outer mesenchyme of the esophagus slice
explants, but there were few or no Hu+ cells outside of
the explants (Figs. 8A,B) and few or no neurites extend-
ing from the explant onto the filter paper. In the presence
of GDNF and neurturin, some neurites extended from the
explants onto the filter paper, but the number of neurites
appeared to be less than that observed on collagen gels.
There were very few Hu+ cells present within the
explants, but many Hu+ cells outside of the explants in
the GDNF and neurturin-treated explants (Figs. 8C,D).
However, in contrast to the slice explants grown on
collagen gel, many of the Hu+ cells outside of the
explants were not found in clumps (Figs. 8C,D). Hence,
the migratory response induced by GDNF and neurturin
from transverse slices of E11.5 esophagus onto a filter
paper substrate is similar to that seen in collagen gels.
Midgut
Slices of midgut grown on filter paper responded to
GDNF family members in a similar manner to that observed
when they were grown on collagen gels (data not shown). In
control conditions, many Hu+ cells were present within the
wall of the midgut slices, but no or few neurites extended
from the explants onto the filter paper. Some control
explants had a small number of Hu+ cells outside, but close
to, the explant. In the presence of artemin, a few neurites
(<10) were observed extending from some explants (3/6)
and a small number of Hu+ cells were associated with these
neurites. Many Hu+ cells were present within the explant
wall of artemin-treated explants. In the presence of GDNF
Fig. 8. Fluorescence images of transverse slices of E11.5 esophagus grown
on filter paper in control condition (A), or with artemin (B), GDNF (C), or
neurturin (D) added to the culture medium for 4 days, and then processed
for immunohistochemistry using an antibody to Hu to label nerve cell
bodies. Under control conditions (A) or in the presence of artemin (B), Hu+
cell bodies were present in the external muscle layer of the slices and few
were present outside of the explants. In the presence of GDNF (C) or
neurturin (D), many Hu+ neurons (arrows) were present on the filter paper,
outside of the explants, and there were very few neurons within the explants
(whose locations are indicated with a dotted line). Scale bars: 50 Am.
H. Yan et al. / Developmental Biology 272 (2004) 118–133128
(n = 6) or neurturin (n = 6), many Hu+ cells were present
both within and outside of midgut slice explants and many
neurites extended from the explant onto the filter paper. The
migrating neurons were closely associated with nerve fibres.
GFRa1 and GFRa3 immunostaining
GFRa1
The experiments described above showed that the
GDNF-induced neurite outgrowth and migratory responses
of neural crest-derived cells in the esophagus declined with
age, whereas the response of midgut neural cells to GDNF
was maintained until at least E14.5. To examine whether the
decline in responsiveness of esophageal neural cells is due
to a down-regulation of GFRa1, we examined the presence
of GFRa1 immunostaining in whole-mount preparations of
esophagus and midgut using an antiserum that has previ-
ously been shown to be specific for GFRa1 (Golden et al.,
2003). At E14.5, neural crest cells (identified by Phox2b
immunostaining) in both the esophagus (Fig. 6E) and
midgut showed strong GFRa1 immunostaining, and there
was no obvious difference in the levels of immunostaining
between cells in the two different gut regions. Thus, the
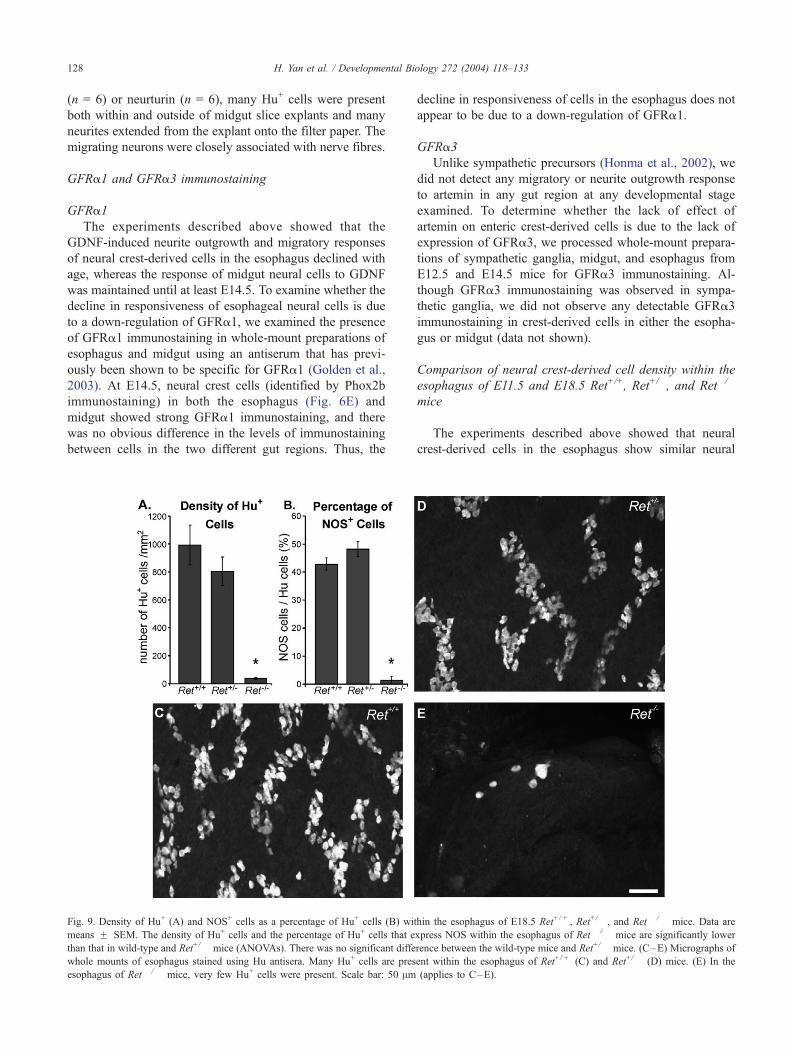
Fig. 9. Density of Hu+ (A) and NOS+ cells as a percentage of Hu+ cells (B) wi
means F SEM. The density of Hu+ cells and the percentage of Hu+ cells that e
