Nestin Expression in Adult and Developing Human Kidney
12
http://jhc.sagepub.com/ Journal of Histochemistry & Cytochemistry http://jhc.sagepub.com/content/55/4/411 The online version of this article can be found at: DOI: 10.1369/jhc.6A7058.2007 2007 55: 411 J Histochem Cytochem Eugenio Bertelli, Marì Regoli, Luciano Fonzi, Rossella Occhini, Susanna Mannucci, Leonardo Ermini and Paolo Toti Nestin Expression in Adult and Developing Human Kidney Published by: http://www.sagepublications.com On behalf of: Official Journal of The Histochemical Society can be found at: Journal of Histochemistry & Cytochemistry Additional services and information for http://jhc.sagepub.com/cgi/alerts Email Alerts: http://jhc.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: What is This? - Apr 1, 2007 Version of Record >> by guest on May 31, 2013 jhc.sagepub.com Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Nestin Expression in Adult and Developing Human Kidney

http://jhc.sagepub.com/Journal of Histochemistry & Cytochemistry
http://jhc.sagepub.com/content/55/4/411The online version of this article can be found at:
DOI: 10.1369/jhc.6A7058.2007
2007 55: 411J Histochem CytochemEugenio Bertelli, Marì Regoli, Luciano Fonzi, Rossella Occhini, Susanna Mannucci, Leonardo Ermini and Paolo Toti
Nestin Expression in Adult and Developing Human Kidney
Published by:
http://www.sagepublications.com
On behalf of:
Official Journal of The Histochemical Society
can be found at:Journal of Histochemistry & CytochemistryAdditional services and information for
http://jhc.sagepub.com/cgi/alertsEmail Alerts:
http://jhc.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
What is This?
- Apr 1, 2007Version of Record >>
by guest on May 31, 2013jhc.sagepub.comDownloaded from

ARTICLE
Nestin Expression in Adult and Developing Human Kidney
Eugenio Bertelli, Marı Regoli, Luciano Fonzi, Rossella Occhini, Susanna Mannucci,Leonardo Ermini, and Paolo Toti
Department of Pharmacology “Giorgio Segre” (EB,MR,LF), Department of Human Pathology and Oncology,Policlinico Le Scotte (RO,SM,PT), and Department of Evolutionary Biology (LE), University of Siena, Siena, Italy
SUMMARY Nestin is considered a marker of neurogenic and myogenic precursor cells. Itsarrangement is regulated by cyclin-dependent kinase 5 (CDK5), which is expressed in murinepodocytes. We investigated nestin expression in human adult and fetal kidney as well asCDK5 presence in adult human podocytes. Confocal microscopy demonstrated that adultglomeruli display nestin immunoreactivity in vimentin-expressing cells with the podocytemorphology and not in cells bearing the endothelial marker CD31. Glomerular nestin-positivecells were CDK5 immunoreactive as well. Western blotting of the intermediate filament-enriched cytoskeletal fraction and coimmunoprecipitation of nestin with anti-CDK5 anti-bodies confirmed these results. Nestin was also detected in developing glomeruli withinimmature podocytes and a few other cells. Confocal microscopy of experiments conductedwith antibodies against nestin and endothelial markers demonstrated that endothelial cellsbelonging to capillaries invading the lower cleft of S-shaped bodies and the immature glo-meruli were nestin immunoreactive. Similar experiments carried out with antibodies raisedagainst nestin and a-smooth muscle actin showed that the first mesangial cells that populatethe developing glomeruli expressed nestin. In conclusion, nestin is expressed in the humankidney from the first steps of glomerulogenesis within podocytes, mesangial, and endothelialcells. This expression, restricted to podocytes in mature glomeruli, appears associated withCDK5. (J Histochem Cytochem 55:411–421, 2007)
KEY WORDS
nestin
podocytes
cyclin-dependent kinase 5
CD31
a-smooth muscle actin
immunohistochemistry
confocal microscopy
THE RENAL CORPUSCLE consists of a tuft of blood capil-laries surrounded by Bowman’s capsule, a bowl-shapeddouble-layered epithelial structure. The two epitheliallayers of the capsule, though continuous at the vascularpole of the corpuscle, are separated by an interval, theurinary space, into which the primary urine is ultra-filtrated. The inner layer of the capsule, tightly appliedto the blood capillaries, is also referred to as the viscerallayer and is formed of highly specialized epithelial cellscalled podocytes because of their peculiar shape. Alongwith the basement membrane and the fenestratedendothelial cells, these cells constitute the glomerularfiltration barrier, whose integrity is essential for thefiltration and proper composition of the primary urine.Several functions have been assigned to podocytes,including key roles in determining the permeability
properties of the glomerular filter, in counteractingintraglomerular hydrostatic pressure, and in the syn-thesis of basement membrane components (Pavenstadtet al. 2003). Even though some of these functions aremore supposed than definitely proven (Pavenstadt et al.2003), the importance of podocytes in maintainingglomerular homeostasis and proper renal function istestified to by the development of proteinuria and pro-gressive glomerulosclerosis upon their injury (Kriz et al.1994; Ruggenenti et al. 2001; Gubler 2003). Podocytesdisplay a very complex morphology that must be pre-served to accomplish their delicate functions (Barietyet al. 1998; Nakahama et al. 1999; Hir et al. 2001;Kerajaschki 2001). The shape of podocytes is suigeneris, with a cell body that protrudes into the capsulelumen and long primary processes anchored to thebasement membrane by extending secondary processesreferred to as foot processes. The foot processes of ad-jacent podocytes interdigitate on the surface of thebasement membrane, leaving a space between them, thefiltration slit, which is occupied by a thin diaphragm.The complex and ramified shape of podocytes has led
Correspondence to: Eugenio Bertelli, Dept. of Pharmacology“Giorgio Segre”, Section of Anatomy, University of Siena, Via AldoMoro 4, I-53100 Siena, Italy. E-mail: [email protected]
Received for publication July 13, 2006; accepted December 20,2006 [DOI: 10.1369/jhc.6A7058.2007].
TheJournal
ofHistoch
emistry&
Cytoch
emistry
C The Histochemical Society, Inc. 0022-1554/07/$3.30 411
Volume 55(4): 411–421, 2007
Journal of Histochemistry & Cytochemistry
http://www.jhc.org
by guest on May 31, 2013jhc.sagepub.comDownloaded from

some investigators to compare them with neurons. In-deed, an increasing number of features are describedas being shared by the two cell types. First, podocytesand neurons are postmitotic terminally differentiatedcells (Nagata et al. 1998; Gubler 2003); second, theyexpress common sets of proteins including synaptopodin(Mundel et al. 1997; Czarnecki et al. 2005), nephrin(Putaala et al. 2001), cyclin-dependent kinase 5 (CDK5)(Griffin et al. 2004), olfactomedin-related glycoprotein(Kondo et al. 2000), a type III receptor protein tyro-sine phosphatase (Beltran et al. 2003), and Rab3A andRabphilin-3a (Rastaldi et al. 2003). Third, some tran-scription factors such as N-myc (Hirvonen et al. 1989)and Wilms’ tumor suppressor protein (Armstrong et al.1992) are at least transiently expressed either in neuronsor podocytes. Finally, if on one hand the mechanism forthe formation of podocyte processes is somehow similarto the development of neuronal dendrites (Kobayashiet al. 2004), on the other hand it has recently been shownthat podocytes possess neuronal-like functional synapticvesicles (Rastaldi et al. 2006). In this respect, the factthat CDK5 has been shown to regulate podocyte mor-phology (Griffin et al. 2004), coupled with the findingthat CDK5 in neuronal progenitor ST15A cells and inC2C12 myoblasts is associated with nestin and governsits arrangement (Sahlgren et al. 2003), prompted us toinvestigate if nestin was expressed by human podocytesas well. Here we report that nestin is present in humanpodocytes, is associated with CDK5 and vimentin, andrepresents an early marker of developing glomeruliwhere it can be detected in the vesicles (the first epithe-lial structure consisting of polarized cells) and in thepodocyte layer of the S-shaped bodies, in endothelialcells of blood capillaries invading the lower cleft of theS-shaped bodies and, finally, in mesangial precursor cells.
