MUSCLE AND ITS NEURAL CONTROL
12
AM. ZOOLOGIST, 2:55-05 (1062). MUSCLE AND ITS NEURAL CONTROL WILLIAM G. VAN DER KI.OOT Department of Physiology and Biophysics, New York University School of Medicine Motion is one of the most imposing of the abilities of living things. Tissues spe- cialized for moving are natural favorites of physiologists, who have studied muscle by following two separate and largely inde- pendent paths. One group studies the chemical events underlying contraction and supplying the energy for contraction. The second group asks how nerves set muscle into contraction and otherwise regulate its activity. The actions of nerves on muscle have helped neurophysiology by serving as accessible models of the behavior of junc- tions between excitable cells. (For a com- parative review see Hoyle, 1957a). VERTKHRATK MUSCLES Frog twitch muscles To begin, consider the system at mechani- cal rest, taking an ordinary frog muscle like the sartorius as a convenient starting point (for a general review, see Fatt, 1959). A mi- croelectrode is slipped through the cell membrane in the immediate neighborhood of a junction between a branch of motor nerve and a muscle fiber. The penetration of the microelectrode is marked by the ab- rupt appearance of the steady resting po- tential—about 90 mV, with the inside of the fiber negative. A closer look, however, shows that all is not quiet (Fatt and Katz, 1952). Every now and again there is a tiny depolarization of the membrane, of 0.5 mV at most (Fig. 1A). These miniature end- plate potentials come along in a perfectly random time sequence, appearing on the average at a rate of 0.1 to 100 per second, depending on the muscle chosen for study. The miniatures are found only in the re- gion of the endplate, and disappear after degeneration of the motor nerve. The am- plitude of the miniatures fits a normal dis- The original investigations were supported by grants from the Institute of Neurological Diseases and Blindness, Public Health Service. tribution curve with a standard deviation of about 30 per cent of the mean. All of the evidence shows that the miniatures re- sult from the spontaneous release of a packet or quantum of transmitter from the nerve endings. The situation can be thought of by pic- turing the nerve terminals as a tree, spread over a patch of ground representing the endplate. The tree is loaded with fruits, each a packet of acetylcholine. The fruits are not of identical size, but spread in a normal distribution about a mean. Once in a while a fruit drops from one or another branch of the tree and hits the ground, causing a miniature potential. The differ- ent branches of the tree, dropping fruit at random, are a useful part of the analog)', because the nerve ending is branched over the endplate, and it can be shown by lo- calized extracellular recording that the packets of transmitter are released inde- pendently by each branch (del Castillo and Katz, 1956). A single branch releases trans- mitter at a slow rate, and the miniatures re- corded inside the cell are a sum of the ac- tivity of all parts of the nerve terminal. The quanta of transmitter are released more often when the nerve terminals are depolarized, either by an electrical current or by an excess of K + . When the motor nerve is stimulated, the wave of depolariza- tion travels into the fine nerve terminals overlying the endplate. As a result, over 200 packets of acetylcholine are released within 2 msec, (del Castillo and Katz, 1954a). It is just as if a great gust of wind shook the tree, causing many of the branches almost simultaneously to drop a fruit. When this burst of acetylcholine hits the endplate membrane of the muscle fiber, the miniature potentials add together and produce a large endplate potential. The endplate potential itself is seen in isolation only by using special experimental circum- (55) Downloaded from https://academic.oup.com/icb/article/2/1/55/2110825 by guest on 11 January 2022
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of MUSCLE AND ITS NEURAL CONTROL

AM. ZOOLOGIST, 2:55-05 (1062).
MUSCLE AND ITS NEURAL CONTROL
WILLIAM G. VAN DER KI.OOT
Department of Physiology and Biophysics,New York University School of Medicine
Motion is one of the most imposing ofthe abilities of living things. Tissues spe-cialized for moving are natural favorites ofphysiologists, who have studied muscle byfollowing two separate and largely inde-pendent paths. One group studies thechemical events underlying contraction andsupplying the energy for contraction. Thesecond group asks how nerves set muscleinto contraction and otherwise regulate itsactivity. The actions of nerves on musclehave helped neurophysiology by serving asaccessible models of the behavior of junc-tions between excitable cells. (For a com-parative review see Hoyle, 1957a).
VERTKHRATK MUSCLES
Frog twitch muscles
To begin, consider the system at mechani-cal rest, taking an ordinary frog muscle likethe sartorius as a convenient starting point(for a general review, see Fatt, 1959). A mi-croelectrode is slipped through the cellmembrane in the immediate neighborhoodof a junction between a branch of motornerve and a muscle fiber. The penetrationof the microelectrode is marked by the ab-rupt appearance of the steady resting po-tential—about 90 mV, with the inside of thefiber negative. A closer look, however,shows that all is not quiet (Fatt and Katz,1952). Every now and again there is a tinydepolarization of the membrane, of 0.5 mVat most (Fig. 1A). These miniature end-plate potentials come along in a perfectlyrandom time sequence, appearing on theaverage at a rate of 0.1 to 100 per second,depending on the muscle chosen for study.The miniatures are found only in the re-gion of the endplate, and disappear afterdegeneration of the motor nerve. The am-plitude of the miniatures fits a normal dis-
The original investigations were supported bygrants from the Institute of Neurological Diseasesand Blindness, Public Health Service.
tribution curve with a standard deviationof about 30 per cent of the mean. All ofthe evidence shows that the miniatures re-sult from the spontaneous release of apacket or quantum of transmitter from thenerve endings.
The situation can be thought of by pic-turing the nerve terminals as a tree, spreadover a patch of ground representing theendplate. The tree is loaded with fruits,each a packet of acetylcholine. The fruitsare not of identical size, but spread in anormal distribution about a mean. Oncein a while a fruit drops from one or anotherbranch of the tree and hits the ground,causing a miniature potential. The differ-ent branches of the tree, dropping fruit atrandom, are a useful part of the analog)',because the nerve ending is branched overthe endplate, and it can be shown by lo-calized extracellular recording that thepackets of transmitter are released inde-pendently by each branch (del Castillo andKatz, 1956). A single branch releases trans-mitter at a slow rate, and the miniatures re-corded inside the cell are a sum of the ac-tivity of all parts of the nerve terminal.