than that in wild-type and Ret+/� mice (ANOVAs). There was no significant diffe
whole mounts of esophagus stained using Hu antisera. Many Hu+ cells are pres
esophagus of Ret� /� mice, very few Hu+ cells were present. Scale bar: 50 Am
decline in responsiveness of cells in the esophagus does not
appear to be due to a down-regulation of GFRa1.
GFRa3
Unlike sympathetic precursors (Honma et al., 2002), we
did not detect any migratory or neurite outgrowth response
to artemin in any gut region at any developmental stage
examined. To determine whether the lack of effect of
artemin on enteric crest-derived cells is due to the lack of
expression of GFRa3, we processed whole-mount prepara-
tions of sympathetic ganglia, midgut, and esophagus from
E12.5 and E14.5 mice for GFRa3 immunostaining. Al-
though GFRa3 immunostaining was observed in sympa-
thetic ganglia, we did not observe any detectable GFRa3
immunostaining in crest-derived cells in either the esopha-
gus or midgut (data not shown).
Comparison of neural crest-derived cell density within the
esophagus of E11.5 and E18.5 Ret+/+, Ret+/�, and Ret�/�
mice
The experiments described above showed that neural
crest-derived cells in the esophagus show similar neural
thin the esophagus of E18.5 Ret+/ + , Ret+/� , and Ret� /� mice. Data are
xpress NOS within the esophagus of Ret� / � mice are significantly lower
rence between the wild-type mice and Ret+/� mice. (C–E) Micrographs of
ent within the esophagus of Ret+/ + (C) and Ret+/� (D) mice. (E) In the
(applies to C–E).

H. Yan et al. / Developmental Biology 272 (2004) 118–133 129
migration responses to GDNF and neurturin to cells in the
midgut and hindgut. A previous study reported that
enteric neurons are present in the esophagus of Ret�/�
mice (Durbec et al., 1996). However, the number of
neurons present in the esophagus of mice lacking mem-
bers of the GDNF-signalling pathway has not been
examined quantitatively.
E18.5 esophagus
We examined the density of all enteric neurons in the
E18.5 esophagus using Hu antisera. The identity of Ret�/�
mice determined by PCR was confirmed by NADPH
diaphorase staining of samples of duodenum (Ward et al.,
1999). The density of Hu+ cells on the top surface of the
entire esophagus was determined in five Ret�/�, five Ret+/�,
Fig. 10. Low (A, B) and higher (C, D) magnification images showing the distrib
Ret+/� (A, C) and Ret� /� (B, D) mice. Many Phox2b+ cells were present along
number of Phox2b+ cells were present within the wall of esophagus of Ret� /� m
mice were in the rostral region. There were also some Phox2b+ cells in the stom
and five Ret+/+ mice. The density of Hu+ cells in the
esophagus of E18.5 Ret�/� mice was significantly lower
than that in the esophagus of both wild-type and Ret+/�
mice (ANOVA; Fig. 9), and was only about 4% that of wild-
type and heterozygous mice. The density of Hu+ cells in the
esophagus of Ret+/� mice was not significantly different
from wild-type mice (ANOVA; Fig. 9). A major sub-class of
neurons in the mouse esophagus expresses nitric oxide
synthase (NOS) (Sang and Young, 1997; Worl et al.,
2002). The proportion of Hu+ cells that showed NOS im-
munostaining was examined in three Ret�/�, three Ret+/�,
and three Ret+/+ mice. In wild-type and heterozygous mice,
around 40% of Hu+ cells were NOS+ (Fig. 9B). In contrast,
of 121 Hu+ cells examined in the esophagus of Ret�/� mice,
only 1 cell showed NOS immunostaining.
ution of Phox2b+ neural crest-derived cells within the esophagus of E11.5
the entire esophagus of the Ret+/� mouse (A, C). However, only a small
ice (B, D). Most Phox2b+ cells present in the esophagus of E11.5 Ret� / �
ach of Ret� / � mice (B). Scale bars: 50 Am.

