
N100 and P300 amplitude to Go and No–Go variants of the auditory oddball in siblings discordant...
13
N100 and P300 amplitude to Go and No–Go variants of the auditory oddball in siblings discordant for schizophrenia Alex Sumich a, ⁎ , Veena Kumari b,c , Phillipa Dodd b , Ulrich Ettinger d , Catherine Hughes b , Elizabeth Zachariah b , Tonmoy Sharma e a King's College London, Institute of Psychiatry, BIAU, Centre for Neuroimaging Sciences, London SE5 8AF, UK b King's College London, Institute of Psychiatry, Psychological Medicine, London SE5 8AF, UK c King's College London, Institute of Psychiatry, Psychology, London SE5 8AF, UK d King's College London, Institute of Psychiatry, BIAU, Neuroimaging Research Group, Centre for Neuroimaging Sciences, London SE5 8AF, UK e The Cognition Group, Newark, Delaware, 19702, USA Received 21 June 2006; received in revised form 6 September 2007; accepted 16 September 2007 Available online 19 November 2007 Abstract Background: P300 amplitude reduction is reliably seen in schizophrenia. Inconsistent reports of isolated frontal and/or parietal deficits in unaffected family members may be clarified using a task that places greater load on frontal function. Method: Go and No–Go versions of the auditory oddball task were performed by eighteen schizophrenia patients, age-matched unaffected siblings and healthy controls matched closely to unaffected siblings on age, sex, education, socioeconomic-status, handedness and ethnicity. Groups were compared on P300 and N100 amplitude and latency. Spearman correlations were used to test the relationship between ERP amplitudes and neuropsychological measures of executive function and memory. The relationship between schizotypy – as measured using the structured interview – and ERPs was explored in a combined group of siblings and controls. Results: Independent of task, patients had lower P300 than controls and reduced parietal amplitude compared to siblings. Siblings had enhanced frontocentral N100 compared to controls. No–Go P300 amplitude and N100 latency was associated with executive function measures. There were significant intraclass correlations between patients and siblings for No–Go P300 amplitude, particularly at the central midline electrode. Frontocentral N100 and P300 amplitude were positively correlated with anxiety-related aspects of schizotypy. Conclusion: Enhanced N100 is present in unaffected siblings. Parietal P300 is intact in unaffected siblings, but reduced in patients. The No–Go-oddball is more sensitive than the Go-oddball to executive function deficits in patients and as an index of heritability. © 2007 Published by Elsevier B.V. Keywords: Schizophrenia; Siblings; Event-related potentials; N100; P300; Anxiety; Executive function 1. Introduction Patients with schizophrenia demonstrate reduction in auditory P300 amplitude, an event-related-potential (ERP) measure of attention, memory and contextual Available online at www.sciencedirect.com Schizophrenia Research 98 (2008) 265 – 277 www.elsevier.com/locate/schres ⁎ Corresponding author. BIAU, Centre for Neuroimaging Sciences Institute of Psychiatry, London SE5 8AF, UK. Tel.: +44 207 919 3066; fax: +44 207 9192116. E-mail address: [email protected] (A. Sumich). 0920-9964/$ - see front matter © 2007 Published by Elsevier B.V. doi:10.1016/j.schres.2007.09.018
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of N100 and P300 amplitude to Go and No–Go variants of the auditory oddball in siblings discordant...

Available online at www.sciencedirect.com
98 (2008) 265–277www.elsevier.com/locate/schres
Schizophrenia Research
N100 and P300 amplitude to Go and No–Go variants of the auditoryoddball in siblings discordant for schizophrenia
Alex Sumich a,⁎, Veena Kumari b,c, Phillipa Dodd b, Ulrich Ettinger d, Catherine Hughes b,Elizabeth Zachariah b , Tonmoy Sharma e
a King's College London, Institute of Psychiatry, BIAU, Centre for Neuroimaging Sciences, London SE5 8AF, UKb King's College London, Institute of Psychiatry, Psychological Medicine, London SE5 8AF, UK
c King's College London, Institute of Psychiatry, Psychology, London SE5 8AF, UKd King's College London, Institute of Psychiatry, BIAU, Neuroimaging Research Group, Centre for Neuroimaging Sciences, London SE5 8AF, UK
e The Cognition Group, Newark, Delaware, 19702, USA
Received 21 June 2006; received in revised form 6 September 2007; accepted 16 September 2007Available online 19 November 2007
Abstract
Background: P300 amplitude reduction is reliably seen in schizophrenia. Inconsistent reports of isolated frontal and/or parietaldeficits in unaffected family members may be clarified using a task that places greater load on frontal function.Method: Go and No–Go versions of the auditory oddball task were performed by eighteen schizophrenia patients, age-matchedunaffected siblings and healthy controls matched closely to unaffected siblings on age, sex, education, socioeconomic-status,handedness and ethnicity. Groups were compared on P300 and N100 amplitude and latency. Spearman correlations were used totest the relationship between ERP amplitudes and neuropsychological measures of executive function and memory. Therelationship between schizotypy – as measured using the structured interview – and ERPs was explored in a combined group ofsiblings and controls.Results: Independent of task, patients had lower P300 than controls and reduced parietal amplitude compared to siblings. Siblingshad enhanced frontocentral N100 compared to controls. No–Go P300 amplitude and N100 latency was associated with executivefunction measures. There were significant intraclass correlations between patients and siblings for No–Go P300 amplitude,particularly at the central midline electrode. Frontocentral N100 and P300 amplitude were positively correlated with anxiety-relatedaspects of schizotypy.Conclusion: Enhanced N100 is present in unaffected siblings. Parietal P300 is intact in unaffected siblings, but reduced in patients.The No–Go-oddball is more sensitive than the Go-oddball to executive function deficits in patients and as an index of heritability.© 2007 Published by Elsevier B.V.
Keywords: Schizophrenia; Siblings; Event-related potentials; N100; P300; Anxiety; Executive function
⁎ Corresponding author. BIAU, Centre for Neuroimaging SciencesInstitute of Psychiatry, London SE5 8AF, UK. Tel.: +44 207 919 3066;fax: +44 207 9192116.
E-mail address: [email protected] (A. Sumich).
0920-9964/$ - see front matter © 2007 Published by Elsevier B.V.doi:10.1016/j.schres.2007.09.018
1. Introduction
Patients with schizophrenia demonstrate reduction inauditory P300 amplitude, an event-related-potential(ERP) measure of attention, memory and contextual

Table 1Demographic and clinical variables
Patients Siblings Controls Total
Mean (SD) Mean (SD) Mean (SD) Mean (SD)
Age 30.67(6.41)
28.61(6.59)
28.39(6.48)
29.22(6.45)
Years ofeducation
12.72(1.78)
12.72(2.05)
13.33(2.95)
12.93(2.29)
% female 44.4 72.2 72.2 63.0% smoker 66.7 50 66.7 61.1% right handed 77.8 94.4 94.4 88.9% Caucasian 61.1 61.1 61.1 61.1% Afro-Caribbean
33.3 33.3 33.3 33.3
% other 5.6 5.6 5.6 5.6Antipsychoticuse
Atypical n=14 na na naTypical n=3 na na naUnmedicated n=1 na na naIllnessduration
7.7 (5.65) na na na
266 A. Sumich et al. / Schizophrenia Research 98 (2008) 265–277
updating (Coull, 1998; Gruzelier et al., 1999; Bharathet al., 2000; Gruzelier, 2003; Jeon and Polich, 2003;Bramon et al., 2005). Such deficits have been associatedwith impairments in executive function and verballearning (Nieman et al., 2002; Kim et al., 2003), thoughnot in all cases (Kim et al., 2003; Dichter et al., 2006).Findings of reduced P300 amplitude in unaffectedsiblings are inconsistent (Bharath et al., 2000). Someauthors report no difference (Karoumi et al., 2000;Winterer et al., 2001; Bramon et al., 2005), whilst othersfind a decrease (Turetsky et al., 2000) or increase infrontal P300 amplitude (Winterer et al., 2003). Ametaanalysis (Bramon et al., 2005) suggested centraland posterior sites as most suitable for obtaining P300endophenotypes, but did not account for age differencespresent in many family studies (e.g. Frangou et al., 1997).