Materials and Methods
Antibodies
Primary antibodies were purchased as follows: mouse anti-nestinmonoclonal antibody (MAb5326) and rabbit anti-nestinpolyclonal antibody (Ab5922; Chemicon, Temecula, CA);mouse anti-SMA MAb (clone 1A4; Neomarkers, Fremont,CA); mouse anti-CD31 (clone JC70A) and anti-CD34 MAbs(clone QBEnd-10) from DakoCytomation (Glostrup, Den-mark); mouse anti-vimentin MAb (clone V9; Sigma, St Louis,MO); and rabbit anti-CDK5 PAbC-8 and rabbit anti-vimentin(H-84) (Santa Cruz Biotechnology; Santa Cruz, CA).
Secondary antibodies were obtained from the followingsources: double-labeling-grade tetramethylrhodamine iso-thiocyanate (TRITC)-conjugated donkey anti-mouse IgG andTRITC-conjugated donkey anti-rabbit IgG from Chemicon;fluorescein isothiocyanate-conjugated (FITC) goat anti-mouseIgG antibody from Sigma; double-labeling-grade horseradishperoxidase (HRP)-conjugated goat anti-rabbit IgG from Zymed(South San Francisco, CA); and rabbit anti-mouse IgG fromDakoCytomation.
Tissues
Cases and controls were taken consecutively and retrospec-tively from the surgical and autoptic files of the department ofHuman Pathology and Oncology. Adult kidney samples wereobtained from surgical nephrectomy for carcinoma (n510,age range 24–78 years, median age 60 years). Kidneys werefixed in 4%buffered formalin for 24–48 hr, and normal tissue,far from the neoplasia, was sampled and paraffin embedded.
Fetal kidneys were obtained from autopsies for spontane-ous abortion or induced termination of pregnancy (TP)(n523; gestational age range, 16–37 weeks; median gesta-tional age 22 weeks). Macerated and malformed stillbirthswere not included in the study. The cause of spontaneous fetaldeath was placental in origin and included abruptio pla-centae, true cord knot, acute massive placenta infarcts, acutechorioamnionitis; and conditions associated with prolongedstress such as fetal growth restriction and other morpholog-ical signs of chronic suffering, as well as maternal preeclamp-sia and eclampsia, were excluded from this study. TPstemmed from either minor malformations unrelated to alter-ations in the urinary system or maternal (personal) reasons. Acomplete autopsy was carried out between the second andtwelfth hour after fetal expulsion.
Selected samples for biochemical analysis were immediatelyfrozen in liquidnitrogen and stored at280C.Other fresh tissues(only from adult kidneys) were sampled for immunofluores-cence histochemistry, cryoprotected with Killik Frozen Me-dium (Bio Optica; Milan, Italy), and promptly frozen in liquidnitrogen. Ten-mm-thick sections were cut at 225C, air-dried,fixed in acetone at 220C for 10 min, and stored at280C.
Immunohistochemistry
Immunohistochemistry to detect nestin was performed as pre-viously described (Toti et al. 2005). Three- to 5-mm sectionswere cut from each specimen, mounted on electrostaticallycharged slides, and dried. Sections were dewaxed, rehydrated,and washed in TBS. TBS was used for all subsequent washesand for antibody dilutions. Nestin antigenic sites were un-masked by heating sections in 0.01 M citrate buffer (pH 6.0)with a microwave oven (two 5-min cycles at 750 W) and weresubsequently rinsed for 10 min in 3% hydrogen peroxide toblock endogenous peroxidase. Slides were then incubated for1 hr at room temperature with anti-nestin MAb (workingdilution 1:200). The ABC system (Vector Laboratories; Burlin-game, CA) was applied on sections of fetal samples, and thereaction was developed with 1 mg/ml 3,3¶-diaminobenzidinetetrahydrochloride (Sigma) dissolved in TBS containing 0.3%H2O2. Sections of adult kidneys were incubated with a bridgeantibody anti-mouse IgG for 30 min at room temperature, andthe reactions were developed by the alkaline phosphatase–anti-alkaline phosphatase technique (Dakopatts; DakoCytomation)with new fuchsin. Harris hematoxylin was used for nuclearcounterstaining. Negative controls were prepared by substitut-ing the primary antibody with the corresponding preimmuneserum. Cognate sections were stained with hematoxylin andeosin to check for renal pathologies or malformations.
Confocal Microscopy
A TCS 4D Leica (Wetzlar, Germany) confocal microscopewas used to evaluate double-labeling experiments carried out
TheJournal
ofHistoch
emistry&
Cytoch
emistry
412 Bertelli, Regoli, Fonzi, Occhini, Mannucci, Ermini, Toti
by guest on May 31, 2013jhc.sagepub.comDownloaded from

on frozen sections of adult kidney and on formalin-fixed,paraffin-embedded sections of fetal kidney.
Frozen sections were incubated 2 hr at room temperaturewith anti-vimentin MAb (working dilution 1:50) or anti-CD31 MAb (working dilution 1:100) followed by FITC-conjugated anti-mouse IgG for 1 hr at room temperature; thesecond primary antibody, anti-nestin PAb (working dilution1:200), was applied overnight at 4C followed for 1 hr at roomtemperature by the appropriate TRITC-conjugated secondaryantibody (working dilution 1:50). Other experiments werecarried out incubating sections with anti-nestin MAb (work-ing dilution 1:100) or anti-CD31 MAb (working dilution1:100) for 2 hr at room temperature, followed for 1 hr byTRITC-conjugated anti-mouse IgG (working dilution 1:50).Rabbit anti-CDK5 was then applied overnight at 4C, fol-lowed by HRP-conjugated anti-rabbit IgG for 1 hr at roomtemperature. Application of fluorescein-labeled tyramide am-plification reagent (Perkin-Elmer; Boston, MA) catalyzed aHRP-driven reaction that resulted in the deposition of afluorescent precipitate.
Sections of embryonic kidney were dewaxed, rehydrated,and processed as follows: they were incubated with rabbitanti-nestin PAb for 1 hr at room temperature followed by theABC system according to manufacturer’s instructions (VectorLaboratories); the second primary antibody, anti-CD34 MAb(working dilution 1:100) or anti-SMA MAb (working dilu-tion 1:200), was applied overnight at 4C followed for 1 hr atroom temperature by TRITC-conjugated anti-mouse IgG(working dilution 1:50). Final application of fluorescein-labeled tyramide amplification reagent (Perkin-Elmer) cata-lyzed a HRP-driven reaction that resulted in the deposition ofa fluorescent precipitate on nestin antigenic sites.
SDS/PAGE Electrophoresis, Coimmunoprecipitation,and Western Blotting
To confirm our immunohistochemical results, Western blot-ting analysis with anti-nestin antibody was carried out onproteins of the intermediate filament-enriched cytoskeletal frac-tion (IFCF) of adult and fetal kidney, separated by SDS-PAGE.IFCFs were prepared as previously reported (Achtstaetteret al. 1986; Carapelli et al. 2004; Toti et al. 2005) and storedat 280C until use. Just before use, aliquots of IFCF werethawed and mixed with Laemmli sample buffer.
Immunoprecipitation experiments were carried out ontotal homogenates of adult renal cortex. Pieces of renal cortexwere placed in homogenization buffer (50 mM Tris/HCl,150 mM NaCl, 1 mM EDTA, 0.25% sodium deoxycholate,1% NP-40, 1 mM PMSF, 2 mg/ml aprotinin, pH 7.4) andhomogenized with a Dounce homogenizer. The suspensionwas centrifuged at 50003 g for 10 min at 4C, and the proteincontent was measured with the microBCA protein assay re-agent kit (Pierce; Rockford, IL). Samples were stored at280Cuntil use. Five hundred ml of homogenate was precleared withprotein A–Sepharose (Sigma), centrifuged, and the pellet ofSepharose beads was placed in Laemmli sample buffer. Twoml of anti-CDK5 IgG or 3 ml of anti-vimentin PAb was addedto the supernatant and incubated overnight at 4C, followed by2 hr with the protein A–Sepharose. Immunocomplexes werewashed three times with buffer, and the final pellet wasresuspended in Laemmli sample buffer.
