
Myeloid Dendritic Cells from Human Cutaneous Squamous Cell Carcinoma Are Poor Stimulators of T-Cell...
22
Myeloid dendritic cells from human squamous cell carcinoma are poor stimulators of T cell proliferation Mark J. Bluth 1 , Lisa C. Zaba 2 , Dariush Moussai 1 , Mayte Suárez-Fariñas 2,3 , Helen Kaporis 2 , Linda Fan 1 , Katherine C. Pierson 2 , Traci R. White 2 , Alexander Pitts-Kiefer 1 , Judilyn Fuentes- Duculan 2 , Emma Guttman-Yassky 2 , James G. Krueger 2 , Michelle A. Lowes 2 , and John A. Carucci 1,a 1 Department of Dermatology, Weill Medical College of Cornell University, 1305 York Ave, New York, NY, 10021 2 Laboratory for Investigative Dermatology, The Rockefeller University, 1230 York Ave, New York, NY, 10065 3 Center for Clinical and Translational Science, The Rockefeller University, 1230 York Ave, New York, NY, 10065 Abstract In order to determine the phenotype and function of myeloid DCs from human cutaneous squamous cell carcinoma (SCC), we studied their surface marker expression and allo-stimulatory potential ex vivo. There were abundant CD11c + myeloid DCs, as well as TNF and inducible nitric oxide synthase (iNOS)-producing DCs (TIP-DCs), in and around SCC tumor nests. Although myeloid DCs from SCC, adjacent non-tumor bearing skin, and normal skin, were phenotypically similar by flow cytometry, and there was a pronounced genomic signature of mature DCs in SCC, they showed different T cell stimulatory potential in an allo-MLR. Myeloid DCs from SCC were less potent stimulators of allogeneic T cell proliferation than DCs from non-tumor bearing skin. Culture with a DC-maturing cytokine cocktail (IL-1β, IL-6, TNF, and PGE 2 ) enhanced stimulatory potential in DCs from non-tumor-bearing skin, while SCC associated DCs remained poor stimulators of T cell proliferation. The microenvironment associated with SCC showed expression of TGFβ, IL-10 and VEGF-A, factors capable of suppressing DC function. These findings indicate that CD11c + /HLA- DR hi DCs from SCC are mature, but are not potent stimulators of T cell proliferation compared with phenotypically similar DCs isolated from non-tumor-bearing skin. Identification of mechanisms responsible for suppression of tumor-associated DCs may provide insight into evasion of immunosurveillance by SCC. INTRODUCTION Squamous cell carcinoma (SCC) is the second most common human cancer affecting over 250,000 individuals in the United States in 2007 (Weinberg et al., 2007). In most cases, cutaneous SCC is treated successfully by surgery, however it has the potential to behave aggressively and accounts for the majority of ~10,000 non-melanoma skin cancer deaths in the US each year (Thomas et al., 2007). Aggressive behavior in SCC is associated with immune compromise in the host manifested by increased incidence in solid organ transplant recipients and individuals with chronic lymphocytic leukemia (CLL) (Vanacker et al., 2008). Based on aCorresponding author John A. Carucci Department of Dermatology Weill Medical College of Cornell University, 1305 York Ave, New York, NY, 10021 Ph: 646-962-5574 Fax: 646-962-0004 [email protected]. NIH Public Access Author Manuscript J Invest Dermatol. Author manuscript; available in PMC 2010 October 1. Published in final edited form as: J Invest Dermatol. 2009 October ; 129(10): 2451–2462. doi:10.1038/jid.2009.96. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
-
Upload
rockefeller -
Category
Documents
-
view
2 -
download
0
Transcript of Myeloid Dendritic Cells from Human Cutaneous Squamous Cell Carcinoma Are Poor Stimulators of T-Cell...

Myeloid dendritic cells from human squamous cell carcinoma arepoor stimulators of T cell proliferation
Mark J. Bluth1, Lisa C. Zaba2, Dariush Moussai1, Mayte Suárez-Fariñas2,3, Helen Kaporis2,Linda Fan1, Katherine C. Pierson2, Traci R. White2, Alexander Pitts-Kiefer1, Judilyn Fuentes-Duculan2, Emma Guttman-Yassky2, James G. Krueger2, Michelle A. Lowes2, and John A.Carucci1,a1 Department of Dermatology, Weill Medical College of Cornell University, 1305 York Ave, NewYork, NY, 100212 Laboratory for Investigative Dermatology, The Rockefeller University, 1230 York Ave, New York,NY, 100653 Center for Clinical and Translational Science, The Rockefeller University, 1230 York Ave, NewYork, NY, 10065
AbstractIn order to determine the phenotype and function of myeloid DCs from human cutaneous squamouscell carcinoma (SCC), we studied their surface marker expression and allo-stimulatory potential exvivo. There were abundant CD11c+ myeloid DCs, as well as TNF and inducible nitric oxide synthase(iNOS)-producing DCs (TIP-DCs), in and around SCC tumor nests. Although myeloid DCs fromSCC, adjacent non-tumor bearing skin, and normal skin, were phenotypically similar by flowcytometry, and there was a pronounced genomic signature of mature DCs in SCC, they showeddifferent T cell stimulatory potential in an allo-MLR. Myeloid DCs from SCC were less potentstimulators of allogeneic T cell proliferation than DCs from non-tumor bearing skin. Culture with aDC-maturing cytokine cocktail (IL-1β, IL-6, TNF, and PGE2) enhanced stimulatory potential in DCsfrom non-tumor-bearing skin, while SCC associated DCs remained poor stimulators of T cellproliferation. The microenvironment associated with SCC showed expression of TGFβ, IL-10 andVEGF-A, factors capable of suppressing DC function. These findings indicate that CD11c+/HLA-DRhi DCs from SCC are mature, but are not potent stimulators of T cell proliferation compared withphenotypically similar DCs isolated from non-tumor-bearing skin. Identification of mechanismsresponsible for suppression of tumor-associated DCs may provide insight into evasion ofimmunosurveillance by SCC.
INTRODUCTIONSquamous cell carcinoma (SCC) is the second most common human cancer affecting over250,000 individuals in the United States in 2007 (Weinberg et al., 2007). In most cases,cutaneous SCC is treated successfully by surgery, however it has the potential to behaveaggressively and accounts for the majority of ~10,000 non-melanoma skin cancer deaths in theUS each year (Thomas et al., 2007). Aggressive behavior in SCC is associated with immunecompromise in the host manifested by increased incidence in solid organ transplant recipientsand individuals with chronic lymphocytic leukemia (CLL) (Vanacker et al., 2008). Based on
aCorresponding author John A. Carucci Department of Dermatology Weill Medical College of Cornell University, 1305 York Ave, NewYork, NY, 10021 Ph: 646-962-5574 Fax: 646-962-0004 [email protected].
NIH Public AccessAuthor ManuscriptJ Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
Published in final edited form as:J Invest Dermatol. 2009 October ; 129(10): 2451–2462. doi:10.1038/jid.2009.96.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

the potential for host immunity to regulate behavior in SCC, we evaluated the immunemicroenvironment and function of tumor-associated myeloid DCs.
DCs are professional antigen presenting cells that mediate innate and adaptive immunity, andmay play a key role in anti-tumor response (Steinman, 2006). Mature DCs are identified byexpression of HLA-DR, CD86, CD83, CD80, CD40 and other markers, and their ability tostimulate naïve T cell proliferation. There are three major subsets of cutaneous DCs in humans:myeloid DCs, plasmacytoid DCs (pDCs) and Langerhans cells (Zaba et al., 2008b). There havebeen reports in recent years describing tumor-associated DCs and how the numbers and typesof DCs present may serve as a prognostic indicator, including colorectal, cervical carcinoma,esophageal SCC, nasopharyngeal carcinoma, urinary bladder carcinoma and gastric carcinoma.For example, there was a significant increase in the number of immature DCs in late stageovarian carcinoma-associated ascites compared to peripheral blood, and there was asignificantly greater number of ascitic pDCs than myeloid DCs in many of these patients(Siegal et al., 1999; Wertel et al., 2008a; Wertel et al., 2008b). These observations are typicallyindicative of a poor prognosis for these patients (Zou et al., 2001). An increased presence ofpDCs has also been correlated with a poor prognosis for patients with SCC of the head andneck (SCC of the oral and nasal mucosa) (Hartmann et al., 2003; O'Donnell et al., 2007).
In vitro tumor antigen- or tumor lysate-pulsed monocyte-derived myeloid DCs (MoDCs) canbe generated for use as a specific anti-tumor vaccine (Lee et al., 2002). For example, in aclinical trial of 16 patients with stage IV melanoma, two complete, three partial and one minorresponse was reported after treatment with melanoma tumor or peptide pulsed MoDCs (Nestleet al., 1998). This strategy has been variably successful in different cancer settings, but supportsthe anti-tumor potential of these DCs (Dhodapkar et al., 2002; Jeong et al., 2007; Nestle etal., 2001; Pospisilova et al., 2002).