Reduction in the earlier N100 component is also foundin schizophrenia patients, reflecting initial sensory proces-sing and/or early selective attention deficits (Frangou et al.,1997; Coull, 1998; Gallinat et al., 2002; Sumich et al.,2006). However, no difference was seen betweenunaffected relatives and healthy comparisons (Frangou etal., 1997). In contrast, enhanced N100 in a sensory gatingtaskwas seen in unaffected relatives andwas interpreted asreflecting a compensatory mechanism (Waldo et al.,1988). Enhanced N100 is also seen in anxiety disordersand in response to ketamine administration which isthought to model schizophrenia (Oranje et al., 2000;Alcaini et al., 1994). Given previous poor control forconfounding variables such as age and personalitydisorder, the presence of N100 and P300 abnormalitiesin “truly” unaffected relatives remains unclear.
A task that places greater load on frontal functionmay be more sensitive to P300 abnormalities in siblings.Enhanced anterior–central P300 amplitude is seen torare stimuli in a No–Go variant of the oddball taskduring which participants respond to frequent ratherthan rare stimuli (Salisbury et al., 2004). The effect onN100 of the No–Go oddball task is unclear.
The current study investigated N100 and P300amplitudes in probands, unaffected siblings and closelymatched healthy controls in Go and No–Go oddballtasks. Higher P300 amplitudes were expected at frontalsites in the No–Go, compared to the Go task. Comparedto controls, patients were expected to demonstrate areduction in P300 and N100. Siblings were expected todemonstrate a specific frontal P300 reduction. However,whether N100 reduction or enhancement would bepresent in siblings was unclear. The relationshipbetween ERP amplitudes and neuropsychologicalmeasures of executive function and memory recallwere also investigated. It was expected that ERP
responses to the No–Go Oddball would be more closelyassociated with frontal executive processes.
2. Method and materials
2.1. Subject recruitment
Study procedures were approved by the Institute ofPsychiatry and South London and Maudsley Researchethics committee. Following a detailed description ofthe study procedures, all participants provided writteninformed consent to take part. Details on recruitment arepublished elsewhere along with clinical, neuropsycho-logical, eye-movement and sensory gating data on thissample (Hughes et al., 2005; Ettinger et al., 2004).Table 1 shows clinical and demographic data. Briefly,siblings were recruited via probands from in- or out-patient services within and around South London. Of the25 sibling pairs that participated in our previous studies(Hughes et al., 2005; Ettinger et al., 2004), 18 pairsconsented to undergo event-related potential assess-ments. No more than two siblings (1 affected, 1unaffected) came from the same family. The StructuredClinical interview for DSM-IV Axis 1 disorders (SCID1; First et al., 1996a) was used to confirm diagnosis ofschizophrenia in patients. Controls were recruited viaadvertisement placed in a South London newspaper andwere matched to unaffected siblings on age (within5 yrs), gender, ethnicity, years of education (within4 yrs) and handedness (Oldfield, 1971). An additional 4unaffected siblings and controls participated, but are not

267A. Sumich et al. / Schizophrenia Research 98 (2008) 265–277
included in the current study as data from the affectedsibling were not available.
Participants with a history of illicit substance abuse/dependence, mental retardation, neurological disorder, ormedical condition that may affect brain structure orfunction were excluded. Siblings and controls withdiagnosis of axis 1 disorders or personality disorder asassessed by StructuredClinical interview forDSM-IVAxis2 disorders (SCID 2; First et al., 1996b) were excluded.Based the family interview for genetic studies (Gershonand Guroff, 1984), control subjects with a history ofpsychosis in first-degree relatives were excluded.
2.2. Schizotypy assessment
Schizotypal symptoms were assessed in 17 siblingand control pairs using the Structured Interview forSchizotypy (SIS) (Kendler et al., 1989; also see Kumariet al., 2005).
2.3. Neuropsychological assessment
Neuropsychology data from this sample have previ-ously been reported (Hughes et al., 2005). The currentstudy involved measures of memory and executivefunction. Immediate and delayed recall was assessedusing the visual subtest of the Wechsler Memory Scale—Revised (WMS-R) (Wechsler, 1987) and the Rey Verballearning test (Rey, 1964). Executive functions wereassessed using the Wisconsin Card Sorting (Heaton,1993),Verbal fluency (Benton et al., 1983), StroopColourWord (Stroop, 1935), and Trail Making B (Reitan andWolfson, 1985) tests.
2.4. Evoked potentials
2.4.1. TasksSubjects sat alert and still with their eyes closed and
neck well supported by the back of the chair whilstlistening to the stimuli which were presented binaurallythrough insert ear plugs (intensity=65 dB, 10 ms rise/fall; duration=50 ms). Forty rare tones (1000 Hz) wererandomly distributed amongst 160 frequent tones(2000 Hz). Subjects were asked to respond to stimuliby button press with the thumb of the dominant hand asquickly and accurately as possible. The Go conditionrequired response to low tones. The No–Go conditionrequired response to high tones. Before starting, thesubject's ability to discriminate easily between high andlow tones was confirmed. The order of Go and No–Gotasks was quasirandomly assigned. Ten patients, 9siblings and 12 controls completed the Go task first.
2.4.2. RecordingUsing a Nihon Kohden model EEG-4321F/G ampli-
fier and NeuroScan Acquire 4.0 software, electroenceph-alographic activity (EEG) was recorded following theinternational 10–20 system from 15 electrodes referencedto linked earlobes with forehead ground. Only ERPs torare stimuli at F3, F4, Fz, C3, C4, Cz, P3, P4 and Pz areincluded in the current study. EOGv was recorded fromabove and below the left eye and EOGh from bipolarelectrodes at the outer canthi of each eye. Impedance wasmaintained below 5 kΩ. Electrical activitywas sampled at1000 Hz and filtered using a bandpass of 0.1–70 (Hz) tobe compatible with Karoumi et al. (2000).
2.4.3. ERP analysisAnalysis was performed using NeuroScan Edit 4.1.
Ocular artefact reduction (Semlitsch et al., 1986) wasperformed using EOGv. Following a linear detrend onEOGh for the entire sweep, epochs with EOGh activityoutside −60 to +60 μV were rejected. Epochs thatcontained excessive movement artefact between 100 msprestimulus and 800 ms poststimulus were also rejected.Averaged ERPs included at least 30 rare sweeps.
Components were defined as: N100, the most negativepeak occurring between 70 and 150 ms poststimulus; andP300, the most positive peak between 250 and 500 ms.Amplitudes were measured from baseline to peak.
2.4.4. StatisticsStatistical package for social sciences (SPSS) 15.0
was used for all analysis. Unless otherwise stated, alphalevel of significance was maintained at pb .05.
Separate analyses were performed for N100 andP300. A mixed model repeated measures analysis ofvariance was used to investigate differences in amplitudewith diagnosis (patient, sibling, control) as between-groups variable, and task (go, no–go), site (frontal,central, parietal) and hemisphere (left, right, midline) aswithin-groups variables (compound symmetry as-sumed). A random cluster variable (variance compo-nents) based on the patient–sibling–control match wasused in the model. Significant effects were followed bypost-hoc tests using a Bonferroni correction.
To investigate heritability, intraclass correlationcoefficients (Cronbach's alpha) on cluster pairs (pa-tient–sibling, patient–control and siblings–control)were used to test the effects of relatedness on midlineN100 and P300 amplitudes and latencies during Go andNo–Go tasks. Each ERP variable was assessed in aseparate analysis.