IFCFs, immunoprecipitates, and proteins released from theProtein A–Sepharose complexes used for the preclearings wereseparated by electrophoresis through a 6%polyacrylamide geland transferred to nitrocellulose in a Bio-Rad Transblot appa-ratus (BioRad; Segrate, Italy). A 5% (w/v) solution of skimmilk in TBS was used to quench nonspecific protein binding.Membranes were incubated overnight at room temperaturewith 1:2000 anti-nestin MAb in 5% (w/v) solution of skimmilk in TBS. Specific bands were detected using an electro-chemiluminescense kit (Roche Diagnostics; Milan, Italy).
Results
Adult Kidney
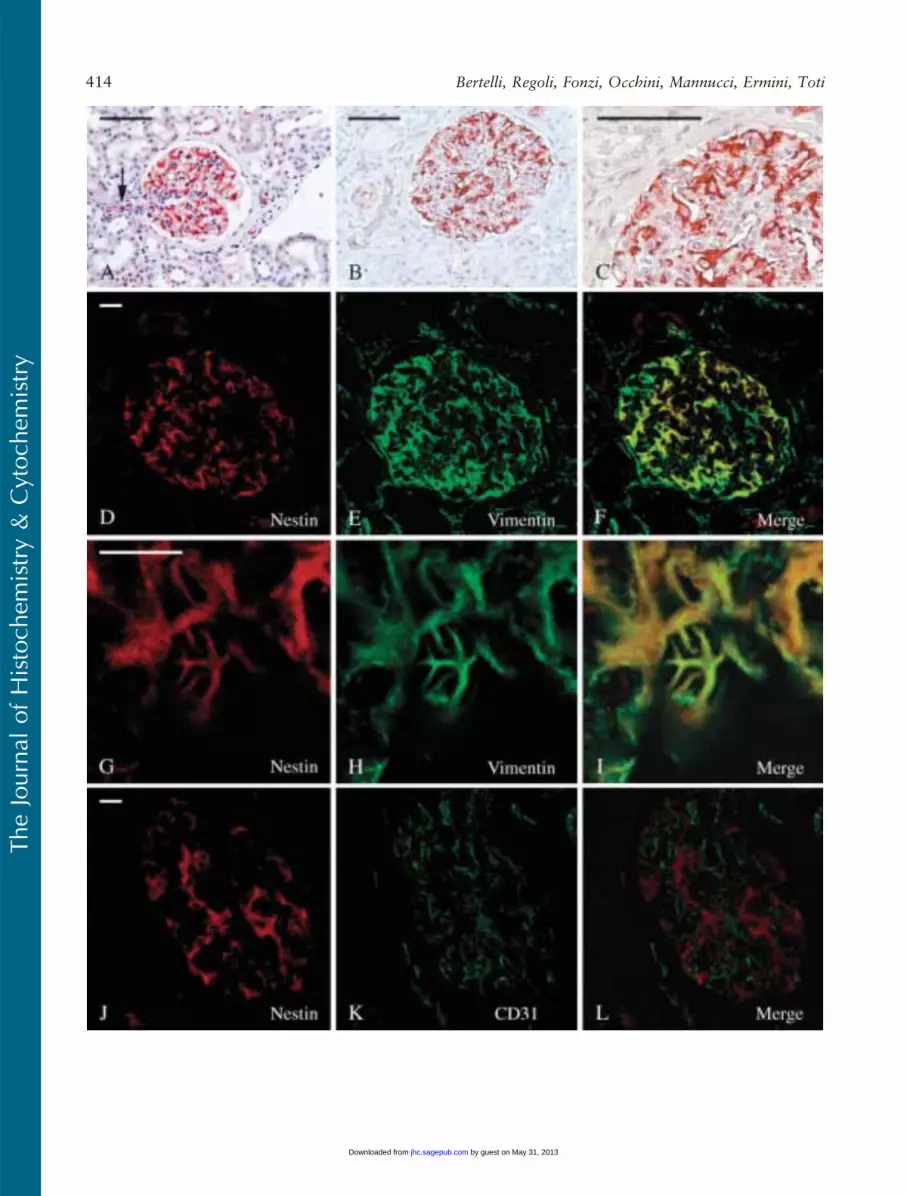
In adult kidney, nestin staining was restricted to glo-meruli, smooth muscle cells of the arterioles, and someendothelial cells (Figures 1A–1C). The highest intensityof staining was referred to podocytes where nestinimmunoreactivity was consistent (Figures 1A–1C) andto the tunica media of some arterioles. Even thoughwith a variable degree of intensity, endothelial cells ofarterioles and of some blood capillaries were nestinlabeled as well (Figure 1A).
Glomerular nestin-expressing cells resembled podo-cytes as to their location and morphology. However,to confirm immunohistochemical data obtained onformalin-fixed, paraffin-embedded material, a series ofdouble-immunofluorescence experiments were per-formed on frozen sections and analyzed with the con-focal microscope. A well-known marker of podocytes,vimentin, was used along with nestin in double-labelingexperiments. In the glomerulus, vimentin is expressedat high levels only within podocytes, mesangial, and en-dothelial cells being reported as showing possibly only amuch weaker labeling (Bachmann et al. 1983; Holthoferet al. 1984; Stamenkovic et al. 1986). Other podocytemarkers like podocin, synaptopodin, nephrin, ZO-1, andpodocalixyn could not be used for our purpose in vivobecause theywere all concentrated prevalently in the footprocesses where intermediate filaments are not present,and their staining hardly overlapped with nestin. Theseexperiences showed that all nestin-immunoreactive cellswere, indeed, highly vimentin-expressing cells as well(Figures 1D–1F). Moreover, nestin/vimentin double-labeled cells clearly resembled podocytes, displaying longimmunoreactive primary processes (Figures 1G–1I).Double-labeling experiments carried out using nestinand the endothelial marker CD31 (Naruse et al. 2000)showed that the relationships occurring between nestin-positive cells and endothelial cells were those expectedbetween podocytes and the glomerular capillaries(Figures 1J–1L).
Because it has been reported that CDK5 is expressedin murine podocytes (Griffin et al. 2004), and that itplays a key role in the regulation of nestin organiza-tion (Sahlgren et al. 2003), we performed CDK5/nestindouble-labeling experiments that showed a certain degree
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Nestin Expression in Human Kidney 413
by guest on May 31, 2013jhc.sagepub.comDownloaded from
TheJournal
ofHistoch
emistry&
Cytoch
emistry
414 Bertelli, Regoli, Fonzi, Occhini, Mannucci, Ermini, Toti
by guest on May 31, 2013jhc.sagepub.comDownloaded from

of colocalization of the two proteins (Figures 2A–2F).Moreover, because CDK5 has been previously reportedin endothelial cells (Sharma et al. 2004), additionalexperiments have been carried out to rule out the pos-sibility that CDK5 staining had to be referred in part tothe glomerular endothelium. Indeed, confocal micros-copy of CDK5/CD31 double-labeled sections showedtwo different staining patterns with no detectable colo-calizations of the two antigens in the renal corpuscles(Figures 2G–2I). Remarkably, CDK5 fluorescence ap-peared frequently arranged parallel to the glomerularendothelium (Figure 2I).