The quanta of transmitter are releasedmore often when the nerve terminals aredepolarized, either by an electrical currentor by an excess of K+. When the motornerve is stimulated, the wave of depolariza-tion travels into the fine nerve terminalsoverlying the endplate. As a result, over200 packets of acetylcholine are releasedwithin 2 msec, (del Castillo and Katz,1954a). It is just as if a great gust of windshook the tree, causing many of thebranches almost simultaneously to drop afruit. When this burst of acetylcholine hitsthe endplate membrane of the muscle fiber,the miniature potentials add together andproduce a large endplate potential. Theendplate potential itself is seen in isolationonly by using special experimental circum-
(55)
Dow
nloaded from https://academ
ic.oup.com/icb/article/2/1/55/2110825 by guest on 11 January 2022

56 WILLIAM G. VAN DER KLOOT
FIG. I-A. Miniature endplate potentials in the sar-torius muscle of the frog. The upper line in eachpair is an intra-cellular recording, the lower line isan extra-cellular recording. The extra-cellular elec-trode samples electrical activity over only 50^ of themembrane; intra-cellular records show activity overmore than a lOOOju of membrane. The different re-sults with the two methods of recording show thateach miniature is produced by the release of trans-mitter by small part of the nerve terminal (fromdel Castillo and Katz, 1956). 1-B. An endplate po-tential in the sartorius muscle of the frog. The am-plitude of the potential was lowered by treating themuscle with d-tubocurarine, so the threshold forthe production of an action potential is not reached(from Fatt and Katz, 1951). 1-C. The response tonerve (N) and to direct stimulation (M) at the end-plate of a muscle fiber. In N notice the initial slantin the depolarization; this is the endplate potentialwhich then triggers the spike. Also notice the de-creased peak and the 'hump' which are caused bythe attenuation of the spike by the 'short-circuit' atthe endplate (from Fatt and Katz, 1951 J. Physiol.)
stances (Fig. IB). Usually the endplate po-tential acts as an electrical stimulus andsets up a conducted action potential in theadjacent muscle membrane. The spike ofthe action potential arises from the ascend-ing portion of the endplate potential, andthereby obscures its normal course (Fig1C). The action potential sweeps over thefiber, triggering the all or nothing contrac-tion. The contraction of the whole muscleis graded according to how many of thefibers are activated.
Obviously, the endplate potential is quitedifferent from an action potential and mustbe generated by a different mechanism. Theendplate potential, if left to itself, wouldreach a maximum depolarization of -15mV. In an action potential, of course, thevoltage across the membrane actually re-verses in direction. Now -15 mV is just thepotential which would exist if the mem-brane of the endplate were suddenly abol-ished and the solutions inside and outsideof the fiber were allowed to begin to mix.The action of acetylcholine is to make themembrane at the endplate highly permeableto all sorts of ions. Technically, there is asudden increase in the conductance of themembrane. The action potential is gener-ated by a selective increase in Na+ conduct-ance; the endplate potential arises by ageneral increase in the conductance of allof the common ions.
Since the endplate membrane stimulatedwith acetylcholine becomes permeable tomany ions, an action potential across thenearby muscle membrane will be distortedand diminished by currents which flowthrough the "short circuit" at the endplate.This accounts for the difference betweenthe signal recorded at the endplate when itis activated by acetylcholine and when it isstimulated directly by a current (Fig. 1C;Fatt and Katz, 1951). Now suppose that themuscle fiber is excited far away from anendplate. The spike travels toward the end-plate and at the same time the motor nervecan be stimulated. The results show thatwhen the spike is in the endplate region itwill always be distorted by the increasedconductance at an activated endplate. Thismeans that the endplate is never refractory
Dow
nloaded from https://academ
ic.oup.com/icb/article/2/1/55/2110825 by guest on 11 January 2022

MUSCLE AND ITS NEURAL CONTROL 57
10 mV
FIG. 2. A junctional potential recorded in a tonicmuscle fiber of the frog. The time marks are 20msec, apart. (From KufHer and Vaughan Williams,1953 J. Physiol.)
to acetylcholine stimulation, and this sug-gests that the spike does not invade the end-plate itself. Therefore, the endplate standsin sharp contrast to the surrounding musclemembrane: they are different structureswith quite different properties.
When the motor nerve is stimulatedtwice in close succession, the second depo-larization of the nerve terminals results inthe release of more quanta of transmitter,and thereby leads to a larger endplate po-tential (del Castillo and Katz, 1954b).With continued repetitive stimulation, theendplate potentials increase in size until ahigh steady level is reached. This facilita-tion is of little physiological significance inordinary frog twitch muscles, because theusual endplate potential is quite adequateto trigger a conducted action potential. Inother muscles, facilitation becomes of greatimportance.
Tonic or slow fibers
Many muscles of the frog contain a fewslow or tonic fibers. The tonic fibers aredistinguished by having an endplate everylew millimeters; each endplate is inner-vated by a branch from the motor axon,and the innervation pattern is thereforecalled multi-terminal. The tonic fibers arealso marked by a complete inability to con-duct an action potential. Contraction istriggered solely by the small endplate po-tentials (Fig. 2). When the motor axon isstimulated, the entire fiber is slightly de-
polarized because the endplates are so closeto one another. With repetitive nerve stimu-lation, the endplate potentials add togetherto produce a pronounced depolarization ac-companied by a sustained contracture ofthe tonic fibers (Kuffler and Vaughan Wil-liams, 1953). The response of the individ-ual tonic fiber is graded by the frequencyof stimulation. •
Birds
Most bird muscles seem to be made up oftwitch fibers. But some of the fibers have amulti-terminal innervation, like frog tonicfibers, even though they conduct normal ac-tion potentials (Ginsborg, 1960).
Man
Recently, intercostal muscles removedduring thoracic surgery were studied withmicroelectrodes. These muscle fibers arcquite like the frog twitch fibers (Elmquist,Johns, and Thesleff, 1960). Muscles frompatients suffering from niyasthenia gravisshow few miniature potentials, and the end-plate potentials fluctuate widely becauseless than the normal number of quanta oftransmitter are released when the motornerve is stimulated. The symptoms of my-asthenia occur because either the synthesisor the release of the transmitter is depressed(Dahlbach, Elmquist, Johns. Radncr, andThesleff. 1961).