H. Yan et al. / Developmental Biology 272 (2004) 118–133130
E11.5 esophagus
To determine whether the decrease in the density of
neurons in the esophagus of E18.5 mice is due to failure
of the neural crest cells to colonize the esophagus or a
failure to survive and differentiate, we examined the immu-
nolocalization of Phox2b in the E11.5 gastrointestinal tract.
Phox2b appears to be expressed by all neural crest-derived
cells within the gastrointestinal tract (Young et al., 2003). In
the esophagus of E11.5 wild-type (n = 7) and Ret+/� (n = 6)
embryos, many Phox2b+ cells were present along the whole
esophagus. However, in the esophagus of homozygous
mutant E11.5 embryos (n = 7), only sparse Phox2b+ cells
were seen within the wall of esophagus (Fig. 10). Most
Phox2b+ cells present in the esophagus of E11.5 Ret�/�
mice were in the rostral esophagus (data not shown). In
E11.5 homozygous mutant mice, Phox2b+ cells were ob-
served outside of the esophagus along the pathway of the
vagus nerve, and there were also Phox2b+ cells in the
stomach, but in vastly reduced numbers compared to wild-
type or Ret+/� embryos (Fig. 10).
Discussion
The results of this study show that (i) neurturin, like
GDNF, is chemoattractive to enteric neural crest-derived
cells and also induces neurite outgrowth; (ii) GDNF and
neurturin promote the migration of crest-derived cells from
the esophagus as well as the midgut and hindgut; (iii)
artemin has no detectable effect on migration or neurite
outgrowth in any region examined; (iv) the migratory
responses induced by GDNF or neurturin decline with
age; and (v) the development of esophageal neurons is
largely Ret-dependent.
Neurturin has similar effects to GDNF on migration and
neurite outgrowth
A previous study has shown that GDNF and neurturin
have similar effects on the survival, proliferation, and
differentiation of neural crest-derived cells isolated from
the gut (Taraviras et al., 1999). Our study showed that
exogenous neurturin also has similar effects to GDNF on the
migration of enteric crest-derived cells, and both neurturin
and GDNF were chemoattractive to crest-derived cells in
esophagus, mid-, and hindgut explants. The current study
also showed that both neurturin and GDNF induced neurite
outgrowth from explants of embryonic gut.
Neurturin is not essential for the migration of neural
precursors into and along the developing gut in vivo. Mice
lacking neurturin or its ligand binding molecule, GFRa2,
possess an enteric nervous system along the entire length of
the gastrointestinal tract (Gianino et al., 2003; Heuckeroth et
al., 1999; Rossi et al., 1999, 2003). Furthermore, although
there is some expression of neurturin in the mucosa of E12
mouse gut, neurturin expression is not detected in the gut
mesenchyme (through which neural crest cells migrate) until
E14 (Golden et al., 1999); this is after the colonization of the
embryonic mouse gut by crest-derived cells is complete
(Kapur et al., 1992). In contrast, GDNF signalling is
necessary for neural cell migration in the intestine (Cacalano
et al., 1998; Enomoto et al., 1998; Moore et al., 1996; Pichel
et al., 1996; Sanchez et al., 1996; Tomac et al., 2000), and
GDNF is expressed by the gut mesenchyme before the entry
of neural crest-derived cells (Natarajan et al., 2002). How-
ever, mice lacking neurturin or GFRa2, which forms part of
the receptor complex for neurturin, have fewer nerve fibres
in the circular muscle (Heuckeroth et al., 1999; Rossi et al.,
1999, 2003), where neurturin is normally primarily
expressed. Thus, neurturin seems to be important for induc-
ing axon outgrowth into the circular muscle layer.
Studies using cell lines have shown that neurturin can
activate Ret by binding to either GFRa1 or GFRa2 (Cree-
don et al., 1997; Jing et al., 1997). Hence, the responses
induced by neurturin described in the current study could be
due to neurturin binding to either GFRa1 or GFRa2, or to
both.
Neural cell migration and axon outgrowth have many
features in common (Rakic, 1999), and it is likely that the
response (migration or axon extension) induced by activa-
tion of Ret by GDNF or neurturin will depend on the age
and state of differentiation of the enteric crest-derived cells.
Ret activation in younger or undifferentiated enteric crest-
derived cells may bias the response toward migration,
whereas Ret activation in older, post-migratory cells may
bias the response toward neuronal differentiation and axon
outgrowth (Hearn et al., 1998; Wu et al., 1999).