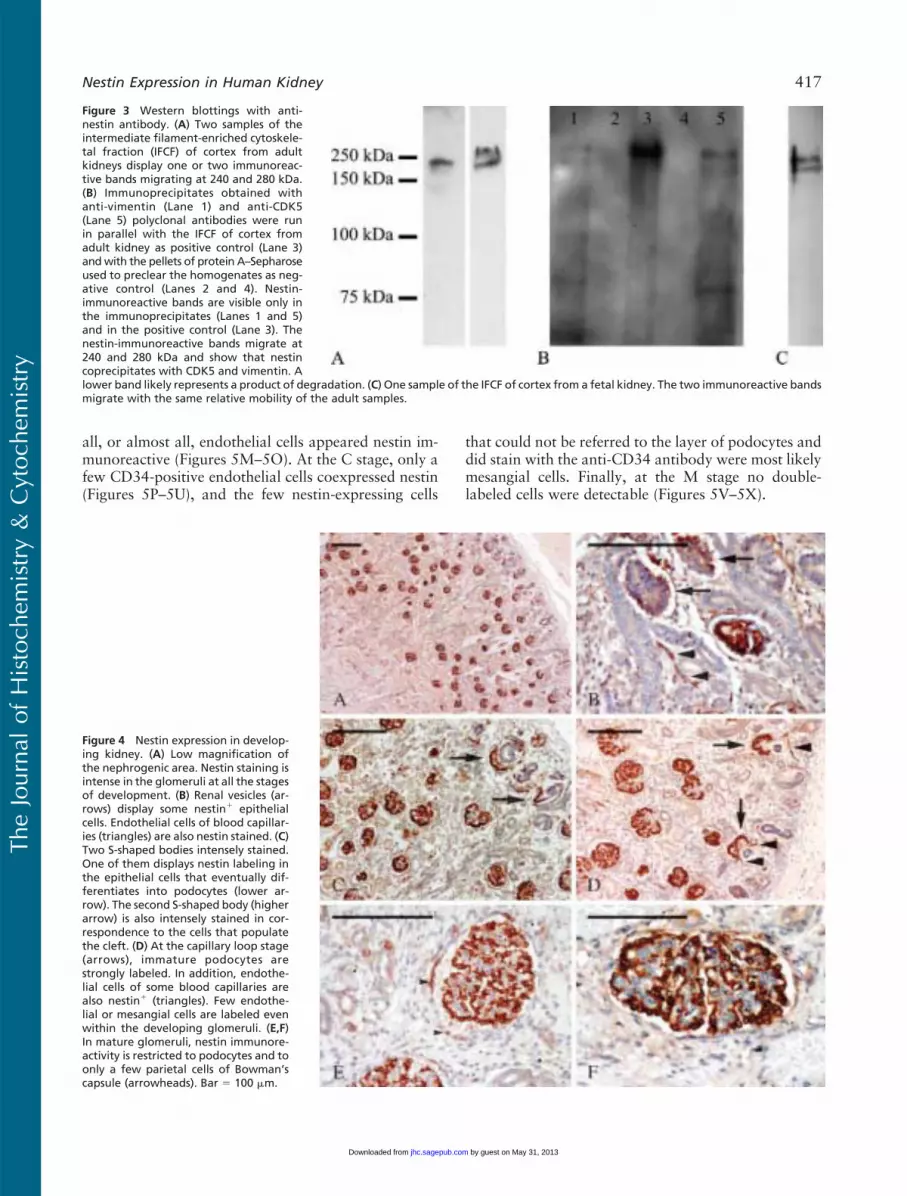
To confirm that the antibodies used in this studyrecognized actual nestin, we carried out Western blotanalysis with anti-nestin MAb on renal IFCF of adultindividuals from total homogenates of adult kidneys.Depending on samples, one or two nestin-immuno-reactive bands could be unveiled in the IFCF migratingat 240 and 280 kDa (Figure 3A), the lower band beingconstant. Moreover, to confirm that CDK5/nestin andnestin/vimentin colocalizations were due to their actualassociation, additional Western blotting experimentswith anti-nestin MAb were carried out on CDK5 andvimentin immunoprecipitates. The presence of a double-nestin-immunoreactive band of the appropriate molec-ular mass (240 and 280 kDa) in the immunoprecipitatesdemonstrated that nestin coimmunoprecipitated withvimentin and CDK5, confirming on the one hand thatnestin and vimentin coassemble together even in podo-cytes and suggesting on the other hand that the asso-ciation of nestin andCDK5observed in C2C12myoblastsand ST15A neuronal precursors (Sahlgren et al. 2003)occurs in human podocytes as well (Figure 3B).
Fetal Kidney
Development of human metanephric kidney begins atgestational week 4–5 and is said to terminate after the35th week of gestation (Nagata et al. 1993). In thisperiod a nephrogenic zone located under the renal cap-sule continues to add new developing glomeruli, theolder being located deeper in the kidney and the newerjust under the capsule. For this reason, in a single sec-tion of fetal kidney it is possible to appreciate all thestages of glomerular development in a sort of gradientof maturation going from the surface to the depth of theorgan. The steps of glomerular maturation are mor-phologically divided into the following stages: vesicle
(V), S-shaped body (S), capillary loop (C), andmaturingglomerulus (M) (Naruse et al. 2000; Pavenstadt et al.2003). At low magnification, nestin staining on fetalkidney sections clearly appeared located in glomeruli atall the maturation stages (Figure 4A). Although lessconstantly, a certain degree of staining was apparenteven at the V stage (Figure 4B). Then, at the S stage,nestin labeling became consistent and intense within thecell layer that eventually would differentiate into podo-cytes (Figure 4C). At the same stage, nestin was alsoclearly locatedwithin cells that populated the cleft of theS-shaped body, from which both mesangial and endo-thelial glomerular cells are known to differentiate.At the C stage, podocytes were the only glomerularcell elements incontrovertibly nestin immunoreactive(Figure 4D). However, a certain degree of nestin stainingin presumptive mesangial cells was also seen at thisstage. At the M stage, some cells of the parietal layerof the Bowman’s capsule were nestin immunoreactive(Figure 4E), whereas all podocytes were morphologi-cally well developed and maintained their intense nestinstaining (Figures 4E and 4F). Outside the glomeruli,nestin was expressed by capillary endothelial cells(Figures 4B and 4D) and smooth muscle cells of thearterial wall (data not shown). To corroborate nestinexpression in developing kidneys, the IFCFs of fetalkidneyswere immunoblottedwith aMAbagainst nestin.A double-immunoreactive band comigrating with nestinfrom adult kidneys was observed in several preparations(Figure 3C). When bands of the proper molecular masswere not detectable, they were always substituted byimmunoreactive bands of lower molecular mass (datanot shown), likely representing degradation products aspreviously reported (Sahlgren et al. 2001).
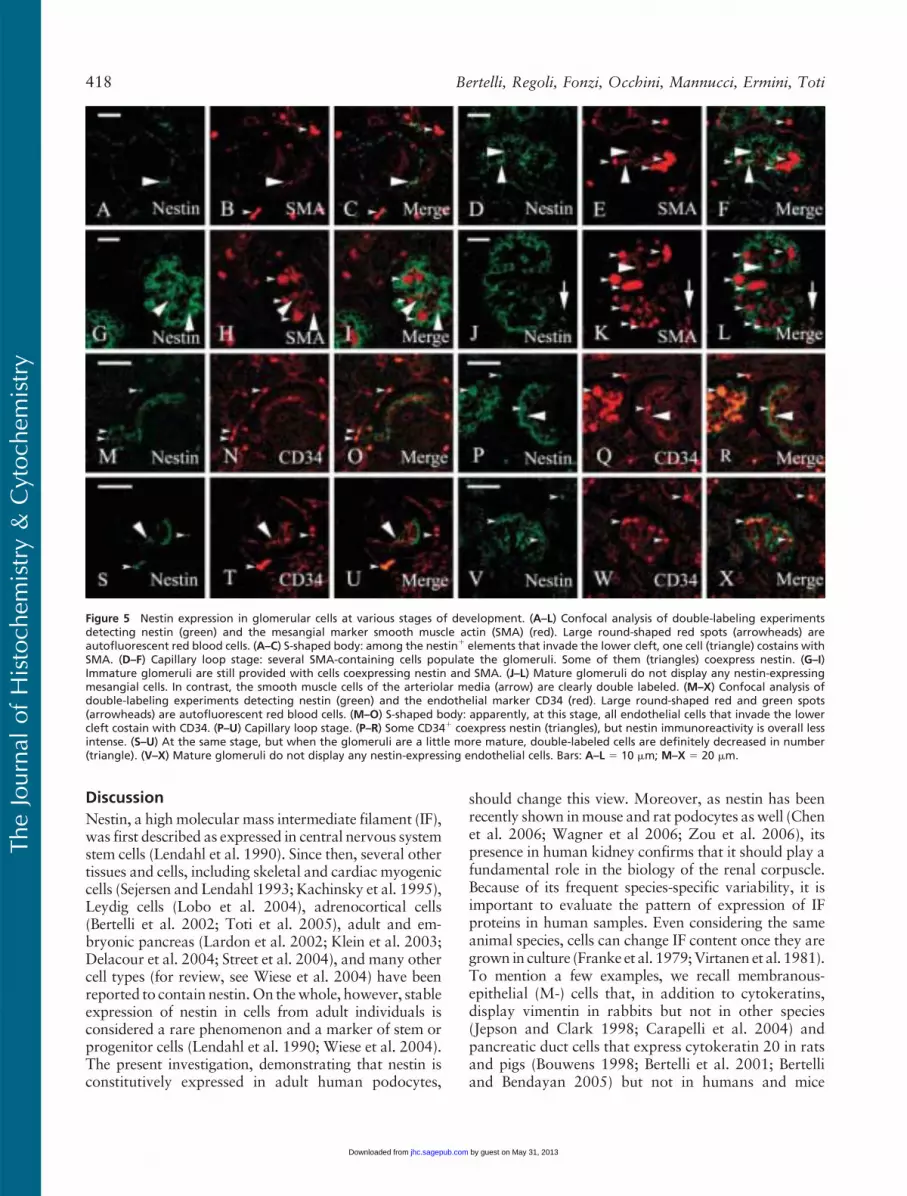
Confocal microscopy analysis of double-labeling ex-periments was carried out to identify nestin1 cells in theearly stages of nephrogenesis. As a mesangial marker,we chose a-smooth muscle actin (a-SMA), which hasbeen reported to be expressed in activated and fetalmesangial cells (Alpers et al. 1992; Naruse et al. 2000),whereas we used CD34 as a marker for the endothelium(Fina et al. 1990; Ito et al. 1995; Naruse et al. 2000).a-SMAwas constantly expressed in the S (Figure 5B), C(Figure 5E), and M (Figures 5H and 5K) stages. Nestin/SMA double-labeled cells were visible at the S (Figures5A–5C) and C (Figures 5D–5F) stages and within thesmaller glomeruli (Figures 5G–5I) at the M stage. Mid-
’
Figure 1 Nestin expression in adult kidney. (A–C) Nestin staining is located within glomeruli. (A) A glomerulus with an arteriole at the vascularpole (arrow). Staining is observed in the endothelial cells of the arteriole. (B,C) A glomerulus at low (B) and higher magnification (C). Staining isrestricted to cells with the morphological features of podocytes. (D–L) Confocal microscopy analysis of nestin/vimentin (D–I) and nestin/CD31(J–L) double-labeling experiments. (D–F) All nestin1 cells (red) costain with vimentin (green). (G–I) The morphology of nestin/vimentin double-labeled cells is typical of podocytes, with long and branched primary processes. (J–L) Endothelial CD311 cells do not costain with nestin. Bars:A–C 5 100 mm; D–L 5 10 mm.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Nestin Expression in Human Kidney 415
by guest on May 31, 2013jhc.sagepub.comDownloaded from

sized glomeruli did not show any overlapping betweenSMA and nestin stainings, whereas smooth muscle cellsof the afferent and efferent arterioles expressed bothantigens (Figures 5J–5L).
CD34 appeared early in the developing glomeruliat the S stage. It is believed that as soon as the lowercleft of the S-shaped body is formed, blood capillaries
invade it or differentiate in loco from mesenchymalprecursors (Naruse et al. 2000). In our preparations,CD34-positive blood capillaries were already visible atthis stage (Figure 5N). Glomeruli at later stages of matu-ration displayed an increasing complex tuft of capillaryloops (Figures 5Q and 5T). Double-labeled CD34/nestin cells were detected in the S-shaped bodies where
Figure 2 Nestin/CDK5 colocalization. Confocal microscopy of sections from adult human kidney. (A–F) Nestin/CDK5 double labeling. (A–C)Low magnification showing an entire glomerulus. CDK5 appears expressed in nestin1 cells. (D–F) Higher magnification. Apparently, whereasnestin staining is restricted to the cell bodies and primary processes of podocytes, CDK5 labeling is extended throughout their entire cytoplasm,probably involving foot processes that cover capillary loops. (G–I) CDK5/CD31 double labeling. The stainings do not overlap. On the abluminalside of the glomerular endothelium, a thin CDK51 thread occupying the typical position of pedicels covers the capillary loops. Bar 5 10 mm.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
416 Bertelli, Regoli, Fonzi, Occhini, Mannucci, Ermini, Toti
by guest on May 31, 2013jhc.sagepub.comDownloaded from