Smooth muscles
Burnstock and Holman (1961) succeededin recording from a nerve and smoothmuscle cell preparation, using the isolatedvas deferens of the guinea pig and stimulat-ing the hypogastric nerve. When there isno stimulation, there is a miniature junc-tional potential once or twice every 2 sec-onds. (The term junctional potential isused when there does not seem to be a dis-tinct morphological endplate.) Every cellshows miniatures, suggesting that every cellreceives a motor nerve. Nerve stimulationproduces a junctional potential rangingfrom 1 to 20 mV, depending on the numberof postganglionic nerve libers stimulated.The variation in the height of the June-
Dow
nloaded from https://academ
ic.oup.com/icb/article/2/1/55/2110825 by guest on 11 January 2022
58 WILLIAM G. VAN DER KLOOT
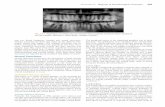
FIG. 3. Jiinctional potentials leading to a spike re-corded in a smooth muscle cell of the guinea pigvas deferens. Calibrations, 100 msec, and 50 mV(From Rurnstock and Holnian, 1961 |. I'hysiol.)
tional potential shows that a number ofdifferent axons converge on each smoothmuscle cell, so there is a poly-neuronal in-nervation. If a series of junctional poten-tials succeeds in depolarizing a cell to -37mV, a spike is elicited (Fig. 3). There is amarked facilitation of the junctional poten-tials with repetitive stimulation. This junc-tion, in short, is built on the same funda-mental plan as the striated muscle endplate.However, spontaneous electrical and me-chanical activity is a prominent feature ofmany smooth muscles.
ARTHROPODS
Crustaceans
Aside from the frog, custacean muscle isthe best understood (see Furshpan, 1959;Wiersma, 1960). All of the crustaceanmuscles have multi-terminal innervation.But the situation is more complex than thatin frog tonic muscles, because often eachliber is innervated by a number of axonsand each axon produces a distinct effect.Some muscle fibers in the main flexor of thecarpopodite of decapods receive branchesfrom 4 motor neurons (Van Harreveld andWiersma, 1939). The four motor neuronscomprise the total motor innervation forthe entire muscle. The contraction of themuscle must be primarily regulated by howmuch the individual fibers contract.
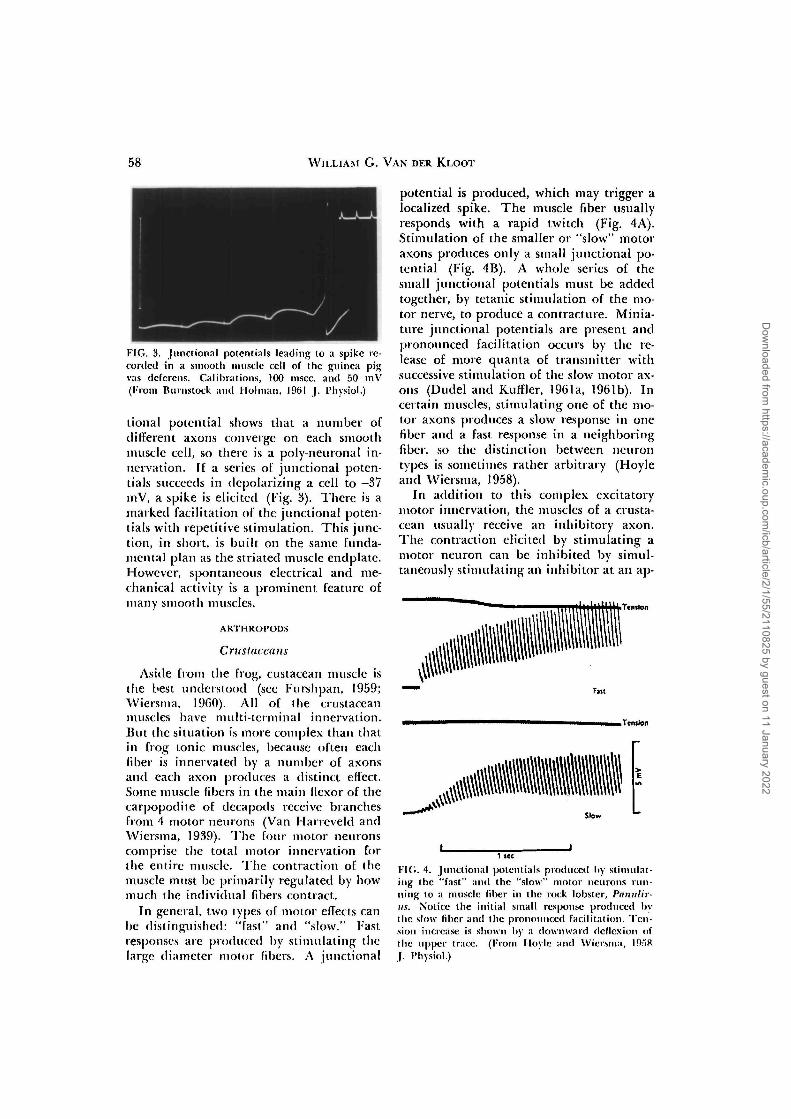
In general, two types of motor effects canbe distinguished: "fast" and "slow." Fastresponses are produced by stimulating thelarge diameter motor fibers. A junctional
potential is produced, which may trigger alocalized spike. The muscle fiber usuallyresponds with a rapid twitch (Fig. 4A).Stimulation of the smaller or "slow" motoraxons produces only a small junctional po-tential (Fig. 4B). A whole series of thesmall junctional potentials must be addedtogether, by tetanic stimulation of the mo-tor nerve, to produce a contracting. Minia-ture junctional potentials are present andpronounced facilitation occurs by the re-lease of more quanta of transmitter withsuccessive stimulation of the slow motor ax-ons (Dudel and Kuffler, 1961a, 1961b). Incertain muscles, stimulating one of the mo-tor axons produces a slow response in onefiber and a fast response in a neighboringfiber, so the distinction between neurontypes is sometimes rather arbitrary (Hoyleand Wiersma, 1958).
In addition to this complex excitatorymotor innervation, the muscles of a crusta-cean usually receive an inhibitory axon.The contraction elicited by stimulating amotor neuron can be inhibited by simul-taneously stimulating an inhibitor at an ap-
iTenikMi
• Tension
Slow
1 KC
FIG. 4. Junclional potentials produced hy stimulat-ing the "fast" and the "slow" motor neurons run-ning to a muscle fiber in the rock lobster, Panulir-us. Notice the initial small response produced bythe slow fiber and the pronounced facilitation. Ten-sion increase is shown by a downward deflexion ofthe upper trace. (From Hoylc and Wiersma, l!).ri8J. I'hysiol.)