We have previously characterized the immune infiltrate in basal cell carcinoma (BCC) (Kaporiset al., 2007). That study showed that there was an increased number of CD11c+ DCs in theBCC tumor microenvironment compared to normal skin, suggesting a role for DCs in this typeof cancer. We have now turned our attention to DCs in cutaneous SCC, as their potential roleand function in this human epithelial cancer remains undefined. We studied the DCmicroenvironment in human SCC and evaluated the populations, numbers, phenotypes andstimulatory capacity of DCs from SCC, peritumoral non-lesional skin (PTNL), and normalskin. We found the following: 1) SCC is associated with appreciable numbers of CD1a+ cells,Langerin+ cells, CD11c+ cells, and BDCA1+ cells; 2) the SCC microenvironment is alsoassociated with TNF and inducible nitric oxide-producing myeloid DCs (TIP-DCs); 3)CD11c+/HLA-DRhi myeloid DCs from SCC express maturity markers at levels similar to thosefrom patient-matched, site-matched PTNL skin, and normal skin from tumor-free subjects; 4)SCC is associated with a mature DC gene expression signature; 5) myeloid DCs from SCC arepoor stimulators of allogeneic T cell proliferation compared to myeloid DCs from PTNL skinand normal skin; 6) and the tumor microenvironment is associated with an increased expressionof immunosuppressive cytokines.
RESULTSHuman cutaneous SCC is associated with intratumoral CD1a+/Langerin+ Langerhans cellsand juxtatumoral CD11c+ myeloid DCs and BDCA2+ pDCs
SCC and normal skin were evaluated for the presence of DCs by immunohistochemistry.Representative immunohistochemistry is shown for each antigen analyzed, in normal skin andSCC and (n=10-12 for each group), and cell counts of all the cases are shown (Figure 1, right).We determined the numbers of DCs within the SCC microenvironment based on those countedwithin the tumor nodules and in the papillary dermis 100μm immediately around the tumor
Bluth et al. Page 2
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

nodules (juxtatumoral). The numbers of DCs in normal skin were evaluated based on thoseobserved in the epidermis and those found in the normal papillary dermis 100μm deep to thebasement membrane.
In SCC, CD1a+ and Langerin+ cells were infiltrating epithelial tumor aggregates at numberslower than those per unit area of normal epidermis (p=0.016 and p=0.003 respectively) (Figure1A, and B). There were even less Langerhans cells in juxtatumoral skin compared to SCC(p<0.001). There were greater numbers of CD11c+ myeloid DCs in the juxtatumoral dermiscompared to SCC tumor nodules (p<0.001), with relatively few CD11c+ myeloid DCs in SCCscompared to normal skin (p=0.0012). (Figure 1C). There were significantly greater numbersof BDCA-1+ cells observed in both normal dermis (p<0.05) and juxtatumoral dermis comparedto SCC (p<0.005) (Figure 1D). There were higher numbers of BDCA-2+ pDCs in thejuxtatumoral dermis compared to normal papillary dermis (p<0.002), fewer pDCs infiltratingSCC tumor nodules compared to juxtatumoral dermis (p<0.025), but greater numbers of pDCswere observed in SCC tumor nodules than in normal papillary dermis (p<0.05) (Figure 1E).
TNFα and inducible nitrous oxide (iNOS) -producing (TIP) DCs are associated with the SCCmicroenvironment
TIP-DCs have been described in imiquimod-treated BCC (Stary et al., 2007), and recent studiesfrom our group described the presence of TIP-DCs in psoriasis (Lowes et al., 2005). Treatmentof psoriasis patients with efalizumab (anti-CD11a, Raptiva) strongly reduces infiltration bythese DCs in patients responding to this agent, suggesting that TIP-DCs may play a role inkeratinocyte hyperproliferation. Since SCC is also a condition involving keratinocytehyperproliferation, we wanted to determine if TIP-DCs were present in the SCCmicroenvironment. Triple-label immunoflourescence shows that there are CD11c+ (blue)myeloid DCs that also express TNFα (green) and iNOS (red), which appear white in color(Figure 2, representative image).
CD11c+/HLA-DRhi myeloid DCs from human SCC express maturity markersWe wanted to characterize the surface phenotype of CD11c+ myeloid DCs associated withSCC. CD11c+/HLA-DRhi myeloid DCs from normal skin, PTNL skin and SCC were evaluatedfor expression of co-stimulatory and maturity markers DC-LAMP/CD208, DC-SIGN/CD209,DEC-205/CD205, CD83, CD86, and CD80 by flow cytometry. Representative FACShistograms for selected markers are shown; MFI is mean of patients studied (n=3) (Figure 3).
Expression of DC-LAMP was slightly lower in SCC-derived myeloid DCs (MFI=1068)compared to those from PTNL and normal skin (MFI=1228 and 1492 respectively). DC-SIGNexpression was slightly higher in myeloid DCs from SCC (MFI=1938) compared to those fromPTNL skin, but lower than those from normal skin (MFI=1759 and 2278 respectively). MyeloidDCs from SCC expressed greater levels of DEC-205 (MFI=32963) than PTNL and normalskin (MFI=27513 and 18616 respectively). SCC-associated myeloid DCs expressed CD83(MFI=2963) but at slightly lower levels than those from site-matched PTNL and normal skin(MFI=3959 and 3527 respectively). Myeloid DCs from SCC expressed higher levels of CD86(MFI=788) compared with site matched PTNL and normal skin MFI=566 and 414respectively). Myeloid DCs from SCC express lower levels of CD80 (MFI=283) than thosefrom site-matched PTNL and normal skin (MFI=387 and 662 respectively). Based on theseresults it appears that SCC-associated myeloid DCs achieved slightly increased expression ofmaturation marker DEC-205 and co-stimulatory molecule CD86 than PTNL skin and normalskin, but all three groups have a similar overall phenotype.
Bluth et al. Page 3
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

SCC is associated with a mature DC gene expression signatureThe genomic signature of DC genes was analyzed in our samples, since the surface phenotypeof myeloid DC in SCCs was relatively mature and similar to DCs in adjacent PTNL skin. Thefold change for DC genes in four lists was evaluated, as described in material and methods.Figure 4 shows the mean fold change of DC gene sets for SCC vs. normal skin (black bars)and PTNL vs. normal skin (gray bars). Mature DC genes showed increased mean expressionin SCC (p<0.004). In PTNL vs. normal skin, all gene lists showed increased expression(p<0.05). The significance of those findings was also tested using Gene Set EnrichmentAnalysis (GSEA) (Table 1). DC gene expression was significantly enriched in PTNL skincompared to normal skin (Hohenkirk_DC_Up, p=0.024), while in SCC, enrichment of matureDC genes is increased (NES =1.25) but did not reach statistical significance (p=0.12). Overall,these data indicate DC gene activation in and around human SCC.
DCs from SCC are poor stimulators of allogeneic T cell proliferationSince CD11c+/HLA-DRhi DCs from SCC and PTNL skin expressed similar levels of maturitymarkers we measured the capacity of these cells to stimulate allogeneic T cell proliferation inan allo-MLR. DCs were isolated from SCC and adjacent PTNL skin and co-cultured withallogeneic T cells for 8 days. Representative dot plots for CFSE vs. CD3 are shown for controlsand each condition, and the gate for proliferating cells is shown (Figure 5A). A summary ofall MLR data is shown in Figure 5B, and results tabulated in Table 2. On day 8, allogeneic Tcell proliferation determined by CFSE dilution showed that SCC-derived myeloid DCs were2-3 fold weaker stimulators of allogeneic T cell proliferation compared to DCs derived frommatched PTNL, but they were similar to normal skin (Figure 5A). Treatment with a maturationcocktail containing cytokines including: IL1β, IL6, TNFα and PGE2, increased stimulatorypotential of peritumoral DCs slightly, but was less effective in enhancing stimulatory potentialof DCs from SCC than normal skin (Figure 5, right lower three panels).
The SCC microenvironment is associated with an increased expression of cytokines thatmay suppress DC function
Since myeloid DCs from SCC display maturity markers, yet lack appreciable allostimulatorypotential, the level of expression of soluble immunosuppressive factors, IL-10, TGF-β andVEGF-A, was evaluated in the tumor microenvironment compared to PTNL and normal skin.The mean IL-10 mRNA expression was increased in both SCC and PTNL skin compared tonormal skin. However, these increases were not significant (p=0.086 and p=0.199,respectively) (Figure 6, left panel). The mean expression of TGF-β mRNA was significantlyincreased in SCC compared with both PTNL and normal skin (p<0.027 and p<0.001,respectively) (Figure 6, middle panel) and in PTNL compared to normal skin (p<0.001). Therewas also a significantly higher mean level of expression of VEGF-A mRNA in both SCC andPTNL skin compared to normal skin (p<0.001 for both) and the mean level of VEGF-A mRNAwas significantly increased in PTNL skin compared to SCC (p<0.041) (Figure 6, right panel).
DISCUSSIONSeveral findings stand out from the results reported here: 1) human cutaneous SCC wasassociated with CD11c+ and BDCA1+ cells in the juxtatumoral dermis; 2) CD1a+ andLangerin+ Langerhans cells were found in epithelial tumor nests, but at lower concentrationsper unit area than normal epithelium; 3) the SCC microenvironment was associated withCD11c+ TIP DCs; 4) CD11c+/HLA-DRhi myeloid DCs from SCC and site-matchedperitumoral skin express fundamentally similar levels of maturation markers; 5) expressionarray analysis indicated a mature DC pathway signature in SCCs; 6) myeloid DC from SCCwere poor stimulators of allogeneic T cell proliferation compared with DCs from adjacentPTNL skin; 7) treatment with maturation cytokines enhanced stimulatory potential of DCs
Bluth et al. Page 4
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

from normal skin and peritumoral skin but had less effect on DCs from SCC; and 8) the SCCtumor microenvironment was associated with an increased expression of immunosuppressivecytokines.