all, or almost all, endothelial cells appeared nestin im-munoreactive (Figures 5M–5O). At the C stage, only afew CD34-positive endothelial cells coexpressed nestin(Figures 5P–5U), and the few nestin-expressing cells
that could not be referred to the layer of podocytes anddid stain with the anti-CD34 antibody were most likelymesangial cells. Finally, at the M stage no double-labeled cells were detectable (Figures 5V–5X).
Figure 3 Western blottings with anti-nestin antibody. (A) Two samples of theintermediate filament-enriched cytoskele-tal fraction (IFCF) of cortex from adultkidneys display one or two immunoreac-tive bands migrating at 240 and 280 kDa.(B) Immunoprecipitates obtained withanti-vimentin (Lane 1) and anti-CDK5(Lane 5) polyclonal antibodies were runin parallel with the IFCF of cortex fromadult kidney as positive control (Lane 3)andwith the pellets of protein A–Sepharoseused to preclear the homogenates as neg-ative control (Lanes 2 and 4). Nestin-immunoreactive bands are visible only inthe immunoprecipitates (Lanes 1 and 5)and in the positive control (Lane 3). Thenestin-immunoreactive bands migrate at240 and 280 kDa and show that nestincoprecipitates with CDK5 and vimentin. Alower band likely represents a product of degradation. (C) One sample of the IFCF of cortex from a fetal kidney. The two immunoreactive bandsmigrate with the same relative mobility of the adult samples.
Figure 4 Nestin expression in develop-ing kidney. (A) Low magnification ofthe nephrogenic area. Nestin staining isintense in the glomeruli at all the stagesof development. (B) Renal vesicles (ar-rows) display some nestin1 epithelialcells. Endothelial cells of blood capillar-ies (triangles) are also nestin stained. (C)Two S-shaped bodies intensely stained.One of them displays nestin labeling inthe epithelial cells that eventually dif-ferentiates into podocytes (lower ar-row). The second S-shaped body (higherarrow) is also intensely stained in cor-respondence to the cells that populatethe cleft. (D) At the capillary loop stage(arrows), immature podocytes arestrongly labeled. In addition, endothe-lial cells of some blood capillaries arealso nestin1 (triangles). Few endothe-lial or mesangial cells are labeled evenwithin the developing glomeruli. (E,F)In mature glomeruli, nestin immunore-activity is restricted to podocytes and toonly a few parietal cells of Bowman’scapsule (arrowheads). Bar 5 100 mm.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Nestin Expression in Human Kidney 417
by guest on May 31, 2013jhc.sagepub.comDownloaded from
Discussion
Nestin, a high molecular mass intermediate filament (IF),was first described as expressed in central nervous systemstem cells (Lendahl et al. 1990). Since then, several othertissues and cells, including skeletal and cardiac myogeniccells (Sejersen and Lendahl 1993; Kachinsky et al. 1995),Leydig cells (Lobo et al. 2004), adrenocortical cells(Bertelli et al. 2002; Toti et al. 2005), adult and em-bryonic pancreas (Lardon et al. 2002; Klein et al. 2003;Delacour et al. 2004; Street et al. 2004), and many othercell types (for review, see Wiese et al. 2004) have beenreported to contain nestin.On thewhole, however, stableexpression of nestin in cells from adult individuals isconsidered a rare phenomenon and a marker of stem orprogenitor cells (Lendahl et al. 1990; Wiese et al. 2004).The present investigation, demonstrating that nestin isconstitutively expressed in adult human podocytes,
should change this view. Moreover, as nestin has beenrecently shown inmouse and rat podocytes aswell (Chenet al. 2006; Wagner et al 2006; Zou et al. 2006), itspresence in human kidney confirms that it should play afundamental role in the biology of the renal corpuscle.Because of its frequent species-specific variability, it isimportant to evaluate the pattern of expression of IFproteins in human samples. Even considering the sameanimal species, cells can change IF content once they aregrown in culture (Franke et al. 1979;Virtanen et al. 1981).To mention a few examples, we recall membranous-epithelial (M-) cells that, in addition to cytokeratins,display vimentin in rabbits but not in other species(Jepson and Clark 1998; Carapelli et al. 2004) andpancreatic duct cells that express cytokeratin 20 in ratsand pigs (Bouwens 1998; Bertelli et al. 2001; Bertelliand Bendayan 2005) but not in humans and mice
Figure 5 Nestin expression in glomerular cells at various stages of development. (A–L) Confocal analysis of double-labeling experimentsdetecting nestin (green) and the mesangial marker smooth muscle actin (SMA) (red). Large round-shaped red spots (arrowheads) areautofluorescent red blood cells. (A–C) S-shaped body: among the nestin1 elements that invade the lower cleft, one cell (triangle) costains withSMA. (D–F) Capillary loop stage: several SMA-containing cells populate the glomeruli. Some of them (triangles) coexpress nestin. (G–I)Immature glomeruli are still provided with cells coexpressing nestin and SMA. (J–L) Mature glomeruli do not display any nestin-expressingmesangial cells. In contrast, the smooth muscle cells of the arteriolar media (arrow) are clearly double labeled. (M–X) Confocal analysis ofdouble-labeling experiments detecting nestin (green) and the endothelial marker CD34 (red). Large round-shaped red and green spots(arrowheads) are autofluorescent red blood cells. (M–O) S-shaped body: apparently, at this stage, all endothelial cells that invade the lowercleft costain with CD34. (P–U) Capillary loop stage. (P–R) Some CD341 coexpress nestin (triangles), but nestin immunoreactivity is overall lessintense. (S–U) At the same stage, but when the glomeruli are a little more mature, double-labeled cells are definitely decreased in number(triangle). (V–X) Mature glomeruli do not display any nestin-expressing endothelial cells. Bars: A–L 5 10 mm; M–X 5 20 mm.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
418 Bertelli, Regoli, Fonzi, Occhini, Mannucci, Ermini, Toti
by guest on May 31, 2013jhc.sagepub.comDownloaded from

(Bouwens 1998). However, many other cases could belisted and even podocytes differ in IF content from oneanimal species to another (Yaoita et al. 1999; Zou et al.2006) showing, accordingly, that the molecular arrange-ment of the IF network can be quite diverse, depending onthe animals. Nestin is expressed in undifferentiated anddividing cells in the central nervous system and in myo-genic tissues (Lendahl et al. 1990; Sejersen and Lendahl1993). Upon differentiation, nestin is downregulated andreplaced by other IFs. Nevertheless, it is re-expressedupon injury and in regenerating phenomena (Frisen et al.1995; Vaittinen et al. 2001). These features have led someinvestigators to propose nestin as amarkerofmultilineageprogenitor cells (Wiese et al. 2004). Nestin expression hasalso been correlated to increased motility and tumorinvasiveness (Dahlstrand et al. 1992; Thomas et al. 2004).It is evident that podocytes cannot be placed into such aframework. Podocytes are non-motile, postmitotic termi-nally differentiated cells that re-enter the cell cycle onlyin a limited set of diseases, namely, HIV glomerulopathy,collapsing glomerulopathy, and the cellular variant of focalsegmental glomerulosclerosis (Pavenstadt et al. 2003).