Dow
nloaded from https://academ
ic.oup.com/icb/article/2/1/55/2110825 by guest on 11 January 2022

MUSCLE AND ITS NEURAL CONTROL
FIG. 5. Junctional potentials produced by stimulat-ing the inhibitory neuron running a muscle fiberin the crayfish. The membrane potential is shiftedfrom 80 mV to 61 mV and back again by passingcurrent through a second micro-electrode insertedinto the cell. The arrows mark the reversal poten-tial for the inhibitory potential which is at 22 mV.(From Dudel and Kuffier, 1961c J. Physiol.)
propriate rate. It now seems that at leastthree different mechanisms are involved ininhibiting contraction. First, stimulationof the inhibitory neuron increases the Cl"permeability (or conductance) of the junc-tional membrane (Boistel and Fatt, 1958).The normal resting potential is close to theequilibrium potential for Cl". Therefore,an increase in Cl" conductance will be aconservative force, tending to hold themembrane potential close to the restingvalue and to oppose an excitatory junc-tional potential which would normally de-polarize the membrane. When the inhibi-tory neuron is stimulated, the electrical re-sponse depends upon the resting potentialof the muscle fiber. If the resting potentialis at the Cl" equilibrium value, there willbe no change. If the resting potential is be-low the Cl" equilibrium, the fiber will hy-perpolarize. If the resting potential is abovethe Cl" equilibrium, the fiber will depolar-ize (Fig. 5). Secondly, during inhibitorynerve stimulation there is a decrease in thenumber of quanta of transmitter releasedby the motor nerve terminals (Dudel andKuffier, 1961c). This is an example of apre-junctional inhibition. And finally, insome crabs, inhibitory nerve stimulation de-creases contraction without seeming to lowerthe excitatory junctional potentials (Floreyand Hoyle, 1961). In this case mechanicalinhibition may be produced by a direct ef-fect on the contractile mechanism.
Insects
Many insect muscles are innervated byboth a fast and a slow motor neuron, quite
like the crustacean plan. Certain musclesare also innervated by a third neuron.Stimulation of the third neuron produces aslight hyperpolarization of the cell, but atthe same time potentiates contractionselicited by motor nerve stimulation. Hoyle(1957b) suggests that the third axon is anevolutionary relic of the inhibitory systemwhich was preserved in the crustaceans.
The wings of the higher Diptera maybeat more than 220 times per second, fourtimes as fast as the wings of a humming-bird. The tymbal muscles (the sound pro-ducers) of cicadids work at frequencies of390 per second. These high frequencieswere hard to understand, because even fast-moving mammalian muscles enter a smoothtetanus at much lower rates of stimulation.The mystery was compounded when it wasfound that there are many more movementsof the muscles than there are action poten-tials. In Culliphora flight muscle, there arethree action potentials per second when thewings are beating at 120 per second (Prin-gle, 1949; Roeder, 1951). In tymbal muscle,each stimulus of the motor nerve may elicitnine contractions by the muscle (Pringle,1954). If, however, the muscles are cut freefrom their cuticular attachments and fas-tened to a lever, each stimulus gives onlyone contraction. In the insect, the mechani-cal set-up of the attachments for thesemuscles is in the form of a "click" mecha-nism. When the muscle contracts and in-creases its tension, a critical point is reached.The cuticular attachment then clicks in-ward to a second position. The abrupt mo-tion is like the spring on the "cricket"which a lecturer uses to signal the projec-tionist. The sudden shortening of themuscle when the click mechanism gives wayresults in a "deactivation" of the contractilesystem and the tension exerted by themuscle quickly falls. The attachment thenclicks outward, back to its original position.The muscle begins to pull once again sothat the attachment clicks inward. This se-quence can be repeated time and again dur-ing a single muscle twitch. By taking ad-vantage of these specialized mechanical fea-tures of the muscles, and by evolving clicksystems, these animals produce a whole sc-
Dow
nloaded from https://academ
ic.oup.com/icb/article/2/1/55/2110825 by guest on 11 January 2022

60 WILLIAM G. VAN DER KLOOT
ries of movements during one contractioncycle by the muscle (Pringle, 1957; Boet-tiger, 1957).
Xiphosurans
Limulus muscle fibers are innervated byfast and slow axons; there is no evidencefor an inhibitory innervation (Hoyle, 1958).
OTHER INVERTEBRATE MUSCLES
The other invertebrate phyla usuallyhave uninucleate, non-striated muscle cells.These non-striated cells look much like ver-tebrate smooth muscle, but the comparisoncan be physiologically misleading, becausesome of the invertebrate non-striated cellsare capable of quite rapid contractions (seeProsser, 1960). A fast and a slow innerva-tion are commonly found; the studiedpreparation include cephalopods (Wilson,1960a), polycheates (Wilson, 1960b), ho-lothurians (Prosser, Curtis, and Travis,1951; Pople and Ewer, 1955), and scypho-zoans (Horridge, 1956). In the polycheatesthe fast system fatigues rapidly, so it mayonly be used in startle responses. Indirectevidence suggests that single cells are inner-vated by both fast and slow systems, butthere is direct proof by intracellular record-ing only for the proboscis retractors ofPhascolosoma (Prosser and Sperelakis,quoted by Prosser, 1960).
Particularly interesting unstriated musclesare the foot retractors and the shell adduc-tors of the bivalves. These muscles are thechampion tension developers—about 25 kg/g muscle (the best mammalian muscle de-velops a tension of about 2.5 kg/g muscle).When stimulated with an alternating cur-rent, the molluscan muscles give a briefcontraction. But when stimulated with adirect current for a few seconds, or withacetylcholine, the muscles enter a prolongedcontracture. During the prolonged con-t radii re, the muscles expend little more en-ergy than they do at rest; they behave asthough "caught" in the contracted position.The muscle can be relaxed from the con-tracture by stimulating it with a series ofbrief electrical pulses or will) 5-lmlroxy-(ivplamine (Twaiog. Ht.54).
Although there is sonic disagreement in
the explanations offered for the prolongedcontracture, a plausible theory runs as fol-lows. When the muscle twitches, contrac-tion is produced by the usual actomyosinmechanism. When a muscle is stimulatedto enter a prolonged contracture, it is againcontracted by the actomyosin. But oncecontracted, a second protein, paramyosin,crystallizes within the fibers. Paramyosin isfound only in muscles with the "catch"mechanism and it crystallizes followingsmall shifts in the pH. The crystallizedparamyosin greatly prolongs the relaxationof the contracted muscle. Therefore, withinfrequent nerve activation, the musclemaintains a steady tension (Jewell, 1959;Johnson, Kahn, and Szent-Gyorgyi, 1959;Johnson and Twarog, 1960).
