
Morphological and physiological differences in Scots pine ...
10
Morphological and physiological differences in Scots pine seedlings of six seed origins M. P. PERKS 1 AND H. M. MCKAY 2 1 I.E.R.M., University of Edinburgh, Darwin Building, Mayfield Road, Edinburgh, EH9 3JU, Scotland 2 Northern Research Station, Forestry Commission, Roslin, Midlothian, EH25 9SY, Scotland Summary One-year-old seedlings of Scots pine (Pinus sylvestris L.) of four native seed origins (Loch Maree Islands, Glengarry/Glen Morriston, Glen Affric and Abernethy), a commercial British seedlot, and a seedlot from Hedesunda, in middle Sweden, were compared at monthly intervals from October 1993 to April 1994. Seedling morphology, root condition, root frost hardiness and bud dry matter were determined at each date. There were clear morphological differences among seed origins. Seedlings raised from the commercial seedlot (A70) were larger but had a poorer root:shoot ratio than the other seed ori- gins. Of the native pines tested, the Loch Maree Islands origin allocated a larger proportion of its photosynthate to fine roots and needles and smaller proportion to woody structures. Seedlings raised from the commercial British seedlot tended to have poorer bud lignification than the other origins and also, in autumn, higher electrolyte leakage rates from its fine roots. During winter, the Swedish origin had the lowest fine root electrolyte leakage. Seedlings of all origins showed a progressive increase in fine root hardiness towards mid-winter with maximum hardiness (—7°C) in January. Dehardening occurred over subsequent months reaching -3°C in April. Differences among origins were evident. The Swedish seedlot developed greater frost resis- tance than the other origins, hardening began earlier in autumn and dehardening began later in spring. The commercial seedlot hardened later than the other origins but reached a similar level of frost hardiness by January. Of the native pines, seedlings of the Loch Maree Islands origin were slowest to develop root hardiness. Introduction natura , woodlands > ( AnO n., 1994), is to main- There is currently considerable interest in the tain the genetic integrity of native species, regeneration and expansion of existing remnant including identifiable sub-populations of native populations of Scots pine in Scotland. Current pine. In practice, the introduction of outside policy, outlined in 'The management of semi- seed sources is frequently discouraged but in O Iraritmc at Chartered F o r a t m , 1997 Forcary, Vol. 70, No. 3, 1997 Downloaded from https://academic.oup.com/forestry/article/70/3/223/543881 by guest on 24 June 2022
-
Upload
khangminh22 -
Category
Documents
-
view
3 -
download
0
Transcript of Morphological and physiological differences in Scots pine ...

Morphological and physiologicaldifferences in Scots pine seedlings ofsix seed originsM. P. PERKS1 AND H. M. MCKAY2
1 I.E.R.M., University of Edinburgh, Darwin Building, Mayfield Road, Edinburgh, EH9 3JU, Scotland2 Northern Research Station, Forestry Commission, Roslin, Midlothian, EH25 9SY, Scotland
Summary
One-year-old seedlings of Scots pine (Pinus sylvestris L.) of four native seed origins (Loch MareeIslands, Glengarry/Glen Morriston, Glen Affric and Abernethy), a commercial British seedlot,and a seedlot from Hedesunda, in middle Sweden, were compared at monthly intervals fromOctober 1993 to April 1994. Seedling morphology, root condition, root frost hardiness and buddry matter were determined at each date.
There were clear morphological differences among seed origins. Seedlings raised from thecommercial seedlot (A70) were larger but had a poorer root:shoot ratio than the other seed ori-gins. Of the native pines tested, the Loch Maree Islands origin allocated a larger proportion ofits photosynthate to fine roots and needles and smaller proportion to woody structures.
Seedlings raised from the commercial British seedlot tended to have poorer bud lignificationthan the other origins and also, in autumn, higher electrolyte leakage rates from its fine roots.During winter, the Swedish origin had the lowest fine root electrolyte leakage. Seedlings of allorigins showed a progressive increase in fine root hardiness towards mid-winter with maximumhardiness (—7°C) in January. Dehardening occurred over subsequent months reaching -3°C inApril. Differences among origins were evident. The Swedish seedlot developed greater frost resis-tance than the other origins, hardening began earlier in autumn and dehardening began later inspring. The commercial seedlot hardened later than the other origins but reached a similar levelof frost hardiness by January. Of the native pines, seedlings of the Loch Maree Islands originwere slowest to develop root hardiness.
Introduction n a t u r a , w o o d l a n d s > (AnOn., 1994), is to main-There is currently considerable interest in the tain the genetic integrity of native species,regeneration and expansion of existing remnant including identifiable sub-populations of nativepopulations of Scots pine in Scotland. Current pine. In practice, the introduction of outsidepolicy, outlined in 'The management of semi- seed sources is frequently discouraged but inO Iraritmc at Chartered Foratm, 1997 Forcary, Vol. 70, No. 3, 1997
Dow
nloaded from https://academ
ic.oup.com/forestry/article/70/3/223/543881 by guest on 24 June 2022

224 FORESTRY
some situations this would be beneficial. Withinsmall remnant populations, there may not besufficient genetic variation to warrant their usein regeneration (Ennos, 1991). Furthermore,Forrest and Fletcher (1994) point out that it ishighly probable that gene transfer has alreadytaken place between many relict woodlands andnearby planted stands of continental origin orfrom seed collected in other parts of Scotland sothat seed from these relict woodlands will notrepresent the genotypic structure of the parentstand. They also propose that small remnantsare likely to be the result of long-term removalof trees with good growth rate and form; con-sequently the remaining trees may be poorlyadapted to the site. In such instances, transfer ofseed origins would be appropriate.