Nestin has been functionally linked to CDK5. CDK5seems to play a fundamental role in triggering the exitof neurons from the cell cycle and in inducing differen-tiation (Cicero and Herrup 2005). In addition, CDK5is important in regulating the differentiation of musclecells and the rearrangement of the nestin network(Lazaro et al. 1997; Sahlgren et al. 2003). To carry outits kinase activity, CDK5 requires an activator, p35,which has been reported to associate with nestin as well(Sahlgren et al. 2003). When activated, CDK5 phos-phorylates nestin and, in this way, probably regulatesnestin disassembling because phosphorylated nestinis detected exclusively in the Triton X-100 soluble frac-tion (Sahlgren et al. 2003). Our results demonstratefor the first time that in vivo nestin and CDK5 are co-expressed and associated in adult human podocytes.Moreover, our data show that, whereas CDK5 is dif-fuse throughout the entire cytoplasm, pedicels included,the nestin network is located in the cell body and in theprimary processes, but not in the foot processes. This isin accordance with the notion that the IF network doesnot extend into foot processes (Drenckhahn and Franke1988; Pavenstadt et al. 2003). CDK5 in podocyteslikely plays multiple roles because it is probably respon-sible for the differentiation and exit from the cell cycle(Griffin et al. 2004). However, an additional and pivotalrole could be the regulation of IF network extension,preventing it from invading foot processes. In support ofthis view, nestin disassembling has been reported asregulated by the active complex CDK5/p35 (Sahlgrenet al. 2003), which has been spotted mainly gathered atthe podocyte cell processes (Griffin et al. 2004).
Nestin, however, is not capable of forming IFs on itsown, as it must copolymerize with other type III IF
proteins (Sjoberg et al. 1994; Steinert et al. 1999). Inaccordance with the well-known expression of vimentinin podocytes (Holthofer et al 1984; Stamenkovic et al.1986; Moll et al. 1991), our immunocytochemical andbiochemical results confirm that the nestin partner inhuman podocytes is vimentin. The persistence of nestinexpression in adult postmitotic terminally differentiatedcells, in contrast to the more general rule that wouldrestrict nestin to undifferentiated and dividing cells,implies an involvement of nestin in peculiar roles specificfor podocytes. The possibility of a structural strengthen-ing of the podocyte IF network is not convincing becausenestin, far from reinforcing vimentin IFs, has been shownto make them less stable (Steinert et al. 1999). Nestin/vimentin heterodimers, however, have been proposedto be located at the periphery of a core of vimentinhomodimers with the long nestin carboxyl terminalthat sticks out of the filament. The importance of thisarrangement would reside in the ability of the carboxyldomain to interactwithmicrofilaments andmicrotubulescross-linking the three distinct components of thecytoskeleton (Herrmann and Aebi 2000). These inter-connections could result, therefore, in a net advantage forthemechanical stability of podocytes. Another possibilityis that nestin may serve a regulatory function for theassembly state of vimentin because it has been demon-strated inmitotic cells where vimentin IFs, in spite of theirphosphorylated state, disassemble only when nestin iscoexpressed (Chou et al. 2003). In this respect, based alsoon our results, a functional chain linking CDK5, nestin,and vimentin could be envisioned in podocytes: activeCDK5/p35 complex within foot processes would phos-phorylate nestin which, in turn, could regulate vimentindisassembly, preventing the IF network from extend-ing into the pedicels. This view is supported by theobservation that, in puromomycin-induced nephrosis,nestin and vimentin are both overexpressed (Zou et al.2006) and that foot process effacement is characterizedby the presence of IFs within the cytoplasmic sheet that,replacing pedicels, covers the basement membrane(Kubosawa and Kondo 1994). At any rate, the properarchitecture of the IF network seems essential tomaintainfoot process integrity. This is confirmed by experimentsof nestin silencing with siRNA where the effect of nestinknock-down resulted in a marked decrease of podocyteprocesses (Chen et al. 2006).
The present investigation shows that nestin IF is ex-pressed in human podocytes starting from the very be-ginning of their differentiation. The pattern of nestinexpression during human glomerulogenesis sharessome resemblances with that which has been observedin rat and mouse (Chen et al. 2006; Wagner et al. 2006;Zou et al. 2006) but also shows significant differences.In contrast to mouse developing kidney where nestinhas been detected within podocytes only in matureglomeruli (Chen et al. 2006), in humans nestin appears
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Nestin Expression in Human Kidney 419
by guest on May 31, 2013jhc.sagepub.comDownloaded from

to be expressed from the first stage of glomerulogenesis(V stage). Throughout the following stages of develop-ment, nestin staining becomes more intense, localizingpreferentially in the epithelial cells of the visceral layer.This behavior seems closer to what has been reported inrat developing kidney (Zou et al. 2006). On the otherhand, whereas nestin expression in human differenti-ating glomerular endothelial cells is in accordance withmouse glomerulogenesis (Chen et al. 2006), our findingof a further expression in mesangial precursors seems tobe unique to human immature glomeruli.
In Western blotting, nestin is usually reported as mi-grating as a double-immunoreactive band at 220 and240 kDa (Sultana et al. 1998; Sahlgren et al. 2001;Lobo et al. 2004; Wiese et al. 2004). However, ourevaluation gives considerably higher molecular masses,240 and 280 kDa. Our values, in accordance with otherstudies (Messam et al. 2000; Chou et al. 2003; Totiet al. 2005; Zou et al. 2006), point toward nestin heavyposttranslational modifications as possibly servingparticular functions in podocytes and in a few otherparticular cell types. Otherwise, considering that themolecular mass predicted by the amino acid sequenceshould be only 177 kDa and that posttranslationalmodifications should add 100 kDa to the molecule, theexistence of alternative splicings of nestin gene is apossibility that should also be considered. However,more studies are required to clarify this point.