Enteric neural crest-derived cells did not show a migratory
or neurite outgrowth response to artemin
Artemin appears to play a role in migration and neurite
outgrowth during the development of the sympathetic ner-
vous system (Enomoto et al., 2001; Honma et al., 2002;
Nishino et al., 1999). The only gastrointestinal region that
expresses artemin is the esophagus (Enomoto et al., 2001).
Despite the expression of artemin in the esophagus, enteric
neural crest-derived cells in both the esophagus and the
intestine failed to show a migratory or neurite outgrowth
response to artemin at the ages examined in this study.
However, we also showed that, unlike sympathetic neuron
precursors, enteric neural crest-derived cells do not appear
to express GFRa3, which would account for their lack of
response to artemin.
The migratory response induced by GDNF and neurturin
declines with age
Our data showed that the GDNF-induced migratory
responses in both the esophagus and midgut declined with
age because (a) transverse slices of E11.5 esophagus grown
on collagen gel showed a strong migratory response to

H. Yan et al. / Developmental Biology 272 (2004) 118–133 131
GDNF and neurturin, but E12.5 and E14.5 esophageal slices
showed little or no migratory response; and (b) the chemo-
attractive response to GDNF of crest-derived cells in sus-
pended explants of E12.5 midgut was dramatically lower
than that of E11.5 suspended midgut explants (see Fig. 4).
The earlier age-dependent decline in the esophagus com-
pared to the midgut parallels the earlier colonization of the
esophagus by neural crest cells. In addition, our study
showed that the GDNF-induced neurite outgrowth response
from the esophagus also declined with age. In fetal rats, the
proliferative responses and neuronal differentiation induced
by GDNF or neurturin also decline between E12.5 and
E14.5/E15.5 (Chalazonitis et al., 1998; Taraviras et al.,
1999).
The decline in responsiveness to GDNF does not appear
to be due to a down-regulation of GFRa1, as E14.5
esophageal neural cells showed strong GFRa1 immunos-
taining, but did not show GDNF-induced migratory or
neurite-outgrowth responses. Thus, the age-dependent de-
cline in responsiveness appears to be due to changes
downstream of GFRa1. It is unclear whether the decline
in the GDNF-induced migratory responses with age reflects
an age-dependent decrease in migratory ability, or other
factors such as an increase in cell–cell adhesion, which may
prevent cells from migrating.
Although GDNF induced a migratory response from
transverse slices of E11.5 esophagus grown on collagen
gels or on filter paper, we did not detect a migratory
response from suspended explants of E11.5 esophagus.
We believe this is because the crest-derived cells have to
migrate further to escape the tubular suspended explants
than they do to escape the transverse slices. Because the
esophagus of E10.5 mice is fragile, we could not reliably
set them up as suspended explants for quantitative
analysis.
There was also a decline with age of the migratory
response of neural cells in the esophagus induced by
neurturin. However, unlike GDNF, there was still a robust
neurite outgrowth response from the esophagus and mid-
gut induced by neurturin at E14.5, the oldest age exam-
ined. Because of the thickness of the gut wall, it was not
possible to establish suspension or slice cultures of gut
from older fetuses. However, as neurturin appears to be
involved in inducing axon outgrowth into the circular
muscle, and as the innervation of the circular muscle
continues to develop after birth (Young et al., 1998), it
appears likely that the neurites of enteric neurons will be
responsive to neurturin postnatally.
GDNF and neurturin promote the migration of crest-derived
cells from the esophagus as well as the mid- and hindgut
The rostrocaudal origin of neural crest cells that give rise
to esophageal neurons may vary between birds and rodent
(see Newgreen and Young, 2002). In mice, a DiI-tracing
study showed that esophageal neurons arise from neural
crest cells that emigrate adjacent to somites 6–7, whereas
intestinal neurons arise from neural crest cells that emigrate
from the hindbrain next to the more rostral somites (Durbec
et al., 1996). Neural crest cells adjacent to somites 6–7 also
give rise to dorsal root ganglia, and thus this level of the
neural axis can also be classified as ‘‘truncal’’ (Durbec et al.,
1996). In contrast, Burns et al. (2000) using cell labelling in
birds suggest that all esophageal neuron arise from the
rostral limit of the vagal region, not from the caudal or
truncal limit.
Because (a) esophageal neurons in embryonic mice arise
from a slightly different rostrocaudal level of the neural axis
from intestinal neurons (Durbec et al., 1996), (b) neurons
have been reported to be present in the esophagus and
stomach but absent elsewhere in the gastrointestinal tract in
Ret�/� mice (Durbec et al., 1996), and (c) neurons are absent
from the esophagus but present elsewhere in the gastrointes-
tinal tract of Mash1�/� mice (Guillemot et al., 1993), it
seemed likely that neural crest-derived cells in the esophagus
may have different trophic requirements and respond to
different signaling pathways from other enteric crest-derived
cells. However, the current study showed that, like intestinal
crest-derived cells, crest-derived cells in the esophagus
showed neurite outgrowth and migratory responses to both
neurturin and GDNF. Thus, at least in the assays performed in
this study, esophageal crest-derived cells behaved like other
enteric crest-derived cells, although the decline in the GDNF-
induced migratory response occurred in the esophagus before
more caudal regions of the gut.