SPONTANEOUS ACTIVITY BY STRIATED MUSCLES
Some striated muscles can contract spon-taneously. The classic example is the verte-brate heart, which is driven by a slow, spon-taneous depolarization of the cell mem-brane in the sinus venosus, as shown in Fig.6A (del Castillo and Katz, 1955; Hutterand Trautwein, 1956). Another kind ofspontaneously active muscle is the tinycloser of the spiracular opening in the pupaof the cecropia silkworm. This muscle, iso-lated from the nervous system, also has amembrane which spontaneously depolar-izes thereby causing the muscle to contract(Fig. 6B: Van der Kloot, unpublished ob-servations). Both heart and spiracularmuscle are regulated in the whole animalby nerve impulses originating in the centralnervous system.
EXCITATION AND CONTRACTION
While much effort has been concentratedon the electrical aspects of muscle activa-tion, there is a great mystery about theevents going on between the generation ofthe junctional potential or spike and theonset of contraction. In this field, the com-parative approach promises particularly im-portant rewards. One serious problem ishow the excitation navels from the surfaceof the cell into (he interior. In the fastervertebrate muscles, at least, contractioncomes about too quickh to be explained
Dow
nloaded from https://academ
ic.oup.com/icb/article/2/1/55/2110825 by guest on 11 January 2022

MUSCLE AND ITS NEURAL CONTROL 61
I'IG. 6-A. Spontaneous depolarizations leading toan action potential in the sinus venosus of the frog.Calibrations 10 mV, 100 msec. 6-B. Action poten-tials from the spiracular closing muscle of the Ce-cropia silkworm. The first potential was elicited bystimulating the motor nerve, the second arises by aspontaneous depolarization of the cell membrane.The small spike does not wipe out the spontaneousgenerator potential. Calibrations 25 mV, 100 msec.
by the diffusion of some triggering sub-stance inward from the cell surface (Hill,1949). The answer to this problem maycome from the recent rediscovery of a sys-tem of channels—the sarcoplasmic reticu-lum—which runs inward from the surfaceof the muscle. In a frog twitch fiber, localelectrical stimulation is most effective at aZ line. A localized contraction is elicited ofthe halves of the I band on either side ofthe stimulated Z line. The contraction doesnot spread very far longitudinally down thefiber but it does spread deeply into the cell.This suggests that there is a specialized con-ducting channel running into the musclefiber starting at the Z line, spreading intothe fiber and laterally to the borders of theI band (Huxley and Taylor, 1958). And in-deed at the level of the Z line there is acharacteristic group or "triad" of the vesi-cles of the sarcoplasmic reticulum (Porterand Palade, 1957).
In crab muscle the spots sensitive tostimulation lie close to the boundaries be-tween the A and the I bands, the contrac-tion consists of a movement of the adjacentZ lines toward the A-I boundary. Presum-ably this different result corresponds to adifferent arrangement of the sarcoplasmicreticulum.
Even if these channels into the cytoplasmare important in the activation of a muscle,we still have no idea what travels inwardby this specialized path. Perhaps the logicalway to further consider the question is atthe level of the glycerinated muscle fiber(see Needham, 1959, for a recent review).The glycerinated fiber, in an environmentof proper ionic strength, can be made tocontract by adding a nucleotide triphos-phate. The order of effectiveness for theproduction of tension and also for the rateof enzymatic hydrolysis to the nucleotidediphosphate is:ATP > CTP > UTP > ITP > GTP(Hasselbach, 1956). Mg++ is an essential co-factor.
To relax the glycerinated fiber once it iscontracted, ATP and Mg++ are again re-quired. In addition, the relaxing factor sys-tem, is needed (Marsh, 1952). This systemconsists of a microsomal fraction of muscleplus a dialyzable co-factor. Apparently themicrosomal fraction contains enzymeswhich, in the presence of the co-factor,manufacture a substance (or substances).The product of the relaxing factor systeminhibits the nucleotide triphosphate activ-ity of the actomyosin and therefore leadsto relaxation. The substance causing re-laxation appears to be labile and to bereadily destroyed by enzymes which arepresent even in the glycerinated fiber(Gergely, 1959). The nature of this sub-stance is a central problem for muscle physi-ology.
With this background, drawn from ahighly simplified system, it is possible toget some idea of what must be going on ina muscle. In the resting muscle there isplenty of ATP, and about 80 per cent ofthe magnesium is free (Hasselbach, 1957).In these conditions a glycerinated fiberwould contract—the muscle does not. Pre-
Dow
nloaded from https://academ
ic.oup.com/icb/article/2/1/55/2110825 by guest on 11 January 2022

62 WII.MAM G. VAN DER KLOOT
sumably this is because the relaxing factorsystem is operating and contraction is in-hibited. The question then is how the relax-ing factor system is momentarily slopped.As little as 0.2 HIM Ca++ is a powerful in-hibitor of the relaxing factor system in invitro tests (Bendall, 1953). Frog musclefiber water contains about 1.4 mM/kg ofCa++, but according to Hasselbach (1957)more than 90% is bound. Carnosine is alsoan effective inhibitor of the relaxing factorsystem at concentrations of 2 mM (Gergely,1959; Nagi and Uchida, I960). This is par-ticularly surprising because the concentra-tion of carnosine in the fiber water of afrog muscle is about 14.7 mM/kg (see Con-way, 1957). Other known inhibitors of therelaxing factor system, like DPN and caf-feine, can produce contracture in musclebut, of course, are not normally present inthe cell.
At the moment, calcium or carnosineseem to be of the greatest physiological in-terest. It is known that the microinjectioninto an intact muscle fiber of either calcium(Heilbrunn and Wiercinski, 1946; Neider-gerke, 1955) or of carnosine (Hayashi, 1956)produces a local contraction. Consequently,any increase in the inter-fiber concentra-tion of calcium or of carnosine would be ex-pected to cause contraction.
Contraction is often triggered by the de-polarization of the cell membrane. Oneidea is that the depolarization liberates acalcium compound from the cell mem-brane, and this then diffuses into the in-terior to elicit contraction. However, manyof the experiences of comparative musclephysiologists argue against a direct relation-ship between depolarization and contrac-tion. In the Crustacea, there are muscleswhere the stimulation of the fast axon pro-duces large potentials but no contraction,while stimulation of the slow axon at thesame frequency gives smaller potentials andcontraction (Wiersma and Van Harreveld,1938; Hoyle and Wiersma, 1958). Verte-brate smooth and skeletal muscles, com-pletely depolarized in a Ringer's in whichthe sodium is replaced with K*, continueto contract in response to acetylcholinestimulation (Singh and Acharya, 1957a; Ev-
ans, SchiId and Thesleff, 1958; Jenkinsonand Nicholls, 1900). Therefore, contractiondoes not always go hand-in-hand with de-polarization.