In order to develop transfer guidelines, it isessential to ensure that seed stock has highgenetic variation and is adapted to local envir-onmental conditions. Genetic techniques such asDNA marking, isozyme and terpene analysisgive valuable information concerning geneticcomposition of populations and their genetichistory but no indication of the abundance andpatterns of variation for quantitative charactersconcerned with adaption to particular environ-mental conditions. In upland temperate regions,provenance transfer is often limited by frost(Cannell, 1984). Recent studies have shown thatseedlings of a selected seed orchard source hadlimited ability to develop root frost hardiness(McKay, 1994), suggesting that both shoot and
root frost hardiness should be considered in theevaluation of provenances' suitability for partic-ular sites.
Scots pine (Pinus sylvestris L.) seedlings of sixseed origins were lifted at monthly intervalsbetween October 1993 and April 1994 andassessed for height, diameter, biomass and bio-mass partitioning, bud lignification, root activ-ity and root frost hardiness. The objective ofthis study was to examine differences amongScots pine origins in a range of physiologicaland morphological characteristics.
Materials and methods
There were four native Scottish seed origins (GlenAffric (NT37), Loch Maree Islands (N701), Glen-garry and Glen Morriston (NT39), and Aber-nethy Forest Lodge (N401) ), one commercialBritish seedlot (A70) and one origin fromHedesunda, Sweden (Table 1). Seedlings weregrown in containers for 6 weeks in glasshouseconditions in spring 1993 and then planted in aForestry Commission research nursery at BushEstate, Roslin, Midlothian (03°ll'W, 55°51'N,170 m above sea level). The nursery beds are gen-tly sloping (1°) to the east and have a brownsandy loam soil. The mean annual rainfall is 866mm. Seedlings were planted in seed origin blocksat 10 cm spacing in 10 rows per 1 metre wide bed.
Seedlings were lifted at the beginning of thefirst full week of every month, from October
Table 1: Geographical and climatic descriptors of six Scots pine seed origins
Provenance Location of origin Height above Mean annualsea level rainfall
Origin* Location of provenance
Glen Affric
Loch Maree
Glengarry/Glen Morriston
Abernethy
A70(Commercial)
Hedesunda
04° 55'W 57° 15'N
05° 26'W 57° 39'N
03" 37'W 56° 24'N
03° 39'W 57° 14'N
Range: 55° 28'Nto 57° 41'N
16o50'E60°23'N
180-450m
10-60 m
120-225 m
210-450 m
Varied
105 m
1778 mm
1778-2032 mm
1778 mm
762-889 mm
Varied
Unknown
Scottish
NW ExclusionZone
SW ExclusionZone
Scottish
Scottish
Swedish
Oxendean, Berwickshire,Scotland
Loch Maree Islands,Scotland
Keillour Forest, Perthshire,Scotland
Abernethy, Scotland
Ringwood, Dorset,England
Hedesunda, Sweden
* Exclusion zone indicates provenance thought to have survived last ice-age in refugia. (G.I. Forrest, personal communica-tion).
Dow
nloaded from https://academ
ic.oup.com/forestry/article/70/3/223/543881 by guest on 24 June 2022

MORPHOLOGICAL AND PHYSIOLOGICAL DIFFERENCES IN SCOTS PINE 225
1993 to April 1994. Sixty seedlings of each ori-gin were randomly removed for morphologicaland physiological assessments. In January, lift-ing was not feasible until the second week of themonth as the ground was frozen. After lifting,all plants were stored at 4°C in sealed plasticbags for a maximum of 12 days until physiolog-ical and morphological tests were completed.All tests were conducted on 20 replicates of eachorigin with the exception of the triphenyl tetra-zolium chloride test for which only 10 replicatesper origin were used. Before testing, plant tis-sues were washed carefully in cold tap water toremove soil material.
The morphological characteristics measuredwere height (mm) from root collar to terminalbud, diameter (mm) as the average of two read-ings at 90° to one another, measured midwaybetween root collar and cotyledon scar, and dryweights of fine roots (<2 mm diameter), woodyroots (>2 mm diameter), stem and needles after24 h at 80°C. The level of lignification of the ter-minal bud was assessed by dry matter percentageof the top 20 mm of the leading shoot (Ritchie,1984). Two aspects of the condition of undam-aged fine (i.e. <2 mm diameter) roots wereexamined—membrane function as assessed byelectrolyte leakage and dehydrogenase activity asassessed by tetrazolium chloride reduction. Rootelectrolyte leakage was measured using themethod of McKay (1992); leakage of cell con-tents during 24 h at 20°C was measured by theconductivity of the solution bathing the sampleand expressed as a percentage of the leakagefrom the same sample following autoclaving tolyse all the cells. Triphenyl tetrazolium chloride(TTC) reduction was determined using themethod of LindstrOm and Nystrom (1987) withminor modifications (no wetting agent was usedand samples were only 100 mg ± 0.02 g fineroots). When reduced by the enzyme dehydroge-nase, TTC formed a pink compound which wasextracted and quantified using a spectropho-tometer. Root frost hardiness was assessed bysubjecting rinsed fine root samples to a simu-lated frost in a programmable freezing cabinet.Samples were cooled from 15°C to 1°C in 1 h,cooled to a set minimum temperature at 2°C h~',a rate which avoids rapid cooling injury by intra-cellular ice formation (Levitt, 1980), maintainedat the set temperature for 3 h, warmed at 3°C
h ' to 1°C and taken back to a maintained tem-perature of 15°C. A series of minimum tempera-tures was selected to bracket the temperaturecausing 50 per cent leakage from the root sys-tem. After the freezing cycle was complete, elec-trolyte leakage was measured.