Thus, our initial characterization of the myeloid DCs in SCC demonstrated that while theseDCs were as phenotypically mature as normal skin DCs, they were impaired in their ability toupregulate their allostimulatory capacity compared to those obtained from normal skin. Thedifference in allostimulation is most noticeable when the myeloid DCs from both SCC andnormal skin were treated with maturing cytokines. A similar observation was described byZaba et al (Zaba et al., 2007b) and is due to the “steady-state” myeloid DCs found in normalskin not being fully mature, but may become so when immunologically or otherwisechallenged. We also demonstrated that although located only 2-5mm from the tumor margin,DCs derived from PTNL skin were consistently better stimulators of allogeneic T cells thanthose derived from SCC. Hence, although myeloid DCs from SCC and PTNL skin expressedsimilarly intermediate level of maturity, they differed in allogeneic T cell stimulatory capacity.
However, there was an abundant genomic signature of both immature and mature DCs in SCClesions, so we consider that the tumor may be secreting factors causing this paralysis.Candidates include IL-10 and TGF-β, both of which were elevated in lesional tissue and sun-damaged skin adjacent to the tumor. Both these cytokines may have an immunosuppressiverole (Mimura et al., 2007; Steinbrink et al., 1999). VEGF-A, while not increased in SCC, wasincreased in peritumoral skin compared to normal skin, and has been reported to decrease thestimulatory function of mature MoDCs (Mimura et al., 2007). This observation is noteworthy,because it may help explain why myeloid DCs from peritumoral skin still significantly lackstimulatory capacity compared to those from normal skin even when treated with maturingcytokines. VEGF-A isoforms are designated based on their peptide lengths, the most wellstudied being VEGF-A121, VEGF-A165 and VEGF-A189, all of which have pro-angiogeniceffects. However, Ladomery et al has described the expression of another class of VEGF-Aisoforms derived from alternative 3’ splicing of the mRNAs that correspond to the above-mentioned VEGF-A proteins (Ladomery et al., 2007). These splice variants produce peptidesof nearly identical lengths (i.e. VEGF-A121b, VEGF-A165b and VEGF-A189b), but have anti-angiogenic effects. The splice variant mRNAs for VEGF-A165 and VEGF-A165b producepeptides that are identical except for the last 6 C-terminal amino acids. Since the commerciallyavailable RT-PCR primer-probe set we used may identify both pro- and anti-angiogenicisoforms of VEGF-A, it may be that the increased expression of VEGF-A in normal skinrepresents the anti-angiogenic isoform and the level of expression of VEGF-A in SCCrepresents the level of the pro-angiogenic isoform. Bates et al address this distinction for anumber of normal tissues in their study (Bates et al., 2002). We are currently working to definedifferential VEGF-A isoform expression in the SCC microenvironment. Thus tumors maydevelop many mechanisms by which to slow or disable myeloid DC anti-tumoral activity.
In addition to tumor-derived factors that may impair local DC function, there are otherleukocytes that appear to promote tumor growth, including immature DCs, pDCs, regulatoryDCs, and regulatory T cells (Berger et al., 2002; Dauer et al., 2003; Enk et al., 1997; Talmadgeet al., 2007). There is also a heterogeneous population of myeloid cells that may have tumorpromoting functions, termed myeloid-derived suppressor cells (MDSC). This group of cellsincludes granulocytic, macrophage, DC and early myeloid progenitors (Apolloniet al., 2000;Kusmartsev and Gabrilovich, 2005; Young and Lathers, 1999; Zea et al., 2005). In humansthey are identified as CD3−, CD11b+, CD14−, CD19−, CCD33+, CD56−, and HLA-DR−.MDSCs can inhibit T cell activation and antigen-specific proliferation (Movahedi et al.,2008). Angiogenic factors such as plasminogen activator inhibitor 1 (PAI-1) correlated withan increased number of immature DCs and MDSCs in the blood of patients with advancedmalignancies (Osada et al., 2008). The mechanisms responsible for immunosuppressive
Bluth et al. Page 5
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

functions of MDSC include production of reactive oxygen species (ROS), nitric oxide (NO),L-arginine metabolism, and cytokines. MDSCs have been shown to prevent the developmentof antigen-specific T cells by decreasing the proximal concentration of L-arginine via thearginase and nitric oxide synthase pathways (Bronte et al., 2003). VEGF has also been directlycorrelated with myeloid DC expansion (Ohm and Carbone, 2001). Therefore, not only cantumor cells modulate the immune system to escape immunosurveillance, but they can also takecontrol of immune cells and use them to actively interfere with continued efforts by the immunesystem to destroy it.
In our study, we have observed the presence of TIP-DCs around the SCC tumor nests. We havepreviously described TIP-DCs in the common inflammatory skin disease psoriasis (Lowes etal., 2005). These cells were first described in a murine model of Listeria monocytogenesinfection (Serbina et al., 2003), and their presence in the spleen of infected mice was CCR2-dependant. We have since determined that these TIP-DCs are part of a group of myeloid dermalDCs that we have termed “inflammatory” DCs. These cells are characterized by the followingsurface markers: CD11c+, HLA-DRhi, and they are BDCA-1/CD1c negative, unlike theresident myeloid DCs found in normal skin in steady state which are BDCA-1/CD1c+ (Zabaet al., 2007b; Zaba et al., 2008b). These inflammatory DCs are able to produce cytokines andinflammatory mediators such as TNF, iNOS, IL-20, and IL-23 (Wang et al., 2006; Zaba etal., 2007a; Zaba et al., 2008a; Zaba et al., 2008b). They are also able to induce allogeneic Tcell proliferation, and Th1 and Th17 cell polarization (Zaba et al., 2008a). Thus, while capableof classic antigen-presenting function ex vivo, we hypothesize that the main role of theseinflammatory DCs in the skin may be that of pro-inflammatory cytokine production critical inthe pathogenesis of psoriasis.
Stary et al were the first to show that TIP-DCs were present in a tumor model, in BCCs aftertreatment with the TLR7/8 agonist imiquimod (Stary et al., 2007). Although they did not showstaining of these tumors for these markers before imiquimod, it was presumed that the treatmentinduced these TIP-DCs. In an effort to further characterize the CD11c+ myeloid DCssurrounding the SCCs, we performed immunofluorescence for TNF and iNOS, expecting anabsence of such mediators. It was a surprise to find abundant dermal TNF and iNOS-producingCD11c+ cells. We have previously demonstrated increased iNOS mRNA in SCCs (Haider etal., 2006).
This finding led us to consider the role of these TIP-DCs, and their relationship to the myeloidDCs we obtained for functional assays. The gating strategy for obtaining cells from the SCCtissue for T cell stimulating capability was CD11c+/HLA-DR+, and it is not possible toretrospectively assess if some of the myeloid DCs obtained from the single cell suspensionswere making TNF-α and iNOS. We will address this question in future studies. These TIP-DCs could be part of the host inflammatory response to the tumor, to try and induce a pro-inflammatory environment and stimulate regression. Current dogma is that certaininflammatory leukocytes are good prognostic indicators, reviewed by Talmadge et al(Talmadge, 2007). For example, mature DCs, IFN-γ primed macrophages, and Th1 cells, areall leukocyte populations that may contribute to tumor regression. The human clinicalsituations where TIP-DCs have been described to date are both in the setting of reduced cancer:in psoriasis, where there is little cutaneous malignancy, and imiquimod-induced regressingBCCs. We have not quantitated these cells or examined sufficient SCCs to conclude this, butit is possible that TIP-DCs are also trying to induce tumor regression, but are impaired in theirefforts by immunosuppressive factors produced by the tumor.
Alternatively, it is possible that the TIP-DCs fall into the group of MDSC, as one of theirdefining mediators is iNOS and subsequently NO. This short-lived mediator can haveimmunosuppressive effects such as inhibiting the proliferation of activated helper T cells
Bluth et al. Page 6
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

(Huang et al., 1998; van der Veen et al., 2000). However, against classifying the TIP-DCs asmyeloid DCs is the fact that TIP-DCs are likely to be HLA-DR+, and MDCS were defined asHLA-DR−. Future investigations will further evaluate the phenotype and function of these TIP-DCs, and how they fit into our working model of resident and “inflammatory” dermal DCs(Zaba et al., 2008b), as well as their contribution to tumor growth or suppression.
Some of the major causes for the lack of immunogenicity in cancer are associated withhypoactive DCs and the presence of regulatory T cells (Ishibashiet al., 2006; Kono et al.,2006; Larkin et al., 2006). The cytokine milieu in the tumor microenvironment may help drivethese associative factors, but appears to depend on the concentration of each cytokine at anygiven time. Additionally, expression of the Th17-associated cytokine, IL-22, may contributeto the hyperproliferation of keratinocytes in SCC as has been described in psoriasis (Chan etal., 2006; Fitch et al., 2007; Lowes et al., 2008; Ma et al., 2008; Nickoloff, 2007). Takentogether, all of these conditions pose a formidable challenge for standard chemotheraputic andbiotheraputic modalities in the treatment of malignant cancers of the skin. However, a betterunderstanding of the cytokine milieu, DC function and tumor evasion of immune responsesmay eventually lead to better immunomodulatory drugs and immune-based vaccine therapiesapplicable to carcinomas not amenable to standard treatments.