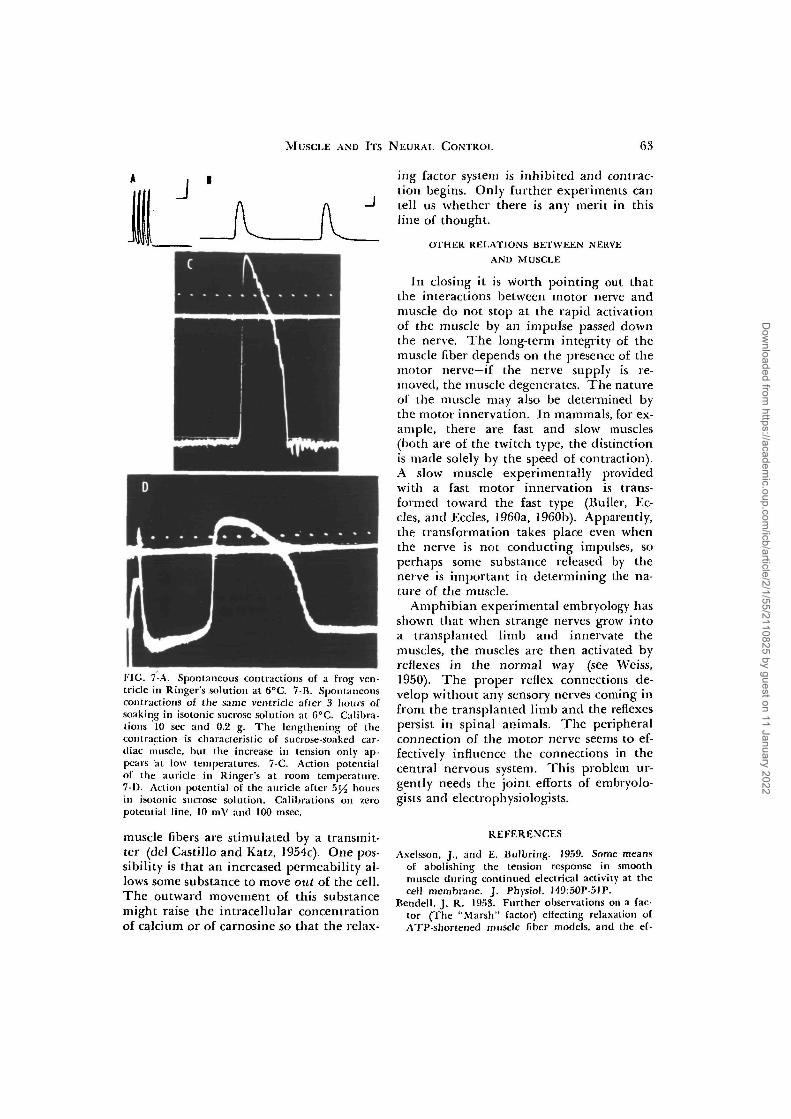
Another idea is that the change in thepermeability of the cell membrane duringexcitation allows an increased inflow of cal-cium. This idea is based on the facts thatthe withdrawal of calcium from the Ring-er's leads to the loss of contractility (Ring-er, 1880; Axelsson and Bulbring, 1959;Frank, 1960; Robertson, I960) and thatcontraction is accompanied by increaseduptake of Ca4"' by skeletal (Bianchi andShanes, 1959) and by heart muscles (Wine-grad, 1960). However, in isotonic sucrosesolutions, both smooth and cardiac musclescontinue to contract for hours (Singh, 1944;Singh et al., 1945) and to conduct action po-tentials (Fig. 7; Kolodny and Van derKloot, 1961; Van der Kloot and Rubin, un-published observations). Moreover, whenthe heart which has been kept in isotonicsucrose finally fails, it does not recoverwhen transferred to a Ringer's solution inwhich SO4" is the anion. The sucrose-soaked hearts do recover in a Cl~ Ringer's.The probable explanation of this experi-ment is that the electrical and mechanicalactivity of the heart fails because of the lossof ions from inside the cells. These ionscannot be replaced from a SO4~ Ringer'ssolution because the cell membrane is rela-tively impermeable to SO4~. In a Cl" Ring-er's solution, the intracellular cations arerapidly replaced. Since the concentrationof calcium in the sucrose solutions is verylow, coining only from impurities in the de-ionized, distilled water or from the reagentgrade sucrose, these experiments argueagainst the idea that the inflow of calciumfrom the external medium is the trigger forcontraction.
At present, no answer to the problem ofexcitation-contraction coupling seems to fitall of the facts. It may even be that alterna-tive mechanisms are operating under dif-ferent experimental conditions. My ownguess is that the change in the permeabilityof the cell membrane during excitation isan essential step. The permeability of themembrane increases even when depolarized
Dow
nloaded from https://academ
ic.oup.com/icb/article/2/1/55/2110825 by guest on 11 January 2022
MUSCLE AND ITS NEURAL CONTROL 63
FIG. 7-A. Spontaneous contractions of a frog ven-tricle in Ringer's solution at 6°C. 7-B. Spontaneouscontractions of the same ventricle after 3 hours ofsoaking in isotonic sucrose solution at 6°C. Calibra-tions 10 sec and 0.2 g. The lengthening of thecontraction is characteristic of sucrose-soaked car-diac muscle, but the increase in tension only ap-pears at low temperatures. 7-C. Action potentialof the auricle in Ringer's at room temperature.7-1). Action potential of the auricle after b]/2 hoursin isotonic sucrose solution. Calibrations on zeropotential line, 10 mV and 100 msec.
muscle fibers are stimulated by a transmit-ter (del Castillo and Katz, 1954c). One pos-sibility is that an increased permeability al-lows some substance to move out of the cell.The outward movement of this substancemight raise the intracellular concentrationof calcium or of carnosine so that the relax-
ing factor system is inhibited and contrac-tion begins. Only further experiments cantell us whether there is any merit in thisline of thought.
OTHER RELATIONS BETWEEN NERVEAND MUSCLE
In closing it is worth pointing out thatthe interactions between motor nerve andmuscle do not stop at the rapid activationof the muscle by an impulse passed downthe nerve. The long-term integrity of themuscle fiber depends on the presence of themotor nerve—if the nerve supply is re-moved, the muscle degenerates. The natureof the muscle may also be determined bythe motor innervation. In mammals, for ex-ample, there are fast and slow muscles(both are of the twitch type, the distinctionis made solely by the speed of contraction).A slow muscle experimentally providedwith a fast motor innervation is trans-formed toward the fast type (Buller, Ec-cles, and Eccles, 1960a, 1960b). Apparently,the transformation takes place even whenthe nerve is not conducting impulses, soperhaps some substance released by thenerve is important in determining the na-ture of the muscle.