Root electrolyte leakage (REL) and TTCvalues of the undamaged seedlings, bud lignifi-cation and morphological measurements(height, diameter, dry weight and dry weightratios) were analysed using SAS (SAS InstituteInc., 1987) analysis of variance program. RELpercentages were first subject to arcsine trans-formations as temperature variances wereunequal. The temperatures giving 50 per centfine root leakage after 24 h were estimated foreach provenance and lifting date by fitting amean linear regression line over the range offreezing temperatures applied. Regression lineswere tested for parallelism by a further statisti-cal test. The estimated temperature causing 50per cent root electrolyte leakage (LT50) was thencalculated from the appropriate slope and inter-cept.
Results
Sampling date had a significant (P<0.05) effecton all parameters. Seed origin significantly(P<0.005) affected all parameters except drymatter content. However, there were significantinteractions between sampling date and originon all parameters except biomass. In caseswhere interactions were significant, generaltrends plus data for significantly different(P<0.05) individual sampling dates and originsare described.
Stem dimensions (Figure 1)
Seedlings from the commercial seedlot weretaller with a greater root collar diameter thanthe other origin. Seedlings of Loch MareeIslands seed origin had the smallest height anddiameter.
Biomass
Across all lifting dates, the commercial seedlothad significantly greater stem and needle
Dow
nloaded from https://academ
ic.oup.com/forestry/article/70/3/223/543881 by guest on 24 June 2022
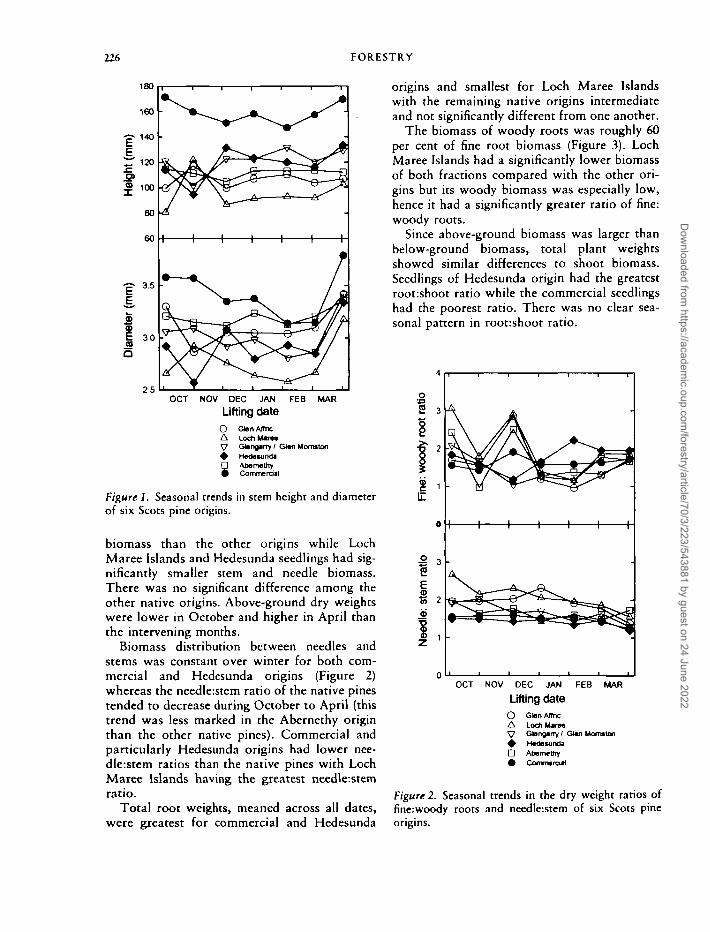
226 FORESTRY
OCT NOV DEC JAN FEB MAR
Lifting dateO GlonAffncA Loch M m *V Gfengsny / G4«fl Moowton• Heduunda
S AijemethyCommercial
Figure 1. Seasonal trends in stem height and diameterof six Scots pine origins.
biomass than the other origins while LochMaree Islands and Hedesunda seedlings had sig-nificantly smaller stem and needle biomass.There was no significant difference among theother native origins. Above-ground dry weightswere lower in October and higher in April thanthe intervening months.
Biomass distribution between needles andstems was constant over winter for both com-mercial and Hedesunda origins (Figure 2)whereas the needle:stem ratio of the native pinestended to decrease during October to April (thistrend was less marked in the Abernethy originthan the other native pines). Commercial andparticularly Hedesunda origins had lower nee-dle:stem ratios than the native pines with LochMaree Islands having the greatest needle:stemratio.
Total root weights, meaned across all dates,were greatest for commercial and Hedesunda
origins and smallest for Loch Maree Islandswith the remaining native origins intermediateand not significantly different from one another.