MATERIALS AND METHODSApproval from the Weill Cornell Medical College Institutional Review Board and writteninformed consent was obtained before enrolling patients to participate in this study, and thestudy was performed with strict adherence to the Declaration of Helsinki Principles.
Samples used for IHCCutaneous SCC samples were obtained during Mohs micrographic surgery. Tumors wereobtained from head and neck, trunk and extremities. Additionally, normal specimens wereobtained via 3-mm punch biopsies from non-sun exposed areas of patients without skin cancer.Some of these samples may have been used in prior studies as normal controls (Guttman-Yassky et al., 2007).
ImmunohistochemistryFrozen tissue sections were stained as previously described (Kaporis et al., 2007). We stainedsections with hematoxylin (Fisher, Fair Lawn, NJ) and eosin (Shandon, Pittsburgh, PA) andwith purified mouse anti-human monoclonal antibodies to CD1a and CD11c, (BD-Pharmingen, San Diego, CA; all 1:100), Langerin, (Immunotech, Marseille, France 1:50),blood derived dendritic cell antigen (BDCA)-1/CD1c and BDCA-2/CD303 (clones AD5-8E7and AC144 respectively, Miltenyi Biotech, Auburn, CA., both 1:100). Biotin-labeled horseanti-mouse antibody (Vector Laboratories, Burlingame, CA) was amplified with avidin–biotincomplex (Vector Laboratories, Burlingame, CA) and developed with chromogen 3-amino-9-ethylcarbazole (Sigma Aldrich, St Louis, MO). Counterstaining was performed with light green(Sigma Aldrich, St Louis, MO). Appropriate isotype controls were used.
Image analysisTumor-bearing skin and non-tumor-bearing skin adjacent to excised tumor (n=10-19), andnormal skin from volunteers (n=10) were evaluated for the expression of a panel of surfacemarkers by immunohistochemistry. The regions examined within SCC samples weredesignated as “tumor” and “juxtatumoral dermis” (the dermis immediately surrounding thetumor). The regions examined within normal skin samples were designated as normalepidermis and normal (papillary) dermis. Normal papillary dermis and juxtatumoral dermiswere compared in this study based on the structural similarities between these regions. Positive
Bluth et al. Page 7
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

cells were counted manually, and area measures were computed by computer-assisted imageanalysis, National Institutes of Health software (NIH IMAGE 6.1 and IMAGE J). Cell countsper unit area (μm2 × 100000) were determined within normal epidermis, normal papillarydermis (100 μm deep to the epidermis), SCC epithelial aggregates and juxtatumoral dermis(100 μm circumferential to tumor nodules). In all cases, tumor sections with representativeresponse were selected for analysis.
ImmunofluorescenceFrozen sections of squamous cell carcinoma were fixed in 100% acetone for five minutes (n=5).Non-specific staining was blocked with 5% normal goat serum and 5% normal chicken serum(both from Vector, Burlingame, CA). Tissue was then stained overnight at 4°C with twoprimary antibodies: (1) murine anti-TNF-FITC (clone 6401.1111, 1:10, BD Biosciences, SanJose, CA), and (2) iNOS affinity purified rabbit antibody (N-20, 1:20, Santa Cruz, Santa Cruz,CA). The following day, slides were washed and labeled with two secondary antibodyconjugates for 30 minutes at room temperature: (1) goat anti-FITC-Alexa-488, and (2) anti-rabbit Alexa-594. Sections were blocked with 10% mouse serum for 30 minutes and stainedovernight with murine anti-CD11c (B-ly6, 1:100, BD Pharmingen, San Diego, CA) labeledwith Zenon Alexa Fluor-647 Mouse IgG1. All amplification/ detection systems were fromInvitrogen/Molecular Probes (Eugene, OR). Images were acquired using appropriate filters ofa Zeiss Axioplan 2 widefield fluorescence microscope fitted with a PlanNeofluar 10X/ 0.30N.A. objective lens and a Hamamatsu Orca ER CCD camera (Hamamatsu, Bridgewater, NJ)controlled by MetaVue software (MDS Analytical Technologies, Downington, PA). TNF wasvisualized using a FITC filter set, iNOS with a Texas red filter set, and CD11c with a Cy5 filterset. Images in the figure are presented both as single monochrome images, and in pseudocolor(green, red and blue) located above the merged image, so that one can appreciate thelocalization of markers on similar or different cells. Red and green overlapping cells appearyellow in color; red and blue appear purple; and green and blue appear aqua; cells labeled withall three stains appear white. Inset shows triple-labeled cell at higher magnification (4 foldincrease in image size). Dermal collagen fibers gave green auto-fluorescence. Size bar =100μm.
Skin sample preparation for flow cytometry analysis and FACSSCC tumors and sight-matched peritumoral skin were obtained at surgery, and normal skinwas obtained as the discarded product of abdominoplastic surgery. Subcutaneous fat wasexcised, and the remaining tissue was washed three times with PBS and then once with PBScontaining 10% gentamicin reagent solution (Invitrogen, Carlsbad, CA). Both the dermal andepidermal layers were heavily scored with a scalpel then transferred to fresh RPMI 1640supplemented with 10% pooled human serum (Mediatech Inc., Manassas, VA), 0.1%gentamicin reagent solution (Invitrogen, Carlsbad, CA), 1% penicillin-streptomycin solution(Sigma Aldrich, St Louis, MO) and 1% 1 M HEPES buffer (Sigma Aldrich, St Louis, MO).The tissue was incubated for 72 hours in 5% CO2 at 37°C. The supernatant was collected andfiltered with 40μm cell strainers (BD Biosciences, San Jose, CA) yielding a single cellsuspension. Cells were then either used immediately (for mixed leukocyte reaction [MLR]) orfrozen in RPMI 1640 (Invitrogen, Carlsbad, CA) with 50% pooled human serum and 10%DMSO (ATCC) for FACS.
Flow cytometryCellular suspensions (SCC and matched PTNL n=3; normal skin n=3) were washed once withPBS and then stained with AquaMarina Live/Dead cell detection according to themanufacturers instructions (Molecular probes, Invitrogen, Carlsbad, CA) and then stained withthe following anti-human, mouse monoclonal antibodies: HLA-DR – Alexa Fluor 700 (L243,
Bluth et al. Page 8
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

IgG2a, 1:50; BioLegend, San Diego, CA), CD11c – PE-Cy7 (3.9, IgG1, 1:20; BioLegend, SanDiego, CA), dendritic cell lysosome-associated membrane protein - PE (DC-LAMP/CD208-PE) (I10-1112, IgG1, 1:20; BD Pharmingen, San Jose, CA), dendritic cell-specific intercellularadhesion moleule-3-grabbing non-integrin – PerCP-Cy5.5 (DC-SIGN/CD209-PerCP-Cy5.5)(DCN46, IgG2b, 1:20; BD Biosciences, San Jose, CA), DEC-205/CD205-Alexa 647(MMRI-7, IgG1, 1:33; BD Pharmingen, San Jose, CA), CD80-PE (L307.4, IgG1, 1:20; BDBiosciences, San Jose, CA), CD86-Pacific Blue (IT2.2, IgG2b, 1:50; BioLegend, San Diego,CA), and CD83-APC (Michel-19, IgG1, 1:33; BD-Pharmingen, San Jose, CA). Briefly, cellswere stained in a total volume of 100μl for 30 minutes at 4°C, washed with FACSwash (PBSwith 0.2% wt/vol BSA, 0.1% wt/vol sodium azide), and resuspended in 1.3% formaldehyde(Thermo Fisher Scientific, Waltham, MA) in FACSwash. Samples were acquired using a flowcytometer (LSR II; BD Biosciences, San Jose, CA) and analyzed with FlowJo software(TreeStar Inc., Ashland, OR). Appropriate isotype controls were used.
Mixed Leukocyte Reaction (MLR)Single-cell suspensions of dermal cell crawl-outs were stained with mouse anti–human,CD11c-PE (S-CHL-3, IgG2b, 1:20, BD Biosciences, San Jose, CA) and HLA-DR – AlexaFluor 700 monoclonal antibodies, and sorted on a FACSAria (BD Biosciences, San Jose, CA)using a low-pressure setting (n=3-4). A population of CD11c+/HLA-DRhi cells was collectedand a post-sort collection was performed to confirm its purity. T cells were obtained from anormal volunteer by treatment of while blood with RosetteSep T cell enrichment cocktail asper the manufacturers instructions (Stem Cell Technologies, Vancouver, BC, Canada) thensubjected to density centrifugation over Ficoll-Paque Plus (Amersham Biosciences,Piscataway, NJ), and subsequently labeled with 10μm CFSE (Carboxyfluorescein diacetate,succinimidyl ester) using the Vybrant CFDA SE Cell Tracer Kit (Invitrogen, Molecular Probes,Carlsbad, CA) in PBS with 0.1% BSA for 15 minutes at 37oC. We have adapted a modifiedallogeneic mixed leukocyte reaction (MLR) described in Zaba et al. (Zaba et al., 2007b).Briefly, CFSE labeled T cells were co-cultured with sorted CD11c+/HLA-DRhi cells for 8 dayswith and without cytokines for maturing DCs ex-vivo (IL-1β, IL-6, TNF-α, and PGE2), at aratio of 1:50. T cells alone were used as a negative control. T cells cultured with CD3/CD28coated beads (Dynal, Invitrogen, Carlsbad, CA), or immature MoDCs were used as positivecontrols. The process for making monocyte-derived DCs was previously described (Dhodapkaret al., 2002; Lee et al., 2002). T cell proliferation was analyzed on day 8 after sorting. Thecultures were harvested, stained with 250ng/ml propidium iodide (PI, BD Phamingen, SanJose, CA [catalog #51-66211E]), to label dead cells, and CD3-APC (BD Biosciences, San Jose,CA) for 20 minutes at room temperature. PI-negative cells were gated and then plotted as CFSEversus CD3+ cells, where proliferating cells diluted their content of CFSE and moved to theleft of the non-proliferating cells. The CFSE-low cells were quantified as a percentage ofproliferating cells in the culture.