In conclusion, the present investigation has shown,for the first time, that human podocytes express nestinand that this is associated in vivo with CDK5, which isalso located within the foot processes where nestin isnot detectable. Moreover, we have demonstrated that,during glomerulogenesis, nestin is expressed in differ-entiating and mature podocytes, in some endothelialcells and, for the first time, in mesangial precursors.
Acknowledgments
This work was supported by PAR2004 (quota servizi) (toEB) and PRIN2004 (to PT).
Literature Cited
Achtstaetter T, Hatzfeld M, Quinlan RA, Parmelee DC, Franke WW(1986) Separation of cytokeratin polypeptides by gel electropho-retic and chromatographic techniques and their identification byimmunoblotting. Methods Enzymol 134:355–371
AlpersCE, Seifert RA,HudkinsKL, JohnsonRJ, Bowen-PopeDF (1992)Developmental patterns of PDGF B-chain, PDGF-receptor, and a-actin expression in human glomerulogenesis. Kidney Int 42:390–399
Armstrong JF, Pritchard-Jones K, Bickmore WA, Hastie ND, BardJBL (1992) The expression of theWilms’ tumour gene,WT1, in thedeveloping mammalian embryo. Mech Dev 40:85–97
Bachmann S, Kriz W, Kuhn C, Franke WW (1983) Differentiationof cell types in the mammalian kidney by immunofluorescencemicroscopy using antibodies to intermediate filament proteins anddesmoplakins. Histochemistry 77:365–394
Bariety J, Nochy D, Mandet C, Jacquot C, Glotz D, Meyrier A (1998)Podocytes undergo phenotypic changes and express macrophagic-associated markers in idiopathic collapsing glomerulopathy. Kid-ney Int 53:918–925
Beltran PJ, Bixby JL, Masters BA (2003) Expression of PTPRO dur-ing mouse development suggests involvement in axono-genesisand differentiation of NT-3 and NGF-dependent neurons. J CompNeurol 465:384–395
Bertelli E, Bendayan M (2005) Association between endocrine pan-creas and ductal system. More than an epiphenomenon of en-docrine differentiation and development? J Histochem Cytochem53:1071–1086
Bertelli E, Regoli M, Lucattelli M, Bastianini A, Fonzi L (2002) Nestinexpression in rat adrenal gland. Histochem Cell Biol 117:371–377
Bertelli E, Regoli M, Orazioli D, Bendayan M (2001) Associationbetween islets of Langerhans and pancreatic ductal system in adultrat. Where endocrine and exocrine meet together? Diabetologia44:575–584
Bouwens L (1998) Cytokeratins and cell differentiation in the pan-creas. J Pathol 184:234–239
Carapelli A, Regoli M, Nicoletti C, Ermini L, Fonzi L, Bertelli E(2004) Rabbit tonsil-associated M cells express cytokeratin 20 andtake up particulate antigen. J Histochem Cytochem 52:1323–1331
Chen J, Boyle S, ZhaoM, SuM, Takahashi K, Davis L, DeCaesteckerM, et al. (2006) Differential expression of the intermediate fila-ment protein nestin during renal development and its localizationin adult podocytes. J Am Soc Nephrol 17:1283–1291
Chou Y-H, Khuon S, Herrmann H, Goldman RD (2003) Nestinpromotes the phosphorylation-dependent disassembly of vimen-tin intermediate filaments during mitosis. Mol Biol Cell 14:1468–1478
Cicero S, Herrup K (2005) Cyclin-dependent kinase 5 is essentialfor neuronal cell cycle arrest and differentiation. J Neurosci 25:9658–9668
Czarnecki K, Haas CA, Bas Orth C, Deller T, Frotscher M (2005)Postnatal development of synaptopodin expression in the rodenthippocampus. J Comp Neurol 490:133–144
Dahlstrand J, Collins VP, Lendahl U (1992) Expression of the class VIintermediate filament nestin in human central nervous system tu-mors. Cancer Res 52:5334–5341
Delacour A, Nepote V, Trump A, Herrera PL (2004) Nestin expres-sion in pancreatic exocrine cell lineages. Mech Dev 121:3–14
Drenckhahn D, Franke R-P (1988) Ultrastructural organization ofcontractile and cytoskeletal proteins in glomerular podocytes ofchicken, rat and man. Lab Invest 59:673–682
Fina L, Molgaard HV, Robertson D, Bradley NJ, Monaghan P, DeliaD, Sutherland DR, et al. (1990) Expression of the CD34 gene invascular endothelial cells. Blood 75:2417–2426
Franke WW, Schid E, Winter S, Osborn M, Weber K (1979) Wide-spread occurrence of intermediate-sized filaments of the vimentin-type in cultured cells from diverse vertebrates. Exp Cell Res 123:25–46
Frisen J, Johansson CB, Torok C, RislingM, Lendahl U (1995) Rapid,widespread and long-lasting induction of nestin contributes to thegeneration of glial scar tissue after CNS injury. J Cell Biol 131:453–464
Griffin SV, Hiromura K, Pippin J, Petermann AT, Blonski MJ, KrofftR, Takahashi S, et al. (2004) Cyclin-dependent dinase 5 is a regu-lator of podocyte differentiation, proliferation, and morphology.Am J Pathol 165:1175–1185
Gubler M-C (2003) Podocyte differentiation and hereditary protein-uria/nephrotic syndromes. J Am Soc Nephrol 14:S22–S26
Herrmann H, Aebi U (2000) Intermediate filaments and their as-sociates: multi-talented structural elements specifying cytoarchitec-ture and cytodynamics. Curr Opin Cell Biol 12:79–90
Hir ML, Keller C, Eschmann V, Hahnel B, Hosser H, Kriz W (2001)Podocytes bridges between the tuft and Bowman’s capsule: anearly event in experimental crescentic glomerulonephritis. J AmSoc Nephrol 12:2060–2071
Hirvonen H, Sandberg M, Kalimo H, Hukkanen V, Vuorio E, SalmiTT, Alitalo K (1989) The N-myc proto-oncogene and IGF-IIgrowth factor mRNAs are expressed by distinct cells in humanfetal kidney and brain. J Cell Biol 108:1093–1104
Holthofer H, Miettinen A, Letho VP, Lehtonen E, Virtanen I (1984)Expression of vimentin and cytokeratin types of intermediate fila-
TheJournal
ofHistoch
emistry&
Cytoch
emistry
420 Bertelli, Regoli, Fonzi, Occhini, Mannucci, Ermini, Toti
by guest on May 31, 2013jhc.sagepub.comDownloaded from

ment proteins in developing and adult human kidneys. Lab Invest50:552–559
Ito A, Nomura S, Hirota S, Suda J, Suda T, Kitamura Y (1995) En-hanced expression of CD34 messenger RNA by developing endo-thelial cells of mice. Lab Invest 72:532–538
Jepson MA, Clark MA (1998) Studying M cells and their role ininfection. Trends Microbiol 6:359–365
Kachinsky AM, Dominov JA,Miller JB (1995) Intermediate filamentsin cardiac myogenesis: nestin in the developing mouse heart. JHistochem Cytochem 43:843–847
Kerajaschki D (2001) Caught flat-footed: podocyte damage and the mo-lecular bases of focal glomerulosclerosis. J Clin Invest 108:1583–1587
Klein T, Ling Z, Heimberg H, Madsen OD, Heller RS, Serup P (2003)Nestin is expressed in vascular endothelial cells in the adult humanpancreas. J Histochem Cytochem 51:697–706
Kobayashi N, Gao S, Chen J, Saito K, Miyawaki K, Li C, Pan L, et al.(2004) Process formation of the renal glomerular podocyte: is therecommon molecular machinery for processes of podocytes andneurons? Anat Sci Int 79:1–10
Kondo D, Yamamoto T, Yaoita E, Danielson PE, Kobayashi H,Ohshiro K, Funaki H, et al. (2000) Localization of olfactomedin-related glycoprotein isoform (BMZ) in the golgi apparatus of glo-merular podocytes in rat kidneys. J Am Soc Nephrol 11:803–813