Univariate analyses of variance (ANOVA) were usedto test for differences between siblings and controls in
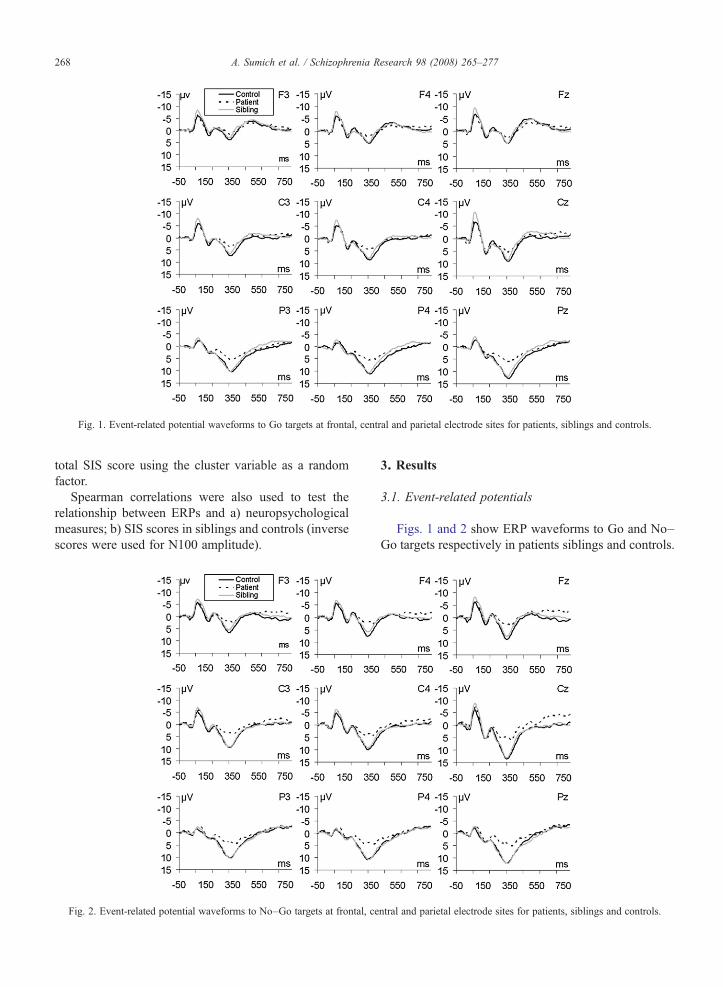
Fig. 1. Event-related potential waveforms to Go targets at frontal, central and parietal electrode sites for patients, siblings and controls.
268 A. Sumich et al. / Schizophrenia Research 98 (2008) 265–277
total SIS score using the cluster variable as a randomfactor.
Spearman correlations were also used to test therelationship between ERPs and a) neuropsychologicalmeasures; b) SIS scores in siblings and controls (inversescores were used for N100 amplitude).
Fig. 2. Event-related potential waveforms to No–Go targets at frontal, ce
3. Results
3.1. Event-related potentials
Figs. 1 and 2 show ERP waveforms to Go and No–Go targets respectively in patients siblings and controls.
ntral and parietal electrode sites for patients, siblings and controls.
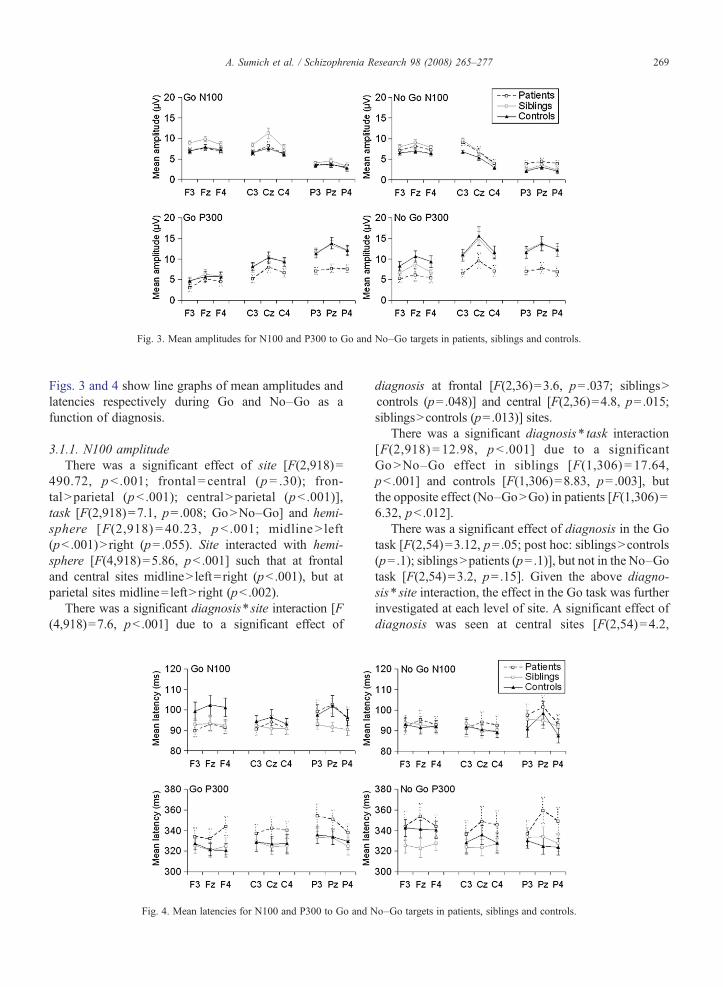
Fig. 3. Mean amplitudes for N100 and P300 to Go and No–Go targets in patients, siblings and controls.
269A. Sumich et al. / Schizophrenia Research 98 (2008) 265–277
Figs. 3 and 4 show line graphs of mean amplitudes andlatencies respectively during Go and No–Go as afunction of diagnosis.
3.1.1. N100 amplitudeThere was a significant effect of site [F(2,918)=
490.72, pb .001; frontal = central (p= .30); fron-talNparietal (pb .001); centralNparietal (pb .001)],task [F(2,918)=7.1, p=.008; GoNNo–Go] and hemi-sphere [F(2,918) = 40.23, p b .001; midlineN left(pb .001)N right (p=.055). Site interacted with hemi-sphere [F(4,918)=5.86, pb .001] such that at frontaland central sites midlineN left= right (pb .001), but atparietal sites midline= leftN right (pb .002).
There was a significant diagnosis⁎site interaction [F(4,918)=7.6, pb .001] due to a significant effect of
Fig. 4. Mean latencies for N100 and P300 to Go and N
diagnosis at frontal [F(2,36)=3.6, p=.037; siblingsNcontrols (p=.048)] and central [F(2,36)=4.8, p=.015;siblingsNcontrols (p=.013)] sites.
There was a significant diagnosis⁎ task interaction[F(2,918) = 12.98, pb .001] due to a significantGoNNo–Go effect in siblings [F(1,306) = 17.64,pb .001] and controls [F(1,306)=8.83, p= .003], butthe opposite effect (No–GoNGo) in patients [F(1,306)=6.32, pb .012].
There was a significant effect of diagnosis in the Gotask [F(2,54)=3.12, p=.05; post hoc: siblingsNcontrols(p=.1); siblingsNpatients (p=.1)], but not in the No–Gotask [F(2,54)=3.2, p=.15]. Given the above diagno-sis⁎ site interaction, the effect in the Go task was furtherinvestigated at each level of site. A significant effect ofdiagnosis was seen at central sites [F(2,54)=4.2,
o–Go targets in patients, siblings and controls.

270 A. Sumich et al. / Schizophrenia Research 98 (2008) 265–277
p=.02] due to higher amplitudes in siblings compared tocontrols (p=.037) and patients (p=.06). At frontal sitesa trend for diagnosis was seen [F(2,54)=3.0, p=.058]which was mainly due to differences between patientsand siblings (p=.09). The effect at parietal sites was notsignificant.
3.1.1.1. N100 latency. There were significant effectsof hemisphere [F(2,918)=66.84, pb .001] and site [F(2,918)=22.3, p=.007], as well as hemisphere⁎ site. [F(4,918)=10.0, pb .001], task⁎ site [F(4,918)=12.7,pb .001] and hemisphere⁎ task⁎ site [F(4,918)=11.2,pb .001] interactions. Post-hoc tests for hemisphereshowed longer latencies at midline than left or right sites(both pb .001). Post-hoc tests for site showed longerlatencies at parietal compared to frontal and central sites(both pb .001) with no difference between frontal andcentral sites. This effect was significant at midline sites,but was not present in the left hemisphere. In the righthemisphere, latency was later at parietal compared tocentral sites, but not frontal sites.
The task⁎ site and hemisphere⁎ task⁎ site interac-tions were such that effects of taskwere seen at central[F(1,270)=13.8, pb .001; GobNo–Go] and parietal[F(1,270)=10.04, p= .002; No–GobGo] sites only.The former effect was significant for midline sites only[F(1,54) =45.37, pb .001], whilst the parietal effectwas seen at midline [F(1,54) =13.78, pb .001] and lefthemisphere [F(1,54)=4.09, p=.048] sites. The Go taskshowed a frontal=centralbparietal pattern (pb .001),whilst in the No–Go task frontal latencies were earlierthan central (p=.056) and parietal (p=.015).