The biomass of woody roots was roughly 60per cent of fine root biomass (Figure 3). LochMaree Islands had a significantly lower biomassof both fractions compared with the other ori-gins but its woody biomass was especially low,hence it had a significantly greater ratio of fine:woody roots.
Since above-ground biomass was larger thanbelow-ground biomass, total plant weightsshowed similar differences to shoot biomass.Seedlings of Hedesunda origin had the greatestroot:shoot ratio while the commercial seedlingshad the poorest ratio. There was no clear sea-sonal pattern in root:shoot ratio.
OCT NOV DEC JAN FEB MAR
Lifting dateO GlenAITncA LodiMjreeV Gtonfliny / Gt«n Uomttm• HtOemndaD Atjemettiy• Commarcul
Figure 2. Seasonal trends in the dry weight ratios offine:woody roots and needle:stcm of six Scots pineorigins.
Dow
nloaded from https://academ
ic.oup.com/forestry/article/70/3/223/543881 by guest on 24 June 2022

MORPHOLOGICAL AND PHYSIOLOGICAL DIFFERENCES IN SCOTS PINE 227
OCT NOV DEC JAN FEB MAR
Lifting dateO GtenAflncA Loch M*TMV Gttngirry / Glen MoniKon4> HedewndaD Abemothy4} Commercial
Figure 3. Seasonal trends in the dry weight of fineand woody roots of six Scots pine origins.
Dry matter content of the terminal bud
In general, dry matter content increased rapidlyduring October and then more slowly duringNovember (Figure 4). Maximum values tendedto be reached in early December or early Janu-ary before decreasing during January and Feb-ruary. In October, Abernethy origin had thegreatest dry matter content. Initially, Hedesundaorigin had a low degree of bud lignification butdry matter content showed a rapid earlyincrease and reached its maximum in earlyNovember while it had a lower value of drymatter content than the other origins in April.Over November to February, Glen Affric origintended to have a higher degree of lignificationwhile the commercial seedlings tended to have alower value than the other origins.
OCT NOV FEB MARDEC JAN
Lifting dateO GlenABncA lodiMareeV Gtengmy/Glenlylarirtjn• HtdnundaD Abemaihy• Commercial
Figure 4. Seasonal trends in the terminal bud drymatter percentage of six Scots pine origins.
Condition of undamaged fine roots
The general seasonal trend was for REL valuesto decrease slightly from c. 20 per cent in Octo-ber to a minimum of 15 per cent in Decemberbefore increasing during January and February(Figure 5). The low value in April is not dis-cussed further because of methodological prob-lems on that date. Three reasons are suggestedfor the significant lifting date X origin interac-tion. First, Glen Affric seedlings, compared withthe other origins, had lower leakage rates inOctober and a higher rate in January so thatthey did not show the general pattern ofdecreasing REL during October to January.Second, seedlings of the commercial British seedorigin had, in general, slightly higher leakagerates than the other origins during October.Third, seedlings of Hedesunda provenance hadlower leakage rates during mid-January to mid-March.
Estimates of dehydrogenase activity were veryvariable from lift to lift (Figure 6). Althoughthere were some significant differences among
Dow
nloaded from https://academ
ic.oup.com/forestry/article/70/3/223/543881 by guest on 24 June 2022
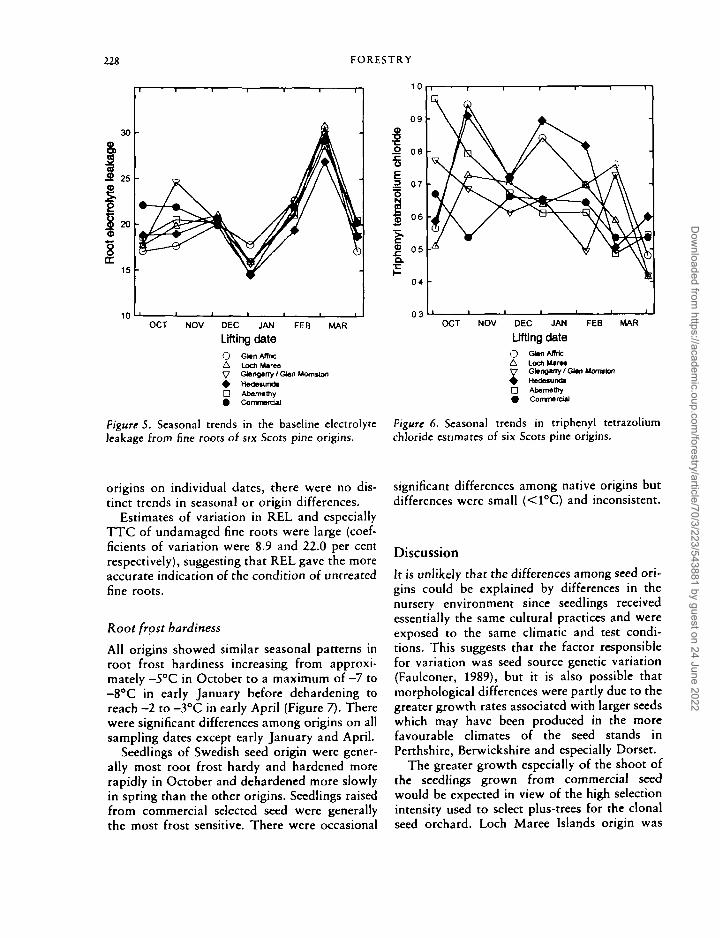
228 FORESTRY
OCT NOV DEC JAN FEB MAR
Lifting dateO Gkn AffncA Loch MlreeV Gtanfliny/Glefi Monwton+ HedwundiD Abenwttiy# Corrwmfdal
Figure 5. Seasonal trends in the baseline electrolyteleakage from fine roots of six Scots pine origins.