RNA IsolationTotal RNA isolation and RT-PCR reactions were performed as previously described (Kaporiset al., 2007). Briefly, SCC tumor samples, removed at Mohs surgery (n=10-12), and patient-matched site-matched peritumoral non-lesional (PTNL) skin (n=10-12), were obtained at thetime of repair after clear margins were achieved. Normal skin was obtained as the waste fromabdominoplastic surgery (n=4). All samples were snap-frozen and stored in liquid nitrogen.Individual frozen samples were placed in 1ml of room temperature RLT buffer with 1% β-mecaptoethanol (Qiagen, Valencia, CA) and immediately homogenized at full power for 30seconds using a PowerGen 1000 homogenizer (Fisher Scientific, Pittsburgh, PA).Homogenates were sonicated on ice for 20 seconds at full power. DNA was removed with on-column DNAse digestion using an RNAse-free DNAse Set (Qiagen, Valencia, CA). RNA wasisolated using the RNeasy Mini Kit (Qiagen, Valencia, CA) according to manufacturer's
Bluth et al. Page 9
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

recommendations. Total RNA concentration and purity was evaluated using an Ultraspec 2100pro-spectrophotometer (Amersham Biosciences, Piscataway, NJ).
RT-PCRThe primers and probes for IL-10 (Hs00174086_m1), TGF-β (Hs99999918_m1) and VEGF-A (Hs00900054_m1) were TaqMan RT-PCR individual assays designed by AppliedBiosystems (Foster City, CA). The sequences of the primers and probe for HARP are: HARP-forward, CGCTGCTGAACATGCTCAA, HARP reverse, TGTCGAACACCTGCTGGATG;HARP-probe, 6-FAM-TCCCCCTTCTCCTTTGGGCTGG-TAMRA (GenBank accession no.NM-001002). The RT-PCR reaction was performed using 5ng total RNA and EZ PCR CoreReagents (Applied Biosystems, Foster City, CA) according to the manufacturer's directions.The samples were amplified and quantified on an Applied Biosystems PRISM 7900 HT usingthe following thermal cycler conditions: 2 minutes at 50°C, 30 minutes at 60°C, 5 minutes 95°C; and 40 cycles of 15 seconds at 95°C followed by 60 seconds at 60°C. Each sample and genewas normalized to the human acidic ribosomal protein gene, a housekeeping gene. The datawere analyzed and samples quantified by the software provided with the Applied BiosystemsPRISM 7900 HT.
DC maturity pathway analysisSCC microarray data has been previously published (Haider et al., 2007). We wanted toevaluate the fold change for DC genes published in four lists: (1) immature DC upregulatedgenes (Haider et al., 2007), (2) mature DC up-regulated genes (Haider et al., 2007), (3)Lindstedt (Lindstedt et al., 2002) and (4) Hohenkirk monocyte-derived DCs “up” (Le Naouret al., 2001). These lists were similar: ten of the genes in the Hohenkirk list were in the immatureDC list (p< 0.0001), and 9 of the Lindstedt and 11 of the Hohenkirk were in the mature DClist (p< 0.0001 for both) (analyzed in http://www.broad.mit.edu/gsea/index.jsp). We also testedthe significance of gene expression findings using Gene Set Enrichment Analysis (GSEA)(Subramanian et al., 2007). GSEA is a method that determines whether a previously definedset of genes (in our case the DC gene lists) shows statistically significant differences betweentwo biological states (skin samples, for example SCC vs. normal skin) using rank statistics. Ifthe up-regulated genes in the DC list are ranked high in “SCCs vs. normal fold change”, theES score will be near 1, if the opposite effect happens, then the value approaches -1. Becausewe were working with a several lists, a normalized enrichment score (NES) allowed us tocompare the enrichment score between lists.
Statistical analysisStatistical comparisons of relative cell counts per μm2 ×100000 was performed using an Anovamixed model in log2 scale, with P<0.05 considered significant. Statistical comparisons ofmRNA expression levels was performed also using the same Anova mixed model in log2 scalewith P<0.05 considered significant.
AcknowledgmentsResearch was supported by the Dana Foundation (Human Immunology Consortium Grant), which supports JAC, DM,KCP, LF and AP-K. MJB is supported by National Institutes of Health (NIH) grant T32-HL07423, LCZ is supportedby NIH MSTP grant GM07739, MAL is supported by NIH grant 1 K23 AR052404-01A1, and MS-F is partiallysupported by NIH grant UL1 RR024143 from the National Center for Research Resources (NCRR). We thank plasticsurgeons Drs. AN LaBruna and DM Senderoff for their generous donation of abdominoplasty surgical waste. We alsoappreciate the assistance and advice of the Flow Cytometry Core Facility (Dr. S. Mazel, Dr. X. Fan and C. Bare) andBio-imaging Resource Center (Dr. A. North) at Rockefeller University. The authors do not have any conflict of interestrelated to this work.
Bluth et al. Page 10
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

ABBREVIATIONS
SCC squamous cell carcinoma
TIP-DCs TNF- and inducible nitric oxide synthase iNOS-producing DCs
PDCs plasmacytoid DCs
MoDCs monocyte-derived DCs
MDSC myeloid-derived suppressor cells
PTNL peritumoral non-lesional
BCC basal cell carcinoma
REFERENCESApolloni E, Bronte V, Mazzoni A, Serafini P, Cabrelle A, Segal DM, et al. Immortalized myeloid
suppressor cells trigger apoptosis in antigen-activated T lymphocytes. J Immunol 2000;165:6723–6730. [PubMed: 11120790]
Bates DO, Cui TG, Doughty JM, Winkler M, Sugiono M, Shields JD, et al. VEGF165b, an inhibitorysplice variant of vascular endothelial growth factor, is down-regulated in renal cell carcinoma. CancerRes 2002;62:4123–4131. [PubMed: 12124351]
Berger CL, Hanlon D, Kanada D, Dhodapkar M, Lombillo V, Wang N, et al. The growth of cutaneousT-cell lymphoma is stimulated by immature dendritic cells. Blood 2002;99:2929–2939. [PubMed:11929784]
Bronte V, Serafini P, Mazzoni A, Segal DM, Zanovello P. L-arginine metabolism in myeloid cells controlsT-lymphocyte functions. Trends Immunol 2003;24:302–306. [PubMed: 12810105]
Chan JR, Blumenschein W, Murphy E, Diveu C, Wiekowski M, Abbondanzo S, et al. IL-23 stimulatesepidermal hyperplasia via TNF and IL-20R2-dependent mechanisms with implications for psoriasispathogenesis. J Exp Med 2006;203:2577–2587. [PubMed: 17074928]
Dauer M, Pohl K, Obermaier B, Meskendahl T, Robe J, Schnurr M, et al. Interferon-alpha disablesdendritic cell precursors: dendritic cells derived from interferon-alpha-treated monocytes are defectivein maturation and T-cell stimulation. Immunology 2003;110:38–47. [PubMed: 12941139]
Dhodapkar MV, Krasovsky J, Olson K. T cells from the tumor microenvironment of patients withprogressive myeloma can generate strong, tumor-specific cytolytic responses to autologous, tumor-loaded dendritic cells. Proc Natl Acad Sci U S A 2002;99:13009–13013. [PubMed: 12235374]
Enk AH, Jonuleit H, Saloga J, Knop J. Dendritic cells as mediators of tumor-induced tolerance inmetastatic melanoma. Int J Cancer 1997;73:309–316. [PubMed: 9359474]