Most esophageal neurons are dependent on Ret
The reported presence of neurons only in the esophagus
of Ret�/� mice, and the absence of neurons only from the
esophagus of Mash1�/� mice (Guillemot et al., 1993), has
lead to the idea that neural crest cells that migrate into the
esophagus have different trophic requirements from neural
crest cells that colonize the rest of the gut (Durbec et al.,
1996). However, Ret appears to be downstream of Mash1 in
signaling cascades (Lo et al., 1998), and if the development
of esophageal neurons were Mash1-dependent but Ret-
independent, it would suggest that there is a signaling
pathway downstream of Mash1, other than Ret, that is
essential for esophageal neuron development. However,
our study showed that there are very few neurons in the
esophagus of E18.5 Ret�/� mice and very few neural
precursors (Phox2b+ cells) in the esophagus of E11.5
Ret�/� mice. It therefore seems likely that few neural crest
cells ever migrate into the esophagus of embryonic Ret�/�
mice, and hence Ret signaling is required by essentially all
neural crest-derived cells destined to colonize the gut. The
small number of neural crest cells that reach the esophagus
and stomach probably only do so because they do not have
to migrate very far to reach these gut regions. Interestingly,
although gut neural crest stem cells are present in the
esophagus of Ret�/� mice, there are four times more stem

H. Yan et al. / Developmental Biology 272 (2004) 118–133132
cells in the esophagus of wild-type mice (Iwashita et al.,
2003).
There are different types of myenteric neurons in the
esophagus. In the current study, we showed that 40% of Hu+
cells expressed NOS in E18.5 heterozygote and wild-type
mice. However, of a total of 121 Hu+ neurons observed in
the esophagus of three Ret�/� mice, only 1 was NOS+. This
suggests either that the differentiation of all esophageal
neurons is delayed in the absence of Ret signaling, or that
Ret is specifically required for the development of NOS
neurons. In the developing esophagus, other major classes
of myenteric neurons (e.g., cholinergic neurons) do not
develop until after birth (Sang et al., 1999), so we were
unable to examine the appearance of other sub-populations
of neurons in Ret null mice.
The main difference between esophageal neurons and
other gastrointestinal neurons appears to be their dependen-
cy on Mash1; the development of all esophageal neurons is
Mash1-dependent, whereas only some sub-populations of
neurons in the intestine require Mash1 for their development
(Blaugrund et al., 1996). In addition to Mash1, Ret can also
be activated by Phox2b (Lo et al., 1998; Pattyn et al., 1999).
Phox2b�/� mice lack enteric neurons in all regions of the
gastrointestinal tract, including the esophagus (Pattyn et al.,
1999). It therefore seems likely that both Mash1 and Phox2b
are required for Ret activation in esophageal neuron pre-
cursors, but only Phox2b is required for Ret activation in
most intestinal neuron precursors.
Acknowledgments
This work was supported by the National Health and
Medical Research Council of Australia. We thank Drs. Jean-
Francois Brunet, Piers Emson, and Miles Epstein for kindly
providing antisera.
References
Airaksinen, M.S., Saarma, M., 2002. The gdnf family: signalling, biolog-
ical functions and therapeutic value. Nat. Rev. Neurosci. 3, 383–394.
Airaksinen, M.S., Titievsky, A., Saarma, M., 1999. GDNF family neuro-
trophic factor signaling: four masters, one servant? Mol. Cell. Neurosci.
13, 313–325.
Barlow, A., de Graaff, E., Pachnis, V., 2003. Enteric nervous system
progenitors are coordinately controlled by the G protein-coupled re-
ceptor EDNRB and the receptor tyrosine kinase RET. Neuron 40,
905–916.
Blaugrund, E., Pham, T.D., Tennyson, V.M., Lo, L., Sommer, L., Ander-
son, D.J., Gershon, M.D., 1996. Distinct subpopulations of enteric neu-
ronal progenitors defined by time of development, sympathoadrenal
lineage markers and Mash-1-dependence. Development 122, 309–320.
Burns, A.J., Le Douarin, N.M., 1998. The sacral neural crest contributes
neurons and glia to the post-umbilical gut: spatiotemporal analysis of
the development of the enteric nervous system. Development 125,
4335–4347.
Burns, A.J., Champeval, D., Le Douarin, N.M., 2000. Sacral neural crest
cells colonise aganglionic hindgut in vivo but fail to compensate for
lack of enteric ganglia. Dev. Biol. 219, 30–43.