The hemisphere⁎ task⁎ site interaction was alsodue to a significant task⁎ site interaction at midlinesites [F(1,270)=23.65, pb .001]. Significant effects ofsite were seen for the right hemisphere during the No–Go task [F(2,108)=5.48, p= .005; parietalN frontal( p=.065) parietalNcentral ( p=.005)] and for midlinesites in both tasks [Go: F(2,108)=45.68, pb .001;parietalN frontal ( pb .001), parietalNcentral (pb .001);No–Go: F(2,108) = 8.30, pb .001; centralN frontal( pb .001), parietalN frontal (p= .025)].
Diagnosis interacted with task [F(2,918)=3.07,p=.047] and hemisphere [F(2,918)=2.39, p=.049].The former was due to a significant effect of task incontrols [F(1,306)=7.40, p= .007; GoNNo–Go] thatwas absent in both patients and siblings.
3.1.2. P300 amplitudeThere were significant effects of site [F(2,918)=
190.75, pb .001; post hoc: frontalbcentralbparietal (allpb .001)], task [F(1,918)=71.3, pb .001; No–GoNGo]
and hemisphere [F(1,918)=42.8, pb .001; leftb right(p=.015)bmidline ( pb .001)].
There was a significant task⁎ site interaction [F(2,918)=16.8, pb .001] due to a significant effect oftask (No–GoNGo) at frontal [F(1,270) = 67.28,pb .001] and central [F(1,270)=39.93, p=.002] sitesonly. Furthermore, in the Go task the effect of site [F(2,432) = 264.35, p b .001] was due to fron-talbcentralbparietal (all pb .001) effects, whilst theeffect of site in the No–Go task [F(2,432)=97.89,pb .001] was due to frontalbcentral (pb .001)=parietaleffects.
Hemisphere interacted with task [F(1,918)=2.95,p=.05] such that the effect in the Go task [F(2,432)=31.18, pb .001] was due to leftb rightbmidline effects(all pb .001). During the No–Go task, the effect of he-misphere [F(2,432)=45.57, pb .001] was due to higheramplitudes at midline than left and right sites (bothpb .001), but no difference was seen between left andright hemispheres (p=.74).
Hemisphere also interacted with site [F(1,918)=3.07, p= .016]. This was because the effects at left [F(2,270)=93.89, pb .001] and right [F(2,270)=68.14,pb .001] sites were due to frontalbcentralbparietaleffects (all pb .001), but at midline the effect of site [F(2,270)=48.42, pb .001] was seen as frontalbcentral(pb .001)=parietal.
There was a significant effect of diagnosis [F(2,54)=5.30, pb .001; post hoc: controlsNpatients (p=.014);siblingsNpatients (p=.031)]. Significant interactionswere seen between diagnosis⁎ task [F(2,918)=6.90,p = .001] and diagnosis ⁎ site [F(4,918) = 10.38,pb .001]. The former was due to a weaker effect ofdiagnosis in the Go [F(2,54)=3.39, p=.041] thanNo=Go [F(2,54)=5.1, p=.009] task. Post-hoc testsfor the No-Go task were significant [patientsb siblings(p=.048); patientsbcontrols (p=.012)], but reachedtrend level only in the Go task [patientsb siblings(p= .083); patientsbcontrols (p= .087)]. The meandifference between Go and No–Go conditions wasgreatest in controls (2.5 μV) and least in patients(.74 μV), whilst siblings lay midway (1.7 μV).
The site⁎diagnosis interaction was due to a signif-icant effect of diagnosis at parietal [F(2,54)=8.6,p=.001] and central [F(2,54)=4.55, p=.015] sites.Compared to patients, higher parietal amplitudes wereseen in both siblings (p= .002) and controls (p=.002) toa similar degree. At central sites a stronger effect wasseen in controls vs patients (p=.026) than siblings vspatients (p=.049). Also contributing to the site⁎diag-nosis interaction both siblings and controls hadsignificantly higher amplitudes at parietal than central
271A. Sumich et al. / Schizophrenia Research 98 (2008) 265–277
sites (pb .001). However, patients failed to show thiseffect.
Given the site⁎diagnosis and task⁎diagnosis inter-action and our hypothesis of anterior No–Go deficits insiblings and patients, the effect of diagnosis wasinvestigated at each level of task and site. Significanteffects were seen for the No–Go task at frontal [F(2,54)=8.3, p= .042], central [F(2,54)=4.5, p= .015] andparietal [F(2,54)=6.89, p=.002] sites; whilst in the Gotask significant effects were only seen at parietal sites [F(2,54)=8.3, p= .001]. Post-hoc tests for the No–Go taskshowed higher amplitudes in controls than patients atfrontal (p=.037), central (p=.023) and parietal (p=.003)sites, whilst siblings had higher amplitudes than patientsat parietal (p=.002), but not central (p=.06) or frontal(p=.62) sites.
3.1.2.1. P300 latency. There were significant effectsof task [F(1,918)=4.58, p=.033; GobNo–Go], site [F(2,918)=9.18, pb .001; frontalbcentral (p=.049) fron-talbparietal (pb .001)] and hemisphere [F(2,918)=9.03,pb .001; left= rightNmidline (both pb .001)]. Signifi-cant interactions were seen for task⁎ site [F(2,918)=3.44, p=.032] and site⁎hemisphere [F(4,918)=3.23,p=.012] such that the effect of site was only seen in theGo task [F(2,432)=15.89, pb .001; frontalbcentral(p=.001) frontalbparietal (pb .001)] or at midline sites
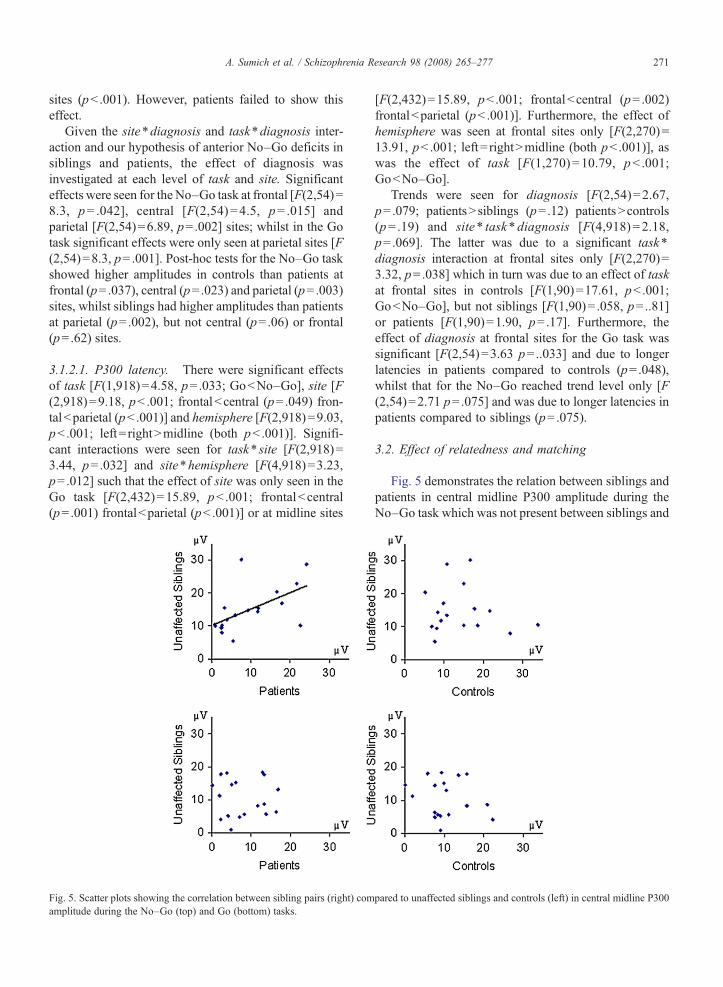
Fig. 5. Scatter plots showing the correlation between sibling pairs (right) comamplitude during the No–Go (top) and Go (bottom) tasks.
[F(2,432)=15.89, pb .001; frontalbcentral (p= .002)frontalbparietal (pb .001)]. Furthermore, the effect ofhemisphere was seen at frontal sites only [F(2,270)=13.91, pb .001; left= rightNmidline (both pb .001)], aswas the effect of task [F(1,270)=10.79, pb .001;GobNo–Go].