Fitch E, Harper E, Skorcheva I, Kurtz SE, Blauvelt A. Pathophysiology of psoriasis: recent advances onIL-23 and Th17 cytokines. Curr Rheumatol Rep 2007;9:461–467. [PubMed: 18177599]
Guttman-Yassky E, Lowes MA, Fuentes-Duculan J, Whynot J, Novitskaya I, Cardinale I, et al. Majordifferences in inflammatory dendritic cells and their products distinguish atopic dermatitis frompsoriasis. J Allergy Clin Immunol 2007;119:1210–1217. [PubMed: 17472813]
Haider AS, Lowes MA, Gardner H, Bandaru R, Darabi K, Chamian F, et al. Novel insight into theagonistic mechanism of alefacept in vivo: differentially expressed genes may serve as biomarkers ofresponse in psoriasis patients. J Immunol 2007;178:7442–7449. [PubMed: 17513795]
Haider AS, Peters SB, Kaporis H, Cardinale I, Fei J, Ott J, et al. Genomic analysis defines a cancer-specific gene expression signature for human squamous cell carcinoma and distinguishes malignanthyperproliferation from benign hyperplasia. J Invest Dermatol 2006;126:869–881. [PubMed:16470182]
Hartmann E, Wollenberg B, Rothenfusser S, Wagner M, Wellisch D, Mack B, et al. Identification andfunctional analysis of tumor-infiltrating plasmacytoid dendritic cells in head and neck cancer. CancerRes 2003;63:6478–6487. [PubMed: 14559840]
Bluth et al. Page 11
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Huang FP, Niedbala W, Wei XQ, Xu D, Feng GJ, Robinson JH, et al. Nitric oxide regulates Th1 celldevelopment through the inhibition of IL-12 synthesis by macrophages. Eur J Immunol1998;28:4062–4070. [PubMed: 9862342]
Ishibashi Y, Tanaka S, Tajima K, Yoshida T, Kuwano H. Expression of Foxp3 in non-small cell lungcancer patients is significantly higher in tumor tissues than in normal tissues, especially in tumorssmaller than 30 mm. Oncol Rep 2006;15:1315–1319. [PubMed: 16596204]
Jeong HS, Lee H, Ko Y, Son YI. Vaccinations with dendritic cells primed with apoptotic tumor cells canelicit preventive antitumor immunity in a poorly immunogenic animal model of squamous cellcarcinoma. Laryngoscope 2007;117:1588–1593. [PubMed: 17597627]
Kaporis HG, Guttman-Yassky E, Lowes MA, Haider AS, Fuentes-Duculan J, Darabi K, et al. Humanbasal cell carcinoma is associated with Foxp3+ T cells in a Th2 dominant microenvironment. J InvestDermatol 2007;127:2391–2398. [PubMed: 17508019]
Kono K, Kawaida H, Takahashi A, Sugai H, Mimura K, Miyagawa N, et al. CD4(+)CD25high regulatoryT cells increase with tumor stage in patients with gastric and esophageal cancers. Cancer ImmunolImmunother 2006;55:1064–1071. [PubMed: 16328385]
Kusmartsev S, Gabrilovich DI. STAT1 signaling regulates tumor-associated macrophage-mediated Tcell deletion. J Immunol 2005;174:4880–4891. [PubMed: 15814715]
Ladomery MR, Harper SJ, Bates DO. Alternative splicing in angiogenesis: the vascular endothelialgrowth factor paradigm. Cancer Lett 2007;249:133–142. [PubMed: 17027147]
Larkin J, Tangney M, Collins C, Casey G, O'Brien MG, Soden D, et al. Oral immune tolerance mediatedby suppressor T cells may be responsible for the poorer prognosis of foregut cancers. Med Hypotheses2006;66:541–544. [PubMed: 16288967]
Le Naour F, Hohenkirk L, Grolleau A, Misek DE, Lescure P, Geiger JD, et al. Profiling changes in geneexpression during differentiation and maturation of monocyte-derived dendritic cells using botholigonucleotide microarrays and proteomics. J Biol Chem 2001;276:17920–17931. [PubMed:11279020]
Lee AW, Truong T, Bickham K, Fonteneau JF, Larsson M, Da Silva I, et al. A clinical grade cocktail ofcytokines and PGE2 results in uniform maturation of human monocyte-derived dendritic cells:implications for immunotherapy. Vaccine 2002;20(Suppl 4):A8–A22. [PubMed: 12477423]
Lindstedt M, Johansson-Lindbom B, Borrebaeck CA. Global reprogramming of dendritic cells inresponse to a concerted action of inflammatory mediators. Int Immunol 2002;14:1203–1213.[PubMed: 12356685]
Lowes MA, Chamian F, Abello MV, Fuentes-Duculan J, Lin SL, Nussbaum R, et al. Increase in TNF-alpha and inducible nitric oxide synthase-expressing dendritic cells in psoriasis and reduction withefalizumab (anti-CD11a). Proc Natl Acad Sci U S A 2005;102:19057–19062. [PubMed: 16380428]
Lowes MA, Kikuchi T, Fuentes-Duculan J, Cardinale I, Zaba LC, Haider AS, et al. Psoriasis VulgarisLesions Contain Discrete Populations of Th1 and Th17 T Cells. J Invest Dermatol. 2008
Ma HL, Liang S, Li J, Napierata L, Brown T, Benoit S, et al. IL-22 is required for Th17 cell-mediatedpathology in a mouse model of psoriasis-like skin inflammation. J Clin Invest 2008;118:597–607.[PubMed: 18202747]
Mimura K, Kono K, Takahashi A, Kawaguchi Y, Fujii H. Vascular endothelial growth factor inhibits thefunction of human mature dendritic cells mediated by VEGF receptor-2. Cancer ImmunolImmunother 2007;56:761–770. [PubMed: 17086423]
Movahedi K, Guilliams M, Van den Bossche J, Van den Bergh R, Gysemans C, Beschin A, et al.Identification of discrete tumor-induced myeloid-derived suppressor cell subpopulations withdistinct T cell-suppressive activity. Blood 2008;111:4233–4244. [PubMed: 18272812]
Nestle FO, Alijagic S, Gilliet M, Sun Y, Grabbe S, Dummer R, et al. Vaccination of melanoma patientswith peptide- or tumor lysate-pulsed dendritic cells. Nat Med 1998;4:328–332. [PubMed: 9500607]
Nestle FO, Banchereau J, Hart D. Dendritic cells: On the move from bench to bedside. Nat Med2001;7:761–765. [PubMed: 11433329]
Nickoloff BJ. Cracking the cytokine code in psoriasis. Nat Med 2007;13:242–244. [PubMed: 17342112]O'Donnell RK, Mick R, Feldman M, Hino S, Wang Y, Brose MS, et al. Distribution of dendritic cell
subtypes in primary oral squamous cell carcinoma is inconsistent with a functional response. CancerLett 2007;255:145–152. [PubMed: 17574329]
Bluth et al. Page 12
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Ohm JE, Carbone DP. VEGF as a mediator of tumor-associated immunodeficiency. Immunol Res2001;23:263–272. [PubMed: 11444391]
Osada T, Chong G, Tansik R, Hong T, Spector N, Kumar R, et al. The effect of anti-VEGF therapy onimmature myeloid cell and dendritic cells in cancer patients. Cancer Immunol Immunother. 2008
Pospisilova D, Borovickova J, Polouckova A, Spisek R, Sediva A, Hrusak O, et al. Generation offunctional dendritic cells for potential use in the treatment of acute lymphoblastic leukemia. CancerImmunol Immunother 2002;51:72–78. [PubMed: 11904731]
Serbina NV, Salazar-Mather TP, Biron CA, Kuziel WA, Pamer EG. TNF/iNOS-producing dendritic cellsmediate innate immune defense against bacterial infection. Immunity 2003;19:59–70. [PubMed:12871639]
Siegal FP, Kadowaki N, Shodell M, Fitzgerald-Bocarsly PA, Shah K, Ho S, et al. The nature of theprincipal type 1 interferon-producing cells in human blood. Science 1999;284:1835–1837. [PubMed:10364556]
Stary G, Bangert C, Tauber M, Strohal R, Kopp T, Stingl G. Tumoricidal activity of TLR7/8-activatedinflammatory dendritic cells. J Exp Med 2007;204:1441–1451. [PubMed: 17535975]