Cacalano, G., Farinas, I., Wang, L.C., Hagler, K., Forgie, A., Moore, M.,
Armanini, M., Phillips, H., Ryan, A.M., Reichardt, L.F., Hynes, M.,
Davies, A., Rosenthal, A., 1998. GFRalpha1 is an essential receptor
component for GDNF in the developing nervous system and kidney.
Neuron 21, 53–62.
Chalazonitis, A., Rothman, T.P., Chen, J., Gershon, M.D., 1998. Age-
dependent differences in the effects of GDNF and NT-3 on the devel-
opment of neurons and glia from neural crest-derived precursors immu-
noselected from the fetal rat gut: expression of GFRalpha-1 in vitro and
in vivo. Dev. Biol. 204, 385–406.
Creedon, D.J., Tansey, M.G., Baloh, R.H., Osborne, P.A., Lampe, P.A.,
Fahrner, T.J., Heuckeroth, R.O., Milbrandt, J., Johnson Jr., E.M.,
1997. Neurturin shares receptors and signal transduction pathways with
glial cell line-derived neurotrophic factor in sympathetic neurons. Proc.
Natl. Acad. Sci. U. S. A. 94, 7018–7023.
Durbec, P.L., Larsson-Blomberg, L.B., Schuchardt, A., Costantini, F.,
Pachnis, V., 1996. Common origin and developmental dependence on
c-ret of subsets of enteric and sympathetic neuroblasts. Development
122, 349–358.
Enomoto, H., Araki, T., Jackman, A., Heuckeroth, R.O., Snider, W.D., John-
son Jr., E.M., Milbrandt, J., 1998. GFR alpha1-deficient mice have def-
icits in the enteric nervous system and kidneys. Neuron 21, 317–324.
Enomoto, H., Crawford, P.A., Gorodinsky, A., Heuckeroth, R.O., Johnson
Jr., E.M., Milbrandt, J., 2001. RET signaling is essential for migration,
axonal growth and axon guidance of developing sympathetic neurons.
Development 128, 3963–3974.
Fairman, C.L., Clagett-Dame, M., Lennon, V.A., Epstein, M.L., 1995.
Appearance of neurons in the developing chick gut. Dev. Dyn. 204,
192–201.
Focke, P.J., Schiltz, C.A., Jones, S.E., Watters, J.J., Epstein, M.L., 2001.
Enteric neuroblasts require the phosphatidylinositol 3-kinase pathway
for GDNF-stimulated proliferation. J. Neurobiol. 47, 306–317.
Focke, P.J., Swetlik, A.R., Schilz, J.L., Epstein, M.L., 2003. GDNF and
insulin cooperate to enhance the proliferation and differentiation of
enteric crest-derived cells. J. Neurobiol. 55, 151–164.
Gianino, S., Grider, J.R., Cresswell, J., Enomoto, H., Heuckeroth, R.O.,
2003. GDNF availability determines enteric neuron number by control-
ling precursor proliferation. Development 130, 2187–2198.
Golden, J.P., DeMaro, J.A., Osborne, P.A., Milbrandt, J., Johnson Jr., E.M.,
1999. Expression of neurturin, GDNF, GDNF family-receptor mRNA
in the developing and mature mouse. Exp. Neurol. 158, 504–528.
Golden, J.P., Milbrandt, J., Johnson Jr., E.M., 2003. Neurturin and perse-
phin promote the survival of embryonic basal forebrain cholinergic
neurons in vitro. Exp. Neurol. 184, 447–455.
Guillemot, F., Lo, L.C., Johnson, J.E., Auerbach, A., Anderson, D.J.,
Joyner, A.L., 1993. Mammalian achaete-scute homolog 1 is required
for the early development of olfactory and autonomic neurons. Cell
75, 463–476.
Hearn, C.J., Murphy, M., Newgreen, D., 1998. GDNF and ET-3 differen-
tially modulate the numbers of avian enteric neural crest cells and
enteric neurons in vitro. Dev. Biol. 197, 93–105.
Hearn, C.J., Young, H.M., Ciampoli, D., Lomax, A.E., Newgreen, D.,
1999. Catenary cultures of embryonic gastrointestinal tract support or-
gan morphogenesis, motility, neural crest cell migration, cell differen-
tiation. Dev. Dyn. 214, 239–247.
Heuckeroth, R.O., Lampe, P.A., Johnson, E.M., Milbrandt, J., 1998. Neur-
turin and GDNF promote proliferation and survival of enteric neuron
and glial progenitors in vitro. Dev. Biol. 200, 116–129.
Heuckeroth, R.O., Enomoto, H., Grider, J.R., Golden, J.P., Hanke, J.A.,
Jackman, A., Molliver, D.C., Bardgett, M.E., Snider, W.D., Johnson Jr.,
E.M., Milbrandt, J., 1999. Gene targeting reveals a critical role for
neurturin in the development and maintenance of enteric, sensory, para-
sympathetic neurons. Neuron 22, 253–263.