OCT NOV DEC JAN
Lifting dateFEB MAR
O G)«nAltricA LochUvMy Gtengany / Ql*n Montrton+ HadetundaD Abemathy• Commero»l
Figure 6. Seasonal trends in triphenyl tetrazoliumchloride estimates of six Scots pine origins.
origins on individual dates, there were no dis-tinct trends in seasonal or origin differences.
Estimates of variation in REL and especiallyTTC of undamaged fine roots were large (coef-ficients of variation were 8.9 and 22.0 per centrespectively), suggesting that REL gave the moreaccurate indication of the condition of untreatedfine roots.
Root frost hardiness
All origins showed similar seasonal patterns inroot frost hardiness increasing from approxi-mately —5°C in October to a maximum of -7 to-8°C in early January before dehardening toreach -2 to -3°C in early April (Figure 7). Therewere significant differences among origins on allsampling dates except early January and April.
Seedlings of Swedish seed origin were gener-ally most root frost hardy and hardened morerapidly in October and dehardened more slowlyin spring than the other origins. Seedlings raisedfrom commercial selected seed were generallythe most frost sensitive. There were occasional
significant differences among native origins butdifferences were small (<1°C) and inconsistent.
Discussion
It is unlikely that the differences among seed ori-gins could be explained by differences in thenursery environment since seedlings receivedessentially the same cultural practices and wereexposed to the same climatic and test condi-tions. This suggests that the factor responsiblefor variation was seed source genetic variation(Faulconer, 1989), but it is also possible thatmorphological differences were partly due to thegreater growth rates associated with larger seedswhich may have been produced in the morefavourable climates of the seed stands inPerthshire, Berwickshire and especially Dorset.
The greater growth especially of the shoot ofthe seedlings grown from commercial seedwould be expected in view of the high selectionintensity used to select plus-trees for the clonalseed orchard. Loch Maree Islands origin was
Dow
nloaded from https://academ
ic.oup.com/forestry/article/70/3/223/543881 by guest on 24 June 2022
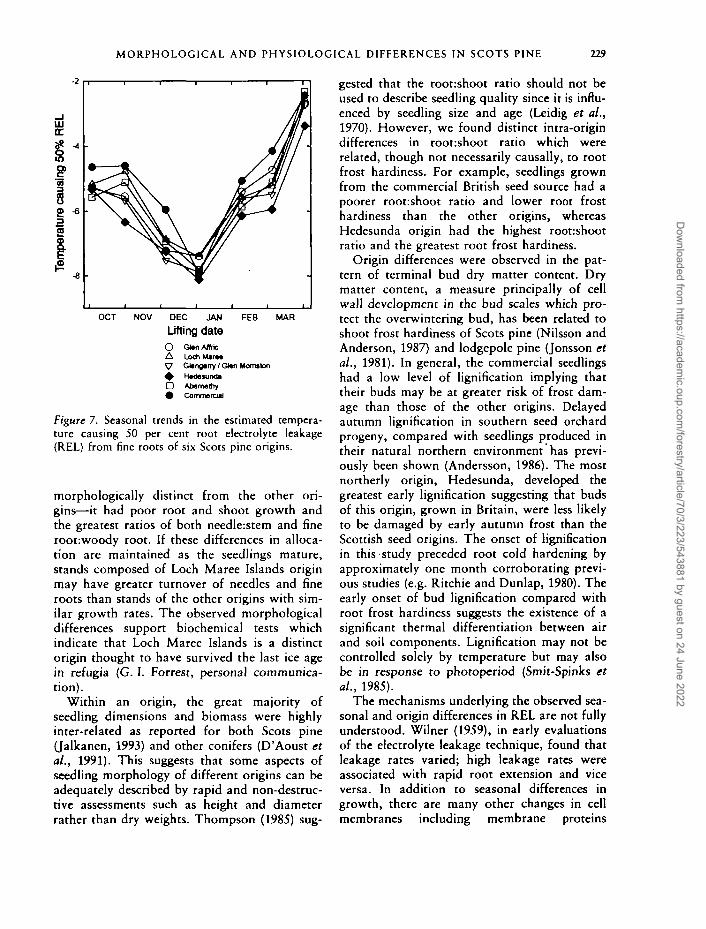
MORPHOLOGICAL AND PHYSIOLOGICAL DIFFERENCES IN SCOTS PINE 229
OCT NOV DEC JAN FEB MAR
Lifting dateO GI«nAffricA LochMvt*V Gttngtny / Gltn Momsion+ HwltiutKtaD Abemethy# CommercuJ
Figure 7. Seasonal trends in the estimated tempera-ture causing 50 per cent root electrolyte leakage(REL) from fine roots of six Scots pine origins.
morphologically distinct from the other ori-gins—it had poor root and shoot growth andthe greatest ratios of both needle:stem and fineroot:woody root. If these differences in alloca-tion are maintained as the seedlings mature,stands composed of Loch Maree Islands originmay have greater turnover of needles and fineroots than stands of the other origins with sim-ilar growth rates. The observed morphologicaldifferences support biochemical tests whichindicate that Loch Maree Islands is a distinctorigin thought to have survived the last ice agein refugia (G. I. Forrest, personal communica-tion).