KrizW, Hackenthal E, Nobiling R, Sakai T, ElgerM (1994) A role forpodocytes to counteract capillary wall distension. Kidney Int 45:369–376
Kubosawa H, Kondo Y (1994) Modulation of cytoskeletal organi-zation of podocytes during the course of aminonucleoside nephro-sis in rats. Pathol Int 44:578–586
Lardon J, Rooman I, Bouwens L (2002) Nestin expression in pan-creatic stellate cells and angiogenic endothelial cells. HistochemCell Biol 117:535–540
Lazaro JB, Kitzmann M, Poul MA, Vandromme M, Lamb NJ,Fernandez A (1997) Cyclin dependent kinase 5, cdk5, is a positiveregulator ofmyogenesis inmouse C2 cells. J Cell Sci 110:1251–1260
Lendahl U, Zimmerman LB, McKay RD (1990) CNS stem cells ex-press a new class of intermediate filament protein. Cell 60:585–595
Lobo MV, Arenas MI, Alonso FJ, Gomez G, Bazan E, Paino CL,Fernandez E, et al. (2004) Nestin, a neuroectodermal stem cellmarker molecule, is expressed in Leydig cells of the human testisand in some specific cell types from human testicular tumours. CellTissue Res 316:369–376
MessamCA, Hou J,Major EO (2000) Coexpression of nestin in neuraland glial cells in the developing human CNS defined by a human-specific anti-nestin antibody. Exp Neurol 161:585–596
Moll R, Hage C, Thoenes W (1991) Expression of intermediate fil-ament proteins in fetal and adult human kidney: modulations ofintermediate filament patterns during development and in dam-aged tissue. Lab Invest 65:74–86
Mundel P, Heid HW,Mundel TM, KrugerM, Reiser J, KrizW (1997)Synaptopodin: an actin-associated protein in telencephalic den-drites and renal podocytes. J Cell Biol 139:193–204
NagataM, Kakayama K, Terada Y, Hoshi S, Watanabe T (1998) Cellcycle regulation and differentiation in the human podocyte line-age. Am J Pathol 153:1511–1520
NagataM, Yamaguchi Y, Ito K (1993) Loss of mitotic activity and theexpression of vimentin in glomerular epithelial cells of developinghuman kidneys. Anat Embryol (Berl) 187:175–179
Nakahama H, Obata K, Matsuyama T, Sugita M, Horio M, Oka K,Moriyama T (1999) Effect of a novel immunosuppressant, FK506,on autoimmune glomerulonephritis in Brown Norway rats.Nephron 81:215–220
Naruse K, FujiedaM,Miyazaki E, Hayashi Y, ToiM, Fukui T, KurodaN, et al. (2000) An immunohistochemical study of developingglomeruli in human fetal kidneys. Kidney Int 57:1836–1846
Pavenstadt H, Kriz W, Kretzler M (2003) Cell biology of the glo-merular podocyte. Physiol Rev 83:253–307
Putaala H, Soininen R, Kilpelainen P, Wartiovaara J, Tryggvason K(2001) The murine nephrin gene is specifically expressed in kidney,brain and pancreas: inactivation of the gene leads to massiveproteinuria and neonatal death. Hum Mol Genet 10:1–8
Rastaldi MP, Armelloni S, Berra S, Calvaresi N, Corbelli A, GiardinoLA, Li M, et al. (2006) Glomerular podocytes contain neuron-likefunctional synaptic vesicles. FASEB J 20:976–978
Rastaldi MP, Armelloni S, Berra S, Li M, Pesaresi M, Poczewski H,Langer B, et al. (2003) Glomerular podocytes possess the synap-tic vesicle molecule Rab3A and its specific effector Rabphilin-3a.Am J Pathol 163:889–899
Ruggenenti P, Schieppati A, Remuzzi G (2001) Progression, remission,regression of chronic renal diseases. Lancet 357:1601–1608
Sahlgren CM, Mikhailov A, Hellman J, Chou Y-H, Lendahl U,Goldman RD, Eriksson JE (2001) Mitotic reorganization of theintermediate filament protein nestin involves phosphorylation bycdc2 kinase. J Biol Chem 276:16456–16463
Sahlgren CM,Mikhailov A, Vaittinen S, Pallari H-M, Kalimo H, PantHC, Eriksson JE (2003) Cdk5 regulates the organization of nestinand its association with p35. Mol Cell Biol 23:5090–5106
Sejersen T, Lendahl U (1993) Transient expression of the intermediatefilament nestin during skeletal muscle development. J Cell Sci106:1292–1300
Sharma MR, Tuszynski GP, Sharma MC (2004) Angiostatin-inducedinhibition of endothelial cell proliferation/apoptosis is associatedwith the down-regulation of cell cycle regulatory protein cdk5. JCell Biochem 91:398–409
Sjoberg G, Jiang WQ, Ringertz NR, Lendahl U, Sejersen T (1994)Colocalization of nestin and vimentin/desmin in skeletal musclecells demonstrated by three-dimensional fluorescence digital im-aging microscopy. Exp Cell Res 214:447–458
Stamenkovic I, Skalli O, Gabbiani G (1986) Distribution of inter-mediate filament proteins in normal and diseased human kidney.Am J Pathol 125:465–475
Steinert PM, Chou Y-H, Prahlad V, Parry DAD, Marekov LN, WuKC, Jang S-I, et al. (1999) A high molecular weight intermediatefilament-associated protein in BHK-21 cells is nestin, a type VIintermediate filament protein. J Biol Chem 274:9881–9890
Street CN, Lakey JR, Seeberger K, Helms L, Rajotte RV, Shapiro AM,Korbutt GS (2004) Heterogenous expression of nestin in humanpancreatic tissue precludes its use as an islet precursor marker. JEndocrinol 180:213–225
Sultana S, Zhou R, Sadagopan MS, Skalli O (1998) Effects ofgrowth factors and basement membrane proteins on the pheno-type of U-373 MG glioblastoma cells as determined by the ex-pression of intermediate filament proteins. Am J Pathol 153:1157–1168
Thomas SK, Messam CA, Spengler BA, Biedler JL, Ross RA (2004)Nestin is a potential mediator of malignancy in human neuroblas-toma cells. J Biol Chem 279:27994–27999
Toti P, Regoli M, Nesi G, Occhini R, Bartolommei S, Fonzi L, BertelliE (2005) Nestin expression in normal adrenal gland and adreno-cortical tumors. Histol Histopathol 20:1115–1120
Vaittinen S, Lukka R, Sahlgren CM, Hurme T, Rantanen J, LendahlU, Eriksson JE, et al. (2001) The expression of the intermediatefilament nestin as related to vimentin and desmin in regeneratingskeletal muscle. J Neuropathol Exp Neurol 60:588–597
Virtanen I, Letho V-P, Lethonen E, Varto T, Stenman S, Kurki P,Wager O, et al. (1981) Expression of intermediate filaments incultured cells. J Cell Sci 50:45–63
Wagner N, Wagner KD, Scholz H, Kirschner KM, Schedl A (2006)The intermediate filament protein nestin is expressed in the de-veloping kidney and heart and might be regulated by the Wilms’tumor suppressor Wt1. Am J Physiol Regul Integr Comp Physiol291:R779–787
Wiese C, Rolletschek A, Kania G, Blyszczuk P, Tarasov KV, TarasovaY, Wersto RP, et al. (2004) Nestin expression—a property ofmulti-lineage progenitor cells? Cell Mol Life Sci 61:2510–2522
Yaoita E, Franke WW, Yamamoto T, Kawasaki K, Kihara I (1999)Identification of renal podocytes in multiple species: higher ver-tebrates are vimentin positive/lower vertebrates are desmin posi-tive. Histochem Cell Biol 111:107–115
Zou J, Yaoita E, Watanabe Y, Yoshida Y, Nameta M, Li H, Qu Z,et al. (2006) Upregulation of nestin, vimentin, desmin in ratpodocytes in response to injury. Virchows Arch 448:485–492
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Nestin Expression in Human Kidney 421
by guest on May 31, 2013jhc.sagepub.comDownloaded from