Honma, Y., Araki, T., Gianino, S., Bruce, A., Heuckeroth, R., Johnson, E.,
Milbrandt, J., 2002. Artemin is a vascular-derived neurotropic factor for
developing sympathetic neurons. Neuron 35, 267–282.
Iwashita, T., Kruger, G.M., Pardal, R., Kiel, M.J., Morrison, S.J., 2003.

H. Yan et al. / Developmental Biology 272 (2004) 118–133 133
Hirschsprung disease is linked to defects in neural crest stem cell func-
tion. Science 301, 972–976.
Jing, S., Yu, Y., Fang, M., Hu, Z., Holst, P.L., Boone, T., Delaney, J.,
Schultz, H., Zhou, R., Fox, G.M., 1997. GFRalpha-2 and GFRalpha-
3 are two new receptors for ligands of the GDNF family. J. Biol. Chem.
272, 33111–33117.
Kapur, R.P., Yost, C., Palmiter, R.D., 1992. A transgenic model for study-
ing development of the enteric nervous system in normal and agan-
glionic mice. Development 116, 167–175.
Kruger, G.M., Mosher, J.T., Tsai, Y.H., Yeager, K.J., Iwashita, T., Gariepy,
C.E., Morrison, S.J., 2003. Temporally distinct requirements for endo-
thelin receptor B in the generation and migration of gut neural crest
stem cells. Neuron 40, 917–929.
Le Douarin, N.M., Teillet, M.A., 1973. The migration of neural crest cells
to the wall of the digestive tract in avian embryo. J. Embryol. Exp.
Morphol. 30, 31–48.
Lo, L., Tiveron, M.C., Anderson, D.J., 1998. MASH1 activates expression
of the paired homeodomain transcription factor Phox2a, and couples
pan-neuronal and subtype-specific components of autonomic neuronal
identity. Development 125, 609–620.
Moore, M.W., Klein, R.D., Farinas, I., Sauer, H., Armanini, M., Phillips, H.,
Reichardt, L.F., Ryan, A.M., Carver-Moore, K., Rosenthal, A., 1996.
Renal and neuronal abnormalities in mice lacking GDNF. Nature 382,
76–79.
Natarajan, D., Marcos-Gutierrez, C., Pachnis, V., de Graaff, E., 2002. Re-
quirement of signalling by receptor tyrosine kinase RET for the directed
migration of enteric nervous system progenitor cells during mammalian
embryogenesis. Development 129, 5151–5160.
Newgreen, D., Young, H.M., 2002. Enteric nervous system: develop-
ment and developmental disturbances—Part 2. Pediatr. Dev. Pathol.
5, 329–349.
Nishino, J., Mochida, K., Ohfuji, Y., Shimazaki, T., Meno, C., Ohishi, S.,
Matsuda, Y., Fujii, H., Saijoh, Y., Hamada, H., 1999. GFR alpha3, a
component of the artemin receptor, is required for migration and sur-
vival of the superior cervical ganglion. Neuron 23, 725–736.
Norris, P.J., Charles, I.G., Scorer, C.A., Emson, P.C., 1995. Studies on the
localization and expression of nitric oxide synthase using histochemical
techniques. Histochem. J. 27, 745–756.
Pattyn, A., Morin, X., Cremer, H., Goridis, C., Brunet, J.F., 1997.
Expression and interactions of the two closely related homeobox
genes Phox2a and Phox2b during neurogenesis. Development 124,
4065–4075.
Pattyn, A., Morin, X., Cremer, H., Goridis, C., Brunet, J.F., 1999. The
homeobox gene Phox2b is essential for the development of autonomic
neural crest derivatives. Nature 399, 366–370.
Pichel, J.G., Shen, L., Sheng, H.Z., Granholm, A.C., Drago, J., Grinberg,
A., Lee, E.J., Huang, S.P., Saarma, M., Hoffer, B.J., Sariola, H., West-
phal, H., 1996. Defects in enteric innervation and kidney development
in mice lacking GDNF. Nature 382, 73–76.
Rakic, P., 1999. Neurobiology. Discriminating migrations. Nature 400,
315–316.
Rossi, J., Luukko, K., Poteryaev, D., Laurikainen, A., Sun, Y.F., Laakso,
T., Eerikainen, S., Tuominen, R., Lakso, M., Rauvala, H., Arumae,
U., Pasternack, M., Saarma, M., Airaksinen, M.S., 1999. Retarded
growth and deficits in the enteric and parasympathetic nervous sys-
tem in mice lacking GFR alpha2, a functional neurturin receptor.
Neuron 22, 243–252.
Rossi, J., Herzig, K.H., Voikar, V., Hiltunen, P.H., Segerstrale, M., Air-
aksinen, M.S., 2003. Alimentary tract innervation deficits and dys-
function in mice lacking GDNF family receptor alpha2. J. Clin.