Amphibian experimental embryology hasshown that when strange nerves grow intoa transplanted limb and innervate themuscles, the muscles are then activated byreflexes in the normal way (see Weiss,1950). The proper reflex connections de-velop without any sensory nerves coming infrom the transplanted limb and the reflexespersist in spinal animals. The peripheralconnection of the motor nerve seems to ef-fectively influence the connections in thecentral nervous system. This problem ur-gently needs the joint efforts of embryolo-gists and electrophysiologists.
REFERENCES
Axelsson, J., and E. Bulbring. 1959. Some meansof abolishing the tension response in smoothmuscle during continued electrical activity at thecell membrane. J. Physiol. 149:50P-51P.
Bendell, j . R. 1953. Further observations on a fac-tor (The "Marsh" factor) effecting relaxation ofATP-shortened muscle fiber models, and the ef-
Dow
nloaded from https://academ
ic.oup.com/icb/article/2/1/55/2110825 by guest on 11 January 2022

64 WILLIAM G. VAN DER KLOOT
feet of Ca and Mg ions upon it. J. Physiol. 121:232-254.
Bianchi, C. P., and A. M. Shanes. 1959. Calciuminflux in skeletal muscle at rest, during activity,and during potassium contracture. J. Gen. I'hysi-ol. 42:803-815.
Boetliger, E. G. 1957. The machinery of insectflight, p. 117-142. In B. T. Scheer (Ed.), Recentadvances in invertabrate physiology. Universityof Oregon Publications, Eugene, Oregon.
Boistel, J., and P. Fatt. 1958. Membrane permea-bility change during inhibitory transmitter actionin crustacean muscle. J. Physiol. 144:176-191.
Buller, A. J., J. C. Ecdes, and R. M. Eccles. 1960a.Differentiation of fast and slow muscles in the cathind limbs. J. Physiol. 150:399-416.
. 1960b. Interactions between motorneuronesand muscles in respect to the characteristic speedsof their responses. J. Physiol. 150:417-439.
Burnstock, G., and M. E. Holman. 1961. The trans-mission of excitation from autonomic nerve tosmooth muscle. J. Physiol. 155.115-133.
del Castillo, J., and B. Katz. 1954a. Statistical fac-tors involved in neuromuscular facilitation anddepression. J. Physiol. 124:574-585.
. 1954b. Quantal components of the endplatepotential. J. Physiol. 124:560-573.
. 1954c. The membrane change produced bythe neuromuscular transmitter. J. Physiol. 125:546-565.
. 1955. Production of membrane potentialchanges in the frog's heart by inhibitory nerveimpulse. Nature 175:1035.
. 1956. Localization of active spots withinthe neuromuscular junction of the frog. J. Physiol.
132:630-649.Comvay, E. J. 1957. Nature and significance of con-
centration relations of potassium and sodium ionsin skeletal muscle. Physiol. Rev. 37:84-132.
Dahlbach, O., D. Elmquist, J. R. Johns, S. Radner,and S. Thesleff. 1961. An electrophysiologic studyof the neuro-muscular junction in myastheniagravis. J. Physiol. 156:336-343.
Dudel, J., and S. W. Kuffler. 1961a. The quanta]nature of transmission and spontaneous minia-ture potentials at the crayfish neuro-muscularjunction. J. Physiol. 155:514-529.
. 1961b. Mechanism of facilitation at thecrayfish neuromuscular junction. J. Physiol. 155:530-542.
. 1961c. Presynaptic inhibition at the cray-fish neuromuscular junction. J. Physiol. 155:543-562.
Elmquist, D., T. R. Johns, and J. Thesleff. 1960. Astudy of some electrophysiological properties ofhuman intercostal muscle. J. Physiol. 154:602-607.
Evans, D. H. L., H. O. Schild, and S. Thesleff. 1958.Effects of drugs on depolarized plain muscle. J.Physiol. 143:474-485.
Fatt, P. 1959. Handbook of physiology, p. 199-213.In John Field. (Ed), Section 1: Neurophysiology.American Physiological Society, Washington, Vol.I.
Fatt, P., and B. Katz. 1951. An analysis of the end-plate potential recorded with an intra-cellularelectrode. J. Physiol. 115:320-370.
. 1952. Sub-threshold activity at motor nerve-endings. J. Physiol. 117:109-128.
Florey, E., and G. Hoyle. Nervous inhibition. E.Florey, (Ed.), Pergamon Press, London, 105-110.
Frank, G. B. 1960. Effects of changes in extra-cellular calcium concentration on the potassium-induced contracture of frog's skeletal muscle. J.Physiol. 151:518-538.
Furshpan, E. J. 1959. Handbook of Physiology.Section 1: Neurophysiology, John Field, (Ed.),American Physiological Society, Washington, Vol.I, Chapt. VII, 239-254.
Gergely, J. 1959. The relaxing factor of muscle.Ann. N. Y. Acad. Sciences 81:490-504.
Ginsborg, B. L. 1960. Some properties of avianskeletal muscle fibers with multiple neuromuscu-lar junctions. J. Physiol. 154:581-598.
Harreveld, A. von, and C. A. G. Wiersma. 1939.The function of the quintuple innervation of acrustacean muscle. J. Exp. Biol. 16:121-133.
Hasselbach, W. 1956. Die wechselwirkung ver-schiedener Nukleosidtriphosphate mit Aktomyo-sin im Gelzustand. Biochim. et Biophys. Acta 20:355-368.
. 1957. Die Bindung von Adenosindiphos-phat, von anorganischem Phosphat und von Er-dalkalien an die Structurproteine des Muskels.Biochem. Biophysica Acta 25:562-574.
Hayashi, T. 1956. The chemical physiology of ex-citation in muscle and nerve. Nakayama-shioten,Tokyo, 152 pp.
Heilbrunn, L. V., and F. J. Wiercinski. 1946. Theaction of various cations on muscle protoplasm.Biol. Bull. 91:237.
Hill, A. V. 1949. On the time required for diffu-sion and its relation to processes in muscle. Proc.Roy. Soc. (London), B, 135:446-453.