Trends were seen for diagnosis [F(2,54)=2.67,p=.079; patientsN siblings (p=.12) patientsNcontrols(p= .19) and site⁎ task⁎diagnosis [F(4,918)=2.18,p=.069]. The latter was due to a significant task⁎
diagnosis interaction at frontal sites only [F(2,270)=3.32, p=.038] which in turn was due to an effect of taskat frontal sites in controls [F(1,90)=17.61, pb .001;GobNo–Go], but not siblings [F(1,90)= .058, p=..81]or patients [F(1,90)=1.90, p=.17]. Furthermore, theeffect of diagnosis at frontal sites for the Go task wassignificant [F(2,54)=3.63 p=..033] and due to longerlatencies in patients compared to controls (p=.048),whilst that for the No–Go reached trend level only [F(2,54)=2.71 p=.075] and was due to longer latencies inpatients compared to siblings (p=.075).
3.2. Effect of relatedness and matching
Fig. 5 demonstrates the relation between siblings andpatients in central midline P300 amplitude during theNo–Go task which was not present between siblings and
pared to unaffected siblings and controls (left) in central midline P300

272 A. Sumich et al. / Schizophrenia Research 98 (2008) 265–277
controls or during the Go P300. Significant relationshipsbetween patient–sibling pairs were seen for No–GoP300 amplitude [central–central r=.57, CIs= .15–.81,p=.006; parietal–frontal r=.5, CIs= .06–.78, p=.014].Weaker relationships between patient–sibling pairswere also seen for Go N100 [frontal–frontal r=.45,CIs=− .008–.75, p= .027]; No–Go N100 [central–frontal r= .35, CIs=− .13–.70, p=.07; parietal–centralr=.42, CIs=− .05–.73, p=.039]; No–Go P300 ampli-tude [frontal–frontal r=.42, CIs=− .04–.74, p=.036;frontal–central r=.44, CIs=− .019–.75, p=.030; cen-tral–frontal r=.46, CIs= .004– .76, p=.024; parietal–central r=.47, CIs= .01–.76, p=.022; parietal–parietalr= .35, CIs=− .12–.70, p= .069] and No–Go P300latency [frontal–parietal r= .40, CIs =− .07–.73,p= .044]. A weak relationship was seen betweencontrol–sibling pairs for No–Go N100 parietal–central[r=.40, CIs=− .07–.72, p= .047].
3.3. ERP relationship to neuropsychology
No–Go frontal N100 amplitude (inverse value) wasnegatively associated with verbal recall, both immediate[right(r=− .29, p=.034), midline(r=− .27, p=.047), left(r=− .24, p=.085)] and delayed [right(r=− .27, p=.05),midline(r=− .26, p=.055), left(r=− .24, p= .077)]. Asimilar effect was seen between right central amplitudeand immediate verbal recall (r=− .28, p=.038), whilstleft central amplitude was associated with good Stroopperformance (r= .27, p=.05). Parietal N100 amplitudewas associated with immediate [left(r=.−34, p=.013);right(r=− .29, p= .034)] and delayed [left(r=− .32,p=.017); right(r=− .25, p=.073)] visual memory.
In the No–Go task, parietal N100 latency wassignificantly associated with verbal fluency (categoriesonly) [midline(r=− .35, p=.01)]; Stroop [midline(r=− .32, p= .02), left(r=− .26, p= .058), right(r=− .27,p=.046)]; WCST perseverative errors [midline(r=.32,p=.02), left(r=− .27, p=.047)]; Trails-B completion time[midline(r=.39, p=.003), left(r=− .35, p=.009), right(r=− .37, p=.007)] and immediate verbal recall [midline(r=− .38, p=.004), left(r=− .27, p=.05)]. Left centralN100 latency was associated with WCST perseverativeerrors (r=.31, p=.02), Trails-B completion time (r=.29,p=.02) and immediate verbal recall (r=− .27, p=.049).Frontal latency was associated with Trails-B completiontime [midline(r=.29, p=.03), right(r=.28, p=.043)];immediate visual memory [right(r=− .30, p=.027)] andverbal learning [left(r=.27, p=.047)]. In the Go task, leftparietal latency was associated with WCST perseverativeerrors (r=.29, p=.033), Trails-B completion time (r=.30,p=.025) and immediate verbal recall (r=− .26, p=.06).
3.3.1. P300Stroop performance was positively correlated with
left central and posterior P300 amplitude in the Go task[central(left r= .27, p= .046; parietal(left: r= .36,p= .007; midline r= .37, p= .006; right r= .34,p=.013)], and at all sites in the No–Go task [frontal(left: r=.34, p=.013; right r=.28, p=.037, midliner= .30, p= .029); central(left r= .33, p= .015; rightr=.30, p=.028; midline r=.34, p=.011) and parietal(left r=.28, p=.040; right r=.30, p=.029; midliner=.30, p=.025)]. Frontal No–Go amplitude was alsoassociated with delayed [left(r=.31, p=.024); right(r=.31, p=.025), midline (r=.29, p=.037)] and imme-diate [left(r= .26, p= .062), right(r= .25, p= .065);midline(r= .25, p=.066)] visual memory.
In the No–Go task, left frontal latency was positivelyassociated with immediate verbal recall (r=.3, p=.02) aswas midline frontal (r=.26, p=.06) and midline central(r=.27, p=.05) latency, whilst right frontal latency wasnegatively associated with verbal learning (r=− .27,p=.05).
3.4. Schizotypy
Controls had significantly higher SIS scores (i.e.lower schizotypy) than siblings [F(1,15)=9.0, p=.009].
Higher schizotypy was positively correlated withamplitude for frontal N100 [Go left (r=− .34, p=.05);midline (r=− .35, p= .042); No–Go left (r=− .37,p=.032); midline (r=− .34, p=.048)], central Go P300[left (r=− .34, p=.049); right (r=− .44; p=.01); midline(r=− .39, p=.022)] and parietal P300 [left (r=− .33,p=.053)].
Further analysis of the relationship between frontocen-tral components and SIS subscores revealed N100 wasmost significantly correlated with childhood isolation[central Go (left r=− .54, p=.001; right r=− .46, p=.007,midline r=− .53, p=.001); central No–Go (midline r=− .55, p=.001)]; sensitivity [frontal Go (left r=.47,p= .006; right r=− .43, p= .011; midline r=− .51,p=.002); central Go (right r=− .44, p=.009; midline r=− .47, p=.006); frontal No–Go (left r=− .43, p=.012);central No–Go (midline r=− .48, p=.004); social anxiety[frontal Go (midline r=− .44, p=.009); central Go (left r=− .47, p=.006; right r=− .48, p=.004; midline r=− .46,p=.007); central No–Go (midline r=− .47, p=.006)] andrestricted emotion [central Go (right r=− .48, p=.004).
Frontocentral P300 was most significantly associatedwith social anxiety [frontal Go (left r=− .51, p=.002;right r=− .47, p=.005; midline r=− .51, p=.002); centralGo (left r=− .58, pb .001; right r=− .62, pb .001; midliner=− .48, p=.004); central No–Go (left r=− .44, p=.009;

273A. Sumich et al. / Schizophrenia Research 98 (2008) 265–277
right r=− .46, p=.007; midline r=− .50, p=.003)] andsensitivity [central Go (right r=− .42, p=.013). Therewere a number of trend effects (.01bpb .05) not currentlyreported.
Analysis by gender found highly significant correla-tions in men for Go N100 at frontal [left(r=− .72,p= .030); right(r=− .93, pb .001); midline(r=− .90,pb .001)]; central [left(r=− .67, p=.05); right(r=− .78,p= .013)] and parietal [left(r=− .72, p= .03)] sites.Similar relationships were seen for No–Go N100 atfrontal [right(r=− .70, p= .036); midline(r=− .85,p=.004)] and central [right(r=− .80, p= .01); midline(r=− .67, p=.05)] sites, as well as for Go P300 at central[left(r=− .80, p=.01); right(r=− .73, p=.025); midline(r=− .90, p=.001)], parietal [left(r=− .68, p=.042);midline(r=− .067, p=.05)] and No–Go P300 at frontal[right(r=− .71, p= .03)] and central[left (r=− .67,p=.05), midline(r=− .73, p=.025)] sites. Significanteffects were not seen in women.