Within an origin, the great majority ofseedling dimensions and biomass were highlyinter-related as reported for both Scots pine(Jalkanen, 1993) and other conifers (D'Aoust etal., 1991). This suggests that some aspects ofseedling morphology of different origins can beadequately described by rapid and non-destruc-tive assessments such as height and diameterrather than dry weights. Thompson (1985) sug-
gested that the root:shoot ratio should not beused to describe seedling quality since it is influ-enced by seedling size and age (Leidig et al.,1970). However, we found distinct intra-origindifferences in root:shoot ratio which wererelated, though not necessarily causally, to rootfrost hardiness. For example, seedlings grownfrom the commercial British seed source had apoorer root:shoot ratio and lower root frosthardiness than the other origins, whereasHedesunda origin had the highest root:shootratio and the greatest root frost hardiness.
Origin differences were observed in the pat-tern of terminal bud dry matter content. Drymatter content, a measure principally of cellwall development in the bud scales which pro-tect the overwintering bud, has been related toshoot frost hardiness of Scots pine (Nilsson andAnderson, 1987) and lodgepole pine (Jonsson etal., 1981). In general, the commercial seedlingshad a low level of lignification implying thattheir buds may be at greater risk of frost dam-age than those of the other origins. Delayedautumn lignification in southern seed orchardprogeny, compared with seedlings produced intheir natural northern environment has previ-ously been shown (Andersson, 1986). The mostnortherly origin, Hedesunda, developed thegreatest early lignification suggesting that budsof this origin, grown in Britain, were less likelyto be damaged by early autumn frost than theScottish seed origins. The onset of lignificationin this study preceded root cold hardening byapproximately one month corroborating previ-ous studies (e.g. Ritchie and Dunlap, 1980). Theearly onset of bud lignification compared withroot frost hardiness suggests the existence of asignificant thermal differentiation between airand soil components. Lignification may not becontrolled solely by temperature but may alsobe in response to photoperiod (Smit-Spinks etal., 1985).
The mechanisms underlying the observed sea-sonal and origin differences in REL are not fullyunderstood. Wilner (1959), in early evaluationsof the electrolyte leakage technique, found thatleakage rates varied; high leakage rates wereassociated with rapid root extension and viceversa. In addition to seasonal differences ingrowth, there are many other changes in cellmembranes including membrane proteins
Dow
nloaded from https://academ
ic.oup.com/forestry/article/70/3/223/543881 by guest on 24 June 2022

230 FORESTRY
(Yoshida and Uemura, 1990). Since proteinsconstitute the specific carriers and channels inthe plasma membrane that mediate movementin and out of cells (Larsson et al., 1990), the dif-ferences among origins in the REL of undam-aged seedlings are thought to reflect geneticallydetermined differences in root activity andpotential hardiness. The low REL of Hedesundaseedlings over December to February may berelated to their more northerly seed origin.Seedlings grown from the commercial seedlothad high REL in October and early Novembersuggesting that their roots continue root growthlonger in autumn than the other origins and thatmembrane changes conferring frost hardinessare delayed. The Glen Affric origin had a dis-tinctive seasonal REL pattern. This may be dueto pollen contamination at the seed orchard inBerwickshire.
The TTC method was less accurate as anindicator of origin differences than the RELmethod. Overall variation in TTC estimateswas high and differences due to sampling datewere small. Problems in the potential accuracyof TTC estimates may have arisen from thesmall sample size—10 individuals per origineach lift date. This importance of sample sizeshows agreement with the findings of LindstrOmand Nystrom (1987).
Electrolyte leakage is a recognized techniquefor quantifying freezing damage to fine roots(Coleman et al., 1992; McKay, 1994). Smit-Spinks et al. (1985) found that electrolyte leak-age was not as accurate an indicator of rootsystem frost damage as visual browning butMcKay (1992, 1994) found that leakage of fineroots was a good objective indicator of plantvitality. In this study, origin differences in frosthardiness of 0.5°C were highly significant(P<0.001).
This study used excised fine roots to assessfrost hardiness. The authors feel that this doesnot invalidate the results since Pellett (1971)reported similar level of hardiness between sev-ered roots and roots from intact plants.
These results tend to confirm that Scots pineas a species has only moderate ability to developfine root hardiness in Scottish winters; it is inter-mediate between Norway spruce, Sitka spruceand Japanese larch, which are more frost hardy,and Douglas fir and Corsican pine which have
very limited ability to harden (LindstrOm andNystrom, 1987; McKay, 1994). All origins had asimilar general sequence of root frost hardi-ness—hardening to a maximum in early Januaryand dehardening during January to March. Thispattern resembled those previously reported inconiferous roots (Smit-Spinks et al., 1985; Cole-man et al., 1992; McKay, 1994).
Among the native Scottish origins examined,differences in root frost hardiness were smalland inconsistent. Significant differences werefound only in January when Loch Maree Islandsand Glen Affric origins were less frost hardythan Abernethy and Glengarry/Glen Morriston.