Steinbrink K, Jonuleit H, Muller G, Schuler G, Knop J, Enk AH. Interleukin-10-treated human dendriticcells induce a melanoma-antigen-specific anergy in CD8(+) T cells resulting in a failure to lyse tumorcells. Blood 1999;93:1634–1642. [PubMed: 10029592]
Steinman RM. Linking innate to adaptive immunity through dendritic cells. Novartis Found Symp2006;279:101–109. discussion 109-113, 216-109. [PubMed: 17278389]
Subramanian A, Kuehn H, Gould J, Tamayo P, Mesirov JP. GSEA-P: a desktop application for Gene SetEnrichment Analysis. Bioinformatics 2007;23:3251–3253. [PubMed: 17644558]
Talmadge JE. Pathways mediating the expansion and immunosuppressive activity of myeloid-derivedsuppressor cells and their relevance to cancer therapy. Clin Cancer Res 2007;13:5243–5248.[PubMed: 17875751]
Talmadge JE, Donkor M, Scholar E. Inflammatory cell infiltration of tumors: Jekyll or Hyde. CancerMetastasis Rev 2007;26:373–400. [PubMed: 17717638]
Thomas CJ, Wood GC, Marks VJ. Mohs micrographic surgery in the treatment of rare aggressivecutaneous tumors: the Geisinger experience. Dermatol Surg 2007;33:333–339. [PubMed: 17338692]
van der Veen RC, Dietlin TA, Dixon Gray J, Gilmore W. Macrophage-derived nitric oxide inhibits theproliferation of activated T helper cells and is induced during antigenic stimulation of resting T cells.Cell Immunol 2000;199:43–49. [PubMed: 10675274]
Vanacker A, Fabre G, Van Dorpe J, Peetermans WE, Maes B. Aggressive Cutaneous Squamous CellCarcinoma Associated with Prolonged Voriconazole Therapy in a Renal Transplant Patient. Am JTransplant. 2008
Wang Q, Liu Y, Wang J, Ding G, Zhang W, Chen G, et al. Induction of allospecific tolerance by immaturedendritic cells genetically modified to express soluble TNF receptor. J Immunol 2006;177:2175–2185. [PubMed: 16887977]
Weinberg AS, Ogle CA, Shim EK. Metastatic cutaneous squamous cell carcinoma: an update. DermatolSurg 2007;33:885–899. [PubMed: 17661931]
Wertel I, Barczynski B, Kotarski J. The role of dendritic cells in cytotoxic immune response regulationin ovarian cancer micro-environment. Front Biosci 2008a;13:2177–2190. [PubMed: 17981701]
Wertel I, Polak G, Bednarek W, Barczynski B, Rolinski J, Kotarski J. Dendritic cell subsets in theperitoneal fluid and peripheral blood of women suffering from ovarian cancer. Cytometry B ClinCytom 2008b;74:251–258. [PubMed: 18302193]
Young MR, Lathers DM. Myeloid progenitor cells mediate immune suppression in patients with headand neck cancers. Int J Immunopharmacol 1999;21:241–252. [PubMed: 10408632]
Zaba LC, Cardinale I, Gilleaudeau P, Sullivan-Whalen M, Suarez Farinas M, Fuentes-Duculan J, et al.Amelioration of epidermal hyperplasia by TNF inhibition is associated with reduced Th17 responses.J Exp Med 2007a;204:3183–3194. [PubMed: 18039949]
Zaba LC, Fuentes-Duculan J, Eungdamrong NJ, Abello MV, Novitskaya I, Pierson KC, et al. PsoriasisIs Characterized by Accumulation of Immunostimulatory and Th1/Th17 Cell-Polarizing MyeloidDendritic Cells. J Invest Dermatol. 2008a
Bluth et al. Page 13
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Zaba LC, Fuentes-Duculan J, Steinman RM, Krueger JG, Lowes MA. Normal human dermis containsdistinct populations of CD11c+BDCA-1+ dendritic cells and CD163+FXIIIA+ macrophages. J ClinInvest 2007b;117:2517–2525. [PubMed: 17786242]
Zaba LC, Krueger JG, Lowes MA. Resident and “Inflammatory” Dendritic Cells in Human Skin. J InvestDermatol. 2008b
Zea AH, Rodriguez PC, Atkins MB, Hernandez C, Signoretti S, Zabaleta J, et al. Arginase-producingmyeloid suppressor cells in renal cell carcinoma patients: a mechanism of tumor evasion. Cancer Res2005;65:3044–3048. [PubMed: 15833831]
Zou W, Machelon V, Coulomb-L'Hermin A, Borvak J, Nome F, Isaeva T, et al. Stromal-derived factor-1in human tumors recruits and alters the function of plasmacytoid precursor dendritic cells. Nat Med2001;7:1339–1346. [PubMed: 11726975]
Bluth et al. Page 14
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. Human cutaneous SCC is associated with intratumoral CD1a+/Langerin+ Langerhanscells, juxtatumoral CD11c+ myeloid DCs, and BDCA2+ plasmacytoid DCsRepresentative immunohistochemstry (left) and summary data with median cell count valuesof (A) CD1a, (B) Langerin, (C) CD11c, (D) BDCA-1, and (E) BDCA-2 cells in normal skin,SCC, and juxtatumoral skin. Asterisks (*) indicate significance where *=p<0.05, **=p<0.01and ***=p<0.005. Scale bar = 100μm.
Bluth et al. Page 15
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2. TNF-α and inducible nitric oxide synthase (iNOS)-producing dendritic cells (TIP-DCs)are found within the SCC microenvironmentRepresentative triple-label immunoflouresence reveals the presence of CD11c+ myeloid DCs(blue) that also express TNF-α (green) and iNOS (red). Triple positive cells appear white (insethigher 4-fold higher magnification). Monochromatic images in upper panel, single pseudocolorcontrols in middle panel, merged image below. Scale bar 100μm.
Bluth et al. Page 16
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. Phenotypically mature myeloid DCs are present in SCC, peritumoral and normal skinCD11c+/HLA-DRhi cells from normal skin, PTNL and SCC express maturation markers DC-LAMP, DC-SIGN, DEC-205, and co-stimulatory markers CD83, CD86 and CD80. There wasslightly increased DEC-205 and CD86 in myeloid DCs from SCC. Blue lines represent isotypecontrol; mean fluorescence intensity (MFI) value is shown in the upper right hand corner ofeach histogram (n=3).
Bluth et al. Page 17
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
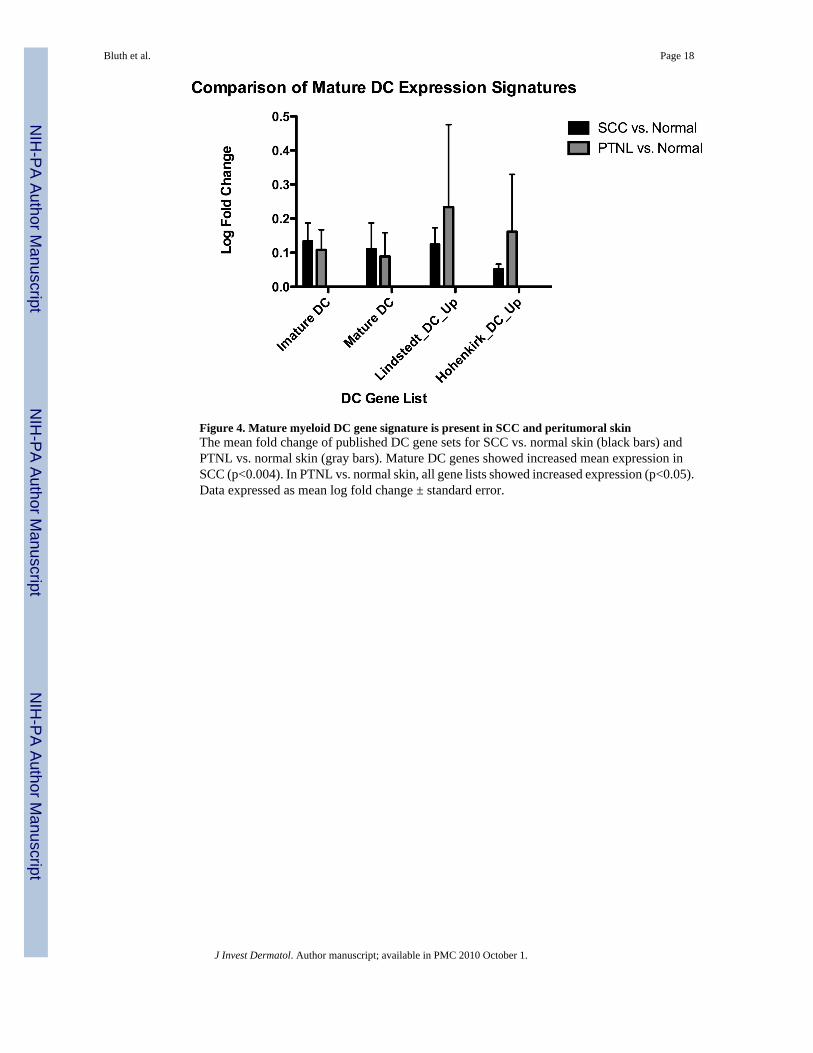
Figure 4. Mature myeloid DC gene signature is present in SCC and peritumoral skinThe mean fold change of published DC gene sets for SCC vs. normal skin (black bars) andPTNL vs. normal skin (gray bars). Mature DC genes showed increased mean expression inSCC (p<0.004). In PTNL vs. normal skin, all gene lists showed increased expression (p<0.05).Data expressed as mean log fold change ± standard error.
Bluth et al. Page 18
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
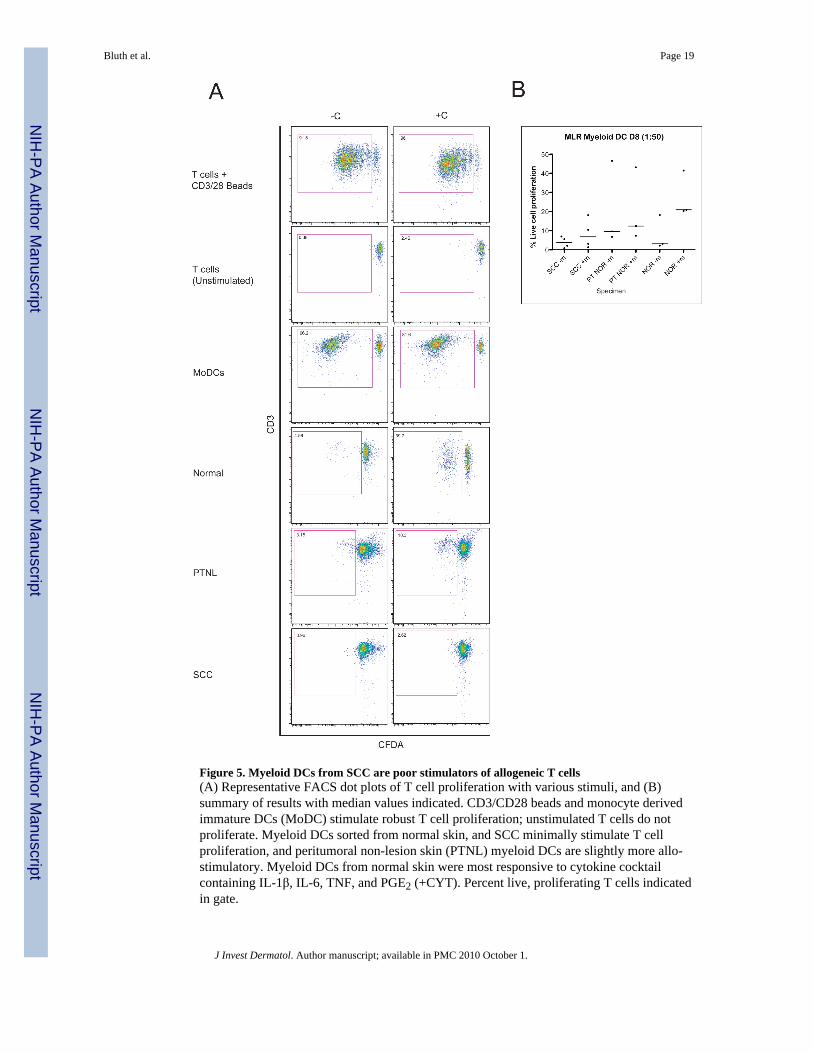
Figure 5. Myeloid DCs from SCC are poor stimulators of allogeneic T cells(A) Representative FACS dot plots of T cell proliferation with various stimuli, and (B)summary of results with median values indicated. CD3/CD28 beads and monocyte derivedimmature DCs (MoDC) stimulate robust T cell proliferation; unstimulated T cells do notproliferate. Myeloid DCs sorted from normal skin, and SCC minimally stimulate T cellproliferation, and peritumoral non-lesion skin (PTNL) myeloid DCs are slightly more allo-stimulatory. Myeloid DCs from normal skin were most responsive to cytokine cocktailcontaining IL-1β, IL-6, TNF, and PGE2 (+CYT). Percent live, proliferating T cells indicatedin gate.