4. Discussion
The current study investigated ERPs in response andresponse-suppression tasks in schizophrenia probands,unaffected siblings and closely matched healthy con-trols. Consistent with previous studies, increasedamplitude, delayed latency and loss of asymmetrywere seen for frontal P300 in the No–Go compared toGo tasks (see Salisbury et al., 2004). Furthermore,higher amplitude and later latency for N100 were seen inthe Go compared to No–Go task. However, the N100amplitude effects were not seen in patients who hadhigher No–Go than Go N100 amplitude. Significantlatency effects (N100 and P300), seen in controls, wereabsent in both patients and unaffected siblings. Siblingshad higher central Go-N100 amplitude than controls andpatients (trend). Patients had lower parietocentral P300than siblings and controls in both tasks and lower frontalNo–Go P300 than controls. Siblings had higher levelsof schizotypy despite being screened for personalitydisorder, consistent with previous studies by our group(Kumari et al., 2005; Ettinger et al., 2004).
Reduced P300 amplitude in patients supportsprevious studies (Frangou et al., 1997; Bharath et al.,2000; Turetsky et al., 2000; Bramon et al., 2005), andwas most prominent at parietal sites. Absence ofsignificant P300 differences between unaffected siblingsand controls is consistent with Karoumi et al. (2000), butcontrasts studies that do not control for age and/orpersonality disorder and were examining families withhigh genetic loading (Frangou et al., 1997; Wintereret al., 2003; Bramon et al., 2005). It is also consistent
with relatively intact neuropsychological functioningand absence of personality disorder in this sample(Hughes et al., 2005).
4.1. No–Go P300 and relatedness
At parietal sites both tasks were equally sensitive todeficits in patients, consistent with suggestions of ageneral cognitive deficit underlying core P300 abnor-malities in schizophrenia, rather than a specific deficit inexecutive function (Schall et al., 1996; Kim et al., 2003).However, the No–Go P300 was more sensitive tofrontal deficits in patients and was most sensitive to therelationship between patients and siblings. Amplitudesin patients at all sites were associated with amplitudes insiblings at frontocentral, but not parietal sites. Thus, agenetic factor may impact on frontocentral executiveP300 mechanisms in both siblings, in addition toposterior mechanisms in probands (Turetsky et al.,2000) — although gender differences between patientsand siblings may have affected these relationships(Turetsky et al., 1998). Impaired frontal function haspreviously been associated with SPEM deficits inschizophrenia (Grawe and Levander, 1995). Thus,consistent with current results we previously reportsignificant within patient–sibling correlations forsmooth pursuit eye movement (SPEM) gain at 10°/s inthis sample (Ettinger et al., 2004).
Our families had a very low genetic loading (only 3families had family history of psychosis; see Hugheset al., 2005) which might explain why we failed toreplicate the finding of significant frontal P300 reduc-tions in unaffected siblings (Turetsky et al., 2000).Nevertheless, siblings lay between patients and controlsfor frontal No–Go P300 amplitude. Thus furtherinvestigation of frontal deficits in unaffected siblingsis clearly warranted and should include study of otheranterior P300 and N100 subcomponents using indepen-dent components analysis. Heritability in frontal GoN100 was seen to a lesser extent and, along with theNo–Go N100 relationship between siblings and con-trols, requires caution until replicated.
4.2. No–Go task as a measure of executive function
Salisbury et al. (2004) suggest the No–Go effect onfrontal P300 is similar to that seen in a silent count taskand may reflect the absence of a negative going waveinvolved in response execution. However, this effectmight also reflect greater involvement of the hippocam-pus in executive (No–Go) and memory (silent count)tasks (Takahashi et al., 2007; Antonova et al., 2004). In

274 A. Sumich et al. / Schizophrenia Research 98 (2008) 265–277
support of this, higher P300 amplitude and longerlatency in the No–Go task was associated with memoryfunctions. Furthermore, higher frontal and right centralNo–Go N100 was associated with poor verbal recall,whilst higher parietal N100 was associated with poorvisual memory. Reduced frontal P300–and enhancedN100–is seen with hippocampal lesions (Knight, 1996;Friedman et al., 2001; Nishitani et al., 1999). Significantrelationships were also seen between the No–Go P300at all sites and Stroop supporting it as an index ofexecutive function. A similar effect was seen for leftcentral and parietal, but not frontal Go-P300 as wasreported in a predominantly male patient group (Niemanet al., 2002). Significant associations between Go-P300amplitude and tests of verbal fluency, visual recall, andverbal learning were unsupported (Nieman et al., 2002;Kim et al., 2003).
Earlier parietal No–Go N100 latency was associatedwith better performance on various executive functions(Stroop, WCST, verbal fluency and Trails B), furthersupporting the No–Go task as an index of executivefunction. Higashima et al. (2005) report a similarrelationship between N100 latency and the differencebetween Trails A and B scores, an index of set-shifting.Our results extend this to include other aspects ofexecutive function. Whilst these are traditionally frontalfunctions, neuroimaging studies elucidate posteriormechanisms, which may be particularly involved inschizophrenia (Moreno-Iniguez et al., 2007; van Veenand Carter, 2005; Vitali et al., 2005).
4.3. Enhanced N100 in unaffected siblings
Enhanced N100 amplitude in siblings is consistentwith a previous study of sensory gating (Waldo et al.,1988) and anterior cingulate hyperactivation in unaffect-ed relatives (Thermenos et al., 2004) given N100generators in this region (Gallinat et al., 2002). However,Frangou et al. (1997) found no difference in N100amplitude between relatives of schizophrenia probandsand significantly younger healthy comparisons. En-hanced N100 is also seen with ketamine or enfluraneadministration, medial temporal lobectomy and/or ob-structive sleep apnea, to which prefrontal and medialtemporal lobes are particularly susceptible (Wong et al.,2006; Bartlett et al., 2004; Oranje et al., 2000; Nishitaniet al., 1999). Given prefrontal and medial temporalregions are implicated in prepulse inhibition, results areconsistent with sensory-motor gating deficits previouslyreported in this sample. Furthermore, high prepulseinhibition was associated with low schizotypy (Kumariet al., 2005).
We hypothesise an anterior cingulate regulatedmechanism indexed by N100 that compensates formedial temporal memory deficits in unaffected siblings.Whilst clearly further studies are needed to confirm thishypothesis, it is somewhat supported by the negativerelationship between left central N100 amplitude andverbal recall, whilst right central N100 amplitude waspositively associated with Stroop performance. Whetherhigh N100 amplitude is associated with incidences ofexceptional creativity and/or intelligence in unaffectedrelatives of schizophrenia patients remains unclear(Gruzelier, 2003). Furthermore, it should be noted thatthe normal ERP intelligence relationship is disrupted inschizophrenia and this may also be the case in siblings(Dichter et al., 2006). Thus a number of alternativeinterpretations for enhanced N100 are discussed belowbased on its relationship to schizotypy.
4.4. ERPs and schizotypy
Consistent with studies of anxiety, N100 and P300amplitudes were positively associated with schizotypy(Clark et al., 1996; Alcaini et al., 1994). In panic disorder,high P300 amplitude was interpreted as reflectinghyperactivation of orientation mechanisms (Clark et al.,1996). In contrast reduced P300 is associated with otheraspects of schizotypy such as negative syndrome,paranormal belief and thought disorder (Sumich et al.,revised submission; Squires-Wheeler et al., 1997; McCo-naghy et al., 1993; Ward et al., 1992). Taken togetherresults support a syndrome-specific relationship betweenschizotypy and brain function (Gruzelier et al., 1999).Similarly, the association between N100 and anxiety-related schizotypy features is consistent with highamplitude N1c in anxiety disorders: thought to reflectincreased arousal due to conflict generated by deviant/rarestimuli (Alcaini et al., 1994). In contrast to N1b, adecrease in N1c is seen with maturation (Bender et al.,2006). Therefore, it is possible that the associationbetween N100 and schizotypy is neurodevelopmental innature and akin to N1c enhancement in anxiety disorders.
Enhancement of the N100/P200 complex in schizoty-pal personality disorders was previously reported in thecontext of intensity dependence, an index of centralserotonergic function (Wang et al., 2006; Hegerl et al.,2001). Our stimuli were presented at 65 dBl which is anintermediate level used in intensity dependence tasks.Nevertheless, future studies should investigate whetherpoor 5-HT function underlies the current results. N100may also have been affected by the use of two oddballtasks which would have heightened habituation effects ontarget stimuli, given an overall greater number of targets.