Horridge, G. A. 1956. The nerves and muscles ofmedusae. V. Double innervation in Scyphozoa.J. Exp. Biol. 33:366-383.
Hoyle, G. 1957a. Comparative physiology of thenervous control of muscular contraction. Cam-bridge University Press, Cambridge, 147 pp.
. 1957b. Nervous control of insect muscles,p. 73-98. In B. T. Scheer (Ed.), Recent advancesin invertebrate physiology. University of OregonPublications, Eugene, Oregon.
. 1958. Studies on neuromuscular transmis-sion in I.imulus. Biol. Bull. 115:209-218.
Hoyle, G., and C. A. G. Wiersma. 1958. Couplingof membrane potential to contraction in crusta-cean muscles. J. Physiol. 143:441-453.
Hutter, O. F.. and W. Trautwein. 1956. Vagal andsympathetic effects on the pacemaker fibers in thesinus venosus of the heart. J. Gen. Physiol. 39:715-733.
Huxley, A. F., and R. E. Taylor. 1958. Local ac-tivation of striated muscle fibers, j . Physiol. 144:426-441.
Jenkinson. I). H., and J. G. Nicholls. 1960. Acetyl-
Dow
nloaded from https://academ
ic.oup.com/icb/article/2/1/55/2110825 by guest on 11 January 2022

MUSCLE AND ITS NEURAL CONTROL 65
choline contractures of denervated skeletal musclein isotonic potassium sulfate. J. Physiol. 152:12P-13P.
Jewell, B. R. 1959. The nature of the phasic andtonic responses of the anterior byssal retractormuscle of Mytilus. J. Physiol. 149:154-177.
Johnson, W. J., J. Kahn, and A. G. Szent-Gyorgyi.1959. Paramyosin and contraction of "CatchMuscles." Science 130:160-161.
Johnson, W. H., and B. M. Twarog. 1960. Thebasis for prolonged contractions in molluscanmuscles. J. Gen. Physiol. 43:941-960.
Kolodny, R. L., and W. G. Van der Kloot. 1961.Contraction of smooth muscles in non-ionic solu-tions. Nature 190:786-788.
Kuffler, S. W., and E. M. Vaughan Williams. 1953.Small-nerve junctional potentials. The distribu-tion of small motor nerves to frog skeletal muscle,and the membrane characteristics of the fibersthey innervate. J. Physiol. 121:289-317.
Marsh, B. B. 1952. The effects of adenosine tri-phosphate on the fiber volume of a muscle ho-mogenate. Biochim. Biophys. Acta 9:247-260.
Nagai, T., and K. Uchida. 1960. Effect of somecontracture-producing agents on glycerol-extractedmuscle fiber relaxed with relaxing factor. Bio-chim. Biophys. Acta 44:334-340.
Needham, D. M. 1960. Biochemistry of muscularaction, p. 55-104. In G. H. Bourne (Ed.), Thestructure and function of muscle. Vol. 2. Bio-chemistry and Physiology, Academic Press, NewYork.
Niedergerke, R. 1955. Local muscle shortening byintra-cellularly applied calcium. J. Physiol. 128:12-13.
Pople, W. and D. W. Ewer. 1955. Studies on themyoneural physiology of Echinodermata. II. Cir-cumoral conduction in Cucumaria. J. Exp. Biol.32:59-69.
Porter, K. K., and G. E. Palade. 1957. Studies onthe endoplasmic reticulum. III. Its form and dis-tribution in striated muscle cells. J. Biophys. Bio-chem. Cytol. 3:269-300.
Pringle, J. W. S. 1949. The excitation and contrac-tion of the flight muscles of insects. J. Physiol.108:226-232.
. 1954. The mechanism of the myogenicrhythm of certain insect striated muscles. J.Physiol. 124:269-291.
. 1957. Myogenic rhythms, p. 99-116. In
B. T. Scheer, (Ed.), Recent advances in inverte-brate physiology. University of Oregon, Eugene,Oregon.
Prosser, C. L. 1960. Comparative physiology of ac-tivation of muscles, with particular attention tosmooth muscles. The structure and function ofmuscle. G. H. Bourne, (Ed.) Academic Press,New York, Vol. II, Chap. 8, 387-434.
Prosser, C. L., H. J. Curtis, and D. M. Travis. 1951.Action potentials from some invertebrate non-stri-ated muscle. J. Cell. Comp. Physiol. 38:299-319.
Ringer, S. 1880. Concerning the influence exertedby each of the constituents of the blood on thecontraction of the ventricle. J. Physiol. 3:380-393.
Robertson, D. A. 1960. Calcium and contractilityin depolarized smooth muscle. Nature 187:316.
Roeder, K. D. 1951. Movements of the thorax andpotential changes in the thoracic muscles of in-sects during flight. Biol. Bull. 100:95-106.
Singh, I. 1944. An electrolyte-free medium for un-striated muscle. Curr. Sci. 10:251-252.
Singh, I., and A. K. Acharya. 1957a. Excitation ofunstriated muscle without any ionic gradient acrossthe membrane. Indian J. Physiol. Pharm. 1:265-269.
Singh, I., K. B. Sehra, and J. I. Singh. 1945. Anelectrolyte-free medium for the frog heart andgraded responses of the heart muscle. CurrentSci. 14:152.
Twarog, B. M. 1954. Responses of a molluscansmooth muscle to acetylcholine and 5-hydroxy-tryptamine. J. Cell. Comp. Physiol. 44:141-163.
Weiss, P. 1950. Experimental analysis of coordina-tion by the disarrangement of control-peripheralrelations. Symposia. Soc. Exp. Biol. 4:92111.
Wiersma, C. A. G. 1961. The neurorauscular sys-tem. In T. H. Waterman, (Ed.), The physiologyof the Crustacea, Vol. II. Academic Press, NewYork.
Wiersma, C. A. G., and A. von Harreveld. 1938.The influence of the frequency of stimulation onthe slow and the fast contraction in crustaceanmuscle. Physiol. Zool. 11:75-81.
Wilson, D. M. 1960a. Nervous control of move-ment in cephalopods. J. Exp. Biol. 37:57-72.
. 1960b. Nervous control of movement inannelids. J. Exp. Biol. 37:46-56.
, Winegrad, J. 1960. The relationship of calcium up-take to contraction in guinea pig atria. ThePhysiologist 3:179.
Dow
nloaded from https://academ
ic.oup.com/icb/article/2/1/55/2110825 by guest on 11 January 2022

Dow
nloaded from https://academ
ic.oup.com/icb/article/2/1/55/2110825 by guest on 11 January 2022