Compared with the native pines, roots ofHedesunda origin hardened earlier in autumnand dehardened later in spring whereas roots ofthe commercial seedlings were later to hardenand earlier to deharden. Although all originshad the same maximum root frost hardiness,differences were evident for most of the period;they could be ranked commercial <native<Hedesunda. Lindstrdm and Nystrom (1987)and McKay (1994) have also found that south-ern seed sources were less root frost hardy thannorthern ones. Aho (1994) using shoot imped-ance measures and Norrell et al. (1986) usingvisual scoring techniques reported significantlygreater shoot damage to southern provenancescompared with northern provenances in Scandi-navia.
Differences between provenances, in freezingtests of plant parts, due to clonal variation overlatitude of origin (Andersson, 1992) andbetween northern and southern P. sylvestrisprogeny (Norrel et al., 1986) have previouslybeen reported in Swedish trials. Andersson(1992) suggested that seed orchards geographi-cally removed from the site of clonal origin cancause maladapted seed stock owing primarily topollen contamination and some environmentalpreconditioning. The later onset of hardeningand earlier de-acclimation of the southernBritain seed orchard progeny noted in this trialare unlikely to be caused by pollen contamina-tion at the seed orchard site (C.J.A. Samuel, per-sonal communication). The prolonged growingseason of this origin is more likely to be due toplus-tree selection by high intensity methodsused for the establishment of this orchard. Selec-tion, for example for height, was negatively
Dow
nloaded from https://academ
ic.oup.com/forestry/article/70/3/223/543881 by guest on 24 June 2022

MORPHOLOGICAL AND PHYSIOLOGICAL DIFFERENCES IN SCOTS PINE 231
related with field survival (Nilsson and Anders-son, 1987). This study suggests that this may bedue to poor root frost hardiness.
Conclusions and practical implications
There were minor differences among nativeScots pine origins in root frost hardiness yetcommercial seedlings from trees of Scottish ori-gin that had undergone one selection for stemgrowth and form were less hardy than the nativepines by approximately 0.5°C. Continued selec-tion for growth may further reduce the rootfrost hardiness and give rise to major establish-ment problems. At present however, differencesamong origins were small. Consequently, differ-ences among origins in survival of transplantedseedlings are more likely to be related to factorsother than root frost hardiness such asroot:shoot ratio and the quantity and conditionof fine roots. Scots pine had only a moderatecapacity to develop root frost hardiness. Thishas several practical implications. Frost is apotential risk to all Scots pine origins examinedin this study. Bare root seedlings should not bestored below 0°C unless very close temperaturecontrol can be guaranteed to keep the tempera-ture above -2°C. Cell-grown seedlings, if over-wintered outside, should be stored on theground, insulated round the edges and, in airtemperatures of -10°C, given overhead protec-tion.
Minor differences were evident among nativeorigins in seedling biomass and its distribution(seedlings of Loch Maree Islands were the mostdistinct) and in root condition (Glen Affric ori-gin had the most distinct seasonal pattern).Greater differences were found between thenon-selected native origins on one hand and, onthe other, both commercial British stock and theSwedish origin. Many of the distinctive featuresof the commercial seedlings were associatedwith their greater shoot extension. Some of theSwedish provenance's characteristics may beassociated with the more severe climate, at itsorigin. Differences among native seed originswere minor and based on a limited sample.Nevertheless, the results do support the currentpolicy of maintaining the genetic integrity ofnative pine populations.
Acknowledgements
This study was supported by the Scottish ForestryTrust. The authors are grateful to all staff at theForestry Commission Northern Research Station. Wethank Ian White for his statistical help and acknow-ledge Mrs Roz Shields for preparation of this manu-script.
References
Aho, M.L. 1994 Autumn frost hardening of one-year-old Pinus sylvestris (L.) seedlings: Effect of originand parent trees. Scan. J. For. Res. 9, 17-24.
Anon. 1994 The management of semi-natural wood-lands. Native pinewoods. Forestry Practice GuideNo 7. Forestry Practice Division, Forestry Com-mission, Edinburgh, 52pp.
Andersson, B. 1986 Freezing tests of Scots pine (Pinussylvestris L.) seed orchard crops. In Provenancesand Forest Tree Breeding for High Latitudes. Lin-dren, D. (ed.). Swed. Univ. Agric. Sci. Dept. For.Genet. Plant Physiol. Rep. No 6, 99-111.
Andersson, B. 1992 Autumn frost hardiness of Pinussylvestris offspring from seed orchard grafts of dif-ferent ages. Scand. J. For. Res. 7, 367-375.
Cannell, M.G.R. 1984 Analysis of the risk of frostdamage to forest trees in Britain. In Crop Physiol-ogy of Forest Trees. P.M.A. Tigerstedt, P. Puttonenand V. Koski (eds). Proceedings of an internationalconference on managing forest trees as cultivatedplants. July 23-28, 1984. Helsinki Univ. Press.
Coleman, M.D., Hinckley, T.M., McNaughton, G.and Smit, B.A. 1992 Root cold hardiness and nativedistribution of sub-alpine conifers. Can. J. For. Res.22, 932-938.
D'Aoust, A.L., Delisle, C , Girovard, R., Gonzalez, A.and Bernier-Cardou, M. 1991 Characterisation ofcontainerised spruce seedlings prior to outplanting.In Proc. 1991 For. Nurs. Assoc. of British Columbia.F.P. Donnelly and H.W. Lessenburg (comp.).Meeting 1991, 23-27 Sept., Prince George, BC. For.Nurs. Assoc. of BC, 59-63.