275A. Sumich et al. / Schizophrenia Research 98 (2008) 265–277
Poor habituation in schizotypal individuals (Vernon et al.,2005) could have thus lead to higher averaged N100.Whilst this is speculative and requires future confirmation,it might also explain the absence of significant differencesbetween patients and controls (Frangou et al., 1997;Sumich et al., 2006). A number of other cognitiveprocesses affecting N100 amplitude, such as consciouseffort, short termmemory, subjective set size (Mulert et al.,2005; Golob and Star 2004), should also be investigated inunaffected siblings and in relationship to schizotypy.
4.5. Limitations
Although we controlled for many potentially con-founding variables, there were clear differences betweenpatients and unaffected siblings in the distribution ofmen and women. Gender-differences were apparent inthe association between ERPs and schizotypy consistentwith previous reports of enhanced P300 in maleunaffected siblings (Winterer et al., 2001, 2003).However, given the current sample of 9 men, resultsshould be treated with caution until replicated. Smallsample size prevented investigation of the effect ofgender on group comparisons.
Current results may have also been influenced by thelarge proportion of probands taking atypical medication.However, the effect of atypical medications on P300 orN100 amplitude is inconclusive (Molina et al., 2004;Nieman et al., 2002). Future studies will require largersamples of closelymatched groups andmore sophisticatedanalysis techniques including source localisationmethods.
4.6. Summary
The current results confirm reduction in P300amplitude in patients, but not age-matched unaffectedsiblings screened for personality disorders. Unaffectedsiblings had higher N100 amplitude than controls whichmay represent a compensatory mechanism for poormedial temporal function and is associated with certainschizotypy features. No–Go auditory P300 amplitude isa more accurate index of executive function deficits inpatients and heritability than Go-P300.
Role of the funding sourceWe would like to thank Psychmed Ltd for financial support. VK is
supported by the Wellcome Trust -067427/z/02/z. UE is funded by aNIHR (National Institute for Health Research) Personal Award. Theviews expressed in this publication are those of the authors and notnecessarily those of the NHS, NIHR or Department of Health.
Conflict of interest
The authors declare no known conflict of interest.AcknowledgementsWe would like to thank the participants in this study for their time
and Daniel Stahl for his statistical advice.
References
Antonova, E., Sharma, T.,Morris, R., Kumari, V., 2004. The relationshipbetween brain structure and neurocognition in schizophrenia: aselective review. Schizophr. Res. 70 (2–3), 117–145.
Alcaini, M., Giard, M.H., Thevenet, M., Pernier, J., 1994. Twoseparate frontal components in the N1 wave of the human auditoryevoked response. Psychophysiology 31 (6), 611–615.
Bartlett, D.J., Rae, C., Thompson, C.H., Byth, K., Joffe, D.A., Enright,T., 2004. Grunstein RR Hippocampal area metabolites relate toseverity and cognitive function in obstructive sleep apnea. SleepMed. Rev. 5 (6), 593–596.
Bender, S., Oelkers-Ax, R., Resch, F., Weisbrod, M., 2006. Frontallobe involvement in the processing of meaningful auditory stimulidevelops during childhood and adolescence. NeuroImage 33 (2),759–773 (Electronic publication 2006 Aug 23).
Benton, A.L., Hamsher, K., Verney, N.R., Spreen, O., 1983.Contributions to Neuropsychological Assessment. Oxford Uni-versity Press, New York.
Bharath, S., Gangadhar, B.N., Janakiramaiah, N., 2000. P300 in familystudies of schizophrenia: review and critique. Int. J. Psychophy-siol. 38, 43–54.
Bramon, E., McDonald, C., Croft, R.J., Landau, S., Filbey, F.,Gruzelier, J.H., Sham, P.C., Frangou, S., Murray, R.M., 2005. Isthe P300 wave an endophenotype for schizophrenia? A meta-analysis and a family study. NeuroImage 27, 960–968.
Clark, C.R., McFarlane, A.C., Weber, D.L., Battersby, M., 1996.Enlarged frontal P300 to stimulus change in panic disorder. Biol.Psychiatry 39 (10), 845–856.
Coull, J.T., 1998. Neural correlates of attention and arousal: insightsfrom electrophysiology, functional neuroimaging and psychophar-macology. Prog. Neurobiol. 55 (4), 343–361.
Dichter, G.S., van der Stelt, O., Boch, J.L., Belger, A., 2006. Relationsamong intelligence, executive function, and P300 event relatedpotentials in schizophrenia. J. Nerv. Ment. Dis. 194 (3), 179–187.
Ettinger, U., Kumari, V., Crawford, T.J., Corr, P.J., Das, M.,Zachariah, E., Hughes, C., Sumich, A.L., Rabe-Hesketh, S.,Sharma, T., 2004. Smooth pursuit and antisaccade eye movementsin siblings discordant for schizophrenia. J. Psychiatr. Res. 38 (2),177–184.
First, M.B., Spitzer, R.L., Gibbon, M., Williams, J.B.W., 1996a.Structured Clinical Interview for DSM-IV Axis I DisordersResearch Version (SCID-I). New York State Psychiatric Institute,Biometrics Research.
First, M.B., Gibbon, M., Spitzer, R.L., Williams, J.B.W., 1996b.Structured Clinical Interview for DSM-IV Axis II DisordersResearch Version (SCID-II). New York State Psychiatric Institute,Biometrics Research.
Frangou, S., Sharma, T., Alarcon, G., Sigmudsson, T., Takei, N.,Binnie, C., Murray, R.M., 1997. The Maudsley Family Study, II:endogenous event-related potentials in familial schizophrenia.Schizophr. Res. 23, 45–53.
Friedman, D., Cycowicz, Y.M., Gaeta, H., 2001. The novelty P3: anevent-related brain potential (ERP) sign of the brain's evaluation ofnovelty. Neurosci. Biobehav. Rev. 25 (4), 355–373.
Gallinat, J., Mulert, C., Bajbouj, M., Herrmann, W.M., Schunter, J.,Senkowski, D., Moukhtieva, R., Kronfeldt, D., Winterer, G., 2002.

276 A. Sumich et al. / Schizophrenia Research 98 (2008) 265–277
Frontal and temporal dysfunction of auditory stimulus processingin schizophrenia. NeuroImage 17, 110–127.
Gershon, E.S., Guroff, J.J., 1984. Information from relatives: diagnosisof affective disorders. Arch. Gen. Psychiatry 41, 173–180.
Golob, E.J., Starr, A., 2004. Serial position effects in auditory event-related potentials during working memory retrieval. J. Cogn.Neurosci. 16 (1), 40–52.
Grawe, R.W., Levander, S., 1995. Smooth pursuit eye movements andneuropsychological impairments in schizophrenia. Acta Psychiatr.Scand. 92 (2), 108–114.
Gruzelier, J.H., 2003. Theory, methods and new directions in thepsychophysiology of the schizophrenic process and schizotypy.Int. J. Psychophysiol. 48 (2), 221–245.
Gruzelier, J., Richardson, A., Liddiard, D., Cheema, S., Puri, B.,McEvedy, C., Rippon, G., 1999. Opposite patterns of P300asymmetry in schizophrenia are syndrome related. Int. J. Psycho-physiol. 34 (3), 275–282.
Heaton, R.K., 1993. Wisconsin Card Sorting Test. PsychologicalAssessment Resources Inc.
Hegerl, U., Gallinat, J., Juckel, G., 2001. Event-related potentials. Dothey reflect central serotonergic neurotransmission and do theypredict clinical response to serotonin agonists? J. Affect. Disord.62 (1–2), 93–100.
Higashima, M., Nagasawa, T., Oka, T., Tsukada, T., Okamoto, T., Komai,Y., Kawasaki, Y., Koshino, Y., 2005. Neuropsychological correlates ofan attention-related negative component elicited in an auditory oddballparadigm in schizophrenia. Neuropsychobiology 51 (4), 177–182.
Hughes, C., Kumari, V., Das, M., Zachariah, E., Ettinger, U., Sumich,A., Sharma, T., 2005. Cognitive functioning in siblings discordantfor schizophrenia. Acta Psychiatr. Scand. 111 (3), 185–192.
Jeon, Y.W., Polich, J., 2003. Meta-analysis of P300 and schizophrenia:patients, paradigms, and practical implications. Psychophysiology40, 684–701.
Karoumi, B., Laurent, A., Rosenfeld, F., Rochet, T., Brunon, A.M.,Dalery, J., d'Amato, T., Saoud, M., 2000. Alteration of eventrelated potentials in siblings discordant for schizophrenia.Schizophr. Res. 41, 325–334.