Bluth et al. Page 19
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
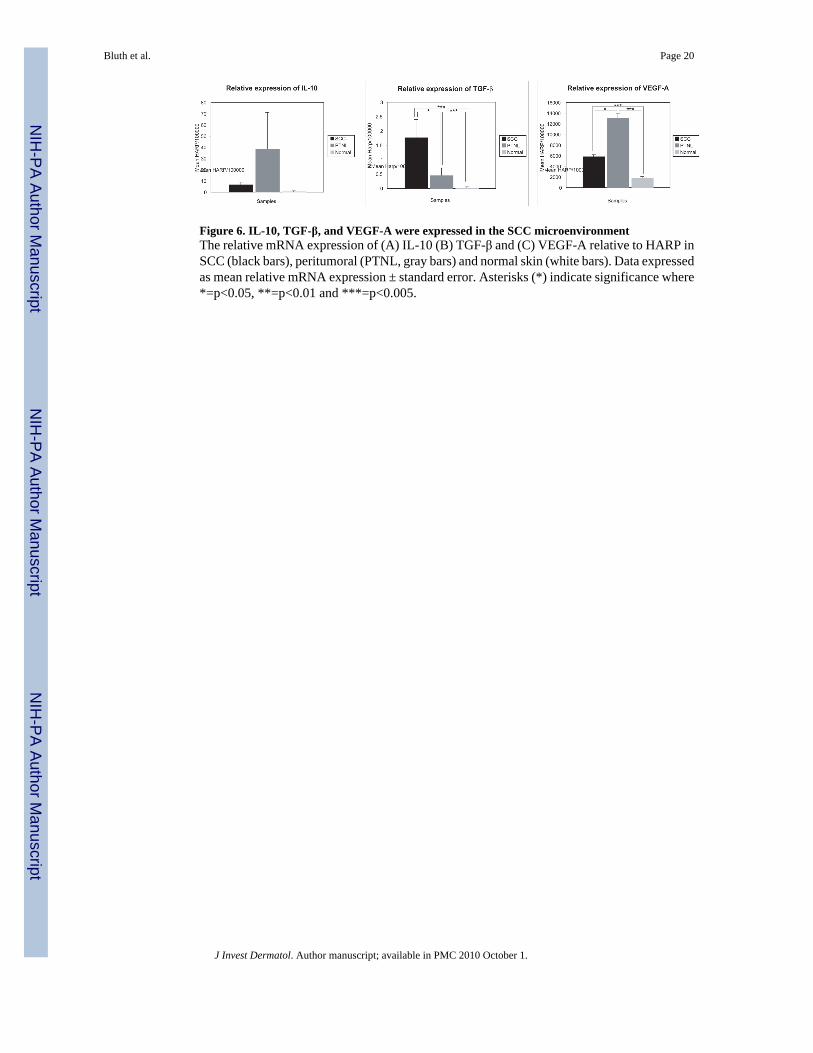
Figure 6. IL-10, TGF-β, and VEGF-A were expressed in the SCC microenvironmentThe relative mRNA expression of (A) IL-10 (B) TGF-β and (C) VEGF-A relative to HARP inSCC (black bars), peritumoral (PTNL, gray bars) and normal skin (white bars). Data expressedas mean relative mRNA expression ± standard error. Asterisks (*) indicate significance where*=p<0.05, **=p<0.01 and ***=p<0.005.
Bluth et al. Page 20
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Bluth et al. Page 21
Table 1Gene Set Enrichment Analysis (GSEA) of DC gene expression in SCC and PTNL skincompared to normal skin
DC gene expression was compared amongst published DC gene lists (Haider et al., 2007)-((Le Naour et al.,2001). DC gene expression was significantly enriched in PTNL skin compared to normal skin(Hohenkirk_DC_Up, p=0.024), while in SCC, enrichment of mature DC genes is increased (NES =1.25) but didnot reach statistical significance (p=0.12).
DC gene lists PTNL VS Normal SCC VS Normal
NES p NES p
Hohenkirk_DC_Up 1.61 0.024 0.94 0.510
Lindstedt_DC_Up 1.54 0.059 0.99 0.464
matureDC 1.34 0.083 1.25 0.122
immatureDC 0.98 0.500 0.82 0.780
NES is normalized enrichment score, p is p value
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Bluth et al. Page 22
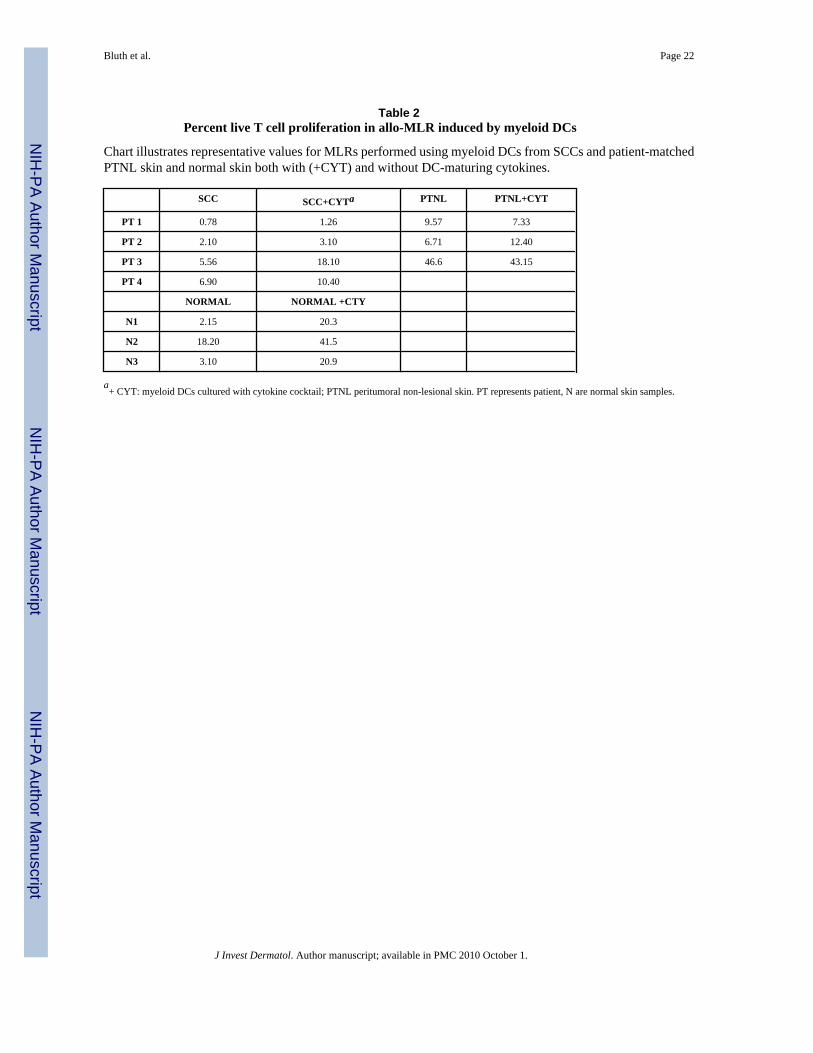
Table 2Percent live T cell proliferation in allo-MLR induced by myeloid DCs
Chart illustrates representative values for MLRs performed using myeloid DCs from SCCs and patient-matchedPTNL skin and normal skin both with (+CYT) and without DC-maturing cytokines.
SCC SCC+CYTa PTNL PTNL+CYT
PT 1 0.78 1.26 9.57 7.33
PT 2 2.10 3.10 6.71 12.40
PT 3 5.56 18.10 46.6 43.15
PT 4 6.90 10.40
NORMAL NORMAL +CTY
N1 2.15 20.3
N2 18.20 41.5
N3 3.10 20.9
a+ CYT: myeloid DCs cultured with cytokine cocktail; PTNL peritumoral non-lesional skin. PT represents patient, N are normal skin samples.
J Invest Dermatol. Author manuscript; available in PMC 2010 October 1.




