Ennos, R.A. 1991 Genetic variation in Caledonianpine populations: origins, exploitation and conser-vation. In Genetic Variation in European Popula-tions of Forest Trees. G. Muller-Starck and M.Ziehe (eds). Saunders Verlag, Frankfurt, 235-249.
Faulconer, R.J. 1989 Using frost hardiness as an indi-cator of seedling condition. Paper presented at thecombined meeting of the Western Forest NurseryCouncil and lntermountain Nurseryman's Assoc,Vernon, BC, 9-11 August, 1988.
Forrest, G.I. and Fletcher, A.M. 1994 Implication ofgenetics research for pinewood conservation. InOur Pinewood Heritage. J. R. Aldhous (ed.),
Dow
nloaded from https://academ
ic.oup.com/forestry/article/70/3/223/543881 by guest on 24 June 2022

232 FORESTRY
Conference Proceedings, Culloden Academy, Inver-ness, 20-22 October, 1994.
Jalkanen, A. 1993 Covariances between morphologi-cal characteristics in bareroot Pinus sylvestris nurs-ery stock. Scan. ]. For. Res. 8, 540-549.
Jonsson, A., Eriksson, G., Dorling, I. and Ifver, J.1981 Studies on frost hardiness of Pinus contortaDougl. seedlings grown in climate chambers. Stud.For. Suec. 157, 47pp.
Larsson, C , Meier, I.M., and Widell, S. 1990 Anintroduction to the plant plasma membrane—itsmolecular composition and organisation. In ThePlant Plasma Membrane. C. Larsson and I. M.Meier (eds). Springer-Verlag, Berlin, Heidelberg.
Leidig, F.T., Borman, F.H. and Wegner, K.F. 1970The distribution of dry matter growth betweenshoot and roots in loblolly pine. Bot. Caz. 131,349-359.
Levitt, J. 1980 Responses of Plants to EnvironmentalStresses. Vol. 1. Chilling, Freezing and High Tem-perature Stresses, 2nd edn. Academic Press, Lon-don, 497pp.
Lindstrom, A. and Nystrom, C 1987 Seasonal varia-tion in root hardiness of container-grown Scotspine, Norway spruce, and lodgepole pine seedlings.Can. J. For. Res. 17, 787-793.
McKay, H.M. 1992 Electrolyte leakage from fineroots of conifer seedlings: a rapid index of plantvitality following cold storage. Can. ) . For. Res. 22,1371-1377.
McKay, H.M. 1994 Frost hardiness and cold-storagetolerance of the root system of Picea sitchensis,Pseudotsuga menziesii, Larix kaempferi and Pinussylvestris. Bare-root seedlings. Scan. J. For. Res. 9,203-213.
Nilsson, J.E. and Andersson, B. 1987 Performance infreezing tests and field experiments of full-sib fam-ilies of Pinus sylvestris (L.). Can. J. For. Res. 17,1340-1347.
Norell, L., Eriksson, G., Ekberg, Z. and Oormling, B.1986 Inheritance of autumn frost hardiness in Pinussylvestris L. seedlings. Theor. Appl. Genet. 72,440-448.
Pellett, H. 1971 Comparison of cold hardiness levelsof root and stem tissues. Can. J. Plant Sci. 51,193-195.
Ritchie, G.A. 1984 Assessing seedling quality. In For-est Nursery Manual: production of bare rootseedlings. Duryea, M.L. and Landis, T.D. (eds).Martinuss Nijhoff and Dr W. Junk Publ.
Ritchie, G.A. and Dunlap, J.R. 1980 Root growthpotential: its development and expression in foresttree seedlings. N.Z. J. For. Sci. 10, 218-248.
SAS Institute Inc. 1987 SAS/STAT Guide for personalcomputers, version 6 Edition. SAS Institute Inc.,Cary, NC. 1028pp. ISBN 1-55544-064-9.
Smit-Spinks, B., Swanson, B.T. and Markhart III,A.H. 1985 The effect of photoperiod and ther-moperiod on cold acclimation and growth of Pinussylvestris. Can. ]. For. Res. 15, 453--460.
Thompson, B.E. 1985 Seedling morphological evalu-ation—What can you tell by looking. In Evaluat-ing Seedling Quality: principles, procedures andpredictive abilities of major tests. Duryea,M.L. (ed.). Oregon State Univ., For. Res. Lab.,Corvallis.
Wilner, J. Note on electrolytic procedure fordifferentiating between frost injury of roots andshoots in woody plants. Can. J. Plant Sci. 39,512-513.
Yoshida, S. and Uemura, M. 1990 Responses of theplasma membrane to cold acclimation and freezingstress. In The Plant Plasma Membrane. C. Larssonand I.M. Meier (Eds). Springer-Verlag, Berlin, Hei-delberg.
Received 5 February 1996
Dow
nloaded from https://academ
ic.oup.com/forestry/article/70/3/223/543881 by guest on 24 June 2022




![Pine Burr [2018] - Internet Archive](https://static.fdokumen.com/doc/165x107/632797d16d480576770d59ea/pine-burr-2018-internet-archive.jpg)