Invest. 112, 707–716.
Sanchez, M.P., Silos-Santiago, I., Frisen, J., He, B., Lira, S.A., Barbacid,
M., 1996. Renal agenesis and the absence of enteric neurons in mice
lacking GDNF. Nature 382, 70–73.
Sang, Q., Young, H.M., 1997. Development of nicotinic receptor clusters
and innervation accompanying the change in muscle phenotype in the
mouse esophagus. J. Comp. Neurol. 386, 119–136.
Sang, Q., Ciampoli, D., Greferath, U., Sommer, L., Young, H.M., 1999.
Innervation of the esophagus in mice that lack MASH1. J. Comp.
Neurol. 408, 1–10.
Schuchardt, A., D’Agati, V., Larsson-Blomberg, L., Costantini, F., Pachnis,
V., 1994. Defects in the kidney and enteric nervous system of mice
lacking the tyrosine kinase receptor Ret. Nature 367, 380–383.
Shen, L., Pichel, J.G., Mayeli, T., Sariola, H., Lu, B., Westphal, H., 2002.
Gdnf haploinsufficiency causes Hirschsprung-like intestinal obstruction
and early-onset lethality in mice. Am. J. Hum. Genet. 70, 435–447.
Taraviras, S., Marcos-Gutierrez, C.V., Durbec, P., Jani, H., Grigoriou, M.,
Sukumaran, M., Wang, L.C., Hynes, M., Raisman, G., Pachnis, V.,
1999. Signalling by the RET receptor tyrosine kinase and its role in
the development of the mammalian enteric nervous system. Develop-
ment 126, 2785–2797.
Tomac, A.C., Grinberg, A., Huang, S.P., Nosrat, C., Wang, Y., Borlongan,
C., Lin, S.Z., Chiang, Y.H., Olson, L., Westphal, H., Hoffer, B.J., 2000.
Glial cell line-derived neurotrophic factor receptor alpha1 availability
regulates glial cell line-derived neurotrophic factor signaling: evidence
from mice carrying one or two mutated alleles. Neuroscience 95,
1011–1023.
Ward, S.M., Ordog, T., Bayguinov, J.R., Horowitz, B., Epperson, A., Shen,
L., Westphal, H., Sanders, K.M., 1999. Development of interstitial cells
of Cajal and pacemaking in mice lacking enteric nerves. Gastroentero-
logy 117, 584–594.
Widenfalk, J., Nosrat, C., Tomac, A., Westphal, H., Hoffer, B., Olsen,
L., 1997. Neuturin and glial cell line-derived neurotrophic factor
receptor-h, novel proteins related to GDNF and GDNFR-a with
specific cellular patterns of expression suggesting roles in the develo-
ping and adult nervous system and in peripheral organs. J. Neurosci.
17, 8506–8519.
Worl, J., Dutsch, F., Neuhuber, W.L., 2002. Development of neuromuscular
junctions in the mouse esophagus: focus on establishment and reduction
of enteric co-innervation. Anat. Embryol. (Berl.) 205, 141–152.
Worley, D.S., Pisano, J.M., Choi, E.D., Walus, L., Hession, C.A., Cate,
R.L., Sanicola, M., Birren, S.J., 2000. Developmental regulation of
GDNF response and receptor expression in the enteric nervous sys-
tem. Development 127, 4383–4393.
Wu, J.J., Chen, J.X., Rothman, T.P., Gershon, M.D., 1999. Inhibition of in
vitro enteric neuronal development by endothelin-3: mediation by endo-
thelin B receptors. Development 126, 1161–1173.
Xian, C.J., Huang, B.R., Zhou, X.F., 1999. Distribution of neurturin
mRNA and immunoreactivity in the peripheral tissues of adult rats.
Brain Res. 835, 247–258.
Yntema, C.L., Hammond, W.S., 1954. The origin of intrinsic ganglia of
trunk viscera from vagal neural crest in the chick embryo. J. Comp.
Neurol. 101, 515–541.
Young, H.M., Newgreen, D., 2001. Enteric neural crest-derived cells: or-
igin, identification, migration, differentiation. Anat. Rec. 262, 1–15.
Young, H.M., Torihashi, S., Ciampoli, D., Sanders, K.M., 1998. Identifi-
cation of neurons that express stem cell factor in the mouse small
intestine. Gastroenterology 115, 898–908.
Young, H.M., Hearn, C.J., Farlie, P.G., Canty, A.J., Thomas, P.Q., New-
green, D.F., 2001. GDNF is a chemoattractant for enteric neural cells.
Dev. Biol. 229, 503–516.
Young, H.M., Bergner, A.J., Muller, T., 2003. Acquisition of neuronal
and glial markers by neural crest-derived cells in the mouse intestine.
J. Comp. Neurol. 456, 1–11.
Copyright © 2022 FDOKUMEN




