Kendler, K.S., Lieberman, J.A., Walsh, D., 1989. The StructuredInterview for Schizotypy (SIS): a preliminary report. Schizophr.Bull. 15, 559–571.
Kim, M.S., Kang, S.S., Youn, T., Kang, D.H., Kim, J.J., Kwon, J.S.,2003. Neuropsychological correlates of P300 abnormalities inpatients with schizophrenia and obsessive–compulsive disorder.Psychiatry Res. 123 (2), 109–123.
Knight, R.T., 1996. Contribution of human hippocampal region tonovelty detection. Nature 383, 256–259.
Kumari, V., Das, M., Zachariah, E., Ettinger, U., Sharma, T., 2005.Reduced prepulse inhibition in unaffected siblings of schizophre-nia patients. Psychophysiology 42 (5), 588–594.
McConaghy, N., Catts, S.V., Michie, P.T., Fox, A., Ward, P.B., Shelley,A.M., 1993. P300 indexes thought disorder in schizophrenics, butallusive thinking in normal subjects. J. Nerv. Ment. Dis. 181 (3),176–182.
Molina, V., Munoz, F., Martin-Loeches, M., Casado, P., Hinojosa, J.A.,Iglesias, A., 2004. Long-term olanzapine treatment and p300parameters in schizophrenia. Neuropsychobiology 50, 182–188.
Moreno-Iniguez, M., Ortuno, F., Bonelli, R., Millan, M., Soutullo, C.,Cervera-Enguiz, S., 2007. Perseverative error in schizophrenia:correlation with cortical blood flow by SPECT. Actas Esp.Psiquiatr. 35 (1), 20–28 (Jan–Feb).
Mulert, C., Menzinger, E., Leicht, G., Pogarell, O., Hegerl, U., 2005.Evidence for a close relationship between conscious effort and
anterior cingulate cortex activity. Int. J. Psychophysiol. 56 (1),65–80.
Nieman, D.H., Koelman, J.H., Linszen, D.H., Bour, L.J., Dingemans,P.M., Ongerboer de Visser, B.W., 2002. Clinical and neuropsy-chological correlates of the P300 in schizophrenia. Schizophr. Res.55 (1–2), 105–113.
Nishitani, N., Ikeda, A., Nagamine, T., Honda, M., Mikuni, N., Taki,W., Kimura, J., Shibasaki, H., 1999. The role of the hippocampusin auditory processing studied by event-related electric potentialsand magnetic fields in epilepsy patients before and after temporallobectomy. Brain 122 (Pt 4), 687–707.
Oldfield, R.C., 1971. The assessment and analysis of handedness: theEdinburgh inventory. Neuropsychologia 9, 97–113.
Oranje, B., van Berckel, B.N., Kemner, C., van Ree, J.M., Kahn, R.S.,Verbaten, M.N., 2000. The effects of a sub-anaesthetic dose ofketamine on human selective attention. Neuropsychopharmacol.22 (3), 293–302.
Reitan, R.M., Wolfson, D., 1985. The Halstead–Reitan Neuropsycho-logical Test Battery — Theory and Clinical Interpretation.Neuropsychology Press, Tuscon, Ariz.
Rey, A., 1964. L'examen clinique en psychologie. Paris.Salisbury, D.F., Griggs, C.B., Shenton, M.E., McCarley, R.W., 2004.
The NoGo P300 ‘anteriorization’ effect and response inhibition.Clin. Neurophysiol. 115, 1550–1558.
Schall, U., Schon,A., Zerbin, D., Eggers, C., Oades, R.D., 1996. Event-related potentials during an auditory discrimination with prepulseinhibition in patients with schizophrenia, obsessive–compulsivedisorder and healthy subjects. Int. J. Neurosci. 84 (1–4), 15–33.
Semlitsch, H.V., Anderer, P., Schuster, P., Presslich, O., 1986. Asolution for reliable and valid reduction of ocular artifacts, appliedto the P300 ERP. Psychophysiology 23, 695–703.
Squires-Wheeler, E., Friedman, D., Amminger, G.P., Skodol, A.,Looser-Ott, S., Roberts, S., Pape, K., Erlenmeyer-Kimling, L.,1997. Negative and positive dimensions of schizotypal personalitydisorder. J. Personal. Disord. 11 (3), 285–300.
Stroop, J.R., 1935. Studies of interference in series verbal reactions.J. Exp. Psychol. 18, 643–662.
Sumich, A., Harris, A., Flynn, G., Whitford, T., Tunstall, N., Kumari,V., Brammer, M., Gordon, E., Williams, L.M., 2006. Event-relatedpotential correlates of depression, insight and negative symptomsin males with recent-onset psychosis. Clin. Neurophysiol. 117 (8),1715–1727.
Sumich, A., Kumari, V., Gordon, E., Tunstall, N., Brammer, M.,revised submission. Event-related potential correlates of paranor-mal ideation and unusual experiences.
Takahashi, H., Kato, M., Hayashi, M., Okubo, Y., Takano, A., Ito, H.,Suhara, T., 2007. Memory and frontal lobe functions; possiblerelations with dopamine D2 receptors in the hippocampus.NeuroImage 34 (4), 1643–1649 (Electronic publication 2006Dec 15).
Thermenos, H.W., Seidman, L.J., Breiter, H., Goldstein, J.M.,Goodman, J.M., Poldrack, R., Faraone, S.V., Tsuang, M.T.,2004. Functional magnetic resonance imaging during auditoryverbal working memory in nonpsychotic relatives of persons withschizophrenia: a pilot study. Biol. Psychiatry 55 (5), 490–500.
Turetsky, B.I., Colbath, E.A., Gur, R.E., 1998. P300 subcomponentabnormalities in schizophrenia: I. Physiological evidence forgender and subtype specific differences in regional pathology.Biol. Psychiatry. 4 (2), 84–96.
Turetsky, B.I., Cannon, T.D., Gur, R.E., 2000. P300 subcomponentabnormalities in schizophrenia: III. Deficits In unaffected siblingsof schizophrenic probands. Biol. Psychiatry 47, 380–390.

277A. Sumich et al. / Schizophrenia Research 98 (2008) 265–277
van Veen, V., Carter, C.S., 2005. Separating semantic conflict andresponse conflict in the Stroop task: a functional MRI study.NeuroImage 27 (3), 497–504.
Vernon, D., Haenschel, C., Dwivedi, P., Gruzelier, J., 2005. Slowhabituation of induced gamma and beta oscillations in associationwith unreality experiences in schizotypy. Int. J. Psychophysiol. 56(1), 15–24 (Electronic publication 2004 Nov).
Vitali, P., Abutalebi, J., Tettamanti, M., Rowe, J., Scifo, P., Fazio, F.,Cappa, S.F., Perani, D., 2005. Generating animal and tool names:an fMRI study of effective connectivity. Brain Lang. 93 (1),32–45.
Waldo, M.C., Adler, L.E., Freedman, R., 1988. Defects in auditorysensory gating and their apparent compensation in relatives ofschizophrenics. Schizophr. Res. 1 (1), 19–24.
Wang, W., Wang, Y., Fu, X., Liu, J., He, C., Dong, Y., Livesley, W.J.,Jang, K.L., 2006. Cerebral information processing in personality
disorders: I. Intensity dependence of auditory evoked potentials.Psychiatry Res. 141 (2), 173–183.
Ward, P.B., Catts, S.V., McConaghy, N., 1992. P300 and conceptualloosening in normals: an event-related potential correlate of“thought disorder?”. Biol. Psychiatry 31 (7), 650–660 (Apr 1).
Wechsler, D., 1987. Wechsler Memory Scale-Revised. The Psycho-logical Corporation, New York.
Winterer, G., Egan, M.F., Radler, T., Coppola, R., Weinberger, D.R.,2001. Event-related potentials and genetic risk for schizophrenia.Biol. Psychiatry 50, 407–417.
Winterer, G., Egan, M.F., Raedler, T., Sanchez, C., Jones, D.W.,Coppola, R., Weinberger, D.R., 2003. P300 and genetic risk forschizophrenia. Arch. Gen. Psychiatry 60, 1158–1167.
Wong, K.K., Grunstein, R.R., Bartlett, D.J., Gordon, E., 2006. Brainfunction in obstructive sleep apnea: results from the Brain ResourceInternational Database. J. Integr. Neurosci. 5 (1), 111–121.




















